
Thyroid hormones and deiodinase activity in plasma and tissues in relation to high levels of...
11
Thyroid hormones and deiodinase activity in plasma and tissues in relation to high levels of organohalogen contaminants in East Greenland polar bears (Ursus maritimus) Kristin Møller Gabrielsen a,n , Julie Stene Krokstad a , Gro Dehli Villanger a,b , David A.D. Blair c,d , Maria-Jesus Obregon e , Christian Sonne f , Rune Dietz f , Robert J. Letcher c,d , Bjørn Munro Jenssen a,nn a Department of Biology, Norwegian University of Science and Technology, NO-7491 Trondheim, Norway b Division of Mental Health, Department of Child Development and Mental Health, Norwegian Institute of Public Health, P.O. Box 4404 Nydalen, 0473 Oslo, Norway c Ecotoxicology and Wildlife Health Division, Science and Technology Branch, Environment Canada, National Wildlife Research Centre, Carleton University, Ottawa, Ontario, Canada K1A 0H3 d Department of Chemistry, Carleton University, Ottawa, Ontario, Canada K1S 5B6 e Instituto de Investigaciones Biomedicas, Centro Mixto from CSIC-UAM, 28029 Madrid, Spain f Department of Bioscience, Arctic Research Centre, Aarhus University, Roskilde, P.O. Box 358, DK-4000, Denmark article info Article history: Received 9 May 2014 Received in revised form 22 September 2014 Accepted 24 September 2014 Keywords: Polar bears East Greenland Thyroid hormones Deiodinase activity Organohalogen contaminants abstract Previous studies have shown relationships between organohalogen contaminants (OHCs) and circulating levels of thyroid hormones (THs) in arctic wildlife. However, there is a lack of knowledge concerning the possible functional effects of OHCs on TH status in target tissues for TH-dependent activity. The relationships between circulating (plasma) levels of OHCs and various TH variables in plasma as well as in liver, muscle and kidney tissues from East Greenland sub-adult polar bears (Ursus maritimus) sampled in 2011 (n ¼7) were therefore investigated. The TH variables included 3.3′,5.5′-tetraiodothyr- onine or thyroxine (T4), 3.3′,5-triiodothyronine (T3) and type 1 (D1) and type 2 (D2) deiodinase activities. Principal component analysis (PCA) combined with correlation analyses demonstrated negative relationships between individual polychlorinated biphenyls (PCBs) and their hydroxylated (OH–) metabolites and T4 in both plasma and muscle. There were both positive and negative relation- ships between individual OHCs and D1 and D2 activities in muscle, liver and kidney tissues. In general, PCBs, OH-PCBs and polybrominated dipehenyl ethers (PBDEs) were positively correlated to D1 and D2 activities, whereas organochlorine pesticides and byproducts (OCPs) were negatively associated with D1 and D2 activities. These results support the hypothesis that OHCs can affect TH status and action in the target tissues of polar bears. TH levels and deiodinase activities in target tissues can be sensitive endpoints for exposure of TH-disrupting compounds in arctic wildlife, and thus, tissue-specific responses in target organs should be further considered when assessing TH disruption in wildlife studies. & 2014 Elsevier Inc. All rights reserved. 1. Introduction Polar bears (Ursus maritimus) from East Greenland, Svalbard and Western Russia have the highest concentrations of organoha- logen contaminants (OHCs) in the Arctic (Letcher et al., 2010; McKinney et al., 2011a; 2011b). Although the levels of some legacy OHCs including polychlorinated biphenyls (PCBs) and many organochlorine pesticides and byproducts (OCPs) are showing decreasing temporal trends in East Greenland polar bears (Dietz et al., 2013b), other contaminants such as polybrominated diphe- nyl ethers (PBDEs) have been reported to continue to increase until recently (Dietz et al., 2013a). However, altered diet in the opportunistic polar bear due to climate change might also cause changes in contaminant levels and patterns of both new and legacy contaminants, as recently shown for the East Greenland polar bears by McKinney et al. (2013). High concentrations of OHCs in polar bears from East Greenland and Svalbard have been associated with alterations of plasma thyroid hormone (TH) homoeostasis (Braathen et al., 2004; Bytingsvik, 2012; Skaare Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/envres Environmental Research http://dx.doi.org/10.1016/j.envres.2014.09.019 0013-9351/& 2014 Elsevier Inc. All rights reserved. n Corresponding author. nn Corresponding author. E-mail addresses: [email protected] (K.M. Gabrielsen), [email protected] (B.M. Jenssen). Environmental Research 136 (2015) 413–423
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Thyroid hormones and deiodinase activity in plasma and tissues in relation to high levels of...

Environmental Research 136 (2015) 413–423
Contents lists available at ScienceDirect
Environmental Research
http://d0013-93
n Corrnn CorE-m
bmj@bi
journal homepage: www.elsevier.com/locate/envres
Thyroid hormones and deiodinase activity in plasma and tissues inrelation to high levels of organohalogen contaminants in EastGreenland polar bears (Ursus maritimus)
Kristin Møller Gabrielsen a,n, Julie Stene Krokstad a, Gro Dehli Villanger a,b,David A.D. Blair c,d, Maria-Jesus Obregon e, Christian Sonne f, Rune Dietz f,Robert J. Letcher c,d, Bjørn Munro Jenssen a,nn
a Department of Biology, Norwegian University of Science and Technology, NO-7491 Trondheim, Norwayb Division of Mental Health, Department of Child Development and Mental Health, Norwegian Institute of Public Health, P.O. Box 4404 Nydalen, 0473 Oslo,Norwayc Ecotoxicology and Wildlife Health Division, Science and Technology Branch, Environment Canada, National Wildlife Research Centre, Carleton University,Ottawa, Ontario, Canada K1A 0H3d Department of Chemistry, Carleton University, Ottawa, Ontario, Canada K1S 5B6e Instituto de Investigaciones Biomedicas, Centro Mixto from CSIC-UAM, 28029 Madrid, Spainf Department of Bioscience, Arctic Research Centre, Aarhus University, Roskilde, P.O. Box 358, DK-4000, Denmark
a r t i c l e i n f o
Article history:Received 9 May 2014Received in revised form22 September 2014Accepted 24 September 2014
Keywords:Polar bearsEast GreenlandThyroid hormonesDeiodinase activityOrganohalogen contaminants
x.doi.org/10.1016/j.envres.2014.09.01951/& 2014 Elsevier Inc. All rights reserved.
esponding author.responding author.ail addresses: [email protected] (K.M. Gabro.ntnu.no (B.M. Jenssen).
a b s t r a c t
Previous studies have shown relationships between organohalogen contaminants (OHCs) and circulatinglevels of thyroid hormones (THs) in arctic wildlife. However, there is a lack of knowledge concerning thepossible functional effects of OHCs on TH status in target tissues for TH-dependent activity. Therelationships between circulating (plasma) levels of OHCs and various TH variables in plasma as wellas in liver, muscle and kidney tissues from East Greenland sub-adult polar bears (Ursus maritimus)sampled in 2011 (n¼7) were therefore investigated. The TH variables included 3.3′,5.5′-tetraiodothyr-onine or thyroxine (T4), 3.3′,5-triiodothyronine (T3) and type 1 (D1) and type 2 (D2) deiodinaseactivities. Principal component analysis (PCA) combined with correlation analyses demonstratednegative relationships between individual polychlorinated biphenyls (PCBs) and their hydroxylated(OH–) metabolites and T4 in both plasma and muscle. There were both positive and negative relation-ships between individual OHCs and D1 and D2 activities in muscle, liver and kidney tissues. In general,PCBs, OH-PCBs and polybrominated dipehenyl ethers (PBDEs) were positively correlated to D1 and D2activities, whereas organochlorine pesticides and byproducts (OCPs) were negatively associated with D1and D2 activities. These results support the hypothesis that OHCs can affect TH status and action in thetarget tissues of polar bears. TH levels and deiodinase activities in target tissues can be sensitiveendpoints for exposure of TH-disrupting compounds in arctic wildlife, and thus, tissue-specific responsesin target organs should be further considered when assessing TH disruption in wildlife studies.
& 2014 Elsevier Inc. All rights reserved.
1. Introduction
Polar bears (Ursus maritimus) from East Greenland, Svalbardand Western Russia have the highest concentrations of organoha-logen contaminants (OHCs) in the Arctic (Letcher et al., 2010;McKinney et al., 2011a; 2011b). Although the levels of some legacyOHCs including polychlorinated biphenyls (PCBs) and many
ielsen),
organochlorine pesticides and byproducts (OCPs) are showingdecreasing temporal trends in East Greenland polar bears (Dietzet al., 2013b), other contaminants such as polybrominated diphe-nyl ethers (PBDEs) have been reported to continue to increaseuntil recently (Dietz et al., 2013a). However, altered diet in theopportunistic polar bear due to climate change might also causechanges in contaminant levels and patterns of both new andlegacy contaminants, as recently shown for the East Greenlandpolar bears by McKinney et al. (2013). High concentrations of OHCsin polar bears from East Greenland and Svalbard have beenassociated with alterations of plasma thyroid hormone (TH)homoeostasis (Braathen et al., 2004; Bytingsvik, 2012; Skaare

K.M. Gabrielsen et al. / Environmental Research 136 (2015) 413–423414
et al., 2001; Villanger et al., 2011), which is considered to beamong one of the most critical targets of endocrine disruption inmammals (Colborn, 2004). Enzymatic biotransformation of OHCsalso result in metabolites with endocrine disrupting potential,such as hydroxylated (OH-) PCBs (OH-PCBs) and PBDEs (OH-PBDEs) and metylsulfonated PCBs (MeSO2-PCBs) (Letcher et al.,2000; Wiseman et al., 2011).
THs are physiologically essential in all mammals, being im-portant for growth, development, reproduction, stress response,tissue repair, metabolism and thermoregulation (Zoeller et al.,2007). They are especially important during growth and neurolo-gical development in prenatal, neonatal and young individuals(Bernal, 2005). Thus, if disruption of the TH system in mammalsoccurs during sensitive windows in pre- and postnatal develop-ment, it can have deleterious effects in later life-stages (Zoelleret al., 2002).
3,3′,5,5′-Tetraiodothyronine or thyroxine (T4) is the major formof TH produced and secreted from the thyroid gland. T4 isdeiodinated to the biologically active 3,3′,5-triiodothyronine (T3)in peripheral tissues (Zoeller et al., 2007). Except for the o0.01%existing in plasma as free hormone, T4 and T3 are bound to theproteins thyroxin binding globulin (TBG), transthyretin (TTR) oralbumin when transported in plasma in mammals (Hulbert, 2000).T3 exerts its effects through either nuclear TH receptors (TRα andTRβ) or non-genomic responses (Cheng et al., 2010). Overall, theTH system is regulated by a negative feedback system underneural control, involving the thyroid stimulating pituitary hor-mone thyrotropin (TSH), which itself is regulated by the releasingfactor thyrotropin-releasing hormone (TRH) from the hypothala-mus (Zoeller et al., 2007).
