
Intra-backbone and Inter-backbone Peering Among Internet Service Providers

Towards a natural classification and backbone treefor Sordariomycetes
Sajeewa S. N. Maharachchikumbura1,4& Kevin D. Hyde2,3,4,5 & E. B. Gareth Jones5 &
Eric H. C. McKenzie6 & Shi-Ke Huang4,7 & Mohamed A. Abdel-Wahab8&
Dinushani A. Daranagama4 & Monika Dayarathne4 & Melvina J. D’souza4 &
Ishani D. Goonasekara4 & Sinang Hongsanan4& Ruvishika S. Jayawardena4,14 &
Paul M. Kirk10& Sirinapa Konta4 & Jian-Kui Liu1,4
& Zuo-Yi Liu1&
Chada Norphanphoun4& Ka-Lai Pang9 & Rekhani H. Perera4 & Indunil C. Senanayake4 &
Qiuju Shang4 & Belle Damodara Shenoy13 & Yuanpin Xiao4,7 & Ali H. Bahkali5 &
Jichuan Kang7 & Sayanh Somrothipol11 & Satinee Suetrong12 & Tingchi Wen7&
Jianchu Xu2,3
Received: 11 February 2015 /Accepted: 7 April 2015# School of Science 2015
AbstractSordariomycetes is one of the largest classes of Ascomycotaand is characterised by perithecial ascomata and inoperculateunitunicate asci. The class includes many important plantpathogens, as well as endophytes, saprobes, epiphytes, andfungicolous, lichenized or lichenicolous taxa. The class in-cludes freshwater, marine and terrestrial taxa and has a
worldwide distribution. This paper provides an updated out-line of the Sordariomycetes and a backbone tree incorporatingasexual and sexual genera in the class. Based on phylogenyand morphology we introduced three subclasses;Diapor thomyce t idae , Lu lwor th iomyce t idae andMeliolomycetidae and five orders; Amplistromatales,Annulatascales , Falcocladiales , Jobellisiales and
* Zuo-Yi [email protected]
1 Guizhou Key Laboratory of Agricultural Biotechnology, GuizhouAcademy of Agricultural Sciences, Guiyang 550006, Guizhou,China
2 Key Laboratory for Plant Biodiversity and Biogeography of EastAsia (KLPB), Kunming Institute of Botany, Chinese Academy ofScience, Kunming 650201, Yunnan, China
3 World Agroforestry Centre, East and Central Asia, 132 Lanhei Road,Kunming 650201, China
4 Institute of Excellence in Fungal Research, and School of Science,Mae Fah Luang University, Chiang Rai 57100, Thailand
5 Department of Botany and Microbiology, College of Science,King Saud University, P.O. Box 2455, Riyadh 11451,Kingdom of Saudi Arabia
6 Manaaki Whenua Landcare Research, Private Bag 92170,Auckland, New Zealand
7 The Engineering and Research Center for SouthwestBio-Pharmaceutical Resources of National Education Ministry ofChina, Guizhou University, Guiyang 550025, Guizhou Province,People’s Republic of China
8 Department of Botany, Faculty of Science, Sohag University,Sohag 82524, Egypt
9 Institute of Marine Biology and Centre of Excellencefor the Oceans, National Taiwan Ocean University,2 Pei-Ning Road, Keelung 20224,Taiwan, Republic of China
10 Mycology Section, Royal Botanic Gardens, Kew,London, UK
11 Microbe Interaction Laboratory (BMIT),National Center for Genetic Engineeringand Biotechnology (BIOTEC),113 Thailand Science Park, Phahonyothin Road, Khlong Nueng,Khlong Luang, Pathum Thani, Thailand
12 Fungal Diversity Laboratory (BFBD), National Centerfor Genetic Engineering and Biotechnology (BIOTEC),113 Thailand Science Park, Phahonyothin Road,Khlong Nueng, Khlong Luang, Pathum Thani, Thailand
13 Biological Oceanography Division, CSIR-National Institute ofOceanography, Dona Paula, Goa 403004, India
14 Institute of Plant and Environment Protection,Beijing Academy of Agriculture and Forestry Sciences,No 9 of ShuGuangHuaYuanZhangLu, Haidian District,Beijing 100097, China
Fungal DiversityDOI 10.1007/s13225-015-0331-z

Togniniales. The outline is based on literature to the end of2014 and the backbone tree published in this paper. Notes for397 taxa with information, such as new family and generanovelties, novel molecular data published since the Outlineof Ascomycota 2009, and new links between sexual and asex-ual genera and thus synonymies, are provided. TheSordariomycetes now comprises six subclasses, 28 orders,90 families and 1344 genera. In addition a list of 829 generawith uncertain placement in Sordariomycetes is also provided.
Keywords Boliniales .Calosphaeriales .
Chaetosphaeriales .Coniochaetales .Coronophorales .
Diaporthales .Glomerellales .Hypocreomycetidae .
Hypocreales .Koralionastetales . Lulworthiales .
Magnaporthales .Melanosporales .Meliolales .
Microascales . Molecular identification . Nomenclature .
Ophiostomatales . Phyllachorales . Sordariales .
Sordariomycetidae . Taxonomy . Trichosphaeriales . Typespecies . Xylariales . Xylariomycetidae
Introduction
Sordariomycetes is the largest class of Ascomycota afterDothideomycetes (Kirk et al. 2008; Hyde et al. 2013). Kirket al. (2008) included 15 orders, 64 families, 1119 genera and10,564 species in Sordariomycetes, while Lumbsch andHuhndorf (2010) included 18 orders, 63 families and 947genera. The majority of the species of Sordariomycetes areterrestrial, while some can be found in aquatic habitats(Hyde and Wong 2000; Samuels and Blackwell 2001; Caiet al. 2006a; Jones et al. 2009a, b, Jones and Pang 2012;Pratibha et al. 2014). They are pathogens of plants, arthropodsand mammals (Sung et al. 2007; Prados-Rosale et al. 2012;Hyde et al. 2014) and have been isolated as endophytes fromvarious plants (Keim et al. 2014). Some taxa are fungicolous(PeiGui et al. 2000; Jaklitsch et al. 2013), while many aresaprobes involved in decomposition and nutrient cycling(Jaklitsch and Voglmayr 2012). Some important plant patho-genic genera belong to Sordariomycetes, includingColletotrichum, Diaporthe, Fusarium, Magnaporthe,Pestalotiopsis and Verticillium (Maharachchikumbura et al.2012, 2013a; Udayanga et al. 2012, 2013; Hyde et al. 2014).Also included are mushroom pathogens, such as someHypocrea and Verticillium species (PeiGui et al. 2000, Zareet al. 2007). Some species of Sordariomycetes (ie. Beauveriabassiana, Trichoderma viride, T. harzianum) are economical-ly important biocontrol agents (Wraight et al. 1998; Kaewchaiet al. 2009; Thiruvudainambi et al. 2010) and as well as im-portant in medicinal and other biotechnological industries(Semenova et al. 2012; Debbab et al. 2013; Xu et al. 2014).
Historically, some of the families of Sordariomycetes suchas Amphisphaeriaceae, Diaporthaceae, Diatrypaceae,
Halosphaeriaceae, Hypocreaceae, Polystigmataceae andSordariaceae, were classified under the order Sphaeriales,an order characterized by sphaerical or flask-shaped fruitingbodies (Müller and von Arx 1962). Chadefaud (1960) recog-nized Diaporthales as a separate order based on the character-istic stromatic tissue. Benny and Kimbrough (1980) intro-duced the orders Microascales , Onygenales andOphiostomatales. A comprehensive taxonomic study of fam-ilies and higher taxa of the Sordariomycetes based on mor-phology was that of Barr (1990). Barr (1990) included theorders Calosphaeriales, Coryneliales, Clavicipitales,Diaporthales, Erysiphales, Eurotiales, Halosphaeriales,Hypocreales, Meliolales, Microascales, Onygenales,Sordariales, Spathulosporales and Xylariales in the classnamed Hymenoascomycetes.
Eriksson and Winka (1997) introduced the classSordariomycetes based on morphological (perithecialascomata, hamathecium composed of paraphyses, osti-o l e s w i t h p e r i p h y s e s a n d u n i t u n i c a t e o rpseudoprotunicate asci) and molecular data and accom-modated three subclasses : Hypocreomycet idae ,Sordariomycetidae and Xylariomycetidae. The subclassSordar iomyce t idae inc luded three orders ( i . e .Diaporthales, Ophiostomatales and Sordariales), whileXylariomycetidae was intoduced to accommodate a sin-gle order Xylariales (Eriksson and Winka 1997).Eriksson and Winka (1997) separated Onygenales andEurotiales from Sordariomycetes and placed them inthe class Eurotiomycetes based on morphology andphylogenetic analyses. A comprehensive study ofSordariomycetes based on both morphologicalcharacters and SSU rDNA sequence data was carriedout by Samuels and Blackwell (2001) and Eriksson(2006). In the classification of Eriksson (2006), the sub-class Hypocreomycetidae comprised 4 orders (i.e.Coronophorales, Halosphaeriales, Hypocreales andMicroascales). Coronophorales, Halosphaeriales andHypocreales are recognized as monophyletic andMicroascales as paraphyletic. Samuels and Blackwell(2001) excluded Erysiphales and Coryneliales sensuBarr (1990) from Sordariomycetes, while Eriksson(2006) placed Erysiphales in the Leotiomycetes, a sistertaxon of the Sordariomycetes. Eriksson (2006) also rec-ognized Melanosporales as a distinct order in theHypocreomycetidae.
A large number of species of Sordariomycetes are charac-terized by non-lichenized, perithecial ascomata andinoperculate unitunicate asci (Zhang et al. 2006) or non-fiscitunicate asci (Kirk et al. 2008). Most members of theXylariomycetidae and some of the Sordariomycetidae havedark perithecia, amyloid asci, true paraphyses and periphysateostioles, while most of the taxa of the Hypocreomycetidaehave light coloured perithecia, nonamyloid apical rings in
Fungal Diversity

the asci when apical rings are present and the absence of trueparaphyses (Zhang et al. 2006).
The most widely used gene regions for phylogenetic stud-ies of Sordariomycetes from the early 1990s were SSUnrDNA and LSU nrDNA (Berbee and Taylor 1992;Spatafora and Blackwell 1993; Spatafora 1995). In additionto these ribosomal genes, the phylogenetic relationshipsamong Sordariomycetes were investigated using partial trans-lation elongation factor 1-alpha (TEF) and the second largestsubunit of RNA polymerase (RPB2) protein-coding genes(Zhang et al. 2006). Tang et al. (2007) evaluated the phyloge-netic utility of the ß-tubulin gene for resolving evolutionaryrelationships in Sordariomycetes.
Pleomorphism arises from the fact that many ascomycetetaxa including species of Sordariomycetes, occur in eithertheir sexual or asexual morphs alone, or in combination(Wingfield et al. 2012). Asexual and sexual morphs togethermake up a whole fungus called the holomorph (Seifert et al.2011). Some taxa also have more than one asexual morph(synanamorph) often linked to unique ecological niches(Wingfield et al. 2012). Sutton (1980) contributed to summa-rize the established links between coelomycetous asexualmorphs and their sexual morphs. Later Seifert et al. (2011)did the same for genera of hyphomycetes. The asexual andsexual morphs of Sordariomycetes often develop separately,or only one morph is formed, and it is therefore often difficultto develop links between the same species but with differentmorphs (Wijayawardene et al. 2014). This is especially truewhen one or both morphs cannot be cultured or induced toform sporulating structures on artificial media. Observation oftwo morphs on the same substrate was an earlier method usedto link morphs, but this does not prove that the morphs arerelated (Shenoy et al. 2007). DNA sequence-data can irrefut-ably link asexual and sexual morphs and provide phylogeneticplacements for asexual taxa within the sexual morph taxo-nomic classification schemes. It is difficult to place asexualgenera into current classification schemes, when asexual gen-era are in fact polyphyletic and paraphyletic. Their typespecies/ specimens need to be recollected to verify their phy-logenetic position (Schoch et al. 2009). In addition to this, itappears that even some concepts of sexual taxa will requireextensive reconsideration (Schoch et al. 2009). DNA sequenc-ing and analysis have established the natural placements ofmany sexual and asexual genera and have also proven linksbetween the sexual and asexual morphs (e.g. Trichoderma–Hypocrea,Dyrithiopsis–Monochaetiopsis,Cylindrocladiella–Nectricladiella, Fusarium–Gibberella, Colletotrichum–Glomerella; Jeewon et al. 2003a; Jaklitsch et al. 2006; Hydeet al. 2009a, b; Rossman et al. 2013), while some links havebeen not accepted (e.g. Didymostilbe–Peethambara,Discostroma–Seimatosporium, Hypomyces–Cladobotyrum;Subramanian and Bhat 1978; Tanaka et al. 2011; Rossmanet al. 2013). Decisions taken in the XVIIIth International
Botanical Congress, in Melbourne, Australia in 2011(Hawksworth 2012) ruled that separate names for asexualand sexual morphs, was no longer allowed; this became effec-tive from 30 July 2011 (Hawksworth 2012; Wingfield et al.2012). Based on this ruling, mycologists now have the task toclear up the historical confusion of dual nomenclature.
In this paper, we deal with the classification of one of thelargest groups of ascomycetes, the class Sordariomycetes.Some asexual genera in this class have been linked to sexualmorphs and some are discussed in Rossman et al. (2013), andthese are followed in this paper. Several groups have not beendealt with as yet and therefore we generally follow the oldername in this paper. A list of Baccepted/ protected names^,Bsuppressed names^ as published for the Dothideomcyetesby Wijayawardene et al. (2014) is still required. The outlinehere uses Lumbsch and Huhndorf (2010) as a starting point.We have then scanned the literature and provide datamostly tothe end of 2014. A note is provided for newly introducedgenera and families and where important molecular data isrecently provided we also provide notes. The outline also fol-lows the results of the phylogenetic data generated in thisstudy. A list of genera presently placed in Sordariomycetesin Index Fungorum (2015) is also included. These were notlisted in Lumbsch and Huhndorf (2010), but most were listedin Kirk et al. (2013). Their status and placement must still beestablished. There are also numerous synonyms that weresynonymised under various genera, possibly without muchthought and it would be interesting to establish if these arealso distinct genera.
Materials and methods
Phylogenetic analyses
Sequences were obtained from GenBank mostly followingprevious literature (Castlebury et al. 2002; Réblová 2003,2008; 2011a, b, Réblová 2013a, b; García et al. 2006;Boonyuen et al. 2011; Mugambi and Huhndorf 2010;Johnson et al. 2009; Campbell et al. 2005, 2009;Thongkantha et al. 2009; Sung et al. 2007) and are listed inTable 1.
This study used LSU, SSU, TEF and RPB2 sequence datain the analyses. Multiple sequence alignments were generatedwith MAFFT v. 7 (http://mafft.cbrc.jp/alignment/server/index.html); then manually corrected to ensure alignmentand to minimize the number of uninformative gaps usingMEGA v. 6.06 (Kumar et al. 2012). The datasets wereproduced to show families and order relationships within theclass Sordariomycetes.
The combined alignments were split between the genera toimprove the robustness of the alignment across the four loci.Phylogenies used Bayesian Inference (BI) and Maximum
Fungal Diversity

Table 1 GenBank accession numbers of isolates includes in this study
Species Culture accession no. GenBank accession
LSU SSU TEF RPB2
Acanthonitschkea argentinensis SMH1395 AY695259 – FJ969042 FJ968943
Achaetomium strumarium IMI 082624 AJ312098 – – –
Albertiniella polyporicola CBS 457.88 AF096185 AF096170 – –
Ambrosiella ferruginea CBS 460.82 EU825651 – – –
Amphisphaeria umbrina AFTOL-ID 1229 FJ176863 FJ176809 FJ238394 FJ238348
Amplistroma caroliniana BEO9923 FJ532377 – – –
Amplistroma erinaceum AH 43902 KC907374 – KC907375 –
Anisogramma anomala AA01 EU683066 – – –
Anisogramma virgultorum AV05 EU683065 – – –
Annulatascus velatisporus HKUCC 3701 AF132320 – – –
Annulusmagnus triseptatus CBS 128831 GQ996540 JQ429242 – –
Apiorhynchostoma curreyi UAMH 11088 JX460989 – – –
Apiosordaria verruculosa F-152365 AY346258 – – AY780150
Apiospora bambusae ICMP 6889 DQ368630 DQ368662 – DQ368649
Apiospora montagnei AFTOL-ID 951 DQ471018 FJ190614 DQ842027 DQ842033
Arecophila bambusae HKUCC 4794 AF452038 AY083802 – –
Arthrinium hydei CBS 114990 KF144936 – KF145024 –
Arthrinium phaeospermum HKUCC 3395 AY083832 AY083816 – –
Ascitendus austriacus MR 2936 GQ996539 GQ996542 – –
Ascopolyporus polychrous PC546 AY886546 – DQ118745 DQ127236
Ascotaiwania lignicola NIL00005 HQ446364 HQ446284 HQ446307 HQ446419
Ascothailandia grenadoidia NB-2010a = SS03615 GQ390267 GQ390252 HQ446309 HQ446420
Ascovaginospora stellipala P5-13A U85088 U85087 – –
Asteridiella obesa VIC 31239 JX096809 – – –
Astrocystis cocoes HKUCC 3441 AY083823 – – –
Australiasca queenslandica BRIP 24607 HM237324 – – –
Bambusicularia brunnea INA-B-92-45 KM484948 – – –
Beltraniella endiandrae CPC 22193 KJ869185 – – –
Bertia moriformis SMH4320 AY695260 – – AY780151
Bertia ngongensis GKM1239 FJ968954 – – –
Bertia tropicalis SMH3513 AY695263 – – –
Bionectria ochroleuca AFTOL-ID 187 DQ862027 DQ862044 DQ862029 DQ862013
Bombardia bombarda AFTOL-ID 967 DQ470970 DQ471021 – DQ470923
Botryotinia fuckeliana AFTOL-ID 59 AY544651 AY544695 DQ471045 DQ247786
Buergenerula spartinae ATCC 22848 DQ341492 DQ341471 JX134692 –
Bussabanomyces longisporus CBS 125232 KM009154 – – –
Cainia graminis CBS 136.62 AF431949 – – –
Cainiella johansonii Kruys 727 (UPS) JF701920 – – –
Camaropella pugillus SMH3846 EU481406 – – –
Camarops ustulinoides AFTOL-ID 72 DQ470941 DQ470989 DQ471050 –
Canalisporium caribense SS03839 GQ390268 GQ390253 – HQ446421
Carpoligna pleurothecii CBS 114211 JQ429235 JQ429249 – JQ429267
Catabotrys deciduum SMH3436 AY346268 – – AY780158
Cephalotheca foveolata UAMH11631 KC40839 – KC408410 KC408404
Ceratocystiopsis minuta CBS 116963 EU913655 – – –
Ceratocystis fimbriata C89 CFU17401 CFU32418 – –
Chaetomidium galaicum CBS 113678 FJ666361 – – FJ666392
Fungal Diversity

Table 1 (continued)
Species Culture accession no. GenBank accession
LSU SSU TEF RPB2
Chaetosphaerella fusca GKML124N FJ968967 – FJ969002 –
Chaetosphaerella phaeostroma SMH4257 AY695264 – FJ969004 FJ968940
Chaetosphaeria innumera SMH 2748 AY017375 – – –
Chromendothia citrina AFTOL-ID 2121 – DQ862046 DQ862031 DQ862015
Claviceps purpurea AEG 97-2 AF543789 AF543765 AF543778 –
Clypeosphaeria uniseptata 6349 (HKUCC) DQ810219 DQ810255 – –
Coccodiella melastomatum CMU78543 – U78543 – –
Coccodiella miconiae ppMP 1342 – GU233440 – –
Colletotrichum asianum LC0037 JN940408 JN940347 – –
Colletotrichum gloeosporioides LC0555 JN940412 JN940356 – –
Colletotrichum fructicola LC0032 JN940418 JN940350 – –
Colletotrichum brevisporum LC0600 JN940398 JN940357 – –
Coniocessia anandra Iran 1468C GU553349 – – –
Coniocessia maxima CBS 593.74 GU553344 – – –
Coniocessia nodulisporioides CBS 281.77 GU553352 AJ875185 – –
Coniochaeta ligniaria C8 AY198388 – – –
Conioscypha japonica CBS 387.84 AY484514 JQ437438 – JQ429259
Conioscyphascus varius CBS 113653 AY484512 – – –
Cordana pauciseptata CBS 121804 HE672160 – – –
Cordyceps militaris OSC 93623 AY184966 AY184977 DQ522332 –
Coronophora gregaria ANM1555 – – FJ969007 FJ968938
Cornipulvina ellipsoides SMH1378 DQ231441 – – –
Corynascella inaequalis CBS 284.82 – – HQ871746 HQ871839
Cosmospora coccinea AR2741 AY489734 AY489702 AY489629 –
Crassochaeta nigrita SMH2931 AY695266 – – –
Creosphaeria sassafras CM AT-018 DQ840056 – – DQ631964
Cryptendoxyla hypophloia WM10.89 HQ014708 – – –
Cryphonectria parasitica CMW 7048 JN940858 JN938760 – –
Cryptadelphia groenendalensis SH12 EU528007 – – –
Cryptadelphia groenendalensis SMH3767 EU528001 – – –
Cryptodiaporthe aesculi AFTOL-ID 1238 DQ836905 – DQ836914 DQ836892
Cryptometrion aestuescens CMW18790 HQ730869 – – –
Cryptosphaerella elliptica SMH4722 FJ968974 – FJ969029 FJ968944
Cryptosporella hypodermia AFTOL-ID 2124 DQ862028 DQ862049 DQ862034 DQ862018
Cumulospora marina MF46 GU252135 GU252136 – –
Custingophora cecropiae CCF 3568 – AM267267 – –
Cytospora elaeagni CFCC 89633 KF765693 – – KF765709
Diaporthales sp. BCC00200 EF622231 EF622228 – –
Diaporthe eres AFTOL-ID 935 – DQ471015 – DQ471161
Diatrype disciformis AFTOL-ID 927 DQ470964 DQ471012 DQ471085 –
Discostroma botan – DQ368629 DQ368660 – DQ368648
Doratomyces stemonitis AFTOL-ID 1380 DQ836907 DQ836901 DQ836916 –
Dothidea sambuci DAOM 231303 AY544681 AY544722 DQ497606 DQ522854
Echinosphaeria canescens SMH4791 AY436403 – – –
Elaphocordyceps capitata OSC 71233 AY489721 AY489689 AY489615 DQ522421
Elaphocordyceps japonica OSC 110991 DQ518761 DQ522547 DQ522330 DQ522428
Endomeliola dingleyae PDD:98304 GU138866 – – –
Fungal Diversity
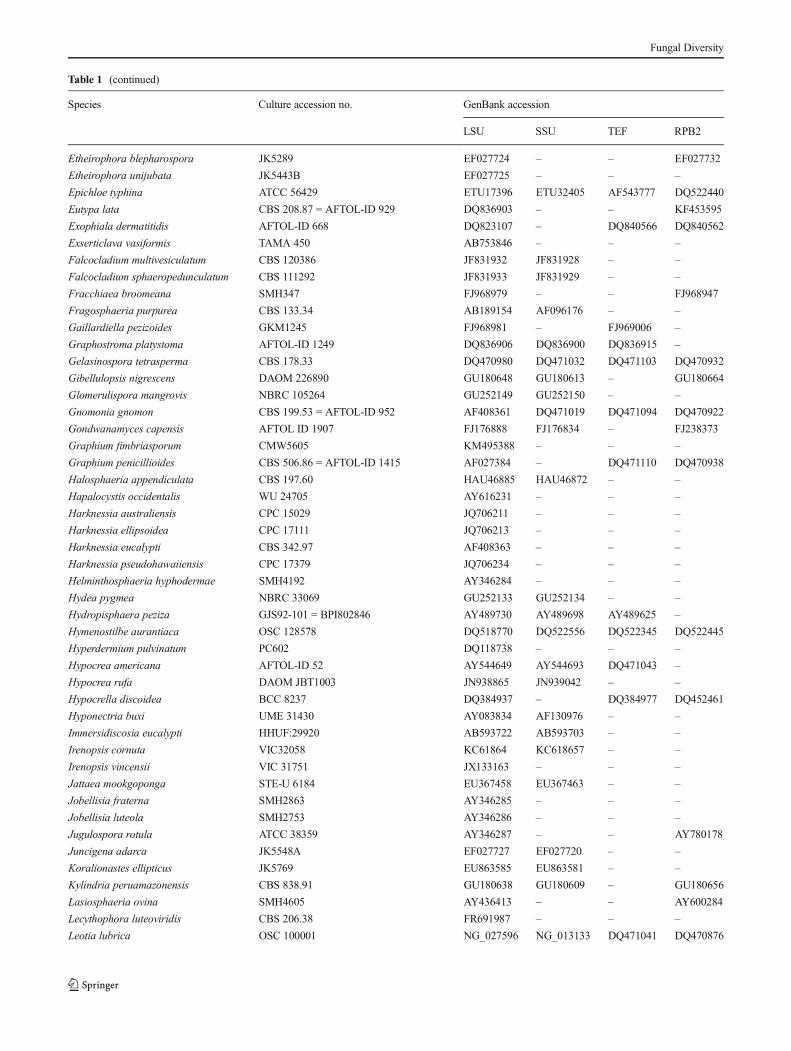
Table 1 (continued)
Species Culture accession no. GenBank accession
LSU SSU TEF RPB2
Etheirophora blepharospora JK5289 EF027724 – – EF027732
Etheirophora unijubata JK5443B EF027725 – – –
Epichloe typhina ATCC 56429 ETU17396 ETU32405 AF543777 DQ522440
Eutypa lata CBS 208.87 = AFTOL-ID 929 DQ836903 – – KF453595
Exophiala dermatitidis AFTOL-ID 668 DQ823107 – DQ840566 DQ840562
Exserticlava vasiformis TAMA 450 AB753846 – – –
Falcocladium multivesiculatum CBS 120386 JF831932 JF831928 – –
Falcocladium sphaeropedunculatum CBS 111292 JF831933 JF831929 – –
Fracchiaea broomeana SMH347 FJ968979 – – FJ968947
Fragosphaeria purpurea CBS 133.34 AB189154 AF096176 – –
Gaillardiella pezizoides GKM1245 FJ968981 – FJ969006 –
Graphostroma platystoma AFTOL-ID 1249 DQ836906 DQ836900 DQ836915 –
Gelasinospora tetrasperma CBS 178.33 DQ470980 DQ471032 DQ471103 DQ470932
Gibellulopsis nigrescens DAOM 226890 GU180648 GU180613 – GU180664
Glomerulispora mangrovis NBRC 105264 GU252149 GU252150 – –
Gnomonia gnomon CBS 199.53 = AFTOL-ID 952 AF408361 DQ471019 DQ471094 DQ470922
Gondwanamyces capensis AFTOL ID 1907 FJ176888 FJ176834 – FJ238373
Graphium fimbriasporum CMW5605 KM495388 – – –
Graphium penicillioides CBS 506.86 = AFTOL-ID 1415 AF027384 – DQ471110 DQ470938
Halosphaeria appendiculata CBS 197.60 HAU46885 HAU46872 – –
Hapalocystis occidentalis WU 24705 AY616231 – – –
Harknessia australiensis CPC 15029 JQ706211 – – –
Harknessia ellipsoidea CPC 17111 JQ706213 – – –
Harknessia eucalypti CBS 342.97 AF408363 – – –
Harknessia pseudohawaiiensis CPC 17379 JQ706234 – – –
Helminthosphaeria hyphodermae SMH4192 AY346284 – – –
Hydea pygmea NBRC 33069 GU252133 GU252134 – –
Hydropisphaera peziza GJS92-101 = BPI802846 AY489730 AY489698 AY489625 –
Hymenostilbe aurantiaca OSC 128578 DQ518770 DQ522556 DQ522345 DQ522445
Hyperdermium pulvinatum PC602 DQ118738 – – –
Hypocrea americana AFTOL-ID 52 AY544649 AY544693 DQ471043 –
Hypocrea rufa DAOM JBT1003 JN938865 JN939042 – –
Hypocrella discoidea BCC 8237 DQ384937 – DQ384977 DQ452461
Hyponectria buxi UME 31430 AY083834 AF130976 – –
Immersidiscosia eucalypti HHUF:29920 AB593722 AB593703 – –
Irenopsis cornuta VIC32058 KC61864 KC618657 – –
Irenopsis vincensii VIC 31751 JX133163 – – –
Jattaea mookgoponga STE-U 6184 EU367458 EU367463 – –
Jobellisia fraterna SMH2863 AY346285 – – –
Jobellisia luteola SMH2753 AY346286 – – –
Jugulospora rotula ATCC 38359 AY346287 – – AY780178
Juncigena adarca JK5548A EF027727 EF027720 – –
Koralionastes ellipticus JK5769 EU863585 EU863581 – –
Kylindria peruamazonensis CBS 838.91 GU180638 GU180609 – GU180656
Lasiosphaeria ovina SMH4605 AY436413 – – AY600284
Lecythophora luteoviridis CBS 206.38 FR691987 – – –
Leotia lubrica OSC 100001 NG_027596 NG_013133 DQ471041 DQ470876
Fungal Diversity

Table 1 (continued)
Species Culture accession no. GenBank accession
LSU SSU TEF RPB2
Lepteutypa cupressi IMI 052255 AF382379 AY083813 – –
Leucostoma niveum AR3413 = BPI 748232 AF362558 NG_013203 EU222015 EU219343
Lignincola laevis JK-5180A U46890 U46873 – –
Lindra thalassiae AFTOL-ID 413 DQ470947 DQ470994 DQ471065 DQ470897
Lopadostoma turgidum LT3 KC774618 – – –
Lulworthia fucicola ATCC 64288 AY878965 AY879007 – –
Luteocirrhus shearii CBS 130776 KC197019 – – –
Magnaporthe salvinii M21 JF414887 – JF710406 –
Marinokulati chaetosa BCRC FU30271 KJ866931 KJ866929 – –
Mazzantia napelli BPI748443 = AR3498 = AFTOL-ID 2126 AF408368 DQ862051 EU222017 EU219345
Melanconis marginalis BPI748446 = AR3442 = AFTOL-ID 2128 AF408373 DQ862053 EU221991 EU219301
Melanconis stilbostoma BPI748447 = AR3501 = CBS 109778 AF408374 NG_013198 EU221886 EU219299
Melanochaeta hemipsila SMH2125 AY346292 – – –
Melanospora tiffanii ATCC15515 AY015630 AY015619 – AY015637
Melanospora zamiae ATCC 12340 AY046579 AY046578 –
Meliola centellae VIC:31244 JQ734545 – – –
Microascus longirostris CBS 267.49 = AFTOL-ID 1237 DQ471172 DQ471026 DQ836913 –
Moheitospora fruticosae EF14 GU252145 GU252146 – –
Monilochaetes infuscans CBS 869.96 GU180639 GU180620 – GU180657
Monosporascus cannonballus FMR6682 – AF340016 – –
Myrothecium roridum ATCC 16297 AY489708 AY489676 AY489603 –
Nectria cinnabarina CBS 114055 = GJS 89-107 NCU00748 NCU32412 AF543785 –
Neofracchiaea callista SMH2689 AY695269 – FJ969020 FJ968941
Neonectria ramulariae CBS 151.29 HM042436 HQ840414 – DQ789792
Neurospora crassa MUCL 19026 AF286411 X04971 – –
Nimbospora effusa JK 5104A NEU46892 NEU46877 – –
Niesslia exilis CBS 560.74 AY489720 AY489688 AY489614 –
Nitschkia tetraspora GKML148N FJ968987 – FJ969011 FJ968936
Ophioceras commune M92 JX134688 JX134662 – –
Ophioceras dolichostomum CBS 114926 JX134689 JX134663 JX134703 –
Ophioceras leptosporum CBS 894.70 JX134690 – – –
Ophiocordyceps sinensis YN09-64 JX968033 JX968028 – JX968013
Ophiodiaporthe cyatheae YMJ 1364 JX570891 JX570890 KC465406 JX570893
Ophiostoma piliferum AFTOL-ID 910 DQ470955 DQ471003 DQ471074 –
Parasarcopodium ceratocaryi CBS 110664 AY425026 – – –
Papulosa amerospora AFTOL-ID 748 DQ470950 DQ470998 DQ471069 –
Peethambara spirostriata CBS 110115 AY489724 AY489692 AY489619 –
Petriella setifera AFTOL-ID 956 DQ470969 DQ471020 DQ836911 –
Phaeoacremonium pallidum STE U 6104 – EU128061 – –
Phialemonium atrogriseum CBS 604.67 HE610470 – – –
Phyllachora graminis UME 31349 – AF064051 – –
Pilidiella diplodiella STE U 3708 AY339284 – – –
Pilidiella wangiensis CPC 19397 JX069857 – – –
Plagiostoma euphorbiae CBS 340.78 = AFTOL-ID 2130 AF408382 DQ862055 GU354016 EU219292
Plectosphaerella cucumerina DAOM 226828 GU180647 GU180612 – GU180663
Pleurostoma ootheca CMU 23858 = CBS 115329 AY761079 AY761074 – HQ878606
Pleurostomophora repens CBS 294.39 AY729813 – – –
Fungal Diversity

Table 1 (continued)
Species Culture accession no. GenBank accession
LSU SSU TEF RPB2
Pleurostomophora richardsiae CBS 270.33 AY249089 – – HQ878607
Conioscyphascus CBS 131271 JQ429240 – – JQ429270
Porosphaerellopsis sporoschismophora ATCC 42528 AY346298 – – –
Pseudohalonectria lignicola M95 JX134691 JX134665 JX134705 –
Pseudomassaria carolinensis 9502 (IFO) DQ810233 DQ810262 – DQ810239
Pseudoneurospora amorphoporcata CBS 626.80 FR774287 – – –
Pseudoplagiostoma eucalypti CBS 124807 GU973606 – – –
Pseudoplagiostoma oldii CBS 124808 GU973609 – – –
Pseudoplagiostoma variabile CBS 113067 GU973611 – – –
Pseudoproboscispora caudae A336-2D AY094192 – – –
Pseudopyricularia kyllingae HYKB202-1-2 KM484992 – – –
Pseudovalsa longipes AR 3541 EU683072 – – –
Pseudovalsa modonia AR 3558 EU683073 – – –
Pyricularia borealis CBS 461.65 DQ341511 – – –
Raffaelea canadensis C2233 = CBS 168.66 EU177458 EU170270 – –
Remispora maritima JPHY-15-038 HQ111012 HQ111002 – HQ111041
Reticulascus clavatus CBS 125296 GU180643 GU180622 – –
Reticulascus clavatus CBS 125239 – GU180615 – –
Rhopalostroma lekae MFLUCC 13-0123 KJ472427 – – KJ472429
Rimaconus coronatus SMH 5212 HM171292 – – –
Rimaconus jamaicensis SMH 4782 HM171293 – – –
Robillarda sessilis BCC13393 FJ825378 FJ825368 – –
Rosellinia necatrix HKUCC 9037 AY083824 – – –
Rossmania ukurunduensis AR 3484 EU683075 – – –
Roumegueriella rufula GJS 91-164 EF469082 EF469129 EF469070 EF469116
Ruzenia spermoides SMH4655 KF765619 – – –
Savoryella lignicola NF00204 HQ446378 HQ446300 HQ446334 –
Schizoparme straminea CBS 149.22 AF362569 – – –
Schizoparme straminea STE-U 3932 AY339296 – – –
Scopinella solani CBS 770.84 AY015632 AY015621 – –
Scortechinia acanthostroma SMH1143 FJ968988 FJ969012 – FJ968948
Scortechiniellopsis leonensis GKM1269 FJ968993 – FJ969021 FJ968933
Seynesia erumpensnew SMH 1291 AF279410 AF279409 – –
Shimizuomyces paradoxus EFCC 6279 EF469084 EF469131 – EF469117
Slopeiomyces cylindrosporus CBS 609.75 KM485040 – – –
Sordaria fimicola CBS 508.50 AY681160 – – –
Sphaerostilbella berkeleyanan CBS 102308 = GJS 82-274 U00756 AF543770 AF543783 DQ522465
Spinulosphaeria nuda SMH1952 FJ968995 – – –
Stachybotrys chlorohalonata UAMH6417 AY489712 AY489680 AY489607 –
Stachylidium bicolor DAOM 226658 GU180651 GU180616 – –
Stegonsporium protopyriforme D30 = CBS 117041 EU039991 – EU040017 –
Stegonsporium pyriforme D2 = CBS 117023 EU039987 – EU040001 –
Stephanonectria keithii GJS92-133 AY489727 AY489695 AY489622 –
Stilbospora macrosperma CBS 121883 JX517299 – – –
Stilbospora longicornuta CBS 122529 KF570164 – KF570232 KF570194
Swampomyces armeniacus JK5325A EF027729 – – –
Swampomyces triseptatus CY2802 AY858953 AY858942 – –
Fungal Diversity
Likelihood (ML) analyses of both the individual data parti-tions as well as the combined aligned dataset. Ambiguouslyaligned regions were excluded from all analyses and gapswere treated as Bmissing data^ in the parsimony analysis.Suitable models for the Bayesian analysis were first selectedusing models of nucleotide substitution for each gene, as de-termined usingMrModeltest v. 2.2 (Nylander et al. 2004), andincluded for each gene partition. The GTR+I+G model withinverse gamma rate were selected for LSU, TEF and RPB2and SYM+I+G model with inverse gamma rate were selectedfor LSU and included for each gene partition. The Bayesiananalyses (MrBayes v. 3.2.1; Ronquist et al. 2012) of foursimultaneous Markov Chain Monte Carlo (MCMC) chainswere run from random trees for 100,000,000 generationsand sampled every 1000 generations. The temperature valuewas lowered to 0.15, burn-in was set to 0.25, and the run wasautomatically stopped as soon as the average standard devia-tion of split frequencies reached below 0.01. A Maximum
Likelihood analysis was performed using raxmlGUI v. 1.3(Silvestro and Michalak 2011). The optimal ML tree searchwas conducted with 100 separate runs, using the default algo-rithm of the program from a random starting tree for each run.The final tree was selected among suboptimal trees from eachrun by comparing likelihood scores under the GTR+GAMMA substitution model. The MP analysis was per-formed with MEGA v. 6.06 using a heuristic search option.Bootstrap analyses were performed using 1000 replications.The resulting trees were printed with FigTree v. 1.4.0 (http://tree.bio.ed.ac.uk software/figtree/) and the layout was donewith Adobe Illustrator CS v. 6.
Results and discussion
The combined LSU, SSU, TEF, and RPB2 gene data setcomprsied 260 taxa, with Botryotinia fuckeliana, Dothidea
Table 1 (continued)
Species Culture accession no. GenBank accession
LSU SSU TEF RPB2
Sydowiella stellatifolii CBS 119342 EU552156 – – –
Synaptospora plumbea SMH3962 KF765621 – – –
Thailandiomyces bisetulosus BCC00018 EF622230 EF622229 – –
Thielaviopsis thielavioides CBS 130.39 AF222480 – – –
Thyridium sp. Mas 56 – HQ993268 – –
Thyridium vestitum AFTOL ID 172 AY544671 AY544715 DQ471058 DQ470890
Tirisporella beccariana BCC36737 JQ655450 JQ655454 – –
Togninia africana STE U 6177 – EU128060 – –
Togninia griseo olivacea STE U 5966 – EU128058 – –
Togniniella acerosa PDD 81432 AY761076 AY761073 – –
Torpedospora radiata AFTOL-ID 751 DQ470951 DQ470999 DQ471070 DQ470902
Torpedospora radiata PP7763 AY858947 AY858939 – –
Torrubiella wallacei CBS 101237 AY184967 AY184978 EF469073 EF469119
Trichoderma viride GJS89-127 AY489726 – – –
Umbrinosphaeria caesariata CBS 102664 AF261069 – – –
Utrechtiana cibiessia CPC 18916 JF951176 – – –
Valsa ambiens AR3516 = CBS 109491 EU255208 DQ862056 EU222019 EU219347
Valsella salicis AR 3514 EU255210 – EU222018 –
Verticillium dahliae ATCC 16535 AY489737 AY489705 AY489632 DQ522468
Vialaea mangifia MFLUCC 12-0808 KF724975 – – –
Vialaea minutella BRIP 56959 KC181924 – – –
Viridispora diparietispora ATCC MYA 627 AY489735 AY489703 AY489630 –
Vittatispora coorgii BICC 7817 DQ017375 – – –
Wallrothiella congregata SMH1760 FJ532375 – – –
Xylaria hypoxylon CBS 122620 KM186301 – – –
a Note provided in this paperbMolecular data used in this studycNo sequence data in GenBank as at date on submission
Fungal Diversity

sambuci, Exophiala dermatitidis and Leotia lubrica as theoutgroup taxa. The combined dataset comprised 4015 charac-ters including gaps. The best scoring RAxML tress are shownin Figs. 1 and 2. The Maximum Parsimony resulted in a treewith the same topology and clades as the Bayesian analysisresulted tree. Bootstrap support (BS) values of MP and ML(equal to or above 50 %) are shown on the upper branches.Values of Bayesian posterior probabilities (equal to or above90 %) from MCMC analyses are shown under the branches.
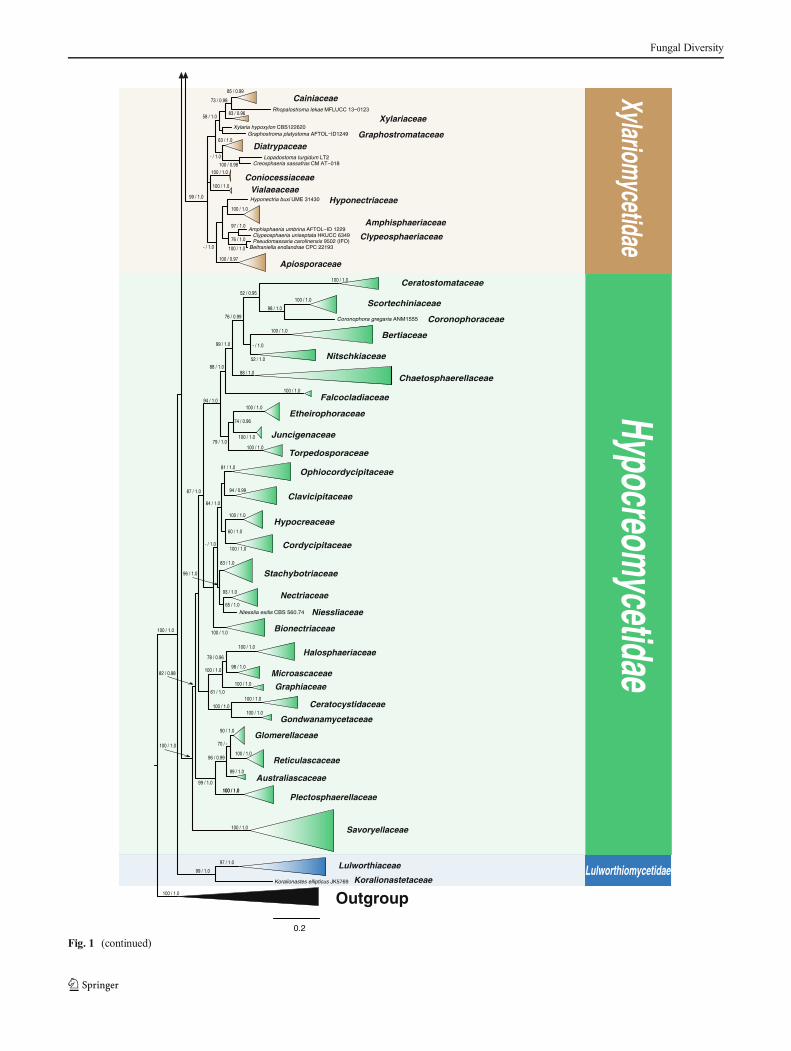
In the phylogenetic trees (Figs. 1 and 2), the 260 strainsof Sordariomycetes included in the analysis cluster into sixsubclasses. Sordariomycetidae, Hypocreomycetidae andXylariomycetidae as in the previous treatment of Lumbschand Huhndorf (2010), Meliolomycetidae as suggested byKirk et al. (2001), while Diaporthomycetidae andLulworthiomycetidae are introduced as new subclasses inthis paper. Figure 1 is a reduced tree of Fig. 2. The arrange-ment of orders and genera within the subclasses can be seenin the outline.
Subclasses of Sordariomycetes
Diaporthomycetidae Senan., Maharachch. & K.D. Hyde,subclass novus
Index Fungorum no: IF 551051; Facesoffungi number:FoF 00594
Pathogenic, parasitic, endophytic, or saprobic, associatedwith plants or pathogens of insects, animals or humans.Sexual morph: Stromata immersed to erumpent, composedof orange, brown or black parenchymatous tissues, ascomataembedded in stromatic tissues. Ascomata solitary or aggregat-ed, immersed, erumpent to superficial, globose to subglobose,rarely pyriform, sometimes carbonaceous, brown to black.Asci 8 or multi-spored, unitunicate, cylindrical, clavate to ob-long, with a nonamyloid refractive apical apparatus.Ascospores 2–3-seriate, variously shaped, aseptate or septate,sometimes with appendages and sheaths. Asexual morph:ceolomycetous or hypomyceteous, when coelomycetous,Conidiomata acervuli or pycnidial. Conidiogenous cellsphialidic or annellidic. Conidia small to large, unicellular tomultiseptate, hyaline, light brown to dark brown. whenhypomyceteous, Conidiophores arising from hyphae.Conidiogenous cells terminal or lateral, sometimes coloured,mostly hyaline. Conidia mostly unicellular, oval, hyaline.
Type order: Diaporthales Nannf.The subclass Diaporthomycetidae comprises ten main
clades which are represented by six existing orders, four new-ly introduced orders and 26 families. The order Diaporthalescomprises 12 clades represented by Cryphonectriaceae,Diaporthaceae , Gnonomiaceae, Harknessiaceae ,Me lan con i da c ea e , Pseudop l a g i o s t oma t a c ea e ,Pseudovalsaceae, Schizoparmeaceae, Stilbosporaceae,Sydowiellaceae , Tirisporellaceae and Valsaceae.
Calosphaeriales comprises the families Calosphaeriaceaeand Pleurostromataceae. The new orders Jobellisiales andTogniniales are introduced for Jobellisiaceae andTogniniaceae, respectively. Magnaporthales is expanded toincorporate Magnaporthaceae, Ophioceraceae andPyriculariaceae.
Type species: Diaporthe eres NitschkeHypocreomycetidae O.E. Erikss. & Winka, Myconet 1(1):
6 (1997)This subclass was introduced by Eriksson and Winka
(1997) and comprises four existing orders, one new orderintroduced in this paper and 28 families. The orderCoronophorales now includes Coronophoraceae ,C e r a t o s t o m a t a c e a e , C h a e t o s p h a e r e l l a c e a eScortechiniaceae, plus Nitschkiaceae and Bertiaceae. TheMelanosporales comprises the family Ceratostomataceae.The order Microascales is enlarged to include the familiesHalosphaeriaceae, Microascaceae, Gondwanamycetaceae,Ceratocystidaceae and Graphiaceae, but with weak support.The order Glomerellales includes Australiascaceae,Glomerellaceae, Plectosphaerellaceae and Reticulascaceae.The order Hypocreales incorporates Bionectriaceae,Clavicipitaceae , Cordycipitaceae , Hypocreaceae ,Nectriaceae, Niessliaceae, Ophiocordyciptiaceae andStachybotriaceae. The new order Falcocladiales is introduceda n d c omp r i s e s o n e f am i l y Fa l c o c l a d i a c e a e .Torpedosporaceae, Juncigenaceae and Etheirophoraceaeare placed in the subclassHypocreomycetidae family incertaesedis. The recently introduced monotypic order Savoryellalesform a basal clade in the Hypocreomycetidae.
Lulworthiomycetidae Dayar., E.B.G. Jones & K.D. Hyde,subclass novus
Index Fungorum number: IF551131; Facesoffungi No.:FoF 00617
Saprobic on wood, sea grasses and marsh plants or para-sites on algae. Sexual morph: Ascomata subglobose to cylin-drical or ovoid to ellipsoidal, immersed or superficial,ostiolate, papillate or epapillate, periphysate. Hamatheciumcomposed of paraphyses or internal structures absent; centruminitially filled with a hyaline pseudoparenchyma, dissolving atmaturity. Asci eight-spored, cylindrical to fusiform or clavateto ellipsoidal, unitunicate, deliquescent. Ascospores filamen-tous or ellipsoidal to fusiform, septate; multiseptate near theapices, or evenly multiseptate, some are non-septate, hyaline,thick- or thin-walled, with or without apical chambers.Asexual morph: hypomycetous, hyphae hyaline, septatae,
�Fig. 1 The best scoring RAxML Sordariomycetes tree (compressedoverview tree) from 260 taxa based on a combined dataset of LSU,SSU, TEF and RPB2 genes with all lineages collapsed to family levelwhere possible. Subclasses are indicated in coloured blocks and RAxMLbootstrap support values (MLB) and Bayesian posterior probabilities (PP)are given at the nodes (MLB/PP). The tree is rooted with Botryotiniafuckeliana,Dothidea sambuci, Exophiala dermatitidis and Leotia lubrica
Fungal Diversity

Cryptosporella hypodermia AFTOL-ID 2124
Papulosa amerospora AFTOL-ID 748
Harknessia australiensis CPC 15029
Catabotrys deciduum SMH3436
Cordana pauciseptata CBS 121804
Harknessia ellipsoidea CPC 17111Harknessia pseudohawaiiensis CPC 17379
Harknessia eucalypti CBS 342.97
Apiosordaria verruculosa F-152365
100 / 1.0
100 / 1.0
100 / 1.0
78 / 1.0
88 / 1.0
60 / 0.96
84 / -
80 / -
87 / -
100 / 1.0
100 / 1.0
100 / 1.0
100 / 1.0
89 / 1.0
94 / 1.00
75 / 1.0
58 / 0.97
84 / 1.0
53 / -
- / 1.0
100 / 1.0
100 / 1.0
100 / 1.0
97 / 1.0
90 / 1.0
52 / 0.9176 / -
99 / 0.98
99 / 0.98
84 / 1.0
89 / 0.91
76 / 0.95
87 / 1.0
69 / 0.97
70 / 0.95
90 / 1.0
100 / 1.0
83 / 1.0
100 / 1.0
100 / 1.0
60 / -
73 / 1.0
62 / 0.93
93 / 0.98
100 / 1.091 / 1.0
88 / 1.0
83 / 1.0
98 / 1.0
83 / -
73 / 0.99
100 / 1.0
76 / -
100 / 1.0
85 /-
100 / 1.0
66 / -
100 / 1.0
62 / 0.95
92 / 1.0
100 / 1.0
100 / 1.0
94 / 1.0
100 / 1.0
Schizoparmeaceae
Cryphonectriaceae
Harknessiaceae
Gnomoniaceae
Melanconidaceae
Valsaceae
Diaporthaceae
Stilbosporaceae
Pseudoplagiostomataceae
Sydowiellaceae
Tirisporellaceae
Pseudovalsaceae
Togniniaceae
Jobellisiaceae
Calosphaeriaceae
Pleurostomataceae
Pyriculariaceae
Magnaporthaceae
Ophioceraceae
Ophiostomataceae
Annulatascaceae
ConiochaetaceaeCordanaceae
TrichosphaeriaceaeThyridiaceae
Papulosaceae
Chaetomiaceae
Lasiosphaeriaceae
Sordariaceae
Boliniaceae
Phyllachoraceae
Chaetosphaeriaceae
Helminthosphaeriaceae
Cephalothecaceae
Meliolaceae
Amplistromataceae
Rimaconus
Catabotrydaceae
Diaporthomycetidae Sordariomycetidae
Meliolomycetidae
Anisogramma
50 / -
Fungal Diversity
0.2
Beltraniella endiandrae CPC 22193
Clypeosphaeria uniseptata HKUCC 6349
Niesslia exilis CBS 560.74
Koralionastes ellipticus JK5769
Rhopalostroma lekae
Xylaria hypoxylon CBS122620
Pseudomassaria carolinensis 9502 (IFO)
Graphostroma platystoma AFTO ID1249
Hyponectria buxi UME 31430
Amphisphaeria umbrina AFTO ID 1229
Lopadostoma turgidum LT2Creosphaeria sassafras CM 018
Coronophora gregaria ANM1555
100 / 0.97
64 / 1.0
100 / 1.0
98 / 1.0
70 / -
94 / 1.0
100 / 1.0100 / 1.0
85 / 0.99
100 / 1.0
76 / 1.0
100 / 1.0
63 / 1.0
100 / 0.98
96 / 0.99
96 / 1.0
100 / 1.0
76 / 0.99
61 / 1.0
100 / 1.0
100 / 1.0
100 / 1.0
100 / 1.0
97 / 1.0
100 / 1.0
100 / 1.0
87 / 1.0
100 / 1.0
99 / 1.0
82 / 0.98
88 / 1.0
90 / 1.0
74 / 0.96
88 / 1.0
100 / 1.0
100 / 1.0
100 / 1.0
99 / 1.0
73 / 0.98
100 / 1.0
79 / 1.0
60 / 1.0
100 / 1.0
83 / 0.96
97 / 1.0
99 / 1.0
94 / 0.99
78 / 0.96
93 / 1.0
98 / 1.0
100 / 1.0
58 / 1.0
81 / 1.0
52 / 1.0
100 / 1.0
100 / 1.0
100 / 1.0
52 / 0.95
65 / 1.0
100 / 1.0
83 / 1.0
100 / 1.0
100 / 1.0
100 / 1.0
99 / 1.0
100 / 1.0
99 / 1.0
Xylariomycetidae Hypocreomycetidae
Lulworthiomycetidae
Cainiaceae
Xylariaceae
Graphostromataceae Diatrypaceae
Coniocessiaceae Vialaeaceae
Hyponectriaceae
Amphisphaeriaceae
Clypeosphaeriaceae
Apiosporaceae
Ceratostomataceae
Scortechiniaceae
Coronophoraceae
Bertiaceae
Nitschkiaceae
Chaetosphaerellaceae
Falcocladiaceae
Etheirophoraceae
Juncigenaceae
Torpedosporaceae
Ophiocordycipitaceae
Clavicipitaceae
Hypocreaceae
Cordycipitaceae
Stachybotriaceae
Nectriaceae
Bionectriaceae
Halosphaeriaceae
Microascaceae
Graphiaceae
Ceratocystidaceae
Gondwanamycetaceae
Glomerellaceae
Reticulascaceae
Australiascaceae
Plectosphaerellaceae
Savoryellaceae
Lulworthiaceae
Koralionastetaceae
Outgroup
Niessliaceae
- / 1.0
- / 1.0
- / 1.0
- / 1.0
Fig. 1 (continued)
Fungal Diversity

branched. Conidiophores micronematous or semimicronematous. Conidia filiform or ellipsoidal, septate ornon septate, some are coiled, when spermatial, spermatia,enteroblastic, subglobose.
Type order: Lulworthiales Kohlm. et al.This new subclass is introduced for a lineage of marine
fungi that are unrelated to Halosphaeriaceae (Microascales,Hypocreomycetidae). It includes the orders Lulworthialeswith the family Lulworthiaceae and Koralionastetales withthe familyKoralionastetaceae. Spatafora et al. (1998) demon-strated that the order Halosphaeriales was polyphyletic andcomprised two distinct l ineages. The first clade,Halosphaeriales including 11 genera, was closely related to,and is now included in the order Microascales, whereas thesecond clade, with Lulworthia and Lindra species, wasassigned to Lulworthiaceae in Lulworthiales (Kohlmeyeret al. 2000). Members of Lulworthiales are saprobes, onwood, sea grasses and marsh plants or parasites on algae(Campbell et al. 2005). Lindra, which is composed of sixspecies, is the only genus in the order Lulworthiales whichdoes not have ascospores with apical chambers filled withmucus. Other genera placed in the Lulworthiales based onmolecular data include Kohlmeyeriella (Campbell et al.2002) and the algicolous genera Spathulospora (Inderbitzinet al . 2004) and Haloguignardia (Harvey 2004).Koralionastes and Pontogeneia are closely related to mem-bers of Lulworthiales, while the differences in morphologicalcharacters are expressed in the ascospores and the presence/absence of periphyses and paraphyses. Phylogenetic studies ofCampbell et al. (2009) showed that some species ofKoralionastes and Pontogeneia that were sequenced groupin a monophyletic clade basal to the clade of Lulworthiales.This clade represents a distinct taxonomic entity at the orderlevel based on both molecular and morphological data. Basedon molecular studies and using morphological characters, thegenera Koralionastes and Pontogeneia were assigned to thenew order Koralionastetales which is a sister group toLu lwor t h i a l e s . Th e o r de r s Lu lwo r t h i a l e s andKoralionastetales clustered together in a well-supported cladebased on combined gene data and a new subclass is introducedhere as Lulworthiomycetidae. The orders in this subclass are,however, well separated.
Type species: Lulworthia fucicola G.K. Sutherl.
Meliolomycetidae P.M. Kirk & K.D. Hyde, subclass novusMeliolomycetidae P.M. Kirk et al., Ainsworth & Bisby’sDictionary of the Fungi, Edn 9 (Wallingford): 314 (2001)[nom. inval.]
This subclass Meliolomycetidae was introduced byKirk et al. (2001) to accomodate the order Meliolales.Justavino et al. (2014) provided a phylogenetic tree(based on analysis of LSU) that included five subclassesin Sordariomycetes, and eight species of Meliolaceae
which were analyzed by neighbor joining analysis. Theresults indicated that the Meliolaceae clade formed themost basal clade in Sordariomycetes with strong support.Hence, they suggested that the order Meliolales whichcomprises Armatellaceae and Meliolaceae should be ac-commodated in a subclass Meliolomycetidae. In ourmulti-gene phylogenetic analysis (Figs. 1 and 2) the sub-class Meliolomycetidae is most closely related toSordariomycetidae as in Justavino et al. (2014). TheMeliolales, clusters as a sister group to the orderChaetosphaeriales, family Cephalothecaceae and genusR i m a c o n u s . T h e p l a c e m e n t o f t h e f a m i l yCephalothecaceae is uncertain during the phylogeneticanalysis and this is probably because the sequences ofCephalothecaceae species are relatively short and do notprovide adequate phylogenetic resolution. The morpholo-gy of taxa of Cephalothecaceae and Meliolales differgreatly. In species of Meliolaceae have a parasitic habitat,forming superficial, web-like, black colonies on leaves,stems or branches, the mycelia are hyphopodiate andspores are mostly brown. Species of Cephalothecaceaeare distinct in having a saprobic habitat, often growingon rotten wood or on other fungi, the mycelia lackhyphopodia and ascomata have a cephalothecoid peridium(von Höhnel 1917c). Hence, we tentatively placeCepha lo thecaceae as fami ly i ncer tae sed i s i nSordariomycetes because of its uncertain phylogeneticplacement and differing morphology. There are few se-quences for species in Meliolomycetidae as they arebiotrophic and cannot presently be isolated into culture.Therefore all sequence data for Meliolales is from DNAextracted from fruiting bodies of taxa on the host.
Sordariomycetidae O.E. Erikss. & Winka, Myconet 1(1): 10(1997)
This subclass was established by Eriksson and Winka(1997) and comprises four orders, eight families and onefamily incertae sedis. The order Chaetosphaerialescomprises the famil ies Chaetosphaer iaceae andHelminthosphaeriaceae. The order Phyllachorales, which ismoved from Sordariomycetes orders incertae sedis toSo rd a r i omy c e t i d a e , c omp r i s e s two f am i l i e s ,Phaeochoraceae and Phyllachoraceae. Hyde et al. (1997)introduced the family Phaeochoraceae in the orderPhyllachorales for a taxon of biotrophic and saprotrophicfungi on palms. The order Boliniales comprises a singlefamily Bolinaceae. The order Sordariales includes threeclades represented by Sordariaceae, Chaetomiaceae andLasiosphaeriaceae, that latter comprises Lasiosphaeriaceaesensu lato and Lasiosphaeriaceae sensu stricto. Batistiaceae,typified by Batistia annulipeswas introduced by Samuels andRodrigues (1989) as a monotypic family. The family wasplaced in the Sordariomycetidae incertae sedis (Kirk et al.
Fungal Diversity

2001). Sequence data published by Huhndorf et al. (2004) hasbeen determined to be contaminated and there is no sequencedata for this family in GenBank.
Xylariomycetidae O.E. Erikss. & Winka, Myconet 1(1): 12(1997)
This subclass was introduced by Eriksson andWinka (1997)and comprises one lagre existing order Xylariales. The orderincludes Amphisphaeriaceae, Apiosporaceae, Cainiaceae,Coniocessiaceae, Diaptrypaceae, Graphostromataceae,Hyponectriaceae, Melanogrammataceae, Vialaeaceae, andXylariaceae.Many species in the subclass may have large stro-mata and are clearly visible and dominating the surface ofdecaying plants.
Orders of SordariomycetesThe phylogenetic trees produced in this study (Figs. 1 and 2)
support several of the existing orders and indicate that severalnew orders should be introduced. Each order is discussed al-phabetically.
Amplistromatales D’souza, Maharachch. & K.D. Hyde, ordonovus
Type family: AmplistromataceaeIndex Fungorum number: IF551156; Facesoffungi No.:
FOF 00632An order comprising families Amplistromataceae and
Catabotrydaceae. Sexual morph: Stromatic or non-stromatic.Stromata if present superficial, obovoid to pulvinate, soft orfirm textured. Ascomata perithecial, rarely subiculate, solitaryor gregarious, globose to sub globose, monostichous or polys-tichous with long necks.Paraphyses abundant, persistent, lon-ger than asci. Asci numerous, unitunicate, stipitate, cylindricalto clavate, with a small, J-, apical ring. Ascospores hyaline,globose, aseptate. Asexual morph: Acrodontium-like forAmplistroma.
Type species: Amplistroma carolinianum Huhndorf et al.Notes: Two genera are included in Amplistromatacaeae
namely Amplistroma Huhndorf et al. and Wallrothiella Sacc.The genus Wallrothiella was redescribed by Reblova andSeifert (2004) along with neotypification of its type speciesWallrothiella congregata (Wallr.) Sacc. Amplistroma has beensegregated from Wallrothiella on the basis of presence ofstromatic ascomata. Catabotydaceae is monotypic withCatabotrys as its type genus. However Catabotrys andWallrothiella do not possess stromatic ascomata, whileAmplistroma is stromatic. LSU sequence data analysis showsthat Amplistromataceae has an unsupported relationship withChaetospheriales and Magnaporthaceae (Huhndorf et al.2009). In our phylogenetic analyses Amplistromatales showsa distant relationship with Meliolomycetidae andSordariomycetidae and thus placed in Sordariomycetes orderincertae sedis until further data is available.
Annulatascales D’souza, Maharachch. & K.D. Hyde, ordonovus
Type family: AnnulatascaceaeIndex Fungorum number: IF551133; Facesoffungi No.:
FoF 00620A monotypic order within the class Sordariomycetes, sub-
class Diaporthomycetidae. Characterized by a dominantfreshwater lifestyle, although some members have been re-ported from terrestrial habitats. Sexual morph: Ascomata uni-locular, rarely clypeate, perithecial. Necks black or hyaline.Peridium coriaceous or membranous. Hamathecium with ta-pering paraphyses. Asci 8-spored, unitunicate, pedicellate,usually with a massive, J-, refractive apical ring. Ascosporesuniseriate, hyaline, sometimes brown, septate or aseptate.Asexual morph: Taeniolella-like for Chaetorostrum.
Type species: Annulatascus velatispora K.D. HydeNotes: This order is characterized by its typical freshwater
habitat, growing on submerged woody substrates, with mostgenera distributed in the tropics. Significant characters of mostgenera in Annulatascales include cylindrical, thin-walled asci,with a massive, J-, refractive, apical apparatus, which assistsin active spore ejection (Tsui et al. 2003). Ascospores areusually equipped with appendages or sheaths. These charac-ters might be important in aquatic habitats where they aid inthe attachment to substrates (Shearer et al. 2007). The orderpresently comprises 18 genera which belong to, or are referredto, the family Annulatascaceae. Many genera lack moleculardata, and it is likely that the family as presently circumscribed ispolyphyletic (Campbell and Shearer 2004; Huhndorf et al. 2004;Vijaykrishna and Hyde 2006; Abdel-Wahab et al. 2011). Theclosest relatives are, Cordanales and Papulosaceae, butAnnulatascales can be easily distinguished by a massive, J-,refractive, apical ring. Ranghoo et al. (1999) analyzed LSUrDNA sequence data to show thatAnnulatascaceae has relation-ships with Sordariales. However, in our phylogenetic analysesthe order Annulatascales is placed in Diaportheomycetidae(Figs. 1 and 2).
Boliniales P.F. Cannon, in Kirk et al., Ainsworth & Bisby’sDictionary of the Fungi, Edn 9 (Wallingford): x (2001)
An order of saprotrophic fungi within the subclassSordariomycetidae, which was introduced by Cannon (Kirket al. 2001). In our phylogenetic analysis (Figs. 1 and 2),Boliniales is closest to Phyllachorales and Sordariales.Boliniales comprises a single family, Boliniaceae, with ninegenera.
�Fig. 2 Maximum likelihood (ML) majority rule combined LSU, SSU,TEF and RPB2 consensus tree for the analyzed Sordariomycetes isolates.Orders are indicated in coloured blocks and RAxML bootstrap supportvalues (MLB) and Bayesian posterior probabilities (PP) are given at thenodes (MLB/PP). The scale bar represents the expected number ofchanges per site. The tree is rooted with Botryotinia fuckeliana,Dothidea sambuci, Exophiala dermatitidis and Leotia lubrica
Fungal Diversity

