
POSSIBLE IMMUNOTOXIC EFFECTS OF ORGANOCHLORINES IN POLAR BEARS ( URSUS MARITIMUS ) AT SVALBARD
17
ELSEVIER PII: S0269-7491(96)00122-4 Environmental Pollution, Vol. 95, No. 2, pp. 159-175, 1997 © 1997 Elsevier Science Ltd All fights reserved. Printed in Great Britain 0269-7491/97 $17.00+0.00 ORGANOCHLORINES IN POLAR BEARS (URSUS MARITIMUS) AT SVALBARD Aksel Bernhoft, a Oystein Wiig b & Janneche Utne Skaare ac aNational Veterinary Institute, PO Box 8156 Dep., N-0033 Oslo, Norway bNorwegian Polar Institute, PO Box 5027 Majorstua, N-0301 Oslo, Norway~Zoological Museum, University of Oslo, Sars gt. 1, N-0562 Oslo, Norway CNorwegian College of Veterinary Medicine, PO Box 8146 Dep., N-0033 Oslo, Norway (Received 18 May 1996; accepted 1 October 1996) Abstract A comprehensive survey on organochlorine (OC) con- taminants in polar bears at Svalbard has been undertaken. Subcutaneous tissue, blood and milk have been sampled from anesthetized free-ranging bears of both sexes and different ages in the period from 1990 to 1994. A number of sexually mature females have been fitted with satellite transmitters which make it possible to follow their repro- ductive behaviour pattern. We report on contamination levels and pattern of PCB congeners, chlordanes, DDT- compounds, HCB and HCH-isomers in the various tis- sues. The relation of age, sex and reproductive status to OC contamination has been described, and the capacity of polar bears in metabolising OCs has been discussed from the isomer/metabolite composition of residues. Finally, the possible association between OC contamination and reproduction success has been assessed. The PCB levels present in polar bears at Svalbard are extremely high. Especially high levels of higher chlorinated PCBs are found, and they accumulate with age, particularly in males. With females, considerable amounts of OCs are transferred to the offspring via milk. However, more effi- cient OC transfer between subcutaneous depot lipid and circulatory lipids than from the circulatory system to milk is found particularly for the most lipophilic compounds. The OC pattern in suckling yearlings reflects the low transfer of the highest chlorinated PCBs into maternal milk. The levels of most other OCs, however, are higher in depot lipid of yearlings than in that of their mothers. The polar bears have high capacity to metabolise several OCs. This may protect them against toxic action of the con- taminants. No relation between OC levels in females and their ability to get pregnant is found in this preliminary investigation on OC effects on the polar bears at Sval- bard. © 1997 Elsevier Science Ltd. All rights reserved INTRODUCTION Large quantities of organochlorines (OCs) including pesticides like chlordanes, dichlorodiphenyltri- chloroethane (DDT), hexachlorocyclohexanes (HCHs), hexachlorobenzene (HCB) and toxaphene related 159 compounds, and industrial chemicals like polychlorina- ted biphenyls (PCBs), have been released to the global environment the last 3-4 decades. Long-range trans- port, via air in particular, takes these persistent and lipophilic compounds to the Arctic (Bowes & Jonkel, 1975; Clausen & Berg, 1976; Muir et al., 1988; Muir et al., 1992; Oehme, 1991; Barrie et al., 1992; Hargrave et al., 1992; Zhu & Norstrom, 1993; Norstrom & Muir, 1994). In arctic areas, particularly in the Western Hemisphere, considerable levels of PCBs and chlor- danes have been found in polar bears (Norstrom et al., 1988; Norstrom et al., 1996; Norheim et al., 1992), the top predator of the Arctic marine food chain. These organochlorines are known to generate adverse effects after chronic administration to laboratory mammals. Both the pesticides and the PCBs may produce a range of effects including neuro, reproductional and immuno toxicities (Hayes, 1982a,b; Safe, 1984; Tilson et al., 1990; Ahlborg et al., 1992; Peterson et al., 1993; Ecobi- chon, 1996). Norheim et al. (1992) found PCB levels in polar bears at Svalbard close to those connected with reproductional disorders in Baltic seals (Helle et al., 1976; Bergman & Olsson, 1985; Olsson et al., 1992). Polar bears mate in the spring (Wiig et al., 1992). Fertilised eggs do not implant before September- October, about the same time that the pregnant female enters the den (Ramsay & Stifling, 1988). According to Sandell (1990), species with delayed implantation appear especially susceptible to deleterious reproductive effects from organochlorine pollution. Polar bears have delayed implantation and Derocher (1991) suggested that organochlorine-induced effects on reproduction may be one factor in the decline in reproductive perfor- mance of the Western Hudson Bay polar bears. Cubs are born around Christmas and the female emerges from the den usually with two cubs in March-April. She has then fasted for about six months. Polar bear milk has a high fat content (Arnould & Ramsay, 1994) and lactation seems to occur more or less until weaning at about 2 ½ years (Oehme et al., 1995). The objectives of this study were to provide compre- hensive baseline data on important OC contaminants in polar bears at Svalbard and to assess the possible
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of POSSIBLE IMMUNOTOXIC EFFECTS OF ORGANOCHLORINES IN POLAR BEARS ( URSUS MARITIMUS ) AT SVALBARD

ELSEVIER P I I : S 0 2 6 9 - 7 4 9 1 ( 9 6 ) 0 0 1 2 2 - 4
Environmental Pollution, Vol. 95, No. 2, pp. 159-175, 1997 © 1997 Elsevier Science Ltd
All fights reserved. Printed in Great Britain 0269-7491/97 $17.00+0.00
ORGANOCHLORINES IN POLAR BEARS (URSUS MARITIMUS) AT SVALBARD
Aksel Bernhoft, a Oystein Wiig b & Janneche Utne Skaare ac
aNational Veterinary Institute, PO Box 8156 Dep., N-0033 Oslo, Norway bNorwegian Polar Institute, PO Box 5027 Majorstua, N-0301 Oslo, Norway~Zoological Museum, University of Oslo, Sars gt. 1,
N-0562 Oslo, Norway CNorwegian College of Veterinary Medicine, PO Box 8146 Dep., N-0033 Oslo, Norway
(Received 18 May 1996; accepted 1 October 1996)
Abstract A comprehensive survey on organochlorine (OC) con- taminants in polar bears at Svalbard has been undertaken. Subcutaneous tissue, blood and milk have been sampled from anesthetized free-ranging bears of both sexes and different ages in the period from 1990 to 1994. A number of sexually mature females have been fitted with satellite transmitters which make it possible to follow their repro- ductive behaviour pattern. We report on contamination levels and pattern of PCB congeners, chlordanes, DDT- compounds, HCB and HCH-isomers in the various tis- sues. The relation of age, sex and reproductive status to OC contamination has been described, and the capacity of polar bears in metabolising OCs has been discussed from the isomer/metabolite composition of residues. Finally, the possible association between OC contamination and reproduction success has been assessed. The PCB levels present in polar bears at Svalbard are extremely high. Especially high levels of higher chlorinated PCBs are found, and they accumulate with age, particularly in males. With females, considerable amounts of OCs are transferred to the offspring via milk. However, more effi- cient OC transfer between subcutaneous depot lipid and circulatory lipids than from the circulatory system to milk is found particularly for the most lipophilic compounds. The OC pattern in suckling yearlings reflects the low transfer of the highest chlorinated PCBs into maternal milk. The levels of most other OCs, however, are higher in depot lipid of yearlings than in that of their mothers. The polar bears have high capacity to metabolise several OCs. This may protect them against toxic action of the con- taminants. No relation between OC levels in females and their ability to get pregnant is found in this preliminary investigation on OC effects on the polar bears at Sval- bard. © 1997 Elsevier Science Ltd. All rights reserved
INTRODUCTION
Large quantities of organochlorines (OCs) including pesticides like chlordanes, dichlorodiphenyltri- chloroethane (DDT), hexachlorocyclohexanes (HCHs), hexachlorobenzene (HCB) and toxaphene related
159
compounds, and industrial chemicals like polychlorina- ted biphenyls (PCBs), have been released to the global environment the last 3-4 decades. Long-range trans- port, via air in particular, takes these persistent and lipophilic compounds to the Arctic (Bowes & Jonkel, 1975; Clausen & Berg, 1976; Muir et al., 1988; Muir et al., 1992; Oehme, 1991; Barrie et al., 1992; Hargrave et al., 1992; Zhu & Norstrom, 1993; Norstrom & Muir, 1994). In arctic areas, particularly in the Western Hemisphere, considerable levels of PCBs and chlor- danes have been found in polar bears (Norstrom et al., 1988; Norstrom et al., 1996; Norheim et al., 1992), the top predator of the Arctic marine food chain. These organochlorines are known to generate adverse effects after chronic administration to laboratory mammals. Both the pesticides and the PCBs may produce a range of effects including neuro, reproductional and immuno toxicities (Hayes, 1982a,b; Safe, 1984; Tilson et al., 1990; Ahlborg et al., 1992; Peterson et al., 1993; Ecobi- chon, 1996). Norheim et al. (1992) found PCB levels in polar bears at Svalbard close to those connected with reproductional disorders in Baltic seals (Helle et al., 1976; Bergman & Olsson, 1985; Olsson et al., 1992).