Activation or inactivation of THs via deiodination is regulatedby type 1 (D1), type 2 (D2) and type 3 (D3) deiodinases (Biancoet al., 2002). These enzymes are essential for regulating the plasmaand intracellular levels of THs. The membrane-bound D1 is themain generator of T3 to the plasma pool by converting T4 to T3 viaouter (5′) ring deiodination (ORD). The enzyme is highly expressedin liver, kidney and thyroid, and the conversion of T4 to T3 isincreased during hyperthyroidism (i.e. excessive levels of T3 andT4) (Köhrle, 1999). The endoplasmic reticulum (ER)-bound D2,similarly to D1, mainly mediates the conversion of T4 to T3 viaORD. The proximity of the ER to the nucleus means that D2 ishighly involved in supplying T3 to the nuclear pool for localrequirements, in contrast to D1 which mainly supplies T3 to theplasma pool (Gereben et al., 2008). Contrary to D1, D2-mediatedconversion of T4 to T3 is decreased by hyperthyroidism andincreased by hypothyroidism (i.e. insufficient levels of T3 and T4)(Köhrle, 1999). D2 is present in the brain, skeletal muscle, heartand brown adipose tissue (BAT), which all exert TH-dependentactions (Bianco et al., 2002). D3 inactivates T3 and T4 by inner ring(5) deiodination (IRD) and is present in brain, skin, placcenta andfoetal tissues (Bianco et al., 2002).
There are in general two main mechanisms suggested toexplain the frequently reported negative associations betweenOHCs and circulating T4 levels in wildlife, humans and laboratorystudies. These are either interference with one or several meta-bolism pathways (glucuronidation, sulphonation or deiodination)or via interactions with TH-binding transport proteins in plasma(Boas et al., 2012; Gutleb et al. 2010; Crofton, 2008). Studies haveshown associations between OHC exposure and alterations inhepatic glucuronidation or sulphonation activity as well as com-petitive binding of OH-PCBs and OH-PBDEs to TTR (as reviewed inBoas et al., 2012). However, due to the multiple target points ofOHCs in the TH systems, other modes of action are also plausible(Crofton, 2008). The deiodinases have received less attentionregarding their role in the TH system in a toxicological context(Zoeller et al., 2007), despite their important role of regulating and
controlling intracellular TH levels in a precise spatio-temporalmanner, independent of plasma levels of THs (Bianco and Kim,2006; Dentice et al., 2013). In fact, few studies, including experi-mental animal studies, have addressed the effects of OHCs intarget tissues for TH-action, including the effects of OHCs ondeiodinases. Thus, there appears to be a knowledge gap regardingthe possible functional effects of OHCs in target tissues, andwhether or not effects observed in plasma can result in anydown-stream biological effects.
Studies of polar bears from Svalbard and East Greenland haveshown that OHCs and their metabolites might affect circulatinglevels of THs, possibly through interaction with TTR or othermechanisms (Braathen et al., 2004; Bytingsvik, 2012; Bytingsviket al., 2013; Simon et al., 2011, 2013; Skaare et al., 2001; Villangeret al., 2011). However, there is currently no evidence in theliterature for polar bears as to whether OHC exposure also affectsthe TH-status in tissues important for deiodination and TH-action,such as muscle, liver and kidney. Since polar bears experienceperiods of extreme low temperatures and prolonged fasting(Stirling, 2002), TH-dependent processes such as thermoregula-tion and fasting metabolism are of particular importance. Func-tional effects of TH-disruptive compounds in target tissues for TH-action in arctic animals might therefore be of concern (Jenssen,2006).
In the present study we therefore hypothesized that there wereassociations between plasma concentrations of OHCs and severalimportant TH variables (T4, T3, D1 and D2 activities) in plasma andliver, kidney, and muscle tissues in sub-adult polar bears from theEast Greenland population. Because blood is a central transportcompartment that maintains the equilibrium between uptake ofcontaminants via diet and the storage and/or metabolisation ofOHCs in various organ-tissues in marine mammals (Boon et al.,1994), it is a an active matrix that reaches target tissues. Plasmaconcentrations of OHCs were therefore applied as a proxy for theexposure status in the polar bears.
2. Animals, materials and methods
2.1. Sampling
The polar bears (n¼7) investigated in the present study wereharvested by local Inuit hunters in February to March 2011 as partof the annual subsistence hunt in the Scoresby Sound (Ittoqqor-toormiit) area, 70°29′N 21°58′W, East Greenland. The polar bearswere 2–4 years of age and defined as sexually immature and henceclassified as sub-adults (Rosing-Asvid et al., 2002). Two of theanimals were males whereas five were females (Table S1). Thepresent study was part of the AURORAE programme led by Prof. D.Sc. Rune Dietz at Aarhus University.
The animals were quickly subjected to post mortem dissectionand sampling (o3 h after death). Sex, standard length, zoologicallength and circumference were noted. Blood samples were col-lected primarily from the heart using a 50 mL syringe andtransferred into Vacuette© heparinized tubes, and then centri-fuged in order to collect plasma. The plasma samples were keptfrozen at �10 °C to �20 °C during the field period and thentransferred to �20 °C until analysis for THs and OHCs. Tissuesamples of liver, kidney and muscle for use in the present studywere sampled and then stored in two different ways. Samples of20–40 g (g) were taken and stored frozen at �10 °C to �20 °Cduring the field period and then transferred to �20 °C untilanalysis. Additional tissue samples (2–5 g) were snap frozen inliquid nitrogen (�196 °C) during the field period and thentransferred to �70 °C until analysis. These nitrogen stored tissueswere the preferred tissues for analyzing levels of TH and D1 and

K.M. Gabrielsen et al. / Environmental Research 136 (2015) 413–423 415
D2 activities. Snap frozen samples were only available from 4 to5 specimens, therefore analyses of regular frozen tissue from liver/kidney and muscle were conducted on 2 and 3 specimens,respectively. However, the low environmental temperature aswas the case herein aided in preserving the samples and sloweddown any degradative processes. This was demonstrated by thefact that the enzyme activities and TH concentrations in the nonsnap-freezed samples were measurable and comparable withthose samples that were snap-freezed. See also Supporting In-formation (SI), Fig S2.
2.2. Contaminant analyses
The analyses of chlorinated and brominated organic com-pounds and their metabolites in plasma were conducted in theLetcher Labs (Organic Contaminants Research Laboratory (OCRL))within the Ecotoxicology and Wildlife Health Division, Environ-ment Canada located at the National Wildlife Research Centre(NWRC), Carleton University, Ottawa, Canada. The method con-sisted of clean-up procedures and extraction based on methods forblood, plasma or serumwith modifications as described elsewhere(Gebbink et al., 2008a, 2008b) before identification and quantifi-cation using gas chromatography (GC) and single quadropole massspectrometry (MS). Further details can be found in the SI.
The individual or co-eluted compounds analysed for in the presentstudy were 74 individual and co-eluting PCB congeners, 20 OCPcompounds, 17 MeSO2-PCBs, bis-2,2-(4-chlorophenyl)-1,1,1-trichlor-oethane (p,p'-DDT) and its metabolites bis-2,2-(4-chlorophenyl)-1,1-dichloroethene (p,p'-DDE) and bis-2,2-(4-chlorophenyl)-1,1-dichlor-oethane (p,p'-DDD), and 35 individual or co-eluting OH-PCB andother chlorinated phenolic congeners, 17 individual and co-elutingPBDEs and non-PBDE brominated flame retardants (BFRs), and 14OH-PBDEs congeners (for details, see SI). The concentrations arereported in pmol/g wet weight (ww), while ng/g ww can be foundin Table S2 in SI.
A blank was included with the batch of extracted samples, andtarget compound concentrations were blank subtracted whentrace amount of target compounds were detected in the blanksamples. Quality control (QC) samples consisted of bovine (Bostaurus) serum spiked with known concentrations of the OHCanalytes. With respect to quality control, mean percent recoveriesin plasma were: 113.1%79.6% for PCBs, 102.8%710.0% for OCPs,99.3%75.2% for PBDEs, 79.9712.3% for MeSO2-PCBs and 102.2%737.2% for OH-PCBs, 111.5%713.6% for OH-PBDEs. Method detec-tion limits were 10� signal to noise ratio for limit of quantifica-tion (MLOQ) and 3� signal to noise ratio for limit of detection(MLOD). MLODs and MLOQ (pmol/g ww), respectively, rangedfrom 0.06 to 0.39 and from 0.21 to 1.29 for PCBs, from 0.12 to 34.38and from 0.41 to 114.61 for OCPs, from 0.02 to 0.31 and from 0.06to 1.04 for PBDEs, from 0.01 to 0.73 and from 0.04 to 2.43 for OH-PCBs and other chlorinated phenolic congeners, from 0.14 to 1.26and from 0.45 to 4.21 for MeSO2-PCB/DDTs, and from 0.13 to 0.47and from 0.43 to 1.58 for OH-PBDEs. MLOQs and MLODs (ng/g ww)are available in SI.
2.3. Analysis of thyroid hormones in plasma
Analyses of total T4 and T3 in plasma (T4P and T3P, respec-tively) were conducted at the Norwegian University of Science andTechnology, Trondheim, Norway using commercially availablesolid phase 125I radioimmune assay (RIA) kits (Coat-A-Count TotalT3, Coat-A-Count Total T4, Siemens Medical Solutions, CA, USA)without modifications. Although developed for human use, com-mercial kits have been used successfully on polar bear plasmapreviously (Braathen et al., 2004; Bytingsvik, 2012). The coeffi-cients of variation (CVs) were acceptable (r15%). Standard
reference material (SRM, Lyphocheks Immunoassay Plus Control,Levels 2 and 3, BIO-RAD, CA, USA) and internal reference materialbovine plasma were within a satisfactory CV (r15%). T4P and T3Pwere converted to per gram (1 mL plasma¼1 g plasma) andreported in pmol/g in order to compare levels across the investi-gated tissues. All TH levels reported are total hormone levels (i.e.protein-boundþ free).
2.4. Analysis of thyroid hormones and deiodinase activity in liver,kidney and muscle
Analyses of TH concentrations and D1 and D2 activities in liver,kidney and muscle were performed at Instituto de InvestigacionesBiomedicas (IIBM), Madrid, Spain. [125I]-T3 and [125I]-T4 were usedfor determination of TH concentrations in RIAs, while [131I]-T4 and[125I]-T3 were used as recovery tracers during the extraction.Further, [125I]-rT3 was used as substrate for the determination ofD1 and D2 activities.
2.4.1. Thyroid hormone concentrations in liver, muscle and kidneyTotal T4 and T3 in liver (T4L, T3L), kidney (T4K, T3K) and muscle
(T4M, T3M) samples were analysed by RIAs after extraction andpurification of the tissue samples, as originally described inMorreale de Escobar et al. (1985) and later in Lavado-Autric et al.(2013). Limits of sensitivity were 2.5 pg T4 and 0.75 pg T3 pertube. Results were given as pg/g and recalculated and reported aspmol/g. For further details, see SI.
2.4.2. Determination of iodothyronine deiodinase activities in liver,kidney and muscle tissues
The analysis of D1 and D2 activities were performed asdescribed in Lavado-Autric et al. (2013). D1 activities in liver andkidney were measured using 400 nM rT3 plus [125I]-rT3(100.000 cpm/tube) as substrate and 2 mM DTT for 10 min at37 °C. The samples were diluted 1:400 giving 10–20 mg protein pertube. D1 and D2 activities in muscle were measured using[125I]-rT3 (100.000 cpm/tube) plus 2 nM rT3 as substrate and71 mM PTU and 20 mM DTT at 37 °C for 1 h using approximately200 mg protein per tube. D1 activity in muscle was found bysubstracting the D2 activity (insensitive to PTU, assayed by addingPTU) from the total 5′-deiodinase activity (no PTU). The results aregiven as pmol/min/mg protein (D1 in liver [D1L] and kidney [D1K])and fmol/h/mg protein (D1 and D2 in muscle [D1M and D2M,respectively]). For further details, see SI.