Stilbospora macrosperma CBS 121883
Pseudovalsa modonia AR 3558
Schizoparme straminea STE-U 3932
Melanconis marginalis AR3442
Thailandiomyces bisetulosus BCC00018
Valsa ambiens AR3516
Chromendothia citrina AFTOL-ID 2121
Ophiodiaporthe cyatheae YMJ 1364
Harknessia australiensis CPC 15029
Togninia africana STE U 6177
Pseudoplagiostoma oldii CBS 124808
Mazzantia napelli AR3498
Melanconis stilbostoma AR3501
Cryptometrion aestuescens CMW18790
Anisogramma virgultorum AV05
Cryptodiaporthe aesculi AFTOL-ID 1238
Pseudoplagiostoma variabile CBS 113067
Rossmania ukurunduensis AR3484
Jobellisia fraterna SMH2863
Togniniella acerosa PDD 81432
Plagiostoma euphorbiae CBS 340.78
Cryptosporella hypodermia AFTOL-ID 2124
Harknessia pseudohawaiiensis CPC 17379
Diaporthe eres AFTOL-ID 935
Pleurostomophora repens CBS 294.39
Pleurostoma ootheca CMU 23858
Jobellisia luteola SMH2753
Stegonsporium protopyriforme CBS 117041
Pseudovalsa longipes AR 3541
Togninia griseo olivacea STE U 5966
Diaporthales sp. BCC00200
Harknessia ellipsoidea CPC 17111
Pilidiella diplodiella STE U 3708
Pleurostomophora richardsiae CBS 270.33
Tirisporella beccariana BCC36737
Pilidiella wangiensis CPC 19397
Sydowiella stellatifolii CBS 119342
Phaeoacremonium pallidum STE U 6104
Jattaea mookgoponga STE-U 6184
Hapalocystis occidentalis WU 24705
Stegonsporium pyriforme CBS 117023
Stilbospora longicornuta CBS 122529
Valsella salicis AR 3514
Pseudoplagiostoma eucalypti CBS 124807
Harknessia eucalypti CBS 342.97
Schizoparme straminea CBS 149.22
Leucostoma niveum AR3413
Cryphonectria parasitica CMW 7048
Gnomonia gnomon CBS 199.53
Cytospora elaeagni CFCC 89633
Anisogramma anomala AA01
Cainiella johansonii Kruys 731 (UPS)
Luteocirrhus shearii CBS 130776
85 /-
98 / 1.0
97 / 1.0
72 / 0.93
77/ 0.99
75 / -
76 / -
73 / 1.0
60 / 0.96
93 / 0.98
68 / -
89 / 0.91
100 / 1.0
100 / 1.0
62 / 0.93
100 / 1.0
100 / 1.0
69 / 0.97
62 / 0.95
99 / 1.0
99 / 0.98
100 / 1.0
50 / -
100 / 1.0
94 / 1.00
100 / 1.0
84 / -
83 / 0.97
89 / 1.0
100 / 1.0
99 / 1.0
100 / 1.0
52 / 0.91
100 / 1.0
70 / 0.95
100 / 1.0
100 / 1.0
Schizoparmeaceae
Cryphonectriaceae
Harknessiaceae
Gnomoniaceae
Melanconidaceae
Valsaceae
Diaporthaceae
Stilbosporaceae
Pseudoplagiostomataceae
Sydowiellaceae
Tirisporellaceae
Pseudovalsaceae
Togniniaceae
Jobellisiaceae
Calosphaeriaceae
Pleurostomataceae
Diaporthales
Togniniales
Jobellisiales
Calosphaeriales
100 / 1.0
100 / 1.0
100 / 1.0
100 / 1.0
- / 1.00
100 / 1.0
Fungal Diversity

Apiorhynchostoma curreyi UAMH 11088
Fragosphaeria purpurea CBS 133.34
Buergenerula spartinae ATCC 22848
Phyllachora graminis UME 31349
Ophiostoma piliferum AFTOL-ID 910
Cryptadelphia groenendalensis SH12
Coniochaeta ligniaria C8
Cornipulvina ellipsoides SMH1378
Ophioceras commune M92
Coccodiella melastomatum CMU78543
Thyridium vestitum AFTOL-ID 172
Cryptadelphia groenendalensis SMH3767
Utrechtiana cibiessia CPC 18916
Lecythophora luteoviridis CBS 206.38
Bambusicularia brunnea INA-B-92-45
Slopeiomyces cylindrosporus CBS 609.75
Jugulospora rotula ATCC 38359
Annulatascus velatisporus HKUCC 3701
Lasiosphaeria ovina SMH4605
Cordana pauciseptata CBS 121804
Corynascella inaequalis CBS 284.82
Ceratocystiopsis minuta CBS 116963
Chaetomidium galaicum CBS 113678
Ophioceras dolichostomum CBS 114926
Coccodiella miconiae ppMP 1342
Ophioceras leptosporum CBS 894.70
Pyricularia borealis CBS 461.65
Pseudohalonectria lignicola M95
Ascovaginospora stellipala P5-13A
Magnaporthe salvinii M21
Achaetomium strumarium IMI 082624
Bussabanomyces longisporus CBS 125232
Raffaelea canadensis C2233
Apiosordaria verruculosa F-152365
Ascitendus austriacus MR 2936
Papulosa amerospora AFTOL-ID 748
Thyridium sp. Mas 56
Neurospora crassa MUCL 19026
Sordaria fimicola CBS 508.50
Camaropella pugillus SMH3846
Pseudopyricularia kyllingae HYKB202-1-2
Camarops ustulinoides AFTOL-ID 72
Pseudoproboscispora caudae A336-2D
Gelasinospora tetrasperma CBS 178.33
Pseudoneurospora amorphoporcata CBS 626.80
Annulusmagnus triseptatus CBS 128831
Bombardia bombarda AFTOL-ID 967
84 / 1.0
83 / 1.0
100 / 1.0
83 / 1.0
79 / 1.0
100 / 1.0
95 / -
100 / 1.0
53 / -
83 / -
87 / 1.0
76 / -
100 / 1.0
67 / -
100 / 1.0
65 / 1.0
63 / 0.96
100 / 1.0
76 / 0.95
100 / 1.0
75 / 1.0
95 / 1.0
88 / 1.090 / 1.0
84 / 1.0
100 / 1.0
100 / 1.0
74 / 0.94
58 / -
90 / 1.0
70 / -
87 / -
78 / 1.0
100 / 1.0
80 / -
81 / 0.95
88 / 1.0
100 / 1.0
93 / 1.0
66 / -
94 / 1.0
Pyriculariaceae
Magnaporthaceae
Ophioceraceae
Ophiostomataceae
Annulatascaceae
Coniochaetaceae
Cordanaceae
Trichosphaeriaceae
Thyridiaceae
Papulosaceae
Chaetomiaceae
Lasiosphaeriaceae
Sordariaceae
Boliniaceae
Phyllachoraceae
Magnaporthales
Ophiostomatales
Annulatascales
Coniochaetales Cordanales
Sordariales
Boliniales
Phyllachorales
- / 0.99
- / 0.98
Trichosphaeriales
Fig. 2 (continued)
Fungal Diversity

Graphostroma platystoma AFTO ID1249
Eutypa lata CBS 208.87
Coniocessia anandra Iran 1468C
Amplistroma erinaceum AH 43902
Diatrype disciformis AFTOL D 927
Ruzenia spermoides SMH 4655
Melanochaeta hemipsila SMH 2125
Apiospora bambusae ICMP 6889
Immersidiscosia eucalypti HHUF 29920
Clypeosphaeria uniseptata HKUCC 6349
Seynesia erumpensnew SMH 1291
Apiospora montagnei AFTOL D 951
Arthrinium hydei CBS 114990
Irenopsis cornuta VIC 32058
Rosellinia necatrix HKUCC 9037
Creosphaeria sassafras CM 018
Arthrinium phaeospermum HKUCC 3395
Pseudomassaria carolinensis 9502 (IFO)
Echinosphaeria canescens SMH 4791
Wallrothiella congregata SMH 1760
Beltraniella endiandrae CPC 22193
Asteridiella obesa VIC 31239
Rhopalostroma lekae
Arecophila bambusae HKUCC 4794
Vialaea minutella BRIP 56959
Discostroma botan
Chaetosphaeria innumera SMH 2748
Coniocessia nodulisporioides CBS 281.77
Synaptospora plumbea SMH 3962
Umbrinosphaeria caesariata CBS 102664
Amplistroma caroliniana BEO9923
Coniocessia maxima CBS 593.74
Robillarda sessilis BCC13393
Cephalotheca foveolata UAMH11631
Amphisphaeria umbrina AFTO ID 1229
Xylaria hypoxylon CBS 122620
Meliola centellae VIC 31244
Catabotrys deciduum SMH3436
Albertiniella polyporicola CBS 457.88
Cainia graminis CBS 136.62
Rimaconus jamaicensis SMH 4782
Lopadostoma turgidum LT2
Vialaea mangifia MFLUCC 12 0808
Phialemonium atrogriseum CBS 604.67
Cryptendoxyla hypophloia WM10.89
Monosporascus cannonballus FMR6682
Rimaconus coronatus SMH 5212
Helminthosphaeria hyphodermae SMH 4192
Exserticlava vasiformis TAMA 450
Endomeliola dingleyae PDD 98304
Astrocystis cocoes HKUCC 3441
Hyponectria buxi UME 31430
Irenopsis vincensii VIC 31751
Lepteutypa cupressi IMI 052255
85 / 0.99
89 / 1.0
50 / -
100 / 1.0
62 / -
- / 1.0
91 / 1.0
90 / 0.98
100 / 1.0
58 / 0.97
91/ 0.98
100 / 1.0
80 / 1.0
63 / 1.0
73 / 0.98
69 / 0.99
50 / -
83 / 0.96
98 / 1.0
76 / 1.0
73 / 0.99
60 / -
99 / 1.0
99 / 0.98
100 / 1.0
100 / 1.0
99 / 1.0
100 / 0.98
97 / 1.0
92 / 1.0
76 / 0.94
100 / 1.0
100 / 1.0
95 / 1.0
100 / 1.0
100 / 0.97
58 / 1.0
100 / 1.0
100 / 1.0
Chaetosphaeriaceae
Helminthosphaeriaceae
Cephalothecaceae
Meliolaceae
Amplistromataceae
Cainiaceae
Xylariaceae
Graphostromataceae
Diatrypaceae
Coniocessiaceae
Vialaeaceae
Hyponectriaceae
Amphisphaeriaceae
Clypeosphaeriaceae
Apiosporaceae
Chaetosphaeriales
Meliolales
Amplistromatales
Xylariales
100 / 1.0
- / 1.0
- / 1.0
- / 1.0
Catabotrydaceae
Fig. 2 (continued)
Fungal Diversity

Swampomyces triseptatus CY2802
Cryptosphaerella elliptica SMH4722
Melanospora tiffanii ATCC15515
Nectria cinnabarina CBS 114055
Hypocrea americana AFTO ID 52
Niesslia exilis CBS 560.74
Fracchiaea broomeana SMH347
Crassochaeta nigrita SMH2931
Torpedospora radiata AFTO ID 751
Torrubiella wallacei CBS 101237
Swampomyces armeniacus JK5325A
Peethambara spirostriata CBS 110115
Falcocladium multivesiculatum CBS 120386
Claviceps purpurea AEG 97 2
Shimizuomyces paradoxus EFCC 6279
Hymenostilbe aurantiaca OSC 128578
Scortechinia acanthostroma SMH1143
Stachybotrys chlorohalonata UAMH6417
Elaphocordyceps capitata OSC 71233
Hypocrea rufa DAOM JBT1003
Ascopolyporus polychrous PC546
Torpedospora radiata PP7763
Melanospora zamiae ATCC 12340
Trichoderma viride GJS
Gaillardiella pezizoides GKM1245
Neofracchiaea callista SMH2689
Glomerulispora mangrovis NBRC105264
Sphaerostilbella berkeleyanan CBS 102308
Moheitospora fruticosae EF14
Marinokulati chaetosa BCRC FU30271
Hypocrella discoidea BCC 8237
Bertia moriformis SMH4320
Hyperdermium pulvinatum PC602
Nitschkia tetraspora GKML148N
Spinulosphaeria nuda SMH1952
Ophiocordyceps sinensis YN09 64
Bertia ngongensis GKM1239
Parasarcopodium ceratocaryi CBS 110664
Cordyceps militaris OSC 93623
Myrothecium roridum ATCC 16297
Etheirophora blepharospor JK5289
Bertia tropicalis SMH3513
Viridispora diparietispora ATCC MYA 627
Epichloe typhina ATCC 56429
Scopinella solani CBS 770.84
Neonectria ramulariae CBS 151.29
Vittatispora coorgii BICC 7817
Acanthonitschkea argentinensis SMH1395
Falcocladium sphaeropedunculatum CBS 111292
Etheirophora unijubata JK5443B
Scortechiniellopsis leonensis GKM1269
Cosmospora coccinea AR2741
Coronophora gregaria ANM1555
Elaphocordyceps japonica OSC 110991
Chaetosphaerella phaeostroma SMH4257
Chaetosphaerella fusca GKML124N
Juncigena adarca K5548A100 / 1.0
100 / 1.0
83 / 1.0
100 / 0.98
81 / 1.0
73 / 0.98
100 / 1.0
100 / 1.0
100 / 1.0
68/ -
79 / 1.0
100 / 1.0
87 / 1.0
74 / 0.96
59 / -
94 / 0.99
88 / 1.0
96 / 1.0
100 / 1.0
83 / -
95 / 1.0
100 / 1.0
64 / 1.0
60 / -
100 / 1.0
98 / 1.0
100 / 1.0
100 / 1.0
66 / -
88 / 1.0
- / 1.0
52 / 1.0
98 / 1.0
88 / 1.0
96 / 1.0
100 / 1.0
76 / 0.99
52 / 0.95
98 / 1.0
93 / 1.0
97 / 1.0
94 / 1.0
94 / 1.0
84/ 1.060 / 1.0
56 / -
- / 1.0
- / 0.98
99 / 1.0
100 / 1.0
65 / 1.0
Ceratostomataceae
Scortechiniaceae
Coronophoraceae
Bertiaceae
Nitschkiaceae
Chaetosphaerellaceae
Falcocladiaceae
Etheirophoraceae
Juncigenaceae
Torpedosporaceae
Ophiocordycipitaceae
Clavicipitaceae
Hypocreaceae
Cordycipitaceae
Stachybotriaceae
Nectriaceae
Niessliaceae
Melanosporales
Coronophorales
Falcocladiales
Hypocreales
Stephanonectria keithii GJS92 133
Bionectria ochroleuca AFTOL D 187
Roumegueriella rufula GJS 91 164
Hydropisphaera peziza GJS92 101100 / 1.0
91 / 1.0
100 / 1.0Bionectriaceae
100 / 1.0
- / 0.97
Fig. 2 (continued)
Fungal Diversity

0.2
Colletotrichum fructicola LC0032
Conioscypha japonica CBS 387.84
Botryotinia fuckeliana AFTOL D 59
Koralionastes ellipticus JK5769
Thielaviopsis thielavioides CBS 130.39
Canalisporium caribense SS038397
Graphium fimbriasporum CMW 5605
Gondwanamyces capensis AFTOL- D 1907
Cumulospora marina MF 46
Monilochaetes infuscans CBS 869.96
Petriella setifera AFTOL D 956
Exophiala dermatitidis AFTOL
Halosphaeria appendiculata CBS 197.60
Porosphaerellopsis sporoschismophora ATCC 42528
Kylindria peruamazonensis CBS 838.91
Pleurothecium semifecundum CBS 131271
Hydea pygmea NBRC 33069
Colletotrichum asianum LC0037
Lindra thalassiae AFTOL D 413
Dothidea sambuci DAOM 231303
Ascothailandia grenadoidia N 2010a
Graphium penicillioides CBS 506.86
Leotia lubrica OSC 100001
Doratomyces stemonitis AFTOL D 1380
Nimbospora effusa JK 5104A
Australiasca queenslandica BR 24607
Carpoligna pleurothecii CBS 114211
Lignincola laevis JK50180
Reticulascus clavatus CBS 125239
Reticulascus clavatus CBS 125296
Remispora maritima
Stachylidium bicolor DAOM 226658
Verticillium dahliae ATCC 16535
Colletotrichum brevisporum LC0600
Microascus longirostris CBS 267.49
Ambrosiella ferruginea CBS 460.82
Custingophora cecropiae CCF 3568
Colletotrichum gloeosporioides LC0555
Plectosphaerella cucumerina DAOM 226828
Ceratocystis fimbriata C89
Savoryella lignicola NF00204
Ascotaiwania lignicola N L00005
Conioscyphascus varius CBS 113653
Gibellulopsis nigrescens DAOM 226890
Lulworthia fucicola ATCC 64288
100 / 1.0
100 / 1.0
100 / 1.0
98 / 1.0
100 / 1.0
82 / 1.0
97 / 1.0
99 / 1.0
100 / 1.0
98 / 1.0
82 / 0.98
100 / 1.0
92 / -
100 / 1.0
96 / -
73 / 0.9
100 / 1.0
99 / 1.0
85 / 0.98
100 / 1.0
100 / 1.0
55 / 0.93
82 / 1.0
68 / -
70 / -
99 / 1.0
61 / 1.0
99 / 1.0
95 / 1.0
100 / 1.0
100 / 1.0
100 / 1.0
96 / 0.99
90 / 1.0
78 / 0.96
99 / 1.0
100 / 1.0
93 / 1.0
Halosphaeriaceae
Microascaceae
Graphiaceae
Ceratocystidaceae
Gondwanamycetaceae
Glomerellaceae
Reticulascaceae
Australiascaceae
Plectosphaerellaceae
Savoryellaceae
Lulworthiaceae
Koralionastetaceae
Microascales
Glomerellales
Savoryellales
Lulworthlales
Koralionastetales
100 / 1.0
100 / 1.0
100 / 1.0
100 / 1.0
Fig. 2 (continued)
Fungal Diversity

Calosphaeriales M.E. Barr, Mycologia 75(1): 11 (1983)This small order was introduced by Barr (1985). It present-
ly comprises Calosphaeriaceae and Pleurostomataceae(Figs. 1 and 2). The taxa belonging to the order mainly com-prise simple, dark perithecia, unitunicate asci, and hyaline toslightly pigmented, ellipsoid to allantoid ascospores (Réblováet al. 2004). It forms a sister clade to the order Jobellisiales(Figs. 1 and 2).
Chaetosphaeriales Huhndorf et al., Mycologia 96(2): 378(2004)
This order was introduced in Sordariomycetidae based onmolecular analysis of LSU nrDNA sequence data byHuhndorf et al. (2004). In our phylogenetic analysis (Fig. 1),Chaetosphaeriales is close to Boliniales and Phyllachorales.The order comprises Chaetosphaeriaceae with 35 genera andHelminthosphaeriaceae with seven genera.
ConiochaetalesHuhndorf et al., Mycologia 96(2): 378 (2004)This order was introduced by Huhndorf et al. (2004) and
incorporates a single family Coniochaetaceae with three gen-era. Cordanales is a sister order and Papulospora(Papulospoaceae) is basal to Coniochaetiaceae and may alsorequire its own order. Coniochaetaceae differs fromSordariaceae and related families in having ascospores withelongate germ slits (Malloch and Cain 1971).
Cordanales Hernández-Restrepo et al., Phytotaxa (in press)This order was introduced by Hernández-Restrepo et al.
(2015) to accommodate the family Cordanaceaewith a singlegenus Cordana. Morphologically it is easily distinguishedfrom its sister order Coniochaetales (Coniochaetaceae) byhaving a basal stroma, ascospores without germ slits andpolyblastic asexual morphs (Hernández-Restrepo et al. 2015).
Coronophorales Nannf., Nova Acta R. Soc. Scient. upsal.,Ser. 4 8(no. 2): 54 (1932)
The wood inhabiting order Coronophorales comprisesBertiaceae, Ceratostomataceae, Chaetosphaerellaceae,Coronophoraceae, Nitschkiaceae and Scortechiniaceae. Thetaxa in the order are characterised by mostly superficialascomata, sometimes with an extensive hyphal subiculum orwell developed basal stroma that often becomes cupulate orcollapsed, and in some cases the ostiolar opening is eitherindistinct or lacking (Mugambi and Huhndorf 2010).
DiaporthalesNannf., Nova Acta R. Soc. Scient. upsal., Ser. 48(no. 2): 53 (1932)
The order Diaporthales comprises 12 families which areCryphonectriaceae, Diaporthaceae, Gnomoniaceae,Harknessiaceae, Melanconidaceae, Pseudoplagiostomataceae,Pseudovalsaceae, Schizoparmeaceae, Stilbosporaceae,Sydowiellaceae, Tirisporellaceae and Valsaceae. The family
Tirisporellaceae will be introduced by Suetrong et al.(2015) to accommodate a basal clade of Diaporthalescomprising the marine ascomycete geneus Tirisporellaand the freshwater genus Thailandiomyces. Members ofDiaporthales are pathogens, parasites, and endophytesof plants, human-animal pathogens, saprobes and soilinhabi tants (Rossman et a l . 2007) . The orderDiaporthales is characterized by perithecia with an elon-gate beak, often formingwithin stromatic tissues(Rossman et al. 2007). Asci generally deliquesce at thebase when mature and have a characteristic refractiveapical annulus.
Falcocladiales R.H. Perera, Maharachch., S. Somrithipol, S.Suetrong & K.D. Hyde, ordo novus
Type family: FalcocladiaceaeIndex Fungorum number: IF551132; Facesoffungi No.:
FoF 00606This monotypic order in the subclass Hypocreomycetidae
is introduced to accommodate the family Falcocladiaceae.Members of this family are saprobes on leaf litter and leavesincluding Eucalyptus grandis and E. camaldulensis in tropi-cal, terrestrial habitats (Crous et al. 1994; Jones et al. 2014).Sexual morph: Undetermined. Asexual morph: Conidiomatahyaline, sporodochial or synnematal, or penicillate, arisingfrom the mycelium or from a stroma or from microsclerotia,thick-walled with non-septate stipe extensions that terminatein thin-walled sphaeropendunculate vesicles. Conidiophoreshyaline, non to multi-septate, branched, forming three seriesof branches per conidioma (primary, secondary and tertiary),subcylindical. Conidiogenous cells phialidic, arranged in 2–6whorls, necks elongate, with minute collarettes. Conidia trans-septate, hyaline, falcate, with short apical and basal append-ages (Jones et al. 2014).
Type species: Falcocladium multivesiculatum S.F.Silveira et al.
Notes: The order presently includes one hyphomycetousasexual genus Falcocladium introduced by Crous et al.(1994). Jones et al. (2014) introduced the monotypic familyFalcocladiaceae based on SSU and LSU sequence data toaccommodate the members of genus Falcocladium and sug-gested further taxon sampling was needed to determine itsordinal status. There is no recorded sexual morph for thisorder. In the molecular analysis (Figs. 1 and 2)Falcocladiales is most closely related to the orderCoronophorales.
Glomerellales Chadef. ex Réblová et al., Stud. Mycol. 68(1):170 (2011a, b)
Chadefaud (1960) had proposed the order BGlomérellales^but without a Latin diagnosis and the name was thus invalid.The order Glomerellales was therefore validly published byRéblová et al. (2011a) in the class Sordariomycetes and
Fungal Diversity

comprised three families Australiascaceae, Reticulascaceaeand Glomerellaceae. The introduction was based on analysisof ITS1, 5.8S rDNA and ITS2 (ITS), nc28S (LSU) rDNA, andnc18S (SSU) rDNA datasets, and a combined data set of LSU-SSU-RPB2.Hypocreales Lindau, in Engler & Prantl, Nat. Pflanzenfam.,Teil. I (Leipzig) 1(1): 343 (1897)
Members of the Hypocreales are highly diverse in the tro-pics and subtropics (Põldmaa 2011). This larger order is rep-resent by Bionectriaceae, Clavicipitaceae, Cordycipitaceae,H y p o c r e a c e a e , N e c t r i a c e a e , N i e s s l i a c e a e ,Oph i o co rd y c i p i t a c e a e , S t a c h y bo t r i a c e a e andTilachlidiaceae. In the phylogenetic analysis (Figs. 1 and 2)i t forms a sis ter clade to Trichosphaeriales andFalcocladiales.
Jobellisiales D’souza & K.D. Hyde, ordo novusType family: JobellisiaceaeIndex Fungorum number: IF551134; Facesoffungi No.:
FoF 00621Jobellisiales is a monotypic order in the class
Sordariomycetes, subclass Diaporthomycetidae. It includes asingle family Jobellisiaceae characterized by saprobiclignicolous taxa found in terrestrial and freshwater habitats.Sexual morph: Ascomata superficial, basally immersed,brown to black, or externally with yellowish pigments, papil-late perithecial. Peridium 3-layered, with an orange, middlewall layer. Hamathecium with paraphyses. Asci unitunicate,cylindrical, with a large, refractive, J-, apical ring. Ascosporesuniseriate, 1-septate, coloured, with germ pores. Asexualmorph: Undetermined.
Type species: Jobellisia luteola (Ellis & Everh.) M.E. BarrNotes: This order encompasses taxa that possess relatively
large, superficial ascomata, a three layered, thick peridium,cylindrical asci and one septate ascospores. Ascomata ofJobellisia luteola (Ellis & Everh.) M.E. Barr are peculiar inappearing orange to yellow brown, whereas those of J. barriiHuhndorf et al. and J. viridifuscaK.M. Tsui &K.D. Hyde havea bright orange middle wall layer (Ranghoo et al. 2001, Liuet al. 2012). The family Jobellisiaceae was introduced byRéblová (2008) as a monotypic family and currently includesseven species. Réblová (2008) using LSU rDNA sequenceanalysis showed that the closest relatives to Jobellisiaceae areDiaporthales, Calosphaeriales and Togniniaceae. No asexualmorph is reported for this order. In our phylogenetic analysis(Figs. 1 and 2) this order is closest to Calosphaeriales.
Koralionastetales Kohlm. et al., Mycol. Res. 113(3): 377(2009)
Based on the combined dataset of SSU and LSU rDNAsequences and morphological characters the generaKoralionastes and Pontogeneia were assigned to the new or-de r Kora l ionas t e ta l e s (Campbe l l e t a l . 2009) .
Koralionastetales shows phylogenetic affinities toLulworthiales, but differences in morphology are expressedin the ascospores and the presence/absence of periphysesand paraphyses (Campbell et al. 2009).
Lulworthiales Kohlm. et al., Mycologia 92(3): 456 (2000)Kohlmeyer et al. (2000) assigned two genera Lulworthia
and Lindra, to this new order and the new family(Lulworthiaceae). Abdel-Wahab et al. (2010) showed that anumber of asxual morphs also belong in this family, ieHalazoon, Hydea and Orbimyces. Members of Lulworthialeslack an apical apparatus in the asci and sexual morphs membersof the Lulworthiales have ascospores with apical chambersfrom which mucilage is released (Campbell et al. 2005) andtogether with the order Koralionastes belong to the new sub-class Lulworthiomycetidae introduced in this study.
Magnaportha`les Thongk. et al., Fungal Diversity 34: 166(2009)
A phylogenetic study of Pyricularia and related genera inMagnaporthales by Klaubauf et al. (2014) resulted in the in-troduction of two new families, Ophioceraceae andPyriculariaceae. Ophioceraceae comprises a single genusOphioceras, containing species that mostly occur on woodsubmerged in freshwater. Magnaporthaceae was consideredto include Buergenerula, Bussabanomyces, Endopyricularia,Gaeumannomyces, Harpophora, Magnaporthiopsis,Nakataea, Omnidemptus, Pyriculariopsis and Slopeiomyceswith Nakataea as the correct name for the type of the family.The new genus Kohlmeyeriopsis was introduced based onmolecular data and morphology. Pyriculariaceae was intro-duced and included Deightoniella and Pyricularia, and then ew g e n e r a Bambu s i c u l a r i a , Ba r re t om y c e s ,Macgarvieomyces, Neopyricularia, Proxipyricularia,Pseudopyricularia and Xenopyricularia.
Melanosporales N. Zhang & M. Blackw., in Hibbett et al.,Mycol. Res. 111(5): 531 (2007)
This order was suggested in Zhang et al. (2006) but notvalidly published; it was formaly introduced by Hibbettet al . (2007) to accommodate Melanospora andSphaerodes in Ceratostomataceae. Melanosporales forms asister clade to Coronophorales in the present phylogeneticanalysis (Figs. 1 and 2).
Meliolales Gäum et al., Syst. Ascom. 5(1): 180 (1986)This o rde r was in t roduced by Er iksson and
Hawksworth (1985) and it accommodates a single familyMeliolaceae, which are the black mildews and comprisemore than 26 genera. This group is unique in producingblack web-like colonies on the host, comprising superfi-cial brown to black mycelium with appressoria(Hosagoudar 2004). Until recently the family was thought
Fungal Diversity

to be a member of Dothideomycetes (Huhndorf andLumbsch 2007).
Microascales Luttr. et al., Mycotaxon 12(1): 40 (1980)The order Microascales was introduced by Benny andKimbrough (1980) to accommodate Chadefaudiellaceae,Microascaceae and Pithoascaceae, and later expanded to in-clude Ceratocyst idaceae , Gondwanamycetaceae ,Halosphaeriaceae and Graphiaceae (Réblová et al. 2011a).
Ophiostomatales Benny & Kimbr., Mycotaxon 12(1): 48(1980)
This order was introduced by Benny and Kimbrough(1980) for the family Ophiostomataceae; plus Kathistaceaewas added byMalloch and Blackwell (1990). In present studyOphiostomatales forms a well supported sister clade toMagnaporthales.
Phyllachorales M.E. Barr, Mycologia 75(1): 11 (1983)The order Phyllachorales was introduced by Barr
(1983) comprising two clades, viz. Phaeochoraceae andPhyllachoraceae. This order is distinctive as the speciesare biotrophs on various hosts, where they form bright orblack stroma (Pearce and Hyde 1994). Asci areunitunicate and ascospores are hyaline or lightlypigmented (Barr 1983; Pearce and Hyde 1993a, b, 1994,Pearce et al. 2001). In our phylogenetic study (Figs. 1 and2), molecular data from a species of Phyllachoraceae con-f i rmed that Phyl lachorales can be inc luded inSordariomycetidae with higher support.
Pisorisporiales Réblová & J. Fourn., Persoonia 34: 43 (2015)This monotypic order was introduced by Réblová et al.(2015) based on LSU, SSU and RPB2 sequence data. Itcurrently includes the family Pisorisporiaceae. In theiranalyses, the Pisorisporiales nested in a weakly supportedc l ad e s i s t e r t o t h e o rd e r s Lu lwor t h i a l e s andKo r a l i o n a s t e t a l e s a n d i s i n c l u d e d h e r e i nSordariomycetes subclass incertae sedis.
Sordariales Chadef. ex D. Hawksw. & O.E. Erikss., Syst.Ascom. 5(1): 182 (1986)
This order was introduced by Eriksson and Hawksworth(1985) and is well-supported in the phylogenetic tree (Figs. 1and 2). It comprises three families viz. Chaetomiaceae,Sordariaceae and Lasiosphaeriaceae sensu lato. Most specieseither grow on dung or decaying substrata such as wood or areaquatic growing on submerged wood (Zhang et al. 2006). TheSordariales are characterized by membraneous or coriaceousascomata, and hyaline or brown ascospores often with ap-pendages or sheaths (Zhang et al. 2006). Our molecular tree(Figs. 1 and 2) places the order in the subclassSordariomycetidae.
Savoryellales Boonyuen et al., in Boonyuen et al., Mycologia103(6): 1368 (2011)
The taxonomic placement of the genus Savoryella has beenwidely debated and Jones et al. (2009a, b) referred it to theSordariales genera incertae sedis. Boonyuen et al. (2011), in acombined phylogenetic analysis of Savoryella species (LSU,SSU, 5.8S rRNA genes, RPB1, RPB2, TEF), showed thatthey formed a monophyletic group in the Sordariomycetes,but showed no affinities with other accepted orders. The orderSavoryellales was introduced to accommodate Savoryellaspecies, along with the genera Ascotaiwania, Ascothailandia(and its asexual morph Canalisporium), as they formed a dis-tinct lineage in the Sordariomycetes (Boonyuen et al. 2011).
Togniniales Senan., Maharachch. & K.D. Hyde, ordo novusType family: Togniniaceae
Index Fungorum no: IF551049; Facesoffungi number: FoF00592
Togniniales is a monotypic order in the classSordariomycetes, subclass Diaporthomycetidae. It includes asingle family Togniniaceae containing species saprobic ondead wood. Sexual morph: Ascomata lacking stromatic tis-sues, perithecial, with 1–3 necks. Hamathecium comprisingparaphyses. Asci 8 or poly-sporous, unitunicate, clavate, withsessile bases, occurring on hyaline and branched ascogenoushyphae. Ascospores hyaline and unicellular. Asexual morph:Conidiophores long or short, branched or unbranched.Conidia hyaline, aseptate.
Type species: Togninia minima (Tul. & C. Tul.) Berl.Notes: This monotypic order is introduced based on its
phylogenetic and morphological distinctiveness to accom-modate a single family, Togniniaceae comprising the sex-ual morph genera Conidiotheca and Togninia and theasexual morph genus Phaeoacremonim (Réblová andMostert 2007). Togniniaceae had been placed inCalosphaeriales based on both unique morphologicalcharacters and phylogeny (Mostert et al. 2003). Mostertet al. (2006) placed Togniniaceae in Diaporthales basedon SSU and LSU rRNA gene analysis. In this studyTogn in iaceae f o rms a d i s t i n c t c l ade ba sa l t oDiaporthales in the subclass Diaportheomycetidae and issister to Diaporthales and Calosphaeriales. Hence, weexclude Togniniaceae from Diaporthales and introduce anew order Togniniales.
Trichosphaeriales M.E. Barr, Mycologia 75(1): 11 (1983)This order was introduced by Barr (1983) based on
Trichosphaeria pilosa (Pers.) Fuckel. The current classifica-tion of Trichosphaeriales recognizes only one family, the ter-restrial Trichosphaeriaceae. Trichosphaeriaceae has affinitieswith Papulosporaceae and Thyridiaceae, however until morecollections and data become available it is better to maintainthese familes.
Fungal Diversity

Xylariales Nannf., Nova Acta R. Soc. Scient. upsal., Ser. 48(no. 2): 66 (1932)
The Xylariales is a large order of perithecial ascomyceteswith eight-spored unitunicate asci, with a J+, apical apparatus(Smith et al. 2003). Presently there are 11 families accepted inXylariales; Amphisphaeriaceae, Apiosporaceae, Cainiaceae,Clypeosphaeriaceae, Coniocessiaceae, Diatrypaceae,Graphostromataceae, Hyponectriaceae, Melogrammataceae,Vialaeaceae and Xylariaceae.
Outline of Sordariomycetes, 31 December 2014
Class SORDARIOMYCETES sensu O.E. Erikss. & WinkaDiaportheomycetidae Senan. et al.
Annulatascales D’souza et al.*Annulatascaceae S.W. Wong et al.
Annulatascus K.D. Hyde$
Annulusmagnus J. Campb. & ShearerAqualignicola V.M. et al.#$
Aquaticola W.H. Ho et al.Ascitendus J. Campb. & ShearerAyria Fryar & K.D. Hyde#
Cataractispora K.D. Hyde et al.$
Chaetorostrum Zelski et al.*#
Clohiesia K.D. Hyde$
Cyanoannulus Raja et al.$
Diluviocola K.D. Hyde et al.#
Fusoidispora D. Vijaykrishna et al.$
Longicollum Zelski et al.*#
Pseudoproboscispora Punith.$
Rhamphoria Niessl.$
Submersisphaeria K.D. Hyde$
Torrentispora K.D. Hyde et al.#
Vertexicola K.D. Hyde et al.$
Calosphaeriales M.E. BarrCalosphaeriaceae Munk
Calosphaeria Tul. & C Tul.*= Calosphaeriophora Réblová et al.
Jattaea Berl*$
= Phragmocalosphaeria Petr.= Wegelina Berl.
Kacosphaeria Speg.#
Sulcatistroma A.W. Ramaley#
Togniniella Réblová et al.*$
= Phaeocrella Réblová et al.Tulipispora Révay & J. Gönczöl, in Révay et al.#
Pleurostomataceae Réblová et al.Pleurostoma Tul. & C. Tul.$
= Pleurostomophora Vijaykr. et al.$
Calosphaeriales genera, incertae sedisEnchnoa Fr.#
Coniochaetales Huhndorf et al.Coniochaetaceae Malloch & Cain
Barrina A.W. RamaleyConiochaeta (Sacc.) Cooke$
Lecythophora Nannf$
Cordanales M. Hern.-Rest. & CrousCordanaceae Nann.
Cordana Preuss$
= Porosphaerella E. Müll. & Samuels
Diaporthales Nannf.Cryphonectriaceae Gryzenh. & M.J. Wingf.
Amphilogia Gryzenh. et al.Aurantiosacculus Dyko & B. Sutton*Aurapex Gryzenh. & M.J. Wingf.Aurifilum Begoude et al.Celoporthe Nakab. et al.Chromendothia Lar.N. Vassiljeva$
Chrysocrypta Crous & Summerell*Chrysoporthe Gryzenh. & M.J. Wingf.*
= Chrysoporthella Gryzenh. & M.J. Wingf.Cryphonectria (Sacc.) Sacc. & D. Sacc.$
Cryptometrion Gryzenh. & M.J. Wingf.*$
Diversimorbus S.F. Chen & J. Roux*Endothia Fr.Endothiella Sacc.Foliocryphia Cheewangkoon & CrousHolocryphia Gryzenh. & M.J. Wingf.Immersiporthe S.F. Chen et al.*Lasmenia Speg.*Latruncellus M. Verm. Et al.Luteocirrhus C.F. Crane & T.I. Burgess*$
Mastigosporella Höhn.Microthia Gryzenh. & M.J. Wingf.Rostraureum Gryzenh. & M.J. Wingf.Ursicollum Gryzenh. & M.J. Wingf.
Diaporthaceae Höhn. ex Wehm.Allantoporthe Petr.#
Apioporthella Petr.#
Clypeoporthella Petr.*#
Diaporthe Nitschke*$
= Phomopsis (Sacc) Bubák*Diaporthella Petr.Diaporthopsis Fabre#
Leucodiaporthe M.E. Barr & Lar.N. Vassiljeva#
Mazzantia Mont.$
Mazzantiella Höhn. #
Fungal Diversity

Ophiodiaporthe Y.M. Ju et al.*$
Pustulomyces D.Q. Dai et al.*
Gnomoniaceae G. WinterAlnecium Voglmayr & JaklitschAmbarignomonia SogonovAmphiporthe Petr.Anisomyces Theiss. & Syd.#
Apiognomonia Höhn.Apioplagiostoma M.E. BarrAsteroma DCBagcheea E. Müll. & R. Menon#
Clypeoporthe Höhn.#
Cryptosporella Sacc.$
Cylindrosporella Höhn.#
Depazea Fr.#
Diplacella Syd.#
= Discosporium Höhn.#
= Discula Sacc.Ditopella De Not.Ditopellopsis J. Reid & C. BoothGloeosporidina Petr.#
Gnomonia Ces. & De Not.$
Gnomoniella Sacc.Gnomoniopsis Berl.Mamiania Ces & De Not.#
Millerburtonia Cif #
Occultocarpon L.C. Mejía & Zhu L. Yang*Ophiognomonia (Sacc.) Sacc.Phragmoporthe Petr.Phylloporthe Syd.#
Plagiostoma Fuckel*$
= Cryptodiaporthe Petr.= Diplodina Westend
Pleuroceras Riess.= Linospora Fuckel
Skottsbergiella Petr.#
Sirococcus Preuss*Spataporthe Bronson et al.*#
Uniseta Ciccar#
Xenotypa Petr.#
Zythia Fr.
Harknessiaceae Crous*Harknessia Cooke*$
Melanconidaceae G. WinterBotanamphora Nograsek & Scheuer#
Ceratoporthe Petr.#
Cytomelanconis Naumov#
Dicarpella Syd. & P. Syd.Dictyoporthe Petr.#
Fremineavia Nieuwl.#
Gibellia Sacc.#
Hypophloeda K.D. Hyde & E.B.G. Jones*#
Kensinjia J. Reid & C. Booth#
Macrodiaporthe Petr.#
Massariovalsa Sacc.#
Mebarria J. Reid & C. Booth#
Melanamphora Lafl.#
Melanconiella Sacc.*Melanconiopsis Ellis & Everh.#*Melanconis Tul. & C. Tul.$
Melanconium LinkPhragmodiaporthe Wehm.#
Plagiophiale Petr.#
Plagiostigme Syd.#
Prosthecium Fresen.Prostratus Sivan. et al.#
Pseudovalsella Höhn.#
Wehmeyera J. Reid & C. Booth#
Wuestneia Auersw. ex FuckelWuestneiopsis J. Reid & Dowsett#
Pseudoplagiostomataceae Cheew. et al.Pseudoplagiostoma Cheew. et al.$
Pseudovalsaceae M.E. BarrApoharknessia Crous & S.J. LeeCoryneum Nees#
Pseudovalsa Ces. & De Not.$
Schizoparmeaceae RossmanConiella Höhn.
= Baeumleria Petr. & Syd.Pilidiella Petr. & Syd.$
Schizoparme Shear$
Stilbosporaceae Link*Natarajania Pratibha & BhatStilbospora Pers.Stegonsporium Corda
Sydowiellaceae Lar.N. VassiljevaCainiella E. Müll.$
Calosporella J. SchrötChapeckia M.E. BarrHapalocystis Auersw. ex Fuckel$
Lambro Racib.#
Rossmania Lar.N. Vassiljeva$
Sillia P. Karst.Stegophora Syd. & P. Syd.#
Sydowiella Petr.$
Uleoporthe Petr.#
Winterella (Sacc.) Kuntze
Fungal Diversity

Tirisporellaceae Suetrong et al.Tirisporella E.B.G. Jones et al.$
Thailandiomyces Pinruan et al.$
Valsaceae Tul. & C. Tul.Amphicytostroma Petr#
Chadefaudiomyces Kamat et al.#
Cryptascoma Ananthap.#
Cytospora Ehrenb.*$
= Leucostoma (Nitschke) Höhn.$
= Valsa Fr.$
= Valsella Fuckel$
= Valseutypella Höhn.Ditopellina J. Reid & C. Booth#
Durispora K.D. Hyde#
Harpostroma Höhn.*#
Hypospilina (Sacc) Traverso#
Kapooria J. Reid & C. Booth#
Leptosillia Höhn.#
Maculatipalma J. Fröhlich & K.D. HydePachytrype Berl. ex M.E. Barr et al.*Paravalsa Ananthapadm.#
Diaporthales, genera incertae sedisArgentinomyces N.I. Peña & Aramb.*#
Anisogramma Theiss. & Syd.$
Anisomycopsis I. Hino & Katum.#
Apiosporopsis (Traverso) Mariani$
= Sphaerognomonia Potebnia ex Höhn.Apomelasmia Grove#
Auratiopycnidiella Crous & Summerell*Bagadiella Cheew. & Crous*Caudospora Starbäck#
Chaetoconis Clem.*Cryptoleptosphaeria Petr.#
Cryptonectriella (Höhn.) Weese#
Cryptonectriopsis (Höhn.) Weese#
Diatrypoidiella Manohar et al.#
Disculoides Crous et al.*Dwiroopa Subram. & Muthumary#
Erythrogloeum Petr.*Exormatostoma Gray#
Greeneria Scribn. & Viala*Gyrostroma Naumov*Hercospora Fr.Hyalorostratum Raja & Shearer*Hypodermina Höhn.#
Keinstirschia J. Reid & C. Booth#
Lollipopaia InderbitzinMacrohilum H.J. SwartMamianiella Höhn.Pseudocryptosporella J. Reid & C. Booth#
Pseudothis Theiss. & Syd.#
Rabenhorstia Fr.Savulescua Petr.#
Stenocarpella Syd. & P. Syd.$
Sphaerognomoniella Naumov & Kusnezowa#
Stioclettia Dennis#
Trematovalsa Jacobesco#
Tubakia B. SuttonValsalnicola D.M. Walker & Rossman*Vismaya V.V. Sarma & K.D. Hyde#
Jobellisiales D’souza & K.D. Hyde*Jobellisiaceae Réblová
Jobellisia M.E. Barr$
Magnaporthales Thongk. et al.*Magnaporthaceae P.F. Cannon
Buergenerula Syd.$
Bussabanomyces Klaubauf et al.$
Ceratosphaerella Huhndorf et al.Ceratosphaeria Niessl.Clasterosphaeria Sivan.#
Clasterosporium Schwein#
Clavatisporella K.D. Hyde#
Gaeumannomyces Arx & D.L. OlivierHarpophora W. GamsHerbampulla Scheuer & Nograsek#
Kohlmeyeriopsis Klaubauf et al.Magnaporthe R.A. Krause & R.K. Webster$
Magnaporthiopsis J. Luo & N. Zhang*Muraeriata Huhndorf et al.Mycoleptodiscus Ostaz.Nakataea HaraOmnidemptus P.F. Cannon & AlcornPhomatospora Sacc.*Pyriculariopsis M.B. EllisSlopeiomyces Klaubauf et al.$
Ophioceraceae Klaubauf et al.*Ophioceras Sacc.*$
Pyriculariaceae Klaubauf et al.Bambusicularia Klaubauf et al.*$
Barretomyces Klaubauf et al.*Deightoniella S. Hughes*
= Utrechtiana Crous & Quaedvl.*$
Macgarvieomyces Klaubauf et al.*Neopyricularia Klaubauf et al.*Proxipyricularia Klaubauf et al.*Pseudopyricularia Klaubauf et al.*$
Pyricularia Sacc.*$
Xenopyricularia Klaubauf et al.*
Fungal Diversity

Magnaporthales, genera incertae sedisPseudohalonectria Minoura & T. Muroi$
Ophiostomatales Benny & Kimbr.Kathistaceae Malloch & M. Blackw.
Kathistes Malloch & M. Blackw.Mattirolella S. Colla#
Termitariopsis M. Blackw. et al.#
Ophiostomataceae Nannf.Ceratocystiopsis H.P. Upadhyay & W.B. Kendr.$
Fragosphaeria Shear$
Hyalobelemnospora Matsush.#
Hyalorhinocladiella H.P. Upadhyay & W.B. Kendr.Klasterskya Petr.#
Leptographium Lagerb. & MelinOphiostoma Syd. & P. Syd.$
Pesotum JL Crane & Schokn.Phialographium H.P. Upadhyay & W.B. Kendr.Raffaelea Arx & Hennebert$
Spumatoria Massee & E.S. Salmon#
Subbaromyces Hesselt.
Ophiostiomatales, genera incertae sedisLanspora K.D. Hyde & E.B.G. Jones*
Togniniales I.C. Senanayake et al.*Togniniaceae Réblová et al.
Conidiotheca Réblová & L. Mostert#
Phaeoacremonium W. Gams et al.*$
Togninia Berl.$
= Romellia Berl.
Trichosphaeriales M.E. BarrTrichosphaeriaceae G. Winter
Acanthosphaeria Kirschst.#
Brachysporium Sacc.#
Collematospora Jeng & Cain#
Coniobrevicolla Réblová#
CresporhaphisM.B. Aguirre#
Cryptadelphia Réblová & Seifert$
Eriosphaeria Sacc.#
Fluviostroma Samuels & E. Müll.#
Kananascus Nag RajKhuskia H.J. Huds.Koorchaloma Subram.Neorehmia Höhn.#
Oplothecium Syd.#
Rizalia Syd. & P. Syd.#
Schweinitziella Speg.#
Setocampanula Sivan. & W.H. Hsieh#
Trichosphaeria FuckelUnisetosphaeria Pinnoi et al.#
Diaportheomycetidae, families incertae sedisPapulosaceae Winka & O.E. Erikss.
Brunneosporella V.M. Ranghoo & K.D. Hyde#
Fluminicola S.W. Wong et al.#
Papulosa Kohlm & Volkm-Kohlm*$
Thyridiaceae O.E. Erikss & J.Z. YueMattirolia Berl. & Bres.#
= Balzania Speg.*#
=Thyronectroidea Seaver#
Pleurocytospora Petr.#
Thyridium Nitschke$
Diaportheomycetidae, genera incertae sedisPlatytrachelon Réblová*
Subclass Hypocreomycetidae O.E. Erikss. & Winka*$
Coronophorales Nannf.*Bertiaceae Smyk*
Bertia De Not.$
Gaillardiella Pat.*$
Chaetosphaerellaceae Huhndorf et al.*Chaetosphaerella E. Müll. & C. Booth$
Crassochaeta Réblová$
Oedemium Link*#
= Veramycina Subram.Spinulosphaeria Sivan.$
Coronophoraceae Höhn.*Coronophora Fuckel$
Nitschkiaceae (Fitzp) Nannf.*Acanthonitschkea Speg.$
Biciliosporina Subram. & Sekar#
Botryola Bat. & J.L. Bezerra#
Fracchiaea Sacc.$
Groenhiella Jørg. Koch et al.*#
Janannfeldtia Subram. & Sekar#
Lasiosphaeriopsis D. Hawksw. & Sivan.#
Loranitschkia Lar.N. Vasiljeva#
Neochaetosphaerella Lar.N. Vassiljeva et al.*#
Neotrotteria Sacc.Nitschkia G.H. Otth ex P. Karst.$
Rhagadostoma Körb.#
Rhagadostomella Etayo#
Tortulomyces Lar.N. Vassiljeva et al.*#
Fungal Diversity

Scortechiniaceae Huhndorf et al.*Biciliospora Petr.*Coronophorella Höhn.Cryptosphaerella Sacc.*$
Euacanthe Theiss.Neofracchiaea Teng$
Scortechinia Sacc.$
Scortechiniella Arx & E. Müll.Scortechiniellopsis Sivan.$
Tympanopsis Starbäck$
Coronophorales, genera incertae sedisPseudocatenomycopsis Crous & L.A. Shuttlew.*$
Falcocladiales R.H. Perera et al.*Falcocladiaceae Somrithipol et al.*
Falcocladium S.F. Silveira et al.*$
Glomerellales Chadef. ex Réblová et al.Australiascaceae Réblová & W. Gams*
Australiasca Sivan. & Alcorn*$
Hyalocylindrophora J.L. Crane & Dumont#
Monilochaetes Halst. ex Harter*$
= Dischloridium B. Sutton
Glomerellaceae Locq. ex Seifert & W. GamsColletotrichum Corda*$
= Glomerella Spauld. & Schrenk
Plectosphaerellaceae W. Gams et al.*Acrostalagmus Corda$
Gibellulopsis Bat. & H. Maia*$
Lectera P.F. Cannon*$
Musicillium Zare & W Gams*$
Plectosphaerella Kleb.*$
= Plectosporium M.E. Palm et al.= Spermosporina U. Braun
Stachylidium Link*$
Verticillium Nees*$
Reticulascaceae Réblová & W. Gams*Cylindrotrichum Bonord.*
= Reticulascus Réblová & W. Gams*$
Kylindria DiCosmo et al.$
Sporoschismopsis Hol-Jech. & Hennebert*= Porosphaerellopsis Samuels & E. Müll.$
Glomerellales, genera, incertae sedisAscocodinaea Samuels et al.
Hypocreales Lindau*Bionectriaceae Samuels & Rossman*
= Spicariaceae Nann.
Acremonium Link*Anthonectria Döbbeler*#
Aphanotria Döbbeler#
Battarrina (Sacc.) Clem. & Shear#
Clonostachys Corda*= Bionectria Speg.$
Bryocentria Döbbeler*Clibanites (P. Karst.) P. Karst.#
Didymostilbe Henn*Dimerosporiella Speg.#
Gliomastix Guég.*Globonectria Etayo#
Gracilistilbella Seifert#
Halonectria E.B.G. Jones*#
Heleococcum P.M. Jørg.*$
Hydropisphaera Dumort*$
Ijuhya Starbäck*Lasionectria (Sacc) CookeMycocitrus Möller#
Nectriella Nitschke ex Fuckel#
Nectriopsis MaireOchronectria Rossman & SamuelsOvicuculospora EtayoParanectria Sacc.#
Peristomialis (W. Phillips) Boud.Pronectria Clem.#
Protocreopsis Yoshim Doi*$
Rhopalocladium Schroers et al.#
Roumegueriella Speg.$
Selinia P. Karst.Spicellum Nicot & RoquebertStephanonectria Schroers & SamuelsStilbocrea Pat.Stromatocrea W.B. Cooke#
Stromatonectria Jaklitsch & H. Voglmayr*Trichonectria Kirschst.Vesicladiella Crous & M.J. Wingf.Verrucostoma Hirooka et al.*Virgatospora Finley
Clavicipitaceae (Lindau) Earle ex RogersonAciculosporium I. Miyake
= Albomyces I. Miyake#
= Mitosporium Clem. & ShearAmphichorda Fr.#
Atkinsonella Diehl.Balansia Speg.
= Dothichloë G.F. Atk.Cavimalum Yoshim. Doi et al.#
Chamaeleomyces SiglerClaviceps Tul.$
Conoideocrella D. Johnson et al.Corallocytostroma Y.N. Yu & Z.Y. Zhang
Fungal Diversity

Diploöspora Grove#
Dussiella Pat.Ephelis Fr.Epichloë (Fr.) Tul. & C. Tul.$
Epicrea Petr.#
Helminthascus Tranzschel#
Heteroepichloë E. Tanaka et al.Hypocrella Sacc.$
= Aschersonia Mont.Konradia Racib.#
Loculistroma F. Patt & Charles#
Metacordyceps G.H. Sung et al.Metarhiziopsis D.W. Li et al.Metarhizium SorokīnMetapochonia Kepler et al.*Moelleriella Bres.Mycomalus Möller#
Myriogenospora G.F. Atk.Neobarya LowenNeoclaviceps J. White et al.Neocordyceps Kobayasi#
Neotyphodium Glenn et al.Nigrocornus Ryley & LangdonNomuraea Maubl.Orbiocrella D. Johnson et al.Parepichloë F.J. White Jr. & ReddyPeriglandula U. Steineret al.*Pochonia Bat. & O.M. FonsecaPseudogibellula Samson & H.C. EvansPseudomeria G.L. Barron#
Regiocrella Chaverri & K.T. Hodge*Romanoa Thirum.#
Rotiferophthora G.L. BarronSamuelsia Chaverri & K.T. Hodge*Shimizuomyces Kobayasi$
Sphacelia Lév.Sphaerocordyceps Kobayasi#
Stereocrea Syd. & P. Syd.#
Tyrannicordyceps Kepler & SpataforaUstilaginoidea Bref.
= Villosiclava E. Tanaka & C. Tanaka
Cordycipitaceae Kreisel ex G.M. Sung et al.Akanthomyces LebertAscopolyporus Möller$
Beauveria Vuill.*Cordyceps (Fr.) Link$
Gibellula Cavara$
Granulomanus de Hoog & Samson#
Hyperdermium J. White et al.$
Isaria Pers.Lecanicillium W. Gams & ZareMicrohilum H.Y. Yip & A.C. Rath
Simplicillium W. Gams & ZareSyspastospora P.F. Cannon & D. Hawksw.Torrubiella Boud.$
Hypocreaceae De Not.= Trichodermataceae Fr.
Aphysiostroma Barrasa et al.Arachnocrea Z. Moravec.Cladobotryum NeesDialhypocrea Speg.#
Escovopsis J.J. Muchovej & Della Lucia*Escovopsioides H.C. Evans & J.O. Augustin*Hypocreopsis P. Karst.*Hypomyces (Fr.) Tul.*Mycogone LinkPayoshaeria W.F. Leong*Protocrea PetchPseudohypocrea Yoshim. Doi#
Rogersonia Samuels & Lodge#
Sepedonium LinkSibirina G.R.W. Arnold#
Sphaerostilbella (Henn.) Sacc. & D. Sacc*$
= Gliocladium CordaSporophagomyces K. Põldmaa & SamuelsStephanoma WallrTrichoderma Pers.*$
= Hypocrea Fr.$
= Sarawakus Lloyd
Nectriaceae Tul. & C. Tul.*= Tuberculariaceae Fr.
Actinostilbe Petch*= Lanatonectria Samuels & Rossman
Albonectria Rossman & SamuelsAllantonectria Earle*Allonectella Petr.#
Antipodium Piroz.#
Atractium Link*Aquanectria L. Lombard & Crous*Baipadisphaeria Pinruan*$
Bisifusarium L. Lombard et al.*Calonectria De Not.*
= Cylindrocladium MorganCalostilbe Sacc. & Syd.#
= Calostilbella Höhn.Campylocarpon Halleen et al.Chaetonectrioides Matsush.Chaetopsina Rambelli$
Chaetopsinectria J Luo & WY ZhuangCoccinonectria Lombard & CrousCorallomycetella Henn.*Corallonectria C. Herrera & P. Chaverri*
Fungal Diversity

Cosmospora Rabenh.*$
Curvicladiella Decock & Crous#
Curvicladium Decock & CrousCyanochyta Höhn.#
Cyanonectria Samuels & ChaverriCyanophomella Höhn.#
Cylindrocladiella Boesew.*= Nectricladiella Crous & C.L. Schoch
Cylindrodendrum Bonord.*Dacryoma Samuels#
Dematiocladium Allegr. et al.Dialonectria (Sacc.) Cooke*Fusarium Link *
= Gibberella Sacc.Fusicolla Bonord*Geejayessia Schroers et al.*Gliocephalotrichum J.J. Ellis & Hesselt.*
= Leuconectria Rossman et al.Gliocladiopsis S.B. Saksena*Glionectria Crous & C.L. SchochHeliscus Sacc*Ilyonectria P. Chaverri & C. Salgado*Macroconia (Wollenw.) Gräfenhan et al.*Mariannaea G. Arnaud ex Samson*Microcera Desm.Nalanthamala Subram.*
= Rubrinectria Rossman & SamuelsNectria (Fr.) Fr.*$
= Tubercularia TodeNeonectria Wollenw.*$
= Cylindrocarpon Wollenw.Neocosmospora E.F. Sm.*
= Haematonectria Samuels & NirenbergOphionectria Sacc.*Paracremonium L. Lombard & Crous*Penicillifer EmdenPersiciospora P.F. Cannon & D. Hawksw.Pleogibberella Sacc.#
Pleonectria Sacc.*= Zythiostroma Höhn ex Falck
Pleurocolla Petr.Pseudocosmospora C. Herrera & P. ChaverriPseudonectria SeaverRectifusarium Lombard et al.Rodentomyces Doveri et al.*Rugonectria P. Chaverri & Samuels*Stachybotryna Tubaki & T. Yokoy#
Stalagmites Theiss. & Syd.#
Stylonectria Höhn.Thelonectria P. Chaverri & C.G. Salgado*Thyronectria Sacc.Viridispora Samuels & Rossman$
Volutella Fr.
= Chaetodochium Höhn.#
Volutellonectria J. Luo & W.Y. Zhuang*Xenoacremonium Lombard & CrousXenocalonectria Crous & C.L. Schoch
= Xenocylindrocladium Decock et al.XenonectriellaWeese#
Niessliaceae Kirschst.Atronectria Etayo#
Circinoniesslia Samuels & M.E. Barr#
Cryptoniesslia Scheuer#
Eucasphaeria Crous*Hyaloseta A.W. Ramaley#
Malmeomyces Starb.#
Melchioria Penz. & Sacc.#
Miyakeomyces Hara#
Myrmaeciella Lindau*#
Niesslia Auersw.*$
Paraniesslia K.M. Tsui et al.#
Pseudonectriella Petr.#
Pseudorhynchia Höhn.#
Rosasphaeria Jaklitsch & Voglmayr*Taiwanascus Sivan. & H.S. Chang#
Trichosphaerella E. Bommer et al.#
Valetoniella Höhn.#
Ophiocordycipitaceae G.H. Sung et al.DrechmeriaW. Gams & H.B. Jansson
= Haptocillium W. Gams & ZareHarposporium Lohde
= Atricordyceps Samuels= Podocrella Seaver= Polyrhina Sorokin
Ophiocordyceps Petch$
= Cordycepioideus Stifler= Didymobotryopsis Henn= Didymobotrys Clem. & Shear= Hirsutella Pat= Hymenostilbe Petch$
= Mahevia Lagarde= Paraisaria Samson & B.L. Brady= Sorosporella Sorokin= Syngliocladium Petch#
= Synnematium Speare= Trichosterigma Petch= Troglobiomyces Pacioni
Polycephalomyces Kobayasi= Blistum B. Sutton
Purpureocillium Luangsa-ard et al.*Tolypocladium W. Gams
= Chaunopycnis W. Gams= Elaphocordyceps G.H. Sung & Spatafora$
Fungal Diversity

Stachybotriaceae L. Lombard & CrousAlbosynnema E.F. Morris$
Myrothecium Tode*$
Parasarcopodium Melnik et al.$
Peethambara Subram. & Bhat*$
Sarcopodium Ehrenb.Scopinella Lév.*$
Stachybotrys Corda*$
= Memnoniella Höhn.= Ornatispora K.D. Hyde et al.= Melanopsamma Niessl.= Valsonectria Speg.
Tilachlidiaceae Lombard & CrousSeptofusidium W. Gams$
Tilachlidium Preuss$
Hypocreales, genera incertae sedisBerkelella (Sacc.) Sacc.#
Bulbithecium Udagawa & T MuroiEmericellopsis JFH Beyma*Fecundostilbum T.P. Devi & Chowdhry#
Geosmithia J. PittGynonectria Döbbeler*#
Hapsidospora Malloch & CainHaptospora G.L. Barron#
Harzia CostantinIllosporiopsis D. Hawksw.#
Illosporium Mart.Kallichroma Kohlm. & Volkm-Kohlm.*Leucosphaerina ArxMetadothella Henn.#
Munkia Speg.#
Mycoarachis Malloch & CainNeomunkia PetrNigrosabulum Malloch & CainPayosphaeria W.F. Leong*#
Peloronectria Möller#
Pseudoidriella Crous & R.G. Shivas*Pseudomeliola Speg.#
Pseudomicrodochium B. Sutton#
Roselliniella Vain*Sarocladium W. Gams & D. Hawksw.*Sedecimiella K.L. Pang et al.*Septomyrothecium Matsush.#
= Sporothrix Hektoen & C.F. PerkinsStanjemonium W. Gams et al.Stilbella LindauTiconectria Döbbeler#
Tilakidium Vaidya et al.#
Trichothecium LinkValetoniellopsis Samuels & M.E. Barr
Melanosporales N. Zhang & M. Blackw.Ceratostomataceae G. Winter*
Acrospeira Berk & Broome#
Arxiomyces P.F. Cannon & D. Hawksw.#
Erythrocarpon Zukal#
Gonatobotrys Corda#
Melanospora Corda$
= Proteophiala Cif.Pteridiosperma J.C. Krug & Jeng#
Pustulipora P.F. Cannon#
Rhytidospora Jeng & Cain#
Setiferotheca Matsush.#
Vittatispora P. Chaudhary et al.$
Melanosporales genera, incertae sedisSphaerodes Clem.*Papulaspora Preuss
Microascales Luttr. ex Benny & Kimbr.= Halosphaeriales Kohlm.
Chadefaudiellaceae Faurel & Schotter ex Benny & Kimbr.Chadefaudiella Faurel & Schotter#
Faurelina Locq-Lin.
Ceratocystidaceae Locq. ex Réblova et al.*Ambrosiella Brader ex Arx & Hennebert$
Ceratocystis Ellis & Halst.$
Chalaropsis PeyronelDavidsoniella Z.W. de Beer et al.Endoconidiophora MünchGrosmannia Goid.Huntiella Z.W. de Beer et al.Thielaviopsis Went.$
Gondwanamycetaceae Réblová et al.*Custingophora Stolk et al.*$
Gondwanamyces Marais & M.J. Wingf.*$
Graphiaceae De BeerGraphium Corda$
Halosphaeriaceae E. Müll & Arx ex Kohlm.Alisea J. Dupont & E.B.G. Jones*Aniptodera Shearer & M. Miller*Anisostagma K.R.L. Petersen & Jørg. Koch*#
Antennospora Meyers*Appendichordella R.G. Johnson et al.*#
Arenariomyces Höhnk*Ascosacculus J J Campb., J.L. Anderson & Shearer*Bathyascus Kohlm.*#
Carbosphaerella I. Schmidt*Ceriosporopsis Linder*
Fungal Diversity

= Bovicornua Jørg Koch & E.B.G. JonesChadefaudia Feldm.-Maz.#
Cirrenalia Meyers & R.T. Moore*Clavatospora Sv Nilsson ex Marvanová & Sv Nilsson*Corallicola Volkm.-Kohlm. & Kohlm.*#
Corollospora Werderm*= Halosigmoidea Nakagiri et al.
Cucullosporella K.D. Hyde & E.B.G. Jones*Ebullia K.L. Pang*Fluviatispora K.D. Hyde*#
Gesasha Abdel-Wahab & Nagah.*Haiyanga K.L. Pang & E.B.G. Jones*Haligena Kohlm.*Halosarpheia Kohlm. & E. Kohlm.*Halosphaeria Linder*$
Halosphaeriopsis T.W. Johnson*$
= Culcitalna Meyers & R.T. MooreHavispora K.L. Pang & VrijmoedIwilsoniella EBG Jones*#
Kitesporella Jheng & K.L. Pang*Kochiella Sakay. et al.*Lautisporiopsis E.B.G. Jones et al.Lignincola Höhnk*$
Limacospora Jørg. Koch & E.B.G. Jones*#
Luttrellia Shearer*#
Magnisphaera J. Campb. et al.*= Matsusphaeria K.L. Pang & E.B.G. Jones
Marinospora A.R. Caval.*= Ceriosporella (Kohlm.) A.R. Caval.
Moana Kohlm. & Volkm.-Kohlm.*#
Morakotiella Sakay.*Nais Kohlm.*Natantispora J. Campb. et al.r*Naufragella Kohlm. & Volkm.-Kohlm.*Nautosphaeria E.B.G. Jones*Neptunella K.L. Pang & E.B.G. Jones*Nereiospora EBG Jones et al.*Nimbospora Jørg. Koch*$
Nohea Kohlm. & Volkm.-Kohlm.*Oceanitis Kohlm.*
= Ascosalsum J. Campb. et al.= Falcatispora K.L. Pang & E.B.G. Jones
Ocostaspora E.B.G. Jones et al.*Okeanomyces K.L. Pang & E.B.G. Jones*Ondiniella E.B.G. Jones et al.*$
Ophiodeira Kohlm. & Volkm.-Kohlm.*Panorbis J. Campb. et al. *Phaeonectriella Eaton & E.B.G. Jones*Pileomyces K.L. Pang & Jheng*Pseudolignincola Chatmala & E.B.G. Jones#
Remispora Linder*$
Saagaromyces K.L. Pang & E.B.G. Jones*= Littispora J. Campb. et al.
Sablicola E B.G. Jones et al.*Thalassogena Kohlm. & Volkm.-Kohlm.*Thalespora Chatmala & E.B.G. Jones*Tirispora E.B.G. Jones & Vrijmoed*Toriella Sakay. et al.*Trailia G.K. Sutherl.*#
Trichomaris Hibbits et al.*#
Tubakiella Sakay. et al.*Tunicatispora K.D. Hyde*#
= Buxetroldia K.R.L. Petersen & Jørg KochVaricosporina Meyers & Kohlm.*
Microascaceae Luttr. ex MallochAnekabeeja Udaiyan & Hosag.#
Brachyconidiellopsis Decock et al.Canariomyces ArxCephalotrichum Link#
Doratomyces Corda$
Echinobotryum Corda#
Enterocarpus Locq.-Lin.Kernia Nieuwl.Knoxdaviesia M.J. Wingf. et al.Lophotrichus R.K. Benj.Microascus Zukal$
Parascedosporium Gilgado et al.*Petriella Curzi$
Pseudallescheria Negroni & I. Fisch.= Petriellopsis Gilgado et al.
Scedosporium Sacc. ex Castell. & Chalm.Scopulariopsis BainierWardomyces F.T. Brooks & Hansf.Wardomycopsis Udagawa & Furuya
Microascales, genera incertae sedisBisporostilbella Brandsb. & E.F. Morris#
Cornuvesica C.D. Viljoen et al.Gabarnaudia Samson & W. GamsSphaeronaemella P. Karst.Sporendocladia G. Arnaud ex Nag Raj & W.B. Kendr.Trichurus Clem.Viennotidia Negru & Verona ex Rogerson#
Vermiculariopsiella Bender
Savoryellales Boonyuen et al.*Savoryellaceae Jaklitsch & Réblová
Ascotaiwania Sivan. & H.S. Chang*$
Canalisporium Nawawi & Kuthub.*$
= Ascothailandia Sri-indr. et al.*$
Savoryella E.B.G. Jones & R.A. Eaton*$
Savoryellales, genera incertae sedisCarpoligna F.A. Fernández & Huhndorf$
Conioscypha Höhn.$*= Conioscyphascus Réblová & Seifert$
Fungal Diversity