Polar bears mate in the spring (Wiig et al., 1992). Fertilised eggs do not implant before September- October, about the same time that the pregnant female enters the den (Ramsay & Stifling, 1988). According to Sandell (1990), species with delayed implantation appear especially susceptible to deleterious reproductive effects from organochlorine pollution. Polar bears have delayed implantation and Derocher (1991) suggested that organochlorine-induced effects on reproduction may be one factor in the decline in reproductive perfor- mance of the Western Hudson Bay polar bears. Cubs are born around Christmas and the female emerges from the den usually with two cubs in March-April. She has then fasted for about six months. Polar bear milk has a high fat content (Arnould & Ramsay, 1994) and lactation seems to occur more or less until weaning at about 2 ½ years (Oehme et al., 1995).
The objectives of this study were to provide compre- hensive baseline data on important OC contaminants in polar bears at Svalbard and to assess the possible

160 A. Bernhoft et al.
influence of age, sex and reproductive status on the levels of these lipohilic compounds, particularly the toxicoki- netic influence of elimination and exposure of OCs with milk. In addition, the possible association between OC levels and reproduction success was investigated.
MATERIALS AND METHODS
Field studies Sampling was done during March and April each year during 1990 to 1994 on land-fast ice, except in 1992 when l0 bears were captured on land during August. Most captures took place in the southern and south- eastern parts of Svalbard, with a few bears captured in the northern parts of the archipelago.
Bears were anesthetized by using remote intramus- cular injection of Zoletil® (tiletamine + zolazepam) from a helicopter (Wiig, 1995). Anesthetized bears were measured and their physical condition assessed. A ves- tigal tooth was extracted for age determination by cementum annuli (Grue & Jensen, 1979). Each bear was assigned a unique number which was tattooed inside the upper lip and also applied to a plastic tag in the ear.
Tissue samples were collected from a total of l l5 individual polar bears of both sexes and different ages. Most bears were sampled once, but 8 bears were recaptured and resampled the following year(s): 6 bears resampled once, 1 bear resampled twice, and 1 bear resampled three times. Thus the total number of samplings was 126 (Table 1). Samples of subcutaneous tissue (ca 0.5 g) were collected from the lateral thigh, blood samples (10-50 ml) from a femural vein and milk samples (10030 ml) were collected by manual milking of the mammae. Milk samples were from females with cubs of the year, except two samples from females with yearlings. Blood samples were placed in heparinized tubes, centrifuged and separated into plasma and cells. Samples were stored in polypropylene containers (-20°C) until analysed. Sample sizes for this study var- ied due to logistics and use in other projects (see Norstrom et al., 1996).
Most adult females were equipped with satellite transmitters (Telonics Inc., Mesa, Arizona, USA) as described by Fancy et al. (1988), Harris et al. (1990), and Wiig (1995). The transmitters were programmed to send information during six hour intervals every six
Table I. The total numbers of captures of polar bears during 1990-94 at Svalbard and types and numbers of collected samples for organochlorine analyses. Six bears were sampled two years, one bear was sampled three years and one bear was sampled four years
Bears Subcutaneous Blood Blood Milk captured t i s s u e plasma cells
1990 19 0 5 19 0 1991 14 0 9 14 0 1992 33 31 19 29 6 1993 34 33 30 32 5 1994 26 25 19 22 3
Total 126 89 82 116 14
days for about two years. Information on the location of the transmitter as well as sensor data on internal transmitter temperature and short- and long-term bear activity were recorded.
Reproductive rates can be estimated from satellite data. Female polar bears normally have a three year reproductive cycle and mate during spring (Ramsay & Stirling, 1988). The cubs are born the next winter and follow their mother for about two and a half years, when the mother is supposed to come in oestrus again. Females with 2-year-olds or without young in spring are available for mating. Females were tagged in spring (or summer) and classified as 'available for mating' or not. Only pregnant females are denning. Data recorded from the satellite transmitter make it possible to register if females are denning or not the preceding winter.
Analyses of organochiorines Samples of subcutaneous tissue (ca 0.5 g), milk (ca 2 g), plasma (ca 8 g) and blood cells (ca 8 g) were weighed and added internal standard TCN (samples from 1990 ° 1993) or PCB-112 (samples from 1994). The samples were extracted twice with cyclohexane and acetone using an ultrasonic homogenizer followed by cleanup with sulfuric acid as described by Brevik (1978), slightly modified by Bernhoft and Skaare (1994). For determi- nation of extractable lipids, blood cell extracts were carefully evaporated to dryness. The extracts of sub- cutaneous tissue, plasma and milk were evaporated to ca 1 ml, then diluted with cyclohexane to fixed volumes (10 ml for subcutis and milk extracts, 5 ml for plasma extracts). Then aliquots (4 ml from subcutis and milk extracts, 2 ml from plasma extracts) were evaporated to dryness for determination of extractable lipid. The samples were automatically injected (Fisons autosam- pier AS 800) on a Carlo Erba, HRGC 5300 Mega Series (Carlo Erba Instrumentation, Milano, Italy) gas chro- matograph, equipped with a split/splitless injector (1 /zl, 1:30) and an electron capture Nickel-63 detector. Hydrogen was used as carrier gas on a SPB-5 60 m capillary column with 0.25 mm i.d. and 0.25/zm film thickness (Supelco Inc., Bellefonte, PA, USA). Makeup gas was 5% methane/95% argon. Chromatographic data were calculated using the software Maxima 820 Chromatography Workstation (Millipore Waters, Mil- ford, MA, USA).
The following OCs were determined: chlordane related compounds: heptachlor, oxychlordane, trans-chlordane, cis-chlordane, trans-nonachlor, and cis-nonachlor; DDT-compounds: p,p-DDE, o,p-DDD, p,p-DDD, o,p- DDT, and p,p-DDT; HCB; HCH-isomers: at-, 13-, and 7- HCH; 22 PCB congeners (UIPAC nos, Ballschmiter & Zell, 1980): 28, 52, 66, 74, 99, 101, 110, 105, 118, 128, 138, 141,149, 153, 156, 157, 170, 180, 187, 194, 206 and 209.
The standard curves used for quantification were made by fitting a straight line through two concentra- tions of the standards and the origin. Quantifications were carried out within the linear range of the detector. Percent recovery was calculated for each sample series of 24 samples by adding a known amount of a standard

Organochlorines in polar bears 161
mixture to two samples of clean material . The recoveries of added s tandard a m o u n t were for PCBs and D D T s 80-110%, for chlordanes (including oxychlordane) 80- 100%, for H C B and H C H s 70-95%. Reproduceabil i ty was continuously tested by analysing a control sample (seal blubber) in each sample series. Detection limits for individual OCs were found between 0.5 and 5 ng g - l in subcutaneous lipid and milk lipid, between 2 and 18 ng g - ~ in p lasma lipid and between 10 and 110 ng g - 1 in blood cell lipid.
