
Tissue-Specific Disruption of Rhythmic Expression of Dec1 and Dec2 in Clock Mutant Mice
16
http://jbr.sagepub.com/ Journal of Biological Rhythms http://jbr.sagepub.com/content/20/5/404 The online version of this article can be found at: DOI: 10.1177/0748730405280195 2005 20: 404 J Biol Rhythms Mitsuhide Noshiro, Masae Furukawa, Sato Honma, Takeshi Kawamoto, Taizo Hamada, Ken-ichi Honma and Yukio Kato Tissue-Specific Disruption of Rhythmic Expression of Dec1 and Dec2 in Clock Mutant Mice Published by: http://www.sagepublications.com On behalf of: Society for Research on Biological Rhythms can be found at: Journal of Biological Rhythms Additional services and information for http://jbr.sagepub.com/cgi/alerts Email Alerts: http://jbr.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: http://jbr.sagepub.com/content/20/5/404.refs.html Citations: What is This? - Nov 2, 2005 Version of Record >> at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from at Presbyterian Univ of East on October 11, 2013 jbr.sagepub.com Downloaded from
-
Upload
hiroshima-u -
Category
Documents
-
view
1 -
download
0
Transcript of Tissue-Specific Disruption of Rhythmic Expression of Dec1 and Dec2 in Clock Mutant Mice

http://jbr.sagepub.com/Journal of Biological Rhythms
http://jbr.sagepub.com/content/20/5/404The online version of this article can be found at:
DOI: 10.1177/0748730405280195
2005 20: 404J Biol RhythmsMitsuhide Noshiro, Masae Furukawa, Sato Honma, Takeshi Kawamoto, Taizo Hamada, Ken-ichi Honma and Yukio Kato
Tissue-Specific Disruption of Rhythmic Expression of Dec1 and Dec2 in Clock Mutant Mice
Published by:
http://www.sagepublications.com
On behalf of:
Society for Research on Biological Rhythms
can be found at:Journal of Biological RhythmsAdditional services and information for
http://jbr.sagepub.com/cgi/alertsEmail Alerts:
http://jbr.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://jbr.sagepub.com/content/20/5/404.refs.htmlCitations:
What is This?
- Nov 2, 2005Version of Record >>
at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from at Presbyterian Univ of East on October 11, 2013jbr.sagepub.comDownloaded from

10.1177/0748730405280195 ARTICLEJOURNALOF BIOLOGICALRHYTHMS / October 2005Noshiro et al. / DEC1 AND DEC2 EXPRESSION IN MICE
Tissue-Specific Disruption of Rhythmic Expressionof Dec1 and Dec2 in Clock Mutant Mice
Mitsuhide Noshiro,*,1,2 Masae Furukawa,†,2 Sato Honma,‡ Takeshi Kawamoto,*
Taizo Hamada,† Ken-ichi Honma,‡ and Yukio Kato*
*Department of Dental and Medical Biochemistry and †Department of Prosthetic Dentistry,Hiroshima University Graduate School of Biomedical Sciences, Hiroshima, Japan,
‡Department of Physiology, Hokkaido University Graduate School of Medicine, Sapporo, Japan
Abstract DEC1 and DEC2—basic helix-loop-helix transcription factors—exhibit a circadian expression in the suprachiasmatic nucleus and other periph-eral tissues and seem to play roles in regulating the mammalian circadianrhythm by suppressing the CLOCK/BMAL1-activated promoters of Per1, Dec1,and Dec2. The authors present data on the expression patterns of mRNAfor Dec1,Dec2, Per2, Dbp, and Npas2 in various tissues of wild-type and homozygous Clockmutant mice (Clock/Clock). The Clock mutation resulted in extreme reduction ofDec1 expression in kidney, heart, and skeletal muscle but not in liver, whereas itstrongly repressed Dec2 expression in liver, kidney, and heart, while Dec2 expres-sion in skeletal muscle remained rhythmic. Per2 also showed the tissue-dependentdisruption of the rhythmicity by Clock mutation, whereas rhythmic expression ofDbp in Clock mutant mice disappeared in all tissues examined. Npas2, a structur-ally and functionally related gene to Clock, showed significant levels of expres-sion in the liver and kidney with a robust rhythmicity, which was also affected byClock mutation. These marked changes in the Dec1 and Dec2 expression, as wellas in the Per2, Dbp, and Npas2 expression in the periphery by Clock mutation,indicated that CLOCK plays a major role in the expression of these genes in mosttissues. However, circadian expression of Dec1 in liver and kidney and that ofDec2 in skeletal muscle of Clock mutant mice suggested that CLOCK-independ-ent circadian regulation operates in some tissues.
Key words circadian rhythm, bHLH transcription factor, Dec1, Dec2, Per2, Dbp, Npas2,Clock mutant mice
DEC1 (BHLHB2/Stra13/Sharp2) and DEC2(BHLHB3/Sharp1)—basic helix-loop-helix (bHLH)transcription factors—have been shown to beinvolved in chondrogenesis (Shen et al., 1997; Shenet al., 2002), neurogenesis (Boudjelal et al., 1997;Rossner et al., 1997), cell growth arrest (Sun andTaneja, 2000), carcinogenesis, and responses tohypoxia (Ivanova et al., 2001; Miyazaki et al., 2002).
Stra13 null mice develop age-induced autoimmunityas a result of impaired T lymphocyte activation (Sunet al., 2001).
Recently, we found that in the SCN, the expressionof Dec1 and Dec2 exhibited a robust circadian rhythmwith a peak in the subjective day, so we speculatedthat Decs are involved in molecular feedback loopsgenerating the circadian rhythms (Honma et al., 2002).
404
1. To whom all correspondence should be addressed: Mitsuhide Noshiro, Department of Dental and Medical Biochemistry,Hiroshima University Graduate School of Biomedical Sciences, Kasumi 1-2-3, Hiroshima 734-8553, Japan; e-mail: [email protected]. These authors contributed equally to this work.
JOURNAL OF BIOLOGICAL RHYTHMS, Vol. 20 No. 5, October 2005 404-418DOI: 10.1177/0748730405280195© 2005 Sage Publications

DEC1 and DEC2 regulate clock-controlled genes suchas Per1 by binding to the CACGTG E-box enhancer(Honma et al., 2002), which is responsible forCLOCK:BMAL1 binding. Furthermore, we demon-strated that the Dec1 and Dec2 genes are regulated bypositive components, such as CLOCK, BMAL1, andBMAL2, and the negative regulators PER1, PER2,CRY1, CRY2, DEC1, and DEC2 through E-box ele-ments (Hamaguchi et al., 2004; Kawamoto et al., 2004;Sato et al., 2004). Thus, multiple feedback loops inter-lock, which may be essential for the finer and morestabilized regulation of the circadian rhythm(Kawamoto et al., 2004; Roenneberg and Merrow,2003). In addition, our recent study (Noshiro et al.,2004) and a study of Dec1 knockout mice (Stra13–/–)(Grechez-Cassiau et al., 2004) indicated that DEC1and DEC2 work as circadian output genes.
The Clock mutant mouse (Clock/Clock) is a usefulanimal model to examine potential up-regulation ofgenes by CLOCK. Homozygous mutant mice exhibitlengthened free-running periods of approximately 27h in wheel running, and they eventually becomearrhythmic in constant darkness (Oishi et al., 2002;Vitaterna et al., 1994). Mutant CLOCK protein lacks51-amino acids because of the deletion of exon 19 andbecomes inactive (Gekakis et al., 1998; King et al.,1997). The expression profiles of all clock genes aredisrupted with reduced peak expression in the SCN ofClock/Clock mice (Jin et al., 1999; Kume et al., 1999;Oishi et al., 2000), and we recently reported that theexpression of Dec1 and Dec2 in the SCN is also dis-rupted (Butler et al., 2004). These findings suggest thatCLOCK activates Dec1 and Dec2 transcription in vivo.
The existence of peripheral clocks has been shownin the liver, kidney, heart, skeletal muscle, and otherorgans, where they serve as endogenous oscillators toregulate the peripheral circadian rhythms—entrainingto various environmental factors such as feedingtime—and are synchronized with the master clock(Panda et al., 2002; Sakamoto et al., 1998; Storch et al.,2002). Circadian expression of clock and clock-controlled genes has been reported in various periph-eral tissues (Oishi et al., 2003; Panda et al., 2002; Uedaet al., 2002). Moreover, our recent study showed thatDec1 and Dec2 are expressed ubiquitously at variousextents with day-night variation (Noshiro et al., 2004).To test the hypothesis that rhythmic expression ofDec1 and Dec2 also requires functional CLOCK in theperiphery, we examined the expression of Dec1, Dec2,and other clock genes in several peripheral tissues of
Clock/Clock and wild-type mice: we found thatperipheral expression profiles of Dec1 and Dec2 wereinfluenced by Clock mutation and became arrhythmicin some, but not all, tissues of Clock/Clock mice. Theseobservations suggest that the peripheral clocks aredriven not only by the CLOCK-dependent mecha-nism but also by some other tissue-dependent mecha-nism(s).