Flammispora Pinruan et al.Sterigmatobotrys Oudem.*
Hypocreomycetidae, families incertae sedisEtheirophoraceae Rungjindamai et al.*
Etheirophora Kohlm. & Volkm.-Kohlm.*$
Swampomyces Kohlm. & Volkm.*$
Juncigenaceae E.B.G. Jones et al.*Juncigena Kohlm et al.*$
Fulvocentrum E.B.G. Jones & Abdel-Wahab*Marinokulati E.B.G. Jones & K.L. Pang*$
Moheitospora Abdel-Wahab et al.*$
Torpedosporaceae E.B.G. Jones & K.L. Pang*Glomerulispora Abdel-Wahab & Nagah.*$
Torpedospora Meyers*$
Hypocreomycetidae, genera incertae sedisMyrmecridium Arzanlou et al.Pleurotheciella Réblová et al.* $
Pleurothecium Höhn.*$
Subclass Lulworthiomycetidae Dayarathne et al.
Koralionastetales Kohlm. et al.Koralionastetaceae Kohlm. & Volkm.-Kohlm.
Koralionastes Kohlm. & Volkm.-Kohlm.$
Lulworthiales Kohlm. et al.Lulworthiaceae Kohlm. et al.
Cumulospora I. Schmidt*$
Equicapillimyces S.S.Y. Wong et al.Halazoon Abdel-Aziz et al.*Haloguignardia A. Cribb & J. Cribb*Hydea K.L. Pang & E.B.G Jones*$
Kohlmeyeriella E B G Jones et al.*Lindra I. Wilson*$
Lulwoana Kohlm. et al.*Lulwoidea Kohlm. et al.*Lulworthia G.K. Sutherl*$
Matsusporium E.B.G. Jones & K.L. Pang*Moleospora Abdel-Wahab et al.*Moromyces Abdel-Wahab et al.*Orbimyces Linder*Rostrupiella Jørg Koch et al.*
Subclass Meliolomycetidae P.M. Kirk & K.D. Hyde*
Meliolales Gäum. ex D. Hawksw. & O.E. Erikss.#
Armatellaceae Hosag.Armatella Theiss. & Syd.#
Meliolaceae G.W. Martin ex Hansf.Amazonia Theiss.#
Appendiculella Höhn.
AsteridiellaMcAlpine$
Basavamyces V.B. Hosag.#
Ceratospermopsis Bat.#
Cryptomeliola S. Hughes & Piroz.Ectendomeliola Hosag. & D.K. Agarwal#
Endomeliola S. Hughes & Piroz.$
Haraea Sacc. & P. Syd.#
Hypasteridium Speg.#
Irenopsis F. Stevens$
Laeviomeliola Bat.#
Leptascospora Speg.#
Meliola Fr.$
Metasteridium Speg.#
Ophiociliomyces Bat. & I.H. Lima#
Ophioirenina Sawada & W. Yamam.#
Ophiomeliola Starb.#
Parasteridium Speg.#
Pauahia F. Stevens#
Pleomeliola (Sacc.) Sacc.#
Pleomerium Speg.#
Prataprajella Hosag.#
Ticomyces Toro#
= Dontuzia L.D. GomezUrupe Viégas#
Xenostigme Syd.#
Subclass Sordariomycetidae O.E. Erikss & Winka
Boliniales P.F. CannonBoliniaceae Rick*
Apiocamarops Samuels & J.D. Rogers#
Apiorhynchostoma Petr.$
Camaropella Lar.N. Vassiljeva$
Camarops P. Karst.*$
Cornipulvina Huhndorf et al.$
Endoxyla FuckelMollicamarops Lar.N. Vassiljeva#
Pseudovalsaria Spooner
Chaetosphaeriales Huhndorf et al.Chaetosphaeriaceae Réblová et al.
Ascochalara Réblová#
Brunneodinemasporium Crous & R.F. Castañeda*Catenularia Grove#
Chaetosphaeria Tul. & C. Tul.*$
Chloridium LinkCodinaea MaireCodinaeopsisMorgan-JonesCraspedodidymum Hol-Jech.#
Cryptophiale Piroz.#
Dendrophoma Sacc.Dinemasporium Lév.*Dictyochaeta Speg.
Fungal Diversity

Dictyochaetopsis Aramb. & Cabello#
Exserticlava S. Hughes$
Gonytrichum Nees & T. Nees#
Hemicorynespora M.B. Ellis#
Kionochaeta P.M. Kirk & B. SuttonInfundibulomyces Plaingam et al.*Lecythothecium Réblová & WinkaMelanopsammella Höhn.Menispora Pers.*PhaeostalagmusW. Gams#
Phialogeniculata Matsush.#
Pseudobotrytis Krzemien. & BaduraPseudolachnea Ranoj.Pyrigemmula D. Magyar & R. Shoemaker*Rattania Prabhug. & Bhat$
Miyoshiella Kawam.#
Sporoschisma Berk. & Broome*= Melanochaeta E. Müll. et al.$
Striatosphaeria Samuels & E. Müll.Tainosphaeria F.A. Fernández & HuhndorfThozetella KuntzeUmbrinosphaeria Réblová$
Zanclospora S. Hughes & W.B. Kendr.#
Zignoëlla Sacc.
Helminthosphaeriaceae Samuels et al.Echinosphaeria A.N. Mill. & Huhndorf*$
Endophragmiella B. Sutton#
Helminthosphaeria Fuckel*$
Hilberina Huhndorf & A.N. Mill.*Ruzenia O. Hilber*$
Synaptospora Cain*$
Tengiomyces Réblová#
Pleurotremataceae Walt. WatsonPleurotrema Müll. Arg.*#
Chaetosphaeriales genera incertae sedisCaudatispora J. Fröhl. & K.D. HydeErythromada Huhndorf et al.Lasiosphaeriella Sivan.*Leptosporella Penz. & Sacc.*Nawawia MarvanováRimaconus Huhndorf et al.*$
Phyllachorales M.E. Barr*Phaeochoraceae K.D. Hyde et al.
Cocoicola K.D. Hyde#
Phaeochora Höhn.#
Phaeochoropsis K.D Hyde & P.F. Cannon#
Serenomyces Petr.*
Phyllachoraceae Theiss. & H. Syd.
Acerviclypeatus Hanlin*#
Apiosphaeria Höhn.#
Ascovaginospora Fallah et al.$
Brobdingnagia K.D. Hyde & P.F. Cannon#
Camarotella Theiss. & Syd.#
Coccodiella Hara$
Cyclodomus Höhn.#
Deshpandiella Kamat & Ullasa#
Diachora Müll. Arg.#
= Diachorella Höhn.Diatractium Syd. & P. Syd.Erikssonia Penz. & Sacc.#
Fremitomyces P.F. Cannon & H.C. Evans#
Geminispora Pat.#
Gibellina Pass. ex Roum.#
Imazekia Tak. Kobay. & Y. Kawabe#
Isothea Fr.#
Lichenochora Hafellner#
Lindauella Rehm#
Linochora Höhn.#
Lohwagia Petr.#
Maculatifrondes K.D. Hyde#
Malthomyces K.D. Hyde & P.F. Cannon#
Muelleromyces Kamat & Anahosur#
Mycohypallage B. Sutton#
Neoflageoletia J. Reid & C. Booth#
Ophiodothella (Henn.). Höhn.*Orphnodactylis Malloch & Mallik#
Oswaldina Rangel#
Oxodeora K.D. Hyde & P.F. Cannon#
Parberya C.A. Pearce & K.D. Hyde#
Petrakiella Syd.#
Phaeochorella Theiss. & Syd.#
Phycomelaina Kohlm.#
Phyllachora Nitschke ex Fuckel$
Phylleutypa Petr.#
Phyllocrea Höhn.#
Polystigma DC.= Polystigmina Sacc.#
Pseudothiella Petr.#
Pseudothiopsella Petr.#
Pterosporidium W.H. Ho & K.D. Hyde#
Rehmiodothis Theiss. & Syd.#
Retroa P.F. Cannon#
Rhodosticta Woron.#
Rikatlia P.F. Cannon#
Schizochora Syd. & P. Syd.#
Sphaerodothella C.A. Pearce & K.D. Hyde#
Sphaerodothis (Sacc. & P. Syd.) ShearStigmatula (Sacc.) Syd. & P. Syd.#
Stigmochora Theiss. & Syd.#
Stromaster Höhn.#
Telimena Racib.#
Fungal Diversity

Telimenella Petr.#
Telimenochora Sivan.#
Trabutia Sacc. & Roum.#
Tribulatia J.E. Taylor et al.#
Uropolystigma Maubl.#
Vitreostroma P.F. Cannon#
Zimmermanniella Henn.#
Phyllachorales genera incertae sedisMarinosphaera K.D. Hyde*#
Sordariales Chad. ex D. Hawksw. & O.E. Erikss.Chaetomiaceae G. Winter
Achaetomium J.N. Rai et al.$
Bommerella Marchal#
Boothiella Lodhi & Mirza#
Botryotrichum Sacc. & MarchalChaetomidium (Zopf) Sacc.$
Corynascella Arx & Hodges$
Chaetomiopsis Mustafa & Abdul-Wahid#
Chaetomium Kunze$
Diplogelasinospora CainEmilmuelleria ArxFarrowia D. Hawksw.GuanomycesM.C. Gonzáles et al.Humicola Traaen*Madurella BrumptMyceliophthora Costantin*
= Corynascus ArxStaphylotrichum J.A. Mey. & Nicot*Subramaniula Arx#
Taifanglania Z.Q. Liang et al.Thielavia ZopfZopfiella G. Winter*
Lasiosphaeriaceae Nannf.*Angulimaya Subram & Lodha#
Anopodium Lundq.*Apiosordaria Arx & W. Gams$
Apodospora Cain & J.H. Mirza*Apodus Malloch & CainArniella Jeng & J.C. Krug#
Arnium Nitschke ex G. Winter*Bellojisia RéblováBiconiosporella Schaumann*#
Bombardia (Fr.) P. Karst.$
Bombardioidea C. Moreau ex N. Lundqv.Camptosphaeria Fuckel#
Cercophora FuckelCladorrhinum Sacc. & MarchalDiffractella Guarro et al.#
Emblemospora Jeng & J.C. Krug#
Eosphaeria Höhn.#
Fimetariella N. Lundq.*Immersiella A.N. Mill. & HuhndorfJugulospora N. Lundq.$
Lacunospora Cailleux#
Lasiosphaeria Ces. & De Not.$
Mammaria Ces. ex Rabenh.Melanocarpus ArxPeriamphispora J.C. Krug#
Podospora Ces.Pseudocercophora Subram. & Sekar#
Schizothecium CordaStrattonia Cif.Thaxteria Sacc.#
Triangularia BoedijnTripterosporella Subram. & Lodha#
Zygopleurage Boedijn$
Zygospermella Cain*
Sordariaceae G. WinterCopromyces N. Lundq.Effetia Bartoli et al.*#
Gelasinospora Dowding$
Guilliermondia Boud.#
Neurospora Shear & B.O. Dodge*$
= Chrysonilia ArxPseudoneurospora Dania García et al.$
Sordaria Ces & De Not. $
Stellatospora T. Ito & A. Nakagiri#
Sordariales, genera incertae sedisAbyssomyces Kohlm*#
Acanthotheciella Höhn.#
Ascolacicola Ranghoo & K.D. HydeAsterosporium KunzeBombardiella Höhn.#
Cancellidium TubakiCoronatomyces Dania García et al.Corylomyces Stchigel et al.Cuspidatispora A. Mill.Globosphaeria D. Hawksw.#
Isia D. Hawksw & Manohar#
Lasiosphaeris Clem.= Lasiadelphia Réblová & W Gams*
Lockerbia K.D. Hyde#
Nitschkiopsis Nannf. & R. Sant.#
Onygenopsis Henn.#
Phaeosporis Clem.#
Ramophialophora M. Calduch et al.Reconditella Matzer & Hafellner#
Rhexodenticula W.A. Baker & Morgan-JonesRhexosporium Udagawa & Furuya#
Roselliniomyces Matzer & Hafellner#
Roselliniopsis Matzer & Hafellner#
Fungal Diversity

Sporidesmiopsis Subram. & BhatUtriascus Réblova#
Ypsilonia Lév.#
Sordariomycetidae, families incertae sedis
Batistiaceae Samuels & K.F. Rodrigues$
Acrostroma Seifert#
Batistia Cif.$
Cephalothecaceae Höhn.*Albertiniella Kirschst.$
Cephalotheca Fuckel$
Cryptendoxyla Malloch & Cain$
Phialemonium W. Gams & McGinnis$
Sordariomycetidae, genera incertae sedisArecacicola Joanne E. Taylor et al.#
Barbatosphaeria Réblová#
Bullimyces A. Ferrer et al.*Brachysporiella Bat.Ceratolenta Réblová*Ceratostomella Sacc.ChaetosphaeridesMatsush.#
Conlarium F. Liu & L. Cai*Garethjonesia K.D. Hyde#
Hanliniomyces Raja & Shearer#
Hydromelitis A. Ferrer et al.*Lentomitella Höhn.Mirannulata Huhndorf et al.Menisporopascus Matsush.*#
Merugia Rogerson & Samuels#
Mycomedusiospora G.C. Carroll & Munk#
Myelosperma Syd. & P. Syd.#
Nigromammilla K.D. Hyde & J. Fröhl.#
Phaeotrichosphaeria Sivan.#
Phragmodiscus Hansf.#
Plagiosphaera Petr.#
Rhodoveronaea Arzanlou et al.Riomyces A. Ferrer et al.Spadicoides S. Hughes*Xylomelasma RéblováWoswasia Jaklitsch et al.
Subclass Xylariomycetidae O.E. Erikss & Winka
Xylariales Nannf.Apiosporaceae K.D. Hyde et al.*$
Appendicospora K.D. HydeArthrinium Kunze*$
= Apiospora Sacc.$
= Cordella Speg.= Pteroconium Sacc. ex Grove
Dictyoarthrinium S. Hughes#
Endocalyx Berk. & Broome#
Scyphospora LA Kantsch#
Spegazzinia Sacc.
Amphisphaeriaceae G. WinterAdisciso Kaz. TanakaAmphisphaerella (Sacc.) Kirschst.#
Amphisphaeria Ces & De Not.*$
Atrotorquata Kohlm. & Volkm.-Kohlm.Bartalinia Tassi*Bleptosporium Steyaert*#
Broomella Sacc*Capsulospora K.D. HydeCeratosporium Schwein.#
Ceriophora Höhn.#
Ceriospora Niessl#
Chitonospora E. Bommer et al.#
Clypeophysalospora H.J. Swart#
Discosia Lib.*Discostroma Clem.*$
Distorimula San Martín et al.#
Doliomyces Steyaert#
Dyrithium M.E. Barr#
Dyrithiopsis L. Cai et al.= Monochaetiopsis L. Cai et al.*
Flagellosphaeria Aptroot#
Frondispora K.D. Hyde#
Funiliomyces Aptroot*Griphosphaerioma Höhn.*#
= Labridella BrenckleHyalotiopsis Punith.*
= Ellurema Nag Raj & W.B. Kendr.Immersidiscosia Kaz. Tanaka et al.*$
Iodosphaeria Samuels et al.#
Labridium Vestergr.Lindquistomyces Aramb. et al.#
Manokwaria K.D. Hyde#
Microdochium Syd.Monochaetia (Sacc) Allesch.*MonochaetinulaMuthumary et al.*#
Monographella Petr.$
Morinia Berl. & Bres.*Mukhakesa Udaiyan & Hosag.#
Neobroomella Petr.#
Neohypodiscus J.D. Rogers et al.#
Neopestalotiopsis Maharachch. et al.*Ommatomyces Kohlm et al.#
Paracainiella Lar.N. Vassiljeva#
Pemphidium Mont.#
Pestalotiopsis Steyaert*= PestalosphaeriaM.E. Barr
Phlogicylindrium Crous et al.*PseudopestalotiopsisMaharachch. et al.*$
Fungal Diversity

Reticulosphaeria Sivan. & Bahekar#
Robillarda Sacc.$
Sarcostroma CookeSeimatosporium Corda
= Vermisporium H.J. Swart & M.A. Will.*Seiridium Nees *
= Blogiascospora Shoemaker et al.= Lepteutypa Petr.$
= Pestalotia De Not.Sporocadus Corda#
Synnemapestaloides T. Handa & Y. Harada#
Truncatella SteyaertUrosporella G.F. Atk.#
Urosporellopsis W.H. Hsieh et al.#
Xylochora Arx & E. Müll.#
Zetiasplozna Nag Raj
Cainiaceae J.C. Krug*Arecophila K.D. Hyde*$
Cainia Arx & E. Müll.*$
Seynesia Sacc.$
Clypeosphaeriaceae G. WinterApioclypea K.D. HydeAquasphaeria K.D. Hyde#
Brunneiapiospora K.D. Hyde et al.Clypeosphaeria Fuckel$
Crassoascus Checa et al.#
Palmomyces K.D. Hyde et al.#
Coniocessiaceae Asgari & Zare*Coniocessia Dania García et al.$
Diatrypaceae NitschkeAnthostoma NitschkeCryptosphaeria Ces & De Not.Cryptovalsa Ces & De Not. ex FuckelDiatrype Fr.$
Diatrypella (Ces & De Not.) De Not.Diatrypasimilis J.J. Zhou & Kohlm., in Chalkleyet al*Echinomyces Rappaz#
Eutypa Tul. & C. Tul.$
Eutypella (Nitschke) Sacc.Leptoperidia Rappaz#
Monosporascus Pollack & Uecker$
Pedumispora K.D. Hyde & E.B.G. Jones*Peroneutypa Berl.#
Phaeoisaria Höhn.Quaternaria Tul. & C. Tul.#
Graphostromataceae M.E. Barr et al.*$
Graphostroma Piroz.$
Hyponectriaceae Petr.Apiothyrium Petr.#
Arecomyces K.D. Hyde#
Arwidssonia B. Erikss.#
Cesatiella Sacc.#
Chamaeascus L. Holm et al.#
Charonectria Sacc.#
Discosphaerina Höhn.*Exarmidium P. Karst.#
Frondicola K D Hyde*#
Hyponectria Sacc.$
Micronectria Speg.#
Papilionovela Aptroot#
Pellucida Dulym. et al.#
Physalospora NiesslPhragmitensis M.K.M. Wong et al.*#
Pseudomassaria Jacz*$
Rhachidicola K.D. Hyde & J. Fröhl.#
Xenothecium Höhn.#
Melogrammataceae G. Winter*Melogramma Fr.
Vialaeaceae P.F. Cannon*Vialaea Sacc.*$
Xylariaceae Tul. & C. Tul.*Amphirosellinia Y.M. Ju et al.*Annulohypoxylon Y.M. Ju et al.*Anthocanalis Daranagama et al.*Anthostomella Sacc.*Appendixia B.S. Lu & K.D. Hyde#
Areolospora S.C. Jong & E.E. Davis#
Arthroxylaria Seifert & W Gams*Ascotricha Berk.*
= Dicyma Boulanger= Puciola De Bert.
Astrocystis Berk & Broome*Barrmaelia RappazBiscogniauxia Kuntze*Brunneiperidium Daranagama et al.*Calceomyces Udagawa & S UedaCamillea Fr.*
= Basidiobotrys Höhn.= Masoniomyces JL Crane & Dumont= Xylocladium P. Syd. ex Lindau
Cannonia J.E. Taylor & K.D. Hyde#
Chaenocarpus Rebent.#
Chlorostroma A.N. Mill. et al.*Collodiscula I Hino & Katum.*
= Acanthodochium Samuels et al.Coniolariella Dania García et al.*
= Coniolaria Seigle-Mur et al.
Fungal Diversity

Creosphaeria Theiss.*$
Cyanopulvis J. Fröhl. & K.D. Hyde#
Daldinia Ces. & De Not.*= Annellosporium M.L. Davey= Versiomyces Whalley & Watling
Durotheca Læssøe et al.*Emarcea Duong et al.Engleromyces Henn.*Entoleuca Syd.Entonaema MöllerEuepixylon FüistingFasciatispora K.D. Hyde*Fassia Dennis#
Gigantospora B.S. Lu & K.D. Hyde#
Guestia G.J.D. Sm. & K.D. Hyde#
Halorosellinia Whalley et al.Helicogermslita Lodha & D. Hawksw.#
Hypocopra (Fr) J. Kickx fHypocreodendron Henn.*
= Discoxylaria J.C. Lindq. & J.E. WrightHypoxylon Bull.*
= Pyrenopolyporus Lloyd= Triplicaria P Karst *
Induratia Samuels, E. Müll. & PetrinJumillera J.D. Rogers et al.#
Kretzschmaria Fr.*= Holttumia Lloyd
Kretzschmariella Viégas#
Leprieuria Laessøe et al.#
Leptomassaria Petr.#
Libertella Desm.Lopadostoma (Nitschke) Traverso*Lunatiannulus Daranagama et al.*Muscodor Worapong et al.*Myconeesia Kirschst.#
Nemania Gray*$
= Geniculisynnema Okane & Nakagiri= Geniculosporium Chesters & Greenh.
Nodulisporium Preuss*= Pleurographium Goid.
Nipicola K.D. Hyde#
Obolarina PouzarOccultitheca J.D. Rogers & Y.M. Ju#
Ophiorosellinia JJ.D. Rogers et al.#
Pandanicola K.D. Hyde#
Paramphisphaeria F.A. Fernández et al.#
Paucithecium Lloyd#
Phylacia Lév.Podosordaria Ellis. & Holw.Poroleprieuria M.C. González et al.#
Poronia Willd.*= Lindquistia Subram. & Chandrash
Pyrenomyxa Morgan
= Pulveria Malloch & RogersonPyriformiascoma Daranagama et al.*Rhopalostroma D. Hawksw.*Rosellinia De Not.*$
= Dematophora R. Hartig= Vrikshopama D. Rao & P.Rag. Rao
Rostrohypoxylon J Fourn & M Stadler*Ruwenzoria J. Fourn. et al.*Sabalicola K.D. Hyde#
Sarcoxylon Cooke#
Sclerodermatopsis Torrens#
Spirodecospora B.S. Lu et al.#
Squamotubera Henn.#
Steganopycnis Syd. & P Syd.#
Stilbohypoxylon Henn.Streblema Chevall.#
Striatodecospora D.Q. Zhou et al.#
Stromatoneurospora S.C. Jong & E.E. DavisThamnomyces Ehrenb.Theissenia Maubl.Thuemenella Penz. & Sacc.Virgaria Nees*
= Ascovirgaria J.D. Rogers & Y.M. JuVivantia J.D. Rogers et al.#
Wawelia Namysl#
Whalleya J.D. Rogers et al.Xylaria Hill ex Schrank*
= Moelleroclavus Henn.= Padixonia Subram.= Pseudoxylaria Boedijn= Xylocoremium J.D. Rogers
Xylocrea Möller#
Xylotumulus J.D. Rogers et al.
Xylariales, genera incertae sedisAdomia S. Schatz#
Ascotrichella Valldos & Guarro#
Beltraniella Subram.$
Diamantinia A.N. Mill. et al.Hadrotrichum FuckelLanceispora Nakagiri et al.*Lasiobertia Sivan.Leiosphaerella Höhn*Linocarpon Syd. & P. Syd.Neolinocarpon K.D. Hyde*Oxydothis Penz. & Sacc.*Palmicola K.D. Hyde#
Pidoplitchkoviella Kiril.Plectosphaera Theiss.Polyancora Voglmayr & YulePulmosphaeria Joanne E. Taylor et al.#
Sporidesmina Subram. & BhatSubramaniomyces Varghese & V.G. Rao
Fungal Diversity

Yuea O.E. Erikss.#
Sordariomycetes, orders incertae sedis
Amplistromatales D’souza et al. *Amplistromataceae Huhndorf et al.
Amplistroma Huhndorf et al.*$
Wallrothiella Sacc.*$
=Pseudogliomastix W. Gams= Zignoina Cooke#
Catabotrydaceae Petr. ex M.E. BarrCatabotrys Theiss. & Syd.$
Pisorisporiales Réblová & J. Fourn.*Pisorisporiaceae Réblová & J. Fourn.*
Achroceratosphaeria Réblová et al.*Pisorisporium Réblová & J. Fourn.*
Spathulosporales Kohlm.Hispidicarpomycetaceae Nakagiri
Hispidicarpomyces Nakagiri*
Spathulosporaceae Kohlm.Retrostium Nakagiri & Tad Ito*Spathulospora A.R. Caval. & T.W. Johnson*
Sordariomycetes, families incertae sedis
Obryzaceae KörbObryzum Wallr.#
Sordariomycetes, genera incertae sedisAcerbiella Sacc.#
Acrospermoides Miller & G.E. Thomps.#
Ameromassaria Hara#
Amphisphaerellula Gucevič#
Amphisphaerina Höhn.#
Amphorulopsis Petr.#
Amylis Speg.#
Anthostomaria (Sacc.) Theiss. & Syd.#
Anthostomellina L.A. Kantsch.#
Apharia Bonord.#
Apodothina Petr.#
Apogaeumannomyces Matsush.#
Aquadulciospora Fallah & Shearer#
Aropsiclus Kohlm. & Volkm.-Kohlm.#
Ascorhiza Lecht.-Trnka#
Ascoyunnania L. Cai & K.D. Hyde#
Assoa Urries#
Aulospora Speg.#
Azbukinia Lar.N. Vassiljeva#
Bactrosphaeria Penz. & Sacc.#
Biporispora J.D. Rogers et al.#
Bombardiastrum Pat.#
Brenesiella Syd.#
Byrsomyces Cavalc.#
Byssotheciella Petr.#
Caleutypa Petr.#
Calosphaeriopsis Petr.#
Caproniella Berl.#
Chaetoamphisphaeria Hara#
Ciliofusospora Bat. & J.L. Bezerra#
Calcarisporium Preuss#
Clypeoceriospora Sousa da Câmara#
Clypeosphaerulina Sousa da Câmara#
Crinigera SchmidtCryptoascus Petri#
Cryptomycella Höhn.#
Cryptomycina Höhn.#
Cucurbitopsis Bat. & Cif.#
Curvatispora V.V. Sarma & K.D. Hyde#
Dasysphaeria Speg.#
Delpinoëlla Sacc.#
Diacrochordon Petr.#
Dryosphaera Jørg. Koch & E.B.G. Jones#
Duradens Samuels & Rogerson$
Ellisembia Subram.Endoxylina Romell#
Esfandiariomyces Ershad#
Frondisphaera K.D. Hyde#
Glabrotheca Chardόn#
Heliastrum Petr.#
Hyaloderma Speg.#
Hydronectria Kirschst.#
Hypotrachynicola Etayo#
Immersisphaeria Jaklitsch.#
Iraniella Petr.#
Imicles Shoemaker & Hambl.#
Konenia Hara#
Kravtzevia Schwartzman#
Kurssanovia Kravtzev#
Lecythiomyces Doweld#
= Lecythium ZukalLeptosacca Syd.#
Leptosphaerella Speg.#
Leptosporina Chardón#
Liberomyces Pažoutová et al.*Lyonella Syd.#
Mangrovispora K.D.Hyde & Nagakiri#
Melanographium Sacc.#
Melomastia Nitschke ex Sacc.Microcyclephaeria Bat.#
Monotosporella S. Hughes
Fungal Diversity

Naumovela Kravtzev#
Natantiella R blováNeocryptospora Petr.#
Neolamya Theiss. & Syd.#
Neothyridaria Petr.#
Nigrospora Zimm.Ophiomassaria Jacz.#
Orcadia GK Sutherl*#
Paoayensis Cabanela et al.Pareutypella Y.M. Ju & J.D. Rogers#
Phialemoniopsis Perdomo et al.*Phomatosporella Tak. Kobay & K. Sasaki#
Phyllocelis Syd.#
Pleocryptospora J. Reid & C. Booth#
Pleosphaeria Speg.#
Pontogeneia Kohlm.#
Porodiscus Lloyd#
Protocucurbitaria Naumov#
Pulvinaria Bon.#
Pumilus Viala & Marsais#
Rehmiomycella E. Müll.#
Rhamphosphaeria Kirschst.#
Rhizophila K.D. Hyde & E.B.G. Jones#
Rhopographella (Henn.) Sacc. & TrotterRhynchosphaeria (Sacc.) Berl.#
Rivulicola K.D. Hyde#
Romellina Petr.#
Saccardoëlla Speg.#
Sarcopyrenia Nyl.#
Sartorya Vuill.#
Scharifia Petr.#
Scoliocarpon Nyl.#
Scotiosphaeria Sivan.#
Selenosporella G. Arnaud ex MacGarvie 1969*Servazziella J. Reid & C. Booth#
Sporoctomorpha J.V. Almeida & Sousada Câmara#
Stanjehughesia Subram.Stearophora L. Mangin & Viala#
Stegophorella Petr.#
Stellosetifera Matsush.#
Stereosphaeria Kirschst.Stomatogenella Petr.#
Stromatographium Höhn.#
Strickeria Körb.#
Sungaiicola Fryar & K.D. Hyde#
Synsphaeria Bon.#
Tamsiniella S.W. Wong et al.#
Tectonidula RéblováTeracosphaeria R blová & SeifertThelidiella Fink.#
Thyridella (Sacc.) Sacc.#
Thyrotheca Kirschst.#
Trichospermella Speg.#
Trichosphaeropsis Bat. & Nasc.#
Tunstallia Agnihothr.#
Vleugelia J. Reid & C. Booth#
Zalerion R.T. Moore & Meyers*DothideomycetesDothidealesThis obviously does not list all genera ofDothideales but onlythose which were considered to be placed in Sordariomycetesin Lumbsch and Huhndorf (2010) and Wijayawardene et al.(2012) and are now excluded.
Dichomera CookeHendersonula Speg.#
Kabatia Bubák#
Kabatiella BubákSarcophoma Höhn.#
Selenophoma Maire
Leotiomycetes O.E. Erikss. & WinkaHelotiales Nannf.This obviously does not list all genera of Helotialesbut only those which were considered to be placed inSordariomycetes in Lumbsch and Huhndorf (2010)and Wijayawardene et al . (2012) and are nowexcluded.
Chalara (Corda) Rabenh.= Chaetochalara B. Sutton & Piroz.#
Diplococcium Grove *Idriella P.E. Nelson & S. Wilh.*
Ascomycota genera incertae sedisThis does not list all genera of Ascomycota genera incertae
sedis but only those which were considered to be placed inSordariomycetes in Lumbsch and Huhndorf (2010) andWijayawardene et al. (2012) and are now excluded. Thereare large numbers of genera listed in Kirk et al. (2013, 2014)that are considered synonyms in Index Fungorum, however itis impossible to check whether these have been correctlysynonymised and these are not listed here.
Acrophialophora EdwardAnguillospora Ingold*Aphanocladium W. GamsBiflua Jørg. Koch & E.B.G. Jones*#
Cacumisporium Preuss#
Didymobotryum Sacc.DumortieriaWestend.#
Eiona Kohlm.*#
Engyodontium de HoogFusichalara S. Hughes & Nag Raj#
Hapsidascus Kohlm. & Volkm.-Kohlm.*#
Kutilakesa Subram.#
Linkosia A. Hern.-Gut. & B. Sutton*
Fungal Diversity

Lunulospora IngoldMenisporopsis S. Hughes*$
Monocillium S.B. Saksena$
Paliphora Sivan. & B. SuttonPhaeocytostroma Petr.Pleurodesmospora Samson et al.Pleurophragmium CostantinRetroconis de Hoog & Bat Vegte$
Septogloeum Sacc.Sporidesmium Link*Trichocladium Harz*Verticimonosporium Matsush.*$
Notes on outline - 2014
AbyssomycesKohlm., Ber. dt. bot. Ges. 83(9–10): 505 (1970)The type species is Abyssomyces hydrozoicus Kohlm.,
which was collected on hydrocaulon, attached to stony corals(Kohlmeyer 1970). The genus was referred by Jones et al.(2009a, b) to Sordariales genera incertae sedis. Further col-lections and molecular study are required to determine itsnatural taxonomic placement.
Achroceratosphaeria Réblová et al., Fungal Diversity 43: 79(2010)
The genus Achroceratosphaeria was introduced fromfreshwater and terrestrial habitats with the type speciesA. potamia Réblová et al. and two other species (Réblováet al. 2010). Achroceratosphaeria is characterized by im-mersed perithecia, with hyaline to subhyaline emergent necksand cylindrical-clavate asci with a distinct apical annulus.Molecular studies shows that Achroceratosphaeria resides ina single branch as a sister to the clade containingLulworthiales and Koralionastetales and was placed inPisorisporiaceae (Réblová et al. 2015).
Acremon ium L ink , Magaz i n d e r Ge s e l l s ch a f tNaturforschenden Freunde Berlin 3: 15 (1809)
Acremonium is generally considered to be a highly poly-phyletic asexual genus containing distantly related fungi(Glenn et al. 1996), with A. alternatum Link, the type species,in the Hypocreales. Acremonium furcatum Moreau & F.Moreau ex Gams has been shown to group in thePlectosphaerellaceae with Gibellulopsis nigrescens(Pethybr.) Zare et al. as a sister group (Réblová et al. 2011a).Summerbell et al. (2011) confirmed the placement ofA. alcalophilum G. Okada, A. brunnescens W. Gams,A. furcatum, A. nepalense W. Gams, A. restrictum (J.F.H.Beyma) W. Gams and A. stromaticum W. Gams & R.H.Stover in the Plectosphaerellaceae, as was already shown byZare et al. (2007).Acremonium apii (M.A. Sm.& Ramsey)W.Gams also has been shown to belong to this family as a syn-onym of Verticillium albo-atrum Reinke & Berthold. The
phytopathogen A. cucurbitacearum Alfaro-García et al. hasalso been shown to belong in Plectosphaerellaceae (Zareet al. 2007). Summerbell et al. (2011) designated CBS407.66 as the epitype of Acremonium alternatum, the typespecies of the genus. ITS sequences analyses by Giraldoet al. (2012) show that the A. alternatum epitype strain (CBS407.66) and a large group of species currently accepted inAcremonium belong in the family Bionectriaceae.
Acrostalagmus Corda, Icones fungorum hucusquecognitorum 2: 15 (1838)
A polyphyletic asexual genus with most species assignedto the Hypocreales; Acrostalagmus cinnabarinus Corda is thetype species. Réblová et al. (2011a) showed Acrostalagmusannulatus (Berk. & M.A. Curtis) Seifert (= Stilbella annulataBerk. & M.A. Curtis) and A. luteoalbus (Link) Zare et al.groups with high support in the Plectosphaerellaceae, basedon nuclear SSU rDNA sequences, with Verticillium dahliaeKleb. as a sister group.
Actinostilbe Petch, Ann. R. bot. Gdns Peradeniya 9: 327(1925)
Actinostilbe appears to be linked to Lanatonectria.However, they are not common genera, and thus Rossmanet al. (2013) proposed the name Lanatonectria be abandonedin favour of the older and more widely used asexual morphgeneric name Actinostilbe. The genus belongs in Nectriaceae.
Alisea J. Dupont & E.B.G. Jones, Mycol. Res. 113(12): 1358(2009)
Monotypic marine taxon growing on submerged wood re-covered at 1000 m depth in the Pacific Ocean with no knownasexual morph. Molecular data confirm its position inHalosphaeriaceae (Dupont et al. 2009).
Allantonectria Earle, in Greene, Plant. Bak. 2(1): 11 (1901)Hirooka et al. (2012) studied the systematics of Nectria
based on a 6-loci phylogenetic analysis. Nectria divided intotwo main clades: the first clade being Nectria sensu strictowhich includes the type species, N. cinnabarina (Tode) Fr.,along with 27 additional species and genera. The second cladecomprised Pleonectria with 26 species some withZythiostroma asexual morphs. Sister to the Pleonectria cladeis Nectria miltina (Mont.) Mont. which has Trichoderma-likeasexual morphs and occurs on monocotyledons. Since thesecharacters are different from Pleonectria, a new genus,Allantonectria, was introduced.
AmphiroselliniaY.M. Ju et al., Mycologia 96(6): 1393 (2004)Amphirosellinia was introduced by Ju et al. (2004) to in-
clude five xylariaceous taxa with erumpent or immersedperithecioid stromata. The name itself refers to its stromatalanatomy which is similar to that of the genus Rosellinia, but
Fungal Diversity

differs in having stromata that develop beneath the host epi-dermis (Ju et al. 2004). The accepted Amphirosellinia speciesconstitute a coherent group that is characterized by severalunique characteristics, and a synnematous asexual morph thatproduces lacrymoid conidia (Ju et al. 2004). A molecular studythat included type strains of A. fushanensis Y.M. Ju et al. andA. nigrosporaY.M. Ju et al. placedAmphirosellinia as a discretemonophyletic group, which appears to be paraphyletic withXylaria digitata and X. montagnei (Hsieh et al. 2010).
Amphisphaeria Ces. & De Not., Comm. Soc. crittog. Ital.1(4): 223 (1863)
The genus Amphisphaeria was introduced by De Notaris(1863) with A. umbrina (Fr.) De Not. as the type species.Based on molecular data, Kang et al. (1999a) determined thatAmphisphaeria is polyphyletic.
Amplistroma Huhndorf et al., Mycologia 101(6): 907 (2009)Amplistroma was described to accommodate seven species
with A. carolinianum Huhndorf et al. as the type species(Huhndorf et al. 2009). According to phylogenetic analysesusing LSU data, Amplistroma is included in the familyAmplistromataceae formed a monophyletic clade with strongbootstrap and Bayesian support (Huhndorf et al. 2009). In ourcombined analysis Amplistroma clusters with Wallrothiellaand is sister to Catabotrys deciduum in Catabotrydaceae(Amplistromatales).
Anguillospora Ingold, Trans. Br. Mycol. Soc. 25 (4): 401(1942)
A polyphyletic genus with species in the familyLulworthiaceae (A. marina Nakagiri & Tubaki), asexualPleosporales (A. crassa Ingold), Hymenoscyphus imberbis(Bull.) Dennis, Helotiaceae (A. fustiformis Marvanová &Descals) and A. coryli (Redhead & G.P. White) U. Braun,asexual Mycosphaerellaceae (Index Fungorum 2015).Anguillospora marina is a synonym of Lindra obtusaNakagiri & Tubaki as established by cultural studies and mo-lecular data (Jones et al. 2009b). The type species,A. longissima (Sacc. & P. Syd.) Ingold, is not linked to anysexual morph and was placed in asexual Ascomycota inShearer et al. (2009); the genus is retained and placed herein Ascomycota genera incertae sedis.Aniptodera Shearer & M.A. Mill., Mycologia 69(5): 893(1977)
A genus of Halosphaeriaceae established by Shearer andMiller (1977) to accommodate the aquatic fungusA. chesapeakensis Shearer & M.A. Mill. Molecular analysisof SSU and LSU rDNA sequence data confirm thatAniptodera is a polyphyletic genus with A. chesapeakensisand A. lignatilisK.D. Hyde placed in one clade, while the restof the species are placed in distant clades (Jones et al. 2009a).Further study is needed to resolve the taxonomy of the species
referred to Aniptodera. No asexual morph is known.
Anisostagma K.R.L. Petersen & Jørg. Koch, Mycol. Res.100(2): 209 (1996)
Monotypic genus of Halosphaeriaceae that wasestablished by Petersen and Koch (1996) to accommodatethe marine species, A. rotundatum K.R.L. Petersen & Jørg.Koch. This infrequently collected species has no known asex-ual morph. A phylogenetic study is required to determine if itdiffers from Thalassogena and confirm its position in theHalosphaeriaceae.
AnopodiumN. Lundq., Botaniska Notiser 117 (4): 356 (1964)A genus of Lasiosphaeriaceae based onmorphological and
molecular data with A. ampullaceum as the type species. Thegenus groups with Cercophora sulphurella (Sacc.) R. Hilberin a well-supported clade and joins the weakly supported cladecontaining Cercophora sparsa (Sacc. & Fairm.) R. Hilber,Corylomyces selenosporus Stchigel et al., Bellojisiarhynchostoma (Höhn.) Réblová and Podospora didyma J.H.Mirza & Cain (Kruys et al. 2015).
Annulohypoxylon Y.M. Ju et al., in Hsieh et al., Mycologia97(4): 855 (2005)
Two sections were recognized in Hypoxylon by Ju andRogers (1996); Hypoxylon sect. Hypoxylon and Hypoxylonsect. Annulata. Based on a phylogenetic analysis, includingprotein coding genes, Annulohypoxylon was introduced as anew genus by Heish et al. (2005) to accommodate Hypoxylonsect. Annulata with A. truncatum (Schwein.) Y.M. Ju et al.designated as the type species. Hsieh et al. (2005) described37 species and varieties belonging to Annulohypoxylon.Several other species have been added to the genus(Fournier et al. 2010; Hladki and Romero 2009; Ju et al.2004; Marincowitz et al. 2008; Pereira et al. 2010;Vassiljeva and Stephenson 2013); Index Fungorum (2015)l i s t s 50 species . Annulohypoxylon species havenodulosporium-like asexual morphs.
Antennospora Meyers, Mycologia 49: 501 (1957)Monotypic marine ascomycete growing on submerged
wood, often in association with calcareous shells. No asexualmorph is known. Molecular data confirms its position in theHa lo sphae r i a c ea e , f o rm ing a s i s t e r g r oup t oPseudolignincola siamensis with high bootstrap support(Pang et al. 2008; Sakayaroj et al. 2011a).
Anthocanalis Daranagama et al., Fungal Diversity (in press)(2015)
The genus Anthocanalis was introduced for A. spartiDaranagama et al. to represent anthostomella-like taxa distin-guished by having ascomata with a reduced clypeus withshiny papilla and limoniform ascospores (Daranagama et al.
Fungal Diversity

2015). Isolates from ascospores of A. sparti produced anodulisporium-like asexual morph in culture. According tothe phylogenetic analysis by Daranagama et al. (2014) usingfour combined genes, A. sparti clustered in the hypoxyloidXylariaceae and was closely related to Rhopalostroma,Ruwenzoria and Thamnomyces.
Anthonectria Döbbeler, Mycologia 102(2): 405 (2010)This monotypic genus was introduced by Döbbeler (2010)
based on Anthonectria mammispora Döbbeler and is stated tobe invalid in Index Fungorum (2014). According to the mor-phology the genus is classified in family Bionectriaceae.
Apiosporaceae K.D. Hyde et al., in Hyde et al., Sydowia50(1): 23 (1998)
Hyde et al. (1998) introduced Apiosporaceae to accommo-date Apiospora and Appendicospora. Smith et al. (2003) con-cluded that Apiosporaceae represented one of the seven fam-ilies in Xylariales. Lumbsch and Huhndorf (2010) listedApiosporaceae as family incertae sedis (Sordariomycetes).Crous and Groenewald (2013) analysed LSU rDNA sequencedata and confirmed the Apiosporaceae belongs in Xylariales,and is a sister group to Amphisphaeriaceae.
Apodospora Cain & J.H. Mirza, Can. Bot. 48 (5): 891 (1970)Monophyletic genus in Lasiosphaeriaceae with
Apodospora simulans Cain & J.H. Mirza as the type species(Kruys et al. 2015). It groups with a number of other taxa withunclear affinities to each other, lacking higher-level support(Kruys et al. 2015).
Appendichordella R.G. Johnson et al., Can. J. Bot. 65(5): 941(1987)
A monotypic lignicolous genus with no known asexualmorph, the type of which was described in Sphaerulina(Kohlmeyer 1962) and has been referred to Haligena(Kohlmeyer and Kohlmeyer 1979). The genus is morpholog-ically well-placed in the family Halosphaeriaceae, but re-quires further collection, isolation and sequencing to confirmits position therein (Jones et al. 2009a).
Aquanectria Lombard et al., in Lombard et al., Stud.Mycol.80: 207 (2015)
Lombard et al. (2015) introduced the genus Aquanectria,typified by Aquanectria penicillioides (Ingold) L. Lombard &Crous, based on a multi-locus gene analysis to accommodatetwo species previously treated as members of the generaFlagellospora and Heliscus. The genus belongs in the familyNectriaceae.
Arecophila K.D. Hyde, Nova Hedwigia 63(1–2): 82 (1996)Arecophila was introduced by Hyde (1996), with the type
species A. gulubilcola K.D. Hyde and six other species. The
genus was originally placed in Amphisphaeriaceae due to amorphological resemblance to Amphisphaeria but was revisedby Kang et al. (1999a) and excluded from Amphisphaeriaceae.Arecophila differs from Amphisphaeria in having a cylindrical,J+ ascus ring and ascomata immersed under a clypeus. Its striateascospores surrounded by a mucilaginous sheath show certainsimilarities to those of Atrotorquata, Cainia, and Ceriophora(Kang et al. 1999b). Due to its close morphological affinitieswith these genera in Cainiaceae, Arecophila was moved toCainiaceae (Kang et al. 1999b). Molecular data is required toresolve its taxonomic position.
ArenariomycesHöhnk, Veröff. Inst. Meeresf. Bremerhaven 3:28 (1954)
A genus of Halosphaeriaceae, the type species of whichhas been variously referred toHalosphaeria (Cribb and Cribb1956), Peritrichospora (Kohlmeyer 1961) and Corollospora(Kohlmeyer 1962), molecular data confirming its position inthis family (Jones et al. 2009b). No asexual morph is known.Five species are referred to the genus (Jones et al. 2009b;Koch 2013).
Argentinomyces N.I. Peña & Aramb., Mycotaxon 65: 333(1997)
A monotypic genus, based on a single collection, of un-known affinities and requiring further study at the molecularlevel. Jones et al. (2009b) referred Argentinomyces to theDiaporthales genera incertae sedis based on its morphology.Although it has features in common with members of theHalosphaeriaceae, it does not belong in the family as it hasa thick-walled ascus apex and persistent asci.
Arnium Nitschke ex G. Winter, Bot. Ztg.: 450 (1873)A genus in the Lasiosphaeriaceae with A. lanuginosum
Nitschke as the type species (Kruys et al. 2015). The genus,as currently circumscribed, comprises 35 species that are mor-phologically diverse. PhylogeneticallyArnium species are dis-persed in three of the four major clades in the family, whichmay be accounted for by the great morphological plasticitywithin the genus. Further studies, including the type species,are required to resolve the phylogenetic relationship ofArnium species.
Arthrinium Kunze, in Kunze & Schmidt, MykologischeHefte (Leipzig) 1: 9 (1817)
This widespread and ecologically diverse genus is commonon grasses and bamboo. Crous and Groenewald (2013) ac-cepted the name Arthrinium over its sexual morph nameApiospora on the basis that the name Arthrinium is older,more commonly encountered, and more frequently used inliterature. Based on morphology and molecular studies ofLSU and ITS rDNA sequence data, the genus is placed in
Fungal Diversity

Apiosporaceae, order Xylariales (Crous and Groenewald2013). The genera Pteroconium andCordella are also reducedto synonymy with Arthrinium, because spore shape and thepresence of setae were rejected as characters of generic signif-icance (Crous and Groenewald 2013).
Arthroxylaria Seifert & W. Gams, in Seifert et al., CzechMycol. 53(4): 299 (2002)
Arthroxylariawas introduced by Seifert et al. (2002) and istypified by A. elegans Seifert & W. Gams. This xylarioidasexual morph has been connected to Xylaria (Stadler et al.2013). The asexual name could be suppressed in favour ofXylaria as it is the older name and the cosmopolitan morphis frequently encountered in nature. Stadler et al. (2013) sug-gested that the name Arthroxylaria can be retained to accom-modate certain species of Xylaria which is thought to be acomparatively diverse genus; a separation at the subgenericlevel may soon occur with the availability of molecular data.
Ascosacculus J. Campb. et al., Mycologia 95(3): 545 (2003)Genus of Halosphaeriaceae that was introduced by
Campbell et al. (2003) to accommodate Ascosacculusheteroguttulatus (S.W. Wong et al.) J. Campb. et al. andA. aquaticus (K.D. Hyde) J. Campb. et al., two species previ-ously placed in Halosarpheia that grow on decaying wood infreshwater habitats; the genus is typified by A. aquaticus. In amolecular study, species of Ascosacculus clustered in a well-supported clade with Aniptodera species (A. chesapeakensisShearer & M.A. Mill. and A. lignatilis K.D. Hyde), that isdistant from H. fibrosa Kohlm. & E. Kohlm., the type speciesof the genus Halosarpheia. Ascosacculus aquaticus produceschains of brown chlamydospores (Campbell et al. 2003),while A. heteroguttulatus has a asexual morph inTrichocladium (Wong et al. 1998a, b).
Ascotaiwania Sivan. & H.S. Chang, Mycol. Res. 96(6): 481(1992)
The genus was introduced to accommodate a freshwaterascomycete (A. lignicola) collected on wood in Taiwan(Sivanesan and Chang 1992) and referred to the Sordarialesby Cai et al. (2006a). Currently 12 species are known fromwood and palm material and are characterised by a conspicu-ous, apical, J- ring in the ascus, reminiscent of that found inAnnulatascus species. A combined phylogenetic analysis ofAscotaiwania species, showed that they grouped inSavoryellales (Hypocreomycetidae), but the position is not fullyresolved. Ascotaiwania sawadae H.S. Chang & S.Y. Hsieh,A. mitriformis Ranghoo & K.D. Hyde and A. lignicola Sivan.&H.S. Chang form awell-supported clade in the Savoryellales,but A. hughesii Fallah et al. does not group with the otherspecies (Boonyuen et al. 2011). Several dematiaceous hypho-mycetous anamorphs have been reported for Ascotaiwania, in-cluding Monotosporella sp. (for Ascotaiwania sawadae,
Sivicha et al. 1998), Monotosporella setosa (Berk. & M.A.Curtis) S. Hughes (for Ascotaiwania sawadae, Ranghoo andHyde 1998), Helicoön farinosum Linder (Ascotaiwaniahughesii, Fallah et al. 1999; Tsui and Berbee 2006). Furthercollections, isolation and sequence data are required to resolvethe taxonomic assignment of individual species in this genus.
Ascotricha Berk., Ann. nat. Hist., Mag. Zool. Bot. Geol. 1:257 (1838)
Dicyma was introduced with D. ampullifera Boulanger1897 as the type species. This asexual genus has been linkedwith Ascotricha by Saccardo (1899) as mentioned in IndexFungorum (2015). The morphological characters of Dicymaampullifera almost perfectly match those of Ascotrichachartarum and Chaetomium chartarum (Saccardo 1899).Stadler et al. (2013) suggested that the older name be givenpriority and that Dicyma placed in synonymy.
Astrocystis Berk. & Broome, J. Linn. Soc., Bot. 14 (no. 74):123 (1873) [1875]
Astrocystis was introduced by Berkeley and Broome(1874) to accommodate species with uniperitheciate or rarelymulti-peritheciate stromata which appear superficial. Pinnoiet al. (2010) introduced a new species Astrocystis eleiodoxaePinnoi et al. and inferred the phylogenetic relationships of thisand related species based on rDNA sequences of the ITS1-5.8S-ITS2 and partial SSU and LSU genes. They observedthat A. eleiodoxae clusters with Stilbohypoxylon elaeicola(Henn.) L.E. Petrini with weak support when ITS and 28SrDNAwere combined. However, multilocus sequence analy-sis confirmed the monophyly of A. eleiodoxae and that it waswell positioned in the family Xylariaceae.
Atractium Link, Mag. Gesell. naturf. Freunde, Berlin 3(1–2):10 (1809)
In the phylogenetic analysis of Gräfenhan et al. (2011) twoAtractium species, including the type species A. stilbasterLink, formed a well-supported monophyletic group inNectriaceae. No sexual morphs are conclusively known forthis genus.
Aurantiosacculus Dyko & B. Sutton, in Dyko et al.,Mycologia 71(5): 922 (1979)
Two new Aurantiosacculus species (A. acutatus Crous &Summerell and A. eucalyptorum Crous & C. Mohammed)clustered in Cryphonectriaceae (Crous et al. 2012a). No sex-ual morph is known for this coelomycetous genus.
Auratiopycnidiella Crous & Summerell, in Crous et al.,Persoonia, Mol. Phyl. Evol. Fungi 28: 69 (2012)
This genus has a unique set of morphological characteris-tics when compared to other genera of coelomycetous
Fungal Diversity

anamorphs treated by Sutton (1980). The genus may be dis-tantly allied to Melanconidaceae, but is presently placed inDiaporthales genera incertae sedis (Crous et al. 2012d). Nosexual morph is known for this monotypic genus.
Aurifilum Begoude et al., in Begoude et al., Antonie vanLeeuwenhoek 98(3): 273 (2010)
The monotypic genus Aurifilum was introduced forA. marmelostoma Begoude et al., a species from bark ofTerminalia ivorensis from the Cameroons (Begoude et al.2010) and was placed in Cryphonectriaceae based on molec-ular data. It has several obvious characteristics that differ fromother genera in Cryphonectriaceae, especially in the asexualmorph. Conidiomata are broadly convex, and wider than sim-ilar structures of Amphilogia and Rostraureum. The presenceof darkened ostiolar openings at the apex of the conidiomata isalso unique to the taxon.
Australiasca Sivan. & Alcorn. Aust. Syst. Bot. 15(5): 741(2002)
Sivanesan and Alcorn (2002) published this genus withAustraliasca queenslandica Sivan. & Alcorn as the typespecies and with Dischloridium camelliae Alcorn & Sivan.asexual morph which is thus a synonym. This genus wastransferred from Chaetosphaeriaceae to Australiascaceae byRéblová et al. (2011a) based on molecular data.
Australiascaceae Réblová & W. Gams, in Réblová et al.,Stud. Mycol. 68(1): 171 (2011)
The family Australiascaceaewas introduced based on LSU,and a combined dataset of LSU, SSU andRPB2 andwas placedin the Glomerellales (Réblová et al. 2011a). The family in-cludes the sexual genus Australiasca and asexual genusMonilochaetes. Currently, the family comprises Australiascaqueenslandica Sivan. & Alcorn, A. laeënsis Réblová & W.G am s , Mon i l o c h a e t e s i n f u s c a n s Ha r t e r a n dM. guadalcanalensis (Matsush.) I.H. Rong & W. Gams. It ac-commodates species with a sexual morph that is similar toChaetosphaeria with indistinguishable perithecia. The asexualmorphs are phialidic dematiaceous and hyphomycetous, whichare also similar to the asexual morphs of Chaetosphaeria.
Baipadisphaeria Pinruan, in Pinruan et al., Mycosphere 1: 58(2011)
Pinruan et al. (2010) introduced this monotypic genus, withB. spathulospora Pinruan et al. as its type, based on a collec-tion from Thailand in a peat swamp forest on a submergedwoody trunk of Licuala longicalycata. Phylogenetic analysesof LSU and SSU rDNA sequence data reveal that the genusbelongs in the Nectriaceae (Hypocreales). where it forms asister clade with Leuconectria.
Bagadiella Cheew. & Crous, Persoonia 23: 59 (2009)
Cheewangkoon et al. (2009) introduced this hyphomy-cetous genus with B. lanata Cheew. & Crous as its type.Currently this genus has three species and the sexual morphis unknown. Bagadiella is similar to Cladorrhinum(Lasiosphaeriaceae) in having pigmented hyphae andpustular-like aggregations of conidiophores. Based on LSUsequence data, the status of this new genus is supported andreferred to Diaporthales genera incertae sedis.
Bambusicularia Klaubauf et al., Stud. Mycol. 79: 105 (2014)Bambusicularia is a monotypic genus of Pyriculariaceae
recently introduced by Klaubauf et al. (2014) to accommodatethe plant pathogenic B. brunnea Klaubauf et al. which wasisolated from Sasa sp. in Japan. It differs from Pyricularia inconidiophore morphology and phylogeny based on LSU, ITS,RPB1, ACT and CAL sequence data (Klaubauf et al. 2014).
Barretomyces Klaubauf et al., Stud. Mycol. 79: 105 (2014)Based on sequence analysis of LSU, ITS, RPB1, actin and
calmodulin genes, Klaubauf et al. (2014) introduced this mono-typic genus, assigned to Pyriculariaceae for Barretomycescalatheae (D.J. Soares et al.) Klaubauf et al., a foliar pathogenof Calathea longifolia from Brazil (Soares et al. 2011).
Bartalinia Tassi, Bulletin Labor. Orto Bot. de R. Univ. Siena3: 4 (1900)
Bartalinia is a coelomycetous anamorph genus which ac-commodates species having conidia with either three or foursepta (Marincowitz et al. 2010). The taxonomic revision ofAndrianova and Minter (2007) provided a key to tenBartalinia species. Marincowitz et al. (2010) introducedB. pondoensis Marinc. et al., which was isolated from SouthAfrica from living leaves of Maytenus abbottii. An epitypewas designated for the type species B. robillardoides Tassiwhich showed the genus to belong in Amphisphaeriaceae.
Bathyascus Kohlm., Revue de Mycologie 41(2): 190 (1977)Genus with no known asexual morph. Although gen-
erally referred to the Halosphaeriaceae, preliminary mo-lecular results indicate it has no affinity with taxa inthis family (Jones et al. 2006). Further collections ofthe five Bathyascus species are required for isolationand sequencing.
Bertiaceae Smyk, Ukr. bot. Zh. 38(6): 47 (1981)Mugambi and Huhndorf (2010) revisited the order
Coronophorales with DNA sequence data and showedBertiaceae to be monophyletic. Bertiaceae was expanded toinclude Gaillardiella and Bertia didyma (Speg.) Mugambi &Huhndorf (Mugambi and Huhndorf 2010).
Biciliospora Petr., Sydowia 6(5–6): 429 (1952)See under Scortechiniaceae.
Fungal Diversity

Biconiosporella Schaumann, Veröff. Inst. Meeresforsch.Bremerhaven 14: 24 (1972)
The genus Biconiosporella was referred to theLasiosphaeriaceae based on morphological observations,but requiring sequence data to confirm its assignment to thefamily (Jones et al. 2009b).
Biflua Jørg. Koch & E.B.G. Jones, Can. J. Bot. 67: 1187(1989)
Monotypic marine genus based on morphological observa-tions, requiring study at the molecular level (Jones et al.2009b) and referred to Ascomycota genera incertae sedis. Itgrows on wood associated with sand.
Bionectriaceae Samuels & Rossman, Stud. Mycol. 42: 15(1999)
Rossman et al. (2013) suggested that Bionectriaceaewhichhas been frequently cited, be retained over Spicariaceae 1934which has rarely been cited. They suggested protectingBionectriaceae, despite the synonymy of Bionectria andClonostachys (syn. Spicaria), and maintaining use of theyounger name.
BiscogniauxiaKuntze, Revis. gen. pl. (Leipzig) 2: 398 (1891)Biscogniauxia Kuntze includes xylariaceous taxa with bi-
partite stromata that do not release KOH extractable pigments(Ju et al. 1998). Their asexual morphs in culture are usuallyconsidered as a variation of nodulisporium-like topericoniella-like (Stadler et al. 2013). Molecular studies,mainly based on ITS and protein coding genes (Hsieh et al.2005; Stadler et al. 2013, 2014), place Biscogniauxia as abasal lineage to other xylariaceous taxa.
Bisifusarium Lombard et al., in Lombard et al., Stud. Mycol.80: 223 (2015)
This genus is introduced by Lombard et al. (2015), typifiedby B. dimerum (Penz.) L. Lombard & Crous, to includefusarium-like species previously classified in the genusFusarium. Species of Bisifusarium can be differentiated fromspecies in Fusarium by their short, (0–) 1–2(−3)-septatemacroconidia and the formation of lateral phialidic pegs aris-ing from the hyphae. Phylogenetic studies revealed thatBisifusarium is closely related to Fusarium in Nectriaceae,but distinct from the Fusarium clade.
Bleptosporium Steyaert, Darwiniana 12: 171 (1961)Bleptosporium includes two species, the generic type
B. montteae Speg. ex Steyaert and B. pleurochaetum (Speg.)B. Sutton. The asexual Bleptosporium has been linked toAmphisphaeria and Discostroma (Hyde et al. 2011, IndexFungorum 2015), but these links have not been proven bymolecular data. Both names should be retained pending freshcollection and molecular study.
Boliniaceae Rick, Brotéria, sér. bot. 25: 65 (1931)The family was emended by Untereiner et al. (2013) based
on molecular data and includes the genera Apiocamarops,Apiorhynchostoma, Camarops, Camaropella, Cornipulvina,Endoxy la , Mol l i camarops , Neohypod i scus andPseudovalsaria. A key to genera was also provided.
Broomella Sacc., Syll. fung. (Abellini) 2: 557 (1883)Broomella species are distinct in having ellipsoid-fusiform,
inequilaterally curved, triseptate ascospores with central browncells and hyaline end cells, with a single appendage at each end(Kang et al. 1999a). Many species in this genus were initiallylinked to Pestalotia asexual morphs (Shoemaker and Müller1963). Based on the conidial forms, Steyaert (1949) splitPestalotia into three genera, namely Pestalotia, Pestalotiopsisand Truncatella (4-celled conidia). However Nag Raj (1985,1993) preferred to adopt a broader concept for Pestalotiopsisto include 3-septate conidial forms. Pestalotiopsis besseyi(Guba) Nag Raj, P. casuarinae (Cooke & Massee) Nag Raj,P. citrina (McAlpine) Nag Raj, P. eupyrena (Tassi) Nag Raj,P. gastrolobi (Tassi) Nag Raj, P. jacksoniae (Henn.) Nag Raj,P. moorei (Harkn.) Nag Raj, P. pestalozzioides (Dearn. &Fairm.) Nag Raj, P. puyae (Henn.) Nag Raj, P. stevensoniiistevensonii (Peck) Nag Raj and P. torrendiii (J.V. Almeida &Sousa da Câmara) Nag Raj are 3-celled conidial forms that NagRaj (1993) placed in Pestalotiopsis, but which actually belongin Truncatella (Maharachchikumbura et al. 2014). LSU se-quence data reveal that Truncatella is a distinct genus in thefamily Amphisphaeriaceae (Jeewon et al. 2002; Barber et al.2011; Maharachchikumbura et al. 2014).
Brunneiperidium Daranagama et al., Fungal Diversity (inpress) (2015)
Brunneiperidium was introduced as a new genus to accom-modate two new speciesB. gracilentumDaranagama et al. (typespecies) and B. involucratum Daranagama et al. (Daranagamaet al. 2015). Phylogenetic analysis places these species withinthe xylarioid clade comprising Xylaria and Kretzschmaria. Thegenus hasmorphological similarities withNipicola, however thephylogenetic relationship of Brunneiperidium and Nipicolacould not be evaluated due to the lack of cultures of the latter.
Brunneodinemasporium Crous & R.F. Castañeda, in Crouset al., Persoonia, Mol. Phyl. Evol. Fungi 28: 128 (2012)
Crous et al. (2012b) studied Dinemasporium and relatedgenera and showed species with tightly aggregatedconidiogenous cells and pale brown conidia belong in themonotypic genus Brunneodinemasporium. They placed thegenus in Chaetosphaeriaceae with B. brasiliense Crous &R.F. Castañeda as the type species.
Brunneosporella Ranghoo & K.D. Hyde, Mycol. Res.105(5): 625 (2001)
Fungal Diversity