The l abora to ry has par t ic ipated in several intercali- b ra t ion tests. G o o d analytical quali ty for de terminat ion o f p , p - D D E , I3-HCH, total PCBs and the ma jo r PCB- congeners in h u m a n milk was confirmed by successful
par t ic ipat ion in in te r labora tory tests organized by The Wor ld Hea l th Organisa t ion /Uni ted Na t ions Environ- menta l P r o g r a m m e ( W H O / U N E P ) in 1982 and 1992. Par t ic ipat ion in the four steps o f the In ternat ional Council for Explora t ion o f the Sea/ In ternat ional Oceanographic Commiss ion /Os lo -Pa r i s Commiss ion ( I C E S / I O C / O S P A R C O M ) test on PCBs in mar ine material , placed the l abora to ry in good and accepta- ble groups compared to the other par t ic ipat ing laboratories .
Data analyses The statistical calculat ions o f da ta were made using J M P statistical software (SAS Inst i tute Inc., Cary, NC, USA).
Table 2. Organochlorine concentrations in subcutaneous lipid (ng g- i ) of polar bears at Svalbard grouped according to age and sex. Group a: young 1-2 years, group b: subadults 3-6 years, group c: adult females 7-15 years, group d: adult males 7-15 years and group e:
old males 16-22 years. Medians (means) and ranges are presented
(a) Young (b) Subadults (c) Adult females (d) Adult males (e) Old males
N 8 25 23 20 9 % extractable lipid 54(51) 45(45) 49(50) 40(40) 51(41)
32-69 20-68 1 0-82 1 0-70 15-59 Chlordanes 3210 (3380) a.e 3340 (3440) a.e 2710 (3150) a,e 1030 (1250) 407 (613)
912-6000 858-8310 364-7550 184-2970 199-1870 DDE 519 (567) 274 (305 227 (372) 255 (340) 202 (342)
287-1120 < 2-859 < 2-1820 58-1490 89-1050 HCB 157 (200) 126 (191) 130 (192) 192 (215) 134 (238)
75-496 64-522 49-461 62-480 34-947 HCHs 258 (233) 212 (197) 172 (163) 335 (385) b," 260 (318)
109-365 < 4-473 < 4-358 212-1150 133-644 PCB-99 1480 (1270) 1080 (1190) 1040 (1200) I100 (1300) 671 (706) 2,4,5,2',4'-pentaCB 379-1890 416-2740 454-2410 248-4 100 346-1650 PCB-105 30 (33) 23 (23) 27 (31) 45 (40) 33 (47) 2,3,4,Y,4'-pentaCB < 3-110 < 3-76 < 3-101 < 3-71 < 3-110 PCB-118 57 (49) 30 (39) 76 (76) 90 (98) b 99 (108) 2,4,5,Y,4'-pentaCB < 3-108 < 3-109 < 3-282 < 3-194 < 3-261 PCB-128 46 (44) 15 (30) 19 (43) 55 (63) b 32 (45) 2,3,4,2',Y,4t-hexaCB < 2-115 < 2-125 < 2-202 < 2-176 < 2-127 PCB-138 1420 (1250) 1040 (1140) 1240 (1330) 1460 (1620) 884 (922) 2,3,4,2',4',5'-hexaCB 553-2010 388 2750 464-2750 306-4670 358-2170 PCB-153 5060 (5020) 6300 (7080) 4450 (5930) 9060 (10800) 4710 (4680) 2,4,5,2',4',5'-hexaCB 1590-8330 1860-14 800 1280-15 100 1570-27 900 1600-8040 PCB-156 138 (132) 191 (189) 152 (166) 231 (271) ~ 173 (193) 2,3,4,5,Y,4'-hexaCB < 3-244 62-478 < 3-400 83-892 72-347 PCB-157 133 (133) 214 (198) 111 (t25) 252 (320) ~,c 230 (254) 2,3,4,3',4~,Y-hexaCB < 4-252 59-505 < 4-237 108-1160 81-418 PCB-170 1140 (1110) 1700 (1820) 1110 (1490) 2670 (3830) ".h," 2380 (2560) 2,3,4,5,2~,3t,4'-heptaCB 471-1670 552-4430 552-3550 879-11 800 797-3740 PCB-180 2010 (1980) 3460 (3720) 2480 (3470) 5380 (7530) ~,C 3780 (4220) 2,3,4,5,2',4',5'-heptaCB 943-3440 921-8410 611-8750 914-22 600 1160-6870 PCB-187 19 (14) 15 (21) 27 (29) 26 (31) 18 (25) 2,3,5,6,2',4',5'-heptaCB < 1-28 < 1-97 < 1-82 < 1-76 < 1-91 PCB-194 323 (373) 788 (828) 743 (1030) a 1160 (1900) ~.b 3040 (2360) ~,b,c 2,3,4,5,2',3',4',5'-octaCB 255-568 198-2940 253-4580 379-5740 746-3410 PCB-206 27 (31) 98 (100) a 141 (173) ~ 158 (236) ~,h 228 (289) ~'b 2,3,4,5,6,2',Y,4',5'-nonaCB < 4-69 < 4-343 29 763 49-827 68-499 PCB-209 7 (11) 31 (35) 68 (114) ~.b 75 (121) a,h 135 (187) ~.b 2,3,4,5,6,2',3P,4',5',6'-decaCB < 5-29 < 5-160 < 5 510 < 5-400 32-390 Sum PCBs 11200 (11 400) 14400 (16400) 13000 (15 700) 21700 (28 100) ~.' 15400 (16600)
4810-18 300 5250-36 700 4790-41 500 6960-80 300 5540-27 700
~Significantly higher than corresponding concentrations of group a (p < 0.05). a'bSignificantly higher than corresponding concentrations of groups a and b (p < 0.05). a'b'cSignificantly higher than corresponding concentrations of groups a, b and c (p < 0.05). a'cSignificantly higher than corresponding concentrations of groups a and c (p < 0.05). bSignificantly higher than corresponding concentrations of group b (p < 0.05). b'cSignificantly higher than corresponding concentrations of groups b and c (p < 0.05). a'eSignificantly higher than corresponding concentrations of groups d and e (p < 0.05).

162 A. Bernhoft et al.
Because of lack of normality in the OC dataset, tested by Shapiro-Wilk W test, the data were log transformed to achieve normal distributions. For observations below detection levels, half of the detection levels were used.
In order to explore the correlations between the OCs in a multivariate space we have applied principal com- ponent analysis (PCA). The OC concentrations in sub- cutaneous lipid of all sampled individual polar bears were both normalized and standardized to unit var- iance. The variables are presented as vectors in the sys- tem of the principal components (PCs) that explain most of the variable variance. By plotting the variables in two dimensions the correlating variables will appear as vectors with similar direction and length.
Comparisons of the OC levels between groups were made using Tukey-Kramer statistical method for com- paring all pairs of groups. The OC levels are mainly presented as median concentrations with ranges. The medians are optimal averages of distributions with positive skewness and observations below detection levels (Altman, 1991).
To explore the relations of the concentrations of OCs in different matrices Pearson product-moment correlations were used. Statistical significant level was set at p < 0.05.
To demonstrate how the OCs vary with age for each sex we used smoothing spline (Eubank, 1988). This is a partial least square method, basically descriptive, to visualize in detail the relation between two variables. A coefficient ~, which determines the stiffness of the line was found optimal at ~,=100. In Figs 1-3 where smoothing spline were used, untransformed values are presented to show the real OC levels on y-axes. Statisti- cal differences of OC levels between age and sex groups were tested elsewhere (Table 2).
For comparison of the OC levels in different matrices sampled from the same individuals, median values at
log-scales were presented. The log-scales show the absolute median levels and also illustrate the relative relation between median levels in the matrices for indi- vidual OCs. Median OC levels in mothers and their off- spring are presented similarly.
