
Tracing long-term tropical montane ecosystem change in the Eastern Arc Mountains of Tanzania
10
Tracing long-term tropical montane ecosystem change in the Eastern Arc Mountains of Tanzania JEMMA FINCH, 1 * ,† MATTHEW WOOLLER 2 and ROB MARCHANT 1 1 York Institute for Tropical Ecosystems (KITE), Environment Department, University of York, Heslington, York, YO10 5DD, UK 2 Water and Environmental Research Center, Institute of Northern Engineering, University of Alaska Fairbanks, 99775, USA Received 25 July 2013; Revised 3 February 2014; Accepted 24 February 2014 ABSTRACT: Pollen, charcoal, stable carbon and nitrogen isotope, and radiocarbon analyses are used to reconstruct a palaeoenvironmental record from the Eastern Arc Mountains (EAM), Tanzania, dating back >45 000 years. A sedimentary hiatus covering the last glacial-interglacial transition (LGIT) and early to mid Holocene follows the lead up to the Last Glacial Maximum (LGM) period. A late Holocene portion of the record is included as a modern analogue. Before 45 000 cal a BP, very low regional pollen counts imply open vegetation surrounding the site. Subsequently, the development of extensive montane forest characterizes a relatively mesic pre-LGM. The transition into the LGM witnessed a marked shift from C 3 - to C 4 -dominated biomass, representing an expansion of open forest dominated by grasses. These changes may be attributed to a combination of enhanced moisture stress and CO 2 limitation, factors that may have been exacerbated by an intensive fire regime, as evident from the charcoal record. This is supported by a decrease in montane forest taxa, coupled with an associated expansion of C 4 graminoids and ericaceous heathland to form a montane grassland mosaic similar to that previously recorded on other East African mountains around the time of the LGM but not, to date, having been shown in the Eastern Arc Mountains. Copyright # 2014 John Wiley & Sons, Ltd. KEYWORDS: carbon dioxide; Eastern Arc Mountains; LGM; pollen; stable isotopes. Introduction A long tradition of palaeoecological research has established the highly responsive nature of East African mountain ecosystems to past climatic changes, notably through the Last Glacial Maximum (LGM; 26 500–19 000 cal a BP; e.g. Coetzee, 1967; Flenley, 1979; Hamilton, 1982). Montane vegetation changes occurred during the lead up to the LGM in response to alterations in pCO 2 , water availability, temperature and fire regime (Coetzee, 1967; Hamilton, 1987; Bonnefille and Riollet, 1988; Jolly et al., 1997; Jolly and Haxeltine, 1997; Marchant et al., 1997; Street-Perrott et al., 1997, 2004, 2007, 2008; Ficken et al., 1998, 2002; Wooller et al., 2000; Olago, 2001; Rucina et al., 2009). The LGM in East Africa has been characterized as cool and dry, as evident from lake lowstands and the replacement of forested montane ecosystems by open grassland (Bonnefille and Riollet, 1988; Jolly et al., 1997; Street-Perrott et al., 1997; Wooller et al., 2000; Ficken et al., 2002). Based on pollen, geochemistry and grass cuticle analysis from Lake Rutundu on Mt Kenya (3078 m above sea level – m a.s.l.), Ficken et al. (2002) interpreted the vegetation of the last glacial period as comprising a mixture of shrubs and open C 4 grassland reflecting a cold environment and an ecosystem optimized for water-use efficiency. Sacred Lake on Mt Kenya (2350 m a.s.l.) was surrounded by C 4 -rich shrub grasslands during the last glacial period (Street-Perrott et al., 2007). Stable carbon isotope data from lowland equatorial Africa also indicate a shift from C 4 dominance during the glacial to a mixed C 3 /C 4 composition in the Holocene (Sinninghe Damste ´ et al., 2011). Grassland expansion and the presence of Afroalpine taxa, accompanied by a reduction of continuous montane forest into isolated refugial patches, have been recorded at Kashiru Swamp (2240 m a.s.l.) from 34 700 to 18 300 cal a BP, indicative of a dry, cool climate (Bonnefille and Riollet, 1988). These interpretations are concordant with pollen data from Lake Naivasha (1890 m a.s.l.), which indicate an open grassland environment and associated arid conditions from 24 200 to 13 400 cal a BP (Maitima, 1991). The desiccation of large parts of Lake Victoria (1135 m a.s.l.) between 20 000 and 15 000 cal a BP exemplifies the extreme aridification recorded for East Africa around the time of the last glacial period (Talbot and Livingstone, 1989; Johnson et al., 1996; Stager and Johnson, 2008). Diatom analysis at Lake Rukwa (793 m a.s.l.) also indicates a pronounced lowstand around the time of the LGM (Barker et al., 2002). Corresponding pollen data reveal a cooler and probably drier environment at the LGM, as evidenced by open woodland and bushland surrounding the lake with poor floristic diversity (Vincens et al., 2005). Contrary to indications for a dry LGM from elsewhere on the African continent, hydrological proxy data from Lake Challa imply wet conditions (Tierney et al., 2011). Pollen studies from the Eastern Arc Mountains (EAM) of Tanzania (Mumbi et al., 2008; Finch et al., 2009; Fig. 1) indicate relatively slight changes in montane ecosystem composition and distribution during the LGM, here termed the ecosystem-stability model. Pollen and stable carbon isotopic data from Dama Swamp at 2100 m a.s.l. in the Udzungwa Mountain block of the EAM (Fig. 1) record a shift from cool and moist conditions towards cool and dry conditions through the LGM (Mumbi et al., 2008). Yet these inferred changes are accompanied by minor changes in floristic composition, an anomaly that has been attributed to the influence of the Indian Ocean as a source of long-term moisture stability (Fjeldsa ˚ and Lovett, 1997; Lovett et al., 2005). A similarly benign response is recorded from a 48 000- year pollen sequence from Deva-Deva Swamp in the Uluguru Mountain block of the EAM (2600m a.s.l.; Finch et al., 2009). Pollen data from Maundi Crater on the south-eastern slope of Mt Kilimanjaro (2780m a.s.l.) also indicate montane forest persistence during the LGM, albeit with a strong reduction in these taxa (Schu ¨ler et al., 2012), a pattern consistent with the ecosystem stability model. Correspondence: J. M. Finch, as above. E-mail: [email protected] † Present address: Discipline of Geography, School of Agricultural, Earth and Environmental Sciences, University of KwaZulu-Natal, Private Bag X54001, Durban, 4000, South Africa. Copyright # 2014 John Wiley & Sons, Ltd. JOURNAL OF QUATERNARY SCIENCE (2014) 29(3) 269–278 ISSN 0267-8179. DOI: 10.1002/jqs.2699
Transcript of Tracing long-term tropical montane ecosystem change in the Eastern Arc Mountains of Tanzania

Tracing long-term tropical montane ecosystem changein the Eastern Arc Mountains of Tanzania
JEMMA FINCH,1*,† MATTHEW WOOLLER2 and ROB MARCHANT11York Institute for Tropical Ecosystems (KITE), Environment Department, University of York, Heslington, York, YO10 5DD, UK2Water and Environmental Research Center, Institute of Northern Engineering, University of Alaska Fairbanks, 99775, USA
Received 25 July 2013; Revised 3 February 2014; Accepted 24 February 2014
ABSTRACT: Pollen, charcoal, stable carbon and nitrogen isotope, and radiocarbon analyses are used toreconstruct a palaeoenvironmental record from the Eastern Arc Mountains (EAM), Tanzania, dating back >45000 years. A sedimentary hiatus covering the last glacial-interglacial transition (LGIT) and early to mid Holocenefollows the lead up to the Last Glacial Maximum (LGM) period. A late Holocene portion of the record is includedas a modern analogue. Before 45 000 cal a BP, very low regional pollen counts imply open vegetation surroundingthe site. Subsequently, the development of extensive montane forest characterizes a relatively mesic pre-LGM. Thetransition into the LGM witnessed a marked shift from C3- to C4-dominated biomass, representing an expansion ofopen forest dominated by grasses. These changes may be attributed to a combination of enhanced moisture stressand CO2 limitation, factors that may have been exacerbated by an intensive fire regime, as evident from thecharcoal record. This is supported by a decrease in montane forest taxa, coupled with an associated expansion ofC4 graminoids and ericaceous heathland to form a montane grassland mosaic similar to that previously recordedon other East African mountains around the time of the LGM but not, to date, having been shown in the EasternArc Mountains. Copyright # 2014 John Wiley & Sons, Ltd.
KEYWORDS: carbon dioxide; Eastern Arc Mountains; LGM; pollen; stable isotopes.