This monotypic genus with Brunneosporella aquaticaRanghoo & K.D. Hyde as type species is known to besaprobic on wood in freshwater habitats. It was placed inAnnulatascaceae based on the black ascomata, cylindrical as-ci with a refractive apical ring and brown uniseptateascospores. Ranghoo et al. (1999) in their phylogenetic studyshowed that Brunneosporella (as Ascobrunneispora) clusterswith Annulatascus in Annulatascaceae. Réblová et al. (2015)showed that Brunneosporella clusters in the familyPapulosaceae in phylogenetic analyses.
Bryocentria Döbbeler, Mycol. Progr. 3(3): 247 (2004)Stenroos et al. (2010) obtained sequence data for this genus
and showed it belongs in Bionectriaceae. The generic type,B. brongniartii Döbbeler, is a frequent biotrophic and host-specific leaf perforator of Frullania dilatata. Currently sevenspecies are assigned to the genus (Index Fungorum 2015).
Bullimyces A. Ferrer et al., Mycologia 104(4): 868 (2012)This genus was introduced by Ferrer et al. (2012), with
Bullimyces communis Ferrer et al. as the type species, basedon a collection of submerged woody debris in freshwater.Molecular analysis showed the genus to belong inSordariomycetidae genera incertae sedis.
Cainia Arx & E. Müll., Acta bot. neerl. 4(1): 111 (1955)Krug (1977) revised the genus and introduced Cainiaceae
to accommodate two species; the type species is Cainiagraminis (Niessl) Arx & E. Müll. According to Kang et al.(1999b) another genus Atrotorquata, which is closely relatedto Cainia in their respective morphologies, did not infer anyphylogenetic affinity with the Amphisphaeriaceae (sensustricto), therefore the genus was excluded from this family.Kang et al. (1999b) pointed out that Atrotorquatamorpholog-ically resembles Cainia.
CalonectriaDeNot., Comm. Soc. crittog. Ital. 2(3): 477 (1867)The asexual morph of the type species of Calonectria, C.
daldiniana (= C. pyrochroa (Desm.) Sacc. 1878) is a speciesof Cylindrocladium. Rossman (1979) established the connec-tion from a culture study. A decision is needed for the usebetween Cylindrocladium and Calonectria. Calonectria isthe older name, a well-known pathogen and has morespecies epithets than Cylindrocladium. Crous et al. (2013)described two new species in Calonectria which only hadCylindrocladium asexual morphs.
Calosphaeria Tul. & C. Tul., Select. fung. carpol. (Paris) 2:108 (1863)
Calosphaeria pulchella (Pers.) J. Schröt. was found to forma distinct acremonium-like asexual morph in culture, for whichthe monotypic genus Calosphaeriophora (Calosphaeriophorapulchella Réblová et al. as type) was proposed (Réblová et al.
2004). It causes almond die back in Iran and has an impact onthe almond industry (Arzanlou and Dokhanchi 2013).Calosphaeriophora becomes a synonym of Calosphaeria.
Camillea Fr., Summa veg. Scand., Section Post. (Stockholm):382 (1849)
The monotypic asexual genus Masoniomyces was intro-duced by Crane and Dumont (1975) and later synonymisedwith Xylocladium. Masoniomyces claviformis J.L. Crane &Dumont, the type, was then named as Xylocladium claviforme(J.L. Crane & Dumont) Arx which is currently in use. Jongand Rogers (1972) investigated the asexual morphs of theCamillea species from culture and later Rogers (1984) report-ed C. broomeana (Berk. & M.A. Curtis) Læssøe et al. to havea Xylocladium asexual morph. Læssøe et al. (1989) observedthe development of a Xylocladium asexual morph in culture ofC. leprieurii Mont., and therefore assigned the asexual genusXylocladium to the sexual genus Camillea. Stadler et al.(2013) mentioned that the asexual morph Xylocladium shouldbe synonymised under Camillea.
Canalisporium (Hol.-Jech. & Mercado) Nawawi & Kuthub.,Mycotaxon 34(2): 479 (1989)
This genus was introduced by Nawawi and Kuthubutheen(1989) to accommodate the type species Canalisporiumcaribense (Hol.-Jech. & Mercado) Nawawi & Kuthub., afreshwater hyphomycetous anamorph growing on wood. Acombined analysis of partial 18S, 28S rDNA and internal tran-scribed spacer 5.8S region showed that seven Canalisporiumspecies formed a highly supported monophyletic clade in theHypocreomycetidae with Ascothailandia, a sexual morph ge-nus. Ascothailandia had been introduced to accommodate afreshwater taxon, Ascothailandia grenadoidia Sri-indr. et al.,collected in Thailand (Sri-indrasutdhi et al. 2010). Ascosporeisolates produced the Canalisporium grenadoidium Sri-indrasutdhi et al. asexual morph in culture. ThusAscothailandia, which is a recent name, becomes a synonymof Canalisporium. Subsequently, the genus Canalisporiumwas referred to the order Savoryellales (Boonyuen et al. 2011).
Cancellidium Tubaki, Trans. Mycol. Soc. Japan 16(4): 357(1975)
Cancellidium applanatum Tubaki is the type species of thegenusCancellidium. Based on a combined analysis of ITS andLSU sequences,Cancellidium lies in Sordariomycetes genera,incertae sedis (Pratibha et al. 2014).
Carbosphaerella I. Schmidt, Feddes Repert. 80: 108 (1969)The genus well-placed at the morphological and molecular
level in the familyHalosphaeriaceae (Sakayaroj et al. 2011a);no asexual morph is known. Molecular data indicates the ge-nus has affinities with the genus Remispora (Sakayaroj et al.2011a).
Fungal Diversity

Carpoligna F.A. Fernández & Huhndorf, in Fernández et al.,Mycologia 91(2): 253 (1999)
The genus Carpoligenawas introduced by Fernández et al.(1999) based on characters of the asexual morph and the anal-ysis of ribosomal DNA data. The characteristics of ascomataand ascospores of the sexual morph resemble typicalChaetosphaeria species, however, the asexual morphs arereadily distinguished by the typical phialidic conidiophoresin Chaetosphaeria species (Fernández et al. 1999).Fernández et al. (1999) noted a high degree of DNA diver-gence between Carpoligena and taxa belonging to the orders,Diaporthales, Hypocreales, Microascales and Sordariales.
Ceratocystidaceae Locq. ex Réblová, W. Gams & Seifert,Stud. Mycol. 68(1): 188 (2011)
The name Ceratocystidaceae (as BCeratocystaceae^) wasproposed by Locquin (1972), but was not validly published.This was rectified in Réblová et al. (2011a, b) with phyloge-netic data and a recent treatment was provided by De Beeret al. (2014). The family Ceratocystidaceae formed a stronglysupported monophyletic group consisting of Ceratocystis,Cornuvesica, Thielaviopsis and the type species ofAmbrosiella (Ambrosiella xylebori Brader ex Arx &Hennebert) and is placed in the order Microascales based onmolecular data (Réblová et al. 2011a, b). De Beer et al. (2014)treated and accepted Ambrosiella, Ceratocystis, Chalaropsis,Da v i d s o n i e l l a ( a n ew l y i n t r o d u c e d g e n u s ) ,Endoconidiophora, Huntiella (a newly introduced genus)and Thielaviopsis in the family. They excluded Cornuvesica,but did not mention which family it should be placed; here werefer it to Microascales genera incertae sedis.
Ceratolenta Réblová, Mycologia 105(2): 466 (2013)Réblová (2013a) described this new genus with a single
species Ceratolenta caudata Réblová based on morphologyand phylogenetic analyses. Phylogenetic analyses of com-bined genes of this wood-inhabiting taxon indicated that thisspecies belongs in a separate monophyletic branch (100/1.0)without close relationship to other morphologically similarfungi of the Sordariomycetidae.
Ceriosporopsis Linder, Farlowia 1: 408 (1945)Genus well-placed at the morphological and molecular lev-
el in the familyHalosphaeriaceae, with no documented asex-ual morph, except it does produce profuse chlamydospores onwood and in culture (Jones et al. 2009a, b). Ceriosporopsisspecies form a well-supported clade that has affinities withOceanitis (Sakayaroj et al. 2011a; Pang et al. 2012). Basedon molecular data the genus Bovicornua is reduced to synon-ymy with Ceriosporopsis (Sakayaroj et al. 2011a).
Chaetoconis Clem., Gen. fung. (Minneapolis): 125, 176(1909)
Chaetoconis is a coelomycetous anamorph proposed toaccommodate Kellermania polygoni, which is a segregate ofKellermania (Nag Raj 1993). Presently there are two species,Chaetoconis polygoni (Ellis & Everh.) Clem. and C. vacciniiMelnik & Nag Raj. De Gruyter et al. (2009) placed the typespecies, C. polygoni, in Diaporthales genera incertae sedisbased on sequence analysis.
Chaetorostrum Zelski et al., Mycosphere 2(5): 594 (2011)This monotypic genus was introduced for Chaetorostrum
quincemilensis Zelski et al., collected on woody debris fromstreams in a lower montaine cloud forest in Peru. The taxon alsoproduces a taeniolella-like asexual morph in culture, which is afirst record of an asexual morph in Annulatascaceae, however,the placement of this genus was confirmed only by morphologyand thus placed in Annulatascaceae sensu lato (Zelski et al.2011a). This genus can be compared to Ascotaiwania, whichwas recently placed in Savoryellales (Boonyuen et al. 2011).
Chaetosphaerellaceae Huhndorf et al., Mycol. Res. 108(12):1387 (2004)
Mugambi and Huhndorf (2010) revisited the orderCoronophorales with DNA sequence data and showedChaetosphaerellaceae to be monophyletic, composed ofChaetosphaerella and Crassochaeta.
Chamaeleomyces Sigler, J. Clin. Microbiol. 48(9): 3186(2010)
Chamaeleomyces was introduced as a new monotypic ge-nus for C. granulomatis Sigler, isolated from the liverChamaeleo calyptratus collected from a zoo (Sigler et al.2010). According to morphological studies and phylogeneticanalyses of nuclear ribosomal rRNA (rDNA) the genus be-longs in Clavicipitaceae. Paecilomyces viridis was concur-rently transferred to the genus as Chamaeleomyces viridis(Segretain et al.) Sigler (Sigler et al. 2010).
Chlorostroma A.N. Mill. et al., Sydowia 59(1): 142 (2007)Chlorostroma was introduced by Miller et al. (2007) as a
monotypic new genus for C. subcubisporum A.N. Mill. et al..This taxon was first collected inhabiting the stromata ofHypoxylon perforatum (Schwein.) Fr. in the USA and resemblesThuemenella cubispora (Ellis & Holw.) Boedijn in stromatalmorphology. Thus, the genus was placed in Xylariaceae byMiller et al. (2007). Læssøe et al. (2010) detected a lepraric acidderivative in the type specimen. Chlorostroma cyaninumLæssøe et al. was described from Thailand and has the samemetabolite profile as Hypoxylon aeruginosum J.H. Mill. sug-gesting therefore concluded that Chlorostroma and Hypoxylonaeruginosum are closely related Læssøe et al. (2010). However,the taxonomic conclusion is not yet established because culturesto study the asexual morphology, secondary metabolites andmolecular phylogeny are lacking (Læssøe et al. 2010).
Fungal Diversity

Chromendothia Lar.N. Vassiljeva, Mikol. Fitopatol. 27(4): 5(1993)
This monotypic genus was introduced by Vasilyeva (1993)for C. appendiculata Lar.N. Vassiljeva. In the present studyC. citrina Lar.N. Vassiljeva clusters in Cryphonectriaceae(Fig. 2).
Chrysocrypta Crous & Summerell, in Crous et al., Persoonia,Mol. Phyl. Evol. Fungi 28: 165 (2012)
This monotypic genus was introduced for a foliar pathogenof Corymbia sp. and is coelomycetous. Crous et al. (2012c)placed it in the Cryphonectriaceae.
Chrysoporthe Gryzenh. & M.J. Wingf., Stud. Mycol. 50(1):129 (2004)
The genus Chrysoporthella is typified by C. hodgesianaGryzenh. & M.J. Wingf., and Chrysoporthe is typified byC. cubens i s (Brune r ) Gryzenh . & M.J . Wingf .Chrysoporthella hodgesiana is recognized as a species ofChrysoporthe based on ITS and β-tubulin data (Gryzenhoutet al. 2004). Phylogenetic analysis showed C. austroafricanaGryzenh. &M.J.Wingf.,C. cubensis,C. doradensisGryzenh.& M.J. Wingf. and Chrysoporthella hodgesiana grouped to-gether as a single genus (Gryzenhout et al. 2004).Chrysoporthe (asexual morph Conoideocrella) is the mostcommonly used name and therefore Chrysoporthella shouldbe considered a synonym.
CirrenaliaMeyers & R.T. Moore, Am. J. Bot. 47: 346 (1960)A genus inHalosphaeriaceae supported by molecular data
(Abdel-Wahab et al. (2010) with C. macrocephala (Kohlm.)Meyers & R.T. Moore as the type species, although the strainsequenced was not ex-type. However, the genus is polyphy-letic with a sexual morph in Juncigera (Microascales;Juncigenaceae), and putative assignments of new genera inLulworthiales (Hydea = Cirrenalia pygmea Kohlm.;Halazoon melhae Abdel-Aziz et al., H. fuscus (I. Schmidt)Abdel-Wahab et al., Abdel-Aziz & E.B.G. Jones;Moleospora maritima Abdel-Wahab et al., Matsusporiumtropicale (Kohlm.) E.B.G. Jones & K.L. Pang = Cirrenaliatropicalis Kohlm.) and Pleosporales (Hiogispora =Cirrenalia japonica Sugiy.) (Abdel-Wahab et al. 2010), basedon molecular data.
Cladobotryum Nees, Syst. Pilze (Würzburg): 56 (1816)[1816–17]See under Hypomyces.
Clavatospora Sv. Nilsson exMarvanová & Sv Nilsson, Trans.Br. Mycol. Soc. 57: 531 (1971)
A polyphyletic genus with species in Halosphaeriaceae(C. bulbosa (Anastasiou) Nakagiri & Tubaki) and
Leotiaceae (C. flagellate J. Gönczöl). Clavatospora bulbosa(= Clavariopsis bulbosa Anastasiou) was shown by culturetechniques to be the asexual morph of Corollospora pulchella(Shearer and Crane 1971). Other species need to be sequencedto determine their phylogenetic position. The type speciesClavatospora longibrachiata (Ingold) Sv. Nilsson exMarvanová & Sv. Nilsson, clusters in Halosphaeriaceae(Duarte et al. 2015).
Clonostachys Corda, Pracht-Fl. Eur. Schimmelbild.: 31(1839)
Schroers (2001) reported a possible link betweenClonostachys and Bionectria, which have consistently beenconsidered congeneric and this is followed in Rossman et al.(2013). Because the name Clonostachys rosea (the generictype) is commonly used in biocontrol studies, Rossman et al.(2013) proposed the protection of the older asexual morph-typified name Clonostachys for this genus.
Clypeoporthella Petr., Annls mycol. 22(1/2): 149 (1924)Clypeoporthella is typified by C. brencklei Petr. which oc-
curs on goldenrod (Solidago) in North America. This may be asynonym ofDiaporthe but needs recollecting and sequencing.
Coccinonectria Lombard & Crous, in Lombard et al., Stud.Mycol. 80: 218 (2015)
The sexual genus Coccinonectria was established byLombard et al. (2015) with C. pachysandricola (B.O.Dodge) L. Lombard & Crous as the type species. It is distin-guished from Pseudonectria by its orange to scarlet ascomatawith short, thick-walled setae extending from the ascomatalsurface. Molecular analysis using combined data sets recog-nized Coccinonectria as a separate genus that is clearly dis-tinct from the genus Pseudonectria (Lombard et al. 2015).
Codinaeopsis Morgan-Jones, Mycotaxon 4(1): 166 (1976)Codinaea gonytrichoides Shearer & Crane was introduced
by Shearer and Crane (1971), was selected as the type of themonotypic genus Codinaeopsis by Morgan-Jones (1976), a ge-nus characterized by its encircling hyphae and conidiogenouscells with collarettes.Whitton et al. (2000) combined this speciesas Dictyochaetopsis gonytrichoides. Crous et al. (2012b) placedCodinaeopsis in the family Chaetosphaeriaceae based on themolecular data. In this study Crous et al. (2012b) is followed.
Colletotrichum Corda, in Sturm, Deutschl. Fl., 3 Abt. (PilzeDeutschl.) 3(12): 41, tab. 21 (1831)
This genus has been placed in Glomerellaceae by Réblováet al. (2011a, b), which is also confirmed in this study (Fig. 2).Significant changes in the understanding of Colletotrichum spe-cies took place following the outline given byHyde et al. (2009a,
Fungal Diversity

b) and recommendation for use of polyphasic approach for spe-cies identification (Cai et al. 2009). Incorporation of these ap-proaches especially the use of multigene phylogenetic analysis,classification and knowledge of species complexes, as well asepitypification, contributed to the better understanding of thegenus Colletotrichum (Cannon et al. 2008, 2012; Crouch 2014;Damm et al. 2012a, b, 2013; Doyle et al. 2013; Weir et al. 2012,Hyde et al. 2014). The epitypification of C. gloeosporioides(Penz.) Penz. & Sacc. (Cannon et al. 2008) was particularlyimportant. Twenty-five Colletotrichum species have now beenepitypified, while one has been neotypified, and three have beenlectotypified (Damm et al. 2009, 2012a, b, 2013; Doyle et al.2013; Liu et al. 2011, 2013; Su et al. 2011; Weir and Johnston2010; Weir et al. 2012).
Cannon et al. (2012) incorporated six gene analyses fornearly all the presently sequenced species of this genus andat least nine clades were revealed. Colletotrichumgloeosporioides (Cannon et al. 2008; Phoulivong et al.2010; Weir et al. 2012), C. acutatum J.H. Simmonds(Marcelino et al. 2008; Shivas and Tan 2009; Damm et al.2012a), C. boninense Moriwaki (Moriwaki et al. 2003; Yanget al. 2009; Damm et al. 2012b), C. destructivum O’Gara(Damm et al. 2014), C. orbiculare Damm et al. (Dammet al. 2013) are well-resolved important species complexesamong the nine clades. Damm et al. (2013) resolvedC. orbiculare and introduced four new species, whileCrouch (2014) introduced a new species complex asC. caudatum (Peck ex Sacc.) Peck with five new speciesfound on warm-season grasses, characterized by spores withthe apex extended to a filiform appendage. Further studies inthe C. gloeosporioides species complex led to recognition ofC. citricola (Huang et al. 2013), C. dianesei (Lima et al. 2013),C. endomangiferae (Vieira et al. 2014), C. endophytica(Manamgoda et al. 2013), C. fructivorum (Doyle et al. 2013),C. melanocaulon (Doyle et al. 2013), C. murrayae (Peng et al.2012), C. syzygicola (Udayanga et al. 2013), C. temperatum(Doyle et al. 2013) and C. viniferum (Peng et al. 2013).Rakotoniriana et al. (2013) introduced a singleton speciesC. gigasporum, Tao et al. (2013) introduced seven new species;four species belonging to the graminicola clade, two speciesbelonging to the Spaethianum clade and one singletonspecies. Yang et al. (2014) introduced a new species belongingto the Spaethianum clade named C. incanum. Colletotrichummelanocaulon was synonymized under C. dianesei (Vieiraet al. 2014). Many recent studies used multi-gene phylogenyto understand the phylogenetic divergence of Colletotrichumspecies. Since it is an expensive procedure as well as timeconsuming, Silva et al. (2012) stressed the need to use ‘power-ful genes’ such as ApMat and Apn25L. The ApMat markerprovides a better resolution, similar to the multi-gene markers,and has been a better gene in resolving species within theC. gloeosporioides species complex (Doyle et al. 2013;
Sharma et al. 2013; Hyde et al. 2014; Silva et al. 2012; Liuet al. 2015).
Von Schrenk and Spaulding (1903) proposed the nameGlomerella as the sexual morph of Colletotrichum and thishas been confirmed in cultural and molecular studies (Sutton1968; Crouch et al. 2009). A single name, eitherColletotrichumorGlomerella, has to be chosen to represent this taxon. Here werecommend the use of Colletotrichum because 1) theColletotrichum name/asexual morph is commonly associatedwith disease symptoms, while many Glomerella sexual morphstend to develop on dead host tissues (Sutton 1992) and thus areunderstudied in comparison to the asexual morphs (Cannonet al. 2012). 2) A comprehensive monograph onColletotrichum (von Arx 1957) and notes on species (Hydeet al. 2009b) has been published, while a comprehensive mono-graph for is not available. 3) There has been little morphologybased comparison of sexual taxa and many claimed asexual-sexual links are not based on authentic/type material. 4) In mostcases the sexual names are not typified according to modernpractices (Cannon et al. 2012). 5) There are no reliable reportsof sexual morph from any taxon within the truncatum clade(Cannon et al. 2012). Further, in the graminicola clade someindividual species such as C. falcatum, and C. graminicola areknown to produce sexual morphs, but other groups seem toform the sexual morph rarely or apparently not at all (Crouchand Beirn 2009; Cannon et al. 2012). 6) Colletotrichum hasmany more species epithets (772) than Glomerella (100) andis also the older generic name. 7) The number of Google scholarhits of Colletotrichum (60,700) is higher than Glomerella (13,500). 8) Colletotrichum is a well-established name in the plantpathological and plant breeder community (Cannon et al. 2012;Hyde et al. 2009a).
Hyde et al. (2009a, b) suggested the adoption of the genericname Colletotrichum in future revisions and this has beenfollowed in most subsequent studies (Cannon et al. 2012;Hyde et al. 2014).Collodiscula I. Hino & Katum., Bull. Faculty of Agriculture,Yamaguchi University 6: 55 (1955)
Collodiscula japonica I. Hino & Katum. is the type speciesof this monotypic genus. Samuels et al. (1987) observed sev-eral specimens that were associated with a conidiomatal fun-gus that is morphologically similar to the ascomatal initial ofC. japonica. This morphological similarity was then con-firmed by isolation of ascospores of C. japonica and the co-nidia of the asexual morph and observing that the culturesformed from both morphs were identical. They were unableto place this new asexual morph in any of the described generaand proposed Acanthodochium to accommodateA. collodisculae Samuels et al. Based on priorityAcanthodochium should be regarded as a synonymCollodiscula I. Hino & Katum. (Jaklitsch and Voglmayr2012; Stadler et al. 2013).
Fungal Diversity

Coniocessiaceae Asgari & Zare, Mycol. Progr. 10(2): 195(2011)
This family was introduced by Asgari and Zare (2011) andplaced in Xylariales and comprises a single genus,Coniocessia, with its type species C. nodulisporioides (D.Hawksw.) Dania García et al. forming with a nodulisporium-like asexual morph. Four new species were added by Asgariand Zare (2011).
Coniolariella Dania García et al., Mycol. Res. 110(11): 1285(2006)
Coniolariella was introduced by Dania García et al. (2006)with C. gamsii (Asgari and Zare) Dania García et al. as thetype species. Phylogenetic analysis placedConiolariellawith-in the Xylariales and its close relationship with rosellinia-likegenera (Checa et al. 2008) and was placed in Xylariaceae inStadler et al. (2013).Coniolariella hispanicaCheca, Arenal &J.D. Rogers and C. limoniispora var. limoniispora (Ellis &Everh.) Checa et al. have sporothrix-or geniculosporium-likeasexual morphs associated with stromata at different stages.
Conioscypha Höhn., Annls mycol. 2(1): 58 (1904)Conioscypha comprises eight terrestrial and freshwater
species found on decayed wood, leaves or bamboo stemsand also from skin scrapings and hair of living animals(Shearer 1973; Matsushima 1980, 1993, 1996). Réblováet al. (2004) introduced Conioscyphascus as the sexual morphof Conioscypha. Phylogenetic analyses confirmed a close re-lationship ofConioscyphawith Ascotaiwania and Carpoligna(Savoryellales), but the family and order relationships remainuncertain. The genus is therefore placed in Savoryellales gen-era incertae sedis.
Conlarium F. Liu & L. Cai, Mycologia 104(5): 1180 (2012)Liu and Cai (2012) described this genus with a single spe-
cies – Conlarium duplumascospora F. Liu & L. Cai based onmorphology andmolecular analyses. It was collected from sub-merged wood in Guangdong Province, China. The genus wasconsidered unique among genera in Annulatascaceae sensulato because of its unusual combination of morphological char-acters. Phylogenetic analysis also indicated that Conlarium hasaffinities with members in Annulatascaceae sensu lato, but thegenus could not be included in any existing genera.
The taxon is placed in Sordariomycetidae genera incertaesedis here.
Corallicola Volkm.-Kohlm. & Kohlm., Mycotaxon 44(2):418 (1992)
A monotypic genus of Halosphaeriaceae that wasestablished by Volkmann-Kohlmeyer and Kohlmeyer (1992)to accommodate the marine fungus,Corallicola nanaVolkm.-Kohlm. & Kohlm. The placement of the genus in theHalosphaeriaceae has not been confirmed at the molecular
level (Jones et al. 2009a). No asexual morph is known. Itcan easily be confused with Arenariomyces but differs in thecentrum pseudoparenchyma in lacking pit connections, asco-spore appendages that do not terminate in a spade-like struc-ture and the fact that it has only been found on dead coral(Volkmann-Kohlmeyer and Kohlmeyer 1992).
Corallomycetella Henn., Hedwigia 43: 245 (1904)This genus was revisited by Herrera et al. (2013a) and
shown to comprise two distinct well-separated clades inNectriaceae. Corallomycetella sensu stricto represented twospecies, C. elegans (Berk. & M.A. Curtis) C. Herrera & P.Chaverri and C. repens (Berk. & Broome) Rossman &Samuels while C. jatrophae (Möller) Rossman & Samuelswas placed in a new genus, Corallonectria, as C. jatrophae(Möller) C. Herrera & P. Chaverri.
Cordana Preuss, Linnaea 24: 129 (1851)Porosphaerella cordanophora E. Müll. & Samuels (gener-
ic type) was isolated in vitro and produced a Cordanapauciseptata Preuss (generic type) asexual morph in culture(Müller and Samuels 1982).Cordana is the older generic nameand Porosphaerella is therefore a synonym. The biological linkbetween Porosphaerella borinquensis F.A. Fernández &Huhndorf and Pseudobotrytis terrestris (Timonin) Subram.was proven by in vitro culture of ascospores (Fernández andHuhndorf 2004; Müller and Samuels 1982). Cordana belongsto the family Cordanaceae and based on sequence analysis,Hernández-Restrepo et al. (2015) recognized Cordanaceae isdistinct at ordinal level established Cordanales.
CorollosporaWerderm., Notizbl. Bot. Mus. Berlin-Dahlem 8:248 (1922)
A monotypic genus of Halosphaeriaceae that wasestablished by Werdermann (1922) to accommodateC. maritimaWerderm.Molecular studies showed that the genusis monophyletic and formed amoderately supported clade with-inHalosphaeriaceae (Campbell et al. 2002; Abdel-Wahab et al.2009; Sakayaroj et al. 2011b). Corollospora is currently themost speciose genus in marine habitats with 23 species. Themost distinctive feature of Corollospora species is their apicaland equatorial ribbon-like secondary appendages, which areformed by the fragmentation and peeling off of the exosporelayer and their associations with sand grains. They have asexualmorphs in species of Clavatospora, Halosigmoidea andVaricosporina (Jones et al. 2009b). Halosigmoidea was intro-duced to accommodate three marine hyphomycetous asexualmorphs that did not group with the type species of SigmoideaJ.L. Crane (type S. prolifera (R.H. Petersen) J.L. Crane);Corollospora has priority and is the accepted name.
Coronophoraceae Höhn., Sber. Akad. Wiss. Wien, Math.-naturw. Kl., Abt. 1 116: 624 (1907)
Fungal Diversity

Mugambi and Huhndorf (2010) revisited the orderCoronophorales using DNA sequence data and showedCoronophoraceae to be monophyletic.
Cosmospora Rabenh., Hedwigia 2: 59 (1862)Gräfenhan et al. (2011) treated Cosmospora in a narrow
sense, limiting it to a clade including the type, C. coccineaRabenh., which have acremonium-like asexual morphs andtend to occur on other fungi. Thus possible synonyms suchas Mariannaea and Volutella are recognised.
Cryptometrion Gryzenh. & M.J. Wingf., Australas. Pl. Path.39(2): 166 (2010)
This monotypic genus was introduced for CryptometrionaestuescensGryzenh. &M.J. Wingf. (Gryzenhout et al. 2010)from bark of Eucalyptus grandis collected in Sumatra. Itclosely related to Cryphonectria, Microthia and Holocryphiaand could be difficult to differentiate morphologically fromspecies in these genera. However, species of Cryptometrioncan be distinguished based on a combination of characteristicsof the stromata, the presence or absence of paraphyses andascospore septation. The morphology is similar to species inCryphonectriaceae with orange, globose to pulvinateconidiomata, and colour and shape of ascospores and conidia(Gryzenhout et al. 2005). ITS and β tubulin data shows thatthis fungus forms a dis t inct clade in the familyCryphonectriaceae (Gryzenhout et al. 2006).
Cryptosphaerella Sacc., Syll. fung. (Abellini) 1: 186 (1882)See under Scortechiniaceae.
Cucullosporella K.D. Hyde & E.B.G. Jones, Bot. Mar. 29(6):491 (1986)
A monotypic genus of Halosphaeriaceae that was in-troduced by Hyde and Jones (1986) to accommodate themarine fungus, C. mangrovei K.D. Hyde & E.B.G. Jones.Molecular phylogenetic analyses of the LSU rDNAshowed that C. mangrovei has affinity with Aniptoderalongispora K.D. Hyde and Antennospora quadricornuata(Jones et al. 2009b). No asexual morph is known (Hydeand Jones 1986).
Cumulospora I. Schmidt, Mycotaxon 24: 420 (1985)A monotypic asexual genus in the order Lulworthiales,
forming a sister clade to three species of Lulworthia (sensulato) with high statistical support (Abdel-Wahab et al. 2010).A second species, C. varia was transferred to a new genusMoromyces Abdel-Wahab et al. (Abdel-Wahab et al. 2010).
Cylindrocladiella Boesew., Can. J. Bot. 60(11): 2289 (1982)The sexual morph-typified genusNectricladiellawas intro-
duced with N. camelliae (Shipton) Crous & C.L. Schoch asthe type species (Schoch et al. 2000). These genera are linked
and Rossman et al. (2013) proposed that Cylindrocladiellashould be used over Nectricladiella, which is followed here.The genus is placed in Nectriaceae.
Cylindrotrichum Bonord., Handb. Allgem. mykol.(Stuttgart): 88 (1851)
The genus Reticulascus (Reticulascaceae), which is similartoChaetosphaeria, was introduced by Réblová et al. (2011a, b)based on multi-gene phylogenetic analysis and comprises twospecies; the type species is R. tulasneorum (Réblová & W.Gams) Réblová &W. Gams. The asexual morph is the generictype of Cylindrotrichum, C. oligospermum (Corda) Bonord.,and thus Cylindrotrichum and Reticulascus should be treatedas congeneric. Cylindrotrichum is the older name and shouldtake priority, and this selection is recommend here.
Cytospora Ehrenb., Sylv. mycol. berol. (Berlin): 2 (1818)Valseutypella and Valsella were synonymised under Valsa,
a genus of Valsaceae. Cytospora is the asexual morph of thesetaxa and the most commonly used name and the name used inrecent publications (Adams et al. 2005). We therefore use theolder name Cytospora in this treatment, but the mycologicalcommunity must decide which name should be used.
Daldinia Ces. & De Not., Comm. Soc. crittog. Ital. 1(4): 197(1863)
Daldinia concentrica (Bolton) Ces. & De Not. (basionymSphaeria concentrica) is the type species of this genus intro-duced by Cesati and De Notaris (1863) and incorporatesxylariaceous fungi with conspicuous glomerate stromata withconcentric zones. Several additional species of Daldinia werelater introduced by Hennings (1898, 1901, 1902) and Lloyd(1919, 1924a). The first successful and accurate monographonDaldiniawas presented by Ju et al. (1997). The presence ofspecific secondary metabolites is a characteristic feature in thisgenus (Hellwig et al. 2005; Stadler and Hellwig 2005).Several chemical metabolites with biological activities havebeen detected from the stromata by analytical methods(Stadler and Hellwig 2005; Stadler et al. 2014). RecentlyStadler et al. (2014) provided a world monograph using apolythetic approach to resolve the taxonomic positions andintergeneric affinities of Daldinia which confirms their place-ment in hypoxyloid Xylariaceae.
Annellosporium was introduced and typified byA. nemorosum M.L. Davey (Davey 2010). The genus wasdescribed based on morphology as well as rDNA ITS se-quence data. Phylogenetic analysis placed A. nemorosumclose to Daldinia loculata (Lév.) Sacc. The culture character-istics include common features with those of the D. petriniaeY.M. Ju et al. species complex (Stadler et al. 2014). Daldiniacultures from Canada were observed by Stadler et al. (2014)and the similarities in morphology with the A. nemorosumtype strain, which were more reminiscent of Daldinia
Fungal Diversity

loculatoides Wollw. & M. Stadler, were reported. Thus, thisnewly introduced monotypic genus can be regarded as a syn-onym of Daldinia (Stadler et al. 2014). Versiomyces was in-troduced by Whalley and Watling (1988) and referred to su-perficially similar toDaldinia, but justified as a distinct genusbecause the type species, Versiomyces cahuchucosusWhalley& Watling, lacks concentric zonations. However, the authorsthemselves mentioned the close relationship with Daldiniaregarding ascospore characters and size and regular shape ofstromata. Stadler et al. (2014) modified the generic descriptionof Daldinia sensu Ju et al. (1997) to accommodate azonatespecies and included Versiomyces as synonym.
Deightoniella S. Hughes, Mycol. Pap. 48: 27. 1952.Deightoniella as presently defined is heterogeneous
(Klaubauf et al. 2014). Klaubauf et al. (2014) based on anal-ysis of combined sequence data derived from D. Africana(from occurring on leaves of Imperata cylindrica var.africana) placed Deightoniella in the Pyriculariaceae.
Dialonectria (Sacc.) Cooke, Grevillea 12(no. 63): 77, 109(1884)
This genus was treated in a narrow sense around its typespecies D. episphaeria (Tode) Cooke by Gräfenhan et al.(2011) and belongs in Nectriaceae. A new species was intro-duced in Gräfenhan et al. (2011) but other species may becorrectly referred to Stylonectria Höhn (1915).
Diaporthe Nitschke, Pyrenomyc. Germ. 2: 240 (1870)With the need for a single scientific name for this pleo-
morphic genus, Diaporthe Nitschke (1870) has relative pri-ority over Phomopsis (Sacc.) Bubák and Kabát (1905), be-ing the older generic name (Wehmeyer 1933; Rossman et al.2007; Wingfield et al. 2012). Nitschke (1870), establishedthe genus Diaporthe based on the sexual morph, which istypified by Diaporthe eres Nitschke. Index Fungorum(2015) and MycoBank (Crous et al. 2004) list 901 and913 names of Diaporthe and 984 and 1040 names ofPhomopsis respectively; thus there is little difference innumbers of species epithets. Diaporthe is also the type ofthe Diaporthaceae and Diaporthales and this adds an extrastrength to the argument for using this name. The only com-prehensive monograph of the genus (Wehmeyer 1933), usedthe name Diaporthe which has been the taxonomic accountfollowed by most mycologists. The arguments in favour ofusing Diaporthe against Phomopsis have been putforwarded in numerous publications (Santos and Phillips2009; Santos et al. 2010; Udayanga et al. 2012; Wingfieldet al. 2012; Gomes et al. 2013; Rossman et al. 2014;Udayanga et al. 2014a, b) and the taxonomy of the genushas progressed considerably in the last few years.
Arguments in favour of Phomopsis against Diaporthehave also been put forward. There are a relative greater
number of applications of Phomopsis in the recent literaturewhich can be seen in Google citations. Some field plantpathologists also prefer Phomopsis. However, most of theimportant phytopathogens are now linked to Diaporthebased on molecular data and there are no apparentproblems in adopting this one name. Saccardo andRoumeguère (1884) defined Phomopsis as a group ofPhoma species that could be separated by having beta co-nidia; he also recognised the link to the sexual morphDiaporthe. Bubák (1905) accepted Saccardo’s concept ofPhomopsis and transferred one species of Phoma (Phomalactucae) to Phomopsis. He did not provide a generic de-scription, but included a detailed description of a specimencollected in Tyrol. Later in the same year Saccardo raisedPhomopsis to generic rank. Although Riedl and Wechtl(1981) formally proposed the retention of Phomopsis againstMyxolibertella Hohnel (1903) with P. lactucae as the generictype, this has not been revisited in recent studies. However, are-definition of the generic type of Diaporthe, D. eresNitschke by Udayanga et al. (2014b), and in parallel, theproposal to conserve the name D. eres by Rossman et al.(2014) over its obscure synonyms will stabilise the use ofthe name. The limits of Diaporthe are now well-definedbased on recent molecular data with respect to its type spe-cies and important phytopathogens (Udayanga et al. 2014b).
Until recently, host affiliated nomenclature has largelybeen applied in naming Diaporthe and Phomopsis species(Uecker 1988). The Phomopsis names are accumulatedmostly in terms of gross morphology, resembling the conid-ial shapes, and some of them also later identified as Phomalike species (Udayanga et al. 2014a). Although the asexualPhomopsis morph is generally found in nature, and routinelyisolated in plant pathological surveys, many of the importantplant pathogens have also been linked to their Diaporthenames in modern treatments . Examples includeD. ampelina (Berk. & M.A. Curtis) R.R. Gomes et al. (deadarm of grapes), D. helianthi Munt.-Cvetk. (sunflower can-ker), D. citri (citrus melanose), D. longicolla (Hobbs) J.M.Santos (soybean seed decay) and D. vaccinii Shear (blueber-ry cankers). Therefore it is practical to adopt in plant patho-logical studies Diaporthe names and it has been practiced inrecent reports (Baumgartner et al. 2013; Elfar et al. 2013).Therefore we suggest using Diaporthe over Phomopsis,which will lead to resolution of nomenclatural problems inmodern treatments largely based on molecular data and spe-cies epithets should be carefully selected (Rossman et al.2013; Udayanga et al. 2014a).
Diatrypasimilis J.J. Zhou & Kohlm., in Chalkley et al.,Mycologia 102(2): 432 (2010)
Chalkley et al. (2010) introduced this genus based on a culturethat was isolated from decaying Rhizophora wood collected inAustralianmangroves. Although the ribosomal DNAgeneswere
Fungal Diversity

sequenced and the fungus was characterized in culture, the au-thors did not fully describe the morphology of the fungus onnatural substrates, apparently because of a lack of material.This species was recently collected on decayed wood in SaudiArabia and fresh cultures established (Abdel-Wahab et al. 2014).LSU sequence data place this species in the Diatrypaceae.
Didymostilbe Henn., Hedwigia 41: 148 (1902)Peethambara sundara Subram. & Bhat, the type species
of Peethambara, has been linked to Didymostilbe sundara(Subram. & Bhat) Seifert by culture of single spores(Subramanian and Bhat 1978). Molecular studies also showedthat Didymostilbe echinofibrosa (Finley) Rossman clusteredtogether with Peethambara sundara in the Bionectriaceae(Rossman et al. 2001). The type of Didymostilbe, D. coffeaeHenn., has however, not been linked toPeethambara and bothnames should therefore be retained pending further analysis.
Dinemasporium Lév., Annls Sci. Nat., Bot., sér. 3 5: 274(1846)
Crous et al. (2012a) restudied the genusDinemasporium andrelated genera with appendaged conidia and placed the genus inChaetosphaeriaceae based onmorphology andmolecular data.They also epitypified the type speies, D. strigosum Lev., thetype of this genus. Brunneodinemasporium, Codinaeopsis,Dendrophoma, Dictyochaeta, Menispora, Pseudolachnea,Pyrigemmula, Thozetella and Zignoella which also clusteredin Chaetosphaeriaceae. Liu et al. (2015) introduced a newspecies, D. nelliana W.J. Li et al.
Diplococcium Grove, J. Bot., Lond. 23: 167 (1885)FiveHelminthosphaeria species have been associated with
Diplococcium asexual morphs (Samuels et al. 1997; Réblová1999a). However, the genus is polyphyletic and has includedspecies having affinities with members of Helotiales,Pleosporales and Venturiales. The type species, D. spicatumGrove clusters within Helotiales, but the sequences are notfrom ex-type strains (Shenoy et al. 2010).Discosia Lib., Pl. crypt. Arduenna, fasc. (Liège) 4: no. 346(1837)
Molecular analysis of combined gene sequence data,showed Discosia was not monophyletic and separated intothree d is t inc t l ineages , Adisc i so , Discos ia andImmersidiscosia (Tanaka et al. 2011).
Discosphaerina Höhn., Sber. Akad. Wiss. Wien, Math.-naturw. Kl., Abt. 1 126(4–5): 353 (1917)
The genus Discosphaerina was introduced by Von Höhnel(1917a, b) based on D. discophora Höhn. The placement ofthis genus is still confused as the unitunicate nature ofD. discophora is unclear and some members of this genushave bitunicate asci (Sivanesan 1984a, b; Thambugala et al.2014). Sivanesan (1984a, b) treated Discosphaerina in
Dothideaceae (Dothideales) and some Discosphaerina spe-cies produce Aureobasidium, Kabatia, Sarcophoma andSelenophoma asexual morphs in culture (Sivanesan 1984a,b; Thambugala et al. 2014). Lumbsch and Huhndorf (2010)included Discosphaerina in Hyponectriaceae. The phyloge-netic placement of Discosphaerina (= Columnosphaeria fagi(H.J. Huds.) M.E. Barr) was confirmed in Dothideaceae(Dothideales) (Schoch et al. 2006, 2009; Zalar et al. 2008;Thambugala et al. 2014). Thambugala et al. (2014) examinedthe types of Discosphaerina and D. fagi in order resolve thisproblem and suggested to place Discosphaerina inHyponectriaceae based on its morphology. No molecular datais available in GenBank except for Discosphaerina fagi.Therefore, recollection, epitypification and molecular analysisare required to confirm the placement of Discosphaerina andits species.
DiscostromaClem., Gen. fung. (Minneapolis): 50, 173 (1909)Species of Discostroma and Seimatosporium have been
linked by molecular data (Tanaka et al. 2011), however themolecular links have not been proven for the types of eithergenera. Both names should therefore be retained pending freshcollections of the types and a molecular study.
Disculoides Crous et al., in Crous et al., Persoonia, Mol. Phyl.Evol. Fungi 28: 71 (2012)
The new coelomycetous genus has two species both caus-ing leaf spots of Eucalyptus. Molecular data places this genusin Diaporthales genera incertae sedis (Crous et al. 2012d).
Diversimorbus S.F. Chen & Jol. Roux, Fungal Biology 117:300 (2014)
Chen et al. (2013a) described this monotypic genus based onD. metrosiderotis S.F. Chen & Jol. Roux. Morphology andmulti-gene phylogenetic analyses show it to group in theCryphonectriaceae (Chen et al. 2014). It was observed on na-tive Metrosideros angustifolia in Western Cape Province ofSouth Africa and resembles a Eucalyptus pathogen. The spe-cies shares characteristics of taxa within Cryphonectriaceae inturning yellow in lactic acid and purple in 3 % KOH(Castlebury et al. 2002; Gryzenhout et al. 2009). In addition,isolates of the new genus had black conidiomata, which distin-guishes it frommost genera in theCryphonectriaceae that haveorange conidiomata (Gryzenhout et al. 2009, 2010; Begoudeet al. 2010; Chen et al. 2011, 2012; Vermeulen et al. 2011).Genera in the Cryphonectriaceae with black conidiomata in-clude Aurapex, Celoporthe, and Chrysoporthe (Gryzenhoutet al. 2005, 2009; Nakabonge et al. 2006). Multi-gene phylo-genetic analyses for this species also clearly represent a previ-ously undescribed genus in Cryphonectriaceae.
DrechmeriaW. Gams & H.-B. Jansson, Mycotaxon 22(1): 36(1985)
Fungal Diversity

This genus was introduced byGams and Jansson (1985). In2014, Quandt et al. showed in nematophagous subclade, thegenus Haptocillium clusters together with Drechmeria andpropose to protect older Drechmeria over Haptocillium.
Durotheca Læssøe et al., IMA Fungus 4(1): 62 (2013)The genus Durotheca was introduced with D. depressa
Læssøe & Srikitikulchai as the type species (Læssøe et al.2013). Two further species described earlier in Theisseniawere transferred as D. comedens (Ces.) Læssøe & Srikitik.and D. rogersii (Y.M. Ju & H.M. Hsieh) Læssøe & Srikitik.in the same study. This genus is characterized by bipartitecarbonaceous stromata, erumpent through wood or bark, cov-ered with white pruina. The asexual morph, where known, isreported as nodulisporium-like (Læssøe et al. 2013). A mo-lecular phylogenetic study placed Theissenia pyrenocrata(Theiss.) Maubl. in a sister clade to isolates of Durotheca asa basal lineage. However, the phylogenetic placement withinXylariaceae in relation to either the subfamilies Xylarioideaeor Hypoxyloideae is yet to be resolved (Læssøe et al. 2013).
Dyrithiopsis L. Cai et al., in Jeewon et al., Mycologia 95(5):912 (2003)
Jeewon et al. (2003a) reported a new genus, Dyrithiopsiswith an asexual morph, Monochaetiopsis (also a new genus)based on cultural studies. We propose to adopt the sexualtypified name over the asexual typified.
Ebullia K.L. Pang, in Chu et al., Mycoscience 56: 40 (2015)Chu et al. (2015) undertook a molecular reappraisal of
Nimbospora, a genus with three marine species, includingrecent collections from Taiwan. The genus was shown to bepolyphyletic with Nimbospora octonae Kohlm. distantlyplaced from the type species N. effusa and a second speciesN. bipolaris. Chu et al. (2015) therefore transferredN. octonaeto the new genus Ebullia (Halosphaeriaceae) as E. octonae(Kohlm.) K.L. Pang. The genus has no known asexual morph.
Echinosphaeria A.N. Mill. & Huhndorf, Mycol. Res. 108(1):29 (2004)
There are seven named species in the genus, but only thetype, Echinosphaeria canescens (Pers.) A.N. Mill. & Huhndorfis represented by gene sequences (Miller et al. 2014) whichplaces the genus in Helminthosphaeriaceae. AlthoughE. canescens has been associated with Endophragmiella andSelenosporella-like synanamorphs (Hughes 1979; Sivanesan1983), there is no molecular data to establish this connection.
Emericellopsis J.F.H. Beyma, Antonie van Leeuwenhoek 6:264 (1940) [1939]
This genus was introduced by Beyma (1940), withEmericellopsis terricola J.F.H. Beyma as the type species,
and placed in the Bionectriaceae (Hypocreales) and has bothmarine and terrestrial species (Rossman et al. 2001).Emericellopsis terricola clustered in a distinct clade withinthe Sclerotigenum/Geosmithia-clade in Hypocreales inSummerbell et al. (2011).
Engleromyces Henn., Bot. Jb. 28(3): 327 (1900)Engleromyces (Xylariaceae) was introduced by Hennings
(1900) for a single species, E. goetzei Henn.; the genus ischaracterized by massive, hard stromata (Whalley et al.2010). Chinese collections, originally identified asE. goetzei, were later found to represent a second species,E. sinensis M.A. Whalley et al. (Whalley et al. 2010).Engleromyces produces cytochalasin D and 19, 20-epoxycytochalasin and a novel metabolite in E. sinensis,neoengleromycin, which is used for cancer treatments(Pedersen et al. 1980; Liu et al. 2002).
Equicapillimyces S.S.Y.Wong et al., VeterinaryMicrobiology115(2–4): 406 (2012)
Wong et al. (2011) described this monotypic genus frombrittle tail syndrome of horses, which causes weakening andbreakage of the tail hairs; the type species is E. hongkongensisS.S.Y. Wong et al. Molecular studies showed that this taxonbelongs in the Ophiostomataceae, however, in our analysis itbelongs in Lulworthiales. This septate branching, hyalinemould, grows optimally at 30 C. Hyphae fill the core of in-fected hair shafts with short necked ascomata containingbanana-shaped septate ascospores.
Erythrogloeum Petr., Sydowia 7(5–6): 378 (1953)Crous et al. (2012d) recollected this genus from South and
Central America where it causes leaf spots on Hymenaea.Molecular data placed the genus in Diaporthales generaincertae sedis. There are presently two species included inthe genus (Index Fungorum 2015).Escovopsis J.J. Muchovej & Della Lucia, Mycotaxon 37: 192(1990)
The phylogenetic analyses of ITS and LSU by Augustinet al. (2013) demonstrate the monophyly of the genusEscovopsis, agreeing with other phylogenetic studies (Currieet al. 2003; Taerum et al. 2007). The type species E. weberiJ.J. Muchovej & Della Lucia and other Escovopsis speciescluster in the Hypocreaceae.
Escovopsioides H.C. Evans & J.O. Augustin, PLoS ONE7(12): e51392, 6 (2013)
This genus was introduced by Augustin et al. (2013) based onEscovopsioides niveaH.C. Evans & J.O. Augustin isolated fromthe fungal garden of Acromyrmex (leaf cutting ants) in Brazil. Inculture it produces a brush-like asexual morph with phialides onwell-defined vesicles in culture. It differs from Escovopsis by the
Fungal Diversity

absence of pigmentation, the lageniform phialides produced onterminal and intercalary, globose vesicles, the hyaline, smoothconidia in long chains, as well as sequence data (Augustin et al.2013). According to the phylogenetic analyses of ITS and LSUsequence data this genus belongs in Hypocreaceae.
Etheirophora Kohlm. & Volkm.-Kohlm., Mycol. Res. 92:414 (1989)
A genus in the Hypocreomycetidae (order incertae sedis)(Jones et al. 2014), initially assigned to the Sphaeriales byKohlmeyer and Volkmann-Kohlmeyer (1989) and to theHalosphaeriales by Hawksworth et al. (1995), and Kirket al. (2001), and the TBM clade (Hypocreomycetidaeincertae sedis) (Sakayaroj et al. 2005; Schoch et al. 2007).
Etheirophoraceae Rungjindamai et al., CryptogamieMycologie 35: 134 (2014)
This family was introduced to accommodate the genera:Etheirophora (E. bijubata Kohlm. & Volkm.-Kohlm.,E. blepharospora (Kohlm. & E. Kohlm.) Kohlm. & Volkm.-Kohlm., E. unijubata Kohlm. & Volkm.-Kohlm.) andSwampomyces (S. armeniacus Kohlm. & Volkm.-Kohlm.,S. triseptatus K.D. Hyde & Nakagiri), that grouped togetherin a well-supported clade based on LSU and SSU sequences(Jones et al. 2014). Further taxon sampling is required to de-termine its ordinal position within the Hypocreomycetidae.
Falcocladiaceae Somrithipol et al., Cryptogamie Mycologie35: 134 (2014)
The family was introduced to accommodate four species ofFalcoc ladium , tha t g rouped wi th o the r taxa inHypocreomycetidae, order incertae sedis (Jones et al. 2014),for which the order Falcocladiales is introduced in this paper.
Falcocladium S.F. Silveira et al., Mycotaxon 50: 447 (1994)Falcocladium was introduced by Crous et al. (1994) with
F. multivesiculatum S.F. Silveira et al. as the type species.Subsequently, three other species were described:F. sphaeropedunculatum Crous & Alfenas (Crous et al.1997), F. thailandicum Crous & Himaman (Crous et al.2007) and F. turbinatum Somrith. et al. (Somrithipol et al.2007). This genus occurs on a wide range of substrata includ-ing Eucalyptus grandis, E. camaldulensis leaves, and leaf lit-t e r co l lec ted f rom trop ica l fo res t s . The fami lyFalcocladiaceae was recently introduced to accommodatethe genus and Falcocladiales is introduced in this paper.
Fimetariella N. Lundq., Botaniska Notiser 117: 239 (1964)A genus of Lasiosphaeriaceae with Fimetariella
rabenhorstii N. Lundq. as the type species. Phylogenetically,based on LSU nrDNA and β-tubulin sequences, the type spe-cies groups on a long branch in a clade with significant
support as a sister taxon to Podospora appendiculata(Auersw. ex Niessl) Niessl and Cercophora scortea (Cain)N. Lundq. (Kruys et al. 2015).
Fluminicola S.W. Wong et al., Fungal Diversity Res. Ser. 2:190 (1999)
Wong et al. (1999) placed this genus in Annulatascaceaebased onmorphology. However, according to phylogenetic anal-yses using a combined LSU, SSU andRPB2 datasetFluminicolaclusters in the family Papulosaceae (Réblová 2013a).
Fluviatispora K.D. Hyde, Mycol. Res. 98(7): 720 (1994)A genus of three species found growing on palm material
in aquatic habitats, with no known asexual morph, and placedin the Halosphaeriaceae. However, molecular data are re-quired to support this placement.
FrondicolaK.D. Hyde, J. Linn. Soc., Bot. 110(2): 100 (1992)A monotypic marine taxon based on morphological obser-
vations with no known asexual morph. Frondicola was intro-duced and placed in Clypeosphaeriaceae by Hyde (1992),while Lumbsch and Huhndorf placed it in Annulatascaceaeand Jones et al. (2009b) placed it in Hyponectriaceae.Molecular data are required to establish its taxonomic posi-tion; Jones et al. (2009b) is followed here.
Fulvocentrum E.B.G. Jones & Abdel-Wahab, CryptogamieMycologie 35(2) 132 (2014)
The genus was introduced by Jones et al. (2014) to accom-modate two species of Swampomyces that did not group withthe type species (S. armeniacus Kohlm. & Volkm.-Kohlm.):F. aegyptiacus (Abdel-Wahab et al.) E.B.G. Jones & Abdel-Wahab and F. clavatisporium (Abdel-Wahab et al.) E.B.G.Jones & Abdel-Wahab. Both species are tropical marine taxaand are very common on intertidal wood of Avicennia marinain Red Sea mangroves, both on the Egyptian coast (Abdel-Wahab 2005) and from Arabian Gulf mangroves of the SaudiArabia coast (Abdel-Wahab et al. 2014). The genus groupswith high statistical support in the family Juncigenaceae,Hypocreomycetidae order incertae sedis, (Jones et al. 2014);it has no known asexual morph.
Funiliomyces Aptroot, Stud. Mycol. 50(2): 309 (2004)The genus Funiliomyces contains a single species,
F. biseptatus Aptroot, which was isolated from aBromeliaceae leaf in Brazil and is characterized by thetorpedo-shaped ascospores with two nearly central septa andone polar and one median appendage (Aptroot 2004). It isplaced inAmphisphaeriaceae based onmolecular data, group-ing with Rosellinia and Arecophila in a moderately supportedsister clade (Aptroot 2004), but further collections are requiredto confirm this position.
Fungal Diversity

Fusarium Link, Mag. Gesell. naturf. Freunde, Berlin 3(1–2):10 (1809)
Rossman et al. (2013) stated that there is no question thatFusarium and Gibberella are synonyms and proposed thatGibberella be suppressed in favour of Fusarium; this isfollowed here. Fusarium belongs in Nectriaceae and com-prises several species complexes, not yet fully resolved(Hyde et al. 2014).
Geejayessia Schroers et al., in Schroers et al., Stud. Mycol.68(1): 124 (2011)
This genus was introduced by Schroers et al. (2011) basedon morphology and phylogenetic analysis and comprises fivespecies, four transferred from Nectria sensu lato, and withGeejayessia cicatricum (Berk.) Schroers as the type species.Based on molecular studies the genus it is placed inNectriaceae.
Gesasha Abdel-Wahab & Nagah., Nova Hedwigia 92(3–4):501 (2011)
This is a genus ofHalosphaeriaceae established by Abdel-Wahab and Nagahama (2011) to accommodate three marinespecies and is typified by G. peditatus Abdel-Wahab &Nagahama. Phylogenetic analyses of the SSU and LSUrDNAgene data placed these species in a well-supported basalclade in the family Halosphaeriaceae along with species ofArenariomyces andCorollospora; no asexual morph is known(Abdel-Wahab and Nagahama 2011).
GibellulopsisBat. & H.Maia, Anais da Sociedade de Biologiade Pernambuco 16 (1): 153 (1959)
The genus was introduced to accommodate Verticilliumpiscis (Bat. & H. Maia) Neish & G.C. Hughes, a fish patho-gen, which did not group in Verticillium sensu stricto (types p e c i e s Ve r t i c i l l i u m d a h l i a e K l e b . ) i n t h ePlectosphaerellaceae. Further species assigned to the genusare G. nigrescens (Pethybr.) Zare et al. (Zare et al. 2007), andG. chrysanthemi Hirooka et al. (Hirooka et al. 2014). Otherspecies may well be related taxa: Acremonium furcatum(Moreau & V. Moreau) ex W. Gams, A. stromaticum W.Gams & R.H . S tove r, A. nepa l ense W. Gams ,Cephalosporium serrae Mafei and Gliocladium cibotiiJ.F.H. Beyma (Zare et al. 2007).
Gliocephalotrichum J.J. Ellis & Hesselt., Bull. Torrey bot.Club 89: 21 (1962)
Leuconectria was introduced as the sexual morph of thetype species of Gliocephalotrichum by Rossman et al.(1993) . Rossman et al . (2013) proposed that asGliocephalotrichum species are widely reported from soilsand that Leuconectria is relatively obscure with only two spe-cies. Leuconectria is suppressed in favour of the older asexualmorph-typified name Gliocephalotrichum.
Gliocladiopsis S.B. Saksena, Mycologia 46: 662 (1954)Rossman et al. (2013) documented the history and us-
age of Gliocladiopsis versus Glionectria and concludedthat the genus name Gliocladiopsis should be protectedover the Glionectria, for these soil-borne fungi (Lombardand Crous 2012).
Gliomastix Guég., Bull. Soc. mycol. Fr. 21: 240 (1905)Summerbell et al. (2011) revised Gliomastix with five spe-
cies based on phylogenetic data which showed 14 strains clus-tering as a distinct clade in the Bionectriaceae.
Glomerellaceae Locq. ex Seifert & W. Gams, in Zhang et al.,Mycologia 98(6): 1083 (2007) [2006]
The family Glomerellaceae was invalidly published byLocquin (1984), validated in Zhang et al. (2006), and it wasaccepted as one of the three families of Glomerellales inRéblová et al. (2011a).
Glomerulispora Abdel-Wahab & Nagah., in Abdel-Wahabet al., Mycol. Progr. 9(4): 552 (2010)
An asexual genus in the family Torpedosporaceae(Hypocreomycetidae order incertae sedis) based on an ex-type culture (Jones et al. 2014). Abdel-Wahab et al. (2010)introduced this genus for species with irregularly helicoidmuriform conidia, which groups with species ofTorpedospora, with variable support. However, it is not con-generic with Torpedospora.
Gondwanamyces G.J. Marais & M.J. Wingf., Mycologia90(1): 139 (1998)
Asexual morphs of Gondwanamyces are distinct inpossessing erect, darkly pigmented conidiophores,monoverticillate or divergently penicillate, with whorls ofphialides, producing hyaline conidia. Gondwanamycesproteae has a Custingophora asexual morph (Mouton et al.1993). Based on SSU and a combined dataset of LSU, SSUand RPB2, species of Gondwanamyces and theirCustingophora asexual morphs form a highly monophyleticclade which is sister to Ceratocystidaceae (Réblová et al.2011a).
Gondwanamycetaceae Réblová et al., Stud. Mycol. 68(1):188 (2011)
This family is placed in Microascales based on SSU and acombined dataset of LSU, SSU and RPB2. Réblová et al.(2011a) includes also the asexual morph genus Custingophora.
Graphiaceae de Beer et al., CBS fungal biodiversity series 12:1–19 (2013)
Based on the phylogenetic distance betweenGraphium andother families of Microascales, the family Graphiaceae wasintroduced by de Beer et al. (2013). The family is
Fungal Diversity

monophyletic (Lackner et al. 2014) and comprisesGraphium,which is a synnematous hypomycete described by Corda(1837) with the type G. penicillioides Corda.
Graphostromataceae M.E. Barr et al., Mycotaxon 48: 533(1993)
This family comprises Graphostroma, a genus intro-duced in the family Calosphaeriaceae by Barr (1985) to-gether with seven other genera. Barr et al. (1993) assimi-lated information on these genera into a revised scheme forthe classification of Calosphaeriaceae which gave rise tothe introduction of a separate monotypic familyGraphostromataceae in the order Xylariales. The presenceof a nodulisporium-like conidial morph and a diatrype-likesexual morph featuring allantoid ascospores supported theintroduction of the new family. Stadler et al. (2013, 2014)used ITS sequence data from Graphostroma platystoma(Schwein.) Piroz., the type species of the genus, to evaluatethe relationship with other representative genera ofXylariaceae and showed it was basal and more closelyrelated to Biscogniauxia and Camillea than to Diatrype(Stadler et al. 2013). However, to clarify the exact phylo-genetic position of this genus further molecular studies arerequired, with inclusion of protein coding genes.
Greeneria Scribn. & Viala, C. r. hebd. Séanc. Acad. Sci., Paris105: 473 (1887)
Tangthirasunun et al. (2014) introduced a new species ofGreeneria, G. saprophytica Tangthirasunun et al., bringingthe number of species in the genus to three (IndexFungorum 2015). Phylogenetic analysis placed the new spe-cies in Diaporthales genera incertae sedis.
Griphosphaerioma Höhn., Ber. dt. bot. Ges. 36: 312 (1918)Shoemaker (1963) established the link between
Griphosphaerioma kansensis (Ellis & Everh.) Shoemakerand its asexual morph Labridella cornu-cervae Brenckle, thetype species of both genera. Griphosphaerioma is the oldestname and should have priority.
Groenhiella Jørg. Koch et al., BotanicaMarina 26: 265 (1983)Monotypic marine genus growing superficially on driftwood
with no known asexual morph. Koch et al. (1983) referred thegenus to the Nitschkiaceae (Coronophorales), but further collec-tions, isolates and sequences are required to confirm thisplacement.
Gynonectria Döbbeler, Mycol. Progr. 11(2): 474 (2012)Gynonectria is a monotypic genus of Hypocreales intro-
duced by Döbbeler (2012) for a species with relatively largeperithecia, forming individually within the perianths of thehepaticOdontolejeunea sp. (Lejeuneaceae, Porellales), which
grow in the phyllosphere of living vascular plants in centralAmerican rainforests.
Gyrostroma Naumov, Bull. Soc. mycol. Fr. 30(3): 386 (1914)Hirooka et al. (2012) examined the type specimen of
Gyrostroma, G. sinosum Naumov, and determined it not tobe a hypocrealean taxon and suggested that it may be a mem-ber of Diaporthales.
Haiyanga K.L. Pang & E.B.G. Jones, The Raffles Bull. Zool.19: 8 (2008)
Haiyanga was introduced to accommodate the taxonArenariomyces salinusMeyers, a species with a much confusedtaxonomy, as shown by its list of synonyms (Index Fungorum2015). A molecular study of the genera Antennospora andHalosphaeria found that it forms a well-supported monophy-letic clade with Arenariomyces trifurcatusHöhnk, the type spe-cies of the genus (Pang et al. 2008). A new genus Haiyangawas established by Pang et al. (2008) for Antennospora salinabased on the morphological differences between it andArenariomyces trifurcatus. Further collection and a wider rangeof genes need to be sequenced to resolve the taxonomic statusof Haiyanga salina. No asexual morph is known.
Halazoon Abdel-Aziz et al., Mycol. Prog. 9: 545 (2010)Asexual genus of Lulworthiaceae that was established by
Abdel-Wahab et al. (2010) with the type species, H. melhaeAbdel-Aziz et al. Molecular study based on SSU and LSUrDNA of an ex-type strain of H. melhae placed it in a well-supported clade within Lulworthiaceae with species ofLulworthia and Cirrenalia fusca I. Schmit. The latter specieswas transferred to Halazoon as H. fuscus (I. Schmidt) Abdel-Wahab et al. Species of Halazoon have been recorded ondriftwood in the intertidal zone in marine habitats (Abdel-Wahab et al. 2010).
Haligena Kohlm., Nova Hedwigia 3: 87 (1961)Monotypic genus ofHalosphaeriaceae that was introduced
by Kohlmeyer (1961) to accommodate the marine fungus,H. elaterophora Kohlm. Currently the genus is monotypicas other species have been transferred to other genera (Joneset al. 2009b). Molecular data confirms its position in the fam-ily Halosphaeriaceae in a basal clade, with Nautosphaeriaand Tubakiella in a sister group (Sakayaroj et al. 2011a;Pang and Jheng 2012a, b).
Haloguignardia Cribb & J.W. Cribb, Univ. Queensl., Pap.Dept. Bot. 3: 97 (1956)
Genus of Lulworthiaceae that was established by Cribband Cribb (1956) to accommodate H. decidua Cribb & J.W.Cribb. and three other species. Another two species wereadded later. Species ofHaloguignardia are parasitic on brownseaweed in the Fucales, especially species of Cystoseira,
Fungal Diversity

Halidrys and Sargassum. Molecular study based on SSUrDNA placed H. irritans (Setch. & Estee) Cribb & J.W.Cribb in a highly supported clade within Lulworthiaceaealong with Lulwoana and Lulworthia species (Inderbitzinet al. 2004; Campbell et al. 2009; Harvey and Goff 2010).
Halonectria E.B.G. Jones, Trans. Br.Mycol. Soc. 48: 287 (1965)Monotypic marine genus in the family Bionectriaceae
(Rossman et al. 1999; Jones et al. 2009b), although not typicalof Hypocreales due to the immersed ascomata, with longnecks and elongate non-septate ascospores. Further studyand sequence data are required to confirm its assignment toBionectriaceae. The asexual morph is pycnidial.
Halosarpheia Kohlm. & E. Kohlm., Trans Br. Mycol. Soc.68: 208 (1977)
The genus was established by Kohlmeyer and Kohlmeyer(1977) to accommodate a marine fungus,H. fibrosaKohlm. &E. Kohlm. Later, similar species with polar uncoiling append-ages were described from both freshwater and marine habitats.Recent molecular phylogenetic studies showed that the genusis polyphyletic and several species were transferred to existingor new genera. Only one species,H. japonicaAbdel-Wahab&Nagah. produced helicoid conidia as an asexual morph. Thesexual/asexual connection was established at morphologicaland molecular level (Abdel-Wahab and Nagahama 2012). Thegenus is based on morphological observations and moleculardata with the three species forming amonophyletic group withLignincola tropica Kohlm. as a sister group (Sakayaroj et al.2011a). Six Halosarpheia species have not been sequenceddue to the lack of cultures. Since other Halosarpheia specieshave been transferred to new genera followingmolecular anal-ysis, we refer to these six species as Halosarpheia sensu latountil further collections, isolations and sequences are generat-ed to test their position in the genus.
Halosphaeria Linder, Farlowia 1:412 (1944)Halosphaeria was introduced by Barghoorn and Linder
(1944a) to accommodate a marine species, H. appendiculataLinder, growing on wood and is the type species of theHalosphaeriaceae. Molecular data show it grouping withLignincola laevis Höhnk with moderate support (Sakayarojet al. 2011a). No asexual morph is known.
Halosphaeriopsis T.W. Johnson, J. Elisha Mitchell scient.Soc. 74: 44 (1958)
A monotypic genus in the Halosphaeriaceae based on mor-phological andmolecular data.Culcitalna is a monotypic genussynonymised under Trichocladium by Dixon (1968), with asexual morph as Halosphaeria mediosetigera Cribb & J.W.Cribb (now Halosphaeriopsis mediosetigera (Cribb & J.W.Cribb) T.W. Johnson). Culcitalna achraspora Meyers & R.T.Moore was described as possessing conidia produced in
sporodochia (Meyers and Moore 1960), but subsequent collec-tions showed they produced conidia freely on the substrate, andthe taxon was thus transferred to Trichocladium alopallonellum(Meyers & R.T.Moore) Kohlm. &Volkm.-Kohlm., the asexualmorph of Halosphaeriopsis mediosetigera based on culturalstudies and molecular data. We therefore treat Culcitalna as asynonym of Halosphaeriopsis.
Hapsidascus Kohlm. & Volkm.-Kohlm., Syst. Ascomycetum10: 113 (1991)
A monotypic marine genus referred to Ascomycota generaincertae sedis (Jones et al. 2009b), which needs to be recol-lected, isolated and sequenced. Ascomata perithecoid, large,deeply embedded in mangrove roots of Rhizophora mangle.May not be a member of the Sordariomycetes.
Harknessiaceae Crous, in Crous et al., Persoonia, Mol. Phyl.Evol. Fungi 28: 55 (2012)
The family Harknessiaceae was introduced to accommo-date species of Harknessia with their Wuestneia-like sexualmorphs and belongs in the order Diaporthales. Byestablishing the family Harknessiaceae, Crous et al. (2012e)avoided replacing Wuestneia, as the family is based on theasexual genus Harknessia. The placement of Wuestneia withits type species, W. aurea Auersw. has not been establishedand is presently placed in Melanconidaceae.
Harposporium Lohde, Tagbl. Versamml. Ges. Deutsch.Naturf. 47: 206 (1874)
Harposporium is the largest and oldest nematode trappingasexual genus in Hypocreales (Quandt et al. 2014). Chaverriet al. (2005) observed the asexual-sexual link betweenHarposporium and Podocrella. Harposporium is the oldestname and considering several other reasons, Quandt et al.(2014) proposed to use Harposporium over Podocrella.
Hispidicarpomyces Nakagiri, Mycologia 85: 639 (1993)M o n o t y p i c m a r i n e g e n u s i n t h e f a m i l y
Hispidicarpomycetaceae that needs recollection, isolationand sequencing to determine it phylogenetic position in theAscomycota (Jones et al. 2009b). Ascomata very large (800–2100 μm), immersed in the thallus of the red alga Galaxurafa lca ta . The genera Spathu lospora , Retros t ium(Spa t h u l o s p o r a c e a e ) a n d Hi s p i d i c a r p omy c e s(Hisp id icarpomyce taceae ) were re fe r red to theSpathulosporales, but molecular data placed the genusSpathulospora (S. adelpha, S. antartica) in the orderLulworthiales (Inderbitzin et al. 2004). Consequently, theplacement of Hispidicarpomyces and Retrostium cannot beresolved until further phylogenetic studies are undertaken.The asexual morph has verticillate spermodochia.
Humicola Traaen, Nytt Mag. Natur. 52: 31 (1914)
Fungal Diversity