MATERIALS AND METHODS
Animals and Isolation of RNA
A breeding colony of Clock mutant mice with aBALB/c background has been developed using miceoriginally supplied by Dr. J. S. Takahashi (Northwest-ern University). Genotypes were determined for eachindividual as described previously (Nakamura et al.,2002). Both Clock mutant and wild-type mice werehoused in clear polycarbonate cages in a 12L:12Dlight-dark cycle (lights on at 0600 h) with ad libitumaccess to food and water. All procedures were per-formed in compliance with principles and guidelinesfor animal research established by the local care anduse committee: “Guide for the Care and Use of Labo-ratory Animals, Hokkaido University GraduateSchool of Medicine.”
Three to 4 mice for each time point were decapi-tated at 4-h intervals beginning at ZT 2 (0800 h), eitherin a normal LD cycle or on the 1st day in DD (i.e.,beginning 14 h after the last lights-off). Adim red light(< 0.1 lx) was placed in the room to allow surgeriesduring the dark periods. The liver, kidney, heart, andskeletal muscle were quickly removed, frozen on dryice, and stored at –80 °C until processing. Total RNAwas extracted by Trizol reagent (Invitrogen). All RNApreparations were obtained separately for individualmice and used for the determination of transcript lev-els of respective genes.
Northern Blot Analysis
Total RNA (10 µg/lane) preparations wereelectrophoresed on 1% agarose gels containing 2.2 Mformaldehyde as described by Thomas (1980) andtransferred to Nytran membranes (Schleicher &Schuell GmbH). The membranes were hybridizedwith the 32P-labeled cDNA probes in hybridizationsolution containing 6 × SSC, 5 × Denhardt’s solution,
Noshiro et al. / DEC1 AND DEC2 EXPRESSION IN MICE 405

10 mM ethylenediaminetetraacetic acid (EDTA), 1%sodium dodecyl sulfate (SDS), and 0.5 mg/mLsonicated salmon sperm DNA at 68 °C overnight.DNA probes were labeled using α-[32P]dCTP (111TBq/mmol) (Du Pont-New England Nuclear) and arandom primer DNA labeling kit (Takara Co., Tokyo,Japan). The membranes were washed with 0.1 × SSCcontaining 0.5% SDS at 50 °C and exposed to KodakBMX films at –80 °C. Radioactivity of the hybridizedareas was quantified using a Bio-imaging AnalyzerSystem BAS2000 (Fuji Photo Film Co. Ltd., Tokyo,Japan). Mouse cDNA clones used as hybridizationprobes for Dec1 and Dec2 were prepared as describedpreviously (Fujimoto et al., 2001; Shen et al., 1997).
Real-Time Quantitative RT-PCR Analysis
Real-time quantitative RT-PCR analysis was per-formed using an ABI PRISM 7900 Sequence DetectionSystem instrument and software (AppliedBiosystems). The principles and protocols for RT-PCRanalyses using this system have been described previ-ously (Gibson et al., 1996). The 1st-strand cDNA wassynthesized using a ReverTra Ace reverse transcrip-
tase kit (Toyobo Co., Osaka, Japan) with total RNA (1µg) preparation. The sequences for the primers andTaqMan™ fluorogenic probes used in these analysesare listed in Table 1. Aliquots of cDNA preparationscorresponding to 20 ng of initial RNA sample weresubjected to amplification. The amplification of theoriginal RNA sample before RT reaction for all genesindicated that the contribution of genomic DNA tofinal amounts of PCR products was negligible (datanot shown). Authentic cDNA standard for all genes ata range of 8 orders of magnitude showed linearity ofamplification to initial amounts of cDNA and wasused to standardize the transcript levels of respectivegenes. Glyceraldehyde-3-phosphate dehydrogenase(GAPDH) was chosen as an internal standard to con-trol for variability in amplification (data not shown).All assays were repeated more than 2 times and gavereproducible results.
Statistical Analysis
Time-series data were analyzed by 1-way analysisof variance (1-way ANOVA) for rhythm. Differencesbetween 2 genotypes or 2 light conditions were tested
406 JOURNAL OF BIOLOGICAL RHYTHMS / October 2005
Table 1. Primers and Probes
Genes Sequences Accession Number
Dec1 NM_011498Forward primer 5′-ATCAGCCTCCTTTTTGCCTTC-3′Reverse primer 5′-AGCATTTCTCCAGCATAGGCAG-3′Probe 5′-FAM-TTCTATCTCATCCCACCATCGGCCAC-TAMRA-3′
Dec2 NM_024469Forward primer 5′-ATTGCTTTACAGAATGGGGAGCG-3′Reverse primer 5′-AAAGCGCGCGAGGTATTGCAAGAC-3′Probe 5′-FAM-CGACTTGGATGCGTTCCACTCGG-TAMRA-3′
Per2 NM_011066Forward primer 5′-CCAGAGGAACTAGCCTATAAGGAACCA-3′Reverse primer 5′-GAACTCGCACTTCCTTTTCAGG-3′Probe 5′-FAM-ATCAGCTGCCTGGACAGTGTCATCAGGTAC-TAMRA-3′
Dbp NM_016974Forward primer 5′-AGGAACTGAAGCCTCAACCAATC-3′Reverse primer 5′-CTCCGGCTCCAGTACTTCTCAT-3′Probe 5′-FAM-TGAAGAAGGCAAGGAAAGTCCAGGTGC-TAMRA-3′
Npas2 U77969Forward primer 5′-GGCATTAGATGGCTTCGTCATC-3′Reverse primer 5′-AAGGAGAGGTGTGATACTGTCGG-3′Probe 5′-FAM-TGACAACAGACGGCAGCATCATCTATGTG-TAMRA-3′
Clock NM_007715Forward primer 5′-CGGCGAGAACTTGGCATT-3′Reverse primer 5′-ATACGATTGTCAGACCCAGAATCTT-3′Probe 5′-FAM-CTCTTCCTGAGACAGCTGCTGACAAAAGC-TAMRA-3′

by 2-way ANOVA. Comparisons with p < 0.05 weretaken as significant.
RESULTS
Effects of the Clock Mutationon Gene Expression in the Liver
Transcripts of Dec1, Dec2, Per2, and Dbp in wild-type and Clock mutant mice were examined by quanti-tative real-time RT-PCR. In wild-type mice, hepaticexpression of Dec1 mRNA showed a circadian rhythmwith peaks at ZT 6 and CT 10 in LD and DD, respec-tively (Fig. 1A) (1-way ANOVA, p < 0.0001 in LD andDD). Differences in the patterns between the 2 lightconditions (LD and DD) were significant (2-wayANOVA, p < 0.01). In Clock/Clock mice, peaks weredelayed by 8 h in both LD and DD compared with therespective peaks in wild-type mice, and their patternsmaintained a significant rhythmicity (1-way ANOVA,p < 0.0001 in LD, p < 0.01 in DD). Differences in the pat-terns between genotypes were significant in both LDand DD (2-way ANOVA, p < 0.05), and the Northernblot analysis confirmed these results (Fig. 1A, bot-tom). To examine whole-day expression level of Dec1,we calculated the area under the curve (AUC) of eachtime series: relative AUC values of hepatic Dec1expression in wild-type mice in LD, wild-type mice inDD, Clock/Clock mice in LD, and Clock/Clock mice inDD were 100, 72, 80, and 51, respectively.
Dec2 mRNA expression in the liver of wild-typemice showed a robust circadian rhythm with peaks atZT 6 and CT 6 in LD and DD, respectively (Fig. 1B) (1-way ANOVA, p < 0.0001 in LD and p < 0.01 in DD). Dif-ferences in the patterns between the 2 light conditionswere not significant. In Clock/Clock mice, the expres-sion of Dec2 mRNAwas depressed in both LD and DD(1-way ANOVA, not significant; 2-way ANOVA, p <0.05 in LD and p < 0.005 in DD). Northern blot analysisconfirmed these results (Fig. 1B, bottom). RelativeAUC values of hepatic Dec2 expression in wild-typemice in LD, wild-type mice in DD, Clock/Clock mice inLD, and Clock/Clock mice in DD were 100, 56, 3, and 2,respectively.