For individual bears sampled two or more years, only the results from the last samplings are included in Tables 2-7, and in all figures, to reduce the influence of sampling dependency. The results from the resampled individuals are presented separately in Table 5.
All the OC levels are given on the basis of extractable lipids.
RESULTS
Types and levels of OCs in subcutaneous lipid The following OCs were present consistently at levels above quantification levels: chlordanes (sum of oxy-, cis-, and trans-chlordane, and trans-nonachlor), DDE, HCB, HCHs (sum of ~- and [3-HCH) and PCBs (14 individual congeners with 5-10 chloro substituents (Table 2). PCBs and chlordanes predominated. The median concentration of sum PCBs in subcutaneous tissue of 85 individual polar bears was 15.5/zg g-1 lipid. PCB nos. 153 and 180 constituted about 62% of sum PCBs. The corresponding median concentration of chlordanes was 2.3 #g g-l lipid. Oxychlordane consti- tuted about 72% of sum chlordanes. The oxychlordane levels may be conservative estimates due to the possibi- lities of losses in partitioning. The median concentra- tions of DDE, HCB and HCHs were 272, 146 and 240 ng g-I lipid, respectively. ~-HCH constituted about 81% of sum HCHs. The chlordane related compounds cis-nonachlor and heptachlor, the DDT compounds p,p- and o,p-DDT, and p,p- and o,p-DDD, the 7-isomer of
Table 3. Correlation coefficients (r) of organochlorine concentrations (log ng g-i fipid) between subcutaneous tissue (subeutis) and plasma, plasma and milk, subeutis and milk, blood cells and plasma and between blood cells and subeutis in polar bears at Svaibard. For
statistical significant correlations (p < 0.05), the coefficients are given in bold
Subcutis-Plasma Plasma-Milk Subcutis-Milk Bl.cells - Plasma Bl.cells - Subcutis
N 64 9 12 79 76 Chlordanes 0.66 0.47 0.91 0.66 0.77 DDE 0.28 -0.20 0.36 0.42 0.58 HCB 0.88 0.91 0.91 0.66 0.71 HCHs 0.37 0.26 0.91 0.55 0.55 PCB-99 0.73 0.90 0.89 0.57 0.71 PCB-105 0.12 0.51 0.39 c.n.c, c.n.c. PCB- 118 0.17 0.10 0.67 c.n.c, c.n.c. PCB- 128 0.27 0.87 0.65 c.n.c, c.n.c. PCB- 138 0.77 0.71 0.89 0.69 0.66 PCB- 153 0.73 0.38 0.93 0.66 0.78 PCB- 156 0.74 0.70 0.91 c.n.c, c.n.c. PCB-157 0.50 -0.31 0.91 c.n.c, c.n.c. PCB- 170 0.79 0.29 0.80 0.69 0.73 PCB- 180 0.58 0.12 0.86 0.49 0.70 PCB- 187 0.05 0.56 0.66 c.n.c, c.n.c. PCB-194 0.81 -0.01 0.51 0.70 0.77 PCB-206 0.71 -0.01 0.27 c.n.c, c.n.c. PCB-209 0.50 c.n.c, c.n.c, c.n.c, c.n.c. Sum PCBs 0.71 0.35 0.92 0.59 0.73
c.n.c. = correlation not calculated.

Organochlorines in polar bears 163
Table 4. Organochlorine concentrations in blood cells (ng g-I lipid) of adult female polar bears at Svalbard grouped according. to offspring status. The ages (years) are also listed. Medians and
ranges are presented
Adult females
with cubs of with with 2-year-olds the year yearlings or without young
N 11 8 16 Age 9 13 11
7-14 10-14 7-32 % extr. lipid 0.20 0.16 0.15
0.13-0.41 0.04-0.38 0.03-0.37 Chlordanes 2500 1030 2840*
722-4800 811-3950 868-7960 DDE 48 68 102
<40-170 <40-430 <40-410 HCB 227 84 171"
33~25 < 10-165 69-832 HCHs 374 129 310"
< 90--687 < 90-464 11 6-2610 PCBs 6460 4520 8640*
2710-11 200 2220-7630 4090-32 900
*Significantly higher than corresponding concentrations of females with yearlings (p < 0.05).
HCH, and the PCBs no. 28 (tri chlorinated) nos. 52, 66 and 74 (tetra chlorinated) nos. 101 and 110 (penta chlorinated), and nos. 141 and 149 (hexa chlorinated) were all below the detection limits of the analytical method in almost all samples.
Intercorrelations of OCs The correlations between concentrations of the different OCs were explored by use of principal components (PCs) (Fig. 1). Four standardized PCs which explained in total 81% of the variance of the OC concentrations express the OC correlations in subcutaneous tissue of all 85 individual bears. PCs 1 and 2 (Fig. I(A)) which explained 60% of the OC variance indicate that the vectors of PCBs 105, 118, 128 and 187, together with that of sum HCHs, make one intercorrelated group, and the vectors of PCBs 99, 138, 153, 156, 157, 170, 180, 194, 206 and 209 make another group. The vector of HCB was somewhat correlated to both the PCB-99 and the sum chlordane vectors and the DDE vector was ungrouped. In PCs 3 and 4 (Fig. 1 (B)), the vectors of the individual OCs were further separated, and the following
Table 5. Organochlorinc concentrations in blood ceils (ng g-t lipid) of individual polar bears at Svalbard sampled two or more years. Offspring status are given for females: 0-- with cubs, 1 = with yearlings, 2 = with 2-year-olds or without young
Year of Age Offspring Chlordanes DDE HCB HCHs PCBs sampling status
Bear 7768 (female) 1990 10 1 2520 64 165 225 7490 1991 2 4060 <40 222 263 20900 1992 a 0 . . . . . 1993 2 2240 61 154 327 5870
Bear 7771 (female) 1990 13 1 1890 56 86 113 7150 1991 2 4270 42 504 116 13000
Bear 7773 (female) 1990 3 2 1310 150 76 168 3950 1991 2 3650 81 186 264 17900 1993 2 1970 110 141 299 6650
Bear 7802 (female) 1991 7 2 3890 110 234 338 9970 1992 0 2290 <40 100 687 11 200
Bear 7803 (female) 1991 6 2 4870 <40 188 247 9570 1992 0 722 <40 42 <90 2710
Bear 7817 (female) 1992 11 I 920 130 80 <90 2220 1993 2 3210 93 338 416 8070
Bear 7822 (~male) 1992 10 1 811 71 81 103 2590 1993 0 2810 61 160 374 6840
Bear 7781 (male) 1992 21 342 <40 138 279 7730 1993 374 <40 73 402 4230
aln 1992, blood was not sampled from bear 7768.
Table 6. Organochlorine concentrations in blood cells (ng g-1 lipid) of young polar bears (3-6 years) at Svalbard during 1990-1994. Medians and ranges are presented. Correlation coefficients (r) between year and organochiorine concentration are included
1990 1991 1992 1993 1994 r
N 5 8 12 10 4 39 Chlordanes 1940 1410 1720 3430 1830 0.10
1310-8850 369-4870 744-11 600 ! 520-6140 689-3770 DDE 120 69 100 115 114 -0.01
75-710 < 40-120 < 40--480 44-260 < 40-158 HCB 294 187 189 190 254 -0.06
76-1400 78-498 54-542 141-937 114-458 HCHs 522* 256 180 503 183 -0.10
168-2620 < 90-669 < 90-525 143-989 < 90-347 PCBs 10 400 6010 6040 11400 5980 -0.03
3950--40 100 2900-17 900 2890-39 400 4820-20 500 3840-8000
*Significantly higher HCH level than in 1992 (p < 0.05).