Introduction
A long tradition of palaeoecological research has establishedthe highly responsive nature of East African mountainecosystems to past climatic changes, notably through the LastGlacial Maximum (LGM; �26 500–19 000 cal a BP; e.g.Coetzee, 1967; Flenley, 1979; Hamilton, 1982). Montanevegetation changes occurred during the lead up to the LGMin response to alterations in pCO2, water availability,temperature and fire regime (Coetzee, 1967; Hamilton, 1987;Bonnefille and Riollet, 1988; Jolly et al., 1997; Jolly andHaxeltine, 1997; Marchant et al., 1997; Street-Perrott et al.,1997, 2004, 2007, 2008; Ficken et al., 1998, 2002; Woolleret al., 2000; Olago, 2001; Rucina et al., 2009). The LGM inEast Africa has been characterized as cool and dry, as evidentfrom lake lowstands and the replacement of forested montaneecosystems by open grassland (Bonnefille and Riollet, 1988;Jolly et al., 1997; Street-Perrott et al., 1997; Wooller et al.,2000; Ficken et al., 2002). Based on pollen, geochemistryand grass cuticle analysis from Lake Rutundu on Mt Kenya(3078m above sea level – m a.s.l.), Ficken et al. (2002)interpreted the vegetation of the last glacial period ascomprising a mixture of shrubs and open C4 grasslandreflecting a cold environment and an ecosystem optimizedfor water-use efficiency. Sacred Lake on Mt Kenya (2350ma.s.l.) was surrounded by C4-rich shrub grasslands during thelast glacial period (Street-Perrott et al., 2007). Stable carbonisotope data from lowland equatorial Africa also indicate ashift from C4 dominance during the glacial to a mixed C3/C4
composition in the Holocene (Sinninghe Damste et al.,2011). Grassland expansion and the presence of Afroalpinetaxa, accompanied by a reduction of continuous montaneforest into isolated refugial patches, have been recorded atKashiru Swamp (2240m a.s.l.) from �34 700 to 18 300 cal a
BP, indicative of a dry, cool climate (Bonnefille and Riollet,1988). These interpretations are concordant with pollen datafrom Lake Naivasha (1890m a.s.l.), which indicate an opengrassland environment and associated arid conditions from�24 200 to 13 400 cal a BP (Maitima, 1991). The desiccationof large parts of Lake Victoria (1135m a.s.l.) between 20 000and 15 000 cal a BP exemplifies the extreme aridificationrecorded for East Africa around the time of the last glacialperiod (Talbot and Livingstone, 1989; Johnson et al., 1996;Stager and Johnson, 2008). Diatom analysis at Lake Rukwa(793m a.s.l.) also indicates a pronounced lowstand aroundthe time of the LGM (Barker et al., 2002). Correspondingpollen data reveal a cooler and probably drier environment atthe LGM, as evidenced by open woodland and bushlandsurrounding the lake with poor floristic diversity (Vincenset al., 2005). Contrary to indications for a dry LGM fromelsewhere on the African continent, hydrological proxy datafrom Lake Challa imply wet conditions (Tierney et al., 2011).Pollen studies from the Eastern Arc Mountains (EAM) of
Tanzania (Mumbi et al., 2008; Finch et al., 2009; Fig. 1)indicate relatively slight changes in montane ecosystemcomposition and distribution during the LGM, here termedthe ecosystem-stability model. Pollen and stable carbonisotopic data from Dama Swamp at 2100m a.s.l. in theUdzungwa Mountain block of the EAM (Fig. 1) record a shiftfrom cool and moist conditions towards cool and dryconditions through the LGM (Mumbi et al., 2008). Yet theseinferred changes are accompanied by minor changes infloristic composition, an anomaly that has been attributed tothe influence of the Indian Ocean as a source of long-termmoisture stability (Fjeldsa and Lovett, 1997; Lovett et al.,2005). A similarly benign response is recorded from a 48 000-year pollen sequence from Deva-Deva Swamp in the UluguruMountain block of the EAM (2600m a.s.l.; Finch et al., 2009).Pollen data from Maundi Crater on the south-eastern slope ofMt Kilimanjaro (2780m a.s.l.) also indicate montane forestpersistence during the LGM, albeit with a strong reduction inthese taxa (Schuler et al., 2012), a pattern consistent with theecosystem stability model.
�Correspondence: J. M. Finch, as above.E-mail: [email protected]†Present address: Discipline of Geography, School of Agricultural, Earth andEnvironmental Sciences, University of KwaZulu-Natal, Private Bag X54001,Durban, 4000, South Africa.
Copyright # 2014 John Wiley & Sons, Ltd.
JOURNAL OF QUATERNARY SCIENCE (2014) 29(3) 269–278 ISSN 0267-8179. DOI: 10.1002/jqs.2699

Additional palaeoecological records are required from theEast African highlands to gain a fuller understanding of therelationships between environmental changes and vegetationecology in the Afrotropics. Such records have the potential toexplain the differential ecosystem response to climatic shiftsevident between existing records. We present a new palae-oecological record (pollen, charcoal, stable carbon andnitrogen isotopes) documenting the period from >45 000 cala BP leading up to the LGM from the Udzungwa Mountainblock (hereafter ‘Udzungwas’; Fig. 1) of the EAM of Kenyaand Tanzania. Although the record after the LGM is followedby a sedimentary hiatus, covering the last glacial-interglacialtransition (LGIT) and early to mid Holocene, we include alate Holocene portion of the record as a ‘modern analogue’with which to compare vegetation that developed leading upto the LGM.
Regional setting and study area
The EAM comprise a chain of isolated highland mountainblocks, which range from the Taita Hills of southern Kenya tothe Makambako Gap in the southern Udzungwa Mountainsof Tanzania (Fig. 1). The mountains are ancient, having beenuplifted at least 30 million years ago as a result of blockfaulting processes (Griffiths, 1993). The Eastern Arc forests areinternationally recognized as one of the world’s most biologi-cally diverse areas and have been designated as a biodiversityhotspot (Myers, 1990; Myers et al., 2000; Mittermeier et al.,2005). The high species richness and endemism of theseforests has been attributed to a long history of geological andclimatic stability, facilitated by the constant moisture supplyof the Indian Ocean (Fjeldsa and Lovett, 1997). Long-termenvironmental stability in the Eastern Arc is supported bypalaeoecological (Mumbi et al., 2008; Finch et al., 2009;Finch and Marchant, 2011), phylogenetic (A.S. Jump, M.Carr, A. Ahrends, R. Marchant, unpublished data) and model-
based (Platts et al., 2008) evidence. However, only threepalaeoecological records have previously been publishedfrom these mountains (Dama, Deva-Deva and KitumbakoSwamps), one of which extends past 40 000 years (Mumbiet al., 2008; Finch et al., 2009; Finch and Marchant, 2011).Our study site is located in the Image Forest Reserve (IFR)
in the southernmost part of the Eastern Arc range, namely theUdzungwas. IFR covers a massive ridge, Mt Selegu, at2450m a.s.l., which runs along the north-eastern extreme ofthe Udzungwas, between Kitonga Gorge and the Great RuahaRiver. The reserve is characterized by extensive areas ofexposed rock faces and boulders, while soils consist of sandyloams over gneiss (Lovett and Congdon, 1990; Lovett andPo�cs, 1993). The coring site is located in the waterloggedMgundamvaha Grassland (2000m a.s.l.; 7˚180S, 36˚13.80E) ina shallow valley surrounded by montane grassland and forest.A small stream drains into the site from the forested side,creating a grass/sedge-dominated marsh. The Mgundamvahasite was selected based on the availability and depth ofsediments for coring.Modern forest distribution in the EAM is determined by
temperature (altitude), rainfall, dry season length and frost(Platts et al., 2008), while rainfall patterns are stronglyinfluenced by sea surface temperature gradients in the IndianOcean (Nicholson, 1996), which brings moisture-laden windsto the eastern slopes. The biannual migration of the Inter-tropical Convergence Zone (ITCZ) results in bimodal rainfallin the northern parts of the mountain range, with brief rainsoccurring between October and December and long rainsfrom March to May (Hastenrath, 2001). Southern areasexperience a single rainy season between November andMay, with a prolonged but variable dry season from June toSeptember. IFR receives an estimated annual rainfall of1500mm a�1 (Lovett and Po�cs, 1993).Vegetation descriptions for IFR are provided by Lovett and
Congdon (1990) and Lovett and Po�cs (1993) as follows. The
Figure 1. Map of the Eastern Arc Mountains within Tanzania, indicating the position of the Image Forest Reserve site within the UdzungwaMountains.