For comparison, we also examined the effects ofClock mutation on hepatic expression levels of Per2and Dbp. Hepatic expression of Per2 mRNA in wild-type mice exhibited a robust rhythm with peaks at ZT
10 and CT 10 in LD and DD, respectively (Fig. 1C) (1-way ANOVA, p < 0.0001 in LD and p < 0.01 in DD)(Panda et al., 2002; Ueda et al., 2002), and the levels inLD and DD were approximately equal. In the liver ofClock/Clock mice, the peaks were delayed, and the lev-els decreased in LD and DD (2-way ANOVA, p < 0.05in LD and DD), but a significant rhythmicity remained(1-way ANOVA, p < 0.05 in LD and DD). RelativeAUC values of hepatic Per2 expression in wild-typemice in LD, wild-type mice in DD, Clock/Clock mice inLD, and Clock/Clock mice in DD were 100, 100, 73, and67, respectively.
Wild-type mice showed a robust Dbp rhythm in theliver with peaks at ZT 6 and CT 6-10 in both light con-ditions (Fig. 1D) (1-way ANOVA, p < 0.0001 in LD andDD), as previously reported (Oishi et al., 2003). Dbpexpression was markedly repressed in Clock/Clockmice in both LD and DD, indicating the strongdependency of Dbp expression on the CLOCK func-tion (Oishi et al., 2003; Ripperger et al., 2000). How-ever, the repressed levels of Dbp mRNAin DD showedsignificant rhythmicity (1-way ANOVA, p < 0.05), asshown in the inset of Figure 1D. Relative AUC valuesof hepatic Dbp expression in wild-type mice in LD,wild-type mice in DD, Clock/Clock mice in LD, andClock/Clock mice in DD were 100, 131, 2, and 4,respectively.
Effects of the Clock Mutationon Gene Expression in the Kidney
In the kidney of wild-type mice (Fig. 2A), Dec1mRNA expression in LD and DD showed a circadianrhythm with peaks at ZT 10 and CT 10-14, respectively(1-way ANOVA, p < 0.0001 in LD and p < 0.05 in DD),and differences in the patterns between the 2 light con-ditions were significant (2-way ANOVA, p < 0.05). Inthe kidney of Clock/Clock mice, Dec1 mRNAlevels sig-nificantly decreased (2-way ANOVA, p < 0.01), but theprofiles maintained a rhythmicity in both LD and DD(1-way ANOVA, p < 0.05 in LD and p < 0.0001 in DD).Northern blot analysis confirmed these results (Fig.2A, bottom). Relative AUC values of renal Dec1expression in wild-type mice in LD, wild-type mice inDD, Clock/Clock mice in LD, and Clock/Clock mice inDD were 100, 84, 50, and 58, respectively.
Dec2 mRNA levels in LD and DD in the kidney ofwild-type mice showed a circadian rhythm with peaksat ZT 10 and CT 6-10 (Fig. 2B) (1-way ANOVA, p < 0.05
Noshiro et al. / DEC1 AND DEC2 EXPRESSION IN MICE 407

in LD and p < 0.01 in DD). Differences in the patternsbetween the 2 light conditions were significant (2-wayANOVA, p < 0.01). In the kidney of Clock/Clock mice,Dec2 mRNA levels were markedly repressed (2-way
ANOVA, p < 0.05 in LD and p < 0.01 in DD), andrhythmicity was lost in both light conditions. North-ern blot analysis confirmed these results (Fig. 2B, bot-tom). Relative AUC values of renal Dec2 expression in
408 JOURNAL OF BIOLOGICAL RHYTHMS / October 2005
Figure 1. Gene expressions in the liver of wild-type and Clock/Clock mice. Transcript levels of Dec1 (A), Dec2 (B), Per2 (C), and Dbp (D)were determined by real time RT-PCR for 4 individual mice for each time point. Dbp expression patterns of Clock/Clock mice are alsoshown in the inset of D with an expanded ordinate, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) levels were determined asan internal standard to control for variability in amplification (data not shown). Data are shown as mean ± SEM (n = 4). Gray bars indicatelights-on and lights-off, as well as subjective daytime for LD and DD, respectively. Closed bars indicate dark phases. All values for thesame gene species in Figures 1 to 4 can be quantitatively compared because the data are standardized by the use of mouse cDNA samplesfor the respective genes. The levels of mRNA for DEC1 and DEC2 were also determined by Northern blot analysis using the same RNAsamples of individual mice. Methylene blue staining of the blotting confirmed the equal loading of the RNA sample on each lane (data notshown). Representative sets of film images of Northern blotting are shown at the bottom of panels A and B. The sizes of the hybridizedmRNA are shown on the right side of the respective panels. Time-series data were analyzed by 1-way ANOVA for rhythmicity (*p < 0.05; **p <0.01; ***p < 0.0001; ns, not significant). Differences between 2 genotypes or between 2 light conditions in wild-type mice were tested by 2-way ANOVA (*p < 0.05; **p < 0.01; ***p < 0.0001; ns, not significant). WT, wild-type mice; Ck/Ck, Clock mutant mice.
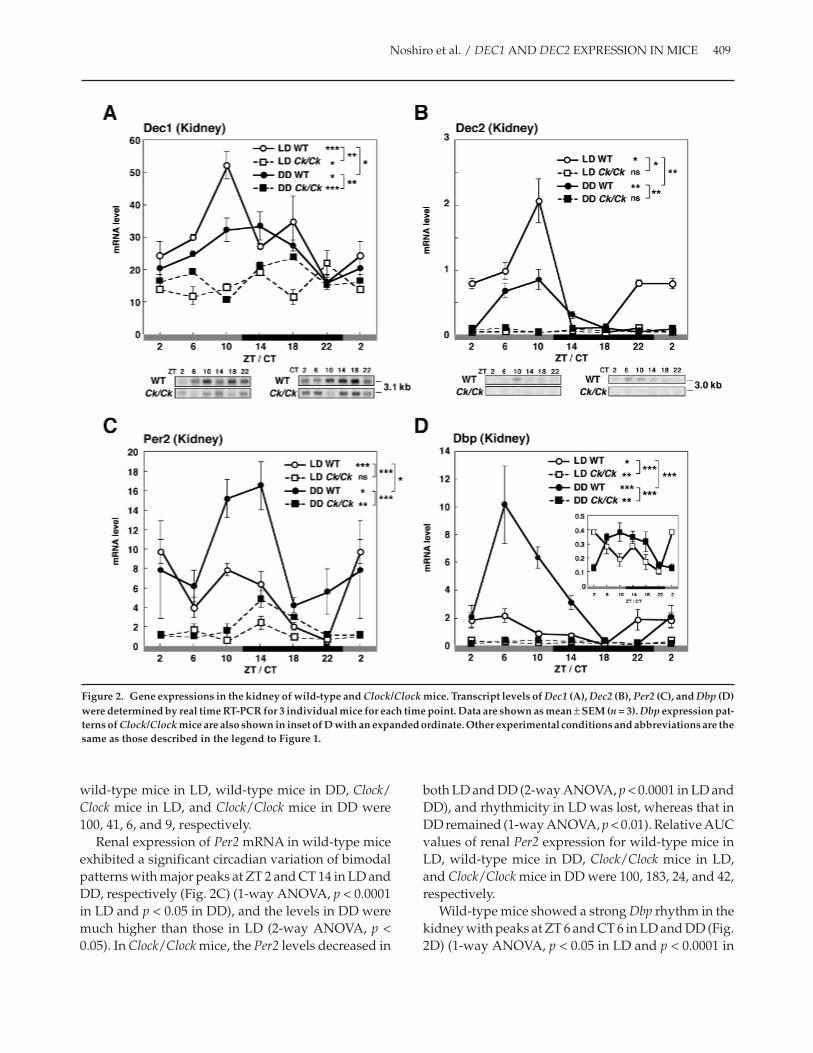
wild-type mice in LD, wild-type mice in DD, Clock/Clock mice in LD, and Clock/Clock mice in DD were100, 41, 6, and 9, respectively.
Renal expression of Per2 mRNA in wild-type miceexhibited a significant circadian variation of bimodalpatterns with major peaks at ZT 2 and CT 14 in LD andDD, respectively (Fig. 2C) (1-way ANOVA, p < 0.0001in LD and p < 0.05 in DD), and the levels in DD weremuch higher than those in LD (2-way ANOVA, p <0.05). In Clock/Clock mice, the Per2 levels decreased in
both LD and DD (2-way ANOVA, p < 0.0001 in LD andDD), and rhythmicity in LD was lost, whereas that inDD remained (1-way ANOVA, p < 0.01). Relative AUCvalues of renal Per2 expression for wild-type mice inLD, wild-type mice in DD, Clock/Clock mice in LD,and Clock/Clock mice in DD were 100, 183, 24, and 42,respectively.