164 A. Bernhoft et al.
A
C)DE ~C~18 128 ~ 187
~ ] 1 5 6
Pc2
Eel
B
j L5 187
PC4
PC3
Fig. 1. Correlations between concentrations of different organochlorines in subcutaneous lipid of polar bears at Svalbard in a principal component (PC)system. The numbers 99, 105, 118, 128, 138, 153, 156, 157, 170, 180, 187, 194, 206 and 209 are IUPAC numbers of PCB congeners. Chls and HCHs are sum of chlordane-related compounds and sum of ct- and ~-HCHs, respectively. DDE is p,p-DDE and HCB is hexachlorobenzene. PCs 1 and 2 (Fig. I(A)) explain 60% of the OC variance. PCs 3 and 4 (Fig. I(B))
explain the next 21% of the variance. Vectors close in direction and length indicate highly correlated compounds.
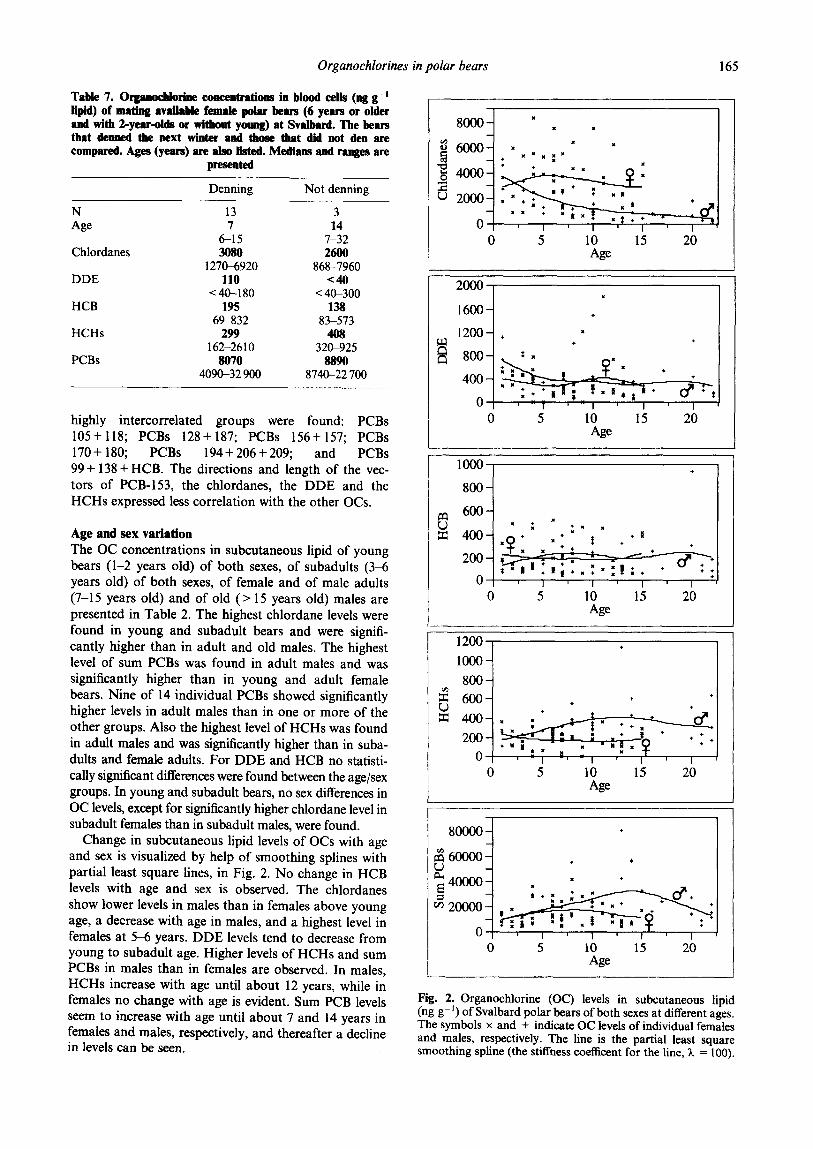
Organochlorines in polar bears 165
Table 7. Orgaaochlorine concentrations in blood cells (ng g-t lipid) of mating available female polar bears (6 years or older and with 2-year-olds or without yowg) at Svalbard. The bears that denned the next winter and those that did not den are compared. Ages (years) are also listed. Medians and ranges are
presented
Denning Not denning
N 13 3 Age 7 14
6-15 7-32 Chlordanes 3080 2600
1270-6920 868-7960 DDE llO < 40
< 40-180 < 40-300 HCB 195 138
69-832 83-573 HCHs 299 408
162-2610 320-925 PCBs 8070 8890
4090-32 900 8740--22 700
highly intercorrelated groups were found: PCBs 105+118; PCBs 128+187; PCBs 156+157; PCBs 170 + 180; PCBs 194 + 206 + 209; and PCBs 99+ 138+HCB. The directions and length of the vec- tors of PCB-153, the chlordanes, the DDE and the HCHs expressed less correlation with the other OCs.
Age and sex variation The OC concentrations in subcutaneous lipid of young bears (1-2 years old) of both sexes, of subadults (3-6 years old) of both sexes, of female and of male adults (7-15 years old) and of old (> 15 years old) males are presented in Table 2. The highest chlordane levels were found in young and subadult bears and were signifi- cantly higher than in adult and old males. The highest level of sum PCBs was found in adult males and was significantly higher than in young and adult female bears. Nine of 14 individual PCBs showed significantly higher levels in adult males than in one or more of the other groups. Also the highest level of HCHs was found in adult males and was significantly higher than in suba- dults and female adults. For DDE and HCB no statisti- cally significant differences were found between the age/sex groups. In young and subadult bears, no sex differences in OC levels, except for significantly higher chlordane level in subadult females than in subadult males, were found.
Change in subcutaneous lipid levels of OCs with age and sex is visualized by help of smoothing splines with partial least square lines, in Fig. 2. No change in HCB levels with age and sex is observed. The chlordanes show lower levels in males than in females above young age, a decrease with age in males, and a highest level in females at 5-6 years. DDE levels tend to decrease from young to subadult age. Higher levels of HCHs and sum PCBs in males than in females are observed. In males, HCHs increase with age until about 12 years, while in females no change with age is evident. Sum PCB levels seem to increase with age until about 7 and 14 years in females and males, respectively, and thereafter a decline in levels can be seen.
8000-
6OOO- § _
~ 4 o o o -
d 200o- O -
x
x •
N x
• X x l X ÷ • 4 ÷ XX X
• ÷ * x l
' i ' I ' i ' I
5 10 15 20 A ~
2000
1600 -
1200 -
8 0 0 -
4 0 0 -
0 0
it +
÷ +
5 10 15 20 Age
1000
800-
600-
2 o o ~
0 0
x
t ~ ÷ ÷ , ÷ II x * ÷ ÷ + +
l[l/l ; - + • II : m C~ v" + +
, . m , . m w . . . . i_. . • + : ' I ' I ' I ' I
5 10 15 20 Age
=
1200
1000
800
600
400
200
0 0
÷ ÷ ÷
5 10 15 20 A ~
80000
60000
E 40000
o~ 20000
0
÷ #
• +
' I ' I ' I ' I
5 10 15 20 Age
Fig. 2. Organochlorine (OC) levels in subcutaneous lipid (ng g-l) of Svalbard polar bears of both sexes at different ages. The symbols x and + indicate OC levels of individual females and males, respectively. The line is the partial least square smoothing spline (the stiffness coeflicent for the line, ~. = 100).