An asexual genus with the type species Humicolafuscoatra Traaen grouping in the Chaetomiaceae, howeverthe genus has been shown to be polyphyletic with sexualmorphs also in Pseudolignincola (Halosphaeriaceae) (Joneset al. 2006), and Eurotiales (Index Fungorum 2015). TheHumicola species referred to the Halosphaeriaceae was notfu l ly iden t i f i ed and was the asexua l morph ofPseudolignincola (Jones et al. 2006).
Hyalorostratum Raja & Shearer, Mycosphere 1: 4 (2010)This genus is described from Alaska and New Hampshire
from freshwater habitats (Raja et al. 2010). The type species,H. brunneisporum Raja & Shearer, is placed in theDiaporthales based on combined SSU and LSU sequence data.
Hyalotiopsis Punith., Mycol. Pap. 119: 12 (1970) [1969]Ellurema was introduced with the type species Ellurema
indica (= Massarina indica Punith. by Nag Raj and Kendrick(1985). Hyalotiopsis subramanianii (Agnihothr. & Luke)Punith. was identified as the asexual morph of Ellurema indicain cultural studies (Nag Raj and Kendrick 1985). Ellurema isplaced in Amphisphaeriaceae in phylogenetic analysis of DNAsequences of the 5.8 S rRNA gene (Kang et al. 1999a).Hyalotiopsis, the older asexual typified genus, has a single spe-cies epithet (Index Fungorum 2015) and Ellurema the youngersexually typified genus has only one species epithet. We pro-pose to adopt the older asexual typified name (i.e.Hyalotiopsis)over sexual typified name (i.e. Ellurema).
HydeaK.L. Pang & E.B.G. Jones, Mycol. Prog. 9: 549 (2010)Monotypic genus of Lulworthiaceae that was introduced
by Abdel-Wahab et al. (2010) to accommodate Cirrenaliapygmea Kohlm. Molecular analysis of SSU and LSU rDNAsequence data placed H. pygmea (Kohlm.) K.L. Pang &E.B.G. Jones in a highly supported basal clade to the generaCumulospora, Kohlmeyeriella, Lindra, Lulworthia andMatsusporium within Lulworthiaceae (Abdel-Wahab et al.2010). The sexual morph is not known (Abdel-Wahab et al.2010).
Hydromelitis A. Ferrer et al., Mycologia 104(4): 876 (2012)This monotypic genus was introduced for a species from
submerged woody debris in freshwater collected in Costa Rica(Ferrer et al. 2012). Molecular analysis placed the genus inSordariomycetidae incertae sedis.
Hypocreodendron Henn., Hedwigia 36: 223 (1897)Hypocreodendronwas introduced based onH. sanguineum
Henn. from Argentina (Hennings 1897). According to theprotologue the genus has robust, highly branched, coralloidstromata and rod-shaped conidia.Discoxylaria is a monotypicgenus introduced for insect-associate xylaroid species; thetype species is Discoxylaria myrmecophila J.C. Lindq. &
J.E. Wright (Lindqvist and J.E. Wright 1964). It was sug-gested that this perithecioid fungus is the sexual morph ofHypocreodendron. Poroniopsis is a possible synonym ofHypocreodendron. Rogers et al. (1995) obtained a collectionfrom Mexico, which bore both the conidial and matureperithecial morphs. They compared the collection with theholotype and also obtained the asexual morph in culture.Hypocreodendron is an earlier name and thereforeDiscoxylaria should be a synonym. According to Stadleret al. (2013) the priority can be given to Hypocreodendronsince it was the first to be described among the two morphs.
Hypomyces (Fr.) Tul. & C. Tul., Annls Sci. Nat., Bot., sér. 413: 11 (1860)
Rossman et al. (2013) discussed the link betweenHypomyces and Cladobotryum and suggested that the use ofHypomyces should be retained even though Cladobotryumhas priority as an older name.We follow this here. The asexualmorph genera Mycogone, Sepedonium and Stephanoma maybe more distantly related to the type species of Hypomycesthan most members of Cladobotryum, and thus may not becongeneric (Rossman et al. 2013), thus we presently retainthese as distinct genera in Nectriaceae.
Hypophloeda K.D. Hyde & E.B.G. Jones, Trans. Mycol. Soc.Japan 30(1): 61 (1989)
Monotypic genus of the Melanconidaceae that was intro-duced by Hyde and Jones (1989) to accommodate the marinespecies H. rhizospora K.D. Hyde & E.B.G. Jones; no asexualmorph was recorded. The species is poorly known and re-quires further collection, isolation and sequencing to deter-mine its taxonomic position.
Hypoxylon Bull., Hist. Champ. Fr. (Paris) 1: 168 (1791)Nodulisporium is typified by N. ochraceum Preuss. Ju and
Rogers (1996) have described the asexual morph of severalxylariaceous taxa as Bnodulisporium-like^ and many authorsavoided giving separate names to the asexual morphs encoun-tered in nature or culture, especially in Hypoxylon. Accordingto Stadler et al. (2013) the possibility of assigningN. ochraceum as the asexual morph of Hypoxylon howeanumwhich tends to be isolated frequently in Germany can be con-sidered in forming the link between the asexual and sexualmorphs. The same study suggested placing Nodulisporium asa synonym of Hypoxylon. Triplicaria with its typeT. hypoxyloides P. Karst. is considered as one of the asexualgenera assigned to the genus Hypoxylon by Stadler et al.(2013). Therefore we consider both asexual genera,Nodulisporium and Triplicaria, are synonyms of Hypoxylon.
Idriella P.E. Nelson & S. Wilh., Mycologia 48(4): 550 (1956)Idriella was described to accommodate an important plant
pathogen, I. lunata P.E. Nelson & S. Wilh. which causes
Fungal Diversity

extensive root rot in strawberry fields in California (Nelsonand Wilhelm 1956). The taxon causes black sunken lesionsand produce dark chlamydospores both in culture and in dis-eased roots. The cultures obtained from chlamydospores pro-duced the asexual conidial morph (Nelson and Wilhelm1956). Wijayawardene et al. (2012) listed Idriella inXylariales, genera incertae sedis as the possible asexualmorph of Phomatospora. However, molecular data shows itto belong in Heliotales.
Ilyonectria P. Chaverri & C. Salgado, in Chaverri et al., Stud.Mycol. 68(1): 69 (2011)
This genus was introduced with I. radicicola(Gerlach & L. Nilsson) P. Chaverri & C. Salgado asthe type species and comprises ten species (Chaverriet al. 2011; Lombard et al. 2013). The genus wasplaced in Nectriaceae in Chaverri et al. (2011), whichwe follow here. Some species have been linked toCylindrocarpon sensu lato asexual morphs.
Immersidiscosia Kaz. Tanaka et al., in Tanaka et al.,Persoonia, Mol. Phyl. Evol. Fungi 26: 94 (2011)
Tanaka et al. (2011) showed Discosia separating in twosegregates in molecular analysis. The second segregate wastherefore renamed as the monotypic genus Immersidiscosiawith the type species I. eucalypti (Pat.) Kaz. Tanaka et al.and placed in Amphisphaeriaceae.
Immersiporthe S.F. Chen et al., in Chen et al., Pl. Path. (2012)This genus was int roduced and typi f ied by
Immersiporthe knoxdaviesiana S.F. Chen et al. (Chenet al. 2012), the causal agent of stem cankers onRapanea melanophloeos. Multi-gene phylogenetic anal-yses showed that the fungus represents a new genus inthe family Cryphonectriaceae. Typical fruiting bodiescharacteristic of Cryphonectriaceae were observed onthe surfaces of cankers (Chen et al. 2013b).
Infundibulomyces Plaingam et al., in Plaingam et al., Can. J.Bot. 81(7): 732 (2003)
The genus Infundibulomyces was introduced by Plaingamet al. (2003) to accommodate a single species, I. cupulataPlaingam et al., which was collected on leaves ofLagerstroemia species. Somrithipol et al. (2008) also intro-duced I. oblongisporus Somrithipol et al. and placement inthe family Chaetosphaeriaceae was confirmed based onanalysis of SSU and LSU gene data.
Iwilsoniella E.B.G. Jones, Syst. Ascomycetum 10: 8 (1991)Monotypic marine genus in the Halosphaeriaceae, grow-
ing on submerged wood (Jones et al. 2009a); no asexualmorph is known. A molecular study is required to confirmits current placement.
Jattaea Berl., Icon. fung. (Abellini) 3(1–2): 6 (1900)Jattaea andWegelina were published by Berlese (1900) as
morphologically similar genera. Réblová (2011b) examinedthe lectotypes of Jattaea algeriensis Berl. and Wegelinadiscreta Berl. and correlated her studies with in vitro studies.Phylogenetic studies show that both species are congeneric.Twelve species are accepted in Jattaea with phialophora-likeasexual morphs produced in culture. In the phylogenies,Jattaea leucospermi Marinc. et al. represents the phenotypecharacters, with septate, allantoid to suballantoid ascospores.This was formerly considered a diagnostic feature of themonotypic genus Phragmocalosphaeria (Petrak 1923).Molecular data confirm that ascospore septation is not a sig-nificant character suitable to distinguish genera in theCalosphaeriales and Phragmocalosphaeria is redued to asynonym of Jattaea.
Juncigena Kohlm. et al., Bot. Mar. 40(4): 291 (1997)Juncigena was introduced to accommodate an ascomycete
growing on the marsh plant Juncus roemerianus withJ. adarca Kohlm. et al. as the type species. DNA sequencesof two ribosomal nuclear loci confirm its position in theJuncigenaceae, Hypocreomycetidae family incertae sedis(Jones et al. 2014). Juncigena forms a well-supported cladewith the marine genera Fulvocentrum, Marinokulati, andMoheitospora.
Juncigenaceae E.B.G. Jones et al. Cryptogamie Mycologie35: 133 (2014)
This family was introduced by Jones et al. (2014) to ac-commodate the genera Fulvocentrum , Juncigena ,Marinokulati and Moheitospora. It is currently placed inHypocreomycetidae family incertae sedis. The asexual morphhas helicoid conidia when present.Kallichroma Kohlm. & Volkm.-Kohlm., Mycol. Res. 97: 759(1993)
Marine genus placed in Hypocreales based on moleculardata (Rossman et al. 2001; Schroers 2001; Jones et al. 2009b;Summerbell et al. 2011). Initially described as Hydronectria,but the type species of Hydronectria is a lichen with the algaphycobiont Trentepohlia. Subsequently, the taxon was trans-ferred to Kallichroma by Kohlmeyer and Volkmann-Kohlmeyer (1993). No asexual morph is known.
Kitesporella J.S. Jheng & K.L. Pang, Bot. Mar. 55(5): 462(2012)
This genus in the familyHalosphaeriaceaewas introducedfor Kitesporella keelungensis Jheng & K.L. Pang collected ondriftwood in Taiwan (Pang and Jheng 2012a). It resemblesAnisostagma, Iwilsoniella and Thalassogena (all membersof the Halosphaeriaceae). Sequence data are required to con-firm the placement of this species in the Halosphaeriaceae(Pang and Jheng 2012a).
Fungal Diversity

Kochiella Sakayaroj et al., Fungal Divers. 46: 96 (2011)A phylogenetic evaluation of Remispora, based on three
loci (nuclear small and large (LSU, SSU), the second largestRNA polymerase II subunit (RPB2)), demonstrated that thegenus is polyphyletic. Remispora maritima Linder (type spe-cies), R. pilleata Kohlm., R. quadri-remis (Höhnk) Kohlm.,R. spitsbergenensis K.L. Pang & Vrijmoed and R. stellataKohlm., form a monophyletic group (Remispora sensustricto) with Sablicola chinensis E.B.G. Jones et al. as as i s te r taxon wi th good suppor t , in the fami lyHalosphaeriaceae (Sakayaroj et al. 2011a). Remisporacrispa Kohlm. was distantly placed from Remispora in aclade with Ocostaspora apilongissima E.B.G. Jones et al.,but they are not considered conspecific. The genusKochiella was therefore introduced to include this speciesas Kochiella crispa (Kohlm.) Sakayaroj et al., based onmorphological and molecular data (Sakayaroj et al. 2011a).
Kohlmeyeriella E.B.G. Jones et al., Bot. Linn. Soc. 87: 210(1983)
Genus of Lulworthiaceae that was introduced by Joneset al. (1983) to accommodate a Corollospora species,C. tubulata Kohlm., based on a TEM study. A phylogeneticstudy based on SSU and LSU rDNA placed K. tubulata(Kohlm.) E.B.G. Jones et al. in Lulworthiaceae, along withLulworthia crassa Nakagiri that has thus been transferred tothis genus (Campbell et al. 2005). No asexual morph has beenrecorded for Kohlmeyeriella species.
Kretzschmaria Fr., Summa veg. Scand., Section Post.(Stockholm): 409 (1849)
Holttumia is typified by H. congregata Lloyd in Lloyd(1924b). This genus is now considered as a synonym ofKretzschmaria (Rogers and Ju 1998). Rogers and Ju (1998)observed the holotype material of Holttumia congregata (lo-cated at BPI) and noted the close morphological characterswith Kretzschmaria. Holttumia is considered as a synonymof Kretzschmaria, while H. congregata is a synonym ofK. macrosperma (Mont.) J.D. Rogers & Y.M. Ju (Rogersand Ju 1998).
Lanceispora Nakagiri et al., Mycoscience 38(2): 208 (1997)Accepted name for an aquatic genus referred to the
Xylariales (Jones et al. 2009b) with recollection, isolationand sequencing required to resolve its taxonomic position.
Lanspora K.D. Hyde & E.B.G. Jones, Can J. Bot. 64: 1581(1986)
Accepted monotypic mar ine genus, placed inOphiostomatales, based onmorphology and preliminary molec-ular data (Schoch pers. comm.), which grows on wood. Initiallyreferred to the Halosphaeriaceae, because of similar morpho-logical features to that family. No asexual morph is known.
Lasiadelphia Réblová & W. Gams, in Réblová et al., FungalDiversity 46(‘1’): 82 (2011)
This monotypic genus was introduced by Réblová et al.(2011b) and is the asexual morph of Lasiosphaeris.
Lasiosphaeriaceae Nannf., Nova Acta Regiae SocietatisScientiarum Upsaliensis 8 (2): 50 (1932)
A large family in the Sordariales with 71 generalisted in MycoBank (October 2014) and we list 35 gen-era. Molecular data indicates that the family isparaphyletic, with many genera polyphyletic e.g.Arnium, and the family requires wider sampling withan increased number of loci sequenced (Kruys et al.2015). A phylogenetic study based on LSU nrDNAand β-tubulin sequences highlights four lineages ofLasiosphaeriaceae (Kruys et al. 2015).
Lasiosphaeriella Sivan., Trans. Br. Mycol. Soc. 64(3): 443(1975)
Lasiosphaeriellawas introduced by Sivanesan (1975) withLasiosphaeriella dennisii Sivan. as the type species. The ge-nus presently includes five species (Index Fungorum 2015).Sequence data is available for three species which show thegenus to cluster as a group in Chaetosphaeriales (Huhndorfand Miller 2011).
Lasmenia Speg., Anal. Soc. cient. argent. 22(4): 199 (1886)Lasmenia causes rot and lesions on leaves and inflo-
rescences of tropical fruits, especially rambutan (Serrato-Diaz et al. 2011). Analysis of combined ITS and LSUsequence data indicates a closer affinity of Lasmenia tothe family Cryphonectriaceae (Serrato-Diaz et al. 2011).
Latruncellus M. Verm. et al., in Vermeulen et al., Mycologia103(3): 562 (2011)
This genus, from Swaziland, on bark of Galpiniatransvaalica, was introduced and typified by Latruncellusaurorae M. Verm. et al. (Vermeulen et al. 2011). The taxonis similar to members of Cryphonectriaceae that have uni-formly orange stromatic tissues for both sexual and asexualmorphs (Gryzenhout et al. 2009). This taxon has many similarcharacters with Aurifilum marmelostoma Begoude et al. andphylogenetically they are also closely related. However,conidiomata of Latruncellus aurorae have uniformly orange,constricted, fattened necks and Aurifilum marmelostoma doesnot form necks and has blackened ostiolar openings. Analysisof LSU and BT exon gene sequences also showedLatruncellus aurorae to be a separate genus in the familyCryphonectriaceae, and closely related to Aurifilummarmelostoma (Vermeulen et al. 2011).
Lautisporiopsis E.B.G. Jones et al., Can. J. Bot. 72l: 1558(1994)
Fungal Diversity

Monotypic marine ascomycete genus inHalosphaeriaceaegrowing on submerged wood (Jones et al. 2009b), variouslyreferred to Halosphaeria (Kohlmeyer 1960) andCeriosporopsis (Jones et al. 1995), but separated from thesegenera by the ultrastructure of its appendaged ascospores(Yusoff et al. 1994a, b). No asexual morph is known, but thisspecies frequently produces chlamydospores.
LecanicilliumW.Gams& Zare, NovaHedwigia 72(3–4): 332(2001)
Zare and Gams (2001b) transferred 12 species formerlyunder Verticillium to the new genus Lecanicillium, and alsointroduced three new species, based on morphology and phy-logenetic analysis. The genus is placed in Cordycipitaceae(Index Fungorum 2015).
Lectera P.F. Cannon, in Cannon et al., MycoKeys 3: 28 (2012)Cannon et al. (2012), based on rDNA ITS and GAPDH
sequences, showed that Lectera colletotrichoides (J.E.Chilton) P.F. Cannon belongs in Plectosphaerellaceae ratherthan Hypocreales, to which other species are assigned.Currently two species are referred to this genus. Lectera ischaracterised by brightly coloured sporodochia surroundedby brown setae, and may be a sister group to Verticillium,but bootstrap support is weak.
Leiosphaerella Höhn., Sber. Akad. Wiss. Wien, Math.-naturw. Kl., Abt. 1 128: 579 (1919)
Jaklitsch and Voglmayr (2012) recollected the typespecies of L. praeclara (Rehm) Höhn. and their se-quence data placed Leiosphaerella in Xylariales.Leiosphaerella sensu stricto and Pseudomassaria sensustricto were shown to be closely related.
Leptosporella Penz. & Sacc., Malpighia 11(9–10): 406 (1897)Leptosporella gregaria Penz. & Sacc., the generic type,
clusters in a well-supported group in Chaetosphaeriales(Huhndorf and Miller 2011).
LignincolaHöhnk, Veröff. Inst. Meeresforch. Bremerhaven 3:216 (1955)
Genus of Halosphaeriaceae with freshwater and marinespecies, growing on submerged wood or palm fronds (Joneset al. 2009a). No sexual morph is known. Phylogenetic studiesplace this genus in Halosphaeriaceae, but further sequencedata is required for some of the species before they can bereferred to the genus with confidence.
Limacospora Jørg. Koch & E.B.G. Jones, Can. J. Bot. 73(7):1011 (1995)
Monotypic marine ascomycete genus growing on sub-merged wood, and referred to the Halosphaeriaceae. Furthercollections, are required for isolation and sequencing to
determine its phylogenetic relationship with the generaCeriosporopsis and Marinsopora.
Lindra I.M. Wilson, Trans. Br. Mycol. Soc. 39: 411 (1956)Genus of Lulworthiaceae introduced by Wilson (1956) to
accommodate L. inflata I.M. Wilson. The genus currently con-tains five marine species, of which L. obtusaNakagiri & Tubakihas Anguillospora marina Nakagiri & Tubaki as its asexualmorph. Preliminary phylogenetic analyses of the SSU andLSU rDNA showed that Lindra species are polyphyletic(Campbell et al. 2005; Jones et al. 2008; Abdel-Wahab et al.2010).
Longicollum Zelski et al., in Zelski et al., Mycosphere 2(5):540 (2011)
Longicollum is a monotypic genus described fromsubmerged woody debris from a river in Peru (Zelskiet al. 2011b). Morphologically, Longicollum exhibits allof the characters of Annulatascaceae and in addition hasascospores with verruculose wall ornamentation.Longicollum biappendiculatum Zelski et al. shares manycharacteristics with Submersisphaeria aquatica K.D.Hyde (Hyde 1996) and shows some similarities to manyof the currently accepted genera in the familyAnnulatascaceae sensu lato (Zelski et al. 2011b). Nomolecular analysis was carried out to support the place-ment of this genus in Annulatascaceae.
Lopadostoma (Nitschke) Traverso, Fl. ital. crypt., Pars 1:Fungi . Pyrenomycetae . Xylariaceae , Valsaceae ,Ceratostomataceae 1(2): 169 (1906)
Genus of Xylariaceae that was first introduced byNitschke (1867) as Anthostoma subg. LopadostomaNitschke and later elevated to generic rank by Traverso(1906). Twelve species were recognized during a revisionof this genus (Jaklitsch et al. 2014). Combined molecularphylogenetic analysis based on ITS, LSU, RPB2 revealed10 distinct taxa within the genus which were also recognizedat the species level. A phylogenetic analysis based on nuLSUrDNA confirmed that the genus appeared as a monophyleticclade within the family Xylariaceae, with moderate bootstrapsupport (Jaklitsch et al. 2014).
Lulwoana Kohlm. et al., Mycol. Res. 109: 62 (2005)Monotypic genus of Lulworthiaceae introduced by
Campbell et al. (2005) to accommodate L. uniseptata(Nakagiri) Kohlm. et al. The asexual morph is Zalerionmaritima (Linder) Anastasiou.
Lulwoidea Kohlm. et al., Mycol. Res. 109: 164 (2005)Monotypic genus of Lulworthiaceae introduced by
Campbell et al. (2005) to accommodate L. lignoarenaria(Jørg. Koch & E.B.G. Jones) Kohlm. et al. Lulwoidea
Fungal Diversity

lignoarenaria along with Orbimyces spectabilis Linderformed a highly supported basal clade in Lulworthiaceae(Campbell et al. 2005). The genus forms black ascomataon sand grains. No asexual morph has been recorded forL. lignoarenaria (Koch and Jones 1984).
Lulworthia G.K. Sutherl., Trans. Br. Mycol. Soc. 5: 259(1916)
Genus of Lulworthiaceae that was introduced bySutherland (1915) to accommodate the type speciesLulworthia fucicola G.K. Sutherl. Several new Lulworthiaspecies were described from marine habitats and the genusbecame one of the largest genera in marine habitats. Theplacement of the genus in Lulworthiaceae, Lulworthialeswas confirmed at the morphological and molecular level(Kohlmeyer et al. 2000). Campbell et al. (2005) re-evaluatedthe taxonomy of the genus at the molecular level and intro-duced two new genera Lulwoana Kohlm. et al. (based onLulworthia uniseptata Nakagiri) and Lulwoidea Kohlm.et al. (based on Lulworthia lignoarenaria Jørg. Koch &E.B.G. Jones). No type material was available for the typespecies L. fucicola Sutherl., therefore, Campbell et al. (2005)neotypified a collection of L. fucicola from Chile growing onwood. No asexual morph has been reported for Lulworthiasensu stricto.
LunatiannulusDaranagama et al., Fungal Diversity (in press)(2015)
Lunatiannulus was introduced by Daranagama et al.(2015) with L. irregularis Daranagama et al. as the typespecies. This genus is morphologically related to Diatrype-like genera in having a libertella-like asexual morph andallantoid ascospores. However, L. irregularis differs in hav-ing 4 or 8-spored asci, whereas most of the diatrypaceousasci are either polysporous or multiples of 8, and ascosporesare falcate and hyaline (Daranagama et al. 2015; Liu et al.2015). Multi-gene analysis shows this new genus has closeaffinities with Eupixylon and Nemania in Xylariaceae(Daranagama et al. 2015).
Luteocirrhus C.F. Crane et al., IMA Fungus 4(1): 115 (2013)Luteocirrhus was isolated from canker lesions in sev-
eral Banksia species plus Lambertia echinata subsp.citrina and was introduced as a monotypic genus withLuteocirrhus shearii C.F. Crane et al. as its type. It isplaced within Cryphonectriaceae (Diaporthales) basedon morphology and molecular data (Crane and Burgess2013). Morphologically it is characterized by pulvinateto globose, black, semi-immersed conidiomata, with pa-raphyses. Its sexual morph is unknown. This pathogenof native Proteaceae causes canker lesions and is con-sidered as an emerging threat towards Banksia speciesin South West Australia.
Luttrellia Shearer, Mycologia 70: 692 (1978)Genus of Halosphaeriaceae (Jones et al. 2009a) that was
established by Shearer (1978) to accommodate the type spe-cies, L. estuarina Shearer from decayed balsa wood sub-merged in Patuxent River, USA. Another three species weredescribed from freshwater habitats (Ferrer and Shearer 2007).No molecular study has been carried out to establish the phy-logenetic position of the genus. Luttrellia is distinguishedfrom other genera in the family by hyaline, phragmoseptateand thick-walled ascospores, with or without a gelatinoussheath. No asexual morph is known.
Macgarvieomyces Klaubauf et al., Stud. Mycol. 79: 107(2014)
MacGarvie introduced the genus Diplorhinotrichum basedon two species occurring on Juncus spp (Klaubauf et al.2014). de Hoog and Van Oorschot (1985) treatedDiplorhinotrichum as a synonym of Dactylaria, and retainedthe species in Pyricularia. However, based on a LSU, ITS,RPB1, actin and calmodulin dataset, Klaubauf et al. (2014)showed the above species do not belong in Pyricularia and anew genus, Macgarvieomyces, was introduced in the familyPyriculariaceae to accommodate them.
Macroconia (Wollenw.) Gräfenhan et al., in Gräfenhan et al.,Stud. Mycol. 68(1): 101 (2011)
A section of Nectria, Macroconia, was raised to genericrank for five species all producing sexual morphs andmacroconidial asexual morphs. The type species isM. leptosphaeriae (Niessl) Gräfenhan & Schroers, which isbased on Nectria leptosphaeriae Niessl. Members of this ge-nus mostly grow on the stromata of other ascomycetes onherbaceous plants or deciduous trees. In the phylogeneticanalyses, Macroconia species are placed in Nectriaceae(Gräfenhan et al. 2011).
Magnaporthiopsis J. Luo & N. Zhang Mycologia 105(4):1024 (2013)
A study on the family Magnaporthaceae by Luo andZhang (2013) based on multiple genes including SSU, ITS,LSU, MCM7, RPB1 and TEF data showed thatMagnaportheand Gaeumannomyces are polyphyletic. Furthermore theirmembers divided into four major groups and consideringmor-phological, biological and molecular data, a new genus,Magnaporthiopsis was introduced. Species in this genus arenecrotrophic parasites infecting roots of grasses (Luo andZhang 2013). The genus is typified with Magnaporthiopsispoae J. Luo &N. Zhang and presently there are four species inMagnaporthiopsis.
Magnisphaera J. Campb. et al., Mycologia 95: 546 (2003)Genus of Halosphaeriaceae introduced by Campbell et al.
(2003) to accommodate the type species,M. spartinae (E.B.G.
Fungal Diversity

Jones) J. Campb. et al.Halosarpheia spartinae (E.B.G. Jones)Shearer & J.L. Crane was transferred toMagnisphaera basedon phylogenetic analyses of SSU and LSU rDNA sequencedata (Campbell et al. 2003). Another new species,M. stevemossago J. Campb. et al. was described from decayedwood in freshwater habitats in the same study. No asexualmorph is known (Campbell et al. 2003). Matsusphaeria(Pang et al. 2004) is a synonym of Magnisphaera J. Campb.et al., which is an earlier name.
Marinokulati E.B.G. Jones & K.L. Pang, CryptogamieMycologie 35: 132 (2014)
A genus introduced to accommodate the marine ascomy-cete Chaetosphaeria chaetosa Kohlm. as Jones et al. (2014)showed thatChaetosphaeriawas polyphyletic, with most spe-cies grouping in the order Chaetosphaeriales. Two sequencesofCh. chaetosa formed a monophyletic group with Juncigenaadarca Kohlm. et al., Moheitospora fruticosa Abdel-Wahabet al. and two Fulvocentrum species, with high bootstrap sup-port in the family Juncigenaceae, Hypocreomycetidae orderincertae sedis (Jones et al. 2014).
Marinosphaera K.D. Hyde, Can J. Bot. 67: 3080 (1989)Monotypic marine genus, whose taxonomic position is not
resolved at the molecular level (Jones et al. 2009b) and furthersampling, is required. Initially referred to the Phyllachoraceae(Hyde 1989), while Jones et al. (2009b) showed it grouping asa separate clade to theMicroascaceae and Halosphaeriaceae,but it shows little affinities with either of these families.However, depending on the molecular analysis of SSUand LSU rRNA gene sequence data the genusMarinosphaera is currently palced in Phyllachorales genusincertae sedis (Jones et al. 2009b). No asexual morph isknown.
Marinospora A.R. Caval., Nova Hedwigia 11: 548 (1966)Genus introduced to accommodate two lignicolous marine
ascomycetes grouping in the Halosphaeriaceae(Microascales) with high support and forming a sister groupto the generaOndiniella and Toriella (Sakayaroj et al. 2011a).No asexual morph is known.
Matsusporium E.B.G. Jones & K.L. Pang, in Abdel-Wahabet al., Mycol. Progr. 9(4): 550 (2010)
Monotypic marine asexual genus in the Lulworthiales(Abdel-Wahab et al. 2010), and forming a sister clade toLulworthia grandispora Meyers (Lulworthia sensu lato).The type species is M. tropicale (Kohlm.) E.B.G. Jones &K.L. Pang, initially described as Cirrenalia tropicalisKohlm., but in a molecular study was found to be distantlyplaced from the type species of Cirrenalia (C. macrocephala(Kohlm.) Meyers & R.T. Moore), Halosphaeriaceae (Abdel-Wahab et al. 2010). No sexual morph is known.
Mattirolia Berl. & Bres., Microm. Trid.: 55 (1889)After examining the type species of Balzania, Mattirolia,
Thyridium, and Thyronectroidea, onlyMattirolia (with 5 spe-cies) and Thyridium (with 4 species) are accepted within theclass Thyridiaceae. Balzania and Thyronectroidea are consid-ered as synonyms of Mattirolia (Checa et al. 2013).
Melanconiella Sacc., Syll. fung. (Abellini) 1: 740 (1882)Voglmayr et al. (2012) confirmed that Melanconiella
is monophyletic and distinct from Melanconis based onLSU sequence data. The type species for Melanconiellais M. spodiaea (Tul. & C. Tul.) Sacc. Melanconiellawas shown to comprise 13 species based on combinedphylogenetic analyses of SSU, ITS, LSU, TEF andRPB2 sequence data. The genus is confined to hostfamily Betulaceae. Melanconiella has previously beenplaced in Melanconidaceae as it was considered as asynonym of Melanconis. A characteristic melanconium-or discosporina-like asexual morph was produced byeach species of Melanconiella.
Melanopsamma Niessl, Verh. nat. Ver. Brünn 14: 200 (1876)Three bitunicate species are exclude from the genus by
Wang (2011).
Melogrammataceae G. Winter [as ‘Melogrameae’], Rabenh.Krypt.-Fl., Edn 2 (Leipzig) 1.2: 797 (1886)
Jaklitsch and Voglmayr (2012) recollected the type species,Melogramma campylosporum Fr., which clustered inXylariales in a monophyletic lineage. The family name wastherefore retrieved to accommodate this sole taxon.
Menisporopsis S. Hughes, Mycol. Pap. 48: 59 (1952)Menisporopsis is typified byMenisporopsis theobromae S.
Hughes and a sequence of this putative name is listed underAscomycota genera incertae sedis in GenBank andChaetothyriaceae in Index Fungorum (2014). Menisporopsiskobensis Matsush. was linked to Menisporopascus kobensisMatsush. (Index Fungorum 2015; Matsushima 2001), howev-er no sequence data is available for these taxa. We treatMenisporopsis in Ascomycota genera incertae sedis pendinga molecular study.
Metapochonia Kepler et al., in Kepler et al., Mycologia106(4): 820 (2014)
This genus was proposed based on molecular analyses ofTUB, RPB1, RPB2 and TEF gene sequences (Kepler et al.2014). No sexual morph is know for this genus.
Mirannulata Huhndorf et al., Sydowia 55(2): 173 (2003)This genus was described by Huhndorf et al. (2003), they
placed this genus in Sordariomycetes genera incertae sedisbased on morphology. Boonyuen et al. (2012) in their
Fungal Diversity

phylogenetic analyses using a LSU and SSU gene datasetplaced it in Sordariomycetidae genera incertae sedis.
Moana Kohlm. & Volkm.-Kohlm., Mycol. Res. 92: 418(1989)
Monotypic genus of Halosphaeriaceae that wasestablished by Kohlmeyer and Volkmann-Kohlmeyer (1989)to accommodate the marine fungus,M. turbinulata Kohlm. &Volkm.-Kohlm. No molecular study has been carried out onthe genus and further studies are required to confirm its as-signment to the Halosphaeriaceae. No asexual morph isknown (Kohlmeyer and Volkmann-Kohlmeyer 1989).
Moheitospora Abdel-Wahab et al., in Abdel-Wahab et al.,Mycol. Progr. 9(4): 551 (2010)
The asexual genus that groups in Juncigenaceae with themarine genera Juncigena, Fulvocentrum, and Marinokulati(Jones et al. 2014). It is distinct from, but forms a sister groupto Juncigena with high support and we maintain these as dis-tinct genera. The fungus has coiled conidia with small cellsthat distinguish it fromCirrenalia species (Abdel-Wahab et al.2010). No sexual morph has been observed.
Moleospora Abdel-Wahab et al., in Abdel-Wahab et al.,Mycol. Progr. 9(4): 547 (2010)
A monotypic marine asexual genus in the Lulworthiales(Abdel-Wahab et al. 2010) isolated from Phragmites australiscollected in Egypt. Morphologically it is similar toHalenospora varia (Anastasiou) E.B.G. Jones andCumulospora marina I. Schmidt, but differs in conidial dimen-sions (Abdel-Wahab et al. 2010). No sexual morph is known.
Monilochaetes Halst. ex Harter, J. Agric. Res., Washington 5:791 (1916)
The type species of Dischloridium, D. laeënse (Matsush.)B. Sutton was synonymised underMonilochaetes, but molec-ular data shows it forms a sister clade to the generic type ofMonilochaetes, M. infuscans Harter in Australiasca (Réblováet al. 2011a).
Monochaetia (Sacc.) Allesch., Rabenh. Krypt.-Fl., Edn 2(Leipzig) 1(7): 665 (1902) [1903]
Steyaert (1949) treated Monochaetia (Sacc.) Allesch. andplaced species with single setula in section Monosetulatae ofPestalotiopsis and Truncatella. However, Monochaetia wasretained as a distinct genus by Guba (1961) based on its singleapical appendage. Monochaetia is not linked to a sexualmorph and so far more than 120 taxa have been described inMonochaetia, and these mostly comprise 4 septate and 5sep t a t e spec i e s . Based on LSU sequence da t aMaharachchikumbura et al. (2014) show that Monochaetiarepresents a genus that is distinct from Pestalotiopsis,Seiridium and Truncatella. However, it is essential to
incorporate molecular data and more taxon sampling in futureanalyses asMonochaetia includes 3-, 4-, and 6-celled conidialforms (Maharachchikumbura et al. 2014).
Monochaetinula Muthumary et al., Trans. Br. mycol. Soc.87(1): 104 (1986)
The genus is typified by M. terminalae (Bat. & Bezerra)Muthumary et al., an asexual coelomycete genus in theAmphisphaeriaceae, which includes M. ampelophila (Speg.)Nag Raj, M. caffra Matsush, M. ceratoniae (Sousa daCâmara) Nag Raj, M. geoffroeana Bianchin. andM. sterculiaeNag Raj. A molecular study is required to ascer-tain its taxonomic position in the Ascomycota.
Morakotiella Sakay., Mycologia 97(4): 806 (2005)Monotypic genus ofHalosphaeriaceae that was introduced
by Sakayaroj (2005) to accommodate Haligena salinaC.A. Farrant & E.B.G. Jones. Phylogenetic analyses ofthe LSU rDNA of the two Haligena species showedthat i t was polyphyletic with the type species(H. elaterophora) forming a basal clade to the family.Subsequently, Sakayaroj et al. (2011a) showed thatMorakotiella salina (C.A. Farrant & E.B.G. Jones)Sakay. grouped as a sister taxon to the generaKochiella and Ocostospora with high statistical support.No asexual morph is known.
Morinia Berl. & Bres., Annuario Soc. Alpinisti Trident.,1887–88: 82 (1889) [1887–88]
The genus Morinia is based onM. pestalozzioides Berl. &Bres. and comprises an asexual fungus forming appendageand muriform conidia in acervular conidiomata (Colladoet al. 2006). ITS sequence data indicate that Morinia belongsin Amphisphaeriaceae with the highest similarity toBartalinia and Truncatella (Collado et al. 2006).
Moromyces Abdel-Wahab et al., in Abdel-Wahab et al.,Mycol. Progr. 9(4): 555 (2010)
Marine monotypic asexual genus in the orderLulworthiales (Abdel-Wahab et al. 2010) forming a sis-ter clade to the monophyletic genus Lulwoana. The ge-nus was introduced for the species Cumulospora variaChatmala & Somrith., as it was distantly placed fromthe type species of Cumulospora (C. marina). No sex-ual morph is known.
Muscodor Worapong et al., Mycotaxon 79: 71 (2001)The genus Muscodor was introduced by Worapong et al.
(2001) for the placement of M. albus Worapong et al., anendophytic fungus that produces volatile antibiotics. It is anasexual genus that has tentatively been referred to theXylariaceae. However, the possibility of the inclusion of thisgenus in the family has been addressed by Stadler et al.
Fungal Diversity

(2013). The phylogenetic analysis by Worapong et al. (2001)was inadequate to support the family placement because theycompared only ribosomal RNA gene sequences of a limitednumber of taxa. Later on, with the availability of reliable se-quence data for important taxa, Stadler et al. (2013) carried outa molecular analysis using only ITS data andMuscodor nestedin the xylarioid Xylariaceae. Thus they represent a monophy-letic basal clade to other xylarioid Xylariaceae especially as anin-group of the heterogeneous group Xylaria.
Musicillium Zare & W. Gams, in Zare et al., Nova Hedwigia85(3–4): 482 (2007)
Verticillium theobromae Turconi is not congeneric withVerticillium sensu stricto, as shown in cladograms based onLSU and ITS sequences, and the genus Musicillium was in-troduced to accommodate it (Zare et al. 2007). The genus isaccommodated in Plectosphaerellaceae. Musicilliumtheobromae (Turconi) Zare & W. Gams is the causal agentof cigar-end rot of bananas.
MyceliophthoraCostantin, C. r. hebd. Séanc. Acad. Sci., Paris114: 849 (1892)
Myceliophthora is typified by Myceliophthora luteaCostantin and Corynascus is typified by Corynascussepedonium (C.W. Emmons) Arx and both genera are linkedbymolecular data (van den Brink et al. 2012).Myceliophthorais the earlier described genus and it is commonly used inpublications. Van den Brink et al. (2012) proposed to placeall Corynascus species under Myceliophthora.
Myrothecium Tode, Fung. mecklenb. sel. (Lüneburg) 1: 25(1790)
Based on DNA sequences analysis, Castlebury et al.(2004) showed that species of Stachybotrys, species ofMyrothecium and two hypocrealean species form a previous-ly unknown monophyletic lineage within the Hypocreales.In recent studies Crous et al. (2014) introduced the familyStachybotriaceae to accommodate this undescribed familyand the genus Myrothecium.
Nais Kohlm., Nova Hedwigia 4: 409 (1962)Genus of Halosphaeriaceae that was established by
Kohlmeyer (1962) to accommodate N. inornata Kohlm.Another species, N. aquatica K.D. Hyde was described froma freshwater habitat. Phylogenetic analyses of the LSU rDNAconfirmed its placement in the family and showed thatN. inornata, has a close relationship with Aniptodera(Sakayaroj 2005, Sakayaroj et al. 2011a). No asexual morphis known (Shearer and Crane 1978).
Nalanthamala Subram., J. Indian bot. Soc. 35: 478 (1956)The genus Nalanthamala was shown to be linked to
Rubrinectria by Schroers et al. (2005) in Nectriaceae. As
Rubrinectria has a single species, Rossman et al. (2013) pro-posed using the older name Nalanthamala with Rubrinectriaas a synonym. This is followed here.
Natantispora J. Campb. et al., Mycologia 95(3): 543 (2003)Genus with two aquatic species that was introduced by
Campbell et al. (2003) to accommodate two Halosarpheia spe-cies, H. lotica Shearer and H. retorquens Shearer & J.L. Craneand another marine species (N. unipolarae) described by K.L.Pang et al. in Liu et al. (2015). The genus formed a well-supported clade that is distant from the type species, H. fibrosaKohlm.&E.Kohlm. (Jones et al. 2009a), based on phylogeneticanalyses of SSU and LSU rDNA. No asexual morph is known.
Naufragella Kohlm. & Volkm.-Kohlm., Syst. Ascomycetum16: 19 (1998)
Abdel-Wahab (2011) concluded, based on LSU sequencedata, that the genera Naufragella and Nohea were congenericand Naufragella spinibarbata (Jørg. Koch) Kohlm. &Volkm.-Kohlm. was consequently transferred to Nohea.However, Chu et al. (2015) in a phylogenetic reappraisal ofNimbospora, showed thatN. spinibarbata formed a sister groupto two Nimbospora species with high statistical support. Theirdata places Nohea umiumiKohlm. & Volkm.-Kohlm. as a sisterclade to the Nimbospora/Naufragella clade with weak support.Unfortunately both studies used different genes and species sothat the placement of these genera needs re-evaluation.Therefore, the transfer of Naufragella spinibarbata to Nohearequires further sampling of related taxa and a wider range ofgenes before the relationships of these genera are resolved.
Nautosphaeria E.B.G. Jones, Trans. Br. Mycol. Soc. 47(1):97 (1964)
Monotypic marine ascomycete genus growing on sub-merged wood, with its placement in the Halosphaeriaceaeconfirmed by 28S rDNA sequences (Sakayaroj 2005,Sakayaroj et al. 2011a). No asexual morph is known.
Nectria (Fr.) Fr., Summa veg. Scand., Section Post.(Stockholm): 387 (1849)
The genus was revisited by Hirooka et al. (2012) withN. cinnabarina (Tode) Fr., the generic type and 29additional species. Tubercularia was considered to be theasexual morph and listed as a synonym. Nectria clusteredwith 16 other genera in Nectriaceae. Rossman et al. (2013)proposed that the generic name Nectria be protected againstTubercularia by suppression of the latter generic name andthis is followed here.
Nectriaceae Tul. & C. Tul. [as ‘Nectriei’], Select. fung. carpol.(Paris) 3: 3 (1865)
Chaverri et al. (2011) showed Nectriaceae to comprise sevengenera. The family was revisited by Hirooka et al. (2012) with a
Fungal Diversity

monograph of Allantonectria, Nectria and Pleonectria. Besidesthese genera, Calonectria, Cosmospora, Cyanonectria,Coral lomycete l la , Lanatonectr ia , Leuconectr ia ,Neocosmospora, Ophionectria, Pseudonectria, Rodentomyces,Rugonectria, Thelonectria and Viridispora clustered inNectriaceae.
NemaniaGray, Nat. Arr. Brit. Pl. (London) 1: 508, 516 (1821)Geniculisynnema termiticola Okane & Nakagiri, was intro-
duced to accommodate the asexual morph of Xylaria angulosaJ.D. Rogers et al. Geniculisynnema termiticola was obtainedfrom a piece of a termite nest incubated in a moist chamberand another fungus isolated on Oatmeal agar (OA) from thesame termite nest produced the sexual morph (Okane andNakagiri 2007). This upright, upper part branched stromatawere identical with X. angulosa found from soil in Indonesiaby Rogers et al. (1987). The sexual morph and the asexualmorph were different in their morphology and colony charac-teristics (Okane and Nakagiri 2007). Geniculisynnematermiticola has never formed its stromata on media.Nevertheless the phylogenetic analysis by Okane and Nakagiri(2007) showed that G. termiticola nested with Nemania, whichhas close affinities to theXylaria.Thus it can be assumed that itssexual morph should be linked to Nemania. As mentioned inStadler et al. (2013) Geniculisynnema can be considered as theyounger synonym of Nemania. The genus Geniculosporiumwas introduced by Chesters and Greenhalgh (1964) and typifiedbyG. serpens Chesters & Greenh., which is currently named asNemania serpens (Pers.) Gray, which is also the type species ofthe sexual genus Nemania S.F. Gray (Stadler et al. 2013).Kenerley and Rogers (1976) found that three Nemania speciesproduced Geniculosporium asexual morphs in culture (Petriniand Rogers 1986). Therefore, Geniculosporium can be sup-pressed in favour of Nemania (Stadler et al. 2013).
Neochaetosphaerella Lar.N. Vassiljeva et al., in Vasiljevaet al., Fungal Diversity 52(1): 192 (2012)
This monotypic genus was introduced by Vasilyeva et al.(2012) based on morphology. The type species isN. thaxteriospora Lar.N. Vassiljeva et al. This genus sharessimilarity to members of Nitschkiaceae sensu lato but no mo-lecular data is available.
NeolinocarponK.D. Hyde, Bot. J. Linn. Soc. 110: 104 (1992)(1992)
Based on morphology, the genus Neolinocarpon cannot beplaced in any family within Xylariales with certainty and thusis placed as Xylariales genera incertae sedis (Jones et al.2009a, b).
NeonectriaWollenw., Annls mycol. 15(1/2): 52 (1917)Chaverri et al. (2011) placed Neonectria sensu stricto in
Nectriaceae with Cylindrocarpon sensu stricto asexual
morphs.Rossman et al. (2013) recommended, given the broadclassical concept of Cylindrocarpon and that it is well-circumscribed and includes a number of plant pathogenic spe-cies, that the generic name Neonectria is protected againstCylindrocarpon. Cylindrodendrum was considered not to becongeneric with Neonectria, while the synonymy of Heliscuswas doubtful.
Neopestalotiopsis Maharachch. et al., Stud. Mycol. 79: 135(2014)
Maharachchikumbura et al. (2014) resolved genera in theAmphisphaeriaceae based on analysis of LSU sequence data.The phylogeny resolved Pestalotiopsis as a distinct clade inAmphisphaeriaceae, with three well-supported groups thatcorrelated with morphology; besides Pestalotiopsis, two newgenera, Neopestalotiopsis and Pseudopestalotiopsis were in-troduced. Neopestalotiopsis protearum (Crous & L. Swart)Maharachch. et al., which was isolated from living leaves ofLeucospermum cuneiforme in Zimbabwe, was assigned as thegeneric type. MorphologicallyNeopestalotiopsis can easily bedistinguished from Pseudopestalotiopsis and Pestalotiopsisby its versicolorous median cells. Furthermore, inNeopestalotiopsis, conidiophores are indistinct and often re-duced to conidiogenous cells (Maharachchikumbura et al.2014). In their phylogenetic analysis Maharachchikumburaet al. (2014) included 24 ex-type/ex-epitype strains for speciesof Neopestalotiopsis.
Neopyricularia Klaubauf et al., Stud. Mycol. 79: 109 (2014)Neopyricularia is a newly introduced genus in the family
Pyriculariaceae, with the type species Pyriculariacommelinicola Klaubauf et al., which was isolated fromCommelina communis in South Korea (Klaubauf et al.2014). Phylogenetically (combined LSU, ITS, RPB1, actinand calmodulin dataset), P. commelinicola does not clusterwithin clades corresponding to species of Pyricularia sensustricto and hence Klaubauf et al. (2014) introduced the genusNeopyricularia.
NeptunellaK.L. Pang & E.B.G. Jones, Mycol. Prog. 2 (1): 35(2003)
A taxon previously referred to Gnomonia (Cribb andCribb 1956) and Lignincola (Kohlmeyer 1984), but mor-phological and molecular data confirms its placement in theHalosphaeriaceae (Pang et al. 2003; Jones et al. 2009a, b).The genus is monotypic and resembles Lignincola. Noasexual morph is known.
Nereiospora E.B.G. Jones et al., Bot. J. Linn. Soc. 87 (2): 204(1983)
An ascomycete genus well placed at the morpholog-ical and molecular level in the Halosphaeriaceae, with amonodictys-like asexual morph (Mouzouras and Jones
Fungal Diversity

1985; Sakayaroj et al. 2011a). Nereiospora forms awel l -suppor ted clade in the Halosphaeriaceae(Sakayaroj et al. 2011a).
Neurospora Shear & B.O. Dodge, J. Agric. Res., Washington34: 1025 (1927)
The type species ofChrysonilia,C. sitophila (Mont.) Arx islinked to the type species ofNeurospora,N. sitophila Shear &B.O. Dodge (Francuz et al. 2010).Neurospora is the older andmore commonly used name and therefore Chrysonilia shouldbe a synonym.
NiessliaAuersw., in Gonnermann&Rabenhorst, Myc. Europ.Pyren. 5–6: 30 (1869)
Whitton et al. (2012) introduced four new species inNiesslia (N. cinctiostiolata Whitton et al., N. kapitiaeWhitton et al., N. pacifica Whitton et al., N. vaginataWhitton et al.) from Pandanaceae, while Etayo et al. (2013)introduced a lichenicolous species N. echinoides Etayo et al.,all based on morphology. There are 11 hits in GenBank for aputative strain of N. exilis (Alb. & Schwein.) G. Winter, butthe generic type species N. exosporioides (Desm.) G. Winter(= N. chaetomium (Corda) Auersw. has not been sequenced.
Nigrocornus Ryley & Langdon, in Ryley, Mycology Series(New York) 19: 266 (2003)
This genus Nigrocornus was introduced to accommo-date a species of Balansia which was significantly dif-ferent from that of the type and all other Balansia spe-cies (Ryley 2003). The genus belongs in the familyClavicipitaceae.
Nimbospora Jørg. Koch, Nordic J. Bot. 2 (2): 166 (1982)Genus in the Halosphaeriaceae based on morphological
and molecular evidence, with Chu et al. (2015) showing twoNimbospora species forming a sister clade to Naufragellaspinibarbata (Jørg. Koch) Kohlm. & Volkm.-Kohlm. withhigh statistical support. No asexual morph is known for spe-cies assigned to this genus.
NitschkiaceaeHöhn., Nova Acta R. Soc. Scient. upsal., Ser. 48(no. 2): 56 (1932)
Mugambi and Huhndorf (2010) revisited the orderCoronophorales with DNA sequence data and showedNitschkiaceae to be paraphyletic with Fracchiaea speciesnot consistently grouping together or with Nitschkia andAcanthonitschkea.
Nohea Kohlm. & Volkm.-Kohlm., Syst. Ascomycetum 19:121 (1991)
Monotypic genus in the Halosphaeriaceae supported bymorphological and molecular data (Sakayaroj et al. 2011a;Chu et al. 2015). The type species (N. umiumi Kohlm. &
Volkm.-Kohlm.) forms a sister clade to the Nimbospora/Naufragella clade with moderate support. A unique featureof Nohea umiumi is the ascospores with two types of append-ages (Kohlmeyer and Volkmann-Kohlmeyer 1991). No asex-ual morph is known.
Occultocarpon L.C. Mejía & ZhuL. Yang, in Mejía et al.,Fungal Diversity 52(1): 101 (2012)
This monotypic genus is placed in Gnomoniaceae,Diaporthales based on a dataset of LSU, RPB2 and TEFgene sequence data (Mejía et al. 2012). The type species isO. ailaoshanense L.C. Mejía & Zhu L. Yang and was iso-lated from the bark of Alnus nepalensis in Yunnan, China(Mejía et al. 2012).
Oceanitis Kohlm., Revue Mycol., Paris 41(2): 193 (1977)Ascosalsum was introduced by Campbell et al. (2003) to
accommodate two Halosarpheia and one Haligena species.Molecular study of SSU and LSU rDNA confirmed thatAscosalsum is congeneric with Oceanitis (Dupont et al.2009). Consequently all species assigned to Ascosalsum werereduced to synonymy in Oceanitis. These species formed ahighly supported clade in the Halosphaeriaceae withOphiodeira monosemeia Kohlm. & Volkm.-Kohlm.
Ocostaspora E.B.G. Jones et al., Bot. Mar. 26(7): 353 (1983)Mono typ i c genus i n the Halosphaer iaceae
(Microascales) with Kochiella as a sister group in astatistically well support clade (Sakayaroj et al. 2011a).No asexual morph is known.
Oedemium Link, in Willdenow, Willd., Sp. pl., Edn 4 6(1): 42(1824)
Oedemium minus (Link) S. Hughes was reported as theasexual morph of the type species of Chaetosphaerella, i.e.C. phaeostroma (Durieu & Mont.) E. Müll. & C. Booth byRéblová (1999b). The conidia ofOedemium minus develop ina cluster at the apex of apical swelling of the terminalpolytretic conidiogenous cell (Hughes and Hennebert 1963).Conidia of O. didymum (J.C. Schmidt) S. Hughes which isreported as the asexual morph of Chaetosphaerella fusca(Fuckel) E. Müll. & C. Booth (Ellis 1971) are produced inacropetal chains of the apex of polytretic conidiogenous cells;not more than two chains occur on a conidiogenous cell(Réblová 1999b). Veramycina elegans Subram. was describedon the basis of culture studies of Oedemium minus(Subramanian 1993). Réblová (1999b) observed that conidi-ophores of Veramycina elegans synanamorphs were foundamong conidiophores of Oedemium minus developing fromthe same mycelium in a collection made on Carpinus betulusin the Czech Republic (Herbarium – M. Réblová 866/96).Since, Veramycina was published as a new genus, based onthe culture of Oedemium minus; we consider O. minus and
Fungal Diversity

Veramycina elegans as synonyms. However, the types of thesegenera have not been linked and therefore Chaetosphaerellaand Oedemium are maintained as distinct genera until shownotherwise.
Okeanomyces K.L. Pang & E.B.G. Jones, Bot. J. Linn. Soc.146(2): 228 (2004)
Monotypic genus in the Halosphaeriaceae, that was intro-duced by Pang et al. (2004) to accommodate Halosphaeriacucullata (Kohlm.) Kohlm. In their study, phylogenetic analy-ses of LSU rDNA placed H. cucullata in a distant clade fromthe type species of the genusHalosphaeria (H. appendiculata).Periconia prolificaAnast. which is not the type of Periconia, isthe asexual morph of the type species,Okeanomyces cucullatus(Kohlm.) K.L. Pang & E.B.G. Jones.
Ondiniella E.B.G. Jones et al., Bot. Mar. 27(3): 136 (1984)Monotypic genus in the Halosphaeriaceae that was intro-
duced by Jones et al. (1984) to accommodate Halosphaeriatorquata Kohlm. The fungus was excluded from the genusHalosphaeria based on the nature and mode of developmentof the appendages at the ultrastructure level. Multi-gene phy-logeny confirmed its position in Halosphaeriaceae withO. torquata in a distant clade from the type species,Halosphaeria appendiculata. Ondiniella torquata (Kohlm.)E.B.G. Jones et al. forms a well supported clade withToriella tubulifera (Kohlm.) Sakay.et al. as a sister group(Sakayaroj et al. 2011a, b). No asexual morph is known.Ophioceraceae Klaubauf et al., Stud. Mycol. 79: 104 (2014)
Ophioceraceae includes the single genus Ophioceras(Klaubauf et al. 2014). Based on the sequence data Klaubaufet al. (2014) showedOphioceras clearly clusters separate fromthe Magnaporthaceae, and hence introduce a new familyOphioceraceae
Ophioceras Sacc., Syll. fung. (Abellini) 2: 358. 1883.Morphologically Ophioceras is somewhat similar to
Gaeumannomyces, however the two genera can easily be dis-tinguished by the aquatic habit of Ophioceras, occurring onwood and herbaceous material, versus the plant pathogenicnature ofGaeumannomyces (Chen et al. 1999; Klaubauf et al.2014). In addition based on sequence analysis of LSU, ITS,RPB1, actin and calmodulin gene data, Klaubauf et al. (2014)showed thatOphioceras clearly clusters separate to the familyMagnaporthaceae and hence a new family, Ophioceraceae,was introduced.
Ophiodeira Kohlm. & Volkm.-Kohlm., Can. J. Bot. 66(10):2062 (1988)
Monotypic genus ofHalosphaeriaceae that was introducedby Kohlmeyer and Volkmann-Kohlmeyer (1988) to accom-modate the marine fungus, O. monosemeia Kohlm. &Volkm.-Kohlm. Multi-gene phylogeny confirms its position
in Halosphaeriaceae with O. monosemeia in a well-supported clade with Oceanitis species as a sister group(Sakayaroj et al. 2011a). No asexual morph is known(Kohlmeyer and Volkmann-Kohlmeyer 1988).
Ophiodiaporthe Y.M. Ju et al., Mycologia 105(4): 868 (2013)This monotypic genus was introduced by Fu et al. (2013)
for a pathogen causing a wilt disease of Cyathea lepifera inTaiwan with Ophiodiaporthe cyatheae Y.M. Ju et al. as thetype species. Phylogenetic analyses based on combined genesequences placed O. cyatheae in Diaporthaceae.
Ophiodothella (Henn.) Höhn., Sber. Akad. Wiss. Wien,Math.-naturw. Kl., Abt. 1 119: 940 [64 repr.] (1910)
Boyd (1934) descr ibed the asexual morph ofOphiodothella vaccinii Boyd as an acervulus, withoutassigning it to a particular genus (Hanlin 1990a). The asexualmorph ofOphiodothella vaccinii Boyd was later assigned to anew monotypic genus Acerviclypeatus as A. poriformansHanlin (Hanlin 1990b). This connection was based on culturalstudies. Barr (1990) indicated that Septoria angustissima Peck(Peck 1911) was probably an earlier name for Ophiodothellavaccinii. Hanlin (2013) examined the type species ofAcerviclypeatus poriformans, Ophiodothella vaccinii andSeptoria angustissima and suggested they represent the sametaxon. Then he suggested a new name as Ophiodothellaangustissima (Peck) Hanlin & M.C. González. Acerviclypeatushas not been linked toO. atromaculans (Henn.) Höhn., which isthe generic type of Ophiodothella and therefore both generashould be currently retained. Ophiodothella is placed inPhyllachoraceae.
Orbimyces Linder, Farlowia 1: 404 (1945)Monotypic asexual marine genus that groups in the
Lulworthiales with low support (Jones et al. 2009b). Furthermolecular studies are required to determine its taxonomic af-finities within the order. The fungus sporulates in culture butno sexual morph was observed (Jones, pers. comm.).
OrbiocrellaD. Johnson et al., Mycol. Res. 113(3): 286 (2009)This genus was introduced to accommodate Torrubiiella
petchiii Hywel-Jones which was recognized as a distinctlineage of pathogens of scale insect in Clavicipitaceae,based on morphological characters and phylogenetic analy-sis of nrSSU, nrLSU, TEF, RPB1 and RPB2 gene data(Johnson et al. 2009).
Orcadia G.K. Sutherl., Trans. Br. mycol. Soc. 5(1): 151(1915) [1914]
Monotypic marine ascomycete genus growing on sea-weeds based on morphological observations, and requiringfurther collection, isolation and sequencing to determine itsrelationship (Jones et al. 2009b). Sutherland (1915) referred
Fungal Diversity

the genus to the Hyponectriaceae (Xylariales), in havinginterthecial elements. Rossman et al. (1999) suggested reten-tion in the Pezizales based on its operculate asci. However,Jones et al. (2009b) decided to place the genus Orcadia asSordariomycetes genera incertae sedis which we follow here.No asexual morph is known.
Oxydothis Penz. & Sacc., Malpighia 11: 505 (1898)Genus of ascomycetes with ascomata immersed in palm
tissues beneath the host epidermis. Kang et al. (1999a) referredthe genus to the Clypeosphaeriaceae, while Jones et al.(2009b) placed marine species in Xylariales genera incertaesedis which we follow here. Further molecular studies arerequired to determine its taxonomic position.
Panorbis J. Campb. et al., Mycologia 95(3): 544 (2003)Monotypic genus in Halosphaeriaceae that was introduced
by Campbell et al. (2003) based on phylogenetic analyses ofSSU and LSU rDNA to accommodate one Halosarpheia spe-cies, H. viscosa (I. Schmidt) Shearer & J.L. Crane that did notgroup with the type species H. fibrosa Kohlm. & E. Kohlm .Sakayaroj et al. (2011a) showed that the species groups withHalosarpheia marina (Cribb & J.W. Cribb) Kohlm. with weaksupport. No asexual morph is known (Campbell et al. 2003).
Papulosa Kohlm. & Volkm.-Kohlm., Syst. Ascomycetum 11:96 (1993)
Monotypic marine genus, referred to the familyPapulosaceae (Diaportheomycetidae, Sordariomycetes) andsupported by morphological and molecular data (Erikssonand Winka 1997).
Paracremonium Lombard & Crous, in Lombard et al.,Studies in Mycology 80: 233 (2015)
The genus Paracremonium was introduced to accommo-date different strains from a group of fungi previously treatedas Acremonium recifei (Leão & Lôbo)W. Gams. This genus ismorphologically different from other acremonium-like generaby the formation of sterile coils from radiate conidiophoresand hyphae with inconspicuously swollen septa (Lombardet al. 2015). Phylogenetic studies showed this genus belongsto family Nectriaceae (Lombard et al. 2015). This genus istypified by P. inflatum L. Lombard & Crous.
Payosphaeria W.F. Leong, Bot. Mar. 33: 511 (1990)Monotypic marine ascomycete genus in Hypocreales, gen-
era incertae sedis, growing on submerged wood and introducedby Leong et al. (1990) based on morphological observations.The species requires further collection, isolation and sequenc-ing to determine its phylogenetic placement in the Ascomycota(Jones et al. 2009b). The type species Payosphaeria minutaW.F. Leong was collected from mangrove wood in Malaysiaand Singapore. No asexual morph is known.
Pedumispora K.D. Hyde & E.B.G. Jones, Mycol. Res. 96: 78(1992)
Pedumispora was introduced by Hyde and Jones (1992) fora marine ascomycete. It was tentatively referred to the orderDiaporthales. Phylogenetic analyses reveal that Pedumisporarhizophorae K.D. Hyde & E.B.G. Jones is distantly placedfrom the Diaporthales, and groups in the Diatrypaceae,Xylariales with strong support (Klaysuban et al. 2014).Pedumispora rhizophorae shares many features with membersof the Diatrypaceae, including its saprobic habitat ondecaying plant material, ascomata embedded in apseudostroma, the presence of paraphyses and pigmentedascospores. However, P. rhizophorae differs from othermembers of the Diatrypaceae in a number of aspects. Inspecies of Diatrypaceae most ascospores are allantoid andunicellular, whereas P. rhizophorae has septate, filiform as-cospores with the apical cells lacking contents andappearing as a hook-like appendage (Klaysuban et al.2014). No asexual morph is known.
Peethambara Subram. & Bhat, RevueMycol., Paris 42(1): 52(1978)
The genus Peethambara as typified by P. sundaraSubram. & Bhat and was isolated from Macaranga indicain India. Peethambara was earlier classified as Hypocrealesorder incertae sedis (Castlebury et al. 2004). In their revi-sion, Crous et al. (2014) showed Peethambara and relatedgenera form a monophyletic lineage with Stachybotrys andintroduced the novel family, Stachybotriaceae and this issupported in Wang et al. (2015).
Periconia Tode, Fung. mecklenb. sel. (Lüneburg) 2: 2 (1791)Periconia prolifica is an asexual morph linked to
Okeanomyces cucullatus (Kohlm.) K.L. Pang & E.B.G.Jones (= Remispora cucullata Kohlm.; Halosphaeriacucullata (Kohlm.) Kohlm.) by cultural observations and mo-lecular data (Sakayaroj et al. 2011a). According to the dataincluded in online databases such as MycoBank (Robert et al.2005) and Index Fungorum, the genus Periconia compriseswith about 183 published names and more than 20 have beentransferred to other genera. Presently only about 40 speciesbelong to this genus. The type species isPericonia lichenoides(Carmarán and Novas 2003; Kirk et al. 2008) and this andsome other species, cluster in the order Pleosporales familyincertae sedis.
Periglandula U. Steiner et al., in Steiner et al., Mycologia103(5): 1137 (2011)
This genus was introduced with Periglandula ipomoeae U.Steiner et al. as the type species which was isolated fromIpomoea asarifolia in Ecuador. Species live on the adaxial leafsurface of plants belonging to the family Convolvulaceae andapparently are epibionts. Phylogenetic study based on sequence
Fungal Diversity

analysis of b-tubulin, RNAPolymerase II subunit 1 (rpbA), andthe mitochondrial gene for ATP synthase F0 subunit A,g r o u p e d t h e P e r i g l a n d u l a i n t h e f a m i l yClavicipitaceae (Steiner et al. 2011).
Pestalotiopsis Steyaert, Bull. Jard. bot. État Brux. 19: 300(1949)
The sexual morph of Pestalotiopsis is Pestalosphaeria andonly 13 species are known as compared to the asexual morph(254 species names). Pestalotiopsis has been related toNeobroomella Petr. which was introduced by Petrak (1947).As such, the primal name is Neobroomella, but no sequencedata has linked with the asexual Pestalotiopsis morph.Maharachchikumbura et al. (2011) pointed out that the com-mon Pestalotiopsis name should be applied to both morphsand this is followed in this paper.
Phaeonectriella Eaton & E.B.G. Jones, Nova Hedwigia 19(3–4): 779 (1971)
A genus ofHalosphaeriaceae based on morphological andmolecular evidence (Sakayaroj et al. 2011a), forming a sistergroup to Panorbis and Halosarpheia. No asexual morph isknown.
Phialemoniopsis Perdomo et al., Mycologia 105(2): 408(2013)
Perdomo et al. (2013) proposed the new genusPhialemoniopsis based on Phialemoniopsis ocularis (Gené &Guarro) Perdomo et al. (= Sarcopodium oculorum Gené &Guarro) and Phialemoniopsis curvata (W. Gams & W.B.Cooke) Perdomo et al. (= Phialemonium curvatum W. Gams&W.B. Cooke and two new species,Phialemoniopsis cornearisPerdomo et al. and P. pluriloculosa Perdomo et al. The genuscomprises human pathogens and can be distinguished fromPhialemonium in having phialides and adelophialides, bothwithcollarettes, and in the development of sporodochium- orpycnidium-like conidiomata, unlike in Phialemonium sensustricto. The relationships of Phialemoniopsis has not beencompletely resolved and the genus is placed inSordariomycetes genera incertae sedis. Molecular analyses in-dicate that Phialemoniopsis species are distant from the typespecies of the genera Phialemonium, Sarcopodium andVolutella, and the orders Sordariales and Hypocreales.
Phlogicylindrium Crous et al., in Summerell et al., FungalDiversity 23: 340 (2006)
The genus Phlogicylindrium was introduced forP. eucalypti Crous et al., a species associated withEucalyptus leaves (Summerell et al. 2006; Crous et al.2011). Phlogicylindrium is characterised by erect flame-likeconidiomatal tufts (Summerell et al. 2006). ITS sequence datashowed it clusters within the family Amphisphaeriaceae(Summerell et al. 2006).
Phomatospora Sacc., Grevillea 3: 22 (1875)Genus of Magnaporthaceae (Magnaporthales) based on
molecular analyses (18S rDNA) (Cai et al. 2006a). This genusis probably polyphyletic and further studies are required witha wider sampling of taxa and genes.
PhragmitensisM.K.M. Wong et al., Bot. Mar. 41: 379 (1998)Genus of Hyponectriaceae (Xylariales) based on morpho-
logical observations and requiring a molecular study to con-firm its taxonomic position. Ascomata occur on Phragmites inthe intertidal zone of salt marshes (Wong et al. 1998b). Noasexual morph is known.
Pileomyces K.L. Pang & Jheng, Botanical Studies 53: 536(2012)
Monotypic marine genus in the Halosphaeriaceae de-scribed from a bamboo culm collected at Yingkeshih,Taiwan (Pang and Jheng 2012b). Ascospores ofP. formosanus K.L. Pang & Jheng are similar to those ofAniptodera and Phaeonectriella. No asexual morph is known.
Pisorisporiaceae Réblová & J. Fourn., Persoonia 34: 43(2015)
Réblová et al. (2015) placed Pisorisporium andAchroceratosphaeria in the family Pisorisporiaceae basedon LSU, SSU and RPB2 sequence data and morphology.This family is placed in the order Pirisporiales.
Pisorisporium Réblová & J. Fourn., Persoonia 34: 45 (2015)This genus has two species which were found on
submerged deciduous wood. Molecular analysis andmorphology studies place this genus in the familyPisorisporiaceae (Réblová et al. 2015). No asexualmorph is known.
Plagiostoma Fuckel, Jb. nassau. Ver. Naturk. 23–24: 118(1870) [1869–70]
The genus was revisited by Mejía et al. (2011) with25 accepted species. The sexual genera Cryptodiaportheand Rostrocoronophora and asexual genus Diplodinawere considered synonyms. Diplodina is the older nameand has priority and thus the mycological communitymust decide which name to retain. The genus is placedin Gnomoniaceae (Fig. 2).
Platytrachelon Réblová, Mycologia 105(2): 466 (2013)Réblová (2013a) introduced this new genus with a single
species, previously named Ceratosphaeria abietis Réblová,based on morphology and phylogenetic analyses. This wood-inhabiting fungus has a simple and inconspicuous morphology.Phylogenetic analyses of combined genes (LSU, SSU andRPB2) indicated that C. abietis has a relationship withPapulosaceae and is better placed in Diaportheomycetidae,
Fungal Diversity

genera incertae sedis.
Plectosphaerella Kleb., Phytopathologische Zeitschrift 1: 43(1930)
A polyphyletic genus with various species assigned toDothideales, Phyllachorales, Pleosporales, and Xylariales(Index Fungorum 2015). Plectosphaerella was introduced toaccommodate the type species Plectosphaerella cucumerisKleb., and currently some 13 species are assigned to the ge-nus. The type species of the asexual genus Spermosporina isS. alismatis (Oudem.) U. Braun with a sexual morph inPlectosphaerella which has priority and is a Nom. rejic., seeArt. 14.7 (Carlucci et al. 2012) and thus Spermosporina istreated as a synonym.
PlectosphaerellaceaeW.Gams et al., NovaHedwigia 85 (3–4):476 (2007)
The family was introduced by Zare et al. (2007) to accom-modate the genera Acrostalagmus, Gibellulopsis andMusicillium based on LSU and ITS sequences, and forms awell-supported clade within the Microascales (Réblová et al.2011a). Subsequently, Stachylidium has also been referred tothe family. The family includes many asexual genera that arelinked to their putative sexual morphs: Acrostalagmus,Plectosporium and Stachylidium.
Pleurotheciella Réblová et al., Mycologia 104(6): 1304(2012)
Réblová et al. (2012) described this genus from decayingwood submerged in freshwater with two new speciesPleurotheciella rivularia Réblová et al. and P. centenariaRéblová et al. Phylogenetic analyses of combined genes indi-cated that Pleurotheciella is closely related to the generaPleurothecium and Sterigmatobotrys, but forms a well-supported monophyletic clade. The genus is placed inHypocreomycetidae genera incertae sedis.
Pleurothecium Höhn., Ber. dt. bot. Ges. 37: 154 (1919)Pleurothecium is considered as a hyphomycete genus, with
six accepted species (Seifert et al. 2011); only P. recurvatumhas a sexual morph, described as Carpoligna pleurothecii(Fernández et al. 1999). Pleurothecium is typified byP. recurvatum (Morgan) Höhn. Réblová et al. (2012) showedthe type species to cluster near Sterigmatobotrys andPhaeo i s a r i a s p e c i e s . Th e g enu s i s p l a c ed i nHypocreomycetidae genera incertae sedis. Pleurotheciumsemifecundum Réblová et al. is placed in Savoryellales familyincertae sedis and the genus is presently polyphyletic.
Poronia Willd., Fl. berol. prodr.: 400 (1787)Rogers and Læssøe (1992) introduced Podosordaria ingii
J.D. Rogers & Læssøe, which was later synonymised asPoronia ingii (J.D. Rogers and Læssøe) J.D. Rogers et al.,
which has an asexual morph belonging to Lindquistia.Lindquistia was introduced to accommodate Lindquistiaindica Subram. & Chandrash. As reported by Rogers andLæssøe (1992) the sexual and asexual morphs are similar.The asexual morph was also produced in cultures from asco-spores. Therefore the sexual–asexual link between Lindquistiaand Poronia ingii were established (Rogers and Læssøe1992). Since, Poronia was introduced before Lindquistia,the latter name should be suppressed (Stadler et al. 2013).
ProxipyriculariaKlaubauf et al., Stud. Mycol. 79: 110 (2014)Proxipyricularia is a plant pathogenic genus in the family
Pyriculariaceae which is typified by P. zingiberis (Klaubaufet al. 2014). Proxipyricularia is morphologically similar toPyricularia, but phylogenetically distinct based on combinedsequence analysis of LSU, ITS, RPB1, actin and calmodulingene data (Klaubauf et al. 2014).
Pseudocatenomycopsis Crous & L.A. Shuttlew., in Crouset al., Persoonia, Mol. Phyl. Evol. Fungi 31: 221 (2013)
This monotypic genus known only by its asexual morph wasintroduced by Crous et al. (2013) with P. rothmanniae Crous &L.A. Shuttlew. as the type species and placed inCoronophorales genera incertae sedis based on sequence data.
PseudocosmosporaC. Herrera & P. Chaverri, Mycologia 105:1291 (2013)
Herrera et al. (2013b) described this genus to accommodatethe type species Pseudocosmospora eutypellae C. Herrera &P. Chaverri and nine additional species based on molecularanalyses (ITS, LSU, MCM7, RPB1, TEF, tub) which indicat-ed this group is phylogenetically distinct from other previous-ly segregated genera under Nectriaceae.
Pseudoidriella Crous & R.G. Shivas, in Crous et al.,Persoonia, Mol. Phyl. Evol. Fungi 27: 135 (2011)
The monotypic genus was introduced by Crous et al.(2011) for an idriella-like taxon, but with similarities toMicrodochium. Based on the sequence data Crous et al.(2011) placed it in incertae sedis.
Pseudomassaria Jacz., Bull. Herb. Boissier 2: 663 (1894)Jaklitsch and Voglmayr (2012) recollected the type spe-
cies of Pseudomassaria, P. chondrospora (Ces.) Jacz., andthe i r mo lecu la r ana lyses p l aced the genus inHyponectriaceae, Xylariales. Pseudomassaria andLeiosphaerella praeclara (Rehm) Höhn. are closely related(Jaklitsch and Voglmayr 2012).
PseudopestalotiopsisMaharachch. et al. Stud. Mycol. 79: 180(2014)
The new genus Pseudopestalotiopsis, which segregatesfrom Pestalotiopsis was proposed based on the type
Fungal Diversity