Wild-type mice showed a strong Dbp rhythm in thekidney with peaks at ZT 6 and CT 6 in LD and DD (Fig.2D) (1-way ANOVA, p < 0.05 in LD and p < 0.0001 in
Noshiro et al. / DEC1 AND DEC2 EXPRESSION IN MICE 409
Figure 2. Gene expressions in the kidney of wild-type and Clock/Clock mice. Transcript levels of Dec1 (A), Dec2 (B), Per2 (C), and Dbp (D)were determined by real time RT-PCR for 3 individual mice for each time point. Data are shown as mean ± SEM (n = 3). Dbp expression pat-terns of Clock/Clock mice are also shown in inset of D with an expanded ordinate. Other experimental conditions and abbreviations are thesame as those described in the legend to Figure 1.
DD), respectively, and the levels in DD were muchhigher than those in LD (2-way ANOVA, p < 0.0001).Dbp expression in the kidney of Clock/Clock mice wasalso markedly repressed but still rhythmic in both LDand DD (1-way ANOVA, p < 0.01 in LD and DD), asshown in the inset of Figure 2D. Relative AUC valuesof renal Dbp expression for wild-type mice in LD,wild-type mice in DD, Clock/Clock mice in LD, andClock/Clock mice in DD were 100, 293, 19, and 22,respectively.
Effects of the Clock Mutationon Gene Expression in Skeletal Muscle
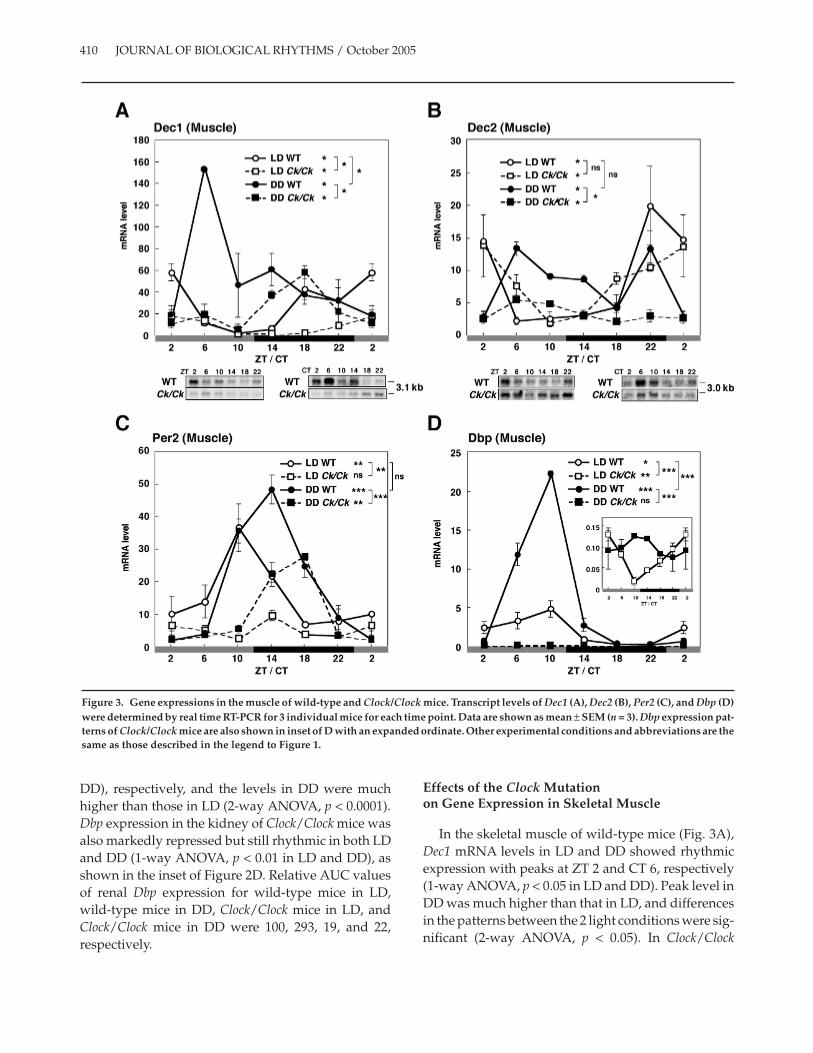
In the skeletal muscle of wild-type mice (Fig. 3A),Dec1 mRNA levels in LD and DD showed rhythmicexpression with peaks at ZT 2 and CT 6, respectively(1-way ANOVA, p < 0.05 in LD and DD). Peak level inDD was much higher than that in LD, and differencesin the patterns between the 2 light conditions were sig-nificant (2-way ANOVA, p < 0.05). In Clock/Clock
410 JOURNAL OF BIOLOGICAL RHYTHMS / October 2005
Figure 3. Gene expressions in the muscle of wild-type and Clock/Clock mice. Transcript levels of Dec1 (A), Dec2 (B), Per2 (C), and Dbp (D)were determined by real time RT-PCR for 3 individual mice for each time point. Data are shown as mean ± SEM (n = 3). Dbp expression pat-terns of Clock/Clock mice are also shown in inset of D with an expanded ordinate. Other experimental conditions and abbreviations are thesame as those described in the legend to Figure 1.

mice, Dec1 mRNA levels decreased (2-way ANOVA,p < 0.05) with a significant rhythmicity in both lightconditions (1-way ANOVA, p < 0.05 in LD and DD).Northern blot analysis confirmed these results (Fig.3A, bottom). Relative AUC values of Dec1 expressionin wild-type mice in LD, wild-type mice in DD, Clock/Clock mice in LD, and Clock/Clock mice in DD were100, 227, 30, and 100, respectively.
Dec2 mRNA levels in the skeletal muscle of wild-type mice (Fig. 3B) showed a peak at ZT 22 in LD and 2peaks at CT 6 and CT 22 in DD (1-way ANOVA, p <0.05 in LD and DD). There was no significant differ-ence between genotypes in Dec2 mRNA expression inLD, and the rhythmicity remained in LD (1-wayANOVA, p < 0.05 in Clock/Clock mice), whereas Dec2mRNA levels significantly decreased in DD Clock/Clock mice (1-way ANOVA, p < 0.05), with a weakrhythmicity (1-way ANOVA, p < 0.05). Northern blotanalysis confirmed these results (Fig. 3B, bottom). Rel-ative AUC values of Dec2 expression in wild-typemice in LD, wild-type mice in DD, Clock/Clock mice inLD, and Clock/Clock mice in DD were 100, 110, 97, and45, respectively, which clearly indicated that the effectof Clock mutation on whole-day expression of Dec2 inthis tissue was much less than that in the other tissues.
The expression of Per2 mRNA in the skeletal mus-cle of wild-type mice exhibited a strong rhythm withpeaks at ZT 10 and CT 14 in LD and DD, respectively(Fig. 3C) (1-way ANOVA, p < 0.01 in LD and p < 0.0001in DD). In Clock/Clock mice, the peaks were delayed,and the levels decreased in both LD and DD (2-wayANOVA, p < 0.01 and p < 0.0001 for LD and DD,respectively). Rhythmicity was disrupted in LD,whereas a significant rhythmicity (1-way ANOVA, p <0.05) remained in DD. Relative AUC values of Per2expression in wild-type mice in LD, wild-type mice inDD, Clock/Clock mice in LD, and Clock/Clock mice inDD were 100, 128, 32, and 66, respectively.
Wild-type mice showed a robust Dbp rhythm in themuscle with peaks at ZT/CT 10 in LD and DD (Fig.3D) (1-way ANOVA, p < 0.05 in LD and p < 0.0001 inDD). Dbp levels in DD were much higher than those inLD (2-way ANOVA, p < 0.0001). Dbp expression in themuscle of Clock/Clock mice was markedly repressed inboth light conditions, indicating the strong depend-ency of muscular Dbp expression on the CLOCK func-tion, but the repressed levels of Dbp mRNA in LDshowed significant rhythmicity (1-way ANOVA, p <0.01 in LD), as shown in the inset of Figure 3D. RelativeAUC values of Dbp expression in wild-type mice in
LD, wild-type mice in DD, Clock/Clock mice in LD,and Clock/Clock mice in DD were 100, 320, 4, and 5,respectively.
Effects of the Clock Mutationon Gene Expression in the Heart
In the heart of wild-type mice (Fig. 4A), Dec1mRNA levels in LD and DD showed a circadianrhythm with peaks at ZT 10 and CT 6, respectively (1-way ANOVA, p < 0.05 in LD and p < 0.01 in DD). Dec1mRNA levels significantly decreased in the heart ofClock/Clock mice (2-way ANOVA, p < 0.0001), andrhythmicity was lost in both light conditions. RelativeAUC values of Dec1 expression in wild-type mice inLD, wild-type mice in DD, Clock/Clock mice in LD,and Clock/Clock mice in DD were 100, 113, 36, and 48,respectively.