2.5. Age and body condition
Age determination was carried out by counting the cementumgrowth layer groups (GLG) of the third incisor tooth (I3) oralternatively the first premolar tooth (P1) as described in Henseland Sorensen (1980) and Dietz et al. (1991). Body condition wascalculated based on the body condition index (BCI) by Cattet et al.(2002), using body mass (BM; calculated according to Derocherand Wiig (2002)) and standard length.
2.6. Data treatment
The present sub-adult female and male polar bears were treated asone group. Only OHCs detected in 470% of the individuals wereincluded in the statistical analyses and reported. Non-detected valueswere replaced with a random value between zero and MLOQ orMLOD. The following compounds had r2 replaced values: PCB-189,-206, pentachlorobenzene, p,p′-DDE, 4-OH-CB159, 3′-OH-180, 3′-OH-CB203/4′-OH-CB198, 3′-MeSO2-CB49, 4-MeSO2-CB64, 4′-MeSO2-CB87(one replaced non-detected value), 1,2,4,5-tetrachlorobenzene, BB-153/BDE-154, 4′-MeSO2-CB101 (two replaced non-detected values).
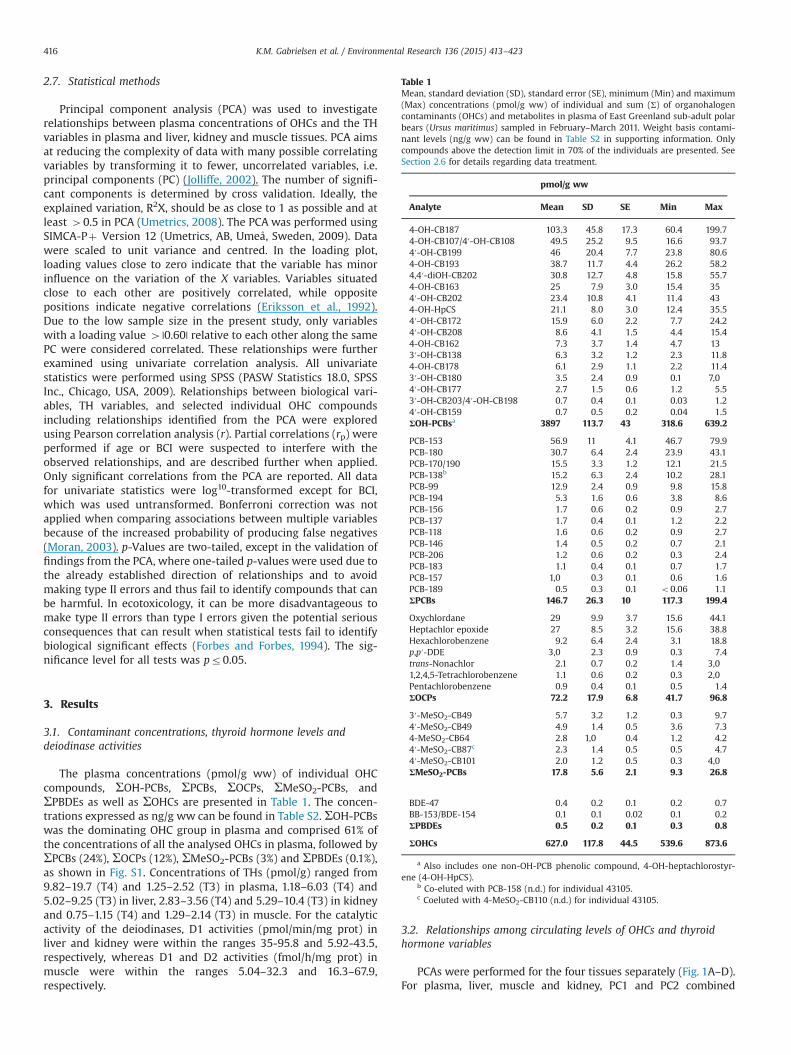
Table 1Mean, standard deviation (SD), standard error (SE), minimum (Min) and maximum(Max) concentrations (pmol/g ww) of individual and sum (Σ) of organohalogencontaminants (OHCs) and metabolites in plasma of East Greenland sub-adult polarbears (Ursus maritimus) sampled in February–March 2011. Weight basis contami-nant levels (ng/g ww) can be found in Table S2 in supporting information. Onlycompounds above the detection limit in 70% of the individuals are presented. SeeSection 2.6 for details regarding data treatment.
pmol/g ww
Analyte Mean SD SE Min Max
4-OH-CB187 103.3 45.8 17.3 60.4 199.74-OH-CB107/4′-OH-CB108 49.5 25.2 9.5 16.6 93.74′-OH-CB199 46 20.4 7.7 23.8 80.64-OH-CB193 38.7 11.7 4.4 26.2 58.24,4′-diOH-CB202 30.8 12.7 4.8 15.8 55.74-OH-CB163 25 7.9 3.0 15.4 354′-OH-CB202 23.4 10.8 4.1 11.4 434-OH-HpCS 21.1 8.0 3.0 12.4 35.54′-OH-CB172 15.9 6.0 2.2 7.7 24.24′-OH-CB208 8.6 4.1 1.5 4.4 15.44-OH-CB162 7.3 3.7 1.4 4.7 133′-OH-CB138 6.3 3.2 1.2 2.3 11.84-OH-CB178 6.1 2.9 1.1 2.2 11.43′-OH-CB180 3.5 2.4 0.9 0.1 7,04′-OH-CB177 2.7 1.5 0.6 1.2 5.53′-OH-CB203/4′-OH-CB198 0.7 0.4 0.1 0.03 1.24′-OH-CB159 0.7 0.5 0.2 0.04 1.5ΣOH-PCBsa 3897 113.7 43 318.6 639.2
PCB-153 56.9 11 4.1 46.7 79.9PCB-180 30.7 6.4 2.4 23.9 43.1PCB-170/190 15.5 3.3 1.2 12.1 21.5PCB-138b 15.2 6.3 2.4 10.2 28.1PCB-99 12.9 2.4 0.9 9.8 15.8PCB-194 5.3 1.6 0.6 3.8 8.6PCB-156 1.7 0.6 0.2 0.9 2.7PCB-137 1.7 0.4 0.1 1.2 2.2PCB-118 1.6 0.6 0.2 0.9 2.7PCB-146 1.4 0.5 0.2 0.7 2.1PCB-206 1.2 0.6 0.2 0.3 2.4PCB-183 1.1 0.4 0.1 0.7 1.7PCB-157 1,0 0.3 0.1 0.6 1.6PCB-189 0.5 0.3 0.1 o0.06 1.1ΣPCBs 146.7 26.3 10 117.3 199.4
Oxychlordane 29 9.9 3.7 15.6 44.1Heptachlor epoxide 27 8.5 3.2 15.6 38.8Hexachlorobenzene 9.2 6.4 2.4 3.1 18.8p,p′-DDE 3,0 2.3 0.9 0.3 7.4trans-Nonachlor 2.1 0.7 0.2 1.4 3,0
K.M. Gabrielsen et al. / Environmental Research 136 (2015) 413–423416
2.7. Statistical methods
Principal component analysis (PCA) was used to investigaterelationships between plasma concentrations of OHCs and the THvariables in plasma and liver, kidney and muscle tissues. PCA aimsat reducing the complexity of data with many possible correlatingvariables by transforming it to fewer, uncorrelated variables, i.e.principal components (PC) (Jolliffe, 2002). The number of signifi-cant components is determined by cross validation. Ideally, theexplained variation, R2X, should be as close to 1 as possible and atleast 40.5 in PCA (Umetrics, 2008). The PCA was performed usingSIMCA-Pþ Version 12 (Umetrics, AB, Umeå, Sweden, 2009). Datawere scaled to unit variance and centred. In the loading plot,loading values close to zero indicate that the variable has minorinfluence on the variation of the X variables. Variables situatedclose to each other are positively correlated, while oppositepositions indicate negative correlations (Eriksson et al., 1992).Due to the low sample size in the present study, only variableswith a loading value 4 |0.60| relative to each other along the samePC were considered correlated. These relationships were furtherexamined using univariate correlation analysis. All univariatestatistics were performed using SPSS (PASW Statistics 18.0, SPSSInc., Chicago, USA, 2009). Relationships between biological vari-ables, TH variables, and selected individual OHC compoundsincluding relationships identified from the PCA were exploredusing Pearson correlation analysis (r). Partial correlations (rp) wereperformed if age or BCI were suspected to interfere with theobserved relationships, and are described further when applied.Only significant correlations from the PCA are reported. All datafor univariate statistics were log10-transformed except for BCI,which was used untransformed. Bonferroni correction was notapplied when comparing associations between multiple variablesbecause of the increased probability of producing false negatives(Moran, 2003). p-Values are two-tailed, except in the validation offindings from the PCA, where one-tailed p-values were used due tothe already established direction of relationships and to avoidmaking type II errors and thus fail to identify compounds that canbe harmful. In ecotoxicology, it can be more disadvantageous tomake type II errors than type I errors given the potential seriousconsequences that can result when statistical tests fail to identifybiological significant effects (Forbes and Forbes, 1994). The sig-nificance level for all tests was pr0.05.
1,2,4,5-Tetrachlorobenzene 1.1 0.6 0.2 0.3 2,0Pentachlorobenzene 0.9 0.4 0.1 0.5 1.4ΣOCPs 72.2 17.9 6.8 41.7 96.8
3′-MeSO2-CB49 5.7 3.2 1.2 0.3 9.74′-MeSO2-CB49 4.9 1.4 0.5 3.6 7.34-MeSO2-CB64 2.8 1,0 0.4 1.2 4.24′-MeSO2-CB87c 2.3 1.4 0.5 0.5 4.74′-MeSO2-CB101 2.0 1.2 0.5 0.3 4,0ΣMeSO2-PCBs 17.8 5.6 2.1 9.3 26.8
BDE-47 0.4 0.2 0.1 0.2 0.7BB-153/BDE-154 0.1 0.1 0.02 0.1 0.2ΣPBDEs 0.5 0.2 0.1 0.3 0.8
ΣOHCs 627.0 117.8 44.5 539.6 873.6
a Also includes one non-OH-PCB phenolic compound, 4-OH-heptachlorostyr-ene (4-OH-HpCS).
b Co-eluted with PCB-158 (n.d.) for individual 43105.c Coeluted with 4-MeSO2-CB110 (n.d.) for individual 43105.
3. Results
3.1. Contaminant concentrations, thyroid hormone levels anddeiodinase activities
The plasma concentrations (pmol/g ww) of individual OHCcompounds, ΣOH-PCBs, ΣPCBs, ΣOCPs, ΣMeSO2-PCBs, andΣPBDEs as well as ΣOHCs are presented in Table 1. The concen-trations expressed as ng/g ww can be found in Table S2. ΣOH-PCBswas the dominating OHC group in plasma and comprised 61% ofthe concentrations of all the analysed OHCs in plasma, followed byΣPCBs (24%), ΣOCPs (12%), ΣMeSO2-PCBs (3%) and ΣPBDEs (0.1%),as shown in Fig. S1. Concentrations of THs (pmol/g) ranged from9.82–19.7 (T4) and 1.25–2.52 (T3) in plasma, 1.18–6.03 (T4) and5.02–9.25 (T3) in liver, 2.83–3.56 (T4) and 5.29–10.4 (T3) in kidneyand 0.75–1.15 (T4) and 1.29–2.14 (T3) in muscle. For the catalyticactivity of the deiodinases, D1 activities (pmol/min/mg prot) inliver and kidney were within the ranges 35-95.8 and 5.92-43.5,respectively, whereas D1 and D2 activities (fmol/h/mg prot) inmuscle were within the ranges 5.04–32.3 and 16.3–67.9,respectively.