Copyright # 2014 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 29(3) 269–278 (2014)
270 JOURNAL OF QUATERNARY SCIENCE
drier western slopes are characterized by miombo woodland,giving way to grassland with scattered forest patches above�2200m a.s.l. Moist forests occur on the wetter easternslopes and can be divided into upper montane and montaneforest types based on altitude. Dominant species of the uppermontane forest zone (>2300m a.s.l.) include Acokantheralaevigata, Aphloia theiformis, Bersama abyssinica, Cassi-pourea gummiflua, Cassipourea malosana, Cussonia spicata,Ekebergia capensis, Hagenia abyssinica, Maesa lanceolata,Morella salicifolia, Nuxia congesta, Podocarpus latifolius,Prunus africana, Rapanea melanophloes and Syzygium corda-tum. The montane forest zone (1500–2300m a.s.l.) is charac-terized by Aningeria adolfi-fredericii, Casearia battiscombei,Chrysophyllum gorungosanum, Croton macrostachys, Ixorascheffleri, Macaranga kilimandscharica, Neoboutonia macro-calyx and Rawsonia reticulata.Montane grassland (2200–2400m a.s.l.) species include
Cyrtanthus breviflorus, Dierama pendulum, Disa nyikensis,Eulophia clavicornis, Eulophia rara, Impatiens gomopho-phylla, Justicia uncinulata, Lobelia goetzei, Moraea irigensisand Oldenlandia rupicola. In the grassland, forest patchescontain Aphloia theiformis, Cassipourea gummiflua, Hageniaabyssinica, Maesa lanceolata, Morella salicifolia, Podocarpuslatifolius, Rapanea melanophloes and Syzygium cordatum.Epiphytic orchids Microholmeria parva, Mystracidium pul-chellum and Polystachya transvaalensis occur in the grass-lands. Miombo woodlands (1800–1900m a.s.l.; westernslopes) are dominated by Brachystegia boehmii, Brachystegiamicrophyllum, Brachystegia spiciformis, Protea spp. andUapaca kirkiana.
Materials and methods
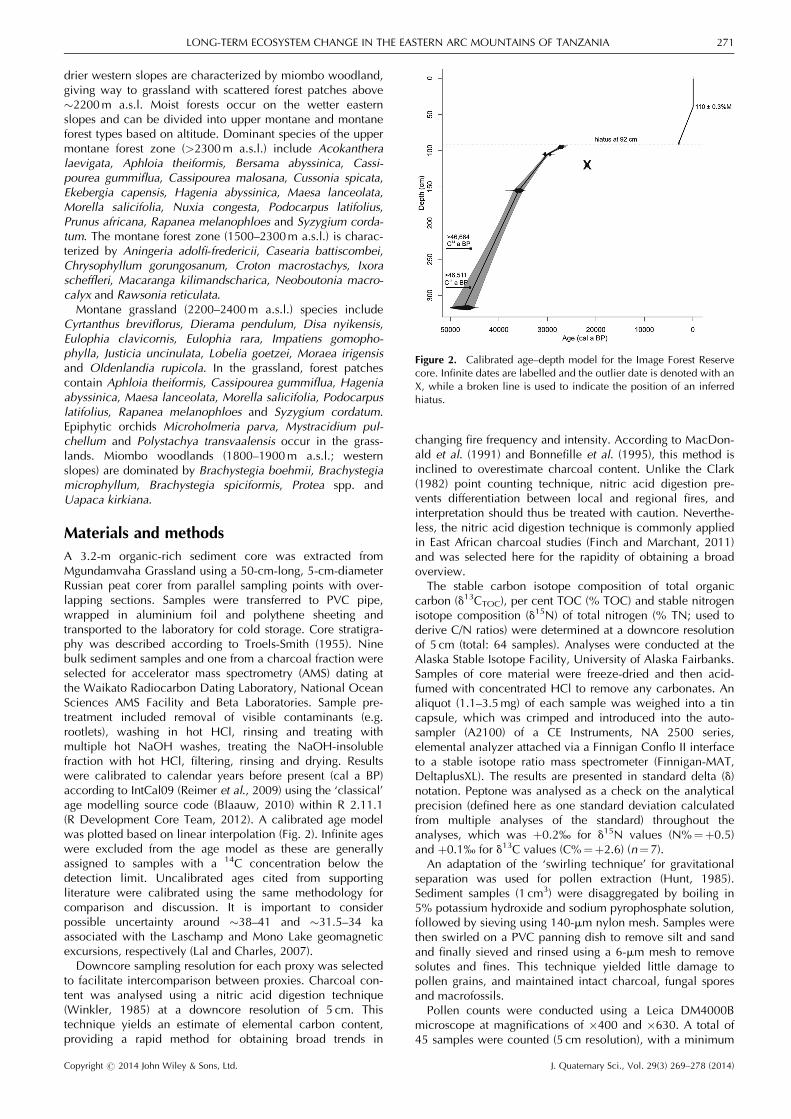
A 3.2-m organic-rich sediment core was extracted fromMgundamvaha Grassland using a 50-cm-long, 5-cm-diameterRussian peat corer from parallel sampling points with over-lapping sections. Samples were transferred to PVC pipe,wrapped in aluminium foil and polythene sheeting andtransported to the laboratory for cold storage. Core stratigra-phy was described according to Troels-Smith (1955). Ninebulk sediment samples and one from a charcoal fraction wereselected for accelerator mass spectrometry (AMS) dating atthe Waikato Radiocarbon Dating Laboratory, National OceanSciences AMS Facility and Beta Laboratories. Sample pre-treatment included removal of visible contaminants (e.g.rootlets), washing in hot HCl, rinsing and treating withmultiple hot NaOH washes, treating the NaOH-insolublefraction with hot HCl, filtering, rinsing and drying. Resultswere calibrated to calendar years before present (cal a BP)according to IntCal09 (Reimer et al., 2009) using the ‘classical’age modelling source code (Blaauw, 2010) within R 2.11.1(R Development Core Team, 2012). A calibrated age modelwas plotted based on linear interpolation (Fig. 2). Infinite ageswere excluded from the age model as these are generallyassigned to samples with a 14C concentration below thedetection limit. Uncalibrated ages cited from supportingliterature were calibrated using the same methodology forcomparison and discussion. It is important to considerpossible uncertainty around �38–41 and �31.5–34 kaassociated with the Laschamp and Mono Lake geomagneticexcursions, respectively (Lal and Charles, 2007).Downcore sampling resolution for each proxy was selected
to facilitate intercomparison between proxies. Charcoal con-tent was analysed using a nitric acid digestion technique(Winkler, 1985) at a downcore resolution of 5 cm. Thistechnique yields an estimate of elemental carbon content,providing a rapid method for obtaining broad trends in
changing fire frequency and intensity. According to MacDon-ald et al. (1991) and Bonnefille et al. (1995), this method isinclined to overestimate charcoal content. Unlike the Clark(1982) point counting technique, nitric acid digestion pre-vents differentiation between local and regional fires, andinterpretation should thus be treated with caution. Neverthe-less, the nitric acid digestion technique is commonly appliedin East African charcoal studies (Finch and Marchant, 2011)and was selected here for the rapidity of obtaining a broadoverview.The stable carbon isotope composition of total organic
carbon (d13CTOC), per cent TOC (% TOC) and stable nitrogenisotope composition (d15N) of total nitrogen (% TN; used toderive C/N ratios) were determined at a downcore resolutionof 5 cm (total: 64 samples). Analyses were conducted at theAlaska Stable Isotope Facility, University of Alaska Fairbanks.Samples of core material were freeze-dried and then acid-fumed with concentrated HCl to remove any carbonates. Analiquot (1.1–3.5mg) of each sample was weighed into a tincapsule, which was crimped and introduced into the auto-sampler (A2100) of a CE Instruments, NA 2500 series,elemental analyzer attached via a Finnigan Conflo II interfaceto a stable isotope ratio mass spectrometer (Finnigan-MAT,DeltaplusXL). The results are presented in standard delta (d)notation. Peptone was analysed as a check on the analyticalprecision (defined here as one standard deviation calculatedfrom multiple analyses of the standard) throughout theanalyses, which was þ0.2‰ for d15N values (N%¼þ0.5)and þ0.1‰ for d13C values (C%¼þ2.6) (n¼7).An adaptation of the ‘swirling technique’ for gravitational
separation was used for pollen extraction (Hunt, 1985).Sediment samples (1 cm3) were disaggregated by boiling in5% potassium hydroxide and sodium pyrophosphate solution,followed by sieving using 140-mm nylon mesh. Samples werethen swirled on a PVC panning dish to remove silt and sandand finally sieved and rinsed using a 6-mm mesh to removesolutes and fines. This technique yielded little damage topollen grains, and maintained intact charcoal, fungal sporesand macrofossils.Pollen counts were conducted using a Leica DM4000B
microscope at magnifications of �400 and �630. A total of45 samples were counted (5 cm resolution), with a minimum
Figure 2. Calibrated age–depth model for the Image Forest Reservecore. Infinite dates are labelled and the outlier date is denoted with anX, while a broken line is used to indicate the position of an inferredhiatus.