Pseudopestalotiopsis theae (Sawada) Maharachchikumburaet al. by Maharachchikumbura et al. (2014). The epitype ofPestalotiopsis theae (Sawada) Steyaert was designated fromfresh leaves of Camellia sinensis collected in Thailand(Maharachchikumbura et al. 2013b). Pseudopestalotiopsiscan be distinguished from Neopestalotiopsis andPestalotiopsis by sequence data and generally dark-colouredconcolourous median cells with indistinct conidiophores(Maharachchikumbura et al. 2014). The genus belongs inthe family Amphisphaeriaceae.
Pseudopyricularia Klaubauf et al., Stud. Mycol. 79: 110(2014)
Based on sequence analysis of a combined dataset ofLSU, ITS, RPB1, ACT and CAL sequences, Klaubaufe t a l . ( 2 0 1 4 ) p l a c e d t h r e e r e l a t e d s p e c i e s(Pseudopyricularia cyperi Klaubauf et al., P. higginsii(Luttr.) Klaubauf et al. and P. kyllingae Klaubaufet al.) in the new genus Pseudopyricularia in the familyPyriculariaceae. Species in Pseudopyricularia are pri-marily distinguished from Pyricularia by having short,determinate, brown conidiophores, with an apical rachiswith flat-tipped denticles (Klaubauf et al. 2014).
Purpureocillium Luangsa-ard et al., in Luangsa-ard et al.,FEMS Microbiol. Lett. 321(2): 144 (2011)
This genus was introduced by Luangsa-ard et al. (2011) toaccommodate a species of Paecilomyceswhich has frequentlybeen found as the causal agent of infections in man and othervertebrates. Molecular data showed it was not related toPaecilomyces but could be placed in Ophiocordycipitaceae.A second species was added by Perdomo et al. (2013).
Pustulomyces D.Q. Dai et al., in Dai et al., Cryptog. Mycol.35(1): 64 (2014)
Pustulomyces is a monotypic genus introduced byDai et al.(2014) with Pustulomyces bambusicola Dai et al., as the typespecies from bamboo. A combined data set of LSU and TEFsequences showed that Pustulomyces is a new genus inDiaporthaceae (Dai et al. 2014).
Pyricularia Sacc., Michelia 2(no. 6): 20. 1880.In the recent revision of Klaubauf et al. (2014), Pyricularia
was placed as the type of the newly introduced familyPyriculariaceae. Species belonging to Pyricularia are plantpathogens (Klaubauf et al. 2014). Presently there are 78 spe-cies epithets in the genus including the type P. grisea Sacc.
Pyriformiascoma Daranagama et al., Fungal Diversity (inpress) (2015)
Pyriformiascoma was introduced to accommodate the typespecies Pyriformiascoma trilobatum Daranagama et al.(Daranagama et al. 2015). Pyriformiascoma produces simple,
brown conidiophores with terminal conidiogenesis cells bearingbrown globose conidia in culture. Phylogenetic analysis placeP. trilobatum as a basal clade to both hypoxyloid and xylaroidXylariaceae parallel toCreosphaeria sassafras (Schwein.) Y.M.Ju et al. and Barrmaelia macrospora (Nitschke) Rappaz withhigh bootstrap support (Daranagama et al. 2015).
Pyrigemmula D. Magyar & Shoemaker, in Magyar et al.,Mycol. Progr. 10(3): 310 (2011)
This monotypic genus was introduced by Magyar et al.(2011) for P. aurantiaca D. Magyar & Shoemaker whichwas collected from inner bark of living woody hosts inHung a r y. Mo l e c u l a r d a t a p l a c e t h e g e nu s i nChaetosphaeriaceae.
Rectifusarium Lombard et al., in Lombard et al., Studies inMycology 80: 229 (2015)
Lombard et al. (2015) introduced this genus to accommo-date fusarium-like species lacking sporodochia, previouslytreated as F. ventricosum. DNA sequence data confirmed thatthey formed a well-supported distinct clade in the familyNectriaceae. The genus Rectifusarium is typified byR. ventricosum (Appel & Wollenw.) L. Lombard & Crous.Regiocrella P. Chaverri & K.T. Hodge, Mycologia 97(6):1232 (2006) [2005]
This genus was described from samples collected inCameroon and China (Chaverri et al. 2005). Morphologicaland molecular evidence place th is genus underClavicipitaceae (Chaverri et al. 2005).
Remispora Linder, Farlowia 1(3): 409 (1944)A phylogenetic evaluation of Remispora, based on three
loci (LSU, SSU and RPB2), demonstrated that the genuswas polyphyletic (Sakayaroj et al. 2011a). Consequently, spe-cies not grouping in Remispora sensu stricto were moved tonew genera (Kochiella, Tubakiella), see elsewhere. This genusis based on the type species R. maritima which forms a well-supported monophyletic clade with Sablicola as a sister clade(Sakayaroj et al. 2011a). No asexual morph is known.
Reticulascaceae Réblová &W. Gams, in Réblová et al., Stud.Mycol. 68(1): 180 (2011)
Cylindrotrichum (= Reticulascus) and Porosphaerellopsisare presently included in the family Reticulascaceae. Thisfamily was introduced by Réblová et al. (2011a) based onmolecular analysis of ITS, LSU, SSU and RPB2 sequencedata. The dematiaceous Cylindrotrichum, Kylindria andSporoschismopsis , l inked as anamorphs with theReticulascaceae (Réblová et al. 2011a).
Retrostium Nakagiri & Tad. Ito, Mycologia 89: 485 (1997)Monotypic marine genus in the Spathulosporaceae, order
incertae sedis, Sordariomycetes (Jones et al. 2009b).
Fungal Diversity

Spathulospora (an algal-inhabiting species) and the type of thefamily has been shown to be well-placed in the Lulworthiales(Inderbitzin et al. 2004). Further collections, isolation and se-quencing required to resolve its taxonomic position.
Rhopalostroma D. Hawksw., Kew Bull. 31(3): 422 (1977)A study conducted by Stadler et al. (2010) showed that the
genus Ropalostroma, exclusively reported from thepalaeotropics, was closely related to the daldinoidXylariaceae and the two predominantly neotropical generaPhylacia and Thamnomyces. The study included the use ofmicroscopic methods, secondary metabolite profiling and ITSnrDNA sequences of cultures obtained from fresh materialcollections of the species R. angolense (Welw. & Curr.) D.Hawksw. Daranagama et al. (2014) studied the affinitiesof R. lekae Whalley et al. with R. angolense and othermembers of Xylariaceae using a polythetic approach.According to the multigene analysis Phylacia ,Rhopalostroma and Thamnomyces form a separate cladeand the monophyletic origin of Thamnomyces andRhopalostroma is supported by high statistical support(Darnagama et al. 2014). Stadler et al. (2010) andDaranagama et al. (2014) observed the nodulisporium–likeasexual morph from both R. angolense and R. lekae.
Rimaconus Huhndorf et al., Mycologia 93(6): 1073 (2001)Rimaconus was introduced by Huhndorf et al. (2001) for
the type species R. jamaicensis (Seaver) Huhndorf et al. andpresently comprises two species (Huhndorf and Miller 2011).Phylogenetic study (Huhndorf and Miller 2011) placedRimaconus in a group close to Helminthosporiaceae and inour analysis (Fig. 2) the genus forms a separate clade close toCephalothecaceae, Chaetosphaeriales and Meliolales.Currently Rimaconus is placed in Chaetosphaeriales generaincertae sedis.
Riomyces A. Ferrer et al., Mycologia 104(4): 876 (2012)This monotypic genus was introduced from submerged
woody debris in freshwater collected in Costa Rica (Ferreret al. 2012). Molecular analysis placed the genus inSordariomycetidae genera incertae sedis.
Rodentomyces Doveri et al., in Doveri et al., Fungal Diversity42: 61 (2010)
This monotypic genus was introduced by Doveri et al.(2010) based on morphology and phylogenetic analysis.Rodentomyces, type species R. reticulatus Doveri et al., isplaced in Nectriaceae, based on ITS and LSU sequence dataanalysis. It has trichothecium-like asexual morphs similar toasexual morphs of a few other hypocrealean taxa.
Rosasphaeria Jaklitsch & Voglmayr, Fungal Diversity 52(1):93 (2012)
This monotypic genus was introduced by Jaklitsch andVoglmayr (2012) based on multi-gene phylogenetic analysesof ITS, LSU, RPB2 and TEF gene data. Its type species isRosasphaeria moravica (Petr.) Jaklitsch & Voglmayr. It hasa pycnidial asexual morph that was seen in culture.Phylogenetically it is close to Eucasphaeria and is placedin the family Niessliaceae.
Rosellinia De Not., G. bot. ital. 1(1): 334 (1844)The genus Rosellinia was introduced by De Notaris (1844)
to accommodate species with uniperitheciate, gregarious,stromata and revised to include species with superficial, con-spicuous, carbonaceous, ostiolate stromata with subicula(Petrini 2013). Their asexual morphs are geniculosporium-like, nodulisporium-like and Dematophora which are typicalof Xylariaceae (Petrini 2013). Petrini (2013) accepted 142species in Rosellinia.
Dematophora was described by Hartig (1883) withD. necatrix R. Hartig as the type species. Viala (1891) andlater Prillieux (1904) observed ascocarps apparently associat-ed with D. necatrix on diseased vine roots. Hansen et al.(1937) described perithecia on diseased apple roots infectedby D. necatrix, which they happened to keep in moist cham-bers for a long time. A connection between germinating co-nidia of D. necatrix and Rosellinia was established in culture.Thus the perithecial morph of D. necatrix was RosellinianecatrixBerl. ex Prill. The same study proved that the conidialmorphs developed in the cultures ofRosellinia buxi Fabre, andwere ei ther mononematus or synnematous as inDematophora. The asexual morph Dematophora is linked tothe sexual morph Rosellinia and was synonymised underRosellinia by Stadler et al. (2013).
Roselliniella Vain., Acta Soc. Fauna Flora fenn. 49(no. 2): 77(1921)
Based on newly obtained LSU sequences from RoselliniellaatlanticaMatzer &Hafellner and R. euparmeliicolaMillanes &D. Hawksw., Hawksworth et al. (2010) noted that Roselliniellahad to be placed inHypocreales genera incertae sedis and not inSordariales genera incertae sedis. Thus, the family placementcould not be resolved from the sequences obtained.
Rostrohypoxylon J. Fourn. & M. Stadler, in Fournier et al.,Fungal Diversity 40: 24 (2010)
This monotypic genus introduced by Fournier et al. (2010)was based on a new combination of asexual-sexual morphcharacters. The type species is R. terebratum J. Fourn. & M.Stadler. The status of this taxon is supported by secondarymetabolite profiling. Morphologically, it is characterized byerumpent, strongly carbonaceous stromata with stout ostiolarnecks and deep cylindrical holes.Rostrohypoxylon produced asporothrix-like to virgariella-like asexual morph in culture. Itis presently placed in family Xylariaceae.
Fungal Diversity

Rostrupiella Jørg. Koch et al., Bot. Mar. 50(5–6): 295 (2007)A monotypic marine genus in Lulworthiales with the type
species R. danica Jørg. Koch et al., grouping with Lulwoanauniseptata (Nakagiri) Kohlm. and various Lulworthia sensulato species as a sister clade (Koch et al. 2007; Jones et al.2009a). No asexual morph is known.
Rugonectria P. Chaverri & Samuels, in Chaverri et al., Stud.Mycol. 68(1): 73 (2011)
This genus was introduced by Chaverri et al. (2011) withR. rugulosa (Pat. & Gaillard) Samuels et al. as the type spe-cies. The genus clustered in Nectriaceae and hascylindrocarpon-like asexual morphs.
Ruwenzoria J. Fourn. et al., in Stadler et al., Mycol. Progr.9(2): 171 (2010)
Ruwenzor ia was in t roduced to accommoda teR. pseudoannulata J. Fourn. et al., which is characterized byhemispherical to peltate, superficial, sessile stromata with in-distinct perithecial outlines and raised ostiolar areas. The asex-ual morph is either sporothrix-like or less frequentlynodulisporium-like (Stadler et al. 2010). Apart from morphol-ogy both chemotaxonomic and ITS sequence data were ob-tained by Stadler et al. (2010). Ruwenzoria belongs to thehypoxyloid Xylariaceae, and has particularly close affinitiesto the genus Daldinia.
RuzeniaO. Hilber, The Genus Lasiosphaeria and Allied Taxa(Kelheim): 7 (2002)
This is a monotypic genus in Helminthosphaeriaceaetypified by Ruzenia spermoides (Hoffm.) O. (Milleret al. 2014).
SaagaromycesK.L. Pang & E.B.G. Jones, Mycol. Prog. 2: 35(2003)
In a phylogenetic evaluation of genera with polar unfurlingappendages the genus Halosarpheia was found to be polyphy-letic and taxa not grouping with the type species H. fibrosaKohlm. & E. Kohlm. were transferred to new genera.Saagaromyces accommodates the marine ascomyceteS. ratnagiriensis (S.D. Patil & Borse) K.L. Pang & E.B.G.Jones (Pang and Jones 2004) in the family Halosphaeriaceae(Jones et al. 2009a). No asexual morph is known. Littispora is asynonym of Saagaromyces, both taxa were described at thesame time, but the latter name has priority (Pang and Jones2004; Jones et al. 2009a).
Sablicola E.B.G. Jones et al., Can. J. Bot. 82: 486 (2004)Monotypic marine ascomycete growing on submerged
wood, with its placement in the Halosphaeriaceae confirmedby 28S rDNA sequences (Pang et al. 2004). The genus forms asister group to Remispora sensu stricto (Pang et al. 2004;Sakayaroj et al. 2011a). No asexual morph is known.
Samuelsia P. Chaverri & K.T. Hodge, in Chaverri et al., Stud.Mycol. 60: 59 (2008)
Chaverri et al. (2008) introduced this new genus based onmorphology and analysis of 28S, TEF and RPB1 gene dataand placed it in Clavicipitaceae. The asexual morph of thisgenus is aschersonia-like (Chaverri et al. 2008).
Sarocladium W. Gams & D. Hawksw., Kavaka 3: 57 (1976)[1975]
Sarocladium was extended by Summerbell et al. (2011) toinclude all members of the Acremonium strictum W. Gamsand A. bacillisporum (Onions &G.L. Barron)W. Gams cladesand groups in Hypocreales genera, incertae sedis.
Savoryella E.B.G. Jones & R.A. Eaton, Trans. Br. mycol. Soc.52(1): 161 (1969)
Awidespread genus of lignicolous ascomycetes in freshwaterand marine habitats, referred variously to Microascales (asHalosphaeriales), Sordariales and Xylariales, and to theHypocreales based on an 18S phylogenetic study (Cai et al.2006a). However, a subsequent study, using awider range of loci(LSU, SSU, 5.8S rRNA genes, RPB1, RPB2, TEF), showed itformed a new lineage in the Hypocreomycetidae(Sordariomycetes). Savoryella species form a monophyleticgroup in the order Savoryellales with high statistical support(Boonyuen et al. 2011). The genus has no known asexual morph.
Savoryellaceae Jaklitsch & Réblová, Index Fungorum 209: 1(2015)
The family Savoryellaceae was introduced by Jaklitschand Réblová (2015). Asexual morphs are dematiaceous hy-phomycetes, e.g., Canalisporium linked to Ascothailandiaand Monotosporella and Helicoon linked to Ascotaiwania.Species of Savoryellaceae are predominantly found in aquat-ic habitats such as freshwater, marine and brackish environ-ments, particularly on submerged wood (Jaklitsch andRéblová 2015). The order Savoryellales was introduced byBoonyuen et al. (2011) to include Ascotaiwania,Canalisporium and Savoryella. Savoryellales is an orderwhich includes family Savoryellaceae. In the current study,the order Savoryellales also includes Carpolignapleurothecii F.A. Fernández & Huhndorf (generic type),Conioscypha japonica Udagawa & Toyaz. and C. variusRéblová & Seifert and Pleurothecium semifecundumRéblová et al., which are placed in Savoryellales familyincertae sedis.
Scortechinia Sacc., Atti Inst. Veneto Sci. lett., ed Arti, Sér. 6 3:713 (1885)See under Scortechiniaceae.
Scortechiniaceae Huhndorf et al., in Huhndorf et al., Mycol.Res. 108(12): 1387 (2004)
Fungal Diversity

Mugambi and Huhndorf (2010) revisited the orderCoronophorales with DNA sequence data and showedScortechiniaceae to be monophyletic with the generaBiciliospora, Cryptosphaerella, Tympanopsis, Scortechinia,Scortechiniella and Scortechiniellopsis. Biciliospora,Scortechiniella and Scortechiniellopsis may be congeneric,but Tympanopsis and Scortechinia are distinct genera(Mugambi and Huhndorf 2010).
Scortechiniella Arx & E. Müll., Beitr. Kryptfl. Schweiz11(no. 1): 382 (1954)
See under Scortechiniaceae.
Sedecimiella K.L. Pang et al., in Pang et al., Bot. Mar. 53(6):495 (2010)
Pang et al. (2010) described Sedecimiella taiwanensisK.L.Pang et al. from twigs of Kandelia obovata collected inTaiwan and a further collection on unidentified mangrovewood at Futian Nature Reserve, Shenzhen, China.Phylogenetic analyses did not resolve its familial position inthe Hypocreales and therefore it is placed in Hypocreales,genera incertae sedis.A distinguishing feature of the ascomy-cete is its cylindrical unitunicate asci with 16 hyaline globoseascospores.
SeiridiumNees, Syst. Pilze (Würzburg): 22 (1816) [1816–17]Sequence data reveal Seiridium to represent a distinct genus
in the family Amphisphaeriaceae, which is characterised by 6-celled conidia (Jeewon et al. 2003b; Maharachchikumburaet al. 2014). Blogiascospora and Lepteutypa have been identi-fied as the sexual morph of Seiridium. However, Seiridium isthe oldest andmost commonly used name and therefore, shouldbe applied to represent both morphs. Maharachchikumburaet al. (2014) suggested the monotypic genus Pestalotia (1839)may be a synonym of Seiridium (1816), since both genera havesimilar morphologies. Presently 39 species epithets forSeiridium in Index Fungorum (2015).
Septofusidium W. Gams, Cephalosporium-art igeSchimmelpilze (Stuttgart): 147 (1971)
Gams (1971) placed the genus Septofusidium in the familyNectriaceae based on morphological characters. Lombardet al. (2015) placed this genus in family Tilachlidiaceae basedon phylogenetic analysis. It is typified by S. elegantulum(Pidopl.) W. Gams.
Simplicillium W. Gams & Zare, Nova Hedwigia 73(1–2): 38(2001)
This genus was introduced based on morphology and phy-logenetic analysis (Zare and Gams 2001a, b). Four taxa and onecombination were placed in this genus because of mainly sol-itary phialides (Zare and Gams 2001b). According to morpho-logical and 28S, ITS rDNA sequences analysis, a novel species
was proposed by Liu and Cai (2012). They also provided a keyfor Simplicillium. Five new species were discovered from soilsamples by Nonaka et al. (2013). The sexual morphs of thisgenus are mostly Torrubiella. species which belong in the fam-ily Cordycipitaceae (Zare and Gams 2001a, b; Liu and Cai2012; Nonaka et al. 2013; Index Fungorum 2015).
Sirococcus Preuss, Linnaea 26: 716 (1855)Rossman et al. (2007) showed, using molecular data that
the genus Sirococcuswith the species S. conigenus (Pers.) P.F.Cannon & Minter (type species), S. piceiola Rossman et al.and S. tsugae Castl. et al. belongs in Gnomoniaceae.
Sodiomyces A.A. Grum-Grzhim. et al., Persoonia 31: 154(2013)
Grum-Grzhimaylo et al. (2013) introduced the genus toaccommodate Helecoccum alkalinum Bilanenko & M.Ivanova which does not group in Hypocreales, based on amulti-locus gene phylogeny (ITS, 5.8S rDNA, 28S rDNA,18S rDNA, RPB2 and TEF). Sodiomyces alkalinus(Bilanenko&M. Ivanova) A.A. Grum-Grzhimaylo is the onlyspecies in the genus, a holomorphic alkaliphilic ascomycetewithin the Plectosphaerellaceae. Many other species in thePlectosphaerellaceae are also known to be alkaliphilic, forexample, Acremonium alcalophilum G. Okada.
Spadicoides S. Hughes, Can. J. Bot. 36: 805 (1958)This asexual genus has been associated with
Tengiomyces (Réblová 1999b), but there is no moleculardata to establish the connection. Spadicoides is polyphy-letic and has affinities with divergent lineages ofDothideomycetes (Pleosporales) and Sordariomycetes. ALSU sequence generated for S. bina (Corda) S. Hughes( the type spec i es ) was found to be s i s t e r toChaetosphaeriales, but the sequence is from a non typestrain (Shenoy et al. 2010). The genus is therefore tenta-tively placed in Sordariomycetidae genera incertae sedis.
Spataporthe Bronson et al., Int. J. Pl. Sci. 174(3): 278–292(2013)
The fossil genus of Gnomoniaceae was introduced byBronson et al. (2013) and is one of the earliest representativesof Diaporthales providing a minimum age of 136 Ma for theorder.
Spathulospora A.R. Caval. & T.W. Johnson, Mycologia 57:927 (1965)
Marine genus with Spathulospora phycophila A.R. Caval. &T.W. Johnson as the type species and referred to theSpathulosporomycetes, Spathulosporomycetidae (Locquin1984), Spathulosporales (Kohlmeyer 1973), Spathulosporaceae(Kohlmeyer 1973) based on morphological observations.However, two Spathulospora species (S. antarctica Kohlm.,
Fungal Diversity

S. adelpha Kohlm.) have been showen to group in the orderLulworthiales with weak support (Inderbitzin et al. 2004;Campbell et al. 2005; Jones et al. 2009b). Further collections,isolation and sequencing are required to determine the phyloge-netic placement of this genus in theAscomycota, especially as thetype species has yet to be sequenced. We presently preferSpathulospora to be placed in Spathulosporaceae until the typespecies has been sequenced.
Sphaerostilbella (Henn.) Sacc. & D. Sacc., Syll. fung.(Abellini) 17: 778 (1905)
Sphaerostilbella and Gliocladium are most likely the samegenera and thus Rossman et al. (2013) proposed to protect theyounger name Sphaerostilbella against Gliocladium, whichthey thought would favour clarity of communication; this isfollowed here. The genus belongs in Hypocreaceae.
Sporidesmium Link, Mag. Gesell. naturf. Freunde, Berlin3(1–2): 41 (1809)
Sporidesmium is in its traditional sense, is a specious genus,with around 330 species (Seifert et al. 2011). Réblová andWinka (2001) employed gene sequence-data to connectEllisembia folliculata (Corda) Subram. (as Sporidesmiumfolliculatum Corda) with Lecythothecium duriligni Réblová& Winka and Stanjehughesia hormiscioides (Corda)Subram. (as Sporidesmium hormiscioides Corda) with(Umbrinosphaeria caesariata (Clinton & Peck) Réblová) inChaetosphaeriaceae. Sporidesmium sensu lato has also beenassociated with other sexual morphs in Asterinaceae (Eupelterapaneae Hansf . , E. amicta Syd. and Placosomanothopanacis Syd.), Chaetosphaeriaceae (Chaetosphaeriacapitata Sivan. & H.S. Chang, Miyoshiella fusisporaKawam., M. larvata Réblová and M. triseptata (Shoemaker& G.P. White) Réblová), Cucurbitariaceae (Cucurbitariavarians Hazsl.),Micropeltidaceae (Akaropeltella kielmeyerae(Bat. & J.L. Bezerra) M.L. Farr) and Trichosphaeriaceae(Eriosphaeria aggregata E. Müll. & Munk) (detailed inShenoy et al. 2006). Some species of Sporidesmium reported-ly produce Chloridium, Idriella, selenosporella-like and un-named phialidic synanamorphs (detailed in Seifert et al.2011). The associations are yet to be supported by DNA-sequence data.
In a phylogenetic study, Shenoy et al. (2006) employedmulti-gene phylogenetic analysis to connect Sporidesmiumand morphologically similar genera such as Ellisembia,Linkos ia , Repe tophragma , Spor idesmie l l a andStanjehughesia with Bsexual morph^ families and their studyconcluded that Sporidesmium is polyphyletic and species arephylogenetically distributed in multiple families ofDothideomycetes and Sordariomycetes. There are nineSporidesmium species, including one undesignated species(Groenewald et al. 2008; Shearer et al. 2009; Shenoy et al.2006) that have been sequenced, but the type species,
Sporidesmium atrum Link is yet to be sequenced and thusthe genus is placed in Ascomycota genera incertae sedis.
Sporoschismopsis Hol.-Jech. & Hennebert, Bull. Jard. Bot.natn. Belg. 42(4): 385 (1972)
Réblová et al. (2014) introduced Sporoschismopsisangustata Réblová with a Porosphaerellopsis sexualmorph. This represents the second known and experimen-tally proven sexual-asexual morph link betweenPorosphaerellopsis and Sporoschismopsis. In the samestudy; Porosphaerellopsis is synonymised underSporoschisma as the latter genus is the older name.Sporoschismopsis was positioned in Reticulascaceaewhere it is transferred here (Réblová 2013b).
Stachybotrys Corda, Icon. fung. (Prague) 1: 21 (1837)Stachybotrys albipes (Berk. & Broome) S.C. Jong &
Davis, has been linked to a sexual morph reported asMelanopsamma pomiformis (Pers.) Sacc. (type) based ongene sequence data (Castlebury et al. 2004). Stachybotrys isthe oldest name and is frequently used in literature andadopted by Wang et al. (2015). Ornatispora is also a syno-nym. The genus Stachybotryswas earlier classified as incertaesedis in the order Hypocreales and based on sequence data.Crous et al. (2014) transferred it to newly established familyStachybotriaceae.
S tachy l i d ium Link , Magaz in de r Gese l l s cha f tNaturforschenden Freunde Berlin 3: 15 (1809)
Stachylidium is an asexual genus variously classified inBionectriaceae, Xylariaceae, and anamorphic Pezizomycotina(Index Fungorum 2015) with Stachylidium bicolor Link, analkalophilic fungus as the type species. Réblová et al. (2011a)s h owed t h a t t h e t y p e s p e c i e s g r o u p s i n t h ePlectosphaerellaceae with high statistical support.
Staphylotrichum J. Mey. & Nicot, Bull. trimest. Soc. mycol.Fr. 72: 322 (1957) [1956]
A new species, Staphylotrichum boninense K. Nonaka et al.and the type species, S. coccosporum J. Mey. & Nicot, werefound to be related to Chaetomiaceae (Nonaka et al. 2012).
Sterigmatobotrys Oudem., Ned. kruidk. Archf, 2 sér. 4: 548(1886)
Réblová and Seifert (2011d) reported the sexual morph ofthe hyphomycete genus Sterigmatrobotrys. Fertile conidio-phores of S. macrocarpa (Corda) S. Hughes. formed on peri-thecial ascomata and an identical asexual morph developedfrom ascospores isolated in axenic culture. Phylogeneticrelationships based on LSU sequence data reveal thatSterigmatrobotrys macrocarpa clustered in a clade withCarpoligna pleurothecii F.A. Fernández & Huhndorf.Carpoligna and Chaetosphaeria are morphologically similar
Fungal Diversity

to Sterigmatobotrys. Réblová et al. (2012) indicated thatPleurotheciella is closely related to Pleurothecium andSterigmatobotrys, and can be placed in Hypocreomycetidaegenera incertae sedis. However, in the current studyCarpoligna pleurothecii and Pleurothecium semifecundumsit in the order Savoryellales.
Stromatonectria Jaklitsch & Voglmayr, Mycologia 103(2):435 (2011)
Jaklitsch and Voglmayr (2011) redescribed Myrmaeciellacaraganae Höhn., in the new genus Stromatonectria asS. caraganae (Höhn.) Jaklitsch & Voglm. based on morphol-ogy and phylogenetic analyses. It was recollected on branchesof Caragana spp., Colutea arborescens and Laburnumanagyroides (Fabaceae) around Vienna, Austria.Phylogenetic analyses of LSU also indicated this genus be-longs to a separate clade in Bionectriaceae.
Swampomyces Kohlm. & Volkm.-Kohlm., Botanica Marina30: 198 (1987)
Sakayaroj (2005) and Schoch et al. (2007) showed that genuswas polyphyletic separating into two clades: Swampomycessensu stricto (type S. armeniacus Kohlm. & Volkm.-Kohlm.groupingwith S. triseptatusK.D.Hyde&Nakagiri) in the familyEtheirophoraceae, Hypocreomycetideae order incertae sedis,while S. aegyptiacus Abdel-Wahab et al. and S. clavatisporaAbdel-Wahab et al. group in the Juncigenaceae for which anew genus Fulvocentrum has been introduced to accommodatethese taxa (Jones et al. 2014).
Synaptospora Cain, Beih. Sydowia 1: 4 (1957) [1956]One of four named species of Synaptospora ,
S. plumbea Huhndorf et al. is represented by gene se-q u e n c e s w h i c h p l a c e t h e g e n u s i nHe lm in t ho sphae r i ac eae (M i l l e r e t a l . 2014 ) .Synaptospora olandica Réblová has been associated witha dactylaria-like asexual morph (Réblová 2002), butthere is no molecular data to prove this connection.
Thalassogena Kohlm. & Volkm.-Kohlm. , Sys t .Ascomycetum 6: 223 (1987)
Monotypic marine ascomycete genus growing on sub-merged wood, with its placement in the Halosphaeriaceae,but confirmation at the molecular level is required. No asexualmorph is known.
ThalesporaChatmala&E.B.G. Jones, NovaHedwigia 83 (1–2):228 (2006)
Monotypic marine ascomycete genus growing on sub-merged wood, with its placement in the Halosphaeriaceaeconfirmed at the molecular level, with Okeanomyces as a sis-ter group (Jones et al. 2006; Sakayaroj et al. 2011a).Thalespora differs from Okeanomyces in having polar
tetraradiate appendages formed after release from theascomata. No asexual morph is known.
Thyronectria Sacc., Grevillea 4(no. 29): 21 (1875)Based on type studies and freshly collected material,
Jaklitsch and Voglmayr (2014) re-instated the genusThyronectria. Molecular phylogenies based on ACT,ITS, LSU rDNA, RPB1, RPB2, TEF, and TUB genes equenc e s a l s o con f i rmed t h e p l a c emen t o fThyronectria in family Nectriaceae (Hypocreales)(Jaklitsch and Voglmayr 2014)
Thelonectria P. Chaverri & C. Salgado, in Chaverri et al.,Stud. Mycol. 68(1): 76 (2011)
This genus was introduced by Chaverri et al. (2011) withThelonectria discophora (Mont.) P. Chaverri & C. Salgado asthe generic type. The genus clusters in Nectriaceae and hascylindrocarpon-like asexual morphs. The genus presentlycomprises 22 species (Chaverri et al. 2011, Salgado-Salazaret al. 2012, Luo and Zhang 2013).
Tilachlidiaceae L. Lombard & Crous, in Lombard et al.,Studies in Mycology 80: 237 (2015)
The family Tilachlidiaceae was introduced based onmulti-gene analysis using combined gene datasets andplaced in the order Hypocreales (Lombard et al.2015). The family comprises two asexual generaSeptofusidium and Tilachlidium, previously classified inthe family Nectriaceae. Phylogenetic analysis showedthat representatives of both genera clustered together ina well-supported clade leading to introduction of thenew family Tilachlidiaceae (Lombard et al. 2015).
Tilachlidium Preuss, Linnaea 24: 126 (1851)The genus Tilachlidium typified by T. pinnatum Preuss was
classified in Hypocreales genus incertae sedis (Gams 1971).This genus was placed in the family Tilachlidiaceae byLombard et al. (2015) based on molecular analysis.
Tirispora E.B.G. Jones & Vrijmoed, Can J. Bot. 72 (9): 1373(1994)
Genus in the Halosphaeriaceae supported by morpholog-ical and molecular data, forming a sister group withHalosarpheia and Panorbis (Jones et al. 2009a, b). It sharessome common features with these genera, in particularly thepolar unfurling appendages (Jones et al. 2009b). No asexualmorph is known.
Togniniella Réblová et al., Stud. Mycol. 50(2): 543 (2004)Réblová et al. (2004) linked and established twomonotypic
genera: Togniniella acerosa Réblová et al. and Phaeocrellaacerosa Réblová et al. using cultural studies. However,T. acerosa is now regarded as a synonym of T. microspora
Fungal Diversity

(Ellis & Everh.) Réblová (Réblová 2011) and Phaeocrella is asynonym. The genus is placed in Calosphaeriaceae.
Tolypocladium W. Gams, Persoonia 6(2): 185 (1971)This genus was introduced by Gams (1971). Based on se-
quence analysis, Quandt et al. (2014), proposed to useTolypocladium over Elaphocordyceps and Chaunopycnis.
Toriella Sakay. et al., Fungal Diversity 46: 99 (2011)Sakayaroj et al. (2011a) carried out a multi-gene phylog-
eny of the Halosphaeriaceae with 36 taxa. Ceriosporopsiswas shown to be polyphyletic and Ceriosporopsis tubuliferaKohlm. did not cluster with the type species of the genus(C. halima Linder). The genus Toriella was introduced toaccommodate C. tubulifera, as Toriella tubulifera (Kohlm.)Sakay. et al. Earlier studies noted that morphologically theascospores did not conform to those described for the typespecies, where the polar appendages pierce through anexosporic sheath (Johnson et al. 1984). No asexual morphis known.
Torpedospora Meyers, Mycologia 49: 496 (1957)The genus has been referred to various higher order place-
ment: Hypocreales incertae sedis (Jones et al. 2009b), a sistergroup to the Bionectriaceae (Sakayaroj et al. 2005) and TBMclade (Schoch et al. 2007). Torpedospora species form a well-supported monophyletic clade in Torpedosporaceae, withGlomerulispora mangrovis (Jones et al. 2014).
Torpedosporaceae E.B.G. Jones & K.L. Pang, CryptogamieMycologie 35: 135 (2014)
The family was introduced to accommodate twoTorpedospora species and Glomerulispora mangrovisAbdel-Wahab & Nagah. that formed a highly supported cladein Hypocreomycetidae family incertae sedis. Asexual morphsare helicoid conidia when present.
Tortulomyces Lar.N. Vassiljeva et al., Mycoscience 54(1):110–115 (2013)
This monotypic genus was introduced for Tortulomycesthailandicus Lar.N. Vassiljeva et al., collected on deadbranches of an unidentified tree in Thailand. Tortulomyces issimilar in appearance to Spinulosphaeria, but a quellkörperand setae on the ascomata are absent. The genus is placed inNitschkiaceae.
Trailia G.K. Sutherl., Trans. Br. mycol. Soc. 5(1): 149 (1915)[1914]
Monotypic genus in the Halosphaeriaceae (Jones et al.2009b) growing on the brown seaweed Ascophyllumnodosum, infrequently collected and its placement in thefamily requires a molecular study. No asexual morph isknown.
Trichocladium Harz, Bull. Soc. Imp. nat. Moscou 44(1): 125(1871)
A polyphyletic genus with some species grouping in theHypocreomycetidae: Trichocladium acharosporum (Meyers& R.T. Moore) M. Dixon was shown by culturing to be theasexual morph of Halosphaeriopsis mediosetigera(Halosphaeriaceae) (Shearer and Crane 1977), while at themolecular level T. melhae E.B.G. Jones et al. also groupedin the Halosphaeriaceae (Abdel-Wahab 2012) withAntennospora salina (Meyers) Yusoff, in a sister clade.Some species have been linked to sexual morphs ofChaetomium. The generic type of Trichocladium ,T. asperum Harz (strain DAOM 232342) was shown to groupin Ascomycota genera incertae sedis by Hambleton et al.(2005) so the name should be retained.
Trichoderma Pers., Neues Mag. Bot. 1: 92 (1794)The type species of the genus Hypocrea (Hypocreaceae),
H. rufa (Pers.) Fr., was re-defined and epitypified using acombination of phenotype and phylogenetic analyses. Itsasexual morph Trichoderma viride Pers., the type species ofTrichoderma, was re-described and epitypified (Jaklitsch et al.2006). Samuels et al. (2012) described the new Trichodermaspecies, T. gillesii Samuels and T. pinnatum Samuels withHypocrea sexual morphs (with culture and molecular data).These genera are clearly linked and Rossman et al. (2013)proposed the use of Trichoderma over Hypocrea, becauseTrichoderma has high usage in the literature and fewHypocrea species have been reported more than once. Wefollow this here.
Trichomaris Hibbits et al., Can. J. Bot. 59(11): 2123 (1981)Monotypic genus requiring further study at the molecular
level and tentatively assigned toHalosphaeriaceae. The asex-ual morph is a coelomycete.
Tubakiella Sakay. et al. Fungal Diversity 46:87–109 (2011)A genus well-supported by molecular data in the
Halosphaeriaceae (Sakayaroj et al. 2011a), with T. galerita(Tubaki) Sakay. et al. (= Remispora galerita Tubaki as the typespecies. A genus segregated from Remispora sensu stricto, anddistantly placedwithin the family, with the two sequenced strainsforming amonophyletic group withNautosphaeria cristaminutaand Haligena elaterophora as a sister group with weak support(Sakayaroj et al. 2011a). No asexual morph is known.
Tunicatispora K.D. Hyde, Aust. Syst. Bot. 3(4): 712 (1990)Monotypic genus referred to the Halosphaeriaceae based
on morphological observations, and requires further study atthe molecular level to confirm its position in the family. Noasexual morph is known. Buxetroldia is a synonym ofTunicatispora based on morphological observations (Joneset al. 2009b).
Fungal Diversity

Tympanopsis Starbäck, Bih. K. svenska VetenskAkad.Handl., Afd. 3 19(no. 3): 24 (1894)
See under Scortechiniaceae.
TyrannicordycepsKepler & Spatafora, Index Fungorum 12: 1(2012)
This genus has five species (Index Fungorum 2015) whichlive on sclerotia of Claviceps. Molecular analysis placed thisgenus inClavicipitaceae in the Plant-Hemiptera clade (Kepleret al. 2012).
Ustilaginoidea Bref., Unters. Gesammtgeb. Mykol. (Liepzig)12: 194 (1895)
Villosiclava was introrcued as the asexual morph ofUstilaginoidea and is thus a synonym (Tanaka et al. 2008).The genus is placed in Clavicipitaceae.
Valsalnicola D.M. Walker & Rossman, Persoonia, Mol. Phyl.Evol. Fungi 29: 149 (2012)
This genus was introduced in Crous et al. (2012f) forValsalnicola oxystoma (Rehm) D.M. Walker & Rossman,and causes linear cankers and lesions on Alnus. Although itresembles Valsa in having allantoid ascospores, the species inValsalnicola are 1-septate, while the majority of Valsa specieshave aseptate spores. Mega blast similarity of ITS sequencewith Genbank place Valsalnicola oxystoma in close affinitywith genera of Gnomoniaceae, Melanconidaceae andValsaceae. However there is no clear phylogenetic familyplacement within Diaporthales and hence it is placed inDiaporthales, genera incertae sedis
VaricosporinaMeyers & Kohlm., Can. J. Bot. 43: 916 (1965)Three Varicosporina species have been shown to be the
asexual morphs of various Corollospora species: C. anglusaAbdel-Wahab & Nagah. (= V. anglusa Abdel-Wahab &Nagah.), C. intermedia E.B.G. Jones (= V. proliferaNakagiri) and Corollospora sp. (= V. ramulosa Meyers &Kohlm.) (Abdel-Wahab 2012). Since the type species,Varicosporina ramulosa Meyers & Kohlm. has not beenlinked to a specific Corollospora species, thus the nameVaricosporina should be retained and the genus is tentativelyplaced in Halosphaeriaceae.
Vermisporium H.J. Swart & M.A. Will., Trans. Br. mycol.Soc. 81(3): 491 (1983)
An asexual genus introduced by Swart and Williamson(1983) for leaf-inhabiting coelomycetous taxa on Eucalyptusleaves, currently with some 13 species (Index Fungorum2015). Vermisporium was chiefly distinguished fromSeimatosporium on the basis of a short exogenous basalappendage, and the absence of a recognisable apicalappendage. Barber et al. (2011) showed the genus to be asynonym of Seimatosporium based on DNA sequence data
analysis of the nrDNA-ITS and 28S nrRNA genes, and trans-ferred eight species to Seimatosporium.
Verrucostoma Hirooka et al., in Hirooka et al., Mycologia102(2): 422 (2010)
Hirooka et al. (2010) described this monotypic genus basedon Verrucostoma freycinetiaeHirooka et al. using morpholog-ical differences and molecular analyses. The taxon was col-lected on dead leaves of Freycinet ia boninensis(Pandanaceae) in Bonin (Ogasawara) Islands, Japan.Molecular analyses indicated that the genus is distinct fromother genera among the nec t r ia- l ike taxa underBionectriaceae.
Verticillium Nees, System der Pilze und Schwämme: 56(1817)
The asexual genus Verticillium comprises a group of plantpathogenic fungi including some 190 documented names andwith V. dahliae Kleb. as the type species. The genus has beenshown to be polyphyletic with various taxa assigned to newgenera: Gibellulopsis, Lectera and Musicillium. Most speciesgroup in the Plectosphaerellaceae based on sequence analysis(Inderbitzin et al. 2011). No sexual morph is known. Hydeet al. (2014) provided a backbone tree for Verticillium species.
Vialaea Sacc., Bull. Soc. mycol. Fr. 12: 66 (1896)A genus introduced by Saccardo (1896) and currently with
five species (Index Fungorum 2015). Senanayake et al. (2014)introduced a new species of Vialaea, V. mangiferae I.C.Senanayake & K.D. Hyde from mango and provide phyloge-netic evidence to support the introduction of this new species.
Vialaeaceae P.F. Cannon Mycological Research 99: 368(1995)
A family introduced by Cannon (1995) for species withisthmoid ascospores. Shoemaker et al. (2013), McTaggartet al. (2013) and Senanayake et al. (2014) provided new dataand phylogenetic evidence to show that this is a well-supported family in Xylariales. The grouped ascomata witha single fused neck, J+, ascal apical ring and unusual asco-spores with two fusiform parts joined by a narrow, long isth-mus are characteristic of the family. The asexual morph iscoelomycetous (Senanayake et al. 2014).
VirgariaNees, Syst. Pilze (Würzburg): 54 (1816) [1816–17]Virgaria nigra (Link) Nees, the type species of the genus is
a commonly encountered hyphomycete. Ascovirgaria was in-troduced to accommodate stromatic Xylariaceae with rudi-mentary blackened stromata in decayed wood. The type spe-cies Ascovirgaria occulta J.D. Rogers & Y.M. Ju was isolatedfrom Hawaii, and produced the asexual morph, Virgaria nigrain culture (Rogers and Ju 2002). According to Rogers and Ju(2002), Virgaria nigra is cosmopolitan and was long
Fungal Diversity

recognized before its respective sexual morph and commonlygrows in the absence of its sexual morph. Therefore, the ear-lier described name Virgaria is used and the sexual morphAscovirgaria becomes a synonym (Rogers and Ju 2002;Stadler et al. 2013).
Volutellonectria J. Luo &W.Y. Zhuang, Phytotaxa 44: 3 (2012)This genus with three new species was introduced in Luo
and Zhuang (2012) with the type based on Dialonectriaconsors Ellis & Everh. (= Cosmospora consors (Ellis &Everh.) Rossman & Samuels). Species have volutella-likeasexual morphs and cosmospora-like sexual morphs, but phy-logenetic analyses of ITS and 28S sequence data showedVolutellonectria and Cosmospora to be distinct genera inNectriaceae.
Woswasia Jaklitsch et al., Mycologia 105(2): 479 (2013)This monotypic genus is represented by Woswasia
atropurpurea Jaklitsch et al. which is a wood-inhabiting andmycotrophic taxon (Jaklitsch et al. 2013). Analysis of sequencedata showed the genus to cluster in Sordariomycetidae close tothe genus Ceratolenta, but could not be placed in any family.
Xenoacremonium Lombard & Crous, in Lombard et al.,Studies in Mycology 80: 234 (2015)
This genus is established by Lombard et al. (2015)based on phylogenetic studies of combined sequencedata to include a group of fungi previously treated asAcremonium reci fe i (Leão & Lôbo) W. Gams.Xenoacremonium species can be distinguish fromParacremonium by a phenomenon of readily releasinga pale luteous to luteous pigment into the growth me-dium and not forming sterile coils in culture. The genusXenoacremonium is typified by X. recifei (Leão &Lôbo) L. Lombard & Crous and clusters in Nectriaceae.
Xenopyricularia Klaubauf et al., Stud. Mycol. 79: 117 (2014)A monotypic genus of Pyriculariaceae that was introduced
by Klaubauf et al. (2014) to accommodate the type speciesXenopyricularia zizaniicola Klaubauf et al., which was isolatedfrom Zizania latifolia in Japan (Klaubauf et al. 2014).Phylogenetic analysis based on a combined dataset LSU, ITS,RPB1, ACT and CAL sequences revealed Xenopyricularia torepresent a distinct genus in Pyriculariaceae (Klaubauf et al.2014).
Xylaria Hill ex Schrank, Baier. Fl. (München) 1: 200 (1789)Padixonia was introduced by Subramanian (1972) to ac-
commodate Padixonia bispora Subram. from Ghana. The au-thor mentioned this hyphomycete as the conidial morph ofXylaria furcata Fr. (Subramanian 1972). Since the older namehas priority, Xylaria should be retained (Stadler et al. 2013).Another asexual genus, Xylocoremium was introduced by
Rogers (1984) to accommodate the coremial conidial morphof Xylaria species. Xylaria cubensis (Mont.) Fr. was identifiedas the corresponding sexual morph of Xylocoremiumflabelliforme (Schwein.) J.D. Rogers (Rogers 1984). Xylariaallantoidea (Berk.) Fr. and X. castorea Berk. have also beencited as connected with Xylocoremium flabelliforme, but notas synonyms. Stadler et al. (2013) suggested priority can begiven to the older name and the asexual name can be sup-pressed. Arthroxylaria was introduced by Seifert et al.(2002), and typified by A. elegans Seifert & W. Gams. Thisxylarioid asexual morph has been connected to Xylaria(Stadler et al. 2013). The asexual name has the possibility tobe suppressed in favour of Xylaria as it is the older name andthe cosmopolitan morph frequently encountered in nature(S tad le r e t a l . 2013) . Möl le r (1901) desc r ibedMoelleroclavus and believed the fungus to be Xylaria.Hennings (1902) introduced the genus Moelleroclavus anddesignated the type species as M. penicilliopsis Henn. and aspecimen from the same locality asMöller’s material, which islisted as the type. Rogers et al. (1997) collected Xylariamoelleroclavus J.D. Rogers et al. from Hawaii, which hasthe asexual morph which is the same as Möller’s descriptionin 1901. Xylaria moelleroclavus was identified as the respec-tive sexual morph because they obtained the asexual morph inculture derived from ascospores (Rogers et al. 1997). Theauthors however believed the name Moelleroclavuspenicilliopsis Henn. should remain available for the asexualmorph of X. moelleroclavus. Stadler et al. (2013) proposedthese names can be either synonymised or retained if the largegenus Xylariawill be split into further groups to accommodatethe certainly diverse Xylaria.
Zalerion R.T. Moore & Meyers, Can. J. Microbiol. 8: 408(1962)
Moore and Meyers (1962) established the genus Zalerionbased on helicoid, subhyaline to fuscous conidia, which arecoiled irregularly in three dimensions and constricted at thesepta. Although they included the two helicoid speciesdescribed by Barghoorn and Linder (1944b) under Helicoma,they did not select one of these as the type species. InsteadZalerion nepura R.T. Moore & Meyers, was selected as thetype species. They described four species based on morpholo-gy, conidial pigmentation and their cellulolytic ability (reducingsugar determinations of Raycord). Herbarium material was de-posited at the Farlow Herbarium (FH), but no cultures weredeposited as far as can be ascertained. Subsequently,Anastasiou (1963) reduced the two marine Helicoma speciesand four Zalerion species under Zalerion maritima (Linder)Anastasiou, and described a second species Z. variaAnastasiou (= Halenospora varia (Anastasiou) E.B.G. Jones).Molecular data generated by Campbell et al. (2005) showedthat Z. maritima grouped with Lulwoana uniseptata(Nakagiri) Kohlm. et al., with 100 % support, with
Fungal Diversity

Moromyces varius (Chatmala & Somrith.) Abdel-Wahab et al.(= Cumulospora varia Chatmala & Somrith.) as a sister group(Abdel-Wahab et al. 2010). This link has also been shown bycultural studies. The name Zalarion should be retained as thereis no molecular data for the type species and the genus is ten-tatively placed in Sordariomycetes genera incertae sedis.
Zopfiella G. Winter, Rabenh. Krypt.-Fl., Edn 2 (Leipzig) 1.2:56 (1884)
A polyphyletic genus with the type species, Zopfiellatabulata (Zopf) G. Winter placed in the familyChaetomiaceae (Cai et al. 2006b). The genus sensu lato iswidely distributed with some 27 records listed in IndexFungorum (2015). Guarro et al. (1999) have reported an asex-ual morph with humicola-like conidia produced on a peg-likestructure on undifferentiated hyphae for marine collections ofZ. latipes (N. Lundq.)Malloch&Cain and Z. marina Furuya&Udagawa.
Zygospermella Cain, Mycologia 27 (2): 227 (1935)Genus in the Lasiosphaeriaceae with Zygospermella
setosa Cain as the type species Three strains of Z. insignis(Mouton) Cain form a monophyletic clade grouping withmoderate support with Cercophora sordarioides based on amolecular study of LSU nrDNA and β-tubulin sequences(Kruys et al. 2015).
Genera of uncertain placement in Sordariomycetes
In this part we list 829 genera that are classified inSordariomycetes in Index Fungorum (2014) as on 31December 2014. These genera were not listed inSordariomycetes in Lumbsch and Huhndorf (2010) and havenot been listed in subsequent publications as belonging inSordariomycetes. The data was extracted from a database pro-vided by Paul Kirk. These genera may need further study.
Abaphospora Kirschst.
Acanthorhynchus Shear
Acanthotheca Clem. & Shear
Acanthothecium Speg.
Acaulium Sopp
Achaetomiella Arx
Acontiopsis Negru
Acrosphaeria Corda
Acrostaphylus G. Arnaud ex Subram.
Acrostaphylus G. Arnaud
Actinodothis Syd. & P. Syd.
Actinonemella Höhn.
Actinopelte Sacc.
Adea Petr.
Ahmadinula Petr.
Akrophyton Lebert
Albocrustum Lloyd
Aleurisma Link
AllantosporaWakker
Allantozythiella Danilova
Allelochaeta Petr.
Allescherina Berl.
Alphitomyces Reissek
Amazoniella Bat. & H. Maia
AmphichaetaMcAlpine
Amphitiarospora Agnihothr.
Amphorula Grove
Ampullaria A.L. Sm.
Andreaea Palm & Jochems
Andreaeana Palm & Jochems
Andreanszkya Tóth
Anisochora Theiss. & Syd.
Anisostomula Höhn.
Anixiella Saito & Minoura
Anixiella Saito & Minoura ex Cain
Antenaglium F.C. Albuq.
Anthasthoopa Subram. & K. Ramakr.
Apiocrea Syd. & P. Syd.
Apiosporella Höhn. ex Theiss.
Apiothecium Lar.N. Vassiljeva
Aplacodina Ruhland
Aponectria (Sacc.) Sacc.
Aporhytisma Höhn.
Aposphaeriella Died.
Aposphaeriopsis Died.
Apotemnoum Corda
Ariefia Jacz.
Arnoldia D.J. Gray & Morgan-Jones
Arnoldiomyces Morgan-Jones
Artallendea Bat. & H. Maia
Aschersoniopsis Henn.
Ascocollumdensa K.D. Hyde et al.
Ascostroma Bonord.
Asordaria Arx, Guarro & Aa
Asteridium (Sacc.) Speg. ex Sacc.
AsterotheciumWallr.
Bacillispora Sv. Nilsson
Bagnisiopsis Theiss. & Syd.
Bahupaathra Subram. & Lodha
Bakeromyces Syd. & P. Syd.
Balansiella Henn.
Balansiopsis Höhn.
Bartaliniopsis S.S. Singh
Barya Fuckel
Baryeidamia H. Karst.
Baryella Rauschert
Fungal Diversity

Basifimbria Subram. & Lodha
Basipilus Subram.
BasisporiumMolliard
Basitorula G. Arnaud
Batschiella Kirschst.
Beejasamuha Subram. & Chandrash.
Belaina Bat. & Peres
Benedekiella Negru & Verona
Berkeleyna Kuntze
Bidenticula Deighton
Bioporthe Petr.
Biotyle Syd.
Bisporomyces J.F.H. Beyma
Bitrimonospora Sivan. et al.
Bivonella (Sacc.) Sacc.
Bizzozeria Speg.
Bizzozeria Sacc. & Berl.
Blennorella Kirschst.
Bolacotricha Berk. & Broome
Bolinia (Nitschke) Sacc.
Bonordenia Schulzer
Bostrychia Fr.
Botryocrea Petr.
Botryodiplis Clem. & Shear
Botryodiplodia (Sacc.) Sacc.
Botryosphaerostroma Petr.
Botrypes Preuss
Bovilla Sacc.
Boydia A.L. Sm.
Brachydesmium (Sacc.) Costantin
Bresadolella Höhn.
Bryonectria Döbbeler
Byssitheca Bonord.
Byssostilbe Petch
Cacosphaeria Speg.
Cainea S. Hughes
Calopactis Syd. & P. Syd.
Calospora Nitschke ex Fuckel
Calospora Sacc.
Calyculosphaeria Fitzp.
Camptoum Link
Campylothecium Ces.
Candelospora Rea & Hawley
Cantharosphaeria Thaxt.
Capnostysanus Speg.
Carnostroma Lloyd
Carothecis Clem.
Castagnella G. Arnaud
Catacauma Theiss. & Syd.
Cateractispora Ranghoo et al.
Caudosporella Höhn.
Caulochora Petr.
Causalis Theiss.
Cephalodiplosporium Kamyschko
Cephalosporiopsis Peyronel
Cephalosporium Corda
Cephalothecium Corda
Cepsiclava J. Walker
Cerastomis Clem.
Ceratocladium Pat.
Ceratopodium Corda
Ceratostoma Pers.
Ceratostoma Fr.
Cercosporula G. Arnaud
Cerillum Clem.
Ceuthocarpon P. Karst.
Chaetapiospora Petr.
Chaetoceratostoma Turconi & Maffei
Chaetoceris Clem. & Shear
Chaetodimerina Hansf.
ChaetolentomitaMaubl.
Chaetomelasmia Danilova
Chaetomeliola (Cif.) Bat. et al.
Chaetomiotricha Peyronel
Chaetonaemosphaera Schwarzman
Chailletia Jacz.
Chailletia Fuckel
Cheilariopsis Petr.
Cheiropodium Syd. & P. Syd.
Chiajaea (Sacc.) Höhn.
Chiloella Syd.
Chilonectria Sacc.
Chitinonectria M. Morelet
Chlorospora Speg.
Chondroplea Kleb.
Chorostate (Nitschke ex Sacc.) Traverso
Chorostella (Sacc.) Clem. & Shear
Chromocrea Seaver
Chromocreopsis Seaver
Chromocytospora Speg.
Chromostylium Giard
Chrysogluten Briosi & Farneti
Ciliciopodium Corda
Ciliciopus Clem. & Shear
Ciliofusa Clem. & Shear
Ciliofusarium Rostr.
Ciliomyces Höhn.
Circinaria Pers.
Circinaria Bonord.
Circinostoma Gray
Cryptoderis Auersw.
Cryptosordaria De Not. ex Sacc.
Cryptosphaerina Lambotte & Fautrey ex Sacc. & P. Syd.
Cryptospora Tul. & C. Tul.
Fungal Diversity

Cryptostictella Grove
Cryptostictis Fuckel
CryptothamniumWallr.
Cryptothecium Penz. & Sacc.
Cucurbitariella Petr.
Cucurbitula Fuckel
Curreyella (Sacc.) Lindau
Cyclocytospora Höhn.
Cyclodomella P.N. Mathur et al.
Cyclophomopsis Höhn.
Cylicogone Emden & Veenb.-Rijks
Cylindrium Bonord.
Cylindrocladiopsis J.M. Yen
Cyphellopycnis Tehon & G.L. Stout
Cytodiplospora Oudem.
Cytophoma Höhn.
Cytosporopsis Höhn.
Dactylina G. Arnaud
Daedala Hazsl.
Dapsilosporium Corda
Dasyphthora Clem.
Debarya Schulzer
Debaryella Höhn.
Dendrodochium Bonord.
Desmotrichum Lév.
Detonina Kuntze
Dialytes Nitschke
Diatrypeopsis Speg.
Dichitonium Berk. & M.A. Curtis
Dichotomella Sacc.
Dicladium Ces.
Didymariopsis Speg.
Didymocladium Sacc.
Didymopsamma Petr.
Diheterospora Kamyschko
Diheterospora Kamyschko ex G.L. Barron & Onions
Dimera Fr.
Diploceras (Sacc.) Died.
Diplocladium Bonord.
Diploplenodomopsis Petr.
Diplosclerophoma Petr.
Diplosporis Clem.
Diplosporium Link
Disaeta Bonar
Discodiaporthe Petr.
Discomycopsella Henn.
Discosiopsis Edward, Kr.P. Singh et al.
Discosiospora A.W. Ramaley
Discosphaera Dumort.
Discostromopsis H.J. Swart
Disperma Theiss.
Dochmolopha Cooke
Dothideovalsa Speg.
Dothidina Theiss. & Syd.
Dozya P. Karst.
Dryadomyces Gebhardt
Dubiomyces Lloyd
Echinodothis G.F. Atk.
Echinopodospora B.M. Robison
Echinospora Mirza
Echusias Hazsl.
Ectomyces P. Tate
Ectosphaeria Speg.
Edmundmasonia Subram.
Eidamia Lindau
Ellisiella Sacc.
Ellisiellina Sousa da Câmara
Ellisiopsis Bat.
Embolidium Bat.
Endocreas Samuels & Rogerson
Endodothella Theiss. & Syd.
Endogloea Höhn.
Endophyllachora Rehm
Endotrabutia Chardón
Engizostoma Gray
Entosordaria Speg.
Entosordaria (Sacc.) Höhn.
Eolichen Zukal
Eolichenomyces Cif. & Tomas.
Ephedrosphaera Dumort.
Epheliopsis Henn.
Ephemeroascus Emden
Epinectria Syd. & P. Syd.
Ergotaetia E.J. Quekett
Eriomene (Sacc.) Clem. & Shear
Eriomenella Peyronel
Erionema Maire
Eriosphaerella Höhn.
Erostella (Sacc.) Sacc.
Erostrotheca G.H. Martin & Charles
Erythrosphaera SorokīnEsfandiaria Petr.
Euhypoxylon Füisting
Euricoa Bat. & H. Maia
Europhium A.K. Parker
Eusordaria Zopf
Eutypopsis P. Karst.
Exomassarinula Teng
Fabreola Kuntze
Fairmania Sacc.
Fellneria Fuckel
Fimetaria D.A. Griffiths & Seaver
Fioriella Sacc.
Fitzpatrickia Cif.
Fungal Diversity