Dec2 mRNA levels in the heart of wild-type miceshowed a circadian rhythm with peaks at ZT 2 and CT6 in LD and DD, respectively (Fig. 4B) (1-wayANOVA, p < 0.05 in LD and DD). Dec2 mRNA levelssignificantly decreased in the heart of Clock/Clockmice (2-way ANOVA, p < 0.05 in LD and p < 0.01 inDD), and rhythmicity was lost in both light condi-tions. We also noted that maximum expression levelsof Dec2 in the skeletal muscle and heart were muchhigher than those in the liver and kidney, as was truein the case of rats (Noshiro et al., 2004). Relative AUCvalues of Dec2 expression in wild-type mice in LD,wild-type mice in DD, Clock/Clock mice in LD, andClock/Clock mice in DD were 100, 116, 48, and 52,respectively.
Expression of Per2 mRNA in the heart of wild-typemice exhibited a strong rhythm with peaks at ZT/CT10 in LD and DD (Fig. 4C) (1-way ANOVA, p < 0.05 inLD and p < 0.01 in DD). In the heart of Clock/Clockmice, the peaks shifted and the levels decreased inboth LD and DD (2-way ANOVA, p < 0.0001 in LD andDD). Rhythmicity was disrupted in LD but not in DD.Relative AUC values of Per2 expression in wild-typemice in LD, wild-type mice in DD, Clock/Clock mice inLD, and Clock/Clock mice in DD were 100, 67, 20, and27, respectively.
Wild-type mice showed a robust Dbp rhythm in theheart with peaks at ZT 6 and CT 10 in LD and DD,respectively (Fig. 4D) (1-way ANOVA, p < 0.05 in LDand p < 0.01 in DD). Dbp levels in DD were muchhigher than those in LD (2-way ANOVA, p < 0.0001).Rhythmic expression of Dbp in the heart of Clock/Clock
Noshiro et al. / DEC1 AND DEC2 EXPRESSION IN MICE 411
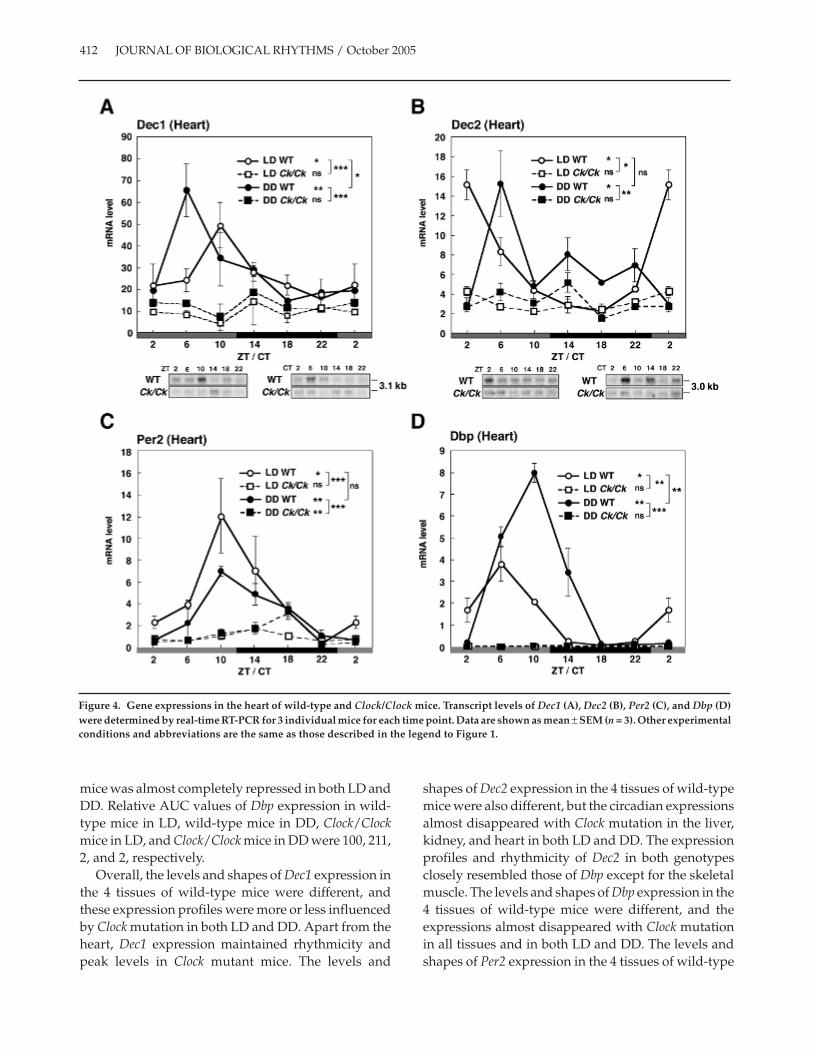
mice was almost completely repressed in both LD andDD. Relative AUC values of Dbp expression in wild-type mice in LD, wild-type mice in DD, Clock/Clockmice in LD, and Clock/Clock mice in DD were 100, 211,2, and 2, respectively.
Overall, the levels and shapes of Dec1 expression inthe 4 tissues of wild-type mice were different, andthese expression profiles were more or less influencedby Clock mutation in both LD and DD. Apart from theheart, Dec1 expression maintained rhythmicity andpeak levels in Clock mutant mice. The levels and
shapes of Dec2 expression in the 4 tissues of wild-typemice were also different, but the circadian expressionsalmost disappeared with Clock mutation in the liver,kidney, and heart in both LD and DD. The expressionprofiles and rhythmicity of Dec2 in both genotypesclosely resembled those of Dbp except for the skeletalmuscle. The levels and shapes of Dbp expression in the4 tissues of wild-type mice were different, and theexpressions almost disappeared with Clock mutationin all tissues and in both LD and DD. The levels andshapes of Per2 expression in the 4 tissues of wild-type
412 JOURNAL OF BIOLOGICAL RHYTHMS / October 2005
Figure 4. Gene expressions in the heart of wild-type and Clock/Clock mice. Transcript levels of Dec1 (A), Dec2 (B), Per2 (C), and Dbp (D)were determined by real-time RT-PCR for 3 individual mice for each time point. Data are shown as mean ± SEM (n = 3). Other experimentalconditions and abbreviations are the same as those described in the legend to Figure 1.

mice were also different, and the expression profileswere influenced by Clock mutation to various degreesin all tissues and in both LD and DD. The effects ofClock mutation on Per2 were similar to those of Dec1.We also determined Per1 expression levels in the 4 tis-sues. The effects of Clock mutation on Per1 expressionrhythms were basically similar to those on Per2 (datanot shown), which was in accordance with the previ-ous findings on their hepatic expression in Clockmutant mice (Oishi et al., 2003).