3.2. Relationships among circulating levels of OHCs and thyroidhormone variables
PCAs were performed for the four tissues separately (Fig. 1A–D).For plasma, liver, muscle and kidney, PC1 and PC2 combined
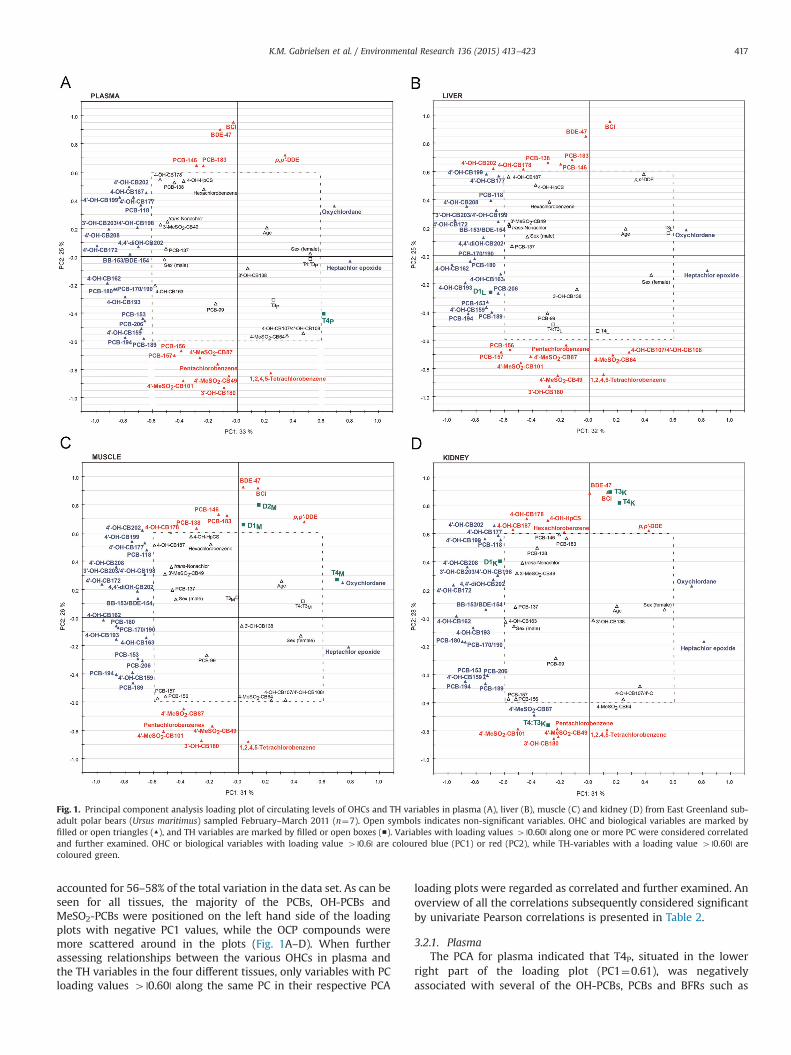
Fig. 1. Principal component analysis loading plot of circulating levels of OHCs and TH variables in plasma (A), liver (B), muscle (C) and kidney (D) from East Greenland sub-adult polar bears (Ursus maritimus) sampled February–March 2011 (n¼7). Open symbols indicates non-significant variables. OHC and biological variables are marked byfilled or open triangles (▲), and TH variables are marked by filled or open boxes (■). Variables with loading values 4 |0.60| along one or more PC were considered correlatedand further examined. OHC or biological variables with loading value 4 |0.6| are coloured blue (PC1) or red (PC2), while TH-variables with a loading value 4 |0.60| arecoloured green.
K.M. Gabrielsen et al. / Environmental Research 136 (2015) 413–423 417
accounted for 56–58% of the total variation in the data set. As can beseen for all tissues, the majority of the PCBs, OH-PCBs andMeSO2-PCBs were positioned on the left hand side of the loadingplots with negative PC1 values, while the OCP compounds weremore scattered around in the plots (Fig. 1A–D). When furtherassessing relationships between the various OHCs in plasma andthe TH variables in the four different tissues, only variables with PCloading values 4 |0.60| along the same PC in their respective PCA
loading plots were regarded as correlated and further examined. Anoverview of all the correlations subsequently considered significantby univariate Pearson correlations is presented in Table 2.
3.2.1. PlasmaThe PCA for plasma indicated that T4P, situated in the lower
right part of the loading plot (PC1¼0.61), was negativelyassociated with several of the OH-PCBs, PCBs and BFRs such as
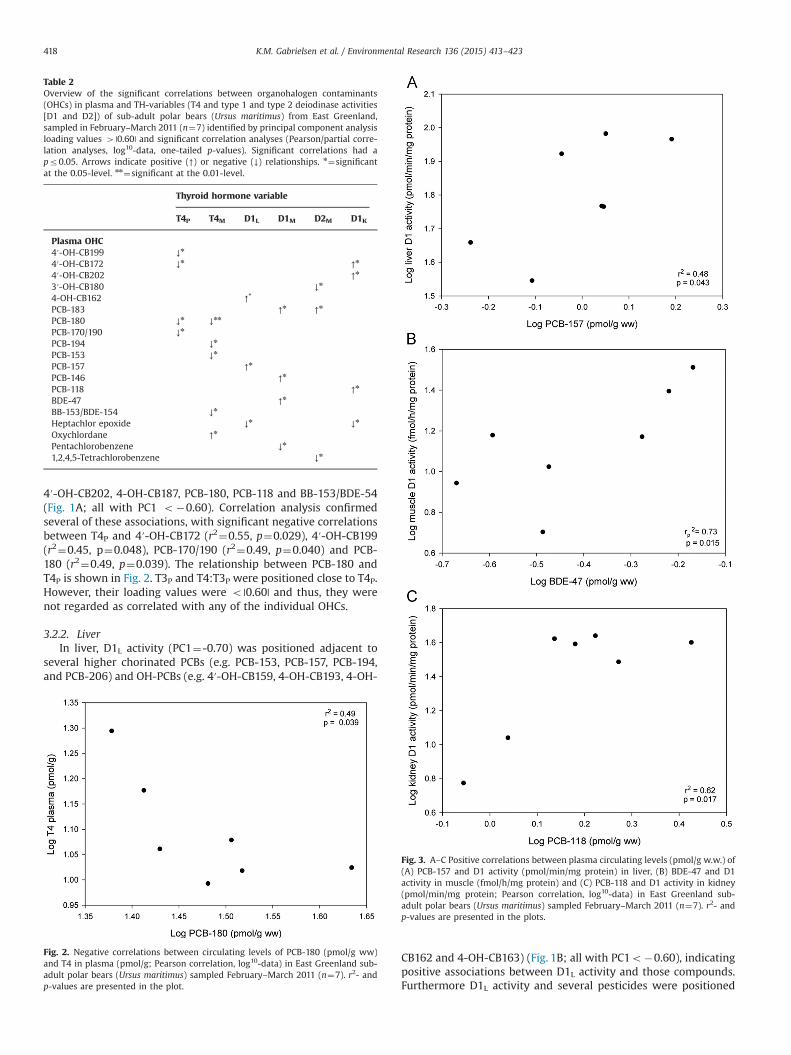
Table 2Overview of the significant correlations between organohalogen contaminants(OHCs) in plasma and TH-variables (T4 and type 1 and type 2 deiodinase activities[D1 and D2]) of sub-adult polar bears (Ursus maritimus) from East Greenland,sampled in February–March 2011 (n¼7) identified by principal component analysisloading values 4 |0.60| and significant correlation analyses (Pearson/partial corre-lation analyses, log10-data, one-tailed p-values). Significant correlations had apr0.05. Arrows indicate positive (↑) or negative (↓) relationships. n¼significantat the 0.05-level. nn¼significant at the 0.01-level.
Thyroid hormone variable
T4P T4M D1L D1M D2M D1K
Plasma OHC4′-OH-CB199 ↓n
4′-OH-CB172 ↓n ↑n
4′-OH-CB202 ↑n
3′-OH-CB180 ↓n
4-OH-CB162 ↑n
PCB-183 ↑n ↑n
PCB-180 ↓n ↓nn
PCB-170/190 ↓n
PCB-194 ↓n
PCB-153 ↓n
PCB-157 ↑n
PCB-146 ↑n
PCB-118 ↑n
BDE-47 ↑n
BB-153/BDE-154 ↓n
Heptachlor epoxide ↓n ↓n
Oxychlordane ↑n
Pentachlorobenzene ↓n
1,2,4,5-Tetrachlorobenzene ↓n
K.M. Gabrielsen et al. / Environmental Research 136 (2015) 413–423418
4′-OH-CB202, 4-OH-CB187, PCB-180, PCB-118 and BB-153/BDE-54(Fig. 1A; all with PC1 o�0.60). Correlation analysis confirmedseveral of these associations, with significant negative correlationsbetween T4P and 4′-OH-CB172 (r2¼0.55, p¼0.029), 4′-OH-CB199(r2¼0.45, p¼0.048), PCB-170/190 (r2¼0.49, p¼0.040) and PCB-180 (r2¼0.49, p¼0.039). The relationship between PCB-180 andT4P is shown in Fig. 2. T3P and T4:T3P were positioned close to T4P.However, their loading values were o |0.60| and thus, they werenot regarded as correlated with any of the individual OHCs.
3.2.2. LiverIn liver, D1L activity (PC1¼-0.70) was positioned adjacent to
several higher chorinated PCBs (e.g. PCB-153, PCB-157, PCB-194,and PCB-206) and OH-PCBs (e.g. 4′-OH-CB159, 4-OH-CB193, 4-OH-
Fig. 2. Negative correlations between circulating levels of PCB-180 (pmol/g ww)and T4 in plasma (pmol/g; Pearson correlation, log10-data) in East Greenland sub-adult polar bears (Ursus maritimus) sampled February–March 2011 (n¼7). r2- andp-values are presented in the plot.
Fig. 3. A–C Positive correlations between plasma circulating levels (pmol/g w.w.) of(A) PCB-157 and D1 activity (pmol/min/mg protein) in liver, (B) BDE-47 and D1activity in muscle (fmol/h/mg protein) and (C) PCB-118 and D1 activity in kidney(pmol/min/mg protein; Pearson correlation, log10-data) in East Greenland sub-adult polar bears (Ursus maritimus) sampled February–March 2011 (n¼7). r2- andp-values are presented in the plots.