Copyright # 2014 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 29(3) 269–278 (2014)
LONG-TERM ECOSYSTEM CHANGE IN THE EASTERN ARC MOUNTAINS OF TANZANIA 271

of 600 pollen grains and spores counted per sample. The lastmetre of the core (220–320 cm) was counted at 10-cmintervals due to low regional pollen counts and poorchronological control associated with this part of the record.The regional pollen sum below 260 cm remains almostconsistently below 20, hampering interpretation of the region-al environmental context. A low pollen count tends to bedominated by common pollen taxa, limiting detection of raretaxa. The uppermost 40 cm of the core was excluded fromthe pollen counts due to dating constraints (see chronologyresults below) and low pollen content.Identification was achieved using a reference collection
(University of York) derived from fresh specimens collected inthe local catchment, in addition to pollen and herbariumspecimens from the National Museums of Kenya and theUniversity of Dar es Salaam Herbarium. Published works andonline databases on East African pollen morphology wereused to aid identification (Bonnefille, 1971; Association desPalynologues de Langue Francaise, 1974; Bonnefille andRiollet, 1980; African Pollen Database, 2013).Pollen and spores were grouped into ecological units to aid
interpretation: upper montane herbs and shrubs, uppermontane forest, montane forest, miombo woodland, aquaticsand undetermined (Table 1). These ecological units are notdiscrete, in particular with regards to species with broadecological tolerances, but serve to aid data description.Interpretation was further aided by calculating percentages inregional and total pollen sums (Table 1). The regional pollensum excluded locally overrepresented, aquatic and undeter-mined taxa, which were reserved for the total pollen sum,including all taxa. To facilitate clear data presentation, raretaxa present at a single depth or contributing <1% of theregional pollen sum were excluded. Pollen data were plottedagainst a dual age/depth axis using Psimpoll 4.26(Bennett, 2005) in conjunction with CorelDRAW X4. Theconstrained incremental sum of squares (CONISS) clustertechnique was applied to the regional pollen data to definepollen zones (Grimm, 1987). Cluster analysis yielded fourpollen zones: I-1 (320–285 cm), I-2 (285–138 cm), I-3 (138–90 cm) and I-4 (90–40 cm).
Results and discussion
Stratigraphy and chronology
The 320-cm core is dominated by organic-rich peat sediments(Fig. 3) becoming more compacted towards the base of thecore. Sediments vary from a dark brown mixture of Substantiahumosa and Argilla staetodes near the base and midsection ofthe core (320–265cm; 220–130 cm) to crumbly dark brownloam layers containing Grana minora (265–250cm; 240–220cm; 130–110cm). There is a discrete grey brown Argillastaetodes layer containing fine grit (250–240cm) and the topof the core is characterized by poorly humified mediumbrown fibrous peat containing Detritus herbosus and Detritusgranosus (110–0 cm) (Troels-Smith, 1955).
Table 1. Pollen and spores identified from the Image Forest Reservesite, and associated ecological groupings (R, regional pollen sum; T,total pollen sum). Rare types excluded from the pollen diagrams aredenoted with an asterisk.
Upper montane herbs and shrubs (R, T)Acanthaceae undiff.�
Acanthaceae: Hypoestes-type�
Acanthaceae: Isoberlinia�
Acanthaceae: Mimulopsis-typeAsteraceae: CarduusAsteraceae: CrassocephalumAsteraceae: Tubuliflorae undiff.
Upper montane forest (R, T)Aquifoliaceae: Ilex mitisCornaceae: Cornus volkensiiEricaceae undiff.Loganiaceae: Nuxia/Ficalhoa-typeOleaceae: Olea�
Rosaceae: Cliffortia nitidulaRosaceae: Hagenia abyssinica
Montane forest (R, T)Araliaceae undiff.Araliaceae: Cussonia-typeAraliaceae: Polyscias fulva-typeAsteraceae: Vernonieae undiff.Balsaminaceae: Impatiens�
Brassicaceae undiff.Celastraceae: Cassine-type�
Combretaceae/Myrsinaceae: Combretum/Myrsine�
Commelinaceae: Commelina forskaeli-type�
Euphorbiaceae: Alchornea-type�
Euphorbiaceae: Croton-typeEuphorbiaceae: Neoboutonia-typeLamiaceae undiff.�
Lamiaceae: Leucas-type�
Lauraceae: Ocotea usambarensisLoganiaceae: Anthocleista-typeLoranthaceae: Englerina woodfordioides�
Malvaceae undiff.Myricaceae: Morella salicifoliaMyrtaceae: Syzygium�
Podocarpaceae: PodocarpusProteaceae: Protea-type�
Sterculiaceae: Dombeya�
Ulmaceae: Celtis�
Miombo woodland (R, T)Anacardiaceae: Ozoroa reticulata-type�
Bombacaceae: Bombax�
Boraginaceae: HeliotropumBuddlejaceae: BuddlejaBurseraceae: CommiphoraCaryophyllaceae undiff.Caryophyllaceae: Cerastium-typeCaryophyllaceae: Silene/Uebelina-type�
Caryophyllaceae: Stellaria mannii-typeConvolvulaceae undiff.Euphorbiaceae: EuphorbiaFabaceae undiff.�
Fabaceae (C): BerliniaFabaceae (C): BrachystegiaHymenocardiaceae: Hymenocardia acidaMimosaceae undiff.�
Mimosaceae: Acacia type III�
Locally common/aquatics (T)Cyperaceae undiff.Eriocaulaceae: Eriocaulon�
Halorrhagaceae: Gunnera perpensaHaloragidaceae: LaurembergiaLycopodiaceae: Lycopodium foveolate-type�
Lycopodiaceae: Lycopodium jussiaei-typePoaceae <40mm
continued
Table 1. (Continued)
Poaceae >40mmPteridophyta: Monoletes undiff.Pteridophyta: Triletes undiff.Restionaceae undiff.�
Undetermined (T)Indeterminable (corroded/broken)Indeterminable (obscured)Undetermined
Copyright # 2014 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 29(3) 269–278 (2014)
272 JOURNAL OF QUATERNARY SCIENCE
Nine AMS ages provide chronological control for therecord (Table 2; Fig. 2). Two infinite ages at 235.5 cm (>46664 14C a BP) and 290 cm (>46 511 14C a BP) introducesome uncertainty as to the age of the basal sediments andemphasize that these are close to or beyond the limit ofradiocarbon dating and should be interpreted with caution.A finite basal age determination of 43 700�1400 14C a BP(45 233–49 709 cal a BP) is obtained at 317.5 cm. Five agedeterminations from 320 to 105 cm provide evidence ofalmost linear accumulation from >45 000 to 27 000 cal aBP, with the exception of a single outlier at 120 cm(18 236� 112 14C a BP; 21 440–22 179 cal a BP). The recentpart of the record from 3000 to 0 cal a BP indicates anincrease in sedimentation rate compared with the oldersection. The age model indicates an erosional hiatus at 92 cmwith a 19 500-year difference in age between 95 and 89 cm,although this does not correspond with an observable,discrete stratigraphic boundary. The lengthy hiatus couldhave arisen from post-depositional erosion, perhaps resultingfrom a wet Holocene scouring event (Marchant et al., 1997),from non-deposition or erosion during a significantly drierperiod surrounding the LGM, or from both. An AMS agedetermination at 40 cm produced a result of 110.8� 0.3%Modern, indicating that sediments at this depth containmodern (post-1954) carbon.