Npas2 mRNA Levels in thePeripheral Tissues of Clock/Clock Mice
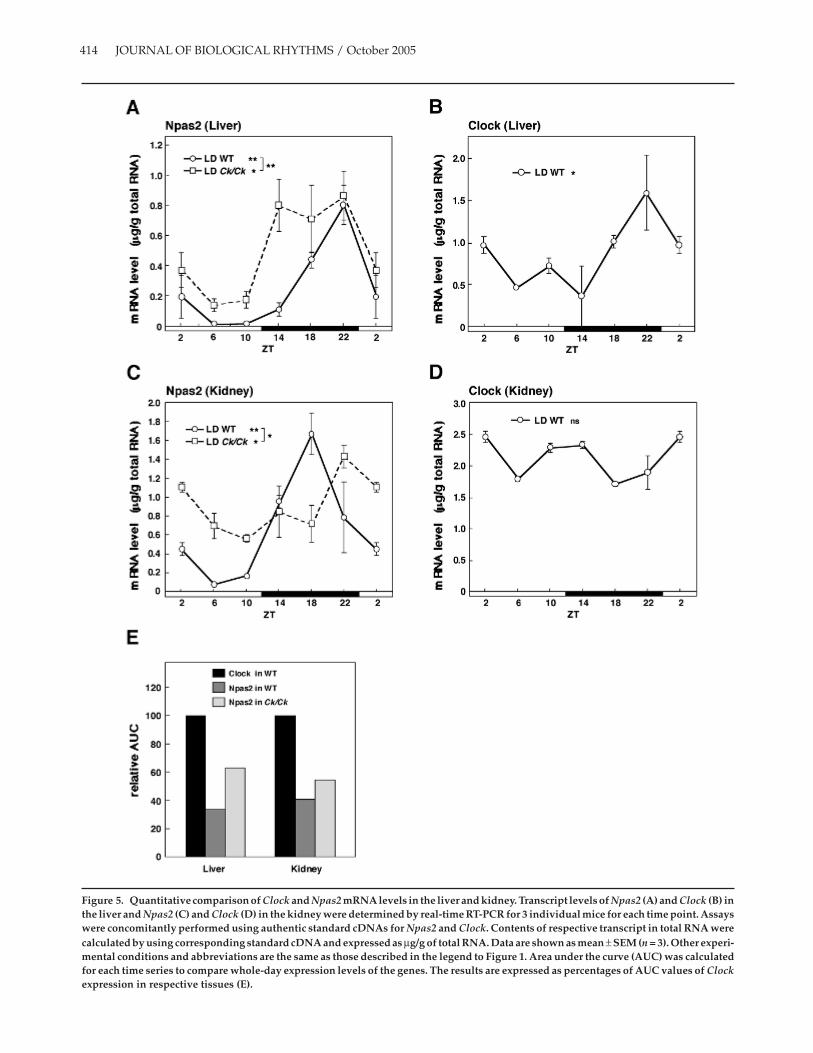
To examine the effects of Clock mutation on theNpas2 expression, we determined the Npas2 mRNAlevel by quantitative real-time RT-PCR using theauthentic standard and compared with that of ClockmRNA. In wild-type mice, hepatic expression ofNpas2 mRNA showed a strong circadian rhythm withpeaks at ZT 22 in LD (Fig. 5A) (1-way ANOVA, p <0.001). Clock/Clock mice also exhibited a robust Npas2rhythm with a broad peak between ZT 14 and 22 (1-way ANOVA, p < 0.05). The difference in the patternsbetween genotypes was significant (2-way ANOVA,p < 0.001). Clock expression in the liver of wild-typemice in LD showed significant circadian rhythm (Fig.5B) (1-way ANOVA, p < 0.05), although with lessamplitude than that of Npas2. Renal expression ofNpas2 in wild-type mice also showed strong circadianrhythm with a peak at ZT 18, whereas Npas2 expres-sion in Clock mutant mice was still rhythmic (1-wayANOVA, p < 0.05) with a peak delayed by 4 h and lessamplitude compared with wild-type mice (Fig. 5C).The difference in the patterns between genotypes wassignificant (2-way ANOVA, p < 0.05). Clock expressionin the kidney of wild-type mice in LD showed no circa-dian rhythm (Fig. 5D). As the authentic standardcDNAs for Npas2 and Clock were concomitantlyassayed, we calculated the AUC for each expressionpattern to compare whole-day expression levels of the2 genes. Results are summarized in Figure 5E. AUC ofClock expression in respective tissue of wild-type micewas used as a control. Relative AUC values of Npas2expression in these tissues of wild-type mice weresimilar (32% and 41% of corresponding Clock expres-sion). Relative AUC values of Npas2 expressionincreased to 63% and 54% in the liver and kidney ofClock mutant mice.
DISCUSSION
Human and mouse promoters of Dec1 and Dec2contain 2 or 3 CACGTG E-box elements responsive tothe activation by CLOCK:BMAL1 heterodimer(Hamaguchi et al., 2004; Kawamoto et al., 2004;Teramoto et al., 2001). In accordance with these facts,the expressions of Dec1 and Dec2 were significantlyaffected by Clock mutation. Dec1 expression becamearrhythmic in the heart, whereas in the liver, kidney,and skeletal muscle, Dec1 expression remained rhyth-mic with phase-delay. The hepatic expression profileof Dec1 in Clock mutant mice was reported by Oishiet al. (2003) and is in accordance with our presentstudy. The expression of Dec2 was also suppressed inClock mutant mice tissue dependently: Dec2 expres-sion became arrhythmic in the liver, kidney, and heartbut maintained rhythmicity in the skeletal muscle.
In addition to Dec1 and Dec2, we examined theeffect of clock mutation on the circadian gene expres-sion profiles of other clock and clock-controlled genes,Per2, Dbp, and Npas2, in the 4 peripheral tissues underLD and DD conditions. We found that Clock mutationaffected the circadian expression patterns tissue spe-cifically and gene specifically. The differential effectssuggest tissue-specific molecular clock mechanismsin each peripheral clock, as well as differential controlof each peripheral clock by the central clock. Basically,Dec1 and Per2 expressions were resistant to the Clockmutation (exceptions were Dec1 in the heart and Per2in the kidney and heart). On the other hand, rhythmicexpressions of Dec2 and Dbp were severely repressedby clock mutation (except for Dec2 in the muscle). Reg-ulatory mechanisms other than transactivationthrough CLOCK:BMAL1 may explain the differencebetween the 2 groups. In addition to the control byCLOCK and BMAL1, Dec1 is regulated by various fac-tors, such as bone morphogenetic protein–2 (Shenet al., 2002), transforming growth factor–β (Shen et al.,2002), retinoic acid (Boudjelal et al., 1997), cyclic AMP(Shen et al., 2001; Shen et al., 1997), insulin (Shen et al.,2002; Yamada et al., 2003), serum starvation (Sun andTaneja, 2000), nerve growth factor, glutamatergic neu-ron stimulation via kainic acid injection (Rossner et al.,1997), hypoxia-inducible factor 1 (Miyazaki et al.,2002), and T cell activation (Sun et al., 2001), whereasPer1 is regulated by CREB, which is thought to triggerthe photic entrainment of the circadian clock(Tischkau et al., 2003; Travnickova-Bendova et al.,
Noshiro et al. / DEC1 AND DEC2 EXPRESSION IN MICE 413
414 JOURNAL OF BIOLOGICAL RHYTHMS / October 2005
Figure 5. Quantitative comparison of Clock and Npas2 mRNA levels in the liver and kidney. Transcript levels of Npas2 (A) and Clock (B) inthe liver and Npas2 (C) and Clock (D) in the kidney were determined by real-time RT-PCR for 3 individual mice for each time point. Assayswere concomitantly performed using authentic standard cDNAs for Npas2 and Clock. Contents of respective transcript in total RNA werecalculated by using corresponding standard cDNA and expressed as µg/g of total RNA. Data are shown as mean ± SEM (n = 3). Other experi-mental conditions and abbreviations are the same as those described in the legend to Figure 1. Area under the curve (AUC) was calculatedfor each time series to compare whole-day expression levels of the genes. The results are expressed as percentages of AUC values of Clockexpression in respective tissues (E).

2002). Tissue-specific expression of these regulatoryfactors may also contribute to the different expressionpatterns in different tissues. Complete SCN lesionresults in a total loss of circadian rhythms in bodilyfunctions, even though each peripheral clock can gen-erate self-sustaining gene expression rhythms, indi-cating that the central clock in the SCN coordinates thephase relationships among peripheral clocks by spe-cific phase-resetting signals, such as humoral andneural signals, or secondary to the behavioralchanges. Single peripheral tissue may have more than2 resetting signals, and the number and sensitivity ofthe receptor may also contribute to the susceptibility.
Most circadian gene expression rhythms thatremained in Clock mutant mice are phase-delayedcompared with those in wild-type mice, which can beexplained by the lengthened circadian period of Clockmutant mice (Daan and Aschoff, 2001; Pittendrighand Daan, 1976). Since Clock mutant mice still exhibitsignificant Per1 and Dec1 expression rhythms withdelayed phase in the SCN—although the level wasgreatly reduced—rhythms in the peripheral tissuesmay be due to a phase setting of the central clock.
In the present experiment, we measured geneexpression levels in the peripheral organs on the 1stday of DD to exclude the desynchronization foundamong Clock mutant mice under prolonged DD. Sincethe Clock mutant mice maintain behavioralrhythmicity on the 1st day of DD, rhythms in theperipheral organs may be secondary to the behavioralrhythms. Circadian signals induced by rhythmic feed-ing, hormonal, or autonomic nervous functions mayinduce expression rhythms of some, but not all, clockgene expression in the peripheral organs (Balsalobreet al., 2000; Damiola et al., 2000; Oishi et al., 2002;Shibata, 2004; Terazono et al., 2003). Therefore, therhythms we observed are not necessarily CLOCK-independent oscillatory mechanisms. Even so, therhythms induced in the peripheral organs areindependent of intact CLOCK.