CB162 and 4-OH-CB163) (Fig. 1B; all with PC1o�0.60), indicatingpositive associations between D1L activity and those compounds.Furthermore D1L activity and several pesticides were positioned
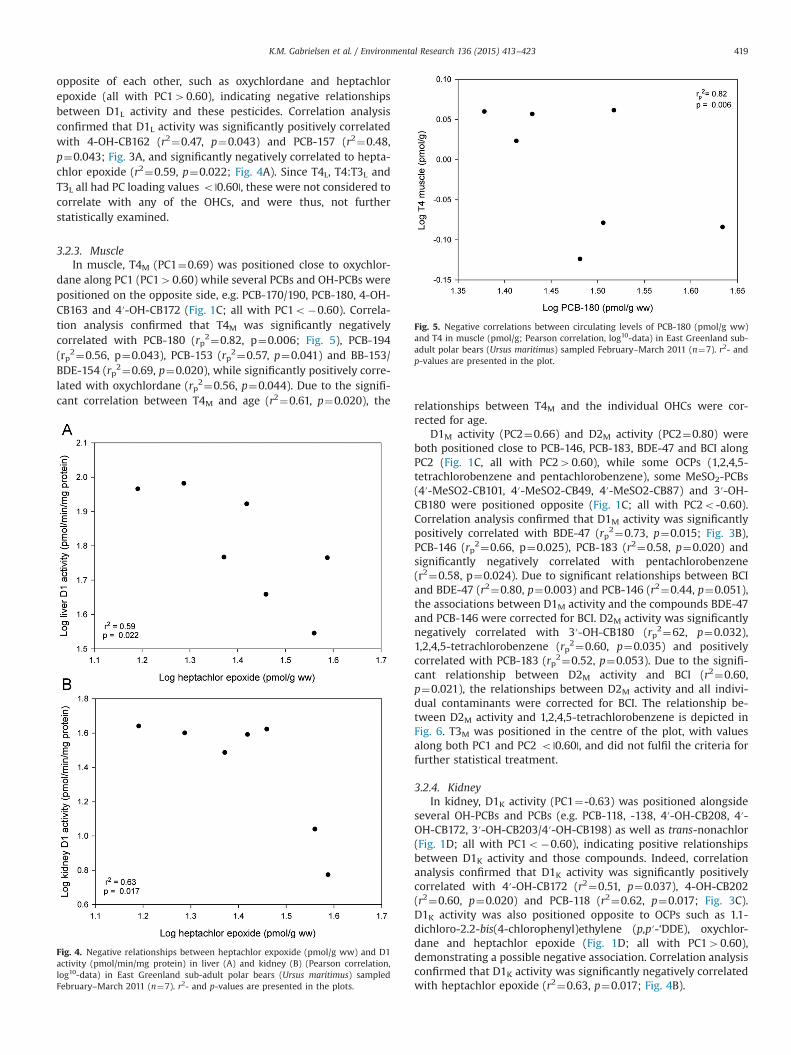
Fig. 5. Negative correlations between circulating levels of PCB-180 (pmol/g ww)and T4 in muscle (pmol/g; Pearson correlation, log10-data) in East Greenland sub-adult polar bears (Ursus maritimus) sampled February–March 2011 (n¼7). r2- andp-values are presented in the plot.
K.M. Gabrielsen et al. / Environmental Research 136 (2015) 413–423 419
opposite of each other, such as oxychlordane and heptachlorepoxide (all with PC140.60), indicating negative relationshipsbetween D1L activity and these pesticides. Correlation analysisconfirmed that D1L activity was significantly positively correlatedwith 4-OH-CB162 (r2¼0.47, p¼0.043) and PCB-157 (r2¼0.48,p¼0.043; Fig. 3A, and significantly negatively correlated to hepta-chlor epoxide (r2¼0.59, p¼0.022; Fig. 4A). Since T4L, T4:T3L andT3L all had PC loading values o |0.60|, these were not considered tocorrelate with any of the OHCs, and were thus, not furtherstatistically examined.
3.2.3. MuscleIn muscle, T4M (PC1¼0.69) was positioned close to oxychlor-
dane along PC1 (PC140.60) while several PCBs and OH-PCBs werepositioned on the opposite side, e.g. PCB-170/190, PCB-180, 4-OH-CB163 and 4′-OH-CB172 (Fig. 1C; all with PC1o�0.60). Correla-tion analysis confirmed that T4M was significantly negativelycorrelated with PCB-180 (rp2¼0.82, p¼0.006; Fig. 5), PCB-194(rp2¼0.56, p¼0.043), PCB-153 (rp2¼0.57, p¼0.041) and BB-153/BDE-154 (rp2¼0.69, p¼0.020), while significantly positively corre-lated with oxychlordane (rp2¼0.56, p¼0.044). Due to the signifi-cant correlation between T4M and age (r2¼0.61, p¼0.020), the
Fig. 4. Negative relationships between heptachlor expoxide (pmol/g ww) and D1activity (pmol/min/mg protein) in liver (A) and kidney (B) (Pearson correlation,log10-data) in East Greenland sub-adult polar bears (Ursus maritimus) sampledFebruary–March 2011 (n¼7). r2- and p-values are presented in the plots.
relationships between T4M and the individual OHCs were cor-rected for age.
D1M activity (PC2¼0.66) and D2M activity (PC2¼0.80) wereboth positioned close to PCB-146, PCB-183, BDE-47 and BCI alongPC2 (Fig. 1C, all with PC240.60), while some OCPs (1,2,4,5-tetrachlorobenzene and pentachlorobenzene), some MeSO2-PCBs(4′-MeSO2-CB101, 4′-MeSO2-CB49, 4′-MeSO2-CB87) and 3′-OH-CB180 were positioned opposite (Fig. 1C; all with PC2o-0.60).Correlation analysis confirmed that D1M activity was significantlypositively correlated with BDE-47 (rp2¼0.73, p¼0.015; Fig. 3B),PCB-146 (rp2¼0.66, p¼0.025), PCB-183 (r2¼0.58, p¼0.020) andsignificantly negatively correlated with pentachlorobenzene(r2¼0.58, p¼0.024). Due to significant relationships between BCIand BDE-47 (r2¼0.80, p¼0.003) and PCB-146 (r2¼0.44, p¼0.051),the associations between D1M activity and the compounds BDE-47and PCB-146 were corrected for BCI. D2M activity was significantlynegatively correlated with 3′-OH-CB180 (rp2¼62, p¼0.032),1,2,4,5-tetrachlorobenzene (rp2¼0.60, p¼0.035) and positivelycorrelated with PCB-183 (rp2¼0.52, p¼0.053). Due to the signifi-cant relationship between D2M activity and BCI (r2¼0.60,p¼0.021), the relationships between D2M activity and all indivi-dual contaminants were corrected for BCI. The relationship be-tween D2M activity and 1,2,4,5-tetrachlorobenzene is depicted inFig. 6. T3M was positioned in the centre of the plot, with valuesalong both PC1 and PC2 o |0.60|, and did not fulfil the criteria forfurther statistical treatment.
3.2.4. KidneyIn kidney, D1K activity (PC1¼-0.63) was positioned alongside
several OH-PCBs and PCBs (e.g. PCB-118, -138, 4′-OH-CB208, 4′-OH-CB172, 3′-OH-CB203/4′-OH-CB198) as well as trans-nonachlor(Fig. 1D; all with PC1o�0.60), indicating positive relationshipsbetween D1K activity and those compounds. Indeed, correlationanalysis confirmed that D1K activity was significantly positivelycorrelated with 4′-OH-CB172 (r2¼0.51, p¼0.037), 4-OH-CB202(r2¼0.60, p¼0.020) and PCB-118 (r2¼0.62, p¼0.017; Fig. 3C).D1K activity was also positioned opposite to OCPs such as 1.1-dichloro-2.2-bis(4-chlorophenyl)ethylene (p,p′-‘DDE), oxychlor-dane and heptachlor epoxide (Fig. 1D; all with PC140.60),demonstrating a possible negative association. Correlation analysisconfirmed that D1K activity was significantly negatively correlatedwith heptachlor epoxide (r2¼0.63, p¼0.017; Fig. 4B).
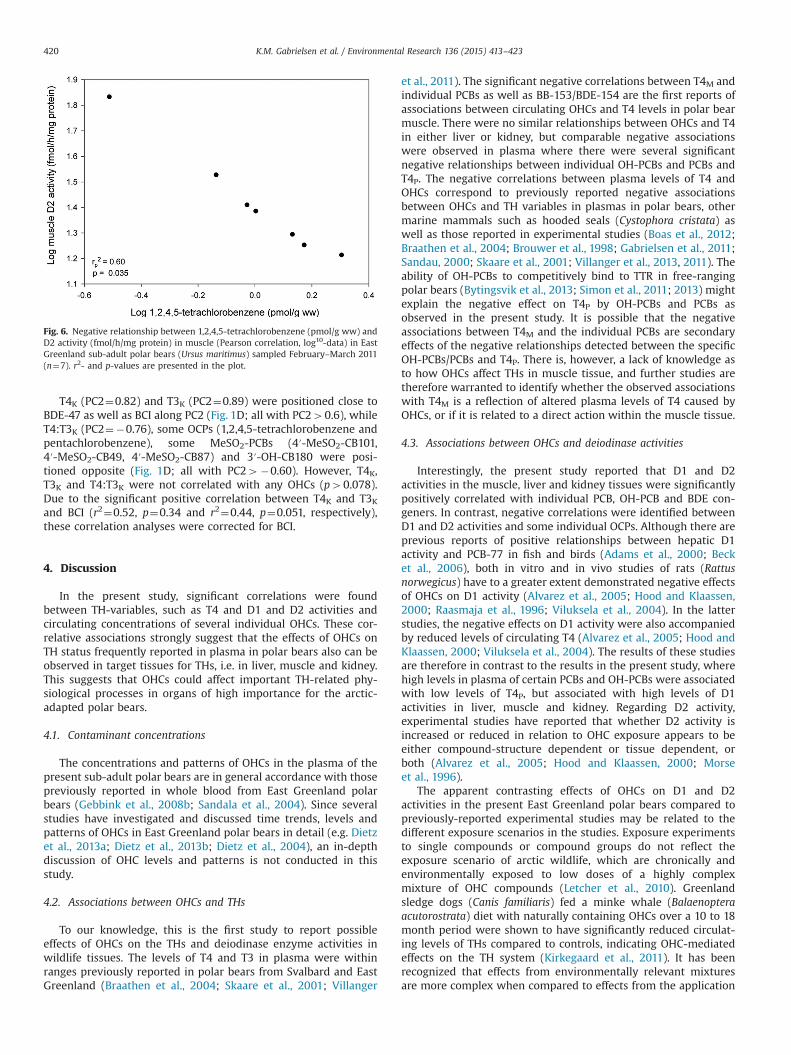
Fig. 6. Negative relationship between 1,2,4,5-tetrachlorobenzene (pmol/g ww) andD2 activity (fmol/h/mg protein) in muscle (Pearson correlation, log10-data) in EastGreenland sub-adult polar bears (Ursus maritimus) sampled February–March 2011(n¼7). r2- and p-values are presented in the plot.
K.M. Gabrielsen et al. / Environmental Research 136 (2015) 413–423420
T4K (PC2¼0.82) and T3K (PC2¼0.89) were positioned close toBDE-47 as well as BCI along PC2 (Fig. 1D; all with PC240.6), whileT4:T3K (PC2¼�0.76), some OCPs (1,2,4,5-tetrachlorobenzene andpentachlorobenzene), some MeSO2-PCBs (4′-MeSO2-CB101,4′-MeSO2-CB49, 4′-MeSO2-CB87) and 3′-OH-CB180 were posi-tioned opposite (Fig. 1D; all with PC24�0.60). However, T4K,T3K and T4:T3K were not correlated with any OHCs (p40.078).Due to the significant positive correlation between T4K and T3Kand BCI (r2¼0.52, p¼0.34 and r2¼0.44, p¼0.051, respectively),these correlation analyses were corrected for BCI.
4. Discussion
In the present study, significant correlations were foundbetween TH-variables, such as T4 and D1 and D2 activities andcirculating concentrations of several individual OHCs. These cor-relative associations strongly suggest that the effects of OHCs onTH status frequently reported in plasma in polar bears also can beobserved in target tissues for THs, i.e. in liver, muscle and kidney.This suggests that OHCs could affect important TH-related phy-siological processes in organs of high importance for the arctic-adapted polar bears.