Charcoal content
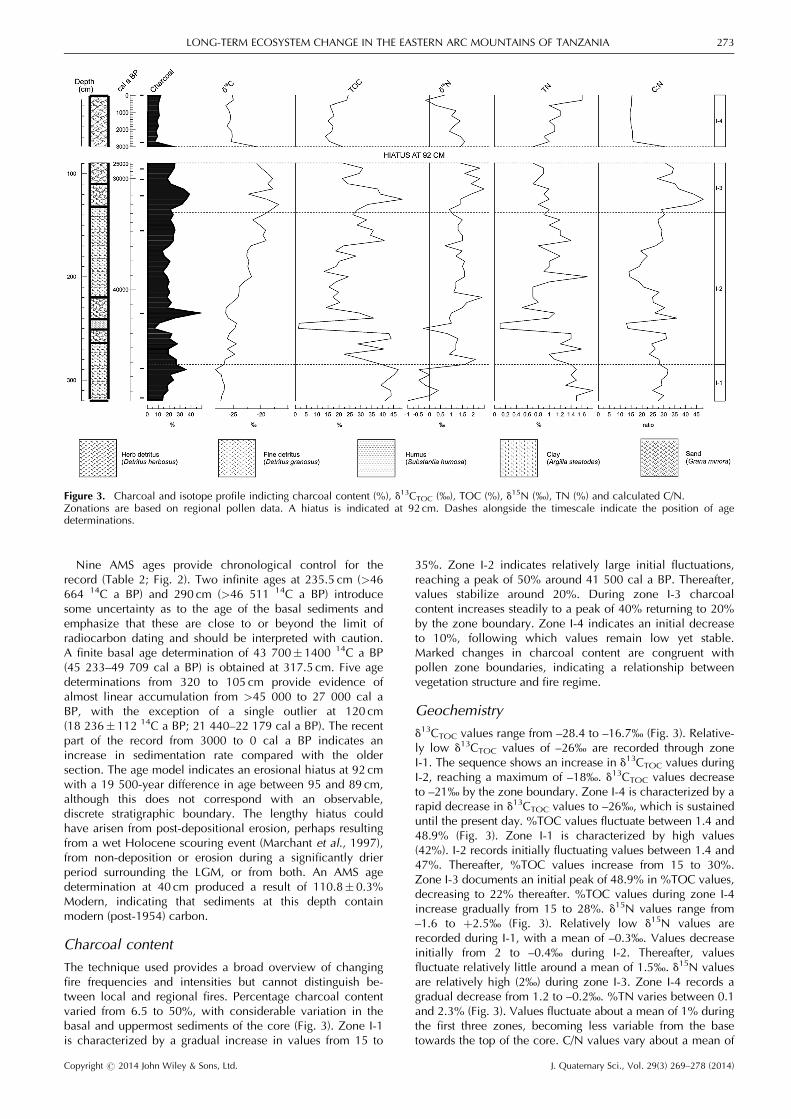
The technique used provides a broad overview of changingfire frequencies and intensities but cannot distinguish be-tween local and regional fires. Percentage charcoal contentvaried from 6.5 to 50%, with considerable variation in thebasal and uppermost sediments of the core (Fig. 3). Zone I-1is characterized by a gradual increase in values from 15 to
35%. Zone I-2 indicates relatively large initial fluctuations,reaching a peak of 50% around 41 500 cal a BP. Thereafter,values stabilize around 20%. During zone I-3 charcoalcontent increases steadily to a peak of 40% returning to 20%by the zone boundary. Zone I-4 indicates an initial decreaseto 10%, following which values remain low yet stable.Marked changes in charcoal content are congruent withpollen zone boundaries, indicating a relationship betweenvegetation structure and fire regime.
Geochemistry
d13CTOC values range from –28.4 to –16.7‰ (Fig. 3). Relative-ly low d13CTOC values of –26‰ are recorded through zoneI-1. The sequence shows an increase in d13CTOC values duringI-2, reaching a maximum of –18‰. d13CTOC values decreaseto –21‰ by the zone boundary. Zone I-4 is characterized by arapid decrease in d13CTOC values to –26‰, which is sustaineduntil the present day. %TOC values fluctuate between 1.4 and48.9% (Fig. 3). Zone I-1 is characterized by high values(42%). I-2 records initially fluctuating values between 1.4 and47%. Thereafter, %TOC values increase from 15 to 30%.Zone I-3 documents an initial peak of 48.9% in %TOC values,decreasing to 22% thereafter. %TOC values during zone I-4increase gradually from 15 to 28%. d15N values range from–1.6 to þ2.5‰ (Fig. 3). Relatively low d15N values arerecorded during I-1, with a mean of –0.3‰. Values decreaseinitially from 2 to –0.4‰ during I-2. Thereafter, valuesfluctuate relatively little around a mean of 1.5‰. d15N valuesare relatively high (2‰) during zone I-3. Zone I-4 records agradual decrease from 1.2 to –0.2‰. %TN varies between 0.1and 2.3% (Fig. 3). Values fluctuate about a mean of 1% duringthe first three zones, becoming less variable from the basetowards the top of the core. C/N values vary about a mean of
Figure 3. Charcoal and isotope profile indicting charcoal content (%), d13CTOC (‰), TOC (%), d15N (‰), TN (%) and calculated C/N.Zonations are based on regional pollen data. A hiatus is indicated at 92 cm. Dashes alongside the timescale indicate the position of agedeterminations.
Copyright # 2014 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 29(3) 269–278 (2014)
LONG-TERM ECOSYSTEM CHANGE IN THE EASTERN ARC MOUNTAINS OF TANZANIA 273

25 during zones I-1 and I-2, followed by an increase to apeak of 48 during I-3. C/N values decrease abruptly to 15 inzone I-4 (Fig. 3).
Pollen data
Pollen preservation was excellent throughout the sequenceand 66 pollen and spore taxa were identified. Where monospe-cific genera are recorded, species names are provided.
I-1 (320–285 cm; >45 000 cal a BP)
This zone is characterized by high variability with fluctua-tions in the relative proportions of the dominant regionalpollen taxa (Fig. 4). The low regional pollen counts towardsthe base of the core suggest a lack of arboreal pollenproducers in the catchment, although low counts should beinterpreted with caution. Podocarpus-dominated montaneforest is highly fluctuating and present at its lowest levelsthroughout the sequence, indicating relatively open vegeta-tion in the catchment. High frequencies of taxa that favour
open ground such as the herb-dominated disturbance indica-tors Tubuliflorae and Vernonieae and open montane foresttaxa such as Neoboutonia suggest this was a relatively mesicperiod. Miombo woodland taxa are sparsely represented.Among local taxa and aquatics, monolete spores dominate,with Cyperaceae subdominant (Fig. 5). Poaceae abundanceremains relatively low (5–10%).
I-2 (285–138 cm; 45 000–34 000 cal a BP)
Podocarpus records a marked recovery at the beginning ofthis zone, increasing from 5 to 80% between 45 000 and 40000 cal a BP. An abrupt decline in Podocarpus to 30% isrecorded at 40 000 cal a BP, although this taxon recoverstowards the zone boundary. A diversity of upper montaneand montane forest taxa is found, the main elements of whichinclude Ericaceae, Hagenia abyssinica, Ilex mitis, Nuxia/Ficalhoa and Malvaceae, supporting the development ofclosed-canopy forest. Hagenia abyssinica becomes estab-lished between 39 000 and 34 000 cal a BP, coincident withCornus volkensii and Nuxia/Ficalhoa. The upper montane
Table 2. Radiocarbon results for the Image Forest Reserve record, indicating calibrated and uncalibrated ages.
Laboratory code Material Depth (cm) d13C (‰) Age (14C a BP) Calibrated age (cal a BP) (2 SD) Method
Wk-23591 Sediment 39.5–40.5 –26.2 110.8�0.3%M � AMSWk-23581 Sediment 88.3–89.3 –24.4 2744�30 2769–2887 (90.9) AMSBeta-274188 Sediment 95–96 –20.4 22 440�120 26 606–27 785 (95) AMSBeta-274189 Sediment 105–106 –19.2 25 650�150 30 216–30 885 (95) AMSBeta-275361 Charcoal fraction 105–106 –17.4 24 630�150 29 097–30 130 (95) AMSWk-23582 Sediment 120–121 –23.4 18 236�112 21 440–22 179 (95) AMSWk-23583 Sediment 155–156 –19.3 31 465�528 34 918–36 907 (95) AMSWk-23584 Sediment 235–236 –29.9 >46 664 � AMSWk-23585 Sediment 289.5–290.5 –28.7 >46 511 � AMSOS-60123 Sediment 317–318 –26.91 43 700�1400 45 233–49 709 (95) Radiometric
Figure 4. Regional pollen profile for the Image Forest Reserve record, indicating core stratigraphy, CONISS-derived zonations, and regional pollensum. A hiatus is indicated at 92 cm. Dashes alongside the timescale indicate the position of age determinations.
Copyright # 2014 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 29(3) 269–278 (2014)
274 JOURNAL OF QUATERNARY SCIENCE

herb taxon Asteraceae increases after 40 000 cal a BP.Miombo taxa Carophyllaceae, Commiphora, Cerastium-typeand Hymenocardia acida become prominent at this time.These changes in regional pollen composition are concurrentwith a peak in the local aquatic Gunnera perpensa. Poaceaeabundance remains relatively low.
I-3 (138–90 cm; �34 000–25 000 cal a BP)
Ericaceae continue to increase, while Podocarpus records acorresponding decline from 80 to 60%. Hagenia abyssinicaand Nuxia/Ficalhoa decrease in the early part of this zoneand closed-canopy taxa Ilex mitis and Cornus volkensii arelargely absent throughout the zone. Cerastium becomesprominent in this zone. The miombo woodland taxonCommiphora records a gradual decrease in the early part ofthis zone. Aquatic weed Laurembergia outcompetes Gunneraperpensa during this zone, dominating the local signal.Poaceae percentages peak around 33 000 cal a BP, butremain relatively high through the remainder of the zone.The I-3/I-4 zone boundary is characterized by major
changes in the frequencies of all pollen taxa, in addition tocharcoal and isotopic values. The lack of coherence betweenadjacent zones provides further evidence for a sedimentaryhiatus at this point in the core.