NPAS2 is structurally related to CLOCK, binds toBMAL1, and can induce transcription of Per1, Per2,and Cry1 genes (Reick et al., 2001). We confirmed thatNPAS2:BMAL1 enhanced the expression from Dec1and Dec2 promoters using luciferase-reporter assays(T. Kawamoto, unpublished results). NPAS2 is abun-dantly expressed in the forebrain, whereas the expres-sion of Npas2 mRNA in several peripheral tissues,including the liver, kidney, and heart, was previouslyreported to be undetectable by Northern blot analysis(Zhou et al., 1997). However, we were able to find the
circadian expression of Npas2 mRNA in the liver andkidney, with Npas2 mRNA levels comparable to thoseof Clock based on AUC values. The expression profilesof Npas2 in these tissues were significantly affected,and their AUC values were increased by the Clockmutation. Expression levels of Npas2 comparable tothat of Clock and in vitro activation of Dec1 and Dec2promoters by NPAS2, together with BMAL1, suggestthat NPAS2 may replace CLOCK to some extent inClock mutant mice. Expressions of Bmal1 mRNA inthese tissues of Clock mutant mice became arrhythmic,exhibiting continuously high levels throughout theday as previously reported (data not shown) (Oishiet al., 2000). Only in the case of Npas2 did Clock muta-tion not repress but enhance the gene expression level,at least in the liver and kidney, suggesting that theremay be compensating mechanisms for Npas2 geneexpression. Yet, functionally, NPAS2 did not compen-sate for mutant CLOCK protein in Dec2 and Dbpexpression. Interference of dominant negativeCLOCK protein (Gekakis et al., 1998) with the inter-action of NPAS2 with BMAL1 may be a plausiblemechanism for these observations. A study on protein-protein interaction using NPAS2 together with nativeand dominant negative CLOCK proteins and BMAL1protein would clarify this discrepancy.
Significant dissociation of expression patterns andlevels between LD and DD of wild-type mice wasobserved for Dec1, Dbp, and Per2, which was probablydue to masking of light. Previously, we reported highexpression levels of Dec1 and Per1 at ZT 2, soon afterlights-on, in the extra SCN brain tissues (Butler et al.,2004). In the present experiment, we also observedincreased gene expression, which suggested the posi-tive masking of light (Dec1 and Dec2 in the liver andkidney). Interestingly, we also observed a significantincrease in AUC after exposure to DD, both in wild-type and Clock mutant mice (Dec1 in the muscle, Per2in the kidney and muscle, DBP in the kidney andheart), suggesting negative masking by light. Accord-ing to the available literature, light stimulation affectsvisceral functions via the SCN (Niijima et al., 1993),and sympathetic innervation was demonstrated forthe regulation of Per expression in the liver (Terazonoet al., 2003), which may indicate that the peripheralclock can be affected by light condition.
In our previous studies with rats (Noshiro et al.,2004), hepatic expression profiles of Dec1 and Dec2showed peaks during night (ZT 14 and ZT 10, respec-tively), whereas in the present study, their expressionsin mouse liver showed peaks during daytime (ZT 6).
Noshiro et al. / DEC1 AND DEC2 EXPRESSION IN MICE 415

However, similar contra-phase expression profilesbetween rat and mouse were known for Dbp: a peak atnight in rat liver (Wuarin and Schibler, 1990) and apeak in the subjective day in mouse liver (Mitsui et al.,2001); these findings are in accord with our presentand previous data (Noshiro et al., 2004). The mecha-nism(s) involved and the physiological significance ofthe discrepancy between rat and mouse are unknown.However, our observations on the discrepancybetween animal species, taken together with the dis-sociation of expression profiles between light condi-tions described above, may be important clues thatcould clarify the function and entrainmentmechanism of peripheral clocks in future studies.
A recent study of Dec1 knockout mice (Stra13–/–)(Grechez-Cassiau et al., 2004) shows that more than 40hepatic genes are affected, compared with wild-typemice. In contrast, DEC2 suppressed the hepaticexpression of CYP7A, CYP8B, and CYP51 when theywere enhanced by DBP (Noshiro et al., 2004). Theseobservations indicate that DEC1 and DEC2 work ascircadian output regulators in some tissues, includingthe liver. Elucidation of the tissue-specific regulatorymechanisms will be necessary to clarify both the rolesof DEC1 and DEC2 in the periphery and the regulationof their target genes in the respective tissues.
ACKNOWLEDGMENTS
This work was supported by grants-in-aid for sci-ence from the Ministry of Education, Culture, Sport,Science and Technology of Japan. We thank J. S.Takahashi and N. Ishida for generous gifts of Clockmutant mice and T. Yasuda for technical assistance.We thank the Research Centre for Molecular Medi-cine, Hiroshima University School of Medicine, for theuse of its facilities.
REFERENCES
Balsalobre A, Brown SA, Marcacci L, Tronche F, KellendonkC, Reichardt HM, Schutz G, and Schibler U (2000) Reset-ting of circadian time in peripheral tissues byglucocorticoid signaling. Science 289:2344-2347.
Boudjelal M, Taneja R, Matsubara S, Bouillet P, Dolle P, andChambon P (1997) Overexpression of Stra13, a novelretinoic acid-inducible gene of the basic helix-loop-helixfamily, inhibits mesodermal and promotes neuronal dif-ferentiation of P19 cells. Genes Dev 11:2052-2065.
Butler M, Honma S, Fukumoto T, Kawamoto T, Fujimoto K,Noshiro M, Kato Y, and Honma K (2004) Dec1 and Dec2
expression is disrupted in the suprachiasmatic nuclei ofClock mutant mice. J Biol Rhythms 19:126-134.
Daan S and Aschoff J (2001) The entrainment of circadiansystem. In Handbook of Behavioral Neurobiology: 12. Circa-dian Clocks, Takahashi JS, Turek FW, Moore RY, eds, pp 7-43, Kluwer Academic/Plenum, New York.
Damiola F, Le Minh N, Preitner N, Kornmann B, Fleury-Olela F, and Schibler U (2000) Restricted feeding uncou-ples circadian oscillators in peripheral tissues from thecentral pacemaker in the suprachiasmatic nucleus. GenesDev 14:2950-2961.
Fujimoto K, Shen M, Noshiro M, Matsubara K, Shingu S,Honda K, Yoshida E, Suardita K, Matsuda Y, and Kato Y(2001) Molecular cloning and characterization of DEC2, anew member of basic helix-loop-helix proteins. BiochemBiophys Res Commun 280:164-171.
Gekakis N, Staknis D, Nguyen HB, Davis FC, WilsbacherLD, King DP, Takahashi JS, and Weitz CJ (1998). Role ofthe CLOCK protein in the mammalian circadian mecha-nism. Science 280:1564-1569.
Gibson UE, Heid CA, and Williams PM (1996). A novelmethod for real time quantitative RT-PCR. Genome Res6:995-1001.
Grechez-Cassiau A, Panda S, Lacoche S, Teboul M, Azmi S,Laudet V, Hogenesch JB, Taneja R, and Delaunay F (2004).The transcriptional repressor STRA13 regulates a subsetof peripheral circadian outputs. J Biol Chem 279:1141-1150.
Hamaguchi H, Fujimoto K, Kawamoto T, Noshiro M,Maemura K, Takeda N, Nagai R, Furukawa M, Honma S,Honma K, et al. (2004). Expression of the gene for Dec2, abasic helix-loop-helix transcription factor, is regulated bya molecular clock system. Biochem J 382:43-50.
Honma S, Kawamoto T, Takagi Y, Fujimoto K, Sato F,Noshiro M, Kato Y, and Honma K (2002). Dec1 and Dec2are regulators of the mammalian molecular clock. Nature419:841-844.
Ivanova AV, Ivanov SV, Danilkovitch-Miagkova A, andLerman MI (2001). Regulation of STRA13 by the vonHippel-Lindau tumor suppressor protein, hypoxia, andthe UBC9/ubiquitin proteasome degradation pathway. JBiol Chem 276:15306-15315.
Jin X, Shearman LP, Weaver DR, Zylka MJ, de Vries GJ, andReppert SM (1999). A molecular mechanism regulatingrhythmic output from the suprachiasmatic circadianclock. Cell 96:57-68.
Kawamoto T, Noshiro M, Sato F, Maemura K, Takeda N,Nagai R, Iwata T, Fujimoto K, Furukawa M, Miyazaki K,et al. (2004). Anovel autofeedback loop of Dec1 transcrip-tion involved in circadian rhythm regulation. BiochemBiophys Res Commun 313:117-124.
King DP, Zhao Y, Sangoram AM, Wilsbacher LD, Tanaka M,Antoch MP, Steeves TD, Vitaterna MH, Kornhauser JM,Lowrey PL, et al. (1997). Positional cloning of the mousecircadian clock gene. Cell 89:641-653.
Kume K, Zylka MJ, Sriram S, Shearman LP, Weaver DR, JinX, Maywood ES, Hastings MH, and Reppert SM (1999).mCRY1 and mCRY2 are essential components of the neg-ative limb of the circadian clock feedback loop. Cell98:193-205.