4.1. Contaminant concentrations
The concentrations and patterns of OHCs in the plasma of thepresent sub-adult polar bears are in general accordance with thosepreviously reported in whole blood from East Greenland polarbears (Gebbink et al., 2008b; Sandala et al., 2004). Since severalstudies have investigated and discussed time trends, levels andpatterns of OHCs in East Greenland polar bears in detail (e.g. Dietzet al., 2013a; Dietz et al., 2013b; Dietz et al., 2004), an in-depthdiscussion of OHC levels and patterns is not conducted in thisstudy.
4.2. Associations between OHCs and THs
To our knowledge, this is the first study to report possibleeffects of OHCs on the THs and deiodinase enzyme activities inwildlife tissues. The levels of T4 and T3 in plasma were withinranges previously reported in polar bears from Svalbard and EastGreenland (Braathen et al., 2004; Skaare et al., 2001; Villanger
et al., 2011). The significant negative correlations between T4M andindividual PCBs as well as BB-153/BDE-154 are the first reports ofassociations between circulating OHCs and T4 levels in polar bearmuscle. There were no similar relationships between OHCs and T4in either liver or kidney, but comparable negative associationswere observed in plasma where there were several significantnegative relationships between individual OH-PCBs and PCBs andT4P. The negative correlations between plasma levels of T4 andOHCs correspond to previously reported negative associationsbetween OHCs and TH variables in plasmas in polar bears, othermarine mammals such as hooded seals (Cystophora cristata) aswell as those reported in experimental studies (Boas et al., 2012;Braathen et al., 2004; Brouwer et al., 1998; Gabrielsen et al., 2011;Sandau, 2000; Skaare et al., 2001; Villanger et al., 2013, 2011). Theability of OH-PCBs to competitively bind to TTR in free-rangingpolar bears (Bytingsvik et al., 2013; Simon et al., 2011; 2013) mightexplain the negative effect on T4P by OH-PCBs and PCBs asobserved in the present study. It is possible that the negativeassociations between T4M and the individual PCBs are secondaryeffects of the negative relationships detected between the specificOH-PCBs/PCBs and T4P. There is, however, a lack of knowledge asto how OHCs affect THs in muscle tissue, and further studies aretherefore warranted to identify whether the observed associationswith T4M is a reflection of altered plasma levels of T4 caused byOHCs, or if it is related to a direct action within the muscle tissue.
4.3. Associations between OHCs and deiodinase activities
Interestingly, the present study reported that D1 and D2activities in the muscle, liver and kidney tissues were significantlypositively correlated with individual PCB, OH-PCB and BDE con-geners. In contrast, negative correlations were identified betweenD1 and D2 activities and some individual OCPs. Although there areprevious reports of positive relationships between hepatic D1activity and PCB-77 in fish and birds (Adams et al., 2000; Becket al., 2006), both in vitro and in vivo studies of rats (Rattusnorwegicus) have to a greater extent demonstrated negative effectsof OHCs on D1 activity (Alvarez et al., 2005; Hood and Klaassen,2000; Raasmaja et al., 1996; Viluksela et al., 2004). In the latterstudies, the negative effects on D1 activity were also accompaniedby reduced levels of circulating T4 (Alvarez et al., 2005; Hood andKlaassen, 2000; Viluksela et al., 2004). The results of these studiesare therefore in contrast to the results in the present study, wherehigh levels in plasma of certain PCBs and OH-PCBs were associatedwith low levels of T4P, but associated with high levels of D1activities in liver, muscle and kidney. Regarding D2 activity,experimental studies have reported that whether D2 activity isincreased or reduced in relation to OHC exposure appears to beeither compound-structure dependent or tissue dependent, orboth (Alvarez et al., 2005; Hood and Klaassen, 2000; Morseet al., 1996).
The apparent contrasting effects of OHCs on D1 and D2activities in the present East Greenland polar bears compared topreviously-reported experimental studies may be related to thedifferent exposure scenarios in the studies. Exposure experimentsto single compounds or compound groups do not reflect theexposure scenario of arctic wildlife, which are chronically andenvironmentally exposed to low doses of a highly complexmixture of OHC compounds (Letcher et al., 2010). Greenlandsledge dogs (Canis familiaris) fed a minke whale (Balaenopteraacutorostrata) diet with naturally containing OHCs over a 10 to 18month period were shown to have significantly reduced circulat-ing levels of THs compared to controls, indicating OHC-mediatedeffects on the TH system (Kirkegaard et al., 2011). It has beenrecognized that effects from environmentally relevant mixturesare more complex when compared to effects from the application

K.M. Gabrielsen et al. / Environmental Research 136 (2015) 413–423 421
of standard toxicity testing protocols using high doses of singlecompounds or compound groups (Vandenberg et al., 2012). Thiscould contribute to explaining the differences observed betweenexperimental studies and the results in the present study. Theremight also be species differences between the experimentalanimals and arctic-adapted polar bears. Insight into TH regulationin marine mammals, which are subjected to very different envir-onmental conditions than experimental animals, including ex-treme low temperatures and long periods of food deprivation, isscarce and incomplete. This was exemplified by Martinez et al.(2013), who recently found an atypical TH response to fasting inelephant seal pups (Mirounga angustirostris) as compared to e.g.humans and rodents.
4.4. Possible mechanisms for disruption of THs and deiodinaseactivities
Whether OHCs can affect deiodinase activities directly orindirectly is not clear. Under normal physiological circumstances,D1 activity in all tissues, except in the thyroid gland, would beinduced by hyperthyroidism and inhibited by hypothyroidism(Köhrle, 1999). Such a corresponding reduction in circulating T4levels and e.g. hepatic D1 activity has been observed in severalexperimental studies, as mentioned above (Alvarez et al., 2005;Hood and Klaassen, 2000; Viluksela et al., 2004). Also, Vilukselaet al. (2004) suggested that the reported lowered D1 activity wassecondary to effects on circulating T4 in TCDD-treated rats andthus, a result of the changes in T4 levels. However, this was not thecase in the present study, where low levels of T4P, but high levelsof D1 activity in liver, muscle and kidney tissues, were associatedwith high levels of certain PCBs and OH-PCBs in plasma. Withregards to effects on D2 activity from OHCs, increased activity ofthis enzyme has mainly been suggested as a compensatorymechanism for the maintenance of T3 levels in critical tissuessuch as the brain following contaminant-induced reduction ofcirculating T4 levels (Hood and Klaassen, 2000; Köhrle, 1999;Morse et al., 1996). This is because D2 activity is induced byhypothyroidism under normal physiological conditions (Köhrle,1999). However, Hood and Klaassen (2000) stated that theimportance of D2 activity to maintain T3 concentrations aftercontaminant-induced changes in T4 remains unclear. It should alsobe noted that thyroid gland D1 activity, which increases inhypothyroidism, together with the D1 activity in the livers of ratshas been suggested to maintain circulating T3 levels despitereduced circulating T4 levels following hexachlorobenzene expo-sure (Alvarez et al., 2005). In contrast to the studies indicatingalterations in deiodinase activity as secondary response, a study byWade et al. (2002) reported that D1 activity appeared to be a moresensitive endpoint than circulating levels of TH in rats after sub-chronic exposure to a complex mixture of TH disrupting chemicals.This supports the reasoning that D1 and D2 activities indeed canbe affected by OHCs without it being merely a secondary responseto alterations in TH levels.
Both OH-PCBs and OH-PBDEs have been shown to bindcompetitively to T4-binding proteins such as TR and TTR(Bytingsvik et al., 2013; Gauger et al., 2007; Gutleb et al., 2010;Meerts et al., 2000; Simon et al., 2011; 2013), and the suggestedinvolvement of D1 in debromination of PBDEs in teleosts indicatesa possible competitive binding interaction between PBDEs and THsfor D1 (Noyes et al., 2011; Stapleton et al., 2004). For the presentstudy, if D1 activities were in fact increased in response to OHCexposure, one could hypothesize an elevated conversion of T4 toT3 with subsequent decreased T4 levels and increased T3 levels.On the other hand, if the enzymatic catalytic activity of D1 isimpaired due to OHC exposure, one of the hypothetical conse-quences is reduced tissue bioavailability of the bioactive T3 due to
reduced deiodination of T4 (Wade et al., 2002). Although T4P andT4M were both negatively associated with individual OHCs in thepresent polar bears, there were no effects on circulating nor tissuelevels of T3. This suggests that the associations between deiodi-nase activity and OHCs reported herein did not jeopardize thebiological activity of T3. This is also consistent with several studiesthat reported non-altered circulating T3 levels despite a reductionin T4 levels and D1 activity following OHC exposure, implying thatthe reduction in T4 levels and D1 activity was not a rate-limitingstep supplying the plasma pool with T3 to maintain homeostasis(Raasmaja et al., 1996; Viluksela et al., 2004). The present studyindicates that this conclusion may be valid for other tissues as wellas plasma. However, it was presently not possible to determine ifthe responses observed for T4P and T4M caused, or were a result of,the observed responses in D1 activity in the present polar bears.Nevertheless, the tightly regulated balance between activation anddeactivation of THs within the targets tissues might still becompromised when deiodinase activity is affected (Brouweret al., 1998). Further studies are therefore needed to elucidatethe order of events to determine whether changes in D1 and D2activities are causative or secondary compensatory effects uponOHC exposure.
Through which mechanisms various compounds might affectthe TH-system, and if they act through additive, antagonistic orsynergistic mechanisms, is to a large extent still unresolved(Crofton et al., 2005; Crofton, 2008). Effects on deiodinase en-zymes could be caused by alteration of gene expression, binding tothe protein, altered TH signaling or other unknown mechanisms(Wade et al., 2002). Effects on enzyme function could also occurdirectly through tissue damage. Tissue effects in liver related tocomplex OHC exposure, where OCPs such as heptachlor epoxidewere included, have been investigated and suggested in EastGreenland polar bears (Sonne et al., 2005) as well as in farmedfoxes (Vulpes lagopus) fed a whale blubber diet rich in OHCs(Sonne et al., 2008). Heptachlor epoxide and its parent compoundheptachlor are primarily regarded as carcinogenic and liver toxiccompounds (ATSDR, 2007; Reuber, 1987). Chlorinated benzenesare also reported to be liver toxic compounds (Alvarez et al., 2005;den Besten et al., 1991). These chlorinated pesticides were allnegatively related to D1 activity in various tissues in the presentstudy. The OH-metabolites of chlorobenzenes however, were alsoreported to reduce circulating TH levels (Alvarez et al., 2005; denBesten et al., 1991), while Villanger et al. (2011) found adiposeconcentrations of heptachlor epoxide to be positively associatedwith circulating total T4 in female polar bears. Thus, whether theobserved negative correlations between the chlorinated pesticidesand D1 activity occur through TH disruptive mechanisms oroverall tissue toxicity is not clear. The complexity of the contami-nant burdens in wildlife species complicates the attempts to shedlight over contaminant-induced mechanisms in field-basedstudies.