Flagellospora Ingold
Flageoletia (Sacc.) Höhn.
Fleischeria Penz. & Sacc.
Fominia Girz.
Fragosoella Petr. & Syd.
Frankia Brunch.
Frankiella Speschnew
Frondisphaeria K.D. Hyde
Fuckelia (Nitschke ex Sacc.) Cooke
Fuckelina Sacc.
Fusicytospora Gutner
Fusidium Link
Fusidomus Grove
Fusisporium Link
Fusisporium Fr.
Gaeumannia Petr.
Galeraicta Preuss
Gamosphaera Dumort.
Gerlachia W. Gams & E. Müll.
Germslitospora Lodha
Gibberellulina Sousa da Câmara
Gibsonia Massee
Gliobotrys Höhn.
Gloeocercospora D.C. Bain & Edgerton ex Deighton
Gloeocercospora D.C. Bain & Edgerton
Gloeosporidium Höhn.
Gloeosporina Höhn.
Gloeosporiopsis Speg.
Glomerularia H. Karst.
Gnomonina Höhn.
Godroniella P. Karst.
Gonatosporium Corda
Gonatotrichum Corda
Gongromeriza Preuss
Gongylocladium Wallr.
Goniosporium Link
Gonytrichella Emoto & Tubaki
Graphilbum H.P. Upadhyay & W.B. Kendr.
Graphiocladiella H.P. Upadhyay
Griphosphaerella Petr.
Griphosphaeria Höhn.
Guttularia W. Oberm.
Halstedia F. Stevens
Hansenia Zopf
Haplochalara Linder
Haplomela Syd.
Haplophoma Riedl & Ershad
Haplostroma Syd. & P. Syd.
Haplothecium Theiss. & Syd.
Haplotrichella G. Arnaud
HartiellaMassee
Harziella Kuntze
Heimiodiplodia Zambett.
Helicosporangium H. Karst.
Hemisphaeria Klotzsch
HendersoniopsisWoron.
Henningsina Möller
Herminia R. Hilber
Heteronectria Penz. & Sacc.
Heteropera Theiss.
Hindersonia Moug. & Nestl. ex J. Schröt.
Hiogispora Abdel-Wahab & Nagah.
Hormosperma Penz. & Sacc.
Hormospora De Not.
Hyalobotrys Pidopl.
Hyaloceras Durieu & Mont.
Hyaloflorea Bat. & H. Maia
Hyalomelanconis Naumov
Hyalopesotum H.P. Upadhyay & W.B. Kendr.
Hyalopus Corda
Hyalostachybotrys Sriniv.
Hyalotia Guba
Hymenopleella Munk
Hymenopodium Corda
Hyphelia Fr.
Hyphoderma Fr.
Hyphonectria (Sacc.) Petch
Hypocreophis Speg.
Hypodiscus Lloyd
Hypoplasta Preuss
Hypopteris Berk.
Hypostegium Theiss.
Hypoxylina Starbäck
Hypoxylum Juss.
Hysterodothis Höhn.
Hystrix Alstrup & Olech
Indiella Brumpt
Innatospora J.F.H. Beyma
Insecticola Mains
Institale Fr.
Irene Theiss., Syd. & P. Syd.
Irenina F. Stevens
Ixodopsis P. Karst.
Jacobaschella Kuntze
Jongiella M. Morelet
Kaskaskia Born & J.L. Crane
Keissleria Höhn.
Kellermanniopsis Edward et al.
KentrosporiumWallr.
Kirschsteinia Syd. & P. Syd.
Knyaria Kuntze
Kommamyce Nieuwl.
Kubinyia Schulzer
Laaseomyces Ruhland
Fungal Diversity

Lachnidium Giard
Laestadia Auersw.
Lageniformia Plunkett
Lamyella Berl.
Lamyella Fr.
Larseniella Munk
Lasiella Quél.
Lasiosordaria Chenant.
Lasiosordariella Chenant.
Lasiosordariopsis Chenant.
Lasiostroma Griffon & Maubl.
Leiosepium Sacc.
Leiostigma Kirschst.
Lennisia Nieuwl.
Lentomita Niessl
Lepidonectria Speg.
Lepra Willd.
Lepteutypella Petr.
Leptina Bat. & Peres
Leptocoryneum Petr.
Leptocrea Syd. & P. Syd.
Leptodiscus Gerd.
Leptosporium (Sacc.) Höhn.
Leucocrea Sacc. & P. Syd. ex Lindau
Leucocytospora (Höhn.) Höhn.
Leucophomopsis Höhn.
Leucosphaera Arx, Mukerji & N. Singh
Leveillea Fr.
Leveillinopsis F. Stevens
Libertina Höhn.
Lichenagaricus P. Micheli
Lichenoverruculina Etayo
Ligniella Naumov
Lilliputia Boud. & Pat.
Linearistroma Höhn.
Linostoma Höhn.
Linostomella Petr.
Lisea Sacc.
Lisiella (Cooke & Massee) Sacc.
Lithomyces Viala & Marsais
Litschaueria Petr.
Lohwagiella Petr.
Lomentospora Hennebert & B.G. Desai
Longoa Curzi
Lophodiscella Tehon
Luxuriomyces R.F. Castañeda
Lysipenicillium Bref.
Macrothelia M. Choisy
Magnusia Sacc.
Malacosphaeria Syd.
Malacostroma Höhn.
Malinvernia Rabenh.
Masonia G. Sm.
Masoniella G. Sm.
Massalongiella Speg.
Massariella Speg.
Massariopsis Niessl
MastigocladiumMatr.
Mastigonetron Kleb.
Maurinia Niessl
Megalonectria Speg.
Megathecium Link
Melanconidium (Sacc.) Kuntze
Melanogone Wollenw. & H. Richt.
Melanopelta Kirschst.
Melanoporthe Wehm.
Melanopsammina Höhn.
Melanosporopsis Naumov
Meliolaster Doidge
Menisporella Agnihothr.
Meringosphaeria Peyronel
Merrilliopeltis Henn.
Mesobotrys Sacc.
Metachora Syd., P. Syd. & E.J. Butler
Microbasidium Bubák & Ranoj.
Micronectriopsis Höhn.
Microphiodothis Speg.
Microstoma Auersw.
Microthecium Corda
Microtypha Speg.
Milowia Massee
Mitrasphaera Dumort.
Miyoshia Kawam.
Moelleria Bres.
Moeszia Bubák
Monoceras Guba
Monochaetina Subram.
Monoconidia Roze
Monopycnis Naumov
Monosporella S. Hughes
Monostachys G. Arnaud
Montemartinia Curzi
Morrisiella Saikia & A.K. Sarbhoy
Mothesia Oddo & Tonolo
Mucrosporium Preuss
Munkiodothis Theiss. & Syd.
Murogenella Goos & E.F. Morris
Mycobanche Pers.
Mycophaga F. Stevens
Mycophycophila Cribb & J.W. Cribb
Mycorhynchella Höhn.
Mycothyridium E. Müll.
Myriogenis Clem. & Shear
Myrmaecium Sacc.
Fungal Diversity

Myrotheciella Speg.
Myxolibertella Höhn.
Myxormia Berk. & Broome
Myxothecium Kunze
NaemasporaWilld.
Neesiella Kirschst.
Nematomyces Faurel & Schotter
Neoarcangelia Berl.
Neohenningsia Koord.
Neokeissleria Petr.
Neoskofitzia Schulzer
Neozimmermannia Koord.
Nephrospora Loubière
NeuronectriaMunk
Nigrosphaeria N.L. Gardner
Notarisiella (Sacc.) Clem. & Shear
Nothopatella Sacc.
Nothopodospora Mirza
Nummularia Tul. & C. Tul.
Nummulariella Eckblad & Granmo
Nummularioidea (Cooke & Massee) Lloyd
Numulariola House
Ochraceospora Fiore
Ollula Lév.
Oostroma Bonord.
Ophiodothis Sacc.
Ophiostomella Petr.
Ophiotexis Theiss.
Ophiovalsa Petr.
Oplotheciopsis Bat. & Cif.
Oramasia Urries
Oswaldia Rangel
Pachnodium H.P. Upadhyay & W.B. Kendr.
Pachybasium Sacc.
Paidania Racib.
Papularia Fr.
ParacesatiellaPe tr.
Paradidymella Petr.
Paradiplodiella Zambett.
Paraeutypa Subram. & Ananthap.
Paragaeumannomyces Matsush.
Paralaestadia Sacc. ex Vain.
Paramazzantia Petr.
Paranthostomella Speg.
Parascorias J.M. Mend.
Parasteridiella H. Maia
Patouillardea Roum.
Peckiella (Sacc.) Sacc.
Pedisordaria Clem.
Peloronectriella Yoshim. Doi
Penzigia Sacc.
Penzigina Kuntze
Peresia H. Maia
Periaster Theiss. & Syd.
Peridoxylon Shear
Peripherostoma Gray
Perisphaeria Roussel
Perisporiella (Henn.) Clem. & Shear
Peristomium Lechmere
Peroneutypella Berl.
Perrotiella Naumov
Pestalozzina (Sacc.) Sacc.
Pestalozzina P. Karst. & Roum.
Petasodes Clem.
Petelotia Pat.
PetriellidiumMalloch
PeyronellulaMalan
Peziotrichum (Sacc.) Lindau
Phaeaspis Kirschst.
Phaeidium Clem.
Phaeoapiospora (Sacc. & P. Syd.) Theiss. & Syd.
Phaeoconis Clem.
Phaeodiaporthe Petr.
Phaeoharziella Loubière
Phaeonectria (Sacc.) Sacc. & Trotter
Phaeophomatospora Speg.
Phaeoscopulariopsis M. Ota
Phaeosperma Nitschke ex Fuckel
Phaeostagonosporopsis Woron.
Phaeostilbella Höhn.
Phaeostoma Arx & E. Müll.
Phaeotrabutia Orejuela
Phaeotrabutiella Theiss. & Syd.
Phaeotrype Sacc.
Phanerocoryneum Höhn.
Phellomyces A.B... Frank
Phenacopodium Debey
Phialicorona Subram.
Phialocladus Kreisel
Phialocorona Subram.
Phialophoropsis L.R. Batra
Philocopra Speg.
Phoenicostroma Syd.
Phoma Fr.
Phomatosporopsis Petr.
Phomopsella Höhn.
Phomopsioides M.E.A. Costa & Sousa da Câmara
Phorcys Niessl
Phragmocarpella Theiss. & Syd.
Phragmocauma Theiss. & Syd.
Phragmodothella Theiss. & Syd.
Phruensis Pinruan
PhysalosporinaWoron.
Physosporella Höhn.
Fungal Diversity

Phytocordyceps C.H. Su & H.H. Wang
Piminella G. Arnaud
Pionnotes Fr.
Pithoascus Arx
PithospermumMont.
Placophomopsis Grove
Placostroma Theiss. & Syd.
Plagiolagynion Schrantz
Plagiostomella Höhn.
Plagiothecium Schrantz
Pleosporopsis Oerst.
Pleurage Fr.
Pleuronaema Höhn.
Pleurosordaria Fernier
Podocrea (Sacc.) Lindau
Podostroma P. Karst.
Pogonospora Petr.
Poikiloderma Füisting
Polistophthora Lebert
Polycytella C.K. Campb.
Polylagenochromatia Sousa da Câmara
Polynema Lév.
Polystigmella Jacz. & Natalina
Poroconiochaeta Udagawa & Furuya
Porodiscella Viégas
Poroniopsis Speg.
Porosphaeria Samuels & E. Müll.
Porphyrosoma Pat.
Prachtflorella Matr.
Proboscispora S.W. Wong & K.D. Hyde
Psalidosperma Syd. & P. Syd.
Psecadia Fr.
Pseudapiospora Petr.
Pseudhaplosporella Speg.
Pseudobasidium Tengwall
Pseudodiplodia Speg.
Pseudodiplodiella Bender
Pseudofusarium Matsush.
Pseudofusidium Deighton
Pseudoguignardia Gutner
Pseudohaplis Clem. & Shear
Pseudomassariella Petr.
Pseudomelasmia Henn.
Pseudomicrocera Petch
Pseudopatella Speg.
Pseudophomopsis Höhn.
Pseudophysalospora Höhn.
Pseudotrype Henn.
Psilobotrys Sacc.
Psilonia Fr.
Psiloniella Costantin
Puiggarina Speg.
Putagraivam Subram. & Bhat
Pycnidiochaeta Sousa da Câmara
Pycnofusarium Punith.
Pycnostroma Clem.
Pyreniopsis Kuntze
Pyrenium Tode
Pyrenodermium Bonord.
Pyrenodochium Bonord.
Racemella Ces.
RacemosporiumMoreau & V. Moreau
Rachidicola K.D. Hyde & J. Fröhl.
Rachisia Lindner
Radulum Fr.
Rehmiella G. Winter
Rehmiomyces (Sacc. & P. Syd.) Syd.
Rhabdostroma Syd. & P. Syd.
Rhaphidophora Ces. & De Not.
Rhaphidospora Fr.
Rheumatopeltis F. Stevens
RhexographiumM. Morelet
Rhinocephalum Kamyschko
Rhodoseptoria Naumov
Rhodothrix Vain.
Rhopalopsis Cooke
Rhopographina Theiss. & Syd.
Rhynchomelas Clem.
Rhynchophoma P. Karst.
Rhynchostomopsis Petr. & Syd.
Ribaldia Cif.
Rinia Penz. & Sacc.
Rinomia Nieuwl.
Rizaliopsis Bat. et al.
Rostrella Zimm.
Rostrella Fabre
Rostrocoronophora Munk
Rostronitschkia Fitzp.
Rostrospora Subram. & K. Ramakr.
Rubromadurella Talice
Saccardomyces Henn.
Sarcopyreniomyces Cif. & Tomas.
Sarcopyreniopsis Cif. & Tomas.
Sarcostromella Boedijn
Saturnomyces Cain
Sauvageautia Har.
Schizocapnodium Fairm.
Schizotrichella E.F. Morris
Sciniatosporium Kalchbr. ex Morgan-Jones
Scirrhiella Speg.
Scleropycnium Heald & C.E. Lewis
Scleroramularia Batzer & Crous
Scolecoccoidea F. Stevens
Scolecodothis Theiss. & Syd.
Fungal Diversity

Scolecodothopsis F. Stevens
Scoleconectria Seaver
Scoptria Nitschke
Scopulina Lév.
Seimatoantlerium Strobel et al.
Seiridina Höhn.
Selenosporium Corda
Seliniana Kuntze
Septomazzantia Theiss. & Syd.
Septomyxa Sacc.
Septorella Allesch.
Sesquicillium W. Gams
Shiraiella Hara
Shropshiria F. Stevens
Simoninus Roum.
Singera Bat. & J.L. Bezerra
Sinosphaeria J.Z. Yue & O.E. Erikss.
Sirentyloma Henn.
Solenoplea Starbäck
Spermatodermia Wallr.
Spermodermia Tode
Spermoedia Fr.
Sphaceliopsis Speg.
Sphaeria Haller
Sphaeria Tode
Sphaeroderma Fuckel
Sphaerodermatella Seaver
Sphaerodermella Höhn.
Sphaeromycetella G. Arnaud
Sphaeronemopsis Speg.
Sphaeropyxis Bonord.
Sphaerosperma Preuss
Sphinctrosporium Kunze
Spilobolus Link
Spirogramma Ferd. & Winge
Sporhelminthium Speg.
SporodermaMont.
Sporophleum Nees ex Link
Sporotheca Corda
Sporotrichella P. Karst.
Sporotrichopsis Guég.
Stagonostroma Died.
Starkeyomyces Agnihothr.
Stegastroma Syd. & P. Syd.
Steirochaete A. Braun & Casp.
Stelechotrichum Ritgen
Stevensiella Trotter
Stictosphaeria Tul. & C. Tul.
Stigmatopsis Traverso
Stilbonectria P. Karst.
Stromateria Corda
Stromatosphaeria Grev.
Stromatostilbella Samuels & E. Müll.
Stromne Clem.
Styloletendraea Weese
Stylonectriella Höhn.
Stysanopsis Ferraris
Stysanus Corda
Subramanella H.C. Srivast.
Sucinaria Syd.
Sulcospora Kohlm. & Volkm.-Kohlm.
Sydowinula Petr.
Synchaetomella Decock & Seifert
Synnemadiplodia Zambett.
Synpenicillium Costantin
Synsporium Preuss
Synsterigmatocystis Costantin
Telemeniella Bat.
Telimenopsis Petr.
Teratonema Syd. & P. Syd.
Termitaria Thaxt.
Tetracytum Vanderw.
Tettigorhyza G. Bertol.
Thielaviella Arx & T. Mahmood
Tolediella Viégas
Thozetellopsis Agnihothr.
Thozetia Berk. & F. Muell.
Thysanopyxis Rabenh. ex Bonord.
Tilakomyces Sathe & Vaidya
Titaeosporina Luijk
Titania Berl.
Tolypomyria Preuss
Tonduzia F. Stevens
Torrubia Lév.
Torsellia Fr.
Torulina Sacc. & D. Sacc.
Torulopsis Oudem.
Trabutiella Theiss. & Syd.
Trabutiella F. Stevens
Trematostoma (Sacc.) Shear
Tretendophragmia Subram.
Trichocollonema Höhn.
Trichoconium Corda
Trichodermia Hoffm.
Trichofusarium Bubák
Trichohleria Sacc.
Trichophysalospora Lebedeva
Trichotheciopsis J.M. Yen
Trigonia J.F.H. Beyma
Tripterospora Cain
Tureenia J.G. Hall
Tylodon Banker
Typhodium Link
Umbellula E.F. Morris
Fungal Diversity

Uncigera Sacc.
Underwoodina Kuntze
Urnularia P. Karst.
Urospora Fabre
Ustilaginoidella Essed
Ustilaginula Clem.
Ustilagopsis Speg.
Ustulina Tul. & C. Tul.
Vakrabeeja Subram.
Vanhallia L. Marchand
Veramyces Subram.
Vermicularia Tode
Vermiculariopsis Höhn.
Verruculina Etayo
Verticicladiella S. Hughes
Verticilliastrum Dasz.
Verticilliodochium Bubák
Verticillis Clem. & Shear
Vestergrenia (Sacc. & P. Syd.) Died.
Vittadinula (Sacc.) Clem. & Shear
Volutina Penz. & Sacc.
Winteria Sacc.
Winterina Sacc.
Woronichina Naumov
Xanthopsora Speg.
Xenostilbum Petr.
Xylariodiscus Henn.
Xylariopsis F.L. Tai
Xylopezia Höhn.
Xylosphaera Dumort.
Yukonia R. Sprague
Zygospermum Cain
Acknow l e d gmen t s Th i s r e s e a r ch was suppo r t ed byFeatured microbial resources and diversity investigation in SouthwestKarst area (2014FY120100). The authors extend their sincere appreciationsto the Deanship of Scientific Research at King SaudUniversity for its fundingthis Prolific Research Group (PRG-1436-09). Kevin D. Hyde thanks theChinese Academy of Sciences, project number 2013T2S0030, for the awardof Visiting Professorship for Senior International Scientists at KunmingInstitute of Botany. B.D. Shenoy acknowledges the funding to visit MaeFah Luang University, Chiang Rai under Indo-Thailand BilateralProgramme (THAI-1205). Y.P. Xiao and T.C. Wen are grateful to TheNational Natural Science Foundation of China (No.31460012 &No.31200016). Shi-Ke Huang and J.C. Kang are grateful to theAgricultural Science and Technology Foundation of Guizhou Province(Nos. NY[2013]3042), the International Collaboration Plan of GuizhouProvince (No. G [2012]7006) and the Innovation Team Construction forScience And Technology of Guizhou Province (No. [2012]4007) from theScience and Technology Department of Guizhou Province, China.
References
Abdel-Wahab MA (2005) Diversity of higher marine fungi fromEgyptian Red Sea mangroves. Bot Mar 48:348–355
Abdel-Wahab MA (2011) Marine fungi from Sarushima Island, Japan,with a phylogenetic evaluation of the genus Naufragella.Mycotaxon 115:443–456
Abdel-Wahab MA, Nagahama T (2011) Gesasha (Halosphaeriales,Ascomycota), a new genus with three new species from theGesashi mangroves in Japan. Nova Hedwigia 92(3–4):497–512
Abdel-Wahab MA, Nagahama T (2012) Halosarpheia japonica sp. nov.(Halosphaeriales, Ascomycota) from marine habitats in Japan.Mycol Prog 11(1):85–92
Abdel-Wahab MA, Nagahama T, Abdel-Aziz FA (2009) Two newCorollospora species and one new anamorph based on morpholog-ical and molecular data. Mycoscience 50:147–155
Abdel-Wahab MA, Pang KL, Nagahama T, Abdel-Aziz FA, Jones EBG(2010) Phylogenetic evaluation of anamorphic species ofCirrenaliaand Cumulosporawith the description of eight new genera and fournew species. Mycol Prog 9(4):537–558
Abdel-wahabMA,Abdel-Aziz FA,Mohamed SS, Abdel-Aziz AE (2011)Annulatascus nilensis sp. nov., a new freshwater ascomycete fromthe River Nile, Egypt. IMA Fungus 2:1–6
Abdel-Wahab MA, HodhodMS, Bahkali AH, Jones EBG (2014) Marinefungi of Saudi Arabia. Bot Mar 57(4):323–335
Adams GC, Wingfield MJ, Common R, Roux J (2005) Phylogeneticrelationships and morphology of Cytospora species fromEucalyptus. Stud Mycol 52:1–146
Anastasiou CJ (1963) The genus Zalerion Moore & Meyers. Can JMicrobiol 41(8):1135–1139
Andrianova TV, Minter DW (2007) New species of Bartalinia andSeptoriella from the Altai Mountains (Russia). Mycotaxon 101:297–313
Aptroot A (2004) Two new ascomycetes with long gelatinous appendagescollected from monocots in the tropics. Stud Mycol 50(2):307–312
ArzanlouM, Dokhanchi H (2013) Calosphaeria canker of almond causedby Calosphaeria pulchella in Iran. Arch Phytopathol Plant Protect46(2):215–226
Asgari B, Zare R (2011) A contribution to the taxonomy of the genusConiocessia (Xylariales). Mycol Prog 10(2):189–206
Augustin JO, Groenewald JZ, Nascimento RJ,Mizubuti ESG, Barreto RW,Elliot SL, Evans HC (2013) Yet more Bweeds^ in the garden: fungalnovelties from nests of leaf-cutting ants. PLoS ONE 8(12), e82265
Barber PA, Crous PW, Groenewald JZ, Pascoe IG, Keane P (2011)Reassessing Vermisporium (Amphisphaeriaceae), a genus of foliarpathogens of eucalypts. Persoonia 27:90–118
Barghoorn ES, Linder DH (1944a) Halosphaeria appendiculata Linder.Farlowia 1(3):412
Barghoorn ES, Linder DH (1944b) Marine fungi: their taxonomy andbiology. Farlowia 1(3):395–467
Barr ME (1983) The ascomycete connection. Mycologia 75:1–13Barr ME (1985) Notes on the Calosphaeriales. Mycologia 77:549–565Barr ME (1990) Prodromus to nonlichenized, pyrenomycetous members
of class Hymenoascomycetes. Mycotaxon 39:43–184Barr ME, Rogers JD, Ju YM (1993) Revisionary studies in the
Calosphaeriaceae. Mycotaxon 48:529–535Baumgartner K, Fujiyoshi PT, Travadon R, Castlebury LA, Wilcox WF,
Rolshausen PE (2013) Characterization of species of Diaporthefrom wood cankers of grape in eastern North American vineyards.Plant Dis 97(7):912–920
Begoude ADB, Gryzenhout M, Wingfield MJ, Roux J (2010) Aurifilum,a new fungal genus in the Cryphonectriaceae from Terminalia spe-cies in Cameroon. Antonie Van Leeuwenhoek 98(3):263–278
Benny GL, Kimbrough JW (1980) A synopsis of the orders and familiesof plectomycetes with keys to genera. Mycotaxon 12:1–91
BerbeeML, Taylor JW (1992) Two ascomycete classes based on fruiting-body characters and ribosomal DNA sequence. Mol Biol Evol 9:278–284
Berkeley MJ, Broome CE (1874) Enumeration of the fungi of Ceylon.Part II. Bot J Linn Soc 14:29–141
Fungal Diversity

Berlese AN (1900) Icones fungorum. Pyrenomycetes. Sphaeriaceae.Allantosporae. Patavii 3:1–120
Beyma FH (1940) Beschreibung einiger neuer Pilzarten aus demCentraalbureau voor Schimmelcultures, Baarn (Nederland). VIMitteilung. Antonie van Leeuwenhoek 6:263–290
Boonyuen N, Chuaseeharonnachai C, Suetrong S, Sri-indrasutdhi V,Sivichai S, Jones EBG, Pang KL (2011) Savoryellales(Hypocreomycetidae, Sordariomycetes): a novel lineage of aquaticascomycetes inferred from multiple-gene phylogenies of the generaAscotaiwania, Ascothailandia, and Savoryella. Mycologia 103(6):1351–1371
Boonyuen N, Sri-indrasutdhi V, Suetrong S, Sivichai S, Jones EBG(2012) Annulatascus aquatorba sp. nov., a lignicolous freshwaterascomycete from Sirindhorn peat swamp forest, Narathiwat,Thailand. Mycologia 104(3):746–757
Boyd ES (1934) A developmental study of a new species ofOphiodothella. Mycologia 26:456–468
Bronson AW, Klymiuk AA, Stockey RA, Tomescu AMF (2013) A peri-thecial sordariomycete (Ascomycota, Diaporthales) from the LowerCretaceous of Vancouver Island, British Columbia, Canada. Int JPlant Sci 174(3):278–292
Bubák F, Kabát JE (1905) Vierter Beitrag zur Pilzflora von Tirol.Österreichische Botanische Zeitschrift 55:73–79
Cai L, Hyde KD, Tsui CKM (2006a) Genera of freshwater fungi. FungalDivers Res Ser 18:1–261
Cai L, Jeewon R, Hyde KD (2006b) Molecular systematics of Zopfiellaand allied genera: evidence from multi-gene sequence analyses.Mycol Res 110(4):359–368
Cai L, Hyde KD, Taylor PWJ, Weir B, Waller JM, Abang MM, Zang JC,Yang YL, Phouliyong S, Prihastuti ZY, Shivas RG,McKenzie EHC,Johnston PR (2009) A polyphasic approach for studyingColletotrichum. Fungal Divers 39:183–204
Campbell J, Shearer CA (2004) Annulusmagnus and Ascitendus, two newgenera in the Annulatascaceae. Mycologia 96:822–833
Campbell J, Shearer CA, Mitchell J, Eaton R (2002) Corollosporarevisited: a molecular approach. In: Hyde KD (ed) Fungi in marineenvironments. Fungal Diversity Press, Hong Kong, pp 15–33
Campbell J, Anderson JL, Shearer CA (2003) Systematics ofHalosarpheia based on morphological and molecular data.Mycologia 95(3):530–552
Campbell J, Volkmann-Kohlmeyer B, Gräfenhan T, Spatafora JW,Kohlmeyer J (2005) A re-evaluation of Lulworthiales: relationshipsbased on 18S and 28S rDNA. Mycol Res 109(5):556–568
Campbell J, Inderbitzin P, Kohlmeyer J, Volkmann-Kohlmeyer B (2009)Koralionastetales, a new order of marine Ascomycota in theSordariomycetes. Mycol Res 113:373–380
Cannon PF (1995) Studies on fungi with isthmoid ascospores: the genusVialaea, with the description of the new family Vialaeaceae. MycolRes 99(3):367–373
Cannon PF, Buddie AG, Bridge PD (2008) The typification ofColletotrichum gloeosporioides. Mycotaxon 104:189–204
Cannon PF, Damm U, Johnston PR, Weir BS (2012) Colletotrichum—current status and future directions. Stud Mycol 73:181–213
Carlucci A, Raimnondo ML, Santos J, Phillips AJL (2012)Plectosphaerella species associated with root and collar rots of hor-ticultural crops in southern Italy. Persoonia 28:34–48
Carmarán CC, NovasMV (2003) A review of Spegazzini taxa ofPericoniaand Sporocybe after over 115 years. Fungal Divers 14:67–76
Castlebury LA, Rossman AY, Jaklitsch WJ, Vasileva LN (2002) A pre-liminary overview of the Diaporthales based on large subunit nucle-ar ribosomal DNA sequences. Mycologia 94:1017–1031
Castlebury LA, Rossman AY, Sung GH, Hyten AS, Spatafora JW (2004)Multigene phylogeny reveals new lineage for Stachybotryschartarum, the indoor air fungus. Mycol Res 108(8):864–872
Cesati V, de Notaris G (1863) Schema di classificazione degle sferiaceiitalici aschigeri piu’ o meno appartenenti al genere Sphaeria
nell’antico significato attribuitoglide Persono. Comment. SocCrittog Ital 1(4):177–420
Chadefaud M (1960) Les végétaux non vasculaires (Cryptogamie). In:Chadefaud M, Emberger L (eds) Traité de botanique systématique,vol 1. Tome I, Masson et Cie, Paris, p 1016
Chalkley DB, Suh SO, Volkmann-Kohlmeyer B, Kohlmeyer J, Zhou JJ(2010) Diatrypasimilis australiensis, a novel xylarialean fungusfrom mangrove. Mycologia 102(2):430–437
Chaverri P, Bischoff JF, Evans HC, Hodge KT (2005) Regiocrella, a newentomopathogenic genus with a pycnidial anamorph and its phyloge-netic placement in the Clavicipitaceae. Mycologia 97(6):1225–1237
Chaverri P, Liu M, Hodge KT (2008) A monograph of the entomopatho-genic genera Hypocrella, Moelleriella, and Samuelsia gen. nov.(Ascomycota, Hypocreales , Clavicipitaceae), and theiraschersonia-like anamorphs in the Neotropics. Stud Mycol 60:1–66
Chaverri P, Salgado C, Hirooka Y, Rossman A, Samuels G (2011)Delimitation of Neonectria and Cylindrocarpon (Nectriaceae,Hypocrea les , Ascomycota ) and re la t ed genera wi thCylindrocarpon-like anamorphs. Stud Mycol 68:35–56
Checa J, Arenal F, Blanco N, Rogers JD (2008)Coniolariella hispanica sp.nov. and other additions to Coniolariella. Mycol Res 112:795–801
Checa J, Blanco MN, Moreno G (2013) Contributions to the familyThyridiaceae. New data on Sphaeriamutabilis. Mycotaxon 125:149–164
Cheewangkoon R, Groenewald JZ, Summerell BA, Hyde KD, To-anunC, Crous PW (2009) Myrtaceae, a cache of fungal biodiversity.Persoonia 23:55–85
Chen W, Shearer CA, Crane JL (1999) Phylogeny of Ophioceras spp.based on morphological and molecular data. Mycologia 91:84–94
Chen SF, Lombard L, Roux J, Wingfield MJ, Zhou XD (2011) Novelspecies of Calonectria associated with Eucalyptus leaf blight inSoutheast China. Persoonia 26:1–12
Chen S, Wingfield MJ, Roets F, Roux J (2012) Identification, pathoge-nicity and population diversity of a new stem canker pathogen in theCryphonectriaceae on Rapanea melanophloeos in South Africa. SAfr J Bot 79:179–180
Chen SF, Wingfield MJ, Roux J (2013a) Diversimorbus metrosiderotisgen. et sp. nov. and three new species of Holocryphia(Cryphonectriaceae) associated with cankers on nativeMetrosideros angustifolia trees in South Africa. Fungal Biol117(5):289–310
Chen SF, Wingfield MJ, Roets F, Roux J (2013b) A serious canker dis-ease caused by Immersiporthe knoxdaviesiana gen. et sp. nov.(Cryphonectriaceae) on native Rapanea melanophloeos in SouthAfrica. Plant Pathol 62(3):667–678
Chen SF, Morgan DP, Michailides TJ (2014) Botryosphaeriaceae andDiaporthaceae associated with panicle and shoot blight of pistachioin California, USA. Fungal Divers 67:157–179
Chesters CGC, Greenhalgh GN (1964) Geniculosporium serpens gen. etsp. nov., the imperfect morph ofHypoxylon serpens. Trans BrMycolSoc 47(3):393–401
Chu Y, Alias SA, Rizman-Idid M, Guo SY, Pang KL (2015) A molecularreappraisal of Nimbospora (Halosphaeriaceae, Microascales) and anew genus Ebullia for N. octonae. Mycoscience 56:34–41
Collado J, Platas G, Bills GF, Basilio A, Vicente F, Tormo JR, Peláez F(2006) Studies on Morinia : Recogni t ion of Morinialongiappendiculata sp. nov. as a new endophytic fungus, and a newcircumscription ofMorinia pestalozzioides. Mycologia 98(4):616–627
Corda A (1837) Icones fungorum hucusque cognitorum, vol 1. J.G.Calve, Prague
Crane C, Burgess TI (2013) Luteocirrhus shearii gen. sp. nov.(Diaporthales, Cryphonectriaceae) pathogenic to Proteaceae in thesouthWestern Australian Floristic Region. IMA Fungus 4(1):111–122
Crane JL, Dumont KP (1975) Hyphomycetes from the West Indies andVenezuela. Can J Bot 53(9):843–851
Fungal Diversity

Cribb AB, Cribb JW (1956) Marine fungi from Queensland II. Pap DeptUniv Queensl 3:97–105
Crouch JA (2014) Colletotrichum caudatum s.l. is a species complex.IMA Fungus 5(1):17–30
Crouch JA, Beirn LA (2009) Anthracnose of cereals and grasses. FungalDivers 39:19–44
Crouch JA, Clarke BB, White JF, Hillman BI (2009) Systematic analysisof the falcate-spored graminicolous Colletotrichum and a descrip-tion of six new species from warm season grasses. Mycologia 101:717–732
Crous PW, Groenewald JZ (2013) A phylogenetic re-evaluation ofArthrinium. IMA Fungus 4(1):133–154
Crous PW, Wingfield MJ, Alfenas AC, Silveira SF (1994)Cylindrocladium naviculatum sp. nov. and two new vesiculate hy-phomycete genera, Falcocladium and Vesicladiella. Mycotaxon 50:441–458
Crous PW, Kendrick WB, Alfenas AC (1997) New species of hyphomy-cetes associated with Eucalyptus. S Afr J Bot 63:286–290
Crous PW, Groenewald JZ, Risède JM, Simoneau P, Hywel-Jones NL(2004) Calonectria species and their Cylindrocladium anamorphs:species with sphaeropedunculate vesicles. Stud Mycol 50:415–430
Crous PW, Groenewald JZ, Himaman W (2007) Falcocladiumthailandicum. Fungal Planet 18
Crous PW, Groenewald JZ, Shivas RG, Edwards J, Seifert KA, AlfenasAC, Alfenas RF, Burgess TI, Carnegie AJ, Hardy GESJ, Hiscock N,Huberli D, Jung T, Louis-Seize G, Okada G, Pereira OL, StukelyMJC, Wang W, White GP, Young AJ, Mc Taggart AR, Pascoe IG,Porter IJ, Quaedvlieg W (2011) Fungal Planet description sheets:69–91. Persoonia 26:108–156
Crous PW, Groenewald JZ, Lombard L, Wingfield MJ (2012a)Homortomyces gen. nov., a new dothidealean pycnidial fungus fromthe Cradle of Humankind. IMA Fungus 3(2):109–115
Crous PW, Verkley GJM, Christensen M, Castañeda-Ruiz RF,Groenewald JZ (2012b) How important are conidial appendages?Persoonia 28:126–137
Crous PW, Summerell BA, Shivas RG, Burgess TI, Decock CA, DreyerLL, Granke LL, Guest DI, Hardy GESJ, Hausbeck MK, Hüberli D,Jung T, Koukol O, Lennox CL, Liew ECY, Lombard L, McTaggartAR, Pryke JS, Roets F, Saude C, Shuttleworth LA, Stukely MJC,Vánky K, Webster BJ, Windstam ST, Groenewald JZ (2012c)Fungal planet description sheets: 107–127. Persoonia 28:138–182
Crous PW, Summerell BA, Alfenas AC, Edwards J, Pascoe IG, Porter IJ,Groenewald JZ (2012d) Genera of diaporthalean coelomycetes as-sociated with leaf spots of tree hosts. Persoonia 28:66–75
Crous PW, Summerell BA, Shivas RG, Carnegie AJ, Groenewald JZ(2012e) A re-appraisal of Harknessia (Diaporthales), and the intro-duction of Harknessiaceae fam. nov. Persoonia 28:49–65
Crous PW, Shivas RG, Wingfield MJ, Summerell BA, Rossman AY,AlvesJL AGC, Barreto RW, Bell A, Coutinho ML, Flory SL,Gates G, Grice KR, Hardy GESJ, Kleczewski NM, Lombard L,Longa CMO, Louis-Seize G, Macedo F, Mahoney DP, Maresi G,Martin-Sanchez PM, Marvanová L, Minnis AM, Morgado LN,Noordeloos ME, Phillips AJL, Quaedvlieg W, Ryan PG, Saiz-Jimenez C, Seifert KA, Swart WJ, Tan YP, Tanney JB, Thu PQ,Videira SIR, Walker DM, Groenewald JZ (2012f) Fungal planetdescription sheets: 128–153. Persoonia 29:146–201
Crous PW, Wingfield MJ, Guarro J, Cheewangkoon R, van der Bank M,Swart WJ, Stchigel AM, Cano-Lira JF, Roux J, Madrid H, Damm U,WoodAR, Shuttleworth LA,Hodges CS,MunsterM,Yáñez-MoralesMJ, Zúñiga-Estrada L, Cruywagen EM, de Hoog GS, Silvera C,Najafzadeh J, Davison EM, Davison PJN, Barrett MD, Barrett RL,Manamgoda DS, Minnis AM, Kleczewski NM, Flory SL, CastleburyLA, Clay K, Hyde KD, Maússe-Sitoe SND, Chen S, Lechat C,Hairaud M, Lesage-Meessen L, Pawłowska J, Wilk M, Śliwińska-Wyrzychowska A, Mętrak M, Wrzosek M, Pavlic-Zupanc D,Maleme HM, Slippers B, Mac Cormack WP, Archuby DI,
Grünwald NJ, Tellería MT, Dueñas M, Martín MP, Marincowitz S,de Beer ZW, Perez CA,Gené J,Marin-Felix Y, Groenewald JZ (2013)Fungal planet description sheets: 154–213. Persoonia 31:188–296
Crous PW, Shivas RG, Quaedvlieg W, van der Bank M, Zhang Y,Summerell BA, Guarro J, Wingfield MJ, Wood AR, Alfenas AC,Braun U, Cano-Lira JF, García D, Marin-Felix Y, Alvarado P,Andrade JP, Armengol J, Assefa A, den Breeÿen A, Camele I,Cheewangkoon R, De Souza JT, Duong TA, Esteve-Raventós F,Fournier J, Frisullo S, García-Jiménez J, Gardiennet A, Gené J,Hernández-Restrepo M, Hirooka Y, Hospenthal DR, King A, LechatC, Lombard L, Mang SM,Marbach PAS,Marincowitz S, Marin-FelixY, Montaño-Mata NJ, Moreno G, Perez CA, Pérez Sierra AM,Robertson JL, Roux J, Rubio E, Schumacher RK, Stchigel AM,Sutton DA, Tan YP, Thompson EH, van der Linde E, Walker AK,Walker DM, Wickes BL, Wong PTW, Groenewald JZ (2014) Fungalplanet description sheets: 214–280. Persoonia 32:184–306
Currie CR, Wong B, Stuart AE, Schultz TR, Rehner SA, Mueller UG,Sung GH, Spatafora JW, Straus NA (2003) Ancient tripartite coevo-lution in the attine ant-microbe symbiosis. Science 299:386–388
Dai DQ, Wijayawardene NN, Bhat DJ, Chukeatirote E, Bahkali AH,Zhao RL, Xu J, Hyde KD (2014) Pustulomyces gen. nov. accom-modated in Diaporthaceae, Diaporthales, as revealed by morphol-ogy and molecular analyses. Cryptogam Mycol 35(1):63–72
Damm U, Woudenberg JHC, Cannon PF, Crous PW (2009)Colletotrichum species with curved conidia from herbaceous hosts.Fungal Divers 39:45–87
Damm U, Cannon PF, Woudenberg JHC, Crous PW (2012a) TheColletotrichum acutatum species complex. Stud Mycol 73:37–113
DammU, Cannon PF,Woudenberg JHC, Johnston PR,Weir BS, Tan YP,Shivas RG, Crous PW (2012b) The Colletotrichum boninense spe-cies complex. Stud Mycol 73:1–36
Damm U, Cannon PF, Liu F, Barreto RW, Guatimosim E, Crous PW(2013) The Colletotrichum orbiculare species complex: importantpathogens of field crops and weeds. Fungal Divers 61(1):29–59
Daranagama DA, Liu XZ, Chamung S, Stadler M, Hyde KD (2014) Amultigene genealogy reveals the phylogenetic position ofRhopalostroma lekae. Phytotaxa 186(4):177–187
DaranagamaDA, Camporesi E, Tian Q, Liu X, Chamyuang S, StadlerM,Hyde KD (2015) Anthostomella is polyphyletic comprising severalgenera in Xylariaceae. Fungal Divers. doi:10.1007/s13225-015-0329-6
Davey ML (2010) Annellosporium nemorosum gen. et sp. nov., anannellidic anamorph with phylogenetic affinities to the genusDaldinia (Xylariales). Karstenia 50:1–10
De Beer ZW, Seifert KA, Wingfield MJ (2013) The ophiostomatoid fun-gi: their dual position in the Sordariomycetes. In: Seifert KA, deBeer ZW,Wingfield MJ (eds) The ophiostomatoid fungi: expandingfrontiers, CBS fungal biodiversity series 12:1–19
De Beer ZW, Duong TA, Barnes I, Wingfield BD, Wingfield MJ (2014)Redefining Ceratocystis and allied genera. Stud Mycol 79:187–219
De Gruyter J, Aveskamp MM, Woudenberg JHC, Verkley GJM,Groenewald JZ, Crous PW (2009) Molecular phylogeny of Phomaand allied anamorph genera: towards a reclassification of the Phomacomplex. Mycol Res 113(4):508–519
de Hoog GS, Van Oorschot CAN (1985) Taxonomy of the Dactylariacomplex, VI. Key to the genera and check–list of epithets. StudMycol 26:97–122
De Notaris G (1839) Micromycetes italiei Dec II. Mere R Acad SciTorino II 3:80–81
De Notaris G (1844) Cenni sulla tribù dei Pirenomiceti sferiacei edescrizione di alcuni generi spettanti alla medesima. G Bot Ital 1:322–355
DebbabA,AlyAH, Proksch P (2013)Mangrove derived fungal endophytes– a chemical and biological perception. Fungal Divers 61(1):1–27
Dixon M (1968) Trichocladium pyriformis sp. nov. Trans Br Mycol Soc51(1):160–164
Fungal Diversity

Döbbeler P (2010) Hyperepiphyllous, perianthicolous Hypocreales -highly specialized ascomycetes of the phyllosphere. Mycologia102(2):404–417
Döbbeler P (2012)Gynonectria intraspora gen. et sp. nov. (Hypocreales),a perianthicolous phyllosphere fungus with a novel ascospore type.Mycol Prog 11:473–481
Doveri F, Pecchia S, Sarrocco S, Minnocci A, Vannacci G (2010)Rodentomyces, a new hypocrealean genus from Italy. FungalDivers 42:57–69
DoyleVP,Oudemans PV,Rehner SA, Litt A (2013)Habitat and host indicatelineage identity in Colletotrichum gloeosporioides s.l. from wild andagricultural landscapes in North America. PLoS ONE 8(5), e62394
Duarte S, Batista D, Bärlocherb F, Cássioa F, Pascoala C (2015) Somenew DNA barcodes of aquatic hyphomycete species. Mycoscience56(1):102–108
Dupont J, Magnin S, Rousseau F, Zbinden M, Frebourg G, Samadi S, deForges BR, Jones EBG (2009) Molecular and ultrastructural char-acterization of two Ascomycota found on sunken wood in the deepPacific Ocean, off Vanuatu Islands. Mycol Res 113(12):1351–1364
Elfar K, Torres R, Díaz GA, Latorre BA (2013) Characterization ofDiaporthe australafricana and Diaporthe spp. associated with stemcanker of blueberry in Chile. Plant Dis 97(8):1042–1050
Ellis MB (1971) Dematiaceous Hyphomycetes. CommonwealthMycological Institute, Kew
Eriksson OE (2006) Outline of Ascomycota - 2006. Myconet 12:1–82Eriksson OE, Hawksworth DL (1985) Outline of the ascomycetes—
1985. System Ascomycetum 4:1–79Eriksson OE, Winka K (1997) Supraordinal taxa of Ascomycota.
Myconet 1(1):1–16Etayo J, Flakus A, Kukwa M (2013) Niesslia echinoides (Niessliaceae,
Ascomycota), a new lichenicolous fungus on Erioderma fromBolivia. Lichenologist 45:21–24
Fallah PM, Crane JL, Shearer CA (1999) Freshwater ascomycetes: twonew species of Ascotaiwania fromNorth America. Can J Bot 77:87–92
Fernández FA, Huhndorf SM (2004) Neotropical pyrenomycetes:Porosphaerella borinquensis sp. nov. and its Pseudobotrytisterrestris anamorph. Fungal Divers 17:11–16
Fernández FA, Lutzoni FM, Huhndorf SM (1999) Teleomorph-anamorphconnections: the new pyrenomycetous genus Carpoligna and itsPleurothecium anamorph. Mycologia 91(2):251–262
Ferrer A, Shearer CA (2007) Three new species of Luttrellia from tem-perate and tropical freshwater habitats. Mycologia 99(1):144–151
Ferrer A, Miller AN, Sarmiento C, Shearer CA (2012) Three new generarepresenting novel lineages of Sordariomycetidae (Sordariomycetes,Ascomycota) from tropical freshwater habitats in Costa Rica.Mycologia 104(4):865–879
Fournier J, Stadler M, Hyde KD, Duong ML (2010) The new genusRostrohypoxylon and two new Annulohypoxylon taxa fromNorthern Thailand. Fungal Divers 40:23–36
Francuz B, Yera H, Geraut L, Bensefa-Colas L, Nghiem ZH, Choudat D(2010) Occupational asthma induced by Chrysonilia sitophila in aworker exposed to coffee grounds. Clin Vaccine Immunol 17(10):1645–1646
Fu CH, Hsieh HM, Chen CY, Chang TT, Huang YM, Ju YM (2013)Ophiodiaporthe cyatheae gen. et sp. nov., a diaporthalean pathogencausing a devastating wilt disease of Cyathea lepifera in Taiwan.Mycologia 105(4):861–872
GamsW (1971) Cephalosporium-artige Schimmeipilze (Hyphomycetes).Gustav Fischer Verlag, Stuttgart
Gams W, Jansson HB (1985) The nematode parasite Meria coniosporaDrechsler in pure culture and its classification. Mycotaxon 22(1):33–38
García D, Stchigel AM, Cano J, Calduch M, Hawksworth DL, Guarro J(2006) Molecular phylogeny of Coniochaetales. Mycol Res110(11):1271–1289
Giraldo A, Gené J, Cano J, de Hoog S, Guarro J (2012) Two new speciesof Acremonium from Spanish soils. Mycologia 104:1456–1465
Glenn AE, Bacon CW, Price R, Hanlin RH (1996) Molecular phylogenyof Acremonium and its taxonomic implications. Mycologia 88(3):369–383
Gomes RR, Glienke C, Videira SIR, Lombard L, Groenewald JZ, CrousPW (2013) Diaporthe: a genus of endophytic, saprobic and plantpathogenic fungi. Persoonia 31(1):1–41
Gräfenhan T, Schroers H-J, Nirenberg HI, Seifert KA (2011) An over-view of the taxonomy, phylogeny and typification of somenectriaceous fungi classifed in Cosmospora, Acremonium,Fusarium, Stilbella and Volutella. Stud Mycol 68:79–113
Groenewald AM, Fullerton JZ, Abeln ECA, Carlier J, Zapater MF,Buddenhagen IW, Viljoen A, Crous PW (2008) Multiple gene ge-nealogies and phenotypic characters differentiate several novel spe-cies ofMycosphaerella and related anamorphs on banana. Persoonia20:19–37
Grum-Grzhimaylo AA, Debets AJM, van Diepeningen AD, GeorgievaML, Bilanenko EN (2013) Sodiomyces alkalinus, a new holomorphicalkaliphilic ascomycete within the Plectosphaerellaceae. Persoonia31:147–158
Gryzenhout M, Myburg H, Van der Merwe NA, Wingfield BD,Wingfield MJ (2004) Chrysoporthe, a new genus to accommodateCryphonectria cubensis. Stud Mycol 50:119–142
Gryzenhout M, Myburg H, Wingfield BD, Montenegro F, Wingfield MJ(2005) Chrysoporthe doradensis sp. nov. pathogenic to Eucalyptusin Ecuador. Fungal Divers 20:39–57
Gryzenhout M, Myburg H, Hodges CS, Wingfield BD, Wingfield MJ(2006) Microthia, Holocryphia and Ursicollum, three new generaon Eucalyptus and Coccoloba for fungi previously known asCryphonectria. Stud Mycol 55:35–52
Gryzenhout M, Wingfield BD, Wingfield MJ (2009) Taxonomy, phylog-eny, and ecology of bark-infecting and tree killing fungi in theCryphonectriaceae. APS Press, St Paul, p 378
Gryzenhout M, Tarigan M, Clegg PA, Wingfield MJ (2010)Cryptometrion aestuescens gen. sp. nov. (Cryphonectriaceae) path-ogenic to Eucalyptus in Indonesia. Australas Plant Pathol 39(2):161–169
Guarro J, Cannon PF, van der Aa HA (1999) A synopsis of the genusZopfiella (Ascomycetes, Lasiosphaeriaceae). Syst Ascom 10:79–112
Guba EF (1961) Monograph of Monochaetia and Pestalotia. HarvardUniversity Press, Cambridge
Hambleton S, Nickerson NL, Seifert KA (2005) Leohumicola, a newgenus of heat-resistant hyphomycetes. Stud Mycol 53:29–52
Hanlin RT (1990a) Icones Ascomycetum Georgiae: Ophiodothellavaccinii. Mycotaxon 39:1–8
Hanlin RT (1990b) Acerviclypeatus, a new genus for the anamorph ofOphiodothella vaccinii. Mycotaxon 37:379–384
Hanlin RT (2013) Ophiodothella angustissima comb. nov, a new namefor Acerviclypeatus poriformans and O. vaccinii. Mycotaxon 123:327–334
Hansen HN, Thomas HE, Thomas HE (1937) The connection betweenDematophora necatrix and Rosellinia necatrix. Hilgardia 10:561–564
Hartig R (1883) Rhizomorpha (Dermatophora) necatrix n. sp. UntersForstbot Inst München 3:95–140
Harvey JBJ (2004) Phylogenetic studies of the marine brown algaeCystoseira and Halidrys including co-evolution with the associatedfungal endophyte Haloguignardia irritans. [Dissertation AbstractsInternational 65(5).] Proquest Information and Learning, AnnArbor, MI
Harvey JBJ, Goff LJ (2010) Genetic covariation of the marine fungalsymbiont Haloguignardia irritans (Ascomycota, Pezizomycotina)with its algal hosts Cystoseira and Halidrys (Phaeophyceae,Fucales) along the west coast of North America. Fungal Biol114(1):82–95
Fungal Diversity

Hawksworth DL (2012) Managing and coping with names of pleomor-phic fungi in a period of transition. Mycosphere 3(2):143–155
Hawksworth DL, Kirk PM, Sutton BC, Pegler DN (1995) Dictionary ofthe fungi. 8th Edn. 616
Hawksworth DL, Millanes AM, Wedin M (2010) Roselliniella revealedas an overlooked genus of Hypocreales, with the description of asecond species on parmelioid lichens. Persoonia 24:12–17
Hellwig V, Ju YM, Rogers JD, Fournier J, Stadler M (2005) Hypomiltin,a novel azaphilone fromHypoxylon hypomiltum, and chemotypes inHypoxylon sect. Hypoxylon as inferred from analytical HPLC pro-filing. Mycol Prog 4(1):39–54
Hennings P (1897) Beiträge zur Pilzflora Sudamerikas. II. Hedwigia 36:190–246
Hennings P (1898) Die in den Gewächshäusern des Berliner BotanischenGartens beobachteten Pilze. Verh Bot Ver Prov Brandenburg 40:109–177
Hennings P (1900) Fungi Africae orientalis. Bot Jahrb Syst 28:318–329Hennings P (1901) Fungi Indiae orientalis. II. Hedwigia 40:323–342Hennings P (1902) Fungi blumenaeviensis. II. a cl. Alfr. Möller lecti.
Hedwigia 41:1–33Hernández-Restrepo M, Groenewald JZ, Crous PW (2015) Neocordana
gen. nov., the causal organism of Cordana leaf spot on banana.Phytotaxa 205(4):229–238
Herrera CS, Rossman AY, Samuels GJ, Lechat C, Chaverri P (2013a)Revision of the genus Corallomycetella with Corallonectria gen.nov. for C. jatrophae (Nectriaceae, Hypocreales). Mycosystema32:518–544
Herrera CS, Rossman AY, Samuels GJ, Chaverri P (2013b)Pseudocosmospora, a new genus to accommodate Cosmosporavilior and related species. Mycologia 105(5):1287–1305
Hibbett DS, Binder M, Bischoff J et al (2007) A higher-level phyloge-netic classification of the Fungi. Mycol Res 111:509–547
Hirooka Y, Kobayashi T, Ono T, Rossman AY, Chaverri P (2010)Verrucostoma, a new genus in the Bionectriaceae from the BoninIslands, Japan. Mycologia 102(2):418–429
Hirooka Y, Rossman AY, Samuels GJ, Lechat C, Chaverri C (2012) Amonograph of Allantonectria, Nectria, and Pleonectria (Nectriaceae,Hypocreales, Ascomycota) and their pycnidial, sporodochial, andsynnematous anamorphs. Stud Mycol 71(1):1–210
Hirooka Y, Kawaradani M, Sato T (2014) Description of Gibellulopsischrysanthemi sp. nov. from leaves of garland chrysanthemum.Mycol Prog 13:13–19
Hladki AI, Romero AI (2009) Taxonomic and nomenclatural aspects ofHypoxylon taxa from Southern South America proposed bySpegazzini. Mycologia 101(5):733–744
Hosagoudar VB (2004) studies on foliicolous fungi – xviClasterosporium flagellatum and its teleomorph. Zoos’ Print J19(4):1437–1438
Hsieh HM, Ju YM, Rogers JD (2005) Molecular phylogeny ofHypoxylon and closely related genera. Mycologia 97(4):844–865
Hsieh HM, Lin CR, Fang MJ, Rogers JD, Fournier J, Lechat C, Ju YM(2010) Phylogenetic status of Xylaria subgenus Pseudoxylariaamong taxa of the subfamily Xylarioideae (Xylariaceae) and phy-logeny of the taxa involved in the subfamily. Mol Phylogenet Evol54(3):957–969
Huang F, Chen GQ, Hou X, Fu YS, Cai L, Hyde KD, Li HY (2013)Colletotrichum species associated with cultivated citrus in China.Fungal Divers 61:61–74
Hughes SJ (1979) Relocation of species of Endophragmia auct. withnotes on relevant generic names. N Z J Bot 17:139–188
Hughes SJ, Hennebert GL (1963) Microfungi X Oedemium, Dimera,Diplosporium,Gongylocladium,Cladotrichum. Can J Bot 41:773–809
Huhndorf SM, Miller AN (2011) A molecular re-appraisal of taxa in theSordariomycetidae and a new species of Rimaconus from NewZealand. Stud Mycol 68:203–210
Huhndorf SM, Fernández FA, Taylor JE, Hyde KD (2001) Two pantrop-ical Ascomycetes: Chaetosphaeria cylindrospora sp. nov. and aRimaconus, a new genus for Lasiosphaeria jamaicensis.Mycologia 93:1072–1080
Huhndorf SH, Fernández FA, Miller AN, Lodge DJ (2003) Neotropicalascomycetes 12. Mirannulata samuelsii gen. et sp. nov. andM. costaricensis sp. nov., new taxa from the Carribean and else-where. Sydowia 55(2):172–180
Huhndorf SM, Miller AN, Fernández FA (2004) Molecular systemtaticsof the Sordariales: the order and the family Lasiosphaeriaceaeredefined. Mycologia 96:368–387
Huhndorf SM,Miller AN, Greif M, Samuels MG (2009) Amplistroma gen.nov. and its relation to Walrothiella, two genera with globose asco-spores and acrodontium-like anamorphs. Mycologia 101(6):904–919
Hyde KD (1989) Intertidal mangrove fungi from north Sumatra. Can JBot 67:3078–3082
Hyde KD (1992) Fungi from decaying inter-tidal fronds of Nypafruticans, including three new genera and four new species. Bot JLinn Soc 110:95–110
Hyde KD (1996) Tropical Australian freshwater fungi. X.Submersisphaeria aquatica gen. et sp. nov. Nova Hedwigia 62:171–175
Hyde KD, Jones EBG (1986) Marine fungi from the Seychelles IV.Cucullospora mangrovei gen. et sp. nov. from dead mangroves.Bot Mar 29:491–495
Hyde KD, Jones EBG (1989) Hypophloeda rhizospora Hyde & Jones,gen. et sp. nov., a new ascomycete from intertidal prop roots ofRhizophora spp. Trans Mycol Soc Jpn 30:61–68
Hyde KD, Jones EBG (1992) Intertidal mangrove fungi: Pedumisporagen. nov. (Diaporthales). Mycol Res 96:78–80
Hyde KD, Wong SW (2000) Annulatascus fusiformis sp. nov., a newfreshwater ascomycete from the Philippines. Mycologia 92(3):353–557
Hyde KD, Cannon PF, Barr ME (1997) Phaeochoraceae, a new familyfrom palms. Syst Ascom 15:117–120
Hyde KD, Fröhlich J, Taylor JE (1998) Fungi from palms. XXXVI.Reflections on unitunicate ascomycetes with apiospores. Sydowia50:21–80
Hyde KD, Cai L, Cannon PF, Crouch JA, Crous PW, DammU, GoodwinPH, Chen H, Johnston PR, Jones EBG, Liu ZY, McKenzie EHC,Moriwaki J, Noireung P, Pennycook SR, Pfenning LH, Prihastuti H,Sato T, Shivas RG, Tan YP, Taylor PWJ, Weir BS, Yang YL, ZhangJZ (2009a) Colletotrichum—names in current use. Fungal Divers39:147–182
HydeKD, Cai L,McKenzie EHC,YangYL, Zhang JZ, Prihastuti H (2009b)Colletotrichum: a catalogue of confusion. Fungal Divers 39:1–17
Hyde KD, McKenzie EHC, KoKo TW (2011) Towards incorporatinganamorphic fungi in anatural classification – checklist and notesfor 2010. Mycosphere 2(1):1–88
Hyde KD, Jones EBG, Liu JK, Ariyawansa H, Boehm E, Boonmee S,Braun U, Chomnunti P, Crous PW, Dai DQ, Diederich P,Dissanayake A, Doilom M, Doveri F, Hongsanan S, JayawardenaR, Lawrey JD, Li YM, Liu YX, Lücking R, Monkai J, Muggia L,Nelsen MP, Pang KL, Phookamsak R, Senanayake IC, Shearer CA,Suetrong S, Tanaka K, Thambugala KM, Wijayawardene NN,Wikee S, Wu HX, Zhang Y, Aguirre-Hudson B, Alias SA, AptrootA, Bahkali AH, Bezerra JL, Bhat DJ, Camporesi E, Chukeatirote E,Gueidan C, Hawksworth DL, Hirayama K, Hoog SD, Kang JC,Knudsen K, Li WJ, Li XH, Liu ZY, Mapook A, McKenzie EHC,Miller AN, Mortimer PE, Phillips AJL, Raja HA, Scheuer C,Schumm F, Taylor JE, Tian Q, Tibpromma S, Wanasinghe DN,Wang Y, Xu JC, Yacharoen S, Yan JY, Zhang M (2013) Familiesof dothideomycetes. Fungal Divers 63:1–313
Hyde KD, Nilsson RH, Alias SA, Ariyawansa HA, Blair JE, Cai L, deCock AWAM, Dissanayake AJ, Glockling SL, Goonasekara ID,Gorczak M, Hahn M, Jayawardena RS, van Kan JAL, Laurence
Fungal Diversity

MH, Lévesque CA, Li XH, Liu JK, Maharachchikumbura SSN,Manamgoda DS, Martin FN, Mckenzie EHC, McTaggart AR,Mortimer PE, Nair PVR, Pawlowska J, Rintoul TL, Shivas RG,Spies CFJ, Summerell BA, Taylor PWJ, Terhem RB, UdayangaD, Vaghefi N, Walther G, Wilk M, Wrzosek M, Xu JC, Yan JY,Zhou N (2014) One stop shop: backbones trees for important phy-topathogenic genera: I. Fungal Divers 67:21–125
Inderbitzin P, Lim SR, Volkmann-kohlmeyer B, Kohlmeyer J, BerbeeML (2004) The phylogenetic position of Spathulospora based onDNA sequences from dried herbarium material. Mycol Res 108(7):737–748
Inderbitzin P, Bostock RM, Davis RM, Usami T, Platt HW, Subbarao KV(2011) Phylogenetics and taxonomy of the fungal vascular wilt path-ogen Verticillium, with the descriptions of five new species. PLoSONE 6(12), e28341
Index Fungorum (2014) http://www.indexfungorum.org/Names/Names.asp, Accession Date −20 December 2014
Jaklitsch WM, Voglmayr H (2011) Stromatonectria gen. nov. and notesonMyrmaeciella. Mycologia 103(2):431–440
Jaklitsch WM, Voglmayr H (2012) Phylogenetic relationships of fivegenera of Xylariales and Rosasphaeria gen. nov. (Hypocreales).Fungal Divers 52(1):75–98
Jaklitsch WM, Voglmayr H (2014) Persistent hamathecial threads in theNectriaceae, Hypocreales: Thyronectria revisited and re-instated.Persoonia 33:182–211
Jaklitsch WM, Réblová M (2015) Index Fungorum 209:1Jaklitsch WM, Samuels GJ, Dodd SL, Lu B-S, Druzhinina IS (2006)
Hypocrea rufa/Trichoderma viride: a reassessment, and descriptionof five closely related species with and without warted conidia. StudMycol 56:137–177
Jaklitsch WM, Réblová M, Voglmayr H (2013) Molecular systematics ofWoswasia atropurpurea gen. et sp. nov. (Sordariomycetidae), afungicolous ascomycete with globose ascospores and holoblasticconidiogenesis. Mycologia 105(2):476–485
Jaklitsch WM, Fournier J, Rogers JD, Voglmayr H (2014) Phylogeneticand taxonomic revision of Lopadostoma. Persoonia 32:52–82
Jeewon R, Cai L, Liew ECY, Zhang K, Hyde KD (2003a) Dyrithiopsislakefuxianensis gen. et sp. nov. from Fuxian Lake, Yunnan, China,and notes on the taxonomic confusion surrounding Dyrithium.Mycologia 95(5):911–920
Jeewon R, Liew ECY, Hyde KD (2003b) Molecular systematics of theAmphisphaeriaceae based on cladistic analyses of partial LSUrDNA gene sequences. Mycol Res 107(12):1392–1402
Johnson RG, Jones EBG, Moss ST (1984) Taxonomic Studies of theHalosphaeriaceae: Remispora Linder, Marinospora Cavaliere andCarbosphaerella Schmidt. Bot Mar 27(12):557–566
Johnson D, Sung GH, Hywel-Jones NL, Luangsa-ard JJ, Bischoff JF,Kepler RM, Spatafora JW (2009) Systematics and evolution of thegenus Torrubiella (Hypocreales, Ascomycota). Mycol Res 113:279–289
Jones EBG, Pang KL (2012) Marine fungi and fungal-like organisms. DeGruyter, Germany
Jones EBG, Johnson RG, Moss ST (1983) Taxonomic studies of theHalosphaeriaceae: Corollospora Werdermann. Bot J Linn Soc87(2):193–212
Jones EBG, Johnson RG, Moss ST (1984) Taxonomic studies of theHalosphaeriaceae: Halosphaeria Linder. Bot Mar 27:129–143
Jones EBG, McKeown TA, Moss ST, Koch J (1995) Limacospora gen.nov. and a reevaluation of Ceriosporopsis cambrensis. Can J Bot73(7):1010–1018
Jones EBG, Chatmala I, Pang KL (2006) Two new genera isolated frommarine habitats in Thailand: Pseudolignincola and Thalespora(Halosphaeriales, Ascomycota). Nova Hedwigia 83(1–2):219–232
Jones EG, Klaysuban A, Pang KL (2008) Ribosomal DNA phylogeny ofmarine anamorphic fungi: Cumulospora varia, Dendryphiella spe-cies and Orbimyces spectabilis. Raff Bull Zool 19:11–18
Jones EBG, Zuccaro A, Mitchell J, Nakagiri A, Chatmala I, Pang KL(2009a) Phylogenetic position of freshwater and marine Sigmoideaspecies: introducing a marine hyphomycete Halosigmoidea gen.nov. (Halosphaeriales). Bot Mar 52(4):349–359
Jones EBG, Sakayaroj J, Suetrong S, Somrithipol S, Pang KL (2009b)Classification of marine Ascomycota, anamorphic taxa andBasidiomycota. Fungal Divers 35:1–187
Jones EBG, Suetrong S, Cheng WH, Rungjindamai N, Sakayaroj J,Boonyuen N, Somrothipol S, Abdelwahab MA, Pang KL (2014)An additional fungal lineage in the Hypocreomycetidae(Falcocladium species) and the taxonomic revaluation ofChaetosphaeria chaetosa and Swampomyces species, based onmor-phology, ecology and phylogeny. CryptogamMycol 35(2):119–138
Jong SC, Rogers JD (1972) Illustrations and descriptions of conidialstates of some Hypoxylon species. Wash State Univ Agric Exp StnTech Bull 77:1–49
Ju YM, Rogers JD (1996) A revision of the genusHypoxylon. MycologiaMemoir No. 20. APS Press, St. Paul, p. 365
Ju YM, Rogers JD, San Martín F (1997) A revision of the genusDaldinia. Mycotaxon 61:243–293
Ju YM, Rogers JD, San Martin F, Granmo A (1998) The genusBiscogniauxia. Mycotaxon 66:1–98
Ju YM, Rogers JD, Hsieh HM, Vasilyeva L (2004) Amphirosellinia gen.nov. and a new species of Entoleuca. Mycologia 96(6):1393–1402
Ju CH, Hsieh HM, Chen CY, Chang TT, Huang YM, Ju YM (2013)Ophiodiaporthe cyatheae gen. et sp. nov., a diaporthalean pathogencausing a devastating wilt disease of Cyathea lepifera in Taiwan.Mycologia 105(4):861–872
Justavino DR, Kirschner R, Piepenbring M (2014) New species and newrecords of Meliolaceae from Panama. Fungal Divers 70(1):73–84
Kaewchai S, Soytong K, Hyde KD (2009) Mycofungicides and fungalbiofertilizers. Fungal Divers 38:25–50
Kang JC, Hyde KD, Kong RYC (1999a) Studies on Amphisphaeriales:the Amphisphaeriaceae (sensu stricto). Mycol Res 103(1):53–64
Kang JC, Hyde KD, Kong RYC (1999b) Studies on Amphisphaeriales:the Cainiaceae. Mycol Res 103:1621–1627
Keim J, Mishra B, Sharma R, Ploch S, Thines M (2014) Root-associatedfungi of Arabidopsis thaliana andMicrothlaspi perfoliatum. FungalDivers 66:99–111
Kenerley CM, Rogers JD (1976) On Hypoxylon serpens in culture.Mycologia 68:688–691
Kepler RM, Sung GH, Harada Y, Tanaka K, Tanaka E, Hosoya T,Bischoff JF, Spatafora JW (2012) Host jumping onto close relativesand across kingdoms by Tyrannicordyceps (Clavicipitaceae) gen.nov. andUstilaginoidea (Clavicipitaceae). Am J Bot 99(3):552–561
Kepler RM, Humber RA, Bischoff JF, Rehner SA (2014) Clarification ofgeneric and species boundaries for Metarhizium and related fungithrough multigene phylogenetics. Mycologia 106(4):811–829
Kirk PM, Cannon PF, David JC, Stalpers JA (2001) Ainsworth andBisby’s dictionary of the fungi, 8th edn. CABI Publishing, London
Kirk PM, Cannon PF, Minter DW, Stalpers JA (2008) Dictionary of thefungi, 10th edn. CABI, Wallingford
Kirk PM, Stalpers JA, Braun U, Crous PW, Hansen K, Hawksworth DL,Hyde KD, Lücking R, Lumbsch TH, Rossman AY, Seifert KA,Stadler M (2013) Awithout-prejudice list of generic names of fungifor protection under the International Code of Nomenclature foralgae, fungi, and plants. IMA Fungus 4:381–443
Klaubauf S, Tharreau D, Fournier E, Groenewald JZ, Crous PW, de VriesRP, Lebrun MH (2014) Resolving the polyphyletic nature ofPyricularia (Pyriculariaceae). Stud Mycol 79:85–120
Klaysuban A, Sakayaroj J, Jones EBG (2014) An additional marine fun-gal lineage in the Diatrypaceae, Xylariales: Pedumisporarhizophorae. Bot Mar 57(5):413–420
Koch K (2013) Arenariomyces truncatellus sp. nov., an ascomycete ondriftwood from the north coast of Zealand, Denmark. Mycotaxon124:69–72
Fungal Diversity