166 A. Bernhoft et al.
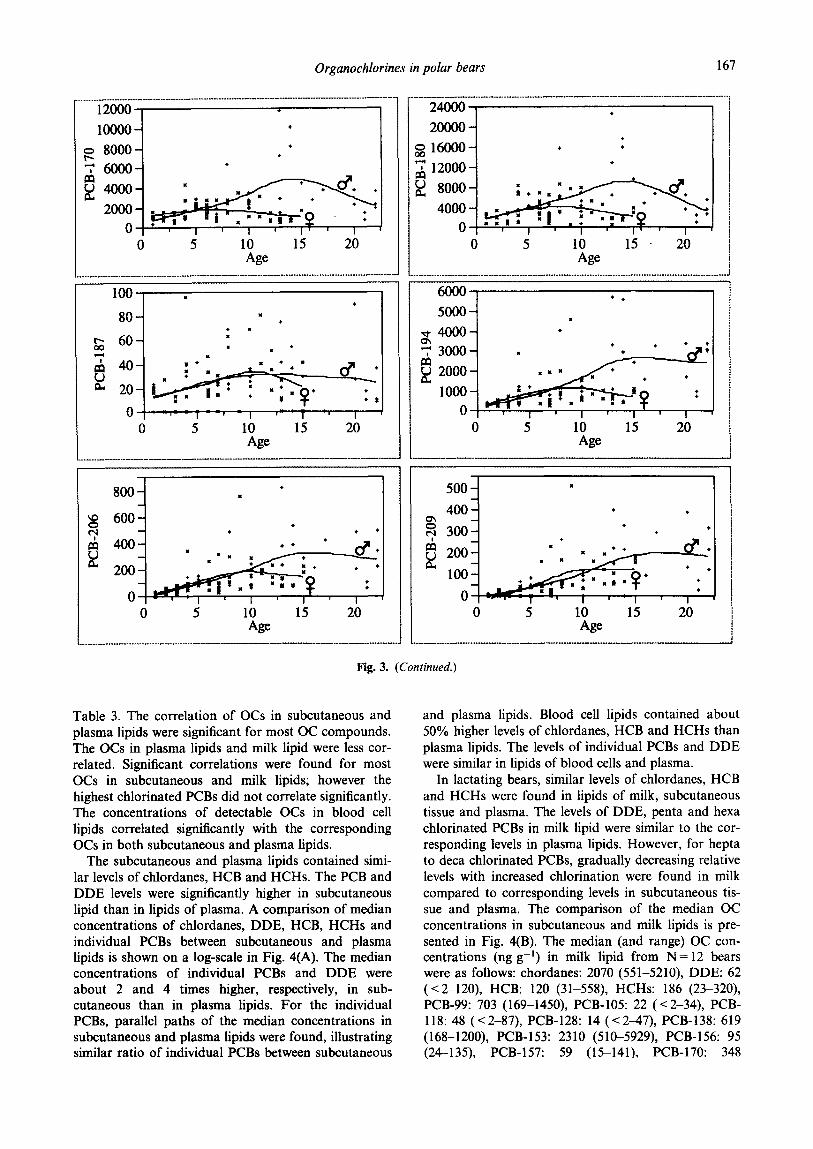
The levels of individual PCBs are similarly visualized in Fig. 3. All congeners except nos. 99 and 138 tend to increase with age in younger males. The increase is more prominent with increased chlorination. The levels of all PCB congeners, except PCBs 105 and 118, tend to reach a plateau or decline in old males. In females, the indivi- dual PCBs tend to follow the corresponding levels in males until adulthood. Then from 7-11 years of age a deviation from the corresponding male level, especially
for the hexa and hepta chlorinated congeners, nos. 153, 156, 157, 170 and 180, can be seen.
T i s s u e d i s tr ibut ion and trans fer to o f f spr ing
Median per cent extractable lipids of blood cells, plasma, milk and subcutaneous tissue were 0.20, 1.1, 25 and 46, respectively.
Correlation coefficients found between OC con- centrations in different types of matrices are shown in
4200 3600- 3000- 2400- 1800- 1200- 600-
0 0
x x
I ' I ' I ' I
5 10 15 20 Age
120
100
8 0 - P:, '7 6 0 -
4 0 -
20-
0 0
u 4. +.
II
II
I t .I. • .I.
x l x I + • + + '+
¥ + .
" " ' ' i ' " "~ , ' + i +' l 5 10 15 20
Age
300
240 - O O
i 180- 120 -
60-
0 0
+
x u • 4. I I 4 *
= = ' - - : i = = ' = I ' -- : i : ' I
5 10 15 20 Age
200 -
160 o o
120
80
40
0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . L . . . . . . . . . . . . . . .
4800
4000 -
3200 + II * u
2400800 " ~ " ~ 1600 , " O .
0 ' I ' I ' I ' I
0 5 10 15 20 Age
800-
600 - ~O z
x '1"
u ~ + * + ÷ ÷
~'~'ft: 1: , r, ; " ' : F : + ' I , n I ' I ' I
5 10 15 20 Age
400
200
M
M m +
+
0 5 10 15 20 Age
28000 24000- •
+
20000- + 16000- : +
J dl 4. M
8000 - 4 0 0 0 - " i ° +
0 ' I ' I ' I ' I
0 5 10 15 20 Age
1200
1000 -
800-
600-
400-
0
$
5 10 15 20 Age
Fig. 3. Levels of individual PCBs in subcutaneous lipid (ng g-Z) of Svaibard polar bears of both sexes at different ages. The sym- bols x and + indicate PCB congener levels of individual females and males, respectively. The line is the partial least square
smoothing spline (the stiffness coefficent for the line, ~, = 100). (Continued.)
Organochlorines in polar bears 167
12000 .
10000-
o 8000- t ~
+
4000 " ~ t ~ i i ~ I 200~ . . . . . . ,.,--.r..i.i..T_ ~ ;
i l I ~ u I I I I
0 5 10 15 20 Age
100
80
60
"7 4 0 -
2 0 -
0
÷
+
÷ •
•
~ , . ' Y ~ m n = * ~; - ' ~ 4, s
' = ,==i ==' = i '" : i =' i
5 10 15 20 Age
24000
20000- +
o 16000- • •
12000-
4000- . ~ 1 ~ 0 I I I
0 5 10 15 20 Age
6000
5000 -
,~ 4000-
3000-
2000
0
+
• +
x x • x ÷ ~ +
' I ' I ' I ' I
5 10 15 20 Age
800-
600- t " , l
@
+ + ÷ +
t~ 400 . ° " , ,~ . • x x •
200 . " " "" -'" ?: 0
0 5 10 15 20 Age
500 -
400 -
¢q 300 -
200 -
100-
0 0
4
X ÷ X I ~
x• x ÷
+
- - - ' - - T - ' I ' : I ' I
5 10 15 20 Age
Fig. 3. (Continued.)
Table 3. The correlation of OCs in subcutaneous and plasma lipids were significant for most OC compounds. The OCs in plasma lipids and milk lipid were less cor- related. Significant correlations were found for most OCs in subcutaneous and milk lipids; however the highest chlorinated PCBs did not correlate significantly. The concentrations of detectable OCs in blood cell lipids correlated significantly with the corresponding OCs in both subcutaneous and plasma lipids.
The subcutaneous and plasma lipids contained simi- lar levels of chlordanes, HCB and HCHs. The PCB and DDE levels were significantly higher in subcutaneous lipid than in lipids of plasma. A comparison of median concentrations of chlordanes, DDE, HCB, HCHs and individual PCBs between subcutaneous and plasma lipids is shown on a log-scale in Fig. 4(A). The median concentrations of individual PCBs and DDE were about 2 and 4 times higher, respectively, in sub- cutaneous than in plasma lipids. For the individual PCBs, parallel paths of the median concentrations in subcutaneous and plasma lipids were found, illustrating similar ratio of individual PCBs between subcutaneous
and plasma lipids. Blood cell lipids contained about 50% higher levels of chlordanes, HCB and HCHs than plasma lipids. The levels of individual PCBs and DDE were similar in lipids of blood cells and plasma.