I-4 (90–40 cm; �3000–0 cal a BP)
The modern-analogue component of this record is characterizedby relatively stable frequencies of a diverse range of upper
montane and montane forest taxa, including Cornus volkensii,Ericaceae, Nuxia–Ficalhoa-type, Vernonieae, Croton-type andMalvaceae. Podocarpus frequencies are moderately high (40%),peaking towards present-day values. Cyperaceae dominate thelocal pollen assemblage.
Environmental reconstruction
Pre-LGM (�50 000–25 000 cal a BP)
Infinite age estimates in the early part of the IFR recordimply that basal sediments are potentially ancient – providingsome long-term context in which to view the remainder ofthe core – but almost certainly cover the pre-LGM period.Before 45 000 cal a BP, proxy data from the site indicate arelatively mesic period characterized by open montane forestwith high levels of ruderal taxa, such as Tubuliflorae andVernonieae (Hamilton et al., 1986). d13CTOC values arerelatively low during this zone, while C/N values vary abouta mean of 30. These proxies suggest a relatively highproportional contribution of organic matter derived from C3
land plants (trees and shrubs) at the site (Meyers andTeranes, 2001; Mackie et al., 2005).A phase of disturbance from 45 000 to 41 000 cal a BP is
inferred from high proportions of ruderal herbs/open grass-land indicators Tubuliflorae and Vernonieae; these could belocally present and reflect an erosional environment alongthe valley bottom. This disturbance is coupled with anincrease in Podocarpus forest (10–80%); Coetzee (1967)
Figure 5. Pollen profile for the Image Forest Reserve record indicating dominant regional taxa and locally common/aquatic taxa.Zonations are based on the regional pollen data. A hiatus is indicated at 92 cm. Dashes alongside the timescale indicate the position of agedeterminations.
Copyright # 2014 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 29(3) 269–278 (2014)
LONG-TERM ECOSYSTEM CHANGE IN THE EASTERN ARC MOUNTAINS OF TANZANIA 275

advocated that values of >20% Podocarpus pollen indicatePodocarpus forest surrounding the site.A peak in charcoal around 41 000 cal a BP may indicate
increased burning, coincident with an abrupt decline inPodocarpus. After 40 000 cal a BP, a phase of forestregeneration is indicated by gradual increases in Podocarpus,concomitant with the establishment of Cornus volkensii,Hagenia abyssinica and Nuxia/Ficalhoa-type. van ZinderenBakker and Coetzee (1988) associated increases in Hageniaabyssinica with a rise in the treeline because of increasingtemperature and humidity. Closure of the forest canopy after�40 000 cal a BP suggests the onset of wetter conditions.This is supported locally by high frequencies of Gunneraperpensa, which grows on the edge of wetland areas, and anincrease in Cyperaceae, indicative of wet stable localconditions. The IFR record indicates a gradual transition inforest composition to include Hagenia abyssinica and Cornusvolkensii, with increasing Podocarpus, during a mesic periodleading up to a relatively arid LGM.Palaeolimnological evidence from Lake Tanganyika points
towards intermediate levels of aridity from 50 000 to 32000 cal a BP, with a more severe arid period from 32 000 to18 000 cal a BP (Felton et al., 2007). In contrast, Kiage andLiu (2006) suggest that cool and moist conditions prevailed inEast Africa after �34 700 cal a BP. Relatively mesic pre-LGMconditions at the IFR are reflected by an inferred increase inhumidity from Maundi Crater, manifested in a spread anddiversification in ericaceous and forest vegetation, includingincreases in Erica, Myrica and Podocarpus (Schuler et al.,2012). Taylor (1990) inferred a slight increase in temperatureand precipitation from upper montane forest expansion atthe expense of ericaceous scrub at Muchoya Swamp (2260ma.s.l.; Rukiga Highlands) during the period leading up to theLGM.The transition period into the LGM (�34 000–25 000 cal
a BP) saw a gradual decline in forest taxa at the IFR,particularly in Hagenia abyssinica and Podocarpus, indicatingan opening up of closed-canopy forest. This is supported by acontemporaneous increase in Tubuliflorae, which may indi-cate more open vegetation. Ericaceae and Poaceae record asteady increase, the latter peaking around �33 000 cal a BP,further implying a transition towards a relatively openenvironment characterized by open grassland and ericaceousheath with much reduced forest cover. After 34 000 cal a BP,ericaceous heath and fire-tolerant C4 grassland expand at theexpense of forest taxa, evidence of whole-scale ecosystemturnover. These changes to ericaceous heath and fire-tolerantC4 grassland, at the expense of forest taxa, could be attributedto cooler and drier conditions and/or increased burning, withcharcoal content peaking about 32 000 cal a BP. The spreadof fire-adaptive ericaceous heathland after 31 000 cal a BPmay indicate environmental conditions becoming cooler anddrier. This phenomenon has been noted for the pre-LGM atSacred Lake, where van Zinderen Bakker and Coetzee (1988)interpret colder and drier conditions from an increase inericaceous heath. Ericaceae act as pioneer species, coloniz-ing disturbed ground (Hamilton, 1982) or even replacingmontane forest following frequent burning. Another possibilityis that the ericaceous belt shifted to lower altitudes in responseto a cooler and drier climate.Based on the d13CTOC values and C/N data, it seems that
the proportional contribution biomass from C4 plants to theorganic matter pool (i.e. grasses and/or sedges; Meyers andTeranes, 2001; Mackie et al., 2005) increased steadilythroughout the pre-LGM period. Despite a minor decrease ind13CTOC values at �32 000 cal a BP, overall values remainrelatively high, indicating a high proportional contribution of
organic matter from C4 plants to the sediment. Relatively highd15N values support a shift towards dry local conditions (e.g.Heaton, 1987; Terwilliger et al., 2008). The local pollen sumis dominated by the aquatic weed Laurembergia, supersedingGunnera perpensa during the initial part of this period.Gunnera perpensa grows on wetland edges while Laurember-gia is semi-aquatic. These findings point to rising water level,with local conditions becoming wetter. After 32 000 cal a BP,Laurembergia begins to decline, suggesting a change towardslocally dry conditions compatible with regional aridification.LGM pollen evidence from highland areas across East
Africa, apart from the EAM closer to the Indian Ocean,suggests an analogous environment to that at the IFR,dominated by grasses and ericaceous scrub with forestmosaic present at low levels (Jolly et al., 1997). A similarexpansion in grasslands with Afroalpine elements is docu-mented for the LGM at Kashiru Swamp in Burundi (Bonnefilleand Riollet, 1988). Results from the Uluguru Mountain block,also part of the Eastern Arc, suggest an increase in Ericaceaefor the duration of the pre-LGM and LGM periods, from 44000 to 19 000 cal a BP, although to a far lesser degree thanrecorded at the IFR. The prevalence of C4 grasslands leadingup to the LGM has been shown in response to sub-ambientpCO2 (Barnola et al., 1987) and enhanced moisture stress(Jolly and Haxeltine, 1997; Street-Perrott et al., 1997; Cerlinget al., 1998), factors which would have been reinforced byincreased burning (Wooller et al., 2000). The LGM ecosystemresponse demonstrated at the IFR does not conform to theproposed model of ‘ecosystem stability’ suggested for othersites in the EAM and in the Uluguru and UdzungwaMountains (Mumbi et al., 2008; Finch et al., 2009). Nor doesthe IFR record show evidence of montane forest persistenceas is recorded from Maundi Crater (Schuler et al., 2012). Thisdifferential response may result from the IFR site being moredistant from the stabilizing influence of the Indian Oceanthan other Eastern Arc mountains with palaeoecologicalrecords.