Koch J, Jones EBG (1984) Lulworthia lignoarenaria, a new marinepyrenomycete from coastal sands. Mycotaxon 20(2):389–395
Koch J, Jones EBG, Moss ST (1983) Groenhiella bivestia, gen. et sp.nov., a lignicolous marine fungus from Denmark. Bot Mar 26:265–270
Koch J, Pang KL, Jones EBG (2007) Rostrupiella danica gen. et sp. nov.,a Lulworthia-like marine lignicolous species from Denmark. BotMar 50:294–301
Kohlmeyer J (1960) Wood-inhabiting marine fungi from the PacificNorth-West, California. Nova Hedwigia 2:293–343
Kohlmeyer J (1961) Pilze von der nördlichen Pazifik-küste der USA.Nova Hedwigia 3:85–91
Kohlmeyer J (1962) Corollospora maritima Werderm.: Ein Ascomycet.Ber Dtsch Bot Ges 74:305–310
Kohlmeyer J (1970) Ein neuer Ascomycet auf Hydrozoen im Südatlantik.Ber Dtsch Bot Ges 83:505–509
Kohlmeyer J (1973) Spathulosporales, a new order and possible missinglink between Laboulbenniales and Pyrenomycetes. Mycologia 65:614–647
Kohlmeyer J (1984) Tropical marine fungi. P.S.Z.N.I. Mar Ecol 5:329–378
Kohlmeyer J, Kohlmeyer E (1977) Bermuda marine fungi. Trans BrMycol Soc 68(2):207–219
Kohlmeyer J, Kohlmeyer E (1979) Marine mycology. The higher fungi.Academic, New York
Kohlmeyer J, Volkmann-Kohlmeyer B (1988) Halographis(Opegraphales), a new endolithic lichenoid from corals and snails.Can J Bot 66(6):1138–1141
Kohlmeyer J, Volkmann-Kohlmeyer B (1989) Hawaiian marine fungi,including two new genera of Ascomycotina. Mycol Res 92(4):410–421
Kohlmeyer J, Volkmann-Kohlmeyer B (1991) Hapsidascus hadrus gen.et sp. nov. (Ascomycota) from mangroves in the Caribbean. SystAscom 10:113–120
Kohlmeyer J, Volkmann-Kohlmeyer B (1993) Observations onHydronectria and Kallichroma gen. nov. Mycol Res 97(6):753–761
Kohlmeyer J, Spatafora JW, Volkmann-Kohlmeyer B (2000)Lulworthiales, a new order of marine Ascomycota. Mycologia92(3):453–458
Krug JC (1977) The genus Cainia and a new family, Cainiaceae.Sydowia 30(1–6):122–133
Kruys A, Huhndorf S, Miller A (2015) Coprophilous contributions to thephylogeny of Lasiosphaeriaceae and allied taxa within Sordariales(Ascomycota, Fungi). Fungal Divers 70(1):101–113
Kumar S, Stecher G, Peterson D, Tamura K (2012) MEGA-CC: comput-ing core of molecular evolutionary genetics analysis program forautomated and iterative data analysis. Appl Note 28(20):2685–2686
Lackner M, de Hoog GS, Yang L, Moreno LF, Ahmed SA et al (2014)Proposed nomenclature for Pseudallescheria, Scedosporium and re-lated genera. Fungal Divers 67:1–10
Læssøe T, Rogers JD,WhalleyAJS (1989)Camillea, Jongiella and light–spored species of Hypoxylon. Mycol Res 93:121–155
Læssøe T, Srikitikulchai P, Fournier J, Köpcke B, Stadler M (2010)Lepraric acid derivatives as chemotaxonomic markers inHypoxylon aeruginosum, Chlorostroma subcubisporum andC. cyaninum, sp. nov. Fungal Biol 114(5–6):481–489
Læssøe T, Srikitikulchai P, Luangsa-ard JJD, StadlerM (2013) Theisseniareconsidered, including molecular phylogeny of the type speciesT. pyrenocrata and a new genus Durotheca (Xylariaceae,Ascomycota). IMA Fungus 4(1):57–69
Leong WF, Tan TK, Hyde KD, Jones EBG (1990) Payosphaeria minutagen. et sp. nov., an ascomycete on mangrove wood. Bot Mar 33:511–514
Lima NB, BatistaMVDA, DeMorais Jr MA, BarbosaMA,Michereff SJ,Hyde KD, Câmara MP (2013) Five Colletotrichum species are
responsible for mango anthracnose in northeastern Brazil. FungalDivers 61:75–88
Lindqvist JC, Wright JE (1964) Discoxylaria genero nuevo la formaperfecta de Hypocreodendron. Darwiniana 13:138–143
Liu F, Cai L (2012) Morphological and molecular characterization of anovel species of Simplicillium fromChina. CryptogamMycol 33(2):137–144
Liu JK, Tan JW, Dong ZJ, Ding ZH, Wang XH, Liu PG (2002)Neoengleromycin, a novel compound from Engleromyces goetzii.Helv Chim Acta 85(5):1439–1442
Liu F, Hyde KD, Cai L (2011) Neotypification of Colletotrichumcoccodes, the causal agent of potato black dot disease and tomatoanthracnose. Mycology 2(4):248–254
Liu F, Hu DM, Cai L (2012) Conlarium duplumascospora gen. et. sp.nov. and Jobellisia guangdongensis sp. nov. from freshwater habi-tats in China. Mycologia 104(5):1178–1186
Liu F, Damm U, Cai L, Crous P (2013) Species of the Colletotrichumgloeosporioides complex associated with anthracnose diseases ofProteaceae. Fungal Divers 61:89–105
Liu JK, Hyde KD, Jones EBG, et al (2015) Fungal diversity notes 1–110:taxonomic and phylogenetic contributions to fungal species. FungalDivers. doi:10.1007/s13225-015-0324-y
Lloyd CG (1919) Synopsis of some genera of the large Pyrenomycetes.(7):16–32
Lloyd CG (1924a) Mycological notes 73. Mycol Writ 7(73):1301–1332Lloyd CG (1924b) Mycological notes 72. Mycol Writ 7(72):1269–1300Locquin MV (1972) Synopsis generalis fungorum, excerpta ex libro’De
Taxia Fungorum’. Revue de Mycologie, SupplémenLocquin M (1984) Mycologie Générale et Structurale. Masson, ParisLombard L, Crous PW (2012) Phylogeny and taxonomy of the genus
Gliocladiopsis. Persoonia 28:25–33Lombard L, Bezuidenhout KM, Crous PW (2013) Novel Ilyonectria
species associated with cutting rot of Proteaceae. Australas PlantPathol 42:337–349
Lombard L, van der Merwe NA, Groenewald JZ, Crous PW (2015)Generic concepts in Nectriaceae. Stud Mycol 80:189–245
Luangsa-ard JJ, Houbraken J, van Doorn T, Hong S-B, Borman AM,Hywel-Jones NL, Samson RA (2011) Purpureocillium, a new genusfor the medically important Paecilomyces lilacinus. FEMSMicrobiol Lett 321:141–149
Lumbsch HT, Huhndorf SM (2010)Myconet volume 14 part one. Outlineof ascomycota-2009. Fieldiana Life Earth Sci 1:1–922
Luo J, Zhang N (2013) Magnaporthiopsis , a new genus inMagnaporthaceae (Ascomycota). Mycologia 105(4):1019–1029
Luo J, Zhuang WY (2012) Volutellonectria (Ascomycota, Fungi), a newgenus with Volutella anamorphs. Phytotaxa 44:1–10
Magyar D, Shoemaker RA, Bobvos J, Crous PW, Groenewald JZ (2011)Pyrigemmula, a novel hyphomycete genus on grapevine and treebark from Hungary. Mycol Prog 10(3):307–314
Maharachchikumbura SSN, Guo LD, Chukeatirote E, Bahkali AH, HydeKD (2011) Pestalotiopsis–morphology, phylogeny, biochemistryand diversity. Fungal Divers 50:167–187
Maharachchikumbura SSN, Guo LD, Cai L, Chukeatirote E, WuWP SX,Crous PW, Bhat DJ,McKenzie EHC, Bahkali AH, Hyde KD (2012)A multi-locus backbone tree for Pestalotiopsis, with a polyphasiccharacterization of 14 new species. Fungal Divers 56:95–129
Maharachchikumbura SS, Guo LD, Chukeatirote E, McKenzie EH, HydeKD (2013a) A destructive new disease of Syzygium samarangensein Thailand caused by the new species Pestalotiopsissamarangensis. Trop Plant Pathol 38:227–235
Maharachchikumbura SSN, Chukeatirote E, Guo LD et al (2013b)Pestalotiopsis species associated with Camellia sinensis (tea).Mycotaxon 123:47–61
Maharachchikumbura SSN, Hyde KD, Groenewald JZ, Xu J, Crous PW(2014) Pestalotiopsis revisited. Stud Mycol 79:121–186
Fungal Diversity

Malloch D, Blackwell M (1990) Kathistes, a new genus of pleomorphicascomycetes. Can J Bot 68:1712–1721
Malloch D, Cain RF (1971) New cleistothecial Sordariaceae and a newfamily, Coniochaetaceae. Can J Bot 49:869–880
Manamgoda DS, Udayanga D, Cai L, Chukeatirote E, Hyde KD (2013)Endophytic Colletotrichum from tropical grasses with a new speciesC. endophytica. Fungal Divers 61:107–115
Marcelino J, Giordano R, Gouli S, Gouli V, Parker BL, Skinner M,TeBeest D, Cesnik R (2008) Colletotrichum acutatum var. fioriniae(teleomorph:Glomerella acutata var. fioriniae var. nov.) infection ofa scale insect. Mycologia 100(3):353–374
Marincowitz S, Crous PW, Groenewald JZ, Wingfield MJ (2008)Microfungi occurring on the Proteaceae in the fynbos. CBSBiodiversity Series 7 CBS Fungal Biodiversity Centre, Utrecht,The Netherlands
Marincowitz S, Gryzenhout M, Wingfield MJ (2010) New and rarecoelomycetes with appendage-bearing conidia from Pondoland,South Africa. Mycotaxon 111(1):309–322
Matsushima T (1980) Matsushima mycological memoirs No. 1.Matsushima, Kobe
Matsushima T (1993) Matsushima mycological memoirs No. 7.Matsushima, Kobe
Matsushima T (1996) Matsushima mycological memoirs No. 9.Matsushima, Kobe
Matsushima T (2001) Matsushima mycological memoirs 9. Matsushimamycological memoirs 10:1–214
McTaggart AR, Grice KR, Shivas RG (2013) First report of Vialaeaminutella in Australia, its association with mango branch diebackand systematic placement of Vialaea in the Xylariales. Aust PlantDis Notes 8:63–66
Mejía LC, Castlebury LA, White JF (2011) A systematic account of thegenus Plagiostoma (Gnomoniaceae, Diaporthales) based on mor-phology, host-associations, and a four-gene phylogeny. Stud Mycol68:211–235
Mejía LC, Rossman AY, Castlebury LA, Yang ZL, White JF Jr (2012)Occultocarpon, a new monotypic genus ofGnomoniaceae on Alnusnepalensis from China. Fungal Divers 52(1):99–105
Meyers SP, Moore RT (1960) Thalassiomycetes II. New genera and spe-cies of Deuteromycetes. Am J Bot 47:345–349
Miller AN, Vasilyeva LN, Rogers JD (2007) Chlorostromasubcubisporum gen. et sp. nov. and notes on the systematic positionof Thuemenella cubispora. Sydowia 59(1):138–147
Miller AN, Huhndorf SM, Fournier J (2014) Phylogenetic relationshipsof five uncommon species of Lasiosphaeria and three new speciesin the Helminthosphaeriaceae (Sordariomycetes). Mycologia106(3):505–524
Möller A (1901) Phycomyceten und Ascomyceten. Untersuchungen ausBrasilien. G. Fischer, Jena
Moore RT, Meyers SP (1962) Thalassiomycetes III. The genus Zalerion.Can J Microbiol 8:407–416
Morgan-Jones G (1976) Notes on Hyphomycetes. X. Codinaeopsis gen.nov. Mycotaxon 4(1):166–170
Moriwaki J, Sato T, Tsukiboshi T (2003) Morphological and molecularcharacterization of Colletotrichum boninense sp. nov. from Japan.Mycoscience 44:47–53
Mostert L, Crous PW, Groenewald JZ, Gams W, Summerbell R (2003)Togninia (Calosphaeriales) is confirmed as teleomorph ofPhaeoacremonium by means of morphology, sexual compatibility,and DNA phylogeny. Mycologia 95:646–659
Mostert L, Groenewald JZ, Summerbell RC, Gams W, Crous PW (2006)Taxonomy and pathology of Togninia (Diaporthales) and itsPhaeoacremonium anamorphs. Stud Mycol 54:1–115
MoutonM,WingfieldMJ, vanWyk PS (1993) Conidium development inthe Knoxdaviesia anamorph of Ceratocystiopsis proteae.Mycotaxon 44:363–370
Mouzouras R, Jones EBG (1985)Monodictys pelagica, the anamorph ofNereiospora cristata (Halosphaeriaceae). Can J Bot 63:2444–2447
Mugambi GK, Huhndorf SM (2010) Multigene phylogeny of theCoronophorales: morphology and new species in the order.Mycologia 102(1):185–210
Müller E, Samuels GJ (1982) Anamorphs of pyremycetous AscomycetesII. Porosphaerella gen. nov. and its Cordana anamorph. Sydowia35:150–154
Müller E, von Arx JA (1962) Die Gattungen der didymosporenPyrenomyceten. Beitr Kryptogamenfl Schweiz 11:1–922
Nag Raj TR (1985) Redisposals and redescriptions in the Monochaetia-Seiridium, Pestalotia-Pestalotiopsis complexes. II. Pestalotiopsisbesseyii (Guba) comb. nov. and Pestalosphaeria varia sp. nov.Mycotaxon 22:52–63
Nag Raj TR (1993) Coelomycetous anamorphs with appendage bearingconidia. Mycologue, Waterloo
Nag Raj T, Kendrick B (1985) Ellurema gen. nov., with notes onLepteutypa cisticola and Seiridium canariense. Sydowia 38:178–193
Nakabonge G, Gryzenhout M, Roux J, Wingfield BD, Wingfield MJ(2006) Celoporthe dispersa gen. et sp. nov. from native Myrtalesin South Africa. Stud Mycol 55:255–267
Nawawi A, Kuthubutheen AJ (1989) Canalisporium, a new genus oflignicolous hyphomycetes from Malaysia. Mycotaxon 34(2):475–487
Nelson PE, Wilhelm S (1956) An undescribed fungus causing a root rotof strawberry. Mycologia 48(4):547–551
Nitschke TRJ (1867) Pyrenomycetes Germanici. Die KernpilzeDeutschlands bearbeitet von Dr. Th. Nitschke 1: i–ii, 1–160.Eduard Trewendt, Breslau
Nitschke T (1870) Pyrenomycetes Germanici. Eduard Trewendt 2:161–320
Nonaka K, Miyazaki H, Iwatsuki M, Shiomi K, Tomoda H, Omura S,Masuma R (2012) Staphylotrichum boninense, a new hyphomycete(Chaetomiaceae) from soils in the Bonin Islands, Japan.Mycoscience 53(4):312–318
Nonaka K, Kaifuchi S, Ōmura S, Masuma R (2013) Five newSimplicillium species (Cordycipitaceae) from soils in Tokyo,Japan. Mycoscience 54(1):42–53
Nylander JA, Ronquist F, Huelsenbeck JP, Nieves-Aldrey JL (2004)Bayesian phylogenetic analysis of combined data. Syst Biol 53(1):47–67
Okane I, Nakagiri A (2007) Taxonomy of an anamorphic xylariaceousfungus from a termite nest found together with Xylaria angulosa.Mycoscience 48(4):240–249
Pang KL, Jheng JS (2012a) A checklist of marine fungi of Taiwan with adescription of Kitesporella keelungensis gen. et sp. Nov. Bot Mar55(5):459–466
Pang KL, Jheng JS (2012b) Pileomyces formosanus gen. et sp. nov.(Halosphaeriaceae, Ascomycota) from a rocky shore of Taiwan.Bot Stud 53:535–539
Pang KL, Jones EBG (2004) Reclassifications in Halosarpheia and re-lated genera with unfurling ascospore appendages. Nova Hedwigia78:269–271
Pang KL, Vrijmoed LLP, Kong RYC, Jones EBG (2003) Lignincola andNais, polyphyletic genera of the Halosphaeriales (Ascomycota).Mycol Prog 2:29–36
Pang KL, Jones EBG, Vrijmoed LLP, Vikineswary S (2004)Okeanomyces, a new genus to accommodate Halosphaeriacucullata (Halosphaeriales, Ascomycota). Bot J Linn Soc 146:223–229
Pang KL, Jones EBG, Vrijmoed LLP (2008) Autecology ofAntennospora (Fungi: Ascomycota: Sordariomycetidae:Halosphaeriales) and its phylogeny. Raffles Bull Zool Suppl 19:1–10
Fungal Diversity

Pang KL, Alias SA, Chiang MWL, Vrijmoed LLP, Jones EBG (2010)Sedecimiella taiwanensis gen. et sp. nov., a marinemangrove fungusin the Hypocreales (Hypocreomycetidae, Ascomycota). BotMar 53:493–498
Pang KL, Chiang WL, Jheng JS (2012) Morphological evaluation ofperidial wall, ascus and ascospore characteristics in the delineationof genera with unfurling ascospore appendages (Halosphaeriaceae).Prog Mol Subcell Biol 53:159–171
Pearce CA, Hyde KD (1993a) The genus Phyllachora from Australia.P. queenslandica and notes on P. apiculata from Neolitsea. MycolRes 97:1328–1332
Pearce CA, Hyde KD (1993b) The genus Ophiodothella from Australia.Mycol Res 97:1272–1276
Pearce CA, Hyde KD (1994) The genus Phyllachora from Australia:observations on taxa from Callistemon species. Mycol Res 98:1393–1401
Pearce CA, Reddell P, Hyde KD (2001) A revision of thePhyllachoraceae (Ascomycota) on the hosts in the angiosperm fam-ily, Proteaceae. Aust Syst Bot 14:283–328
Peck CH (1911) Report of the state botanist. Bull N Y State Mus 150:5–100
Pedersen EJ, Larsen P, Boll PM (1980) Neoengleromycin, a new cyto-chalasin from Engleromyces goetzii. Tetrahedron Lett 21(52):5079–5082
PeiGui L, Doi Y, XiangHua W, QingBin W (2000) The Hypocreaceae ofChina III. Some fungicolous species of the genus Hypocrea.Mycosystema 19(3):317–327
Peng LJ, Yang YL, Hyde KD, Bahkali AH, Liu Z (2012) Colletotrichumspecies on Citrus leaves in Guizhou and Yunnan provinces, China.Cryptogam Mycol 33:267–283
Peng LJ, Sun T, Yang YL, Cai L, Hyde KD, Bahkali HA, Liu ZY (2013)Colletotrichum species on grape in Guizhou and Yuannan prov-inces, China. Mycoscience 54:29–41
PerdomoH, García D, Gené J, Cano J, SuttonDA, Summerbell R, GuarroJ (2013) Phialemoniopsis, a new genus of Sordariomycetes, andnew species of Phialemonium and Lecythophora. Mycologia105(2):398–421
Pereira J, Rogers JD, Bezerra JL (2010) New Annulohypoxylon speciesfrom Brazil. Mycologia 102(1):248–252
Petersen KRL, Koch J (1996) Anisostagma rotundatum gen. et sp. nov., alignicolous marine ascomycete from Svanemøllen Harbour inDenmark. Mycol Res 100:209–212
Petrak F (1923) Beiträge zur Pilzflora von Sternberg in Mähren. AnnMycol 21:107–132
Petrak VF (1947) Neobroomella n. gen., eine neue Gattung derSphaeriales. Sydowia 1(1–3):5
Petrini LE (2013) Rosellinia - a world monograph. Bibl Mycol 205:1–410
Petrini LE, Rogers JD (1986) A summary of the Hypoxylon serpenscomplex. Mycotaxon 26:401–436
Phoulivong S, Cai L, Chen H, McKenzie EHC, Abdelsalam K,Chukeatirote E, Hyde KD (2010) Colletotrichum gloeosporioidesis not a common pathogen on tropical fruits. Fungal Divers 44(1):33–43
Pinnoi A, Phongpaichit S, Jeewon R, Tang A, Hyde KD, Jones EBG(2010) Phylogenetic relationships of Astrocystis eleiodoxae sp.nov. (Xylariaceae). Mycosphere 1(1):1–9
Pinruan U, Rungjindamai N, Sakayaroj J, Lumyong S, Hyde KD, JonesEBG (2010) Baipadisphaeria gen. nov., a freshwater ascomycete(Hypocreales, Sordariomycetes) from decaying palm leaves inThailand. Mycosphere 1:53–63
Plaingam N, Somrithipol S, Jones EBG (2003) Infundibulomyces: a newgenus of coelomycetes from Thailand. Can J Bot 81(7):732–737
Põldmaa K (2011) Tropical species of Cladobotryum and Hypomycesproducing red pigments. Stud Mycol 68:1–34
Prados-Rosale RC, Roldán-Rodríguez R, Serena C, López-Berges MS,Josep Guarro J, Martínez-del-Pozo A, Di Pietro A (2012) A PR-1-like protein of Fusarium oxysporum functions in virulence on mam-malian host. J Biol Chem 287:21970–21979
Pratibha J, Prabhugaonkar A, Hyde KD, Bhat DJ (2014) Phylogeneticplacement of Bahusandhika, Cancellidium and Pseudoepicoccum(asexual Ascomycota). Phytotaxa 176(1):68–80
PrillieuxM (1904) Sur la dehiscence des peritheces duRosellinia necatrix(R. Hart.) Berlese. Bull Soc Mycol Fr 20:34–40
Quandt CA, Kepler RM, Gams W, Araújo JP, Ban S, Evans HC, HughesD, Humber R, Hywel-Jones N, Li Z, Luangsa-ard JJ, Rehner SA,Sanjuan T, Sato H, Shrestha B, Sung GH, Yao YJ, Zare R, SpataforaJW (2014) Phylogenetic-based nomenclatural proposals forOphiocordycipitaceae (Hypocreales) with new combinations inTolypocladium. IMA Fungus 5(1):121
Raja HA, Miller AN, Shearer CA (2010) Freshwater Ascomycetes:Hyalorostratum brunneisporum, a new genus and species in theDiaporthales (Sordariomycetidae, Sordariomycetes) from NorthAmerica. Mycosphere 1(4):275–288
Rakotoniriana EF, Scauflaire J, Rabemanantsoa C, Urveg-RatsimamangaS, Corbisier AM, Quetin-Leclercq J, Declerck S, Munaut F (2013)Colletotrichum gigasporum sp. nov., a new species ofColletotrichum producing long straight conidia. Mycol Prog 12(2):403–412
Ranghoo VM, Hyde KD (1998) Ascomycetes from freshwater habitats:Ascolacicola aquatica gen. et sp. nov. and a new species ofAscotaiwania from wood submerged in reservoir in Hong Kong.Mycologia 90:1055–1062
Ranghoo VM, Hyde KD, Liew ECY, Spatafora JW (1999) Family place-ment of Ascotaiwania and Ascolacicola based on DNA sequencesfrom the large subunit rRNA gene. Fungal Divers 2:159–168
Ranghoo VM, Tsui CKM, Hyde KD (2001) Bruneosporella aquaticagen. et sp. nov., Aqualignicola hyalina gen. et sp. nov., Jobellisiaviridifusca sp. nov. and Porosphaerellopsis bipolaris sp. nov.(ascomycetes) from submerged wood in freshwater habitats.Mycol Res 105:625–633
Réblová M (1999a) Teleomorph-anamorph connections in Ascomycetes3: three new lignicolous species ofHelminthosphaeria. Sydowia 51:233–244
Réblová M (1999b) Studies in Chaetosphaeria sensu lato I. The generaChae tosphaere l la and Tengiomyces gen nov of theHelminthosphaeriaceae. Mycotaxon 70:387–420
Réblová M (2002) Synaptospora olandica, a new species from Sweden.Sydowia 54(2):248–255
Réblová M (2003) Utriascus, a new ascomycetous genus in theSordariales. Mycologia 95(1):128–133
Réblová M (2008) Bellojisia, a new sordariaceous genus for Jobellisiarhynchostoma and a description of Jobellisiaceae fam. nov.Mycologia 100:893–901
Réblová M (2011) New insights into systematics and phylogeny of thegenus Jattaea and similar fungi of the Calosphaeriales. FungalDivers 49:167–198
Réblová M (2013a) Two taxonomic novelties in the Sordariomycetidae:Ceratolenta caudata gen. et sp. nov. andPlatytrachelon abietis gen. etcomb. nov. for Ceratosphaeria abietis. Mycologia 105(2):462–475
Réblová M (2013b) Sporoschismopsis angustata sp nov, a newholomorph species in the Reticulascaceae (Glomerellales), and areappraisal of Sporoschismopsis. Mycol Prog 13(3):671–681
Réblová M, Seifert KA (2004) Cryptadelphia (Trichosphaeriales), a newgenus for holomorphs with Brachysporium anamorphs, and clarifi-cation of the taxonomic status of Wallrothiella. Mycologia 96:343–367
Réblová M, Mostert L (2007) Romellia is congeneric with Togninia, anddescription of Conidiotheca gen. nov. for one species of this genuswith polysporous asci. Mycol Res 111:299–307
Fungal Diversity

Réblová M, Seifert KA (2011) Discovery of the teleomorph of the hy-phomycete, Sterigmatobotry smacrocarpa, and epitypification ofthe genus to holomorphic status. Stud Mycol 68:193–202
RéblováM,Winka K (2001) Generic concepts and correlations in ascomy-cetes based on molecular and morphological data: Lecythotheciumduriligni gen. et sp. nov. with a Sporidesmium anamorph, andAscolacicola austriaca sp. nov. Mycologia 93:479–493
Réblová M, Mostert M, Gams W, Crous PW (2004) New genera in theCalosphaeriales: Togniniella and its anamorph Phaeocrella, andCalosphaeriophora as anamorph of Calosphaeria. Stud Mycol 50:533–550
Réblová M, Fournier J, Hyde KD (2010) Achroceratosphaeria, a newgenus for freshwater and terrestrial fungi (Ascomycetes). FungalDivers 43:75–84
Réblová M, Gams W, Seifert KA (2011a) Monilochaetes and allied gen-era of the Glomerellales, and a reconsideration of families in theMicroascales. Stud Mycol 68:163–191
Réblová M, GamsW, Štěpánek V (2011b) The new hyphomycete generaBrachyalara and Infundichalara, the similar Exochalara and spe-cies of ‘Phialophora sect. Catenulatae’(Leotiomycetes). FungalDivers 46(1):67–86
Réblová M, Seifert KA, Fournier J, Stepánek V (2012) Phylogeneticclassification of Pleurothecium and Pleurotheciella gen. nov. andits dactylaria-like anamorph (Sordariomycetes) based on nuclearribosomal and protein-coding genes. Mycologia 104(6):1299–1314
Réblová M, Stepanek V, Schumacher RK (2014) Xylochrysis lucida gen.et sp. nov., a new lignicolous ascomycete (Sordariomycetidae) withholoblastic conidiogenesis. Mycologia 106(3):564–572
Réblová M, Fournier J, Štěpánek V (2015) Pisorisporiales, a new order ofaquatic and terrestrial fungi for Achroceratosphaeria andPisorisporium gen. nov. in the Sordariomycetes. Persoonia 34:40–49
Riedl H, Wechtl E (1981) Proposal to conserve the name Phomopsis(Sacc.) Bubák (1905) against Myxolibertella Höhnel (1903).Taxon 30(4):826–828
Robert V, Stegehuis G, Stalpers J (2005) TheMycoBank engine and relateddatabases. http://www.mycobank.org. Accessed 15 Dec 2014
Rogers JD (1984) Xylaria cubensis and its anamorph Xylocoremiumflabelliforme, Xylaria allantoidea, and Xylaria poitei in continentalUnited morphs. Mycologia 76:912–923
Rogers JD, Ju YM (1998) The genus Kretszchmaria. Mycotaxon 68:345–393
Rogers JD, Ju YM (2002) Ascovirgaria occulta gen. et sp. nov., Jumillerahawaiiensis sp. nov., and Lopadostoma hawaiianum sp. nov. fromHawaii. Can J Bot 80(5):478–481
Rogers JD, Læssøe T (1992) Podosordaria ingii sp. nov. and itsLindquistia anamorph. Mycotaxon 44(2):435–443
Rogers S, James KS, Butland BK, Etherington MD, O’Brien JR, JonesJG (1987) Effects of fish oil supplement on serum lipids, bloodpressure, bleeding time, haemostatic and rheological variable.Atherosclerosis 63:137–143
Rogers JD, Ju YM, Gonzalez FSM (1995) Discoxylaria myrmecophilaand its Hypocreodendron anamorph. Mycologia 87(1):41–45
Rogers JD, Ju YM, Hemmes DE (1997) Xylaria moelleroclavus sp. nov. anditsMoelleroclavus anamorphic morph. Mycol Res 101(3):345–348
Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Höhna S,Larget B, Liu L, SuchardMA, Huelsenbeck JP (2012)MrBayes 3.2:efficient Bayesian phylogenetic inference and model choice across alarge model space. Syst Biol 61(3):539–542
Rossman AY (1979) Calonectria and its type species, C. daldiniana, alater synonym of C. pyrochroa. Mycotaxon 8:321–328
Rossman AY, Samuels GJ, Lowen R (1993) Leuconectria clusiae gen.nov. and its anamorphGliocephalotrichium bulbilium with notes onPseudonectria. Mycologia 85(4):685–704
Rossman AY, Samuels GJ, Rogerson CT, Lowen R (1999) Genera ofBionectriaceae, Hypocreaceae and Nectriaceae (Hypocreales,Ascomycetes). Stud Mycol 42:3–238
Rossman AY, McKemy JM, Pardo-Schultheiss RA, Schroers HJ (2001)Molecular studies of the Bionectriaceae using large subunit rDNAsequences. Mycologia 93:100–110
Rossman AY, Farr DF, Castlebury LA (2007) A review of the phylogenyand biology of the Diaporthales. Mycoscience 48(3):135–144
Rossman AY, Seifert KA, Samuels GJ, Minnis AM, Schroers HJ,Lombard L, Crous PW, Põldmaa K, Cannon PF, Summerbell RC,Geiser DM, Zhuang W, Hirooka Y, Herrera C, Salgado-Salazar C,Priscila Chaverri P (2013) Genera inBionectriaceae,Hypocreaceae,and Nectriaceae (Hypocreales) proposed for acceptance or rejec-tion. IMA Fungus 4(1):41–51
Rossman AY, Udayanga D, Castlebury LA, Hyde KD (2014) Proposal toconserve the nameDiaporthe eres, with a conserved type, against allo ther compet ing names (Ascomycota , Diapor tha les ,Diaporthaceae). Taxon (In Press)
Ryley MJ (2003) Nigrocornus scleroticus, a common old worldbalansioid fungus. In: White JF et al (eds) Clavicipitalean fungi:evolutionary biology, chemistry, biocontrol and cultural impacts.Marcel-Dekker, Inc., New York, pp 247–271
Saccardo PA (1896) Notes mycologiques (IIe Série). Bull Soc Mycol Fr12(2):64–81
Saccardo PA (1899) Sylloge fungorum ominium hucusque cognitorum,vol 14. P. A. Saccardo, Padova
Saccardo PA, Roumeguère C (1884) Reliquiae mycologicae Libertianae.IV. Rev Mycol (Toulouse) 6:26–39
Sakayaroj J (2005) Phylogenetic relationships of marine Ascomycota.Ph.D. Thesis, Prince of Songkla University, Thailand
Sakayaroj J, Pang KL, Jones EBG, Phongpaichit S, Vrijmoed LLP, Abdel-Wahab MA (2005) A systematic reassessment of the marine ascomy-cetes Torpedospora and Swampomyces. Bot Mar 48:395–406
Sakayaroj J, Pang KL, Jones EBG (2011a) Multi-gene phylogeny of theHalosphaeriaceae: its ordinal status, relationships between generaand morphological character evolution. Fungal Divers 46:87–109
Sakayaroj J, Supaphon O, Jones EBG, Phongpaichit S (2011b) Diversityof higher marine fungi at Hat Khanom-Mu Ko Thale Tai NationalPark, Southern Thailand. Songklanakarin J Sci Technol 33:15–22
Salgado-Salazar C, Rossman AY, Samuels GJ, Capdet M, Chaverri P(2012) Multigene phylogenetic analyses of the Thelonectriacoronata and T. veuillotiana species complexes. Mycologia 104:1325–1350
Samuels GJ, Blackwell M (2001) Pyrenomycetes-fungi with perithecia.In: McLaughlin DJ, McLaughlin EG, Lemke PA (eds) The MycotaVII Part A. Springer, Berlin, pp 221–255
Samuels GJ, Rodrigues KF (1989) Batistia annulipes and its anamorph,Acrostroma anellosynnema. Mycologia 81(1):52–56
Samuels GJ, Rogers JD, Nagasawa I (1987) Studies on theAmphisphaeriaceae (sensu lato) 1. Callodiscula japonica and itsanamorph, Acanthodochium collodisculae. Mycotaxon 28(2):453–459
Samuels GJ, Candoussau F, Magni JF (1997) Fungicolouspyrenomycetes 1. Helminthosphaeria and the new familyHelminthosphaeriaceae. Mycologia 89:141–155
Samuels GJ, Ismaiel A, Mulaw TB (2012) The Longibrachiatum Cladeof Trichoderma: a revision with new species. Fungal Divers 55(1):77–108
Santos JM, Phillips AJL (2009) Resolving the complex of Diaporthe(Phomopsis) species occurring on Foeniculum vulgare in Portugal.Fungal Divers 34:111–125
Santos JM, Correia VG, Phillips AJ (2010) Primers for mating-type di-agnosis inDiaporthe and Phomopsis: their use in teleomorph induc-tion in vitro and biological species definition. Fungal Biol 114(2):255–270
Schoch CL, Crous PW, Wingfield MJ, Wingfield BD (2000) Phylogenyof Calonectria and selected hypocrealean genera with cylindricalmacroconidia. Stud Mycol 45:45–62
Fungal Diversity

Schoch CL, Shoemaker RA, Seifert KA, Hambleton S, Spatafora JW,Crous PW (2006) A multigene phylogeny of the Dothideomycetesusing four nuclear loci. Mycologia 98(6):1041–1052
Schoch CL, Sung GH, Volkmann-Kohlmeyer B, Kohlmeyer J, SpataforaJW (2007) Marine fungal lineages in the Hypocreomycetidae.Mycol Res 111:154–162
Schoch CL, Crous PW, Groenewald JZ, Boehm EWA, Burgess TI, deGruyter J, de Hoog GS, Dixon LJ, Grube M, Gueidan C, Harada Y,Hatakeyama S, Hirayama K, Hosoya T, Huhndorf SM, Hyde KD,Jones EGB, Kohlmeyer J, Kruys Å, Li YM, Lücking R, LumbschHT, Marvanová L, Mbatchou JS, McVay AH, Miller AN, MugambiGK, Muggia L, Nelsen MP, Nelson P, Owensby CA, Phillips AJL,Phongpaichit S, Pointing SB, Pujade-Renaud V, Raja HA, Plata ER,Robbertse, Ruibal C, Sakayaroj J, Sano T, Selbmann L, Shearer CA,Shirouzu T, Slippers B, Suetrong S, Tanaka K, Volkmann-Kohlmeyer B, Wingfield MJ, Wood AR, Woudenberg JHC,Yonezawa H, Zhang Y, Spatafora JW (2009) A class-wide phyloge-netic assessment of Dothideomycetes. Stud Mycol 64:1–15
Schroers HJ (2001) A monograph of Bionectria (Ascomycota,Hypocreales, Bionectriaceae) and its Chonostachys anamorphs.Stud Mycol 46:1–214
Schroers HJ, Geldenhuis MM, Wingfield MJ, Schoeman MH, Yen YF,Shen WC, Wingfield BD (2005) Classification of the guava wiltfungus Myxosporium psidii, the palm pathogen Gliocladiumvermoesenii and the persimmon wilt fungus Acremonium diospyriin Nalanthamala. Mycologia 97(2):375–395
Schroers HJ, Gräfenhan T, Seifert KA, Nirenberg H (2011) Fusariumbuxicola (Cyanonectria) revised and Geejayessia gen. nov. forNectria desmazierii and related species with Fusarium-likeanamorphs. Stud Mycol 68:115–138
Seifert KA, Gams W, Louis-Seize G (2002) Arthroxylaria elegans, anew coprphiolous anamorphic fungus allied with the Xylariaceae,with the notes on the genus Bisporostilbella. Czech Mycol 53(4):297–307
Seifert K, Morgan-Jones G, Gams W, Kendrick B (2011) The Genera ofHyphomycetes. [CBS Biodiversity Series 9]. CBS KNAW FungalBiodiversity Centre, Utrecht
Semenova EF, Shpichka AI, Moiseeva IY (2012) About essential oils bio-technology on the base of microbial synthesis. Pharm Sci 4:29–31
Senanayake IC, Maharachchikumbura SSN, Mortimer PE, Bhat JD,Hyde KD (2014) Vialaeaceae; introducing a novel species Vialaeamangiferae. Sydowia 66(2):203–216
Serrato-Diaz LM, Rivera-Vargas LI, Goenaga R, Verkley GJM, French-Monar RD (2011) First report of a Lasmenia sp. causing rachisnecrosis, flower abortion, fruit rot, and leaf spots on rambutan inPuerto Rico. Plant Dis 95(10):1313
Sharma G, Kumar N,Weir BS, Hyde KD, Shenoy BD (2013) The ApMatmarker can resolve Colletotrichum species: a case study withMangifera indica. Fungal Divers 61:117–138
Sheare CA, Miller M (1977) Fungi of Chesapeake Bay and its tributariesVIII. Ascomycetes with unfuling appendages. Bot Mar 23:607–615
Shearer CA (1973) Fungi of the ChesapeakeBay and its tributaries II. Thegeus Conioscypha. Mycologia 65:128–136
Shearer CA (1978) Fungi of the Chesapeake Bay and its tributaries VII.Luttrellia estuarina gen. et sp. nov. (Ascomycetes). Mycologia70(3):692–697
Shearer CA, Crane JL (1971) Fungi of the Chesapeake Bay and its trib-utaries. I. Patuxent River. Mycologia 63(1):237–260
Shearer CA, Crane JL (1977) Fungi of the Chesapeake Bay and its trib-utaries. VI. Trichocladium achrasporum, the imperfect state ofHalosphaeria mediosetigera, a marine ascomycete. Mycologia 69:1218–1223
Shearer CA, Crane JL (1978) Illinois fungi IX. Zopfiella lundqvistii sp.nov., a new ascomycete from submerged wood. Trans BrMycol Soc70(3):456–459
Shearer CA, Descals E, Kohlmeyer B, Kohlmeyer J, Marvanová L,Padgett D, Porter D, Raja H, Schmit J, Thorton H, Voglymayr H(2007) Fungal biodiversity in aquatic habitats. Biodivers Conserv16:49–67
Shearer CA, Fallah PM, Ferrer A, Raja HA, Schmit JP (2009) Freshwaterascomycetes and their anamorphs. http://fungi.life.illinois.edu/.Accessed Dec 2014
Shenoy BD, Jeewon R, Wu WP, Bhat DJ, Hyde KD (2006) Ribosomaland RPB2 DNA sequence analyses suggest that Sporidesmium andmorphologically similar genera are polyphyletic. Mycol Res 110:916–928
Shenoy BD, Jeewon R, Hyde KD (2007) Impact of DNA sequence-dataon the taxonomy of anamorphic fungi. Fungal Divers 26:1–54
Shenoy BD, Jeewon R, Wang H, Amandeep K, Ho WH, Bhat DJ, CrousPW, Hyde KD (2010) Sequence data reveals phylogenetic affinitiesof fungal anamorphs Bahusutrabeeja, Diplococcium, Natarajania,Paliphora, Polyschema, Rattania and Spadicoides. Fungal Divers44:161–169
Shivas RG, Tan YP (2009) A taxonomic re-assessment ofColletotrichumacutatum, introducing C. fioriniae comb. et stat. nov. andC. simmondsii sp. nov. Fungal Divers 39:111–122
Shoemaker RA (1963) Generic correlations and concepts:Griphosphaerioma and Labridella. Can J Bot 41(10):1419–1423
Shoemaker RA, Müller E (1963) Generic correlations and concepts:Broomella and Pestalotia. Can J Bot 41:1235–1243
Shoemaker RA, Hambleton S, Liu M (2013) Vialaea insculpta revisited.N Am Fungi 8(10):1–13
Sigler L, Gibas CFC, Kokotovic B, Bertelsen MF (2010) Disseminatedmycosis in veiled chameleons (Chamaeleo calyptratus) caused byChamaeleomyces granulomatis, a new fungus related toPaecilomyces viridis. J Clin Microbiol 48(9):3182–3192
Silva DN, Talhinas P, Várzea V, Cai L, Paulo OS, Batista D (2012)Application of the Apn2/MAT locus to improve the systematics ofthe Colletotrichum gloeosporioides complex: an example from cof-fee (Coffea spp.) hosts. Mycologia 104:396–409
Silvestro D, Michalak I (2011) raxmlGUI: a graphical front-end forRAxML. Org Divers Evol. doi:10.1007/s13127-011-0056-0
Sivanesan A (1975) Lasiosphaeriella, a new genus of the familyLasiosphaeriaceae. Trans Br Mycol Soc 64(3):441–445
Sivanesan A (1983) Studies on ascomycetes. Trans Br Mycol Soc 81:313–332
Sivanesan A (1984a) Teleomorph of Clasterosporium anamalum andCyperi. Trans Br Mycol Soc 83:710–718
SivanesanA (1984b) The bitunicate Ascomycetes and their anamorphs. J.Cramer, Vaduz
Sivanesan A, Alcorn JL (2002) Australiasca queenslandica gen et sp nov(Chaetosphaeriaceae: Ascomycota) and its anamorphDischloridiumcamelliae sp nov from Australia. Aust J Bot 15:741–747
S iv ane s an A , Chang HS (1992 ) Asco ta iwan ia , a n ewamphisphaeriaceous ascomycete genus on wood from Taiwan.Mycol Res 96:481–484
Sivicha S, Hywel-Jones NL, Jones EBG (1998) Lignicolous freshwaterAscomycota from Thailand 1. Ascotaiwania sawada and itsanamorph state Monotosporella. Mycoscience 39:307–311
Smith GJD, Liew ECY, Hyde KD (2003) The Xylariales: a monophyleticorder containing 7 families. Fungal Divers 13:185–218
Soares DJ, Rocha FB, Duarte LL, Barreto RW (2011) Pyriculariopsiscalatheae sp. nov., a novel anamorphic hyphomycete from theAtlantic forest of Brazil causing leaf spots on Calathea longifolia.Mycol Prog 10(3):315–321
Somrithipol S, SudhomN, Tippawan S, Jones EBG (2007) A new speciesof Falcocladium with turbinate vesicles from Thailand. Sydowia59(1):148–153
Somrithipol S, Sakayaroj J, Rungjindamai N, Plaingam N, Jones EBG(2008) Phylogenetic relationship of the coelomycete genus
Fungal Diversity

Infundibulomyces based on nuclear rDNA data. Mycologia 100(5):735–741
Spatafora JW (1995) Ascomal evolution of filamentous ascomycetes:evidence from molecular. Can J Bot 73:811–815
Spatafora JW, Blackwell M (1993) Molecular systematics of unitunicateperithecial ascomycetes: the Clavicipitales-Hypocreales connection.Mycologia 85:912–922
Spatafora JW, Volkmann-Kohlmeyer B, Kohlmeyer J (1998) Independentterrestrial origins of the Halosphaeriales (marine Ascomycota). AmJ Bot 85:1569–1580
Sri-indrasutdhi V, Boonyuen N, Sivichai S, Jones EBG (2010) Woodinhabiting freshwater fungi from Thailand: Ascothailandia gen. etsp. nov., Canalisporium grenadoidia sp. nov. with a key toCanalisporium species (Sordariomycetes Ascomycota).Mycoscience 51(6):411–420
Stadler M, Hellwig V (2005) PCR-based data and secondary metabolitesas chemotaxonomic markers in high-throughput screening for bio-active compounds from fungi. Handb Ind Mycol 269
Stadler M, Fournier J, Gardt S, Peršoh D (2010) The phylogenetic posi-tion of Rhopalostroma as inferred from a polythetic approach.Persoonia 25:11–21
Stadler M, Kuhnert E, Peršoh D, Fournier J (2013) The xylariaceae asmodel example for a unified nomenclature following the BOne fun-gus–one name^ (1F1N) concept. Mycology 4(1):5–21
Stadler M, Læssøe T, Fournier J, Decock C, Schmieschek B, Tichy HV,Peršoh D (2014) A polyphasic taxonomy ofDaldinia (Xylariaceae).Stud Mycol 77:1–143
Steiner U, Leibner S, Schardl CL, Leuchtmann A, Leistner E (2011)Periglandula, a new fungal genus within the Clavicipitaceae andits association with Convolvulaceae. Mycologia 103(5):1133–1145
Stenroos S, Laukka T, Huhtinen S, Döbbeler P, Myllys L, Syrjänen K,Hyvönen J (2010) Multiple origins of symbioses between ascomy-cetes and bryophytes suggested by a five–gene phylogeny.Cladistics 26(3):281–300
Steyaert RL (1949) Contribution à l’étude monographique de Pestalotiade Not. et Monochaetia Sacc. (Truncatella gen. nov. et Pestalotiopsisgen. nov.). Bull Jard Bot Brux 19(3):285–354
Su YY, Noireung P, Liu F, Hyde KD, Moslem MA, Bahkali AH,AbdElsalam KA, Cai L (2011) Epitypification of Colletotrichummusae, the causative agent of banana anthracnose. Mycoscience52:376–382
Subramanian CV (1972) Padixonia, a new genus of hyphomycetes. CurrSci 41:282–283
Subramanian CV (1993) Veramyces a new hyphomycete genus fromKumaon Himalayas. Czech Mycol 47:3–5
Subramanian CV, Bhat DJ (1978) Putagraivam, a new genus ofHypomycetes. Proc Indian Acad Sci B 87:99–104
Suetrong S, Klaysuban A, Sakayaroj J, Preedanon S, Chanprasert J,Phongpaichit S, Jones EBG (2015) Tirisporellaceae a new familyin the order Diaporthales (Sordariomycetes). Cryptogamie Mycol(in press)
Summerbell RC, Gueidan C, Schroers HJ, de Hoog GS, Starinck M,Arocha-Rosete Y, Guarro J, Scott JA (2011) Acremonium phyloge-netic overview and revision of Gliomastix, Trichothecium andSarocladium. Stud Mycol 68:139–162
Summerell BA, Groenewald JZ, Carnegie AJ, Summerbell RC, CrousPW (2006) Eucalyptus microfungi known from culture. 2.Alysidiella, Fusculina and Phlogicylindrium genera nova, withnotes on some other poorly known taxa. Fungal Divers 23:323–350
Sung G-H, Hywel-Jones NL, Sung J-M, Luangsaard JJ, Shresthra B,Spatafora JW (2007) Phylogenetic classification of Cordyceps andthe clavicipitaceous fungi. Stud Mycol 57:5–59
Sutherland GK (1915) Additional notes on marine pyrenomycetes. TransBr Mycol Soc 5:257–263
Sutton BC (1968) The appressoria of Colletotrichum graminicola andC. falcatum. Can J Bot 46:873–876
Sutton BC (1980) The Coelomycetes-Fungi imperfecti with Pycnidia.Acervuli and Stromata Commonwealth Mycological Institute, Kew
Sutton BC (1992) The genus Glomerella and its anamorphColletotrichum. In: Bailey JA, Jeger MJ (eds) Colletotrichum.Biology, pathology and control. CAB International, Wallingford,pp 1–26
Swart HJ, Williamson MA (1983) Australian leaf-inhabiting fungi XVI.Vermisporium, a new genus of coelomycetes on Eucalyptus leaves.Trans Br Mycol Soc 81:491–502
Taerum SJ, Cafaro MJ, Little AEF, Schultz TR, Currie CR (2007) Lowhost-pathogen specificity in the leaf-cutting ant-microbe symbiosis.Proc R Soc Lond B Biol Sci 274:1971–1978
Tanaka E, Ashizawa T, Sonoda R, Tanaka C (2008) Villosiclava virensgen. nov., comb. nov., teleomorph of Ustilaginoidea virens, thecausal agent of rice false smut. Mycotaxon 106:491–501
Tanaka K, Endo M, Hirayama K, Okane I, Hosoya T, Sato T (2011)Phylogeny of Discosia and Seimatosporium, and introduction ofAdisciso and Immersidiscosia genera nova. Persoonia 26:85–98
Tang AMC, Jeewon R, Hyde KD (2007) Phylogenetic utility of protein(RPB2, b-tubulin) and ribosomal (LSU, SSU) gene sequences in thesystematic of Sordariomycetes (Ascomycota, Fungi). Antonie VanLeeuwenhoek 91:327–349
Tangthirasunun N, Silar P, Bhat DJ, Maharachchikumbura SSN, HydeKD (2014) Greeneria saprophytica sp. nov. on dead leaves ofSyzygium cumini from Chiang Rai, Thailand. Phytotaxa 184(5):275–282
Tao G, Liu ZY, Liu F, Gao YH, Cai L (2013) Endophytic Colletotrichumspecies from Bletilla ochracea (Orchidaceae), with description ofseven new species. Fungal Divers 61:139–164
Thambugala KM, Ariyawansa HA, Li YM, Boonmee S, Hongsanan S,Tian Q, Singtripop C, Bhat DJ, Camporesi E, Jayawardena R, LiuZY, Chukeatirote XJC, Hyde KD (2014) Dothideales. Fungal Divers68(1):105–158
Thiruvudainambi S, Chandrasekar G, Baradhan G (2010) Potential an-tagonism of Trichoderma sp. against Sclerotium rolfsii Sacc. PlantArch 10(2):617–620
Thongkantha S, JeewonR, VijaykrishnaD, Lumyong S,McKenzie EHC,Hyde KD (2009) Molecular phylogeny of Magnaporthaceae(Sordariomycetes) with a new species Ophioceras chiangdaoensefrom Dracaena loureiroi in Thailand. Fungal Divers 34:157–173
Traverso GB (1906) Flora Italica Cryptogama. Pars 1: Fungi.Pyrenomycetae. Xylariaceae, Valsaceae, Ceratostomataceae 2:1–352. Società Botanica Italiana, Rocca S. Casciano, Italy
Tsui CKM, Berbee ML (2006) Phylogenetic relationships and conver-gence of helicosporous fungi inferred from ribosomal DNA se-quences. Mol Phylogenet Evol 39:587–597
Tsui CKM, Hodgkiss IJ, Hyde KD (2003) Three new species ofAquaticola (Ascomycetes) from tropical freshwater habitats. NovaHedwigia 77:161–168
Udayanga D, Liu X, Crous PW,McKenzie EH, Chukeatirote E, HydeKD(2012) A multi-locus phylogenetic evaluation of Diaporthe(Phomopsis). Fungal Divers 56(1):157–171
Udayanga D,Manamgoda DS, Liu XZ, Chukeatirote E, Hyde KD (2013)What are the common anthracnose pathogens of tropical fruits?Fungal Divers 61:165–179
Udayanga D, Castlebury LA, Rossman AY, Hyde KD (2014a) Specieslimits in Diaporthe: molecular re-assessment of D. citri,D. cytosporella, D. foeniculina and D. rudis. Persoonia 32:83–101
Udayanga D, Castlebury LA, Rossman AY, Chukeatirote E, Hyde KD(2014b) Insights into the genus Diaporthe: phylogenetic speciesdelimitation in the D. eres species complex. Fungal Divers 67(1):203–229
Uecker FA (1988) world list of Phomopsis names with notes on nomen-clature, morphology and biology. Published for the New YorkBotanical Garden in collaboration with the Mycological Society ofAmerica [by] J. Cramer
Fungal Diversity

Untereiner WA, Bogale M, Carter A, Hanson SÅ, Læssøe T, Štěpánek V,Réblová M (2013) Molecular phylogeny of Boliniales(Sordariomycetes) with an assessment of the systematics ofApiorhynchostoma, Endoxyla and Pseudovalsaria. Mycologia105(3):564–588
Van den Brink J, Samson RA, Hagen F, Boekhout T, De Vries RP (2012)Phylogeny of the industrial relevant, thermophilic generaMyceliophthora and Corynascus. Fungal Divers 52:197–207
Vasilyeva LN (1993) Pyrenomycetes of the Russian Far East. I.Gnomoniaceae. Russian Academy of Sciences, Vladivostok
Vas i l yeva LN, Che rnyshev AV, S t ephenson SL (2012 )Neochaetosphaerella thaxteriospora gen. et sp. nov. andTympanopsis texensis sp. nov. (Coronophorales, Ascomycota) fromTexas, USA. Fungal Divers 52(1):191–196
VermeulenM, Gryzenhout M,Wingfield MJ, Roux J (2011) New recordsof the Cryphonectriaceae from southern Africa includingLatruncellus aurorae gen. sp. nov. Mycologia 103(3):554–569
Viala P (1891) Monographie du pourridie (Dermatophora) – These pre-sentee a la faculte des sciences de Paris, p 124
Vieira WAS, Michereff SJ, de Morais Jr MA, Hyde KD, Câmara MPS(2014) Endophytic species ofColletotrichum associated withmangoin northeastern Brazil. Fungal Divers 67(1):181–202
Vijaykrishna D, Hyde KD (2006) Inter- and intra stream variation oflignicolous freshwater fungiin tropical Australia. Fungal Divers21:203–224
Voglmayr H, Rossmann AY, Castlebury LA, Jaklitsch WM (2012)Multigene phylogeny and taxonomy of the genus Melanconiella(Diaporthales). Fungal Divers 57(1):1–44
Volkmann-Kohlmeyer B, Kohlmeyer J (1992) Corallicola nana gen. etsp. nov. and other ascomycetes from coral reefs. Mycotaxon 44(2):417–424
Von Arx JA (1957) Die Arten der Gattung Colletotrichum Cda.Phytopathol Z 29:413–468
Von Arx JA, Müller E (1955) Über die Gattungen Delitschia Auersw.,Trichodelitschia Munk, und Cainia nov.gen. Acta Bot Neerl 4(1):108–115
Von Höhnel F (1903) Mycologische Fragmente (Fortsetzung). AnnMycol 1(6):522–534
von Höhnel F (1915) Fragmente zur Mykologie (XVII. Mitteilung, Nr.876 bis 943). Sitzungsberichte der Kaiserlichen Akademie derWissenschaften Math.-naturw. Klasse Abt I 124:49–159
von Höhnel F (1917a) System der Phacidiales v. H. Sonderabdruck. BerDsch Bot Ges 25:416–422
Von Höhnel F (1917b) Fragmente zur Mykologie (XX Mitteilung, Nr.1031 bis 1057). Sitzungsberichte der Kaiserlichen Akademie derWissenschaften Math.-naturw. Klasse Abt I 126:353–399
von Höhnel FXR (1917c) Mykologische Fragmente [CXX-CXC].Annales Mycologici 15(5):293–383
Von Schrenk H, Spaulding P (1903) The bitter rot of apples. Sci N Y 17:750–751
Wang YZ (2011) Three species excluded from Melanopsamma(Ascomycetes). Mycotaxon 115:365–368
Wang Y, Hyde KD, McKenzie EHC, Jiang YL, Li DW, Zhao DG (2015)Overview of Stachybotrys (Memnoniella) and current species status.Fungal Divers. doi:10.1007/s13225-014-0319-0
Wehmeyer LE (1933) The British species of the genus (Diaporthe) Nits.and its segregates. Trans Br Mycol Soc 17(4):237–295
Weir BS, Johnston PR (2010) Characterisation and neotypification ofGloeosporium kaki Hori as Colletotrichum horii nom. nov.Mycotaxon 111:209–219
Weir BS, Damm U, Johnston PR (2012) The Colletotrichumgloeosporioides species complex. Stud Mycol 73:115–180
Werdermann E (1922) Corollospora maritima Werd., ein salzliebenderPilz von der Nordseeküste. Notizbl Bot Gart Berlin-Dahlem 8:248–250
Whalley AJS, Watling R (1988) Versiomyces cahuchucosus gen. et sp.nov. from Queensland, Australia. Notes R Bot Gard Edinb 45(2):401–404
WhalleyMA, Khalil AMA,Wei TZ, Yao YJ,Whalley AJS (2010) A newspecies of Engleromyces from China, a second species in the genus.Mycotaxon 112:317–323
Whitton SR, McKenzie EHC, Hyde KD (2000) Dictyochaeta andDictyochaetopsis species from the Pandanaceae. Fungal Diver 4:133–158
Whitton S, Hyde KD, McKenzie EHC (2012) Fungi associated withPandanaceae. Fungal Divers Res Ser 21:458
Wijayawardene DNN, McKenzie EHC, Hyde KD (2012) Towards incor-porating anamorphic fungi in a natural classification – checklist andnotes for 2011. Mycosphere 3:157–228
Wijayawardene NN, Crous PW, Kirk PM, Hawksworth DL, Boonmee S,Braun U, Dai DQ, D’souza MJ, Diederich P, Dissanayake A,Doilom M, Hongsanan S, Jones EBG, Groenewald JZ,Jayawardena R, Lawrey JD, Liu JK, Lücking R, Madrid H,Manamgoda DS, Muggia L, Nelsen MP, Phookamsak R, SuetrongS, TanakaK, Thambugala KM,WanasingheDN,Wikee S, ZhangY,Aptroot A, Ariyawansa HA, Bahkali AH, Bhat DJ, Gueidan C,Chomnunti P, De Hoog GS, Knudsen K, Li WJ, McKenzie EHC,Miller AN, Phillips AJL, PiątekM, Raja HA, Shivas RS, Slippers B,Taylor JE, Tian Q,WangY,Woudenberg JHC, Cai L, JaklitschWM,Hyde KD (2014) Naming and outline of Dothideomycetes–2014including proposals for the protection or suppression of genericnames. Fungal Divers 69(1):1–55
Wilson IM (1956) Some new marine Pyrenomycetes on wood or rope:Halophiobolus and Lindra. Trans Br Mycol Soc 39(4):401–415
Wingfield MJ, De Beer ZW, Slippers B, Wingfield BD, Groenewald JZ,Lombard L, Crous PW (2012) One fungus, one name promotesprogressive plant pathology. Mol Plant Pathol 13(6):604–613
Wong SW, Hyde KD, Jones EBG (1998a) Halosarpheia heteroguttulatasp. nov. from submerged wood in streams. Can J Bot 76(11):1857–1862
WongMKM, PoonMOK, Hyde KD (1998b) Phragmitensis marina gen.et sp. nov., an intertidal saprotroph from Phragmites australis inHong Kong. Bot Mar 41:379–382
Wong SW, Hyde KD, Jones EBG (1999) Ultrastructural studies of fresh-water ascomycetes Fluminicola bipolaris gen. et sp. nov. FungalDivers 2:189–197
Wong SS, Ngan AH, Riggs CM, Teng JL, Choi GK, Poon RW, Hui JJ,Low FJ, Luk A, Yuen KY (2011) Brittle tail syndrome is an emerg-ing infection in horses caused by a keratinolytic fungusEquicapillimyces hongkongensis gen. nov., sp. nov. Vet Microbiol155:399–408
Worapong J, Strobel G, Ford EJ, Li JY, Baird G, Hess WM (2001)Muscodor albus anam. gen. et sp nov., an endophyte fromCinnamomum zeylanicum. Mycotaxon 79:67–79
Wraight SP, Carruthers RI, Bradley CA, Jaronski ST, Lacey LA,Wood P,Galaini-Wraight S (1998) Pathogenicity of the entomopathogenicfungi Paecilomyces spp. and Beauveria bassiana against thesilverleaf whitefly, Bemisia argentifolii. J Invertebr Pathol 71:217–226
Xu J, Yang X, Lin Q (2014) Chemistry and biology of Pestalotiopsisderived natural products. Fungal Divers 66:37–68
Yang YL, Liu ZY, Cai L, Hyde KD, Yu ZN, Mckenzie EHC (2009)Colletotrichum anthracnose of Amaryllidaceae. Fungal Divers 39:123–146
YangHC, Haudenshield JS, HartmanGL (2014)Colletotrichum incanumsp. nov., a curved-conidial species causing soybean anthracnose inUSA. Mycologia 106(1):32–42
Yusoff M, Jones EBG, Moss ST (1994a) A taxonomic reappraisal of thegenus Ceriosporopsis based on ultrastructure. Can J Bot 72(10):1550–1559
Fungal Diversity

Yusoff M, Read SJ, Jones EBG, Moss ST (1994b) Ultrastructure ofAntennospora salina comb. nov. Mycol Res 98(9):997–1004
Zalar P, Gostinčar C, De Hoog GS, Uršič V, Sudhadham M, Gunde-Cimerman N (2008) Redefinition of Aureobasidium pullulans andits varieties. Stud Mycol 61:21–38
Zare R, Gams W (2001a) A revision of Verticillium sectionProstrata. VI. The genus Haptocillium. Nova Hedwigia73(3–4):271–292
Zare R, GamsW (2001b) A revision of Verticillium section Prostrata. IV.The genera Lecanicillium and Simplicillium gen. nov. NovaHedwigia 73(1–2):1–50
Zare R, Gams W, Starink-Willemse M, Summerbell RC (2007)Gibellulopsis, a suitable genus for Verticillium nigrescens, and
Musicillium, a new genus for V. theobromae. Nova Hedwigia85(3–4):463–489
Zelski SE, Raja HA, Miller AN, Shearer CA (2011a) Chaetorostrumquincemilensis, gen. et sp. nov., a new freshwater ascomycete andits Taeniolella-like anamorph from Peru. Mycosphere 2(5):593–600
Zelski SE, Raja HA, Miller AN, Barbosa FR, Gusmão LFP, Shearer CA(2011b) Longicollum biappendiculatum gen. et sp. nov., a new fresh-water ascomycete from the Neotropics. Mycosphere 2(5):539–545
Zhang N, Castlebury LA, Miller AN, Huhndorf SM, Schoch CL, SeifertKA, Rossman AY, Rogers JD, Kohlmeyer J, Volkmann-KohlmeyerB, Sung GH (2006) An overview of the systematics of theSordariomycetes based on four-gene phylogeny. Mycologia 98:1077–1088
Fungal Diversity
Copyright © 2022 FDOKUMEN




