In lactating bears, similar levels of chlordanes, HCB and HCHs were found in lipids of milk, subcutaneous tissue and plasma. The levels of DDE, penta and hexa chlorinated PCBs in milk lipid were similar to the cor- responding levels in plasma lipids. However, for hepta to deca chlorinated PCBs, gradually decreasing relative levels with increased chlorination were found in milk compared to corresponding levels in subcutaneous tis- sue and plasma. The comparison of the median OC concentrations in subcutaneous and milk lipids is pre- sented in Fig. 4(B). The median (and range) OC con- centrations (ng g-l) in milk lipid from N = 12 bears were as follows: chordanes: 2070 (551-5210), DDE: 62 (<2-120), HCB: 120 (31-558), HCHs: 186 (23-320), PCB-99:703 (169-1450), PCB-105:22 (<2-34), PCB- 118:48 (<2-87), PCB-128:14 (<2--47), PCB-138:619 (168-1200), PCB-153:2310 (510-5929), PCB-156:95 (24-135), PCB-157: 59 (15-141), PCB-170: 348

168 A. Bernhoft et al.
(97-824), PCB-180:710 (200-1450), PCB-187:5 (< 1- 9), PCB-194:107 (51-164), PCB-206:13 (< 2-22), PCB- 209: < 3 (< 3), Sum PCBs: 5140 (1270-11400).
The OC levels in subcutaneous lipid of yearlings were generally higher than in their mothers. However, the relative levels of the hepta to deca chlorinated biphe- nyls, nos. 187, 194, 206 and 209, in yearlings compared
to their mothers decreased gradually with increased chlorination and were all below maternal levels. PCBs 105 and 118 and DDE were found at similar levels in mothers and yearlings (Fig. 4(C)).
For comparing OC levels between groups of bears pre- sented in Tables 4, 5 and 6, blood cell data were used since these were available from the largest number of bears.
e-
O
r~
flJ
I0000 , N = 6 4
1000
0
100 X
I0-
I
10000". N = '12 O
1000-
O
100"
10"
1
10000- N = 4
i000- 8
I00-
10
I
rj
Ii
0
X
0
X
0
X
0
X 0
0 X 0 X X
0 0
11 x
0
@ 0
0 0 0
0
0
o o Pt o 4, o -~ o
+
0 ¢ 0 @ 0
0 •
• 0 0 0
0
0 0
0
O O
e o o •
A
o Subcutis
0 x Plasma X
0
Z ,Z, 2 2 2 2 2 2 2 2 2 2
B
o Subcutis
Milk
C
o Mother
• Yearling
Fig. 4. The patterns of chlordanes, DDE, HCB, HCHs and individual PCBs in (A) subcutaneous and plasma lipids, (B) subcuta- neous and milk lipids and (C) subcutaneous lipid of mothers and yearlings. The median levels are given on log-scales.

Organochlorines in polar bears 169
The OC levels in adult females (7 years and above) grouped by offspring status are shown in Table 4. Females with cubs of the year, yearlings and with 2-year-olds or no young, are compared. The mothers that had been lactating for about 1 1/4 years contained significantly less chlordanes, HCB, HCHs and sum PCBs than females that had been lactating for about 2 1/4 years or having no young, and showed a ten- dency of lower levels than mothers with cubs of the year.
OC levels in individual bears sampled several years One male and seven female polar bears were sampled two or more years. High individual interyear variation in OC levels was found (Table 5).
Time trends Time trends in OC levels from 1990-1994 were analysed in subadult polar bears (3-6 years) only, to avoid influ- ences of age, sex and reproductive status on the OC levels (Table 6). Interyear fluctuations in levels were found. However, no significant correlations between OC levels and time were found, and the only signifi- cant difference in OC levels between years was found for HCHs as the level in 1990 was higher than in 1992.
Effect on reproduction success The OC levels in sexually mature females were com- pared to their denning status the next winter. As only pregnant females are denning the whole winter, denning status is regarded as a measure of their reproduction success. The OC levels in the denning bears were not significantly different from those not denning (Table 7).
DISCUSSION
Types and levels of OCs The present study confirms high contamination level of particularly the PCBs in polar bears at Svalbard. PCBs are regarded as one of the most toxic groups of pollu- tants of the Arctic environment (Jensen, 1991; Nor- strom & Muir, 1994; Tanabe et al., 1994). The sum PCB level found in subcutaneous lipid from adult male polar bears at Svalbard in the present study is similar to the corresponding level (corrected to adult male level) found in polar bears sampled in the same way at Sval- bard in 1990 (Norstrom et al., 1996). Sum PCB level in adult males at Svalbard is ca 6 and 3 times higher than the averages of corresponding levels in polar bears (corrected to adult male levels) in Alaska and Canada, respectively, sampled during 1989-1993 (Norstrom et al., 1996). Comparison of OC levels between studies should always be done with caution since the groups of animals may not be comparable with respect to sex, age, nutritional and reproductive status. Sampling year and analytical methods may also differ. However, our results
and those of Norstrom et al. (1996) should be compar- able with respect to biological and temporal parameters. Furthermore, some differences in the number of individual PCBs included in sum PCBs between the studies should not be of significance since the ratios of the sum of the two major PCBs, nos. 153 and 180, to the sum PCBs were similar, in average 62% (present study) and Norstrom et al., 1996). Particularly high PCB levels in Svalbard bears were also reported by Norheim et al. (1992) in bears found dead or shot during 1978-1989.
The sum chlordane level in the present study cannot be compared to sum chlordanes reported by Norstrom et al. (1996) in polar bears in Alaska and Canada as somewhat different chlordane related compounds were summed in the two studies. However, oxychlordane is the quantitatively dominating chlordane related com- pound, and quite similar oxychlordane levels seem to be present in adult male bears at Svalbard (present study) and in Alaska and Canada (Norstrom et al., 1996). Similar DDE level was found in Svalbard bears in the present study compared to corresponding Canadian samples, but somewhat higher than in corresponding Alaskan samples (Norstrom et al., 1996). The levels of HCB and HCHs in Svalbard bears seem to be at the lower end of levels reported in Canadian polar bears during 1982-84 (Norstrom et al., 1988). A lower con- tribution of the most persistent HCH isomer, [3-HCH (Hayes, 1982a), to sum HCHs in adipose tissue of the Canadian polar bears (29% of sum HCHs, Norstrom et al., 1988) than in the present study indicates that the Canadian bears were closer to the HCH source, geo- graphically or temporally.
The present and other studies have demonstrated that top predators of the marine food chain at Svalbard are particularly contaminated with PCBs. Thus, high PCB concentrations in Arctic fox (Alopex lagopus) (Wang- Andersen et al., 1993) and Glaucous gull (Larus hyper- boreus) (Gabrielsen et al., 1995; Mehlum & Daelemans, 1995) in the Svalbard area have been found. In ringed seals (Phoca hispida) higher levels of PCBs are found in the Svalbard area than in the Canadian Arctic (Muir et al., 1992; Daelemans et al., 1993). European, East- American and possibly also Eurasian sources, with compounds transported by air and ocean currents, seem to be the major contributors to the OC contamination in the Svalbard area (Oehme, 1991). Higher PCB con- centrations have been found in North Atlantic air and surface seawater, which distribute to the Svalbard area, than in air and surface seawater westwards in the Bering Sea, Gulf of Alaska, and Northern North and North Pacific (Iwata et al., 1993). The North Atlantic levels of chlordanes, DDTs and HCHs, however, were not higher than westwards. The study of Iwata et al. (1993) shows the influence of long range transport of OCs and is consistent with the finding of the particu- larly high PCB levels in predators at Svalbard. How- ever, local sources of PCB contamination from the coal mining sites at Svalbard may also play a role (Mehlum & Daelemans, 1995).