�3000 cal a BP to present
The ‘modern analogue’ portion of the IFR record is character-ized by a relatively stable mosaic of closed-canopy forest,with grassland patches, which is consistent with the vegeta-tion surrounding the site today. Forest composition includeshigh levels of Podocarpus, coupled with a diversity of uppermontane and montane forest taxa, including the moisture-dependent Cornus volkensii. Vernonieae are present at highpercentages in this zone, implying some possible disturbancetaking place, potentially associated with human activity; withthe exception of this possible indicator, we see no evidenceof anthropogenic activity in the record. The absence ofHagenia abyssinica in this zone is probably too early to berelated to logging, although this remains a favoured targetspecies for timber harvesting in the reserve (Lovett andPo�cs, 1993). The lack of human impacts recorded at this site,probably because of temporal resolution, is in contrast tofungal spore and pollen evidence from nearby Dama Swamp,on the south-western edge of the main Udzungwa Mountainblock (Fig. 1). These data suggest pastoralism and selectivelogging of Podocarpus and Psychotria in the late Holocene(Mumbi et al., 2008). Nevertheless, the Holocene section ofour record lacks tight chronological control and shouldtherefore be interpreted with some caution.Our d13C data indicate a switch from C3-dominated
vegetation towards more C4-dominated vegetation during thelast glacial period and a return to C3 dominance in the lateHolocene part of the record. Charcoal content is lower than
Copyright # 2014 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 29(3) 269–278 (2014)
276 JOURNAL OF QUATERNARY SCIENCE

during the lead up to the LGM, but maintains relativelyhigh values at 20%. There is a significant decrease in d13CTOC
values from 3000 cal a BP, reflecting a switch to C3-dominated local vegetation. From the base to the top ofthe core, we see a remarkable overall trend from C3- toC4-dominated vegetation, and then finally back to C3.
Conclusions
The IFR site provides a >45 000-year sedimentary record ofpast vegetation dynamics in the EAM. Combining pollen,charcoal, carbon and nitrogen isotope, and radiocarbondatasets for this new record, from an under-studied area inEast Africa, allows for the following conclusions to be drawn.
1. Before 45 000 cal a BP, poor representation of regionalpollen types suggests open vegetation within the catchment.
2. The pre-LGM period at the IFR was characterized byrelatively mesic conditions, manifested by forest expansion.
3. The transition into the LGM witnessed a marked shift fromC3- to C4-dominated biomass, corresponding with anexpansion of open grassland and ericaceous heathlandcoupled with a retreat in montane forest taxa. Thesechanges may be attributed to a combination of enhancedmoisture stress and CO2 limitation, factors that may havebeen exacerbated by an intensive fire regime.
4. The section after the LGM is followed by a sedimentaryhiatus, covering the early to mid Holocene.
5. The LGM ecosystem-response recorded from the IFR sitedoes not conform to the ‘ecosystem-stability model’proposed for other EAM sites.
6. The pollen record provides no indication of recent humanactivity, despite a long history of regional deforestationdocumented for the area.
Acknowledgements. This work was supported by EU grant EU-MEXT-CT-2004-517098 to R.M., and contributes to the Global LandProject (www.globallandproject.org) and African Pollen Database(APD). Fieldwork permissions were granted by the Tanzania Commis-sion for Science and Technology and supported by the Institute forResource Assessment, University of Dar es Salaam. Field assistancefrom Bruno Mwano is gratefully acknowledged. Jane Wheeler andMargaret Atherden assisted with the swirling technique while RebeccaSutton and Dave Hay provided logistical support in the laboratory.Pollen identification was aided by Annie Vincens, Guillaume Buchet,Cassian Mumbi, Stephen Rucina and Louis Scott. We are grateful tothe anonymous reviewers, whose constructive comments greatlyimproved the manuscript. KITE researchers Antje Ahrends, Phil Plattsand Al Jump provided invaluable support in this research. RuthHowison professionally drafted Figure 1.
Abbreviations. AMS, accelerator mass spectrometry; EAM, Eastern ArcMountains; IFR, Image Forest Reserve; LGM, Last Glacial Maximum;TN, total nitrogen; TOC, total organic carbon.
References
African Pollen Database (APD). 2013. apd.sedoo.fr. [accessed 25July 2013].
Association des Palynologues de Langue Francaise. 1974. Pollen etSpores d’Afrique tropicale. Travaux et Documents de GiographieTropicale 16. Centre Nation de la Recherche Scientifique (CNRS):Talence.
Barker P, Telford R, Gasse F, et al. 2002. Late Pleistocene andHolocene palaeohydrology of Lake Rukwa, Tanzania, inferred fromdiatom analysis. Palaeogeography, Palaeoclimatology, Palaeoecol-ogy 187: 295–306.
Barnola JM, Raynaud D, Korotkevich YS, et al. 1987. Vostok ice coreprovides 160,000 year record of atmospheric CO2. Nature 329:408–414.
Bennett KD. 2005. Documentation for Psimpoll 4.25 and Pscomb1.03; C Programs for Plotting Pollen Diagrams and AnalysingPollen Data. Uppsala Universitet.
Blaauw M. 2010. Methods and code for ‘classical’ age-modelling ofradiocarbon sequences. Quaternary Geochronology 5: 512–518.
Bonnefille R. 1971. Atlas des pollens de Ethiopie. Principales especesdes forets de montagne. Pollen et Spores 13: 15–72.
Bonnefille R, Riollet G. 1980. Pollens des Savanes d’AfriqueOrientale. Editions du Centre National de la Recherche Scientifique(CNRS): Paris.
Bonnefille R, Riollet G. 1988. The Kashiru Pollen sequence (Burundi):palaeoclimatic implications for the last 40,000 yr B.P. in tropicalAfrica. Quaternary Research 30: 19–35.
Bonnefille R, Riollet G, Buchet G, et al. 1995. Glacial/interglacialrecord from intertropical East Africa, high resolution pollen andcarbon data at Rusaka, Burundi. Quaternary Science Reviews 14:917–936.
Cerling TE, Ehleringer JR, Harris JM. et al. 1998. Carbon dioxidestarvation, the development of C4 ecosystems, and mammalianevolution. Philosophical Transactions of the Royal Society ofLondon. Series B, Biological Sciences 353: 159–171.
Clark RL. 1982. Point count estimation of charcoal in pollenpreparations and thin sections of sediments. Pollen et Spores 24:523–535.
Coetzee JA. 1967. Pollen analytical studies in East and SouthernAfrica. In Palaeoecology of Africa 3, van Zinderen Bakker EM (ed.).Balkema: Cape Town; 1–146.
Felton AA, Russell JM, Cohen AS, et al. 2007. Paleolimnologicalevidence for the onset and termination of glacial aridity from LakeTanganyika, Tropical East Africa. Palaeogeography, Palaeoclimatol-ogy, Palaeoecology 252: 405–423.
Ficken KJ, Street-Perrott FA, Perrott RA, et al. 1998. Glacial/intergla-cial variations in carbon cycling revealed by molecular and isotopestratigraphy of Lake Nkunga, Mt Kenya, East Africa. OrganicGeochemistry 29: 1701–1719.
Ficken KJ, Wooller MJ, Swain DL, et al. 2002. Reconstruction of asub-alpine grass dominated ecosystem, Lake, Rutundu, MountKenya: a novel multi-proxy approach. Palaeogeography, Palae-oclimatology, Palaeoecology 177: 137–150.
Finch JM, Leng MJ, Marchant RA. 2009. Vegetation history of abiodiversity hotspot, the Eastern Arc Mountains of Tanzania.Quaternary Research 72: 111–122.
Finch JM, Marchant RA. 2011. A palaeoecological investigation intothe role of fire and human activity in the development of montanegrasslands in East Africa. Vegetation History and Archaeobotany20: 109–124.
Fjeldsa J, Lovett JC. 1997. Geographical patterns of old and young speciesin African forest biota: the significance of specific montane areas asevolutionary centres. Biodiversity and Conservation 6: 325–346.
Flenley JR. 1979. The late Quaternary vegetational history of theequatorial mountains. Progress in Physical Geography 3: 488–509.
Griffiths CJ. 1993. The geological evolution of East Africa. InBiogeography and Ecology of the Rainforests of Eastern Africa, LovettJC, Wasser SK (eds). Cambridge University Press: Cambridge; 9–22.
Grimm EC. 1987. CONISS: a FORTRAN 77 program for strati-graphically constrained cluster analysis by the method of incre-mental sum of squares. Computers and Geosciences 13: 13–35.
Hamilton AC. 1982. Environmental History of East Africa: A Study ofthe Quaternary. Academic Press: London.
Hamilton AC. 1987. Vegetation and climate on Mt Elgon during the latePleistocene and Holocene. Palaeoecology of Africa 18: 283–304.
Hamilton AC, Taylor D, Vogel JC. et al. 1986. Early forest clearanceand environmental degradation in south-west Uganda. Nature 320:164–167.
Hastenrath S. 2001. Variations of East African climate during the pasttwo centuries. Climatic Change 50: 209–217.
Heaton THE. 1987. The 15N/14N ratios of plants in South Africa andNamibia: relationship to climate and coastal/saline environments.Oecologia 74: 236–246.
Hunt CO. 1985. Recent advances in pollen extraction techniques:a brief review. In Palaeobiological Investigations, Fieller NRJ,Gilbertson DD, Ralph NGA (eds). Oxford; British ArchaeologicalReports International Series: 266: 181–187.