4.5. Different effects may be due to different chemical structurs
In the present study, specific OH-PCBs and PCBs congeners, aswell as BDE-47 were positively related to D1 and D2 activities andnegatively associated with T4P and T4M. PCBs and PBDEs con-geners, including their OH-metabolites, are often compared due totheir structural and physiochemical similarities. This also includestheir role as TH disruptors (Boas et al., 2012; Darnerud et al.,2001). The OCPs (two chlorinated benzenes and two epoxides)that were negatively associated with D1 and D2 activities orpositively associated with T4M, are more structurally related toeach other than to PCBs and PBDEs. The apparent differences inthe effects between these groups of OHCs could be related to theirchemical structures. As seen by their opposite positioning in the

K.M. Gabrielsen et al. / Environmental Research 136 (2015) 413–423422
four loading plots (Fig. 1A–D), negative relationships betweenseveral OH-PCBs and PCBs (e.g. 4′-OH-CB172, 4′-OH-CB202 andPCB-183) and some OCPs (e.g. heptachlor epoxide, 1,2,4,5-tetra-chlorobenzene and pentachlorobenzene) were present. This mayhave confounded the interpretation of the reported negative andpositive relationships between individual OHCs and D1 and D2activity. Experimental studies are needed in order to identifywhich of the suggested contaminant groups are responsible fordisruption of deiodinase activities in target tissues for TH-action,and the mechanisms by which they act.
5. Conclusion
The present study reports associations between individual PCBand OH-PCB concentrations and T4P, supporting previous studiesreporting that these compounds are capable of reducing circulat-ing levels of T4. In addition, several negative relationships thatwere discovered between individual PCBs and T4M were found tobe significant. Relationships between individual OHCs and D1 andD2 activities were also reported. It was, however, not possible todrawn any conclusions regarding the role of deiodinase activity inTH disruption in free-ranging East Greenland polar bears. Thepresent study shows that tissue endpoints such as TH levels anddeiodinase activities can be sensitive targets of exposure to TH-disrupting compounds, and that tissue-specific responses in targetorgans should be considered when assessing TH disruption inwildlife studies. Statistical relationships are not evidence per se ofbiological cause-effect relationships, and the authors realize thelimitations of the small sample size in the present study. Thus, weemphasize that the present data set does not allow for any firmconclusions with respect to effects of OHCs on the analysed THvariables. Still, this study presents new data that might add to theweight of evidence that several individual OHCs are capable ofaffecting the TH system. Future studies using a larger sample sizeare necessary to elucidate the effects of individual contaminantson the various TH endpoints.
Acknowledgements
The authors would like to thank all those who contributed tothe field work in the Scoresby Sound area in East Greenland in2011, including the local hunters who provided the polar bears forthis study through their aboriginal hunting quotas. The field workwas funded by the Danish Co-operation for Environment in theArctic (DANCEA). The analysis of thyroid hormones and deiodinaseactivitites in tissues was supported by research grants SAF2012-32491 from MINECO and S2010/BMD-2423 from CAM, Spain. Theauthors would like to thank Sigga Joensen at Aarhus University forparticipating in the ageing of the polar bears and Grethe StavikEggen at the Norwegian University of Science and Technology(NTNU) for help with the thyroid hormone analysis in plasma. Theauthors would also like to thank the rest of the staff at the LetcherLabs-OCRL at EC-NWRC, Canada for help with the OHC analysis.This study was funded by NTNU.
Appendix A. Supporting Information
Supplementary data associated with this article can be found inthe online version at http://dx.doi.org/10.1016/j.envres.2014.09.019.
References
ATSDR, 2007. Toxicological profile for heptachlor and heptachlor epoxide. Depart-ment of Health and Human Services, Public Health Service, Agency for ToxicSubstances and Disease Registry, Atlanta, GA, U.S.
Adams, B.A., Cyr, D.G., Eales, J.G., 2000. Thyroid hormone deiodination in tissues ofAmerican plaice, Hippoglossoides platessoides: characterization and short-termresponses to polychlorinated biphenyls (PCBs) 77 and 126. Comp. Biochem.Physiol. Part C Toxicol. Pharmacol. 127, 367–378.
Alvarez, L., Hernández, S., Martinez-De-Mena, R., Kolliker-Frers, R., Obregón, M.J.,Kleiman De Pisarev, D.L., 2005. The role of type I and type II 5′ deiodinases onhexachlorobenzene-induced alteration of the hormonal thyroid status. Toxicol-ogy 207, 349–362.
Beck, V., Roelens, S.A., Darras, V.M., 2006. Exposure to PCB 77 induces tissue-dependent changes in iodothyronine deiodinase activity patterns in theembryonic chicken. Gen. Comp. Endocrinol. 148, 327–335.
Bernal, J., 2005. Thyroid hormones and brain development. Vitam. Horm. 71,95–122.
Bianco, A.C., Kim, B.W., 2006. Deiodinases: implications of the local control ofthyroid hormone action. J. Clin. Investig. 116, 2571–2579.
Bianco, A.C., Salvatore, D., Gereben, B., Berry, M.J., Larsen, P.R., 2002. Biochemistry,cellular and molecular biology, and physiological roles of the iodothyronineselenodeiodinases. Endocr. Rev. 23, 38–89.
Boas, M., Feldt-Rasmussen, U., Main, K.M., 2012. Thyroid effects of endocrinedisrupting chemicals. Mol. Cell. Endocrinol. 355, 240–248.
Boon, J.P., Oostingh, I., van der Meer, J., Hillebrand, M.T.J., 1994. A model for thebioaccumulation of chlorobiphenyl congeners in marine mammals. Eur. J.Pharm. Environ. 3, 237–251.
Braathen, M., Derocher, A., Wiig, O., Sormo, E., Lie, E., Skaare, J., Jenssen, B., 2004.Relationships between PCBs and thyroid hormones and retinol in female andmale polar bears. Environ. Health Perspect. 112, 826–833.
Brouwer, A., Morse, D., Lans, M., Schuur, A., Murk, A., Klasson-Wehler, E., Bergman,Å., Visser, T.J., 1998. Interactions of persistent environmental organohalogenswith the thyroid hormone system: Mechanisms and possible consequences foranimal and human health. Toxicol. Ind. Health 14, 59–84.
Bytingsvik, J., 2012. Organohalogenated contaminants (OHCs) in polar bear mother-cub pairs from Svalbard, Norway. Norwegian University of Science andTechnology (Doctoral thesis).
Bytingsvik, J., Simon, E., Leonards, P.E.G., Lamoree, M., Lie, E., Aars, J., Derocher, A.E.,Wiig, Ø., Jenssen, B.M., Hamers, T., 2013. Transthyretin-binding activity ofcontaminants in blood from polar bear (Ursus maritimus) cubs. Environ. Sci.Technol. 47, 4778–4786.
Cattet, M.R.L., Caulkett, N.A., Obbard, M.E., Stenhouse, G.B., 2002. A body-conditionindex for ursids. Can. J. Zool. 80, 1156–1161.
Cheng, S.-Y., Leonard, J.L., Davis, P.J., 2010. Molecular aspects of thyroid hormoneactions. Endocr. Rev. 31, 139–170.
Colborn, T., 2004. Neurodevelopment and endocrine disruption. Environ. HealthPerspect. 112, 944–949.
Crofton, K., Craft, E., Hedge, J., Gennings, C., Simmons, J., Carchman, R., 2005.Thyroid-hormone-disrupting chemicals: evidence for dose-dependent additiv-ity or synergism. Environ. Health Perspect. 113, 1549–1554.
Crofton, K.M., 2008. Thyroid disrupting chemicals: mechanisms and mixtures. Int. J.Androl. 31, 209–223.
Darnerud, P.O., Eriksen, G.S., Johannesson, T., Larsen, P.B., Viluksela, M., 2001.Polybrominated diphenyl ethers: occurrence, dietary exposure, and toxicology.Environ. Health Perspect 109, 49–68.
den Besten, C., Vet, J.J.R.M., Besselink, H.T., Kiel, G.S., van Berkel, B.J.M., Beems, R.,van Bladeren, P.J., 1991. The liver, kidney, and thyroid toxicity of chlorinatedbenzenes. Toxicol. Appl. Pharmacol. 111, 69–81.
Dentice, M., Marsili, A., Zavacki, A., Larsen, P.R., 2013. The deiodinases and thecontrol of intracellular thyroid hormone signaling during cellular differentia-tion. Biochim. Biophys. Acta (BBA) Gen. Subj. 1830,, 3937–33945.
Derocher, A.E., Wiig, Ø., 2002. Postnatal growth in body length and mass of polarbears (Ursus maritimus) at Svalbard. J. Zool. 256, 343–349.
Dietz, R., Heide-Jørgensen, M.P., Härkönen, T., Teilmann, J., Valentin, N., 1991. Agedetermination of European harbour seal Phoca vitulina L. Sarsia 76, 17–21.
Dietz, R., Rigét, F.F., Sonne, C., Letcher, R., Born, E.W., Muir, D.C.G., 2004. Seasonaland temporal trends in polychlorinated biphenyls and organochlorine persti-cides in East Greenland polar bears (Ursus maritimus), 1999–2001. Sci. TotalEnviron. 331, 107–124.
Dietz, R., Rigét, F.F., Sonne, C., Born, E.W., Bechshøft, T., McKinney, M.A., Drimmie, R.J., Muir, D.C.G., Letcher, R.J., 2013a. Three decades (1983–2010) of contaminanttrends in East Greenland polar bears (Ursus maritimus). Part 2: brominatedflame retardants. Environ. Int. 59, 494–500.
Dietz, R., Rigét, F.F., Sonne, C., Born, E.W., Bechshøft, T., McKinney, M.A., Letcher, R.J.,2013b. Three decades (1983–2010) of contaminant trends in East Greenlandpolar bears (Ursus maritimus). Part 1: legacy organochlorine contaminants.Environ. Int. 59, 485–493.
Eriksson, L., Sjostrom, M., Wold, S., 1992. Rational ranking of chemicals according toenvironmental risk—an illustration using multivariate biological profiling ofhalogenated aliphatic-hydrocarbons. Chemometr. Intell. Lab. 14, 245–252.
Forbes, V.E., Forbes, T.E., 1994. Ecotoxicology in Theory and Practice.Gabrielsen, K.M., Villanger, G.D., Lie, E., Karimi, M., Lydersen, C., Kovacs, K.M.,
Jenssen, B.M., 2011. Levels and patterns of hydroxylated polychlorinated

K.M. Gabrielsen et al. / Environmental Research 136 (2015) 413–423 423
biphenyls (OH-PCBs) and their associations with thyroid hormones in hoodedseal (Cystophora cristata) mother–pup pairs. Aquat. Toxicol. 105, 482–491.
Gauger, K., Giera, S., Sharlin, D., Bansal, R., Iannacone, E., Zoeller, R., 2007.Polychlorinated biphenyls 105 and 118 form thyroid hormone receptor agonistsafter cytochrome P4501A1 activation in rat pituitary GH3 cells. Environ. HealthPerspect. 115, 1623–1630.
Gebbink, W., Sonne-Hansen, C., Dietz, R., Kirkegaard, M., Born, E.W., Muir, D.C.G.,Letcher, R.J., 2008a. Target tissue selectivity and burdens of diverse classes ofbrominated and chlorinated contaminants in polar bears (Ursus maritimus)from East Greenland. Environ. Sci. Technol. 42, 752–759.
Gebbink, W., Sonne, C., Dietz, R., Kirkegaard, M., Riget, F., Born, E., 2008b. Tissue-specific congener composition of organohalogen and metabolite contaminantsin East Greenland polar bears (Ursus maritimus). Environ. Pollut. 152, 621–629.
Gereben, B., Zavacki, A.M., Ribich, S., Kim, B.W., Huang, S.A., Simonides, W.S., Zeöld,A., Bianco, A.C., 2008. Cellular and molecular basis of deiodinase-regulatedthyroid hormone signaling. Endocr. Rev. 29, 898–938.