416 JOURNAL OF BIOLOGICAL RHYTHMS / October 2005

Mitsui S, Yamaguchi S, Matsuo T, Ishida Y, and Okamura H(2001). Antagonistic role of E4BP4 and PAR proteins inthe circadian oscillatory mechanism. Genes Dev 15:995-1006.
Miyazaki K, Kawamoto T, Tanimoto K, Nishiyama M,Honda H, and Kato Y (2002). Identification of functionalhypoxia response elements in the promoter region of theDEC1 and DEC2 genes. J Biol Chem 277:47014-47021.
Nakamura W, Honma S, Shirakawa T, and Honma K (2002).Clock mutation lengthens the circadian period withoutdamping rhythms in individual SCN neurons. NatNeurosci 5:399-400.
Niijima A, Nagai K, Nagai N, and Akagawa H (1993). Effectsof light stimulation on the activity of the autonomicnerves in anesthetized rats. Physiol Behav 54:555-561.
Noshiro M, Kawamoto T, Furukawa M, Fujimoto K, YoshidaY, Sasabe E, Tsutsumi S, Hamada T, Honma S, Honma K,et al. (2004). Rhythmic expression of DEC1 and DEC2 inperipheral tissues: DEC2 is a potent suppressor forhepatic cytochrome P450s opposing DBP. Genes Cells9:317-329.
Oishi K, Fukui H, Ishida N, Ikeda M, Yu W, Hirai M, EbisawaT, Honma S, Yoshimura K, Honma KI, et al. (2000). Rhyth-mic expression of BMAL1 mRNA is altered in Clockmutant mice: differential regulation in thesuprachiasmatic nucleus and peripheral tissues.Biochem Biophys Res Commun 268:164-171.
Oishi K, Miyazaki K, and Ishida N (2002). FunctionalCLOCK is not involved in the entrainment of peripheralclocks to the restricted feeding: entrainable expression ofmPer2 and BMAL1 mRNAs in the heart of Clock mutantmice on Jcl:ICR background. Biochem Biophys ResCommun 298:198-202.
Oishi K, Miyazaki K, Kadota K, Kikuno R, Nagase T, AtsumiG, Ohkura N, Azama T, Mesaki M, Yukimasa S, et al.(2003). Genome-wide expression analysis of mouse liverreveals CLOCK-regulated circadian output genes. J BiolChem 278:41519-41527.
Panda S, Antoch MP, Miller BH, Su AI, Schook AB, StraumeM, Schultz PG, Kay SA, Takahashi JS, and Hogenesch JB(2002). Coordinated transcription of key pathways in themouse by the circadian clock. Cell 109:307-320.
Pittendrigh CS and Daan S (1976). Functional-analysis of cir-cadian pacemakers in nocturnal rodents: IV. Entrain-ment: pacemaker as clock. J Comp Physiol 106:291-331.
Reick M, Garcia JA, Dudley C, and McKnight SL (2001).NPAS2: an analog of clock operative in the mammalianforebrain. Science 293:506-509.
Ripperger JA, Shearman LP, Reppert SM, and Schibler U(2000). CLOCK, an essential pacemaker component, con-trols expression of the circadian transcription factor DBP.Genes Dev 14:679-689.
Roenneberg T and Merrow M (2003). The network of time:understanding the molecular circadian system. Curr Biol13:R198-R207.
Rossner MJ, Dorr J, Gass P, Schwab MH, and Nave KA(1997). SHARPs: mammalian enhancer-of-split- andhairy-related proteins coupled to neuronal stimulation.Mol Cell Neurosci 10:460-475.
Sakamoto K, Nagase T, Fukui H, Horikawa K, Okada T,Tanaka H, Sato K, Miyake Y, Ohara O, Kako K, et al.(1998). Multitissue circadian expression of rat periodhomolog (rPer2) mRNA is governed by the mammaliancircadian clock, the suprachiasmatic nucleus in the brain.J Biol Chem 273:27039-27042.
Sato F, Kawamoto T, Fujimoto K, Noshiro M, Honda KK,Honma S, Honma K, and Kato Y (2004). Functional analy-sis of the basic helix-loop-helix transcription factor DEC1in circadian regulation: interaction with BMAL1. Eur JBiochem 271:4409-4419.
Shen M, Kawamoto T, Teramoto M, Makihira S, Fujimoto K,Yan W, Noshiro M, and Kato Y (2001). Induction of basichelix-loop-helix protein DEC1 (BHLHB2)/Stra13/Sharp2 in response to the cyclic adenosinemonophosphate pathway. Eur J Cell Biol 80:329-334.
Shen M, Kawamoto T, Yan W, Nakamasu K, Tamagami M,Koyano Y, Noshiro M, and Kato Y (1997). Molecular char-acterization of the novel basic helix-loop-helix proteinDEC1 expressed in differentiated human embryochondrocytes. Biochem Biophys Res Commun 236:294-298.
Shen M, Yoshida E, Yan W, Kawamoto T, Suardita K, KoyanoY, Fujimoto K, Noshiro M, and Kato Y (2002). Basic helix-loop-helix protein DEC1 promotes chondrocyte differen-tiation at the early and terminal stages. J Biol Chem277:50112-50120.
Shibata S (2004). Neural regulation of the hepatic circadianrhythm. Anat Rec A Discov Mol Cell Evol Biol 280:901-909.
Storch KF, Lipan O, Leykin I, Viswanathan N, Davis FC,Wong WH, and Weitz CJ (2002). Extensive and divergentcircadian gene expression in liver and heart. Nature417:78-83.
Sun H, Lu B, Li RQ, Flavell RA, and Taneja R (2001). Defec-tive T cell activation and autoimmune disorder in Stra13-deficient mice. Nat Immunol 2:1040-1047.
Sun H and Taneja R (2000). Stra13 expression is associatedwith growth arrest and represses transcription throughhistone deacetylase (HDAC)-dependent and HDAC-independent mechanisms. Proc Natl Acad Sci USA97:4058-4063.
Teramoto M, Nakamasu K, Noshiro M, Matsuda Y, Gotoh O,Shen M, Tsutsumi S, Kawamoto T, Iwamoto Y, and Kato Y(2001). Gene structure and chromosomal location of ahuman bHLH transcriptional factor DEC1/Stra13/SHARP-2/BHLHB2. J Biochem 129:391-396.
Terazono H, Mutoh T, Yamaguchi S, Kobayashi M, AkiyamaM, Udo R, Ohdo S, Okamura H, and Shibata S (2003).Adrenergic regulation of clock gene expression in mouseliver. Proc Natl Acad Sci USA 100:6795-6800.
Thomas PS (1980). Hybridization of denatured RNA andsmall DNA fragments transferred to nitrocellulose. ProcNatl Acad Sci USA 77:5201-5205.
Tischkau SA, Mitchell JW, Tyan SH, Buchanan GF, andGillette MU (2003). Ca2+/cAMP response element-binding protein (CREB)-dependent activation of Per1 isrequired for l ight- induced signaling in the
Noshiro et al. / DEC1 AND DEC2 EXPRESSION IN MICE 417

suprachiasmatic nucleus circadian clock. J Biol Chem278:718-723.
Travnickova-Bendova Z, Cermakian N, Reppert SM, andSassone-Corsi P (2002). Bimodal regulation of mPeriodpromoters by CREB-dependent signaling and CLOCK/BMAL1 activity. Proc Natl Acad Sci USA 99:7728-7733.
Ueda HR, Chen W, Adachi A, Wakamatsu H, Hayashi S,Takasugi T, Nagano M, Nakahama K, Suzuki Y, Sugano S,et al. (2002). A transcription factor response element forgene expression during circadian night. Nature 418:534-539.
Vitaterna MH, King DP, Chang AM, Kornhauser JM, LowreyPL, McDonald JD, Dove WF, Pinto LH, Turek FW, andTakahashi JS (1994). Mutagenesis and mapping of a
mouse gene, Clock, essential for circadian behavior. Sci-ence 264:719-725.
Wuarin J and Schibler U (1990). Expression of the liver-enriched transcriptional activator protein DBP follows astringent circadian rhythm. Cell 63:1257-1266.
Yamada K, Kawata H, Shou Z, Mizutani T, Noguchi T, andMiyamoto K (2003). Insulin induces the expression of theSHARP-2/Stra13/DEC1 gene via a phosphoinositide 3-kinase pathway. J Biol Chem 278:30719-30724.
Zhou YD, Barnard M, Tian H, Li X, Ring HZ, Francke U,Shelton J, Richardson J, Russell DW, and McKnight SL(1997). Molecular characterization of two mammalianbHLH-PAS domain proteins selectively expressed in thecentral nervous system. Proc Natl Acad Sci USA 94:713-718.
418 JOURNAL OF BIOLOGICAL RHYTHMS / October 2005



