Copyright # 2014 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 29(3) 269–278 (2014)
LONG-TERM ECOSYSTEM CHANGE IN THE EASTERN ARC MOUNTAINS OF TANZANIA 277

Johnson TC, Scholz CA, Talbot MR. et al. 1996. Late Pleistocenedesiccation of Lake Victoria and rapid evolution of cichlid fishes.Science (New York NY) 273: 1091–1093.
Jolly D, Haxeltine A. 1997. Effect of low glacial atmospheric. CO2on tropical African montane vegetation. Science (New York, N.Y.)276: 786–788.
Jolly D, Taylor DM, Marchant RA, et al. 1997. Vegetation dynamicsin central Africa since 18,000 yr BP: pollen records from theinterlacustrine highlands of Burundi, Rwanda and western Uganda.Journal of Biogeography 24: 495–512.
Kiage LM, Liu K. 2006. Late Quaternary paleoenvironmental changesin East Africa: a review of multiproxy evidence from palynology,lake sediments and associated records. Progress in PhysicalGeography 30: 633–658.
Lal D, Charles C. 2007. Deconvolution of the atmospheric radiocar-bon record in the last 50,000 years. Earth and Planetary SciencesLetters 258: 550–560.
Lovett JC, Congdon TCE. 1990. Some notes on a visit to SeleguMountain, Tanzania. East Africa Natural History Society Bulletin20: 4–5.
Lovett JC, Marchant R, Taplin J, et al. 2005. The oldest rainforests inAfrica: stability or resilience for survival and diversity. In Phylogenyand Conservation, Purvis A, Gittleman JL, Brooks TM (eds).Cambridge University Press: Cambridge; 115–125.
Lovett JC, Po�cs T. 1993. Assessment of the Condition of theCatchment Forest Reserves: a Botanical Appraisal, Iringa Region.Catchment Forestry Report 93.3. Forestry and Beekeeping Division:Dar es Salaam.
MacDonald GM, Larsen CPS, Szeicz JM, et al. 1991. The reconstruc-tion of boreal forest fire history from lake sediments: a comparisonof charcoal, pollen, sedimentological and geochemical indices.Quaternary Science Reviews 10: 53–71.
Mackie EAV, Leng MJ, Lloyd JM, et al. 2005. Bulk organic d13C andC/N ratios as palaeosalinity indicators within a Scottish isolationbasin. Journal of Quaternary Science 20: 303–312.
Maitima J. 1991. Vegetation response to climate change in centralRift Valley. Quaternary Research 35: 234–245.
Marchant RA, Taylor D, Hamilton A. et al. 1997. Late Pleistoceneand Holocene history at Mubwindi Swamp, southwest Uganda.Quaternary Research 47: 316–328.
Meyers PA, Teranes JL. 2001. Sediment organic matter. In TrackingEnvironmental Change Using Lake Sediments Volume 2: Physicaland Geochemical Methods, Last WM, Smol JP (eds). KluwerAcademic Publishers: Dordrecht; 239–270.
Mittermeier CG, Lamoreux J, da Fonseca GAB. et al. 2005. HotspotsRevisited: Earth’s Biologically Richest and Most Endangered Terres-trial Ecoregions. Distributed for Conservation International.
Mumbi CT, Marchant R, Hooghiemstra H, et al. 2008. Late Quaterna-ry vegetation reconstruction from the Eastern Arc Mountains,Tanzania. Quaternary Research 69: 326–341.
Myers N. 1990. The biological challenge: extended hot-spots analy-sis. Environmentalist 10: 243–256.
Myers N, Mittermeier RA, Mittermeier CG, et al. 2000. Biodiversityhotspots for conservation priorities. Nature 403: 853–858.
Nicholson SE. 1996. A review of climate dynamics and climatevariability in Eastern Africa. In The Limnology, Climatology, andPaleoclimatology of the East African Lakes, Johnson TC, Odada EO(eds). Gordon and Breach: Amsterdam; 25–56.
Olago DO. 2001. Vegetation changes over palaeo-time scales inAfrica. Climate Research 17: 105–121.
Platts PJ, McClean CJ, Lovett JC, et al. 2008. Predicting treedistributions in an East African biodiversity hotspot: model selec-tion, data bias and envelope uncertainty. Ecological Modelling 218:121–134.
R Development Core Team. 2012. R: a Language and Environmentfor Statistical Computing. R Foundation for Statistical Computing:Vienna. Available at: www.r-project.org/.
Reimer PJ, Baillie MGL, Bard E, et al. 2009. INTCAL04 andMARINE09. Radiocarbon age calibration curves, 0-50,000 years calBP. Radiocarbon 51: 1111–1150.
Rucina SM, Muiruri VM, Kinjanjui RN, et al. 2009. Late Quaternaryvegetation and fire dynamics on Mount Kenya. Palaeogeography,Palaeoclimatology, Palaeoecology 283: 1–14.
Schuler L, Hemp A, Zech W, et al. 2012. Vegetation, climate andfire-dynamics in East Africa inferred from the Maundi crater pollenrecord from Mt Kilimanjaro during the last glacial–interglacialcycle. Quaternary Science Reviews 39: 1–13.
Sinninghe Damste JS, Verschuren D, Ossebaar J, et al. 2011. 25,000-year record of climate-induced changes in lowland vegetation ofeastern equatorial Africa revealed by the stable carbon-isotopiccomposition of fossil plant leaf waxes. Earth and Planetary SciencesLetters 302: 236–224.
Stager JC, Johnson TC. 2008. The late Pleistocene desiccation ofLake Victoria and the origin of its endemic biota. Hydrobiologia596: 5–16.
Street-Perrott FA, Barker PA, Leng MJ, et al. 2008. Towards anunderstanding of late Quaternary variations in the continentalbiogeochemical cycle of silicon: multi-isotope and sediment-fluxdata for Lake Rutundu, Mt Kenya, East Africa, since 38 ka BP.Journal of Quaternary Science 23: 375–387.
Street-Perrott FA, Barker PA, Swain DL, et al. 2007. Late Quaternarychanges in ecosystems and carbon cycling on Mt Kenya, EastAfrica: a landscape-ecological perspective based on lake-sedimentfluxes. Quaternary Science Reviews 26: 1838–1860.
Street-Perrott FA, Ficken KJ, Huang Y, et al. 2004. Late Quaternarychanges in carbon cycling on Mt Kenya, East Africa: an overviewof the d13C record in lacustrine organic matter. Quaternary ScienceReviews 23: 861–879.
Street-Perrott FA, Huang Y, Perrott RA, et al. 1997. Impact of loweratmospheric carbon dioxide on tropical mountain ecosystems.Science 278: 1422–1426.
Talbot MR, Livingstone DA. 1989. Hydrogen index and carbonisotopes of lacustrine organic matter as lake level indicators.Palaeogeography, Palaeoclimatology, Palaeoecology 80: 283–300.
Taylor DM. 1990. Late Quaternary pollen records from two Ugandanmires: evidence for environmental change in the Rukiga Highlandsof southwest Uganda. Palaeogeography, Palaeoclimatology, Palae-oecology 80: 283–300.
Terwilliger VJ, Eshetu Z, Colman A, et al. 2008. Reconstructingpalaeoenvironment from delta C-13 and delta N-15 values of soilorganic matter: a calibration from arid and wetter elevation trans-ects in Ethiopia. Geoderma 147: 197–210.
Tierney JE, Russell JM, Sinninghe Damste JS, et al. 2011. LateQuaternary behavior of the East African monsoon and the impor-tance of the Congo Air Boundary. Quaternary Science Reviews 30:798–807.
Troels-Smith J. 1955. Karakterisering af lose jordarter (Characteriza-tion of unconsolidated sediments). Danmarks Geologiske Under-soegelse IV: 1–73.
van Zinderen Bakker EM, Coetzee JA. 1988. A review of lateQuaternary pollen studies in East, Central and southern Africa.Review of Palaeobotany and Palynology 55: 155–174.
Vincens A, Buchet G, Williamson D, et al. 2005. A 23,000-yr pollenrecord from Lake Rukwa (8˚S, SW Tanzania): new data onvegetation dynamics and climate in Central Eastern Africa. Reviewof Palaeobotany and Palynology 137: 147–162.
Winkler M. 1985. Charcoal analysis for palaeoenvironmental inter-pretation. Quaternary Research 23: 313–326.
Wooller MJ, Street-Perrott FA, Agnew ADQ. et al. 2000. LateQuaternary fires and grassland palaeoecology of Mount Kenya,East Africa: evidence from charred grass cuticles in lake sediments.Palaeogeography, Palaeoclimatology, Palaeoecology 164: 207–230.
Copyright # 2014 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 29(3) 269–278 (2014)
278 JOURNAL OF QUATERNARY SCIENCE




