Gutleb, A., Cenijn, P., van Velzen, M., Lie, E., Ropstad, E., Skaare, J., 2010. In vitroassay shows that PCB metabolites completely saturate thyroid hormonetransport capacity in blood of wild polar bears (Ursus maritimus). Environ.Sci. Technol. 44, 3149–3154.
Hensel, R.J., Sorensen, F.E., 1980. Age determination of live polar bears. In:Proceedings of the International Conference on Bear Research and Manage-ment, vol. 4, pp. 93–100.
Hood, A., Klaassen, C.D., 2000. Effects of microsomal enzyme inducers on outer-ringdeiodinase activity toward thyroid hormones in various rat tissues. Toxicol.Appl. Pharmacol. 163, 240–248.
Hulbert, A.J., 2000. Thyroid hormones and their effects: a new perspective. Biol.Rev. 75, 519–631.
Jenssen, B., 2006. Endocrine-disrupting chemicals and climate change: a worst-casecombination for Arctic marine mammals and seabirds? Environ. HealthPerspect. 114, 76–80.
Jolliffe, I.T., 2002. Principal Component Analysis. Springer-Verlag, NewYork.Kirkegaard, M., Sonne, C., Dietz, R., Letcher, R.J., Jensen, A.L., Stige Hansen, S., Munro
Jenssen, B., Grandjean, P., 2011. Alterations in thyroid hormone status inGreenland sledge dogs exposed to whale blubber contaminated with organo-halogen compounds. Ecotoxicol. Environ. Saf. 74, 157–163.
Köhrle, J., 1999. Local activation and inactivation of thyroid hormones: thedeiodinase family. Mol. Cell. Endocrinol. 151, 103–119.
Lavado-Autric, R., Calvo, R.M., Martinez de Mena, R., Morreale de Escobar, G.,Obregon, M.-J., 2013. Deiodinase activities in thyroids and tissues of iodine-deficient female rats. Endocrinology 154, 529–536.
Letcher, R., Klasson-Wehler, E., Bergman, Å., 2000. Methyl sulfone and hydroxylatedmetabolites of polychlorinated biphenyls. In: Hutzinger, O. (Ed.), The Handbookof Environmental Chemistry. Springer-Verlag, Berlin.
Letcher, R., Bustnes, J., Dietz, R., Jenssen, B., Jorgensen, E., Sonne, C., Verreault, J.,Vijayan, M., Gabrielsen, G., 2010. Exposure and effects assessment of persistentorganohalogen contaminants in Arctic wildlife and fish. Sci. Total Environ. 408,2995–3043.
Martinez, B., Soñanez-Organis, J.G., Vázquez-Medina, J.P., Viscarra, J.A., Mackenzie,D.S., Crocker, D.E., Ortiz, R.M., 2013. Prolonged food deprivation increases mrnaexpression of deiodinase 1 and 2, and thyroid hormone receptor β-1 in afasting-adapted mammal. J. Exp. Biol. 216, 4647–4654.
McKinney, M.A., Letcher, R.J., Aars, J., Born, E.W., Branigan, M., Dietz, R., Evans, T.J.,Gabrielsen, G.W., Muir, D.C.G., Peacock, E., Sonne, C., 2011a. Regional contam-ination versus regional dietary differences: understanding geographic variationin brominated and chlorinated contaminant levels in polar bears. Environ. Sci.Technol. 45, 896–902.
McKinney, M.A., Letcher, R.J., Aars, J., Born, E.W., Branigan, M., Dietz, R., Evans, T.J.,Gabrielsen, G.W., Peacock, E., Sonne, C., 2011b. Flame retardants and legacycontaminants in polar bears from Alaska, Canada, East Greenland and Svalbard,2005–2008. Environ. Int. 37, 365–374.
McKinney, M.A., Iverson, S.J., Fisk, A.T., Sonne, C., Rigét, F.F., Letcher, R.J., Arts, M.T.,Born, E.W., Rosing-Asvid, A., Dietz, R., 2013. Global change effects on the long-term feeding ecology and contaminant exposures of East Greenland polarbears. Glob. Change Biol. 19, 2360–2372.
Meerts, I., van Zanden, J., Luijks, E., van Leeuwen-Bol, I., Marsh, G., Jakobsson, E.,Bergman, A., Brouwer, A., 2000. Potent competitive interactions of somebrominated flame retardants and related compounds with human transthyretinin vitro. Toxicol. Sci. 56, 95–104.
Moran, M.D., 2003. Arguments for rejecting the sequential bonferroni in ecologicalstudies. Oikos 100, 403–405.
Morreale de Escobar, G., Pastor, R., Obregon, M.J., Escobar del Rey, F., 1985. Effects ofmaternal hypothyroidism on the weight and thyroid hormone content of ratembryonic tissues, before and after onset of fetal thyroid function. Endocrinol-ogy 117, 1890–1900.
Morse, D.C., Wehler, E.K., Wesseling, W., Koeman, J.H., Brouwer, A., 1996. Altera-tions in rat brain thyroid hormone status following pre- and postnatal exposureto polychlorinated biphenyls (Aroclor 1254). Toxicol. Appl. Pharmacol. 136,269–279.
Noyes, P.D., Hinton, D.E., Stapleton, H.M., 2011. Accumulation and debromination ofdecabromodiphenyl ether (BDE-209) in juvenile fathead minnows (Pimephalespromelas) induces thyroid disruption and liver alterations. Toxicol. Sci. 122,265–274.
Raasmaja, A., Viluksela, M., Rozman, K.K., 1996. Decreased liver type I 5′-deiodinaseand increased brown adipose tissue type II 5′-deiodinase activity in 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-treated Long Evans rats. Toxicology 114,199–205.
Reuber, M.D., 1987. Carcinogenicity of heptachlor and heptachlor epoxide. J.Environ. Pathol. Toxicol. Oncol. 7, 85–114.
Rosing-Asvid, A., Born, E., Kingsley, M., 2002. Age at sexual maturity of males andtiming of the mating season of polar bears (Ursus maritimus) in Greenland.Polar Biol. 25, 878–883.
Sandala, G., Sonne-Hansen, C., Dietz, R., Muir, D., Valters, K., Bennett, E.,.E., 2004.Hydroxylated and methyl sulfone PCB metabolites in adipose and whole bloodof polar bear (Ursus maritimus) from East Greenland. Sci. Total Environ. 331,125–141.
Sandau, C., 2000. Analytical Chemistry of Hydroxylated Metabolites of PCBs andOther Halogenated Phenolic Compounds in Blood and Their Relationship toThyroid Hormone and Retinol Homeostasis in Humans and Polar Bears.Carleton University (Doctoral thesis).
Simon, E., Bytingsvik, J., Jonker, W., Leonards, P.E.G., De Boer, J., Jenssen, B.M., Lie, E.,Aars, J., Hamers, T., Lamoree, M.H., 2011. Blood plasma sample preparationmethod for the assessment of thyroid hormone-disrupting potency in effect-directed analysis. Environ. Sci. Technol. 45, 7936–7944.
Simon, E., van Velzen, M., Brandsma, S.H., Lie, E., Løken, K., De Boer, J., Bytingsvik, J.,Jenssen, B.M., Aars, J., Hamers, T., Lamoree, M.H., 2013. Effect-directed analysisto explore the polar bear exposome: Identification of thyroid hormonedisrupting compounds in plasma. Environ. Sci. Technol. 47, 8902–8912.
Skaare, J.U., Bernhoft, A., Wiig, O., Norum, K.R., Haug, E., Eide, D.M., Derocher, A.E.,2001. Relationships between plasma levels of organochlorines, retinol andthyroid hormones from polar bears (Ursus maritimus) at Svalbard. J. Toxicol.Environ. Health Part A 62, 227–241.
Sonne, C., Dietz, R., Leifsson, P.S., Born, E.W., Letcher, R.J., Kirkegaard, M., Muir, D.C.G., Riget, F., Hyldstrup, L., 2005. Do organohalogen contaminants contribute tohistopathology in liver from East Greenland polar bears (Ursus maritimus)?Environ. Health Perspect. 113, 1569–1574.
Sonne, C., Wolkers, H., Leifsson, P.S., Jenssen, B.M., Fuglei, E., Ahlstrøm, Ø., Dietz, R.,Kirkegaard, M., Muir, D.C.G., Jørgensen, E., 2008. Organochlorine-inducedhistopathology in kidney and liver tissue from Arctic fox (Vulpes lagopus).Chemosphere 71, 1214–1224.
Stapleton, H.M., Alaee, M., Letcher, R.J., Baker, J.E., 2004. Debromination of theflame retardant decabromodiphenyl ether by juvenile carp (Cyprinus carpio)following dietary exposure. Environ. Sci. Technol. 38, 112–119.
Stirling, I., 2002. Polar bear Ursus maritimus. In: Perrin, W.F., Bernd, W., Thewissen,J.G.M. (Eds.), Encyclopedia of Marine Mammals San Diego. Academic Press,California, USA.
Umetrics, 2008. User guide to Simca-pþ . Version 12. [Online]. Available: ⟨http://www.umetrics.com⟩ [accessed 12.01.11].
Vandenberg, L.N., Colborn, T., Hayes, T.B., Heindel, J.J., Jacobs, D.R., Lee, D.-H., Shioda,T., Soto, A.M., Vom Saal, F.S., Welshons, W.V., Zoeller, R.T., Myers, J.P., 2012.Hormones and endocrine-disrupting chemicals: low-dose effects and nonmo-notonic dose responses. Endocr. Rev. 33, 378–455.
Villanger, G.D., Jenssen, B.M., Fjeldberg, R.R., Letcher, R.J., Muir, D.C.G., Kirkegaard,M., Sonne, C., Dietz, R., 2011. Exposure to mixtures of organohalogen con-taminants and associative interactions with thyroid hormones in East Green-land polar bears (Ursus maritimus). Environ. Int. 37, 694–708.
Villanger, G.D., Gabrielsen, K.M., Kovacs, K.M., Lydersen, C., Lie, E., Karimi, M.,Sørmo, E.G., Jenssen, B.M., 2013. Effects of complex organohalogen contaminantmixtures on thyroid homeostasis in hooded seal (Cystophora cristata) mother–pup pairs. Chemosphere 92, 828–842.
Viluksela, M., Raasmaja, A., Lebofsky, M., Stahl, B.U., Rozman, K.K., 2004. Tissue-specific effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) on the activity of5′-deiodinases I and II in rats. Toxicol. Lett. 147, 133–142.
Wade, M.G., Parent, S., Finnson, K.W., Foster, W., Younglai, E., Mcmahon, A., Cyr, D.G., Hughes, C., 2002. Thyroid toxicity due to subchronic exposure to a complexmixture of 16 organochlorines, lead, and cadmium. Toxicol. Sci. 67, 207–218.
Wiseman, S.B., Wan, Y., Chang, H., Zhang, X., Hecker, M., Jones, P.D., Giesy, J.P., 2011.Polybrominated diphenyl ethers and their hydroxylated/methoxylated analogs:environmental sources, metabolic relationships, and relative toxicities. Mar.Pollut. Bull. 63, 179–188.
Zoeller, R.T., Dowling, A.L.S., Herzig, C.T.A., Iannacone, E.A., Gauger, K.J., Bansal, R.,2002. Thyroid hormone, brain development, and the environment. Environ.Health Perspect. 110, 355–361.
Zoeller, R.T., Tan, S., Tyl, R., 2007. General background on the hypothalamic-pituitary-thyroid (HPT) axis. Crit. Rev. Toxicol. 37, 11–53.




