170 A. Bernhoft et al.
Biomagnitication aspects The polar bears are at the top of the marine food-chain, preying primarily on ringed seals and to a lesser extent on bearded seals (Erignathus barbatus) (Stifling & Archibald, 1977). Often the bears eat only the subcuta- neous fat of the seals they catch. By comparing the pre- sent polar bear OC data (subcutaneous lipid) with available corresponding ringed seal data (blubber) from Svalbard (Oehme et al., 1988; Daelemans et al., 1993), biomagnification of oxychlordane, HCB, sum HCHs and sum PCBs from ringed seal to polar bear is evident with a biomagnification factor of about 10 for sum PCBs. That considerably lower DDE level was found in polar bear as compared to ringed seal is reflecting the capacity of polar bears to metabolise the most persistent DDT compound (Norstrom et al., 1988, 1996; Norheim et al., 1992; Norstrom & Muir, 1994). Furthermore, both HCH and PCB patterns differ between the spe- cies. 13-HCH dominates in polar bear while Qt-HCH was the dominating isomer in ringed seal. The insecticide lin- dane (~-HCH) was detected in ringed seal but not in polar bear. These findings are in accordance with 13- HCH being the most persistent of the HCHs and thus being biomagnified through the food chain (Hayes, 1982a). Metabolism of lower chlorinated PCBs and biomagnification of more highly chlorinated congeners explain the difference in PCB pattern between the spe- cies (Daelemans et al., 1993). Thus, the polar bear seems capable of metabolising a range of congeners that other marine mammals have less ability to eliminate (Nor- strom et al., 1988; Boon et al., 1994; Norstrom & Muir, 1994; Tanabe et al., 1994). Tri and tetra-chlorinated PCBs, in addition to penta and hexa chlorinated PCBs with meta-para adjacent hydrogens, are not found in the polar bears. In other marine mammals, such PCBs are only metabolised to a very limited extent and therefore usually present (Boon et al., 1994; Norstrom & Muir, 1994; Kleivane et al., 1995; Espeland et al., 1997). Fur- thermore, penta chlorinated 105 and 118, hexa chlori- nated 128 and hepta chlorinated 187 are all present at lower relative levels in polar bears compared to ringed seals at Svalbard (Daelemans et al., 1993). The mono- ortho chlorinated 105 and 118 have adjacent ortho-meta hydrogens that seem to be easily available for metabo- lism in polar bear, as reported by Norstrom and Muir (1994) and Letcher et al. (1996). The finding of similar elimination capacity for di-ortho chlorinated 128 may be due to ortho-meta hydrogens available for metabolism on both biphenyl rings. PCB-187 has no adjacent hydrogens but lacks a para chlorine which may make metabolism possible in the polar bear (Norstrom & Muir, 1994).
OC intereorrelations and age/sex variation The study of OC patterns in depot lipid by the use of PC vectors elucidates compounds with similar fates in the polar bear. Thus, the OC vectors that intercorre- late reflect compounds with close toxicokinetics. The highly correlated OCs were found to have common variation with age and sex, as discussed below.
However, the level and pattern of the compounds in polar bear food may also influence the OC intercorrela- tions in the predator.
For the chlordanes, the decrease with age in male bears and the lower level in male than in female adults could be due to sex differences in metabolising capacity for chlordanes in polar bears. Similar tendencies were also found for chlordanes in the polar bear study of Norstrom et al. (1996). The results are in contrast to the common findings of higher chlordane levels in males than in females of other marine mammals (Muir et al., 1992; Norstrom & Muir, 1994).
The tendency of a decreasing DDE level from young to subadults may suggest a lower metabolising capacity for DDE in young than older bears. Similar DDE levels in subadults and adults of both sexes are found both in the present study and the study of Norstrom et al. (1996) and indicate that intake equilibrates with elimi- nation. The findings in polar bears are in contrast to the age dependent accumulation of DDTs commonly found in male cetaceans and pinnipeds (Martineau et al., 1987; Aguilar & Borrell, 1988; Tanabe et al., 1994; Kleivane et al., 1995).
The findings that age and sex did not seem to influ- ence the levels of HCB and PCBs 99 and 138 in polar bear depot lipid, and that a high correlation was found between these compounds, indicate a common tox- icokinetic pattern. Lack of age and sex influence on HCB levels has also been reported for other marine mammals (Muir et al., 1988; Kleivane et al., 1995). In polar bears, the intake of penta chlorinated 99 and hexa chlorinated 138, both with diortho chlorines and adja- cent ortho-meta hydrogens, seems to equal the elimina- tion, indicating metabolism of these PCB structures in this species in contrast to other marine mammals (Boon et al., 1994; Norstrom & Muir, 1994).
The variation with age and sex for the PCBs (exclud- ing 99 and 138) and the HCHs with accumulation with age, particularly in males, is opposite that of the chlor- danes. Age accumulation of HCHs in male bears is somewhat unexpected as the importance of sum HCHs decreases significantly in the Arctic food web above zooplankton, and sum HCH levels do not tend to increase with age in male cetaceans (Kleivane et al., 1995). However, sum HCH accumulation with age may reflect the persistence of the [3-isomer in the polar bear.
In spite of relatively lower levels of PCBs 105, 118, 128 and 187 in polar bears compared to their food (Daelemans et al., 1993), these metabolisable PCBs all tend to increase with age. The increase is difficult to explain and is most pronounced in males. Hexa chlorinated 156 and 157, hepta chlorinated 170 and 180, octa chlorinated 194, nona chlorinated 206 and deca chlorinated 209, accumulate significantly with age in males. A tendency of age accumulation is found for PCB-153. Both the age accumulation in bears and the biomagnification from ringed seal (Daelemans et al., 1993) tend to increase with numbers of chlorines. Only the three highest chlorinated PCBs accumulate signifi- cantly with age in female bears as well. The results are

Organochlorines in polar bears 171
somewhat unexpected for PCB-153 as this congener is often referred in the literature as the most persistent PCB compound in the biota with its unmetabolisable 2,4,5,2',4',5'-chlorine substitution pattern. In polar bears, however, this PCB congener is still the quantita- tively dominating one. Mono-ortho chlorinated 156 and 157 and diortho chlorinated 170, all have ortho-meta adjacent hydrogens which could make metabolism pos- sible; however, the high number of chlorines seems to prevent metabolism.
The tendency of the levels of some PCBs to decrease after a plateau has been reached in old males is difficult to explain.
A comparison of PCB variation with age between the present and the polar bear study of Norstrom et al. (1996) is not possible due to different data presentation in the two studies.
The deviation from the male PCB levels in females from 7-11 years of age coincides with the start of breeding. Female polar bears at Svalbard usually have their first offspring when 6-7 years old (Wiig, 1996). The significant sex-specific PCB levels reflect the fact that in mature female mammals, milk is an important excretion pathway for lipophilic compounds resistant to meta- bolic degradation (Matthews & Dedrick, 1984; Takagi et al., 1986; Addison & Brodie, 1987; Norstrom & Muir, 1994; Espeland et al., 1997). High fat content of polar bear milk and a lactation period up to 2.5 years (Oehme et al., 1995) make OC excretion into milk important for this species. Significantly lower sum PCB levels in adult polar bear females than in adult males are found both in the present study and the study of Norstrom et al. (1996).
Of the individual PCBs, hexa and hepta chlorinated congeners with high age accumulation in males show the greatest differences in levels between male and female adults (PCBs 157, 170, 180). For the more metabolisable PCBs, the metabolic capacity may make excretion into milk relatively less important, and thus the sex differences in the levels less distinct. Lack of significant difference between adult females and males for the highest chlorinated and most persistent PCBs may be due to low rate of transfer into milk of these congeners.
The lower HCH level in female than in male adults corresponds to the finding of relatively high HCH level in milk. In addition, the high level of the stable ~-isomer in polar bear may make the ratio of milk HCH excre- tion to the whole HCH elimination more important in polar bears than in cetaceans and pinnipeds, where similar male and female HCH levels seem to be the common finding in most studies (Muir et al., 1992; Norstrom & Muir, 1994). High excretion into milk is also found for chlordanes and HCB in polar bears. However, metabolism pathways seem more important for their elimination than milk excretion, as adult female levels are not lower than those of adult male bears. Both low level in milk and relatively low persis- tence of DDE in polar bears may explain the lack of sex difference in DDE level.
Tissue distribution and transfer to offspring The significant correlations of OCs between sub- cutaneous depot lipid and blood plasma lipid indicate that the compounds are readily exchanged between the two compartments. Lack of statistically significant cor- relation of levels of PCBs 105, 118 and 187 between subcutaneous and plasma lipids may be due to levels below detection limit in several bears. The dynamic dis- tribution of the OCs between depot lipid and plasma lipid may be explained by simple equilibration-parti- tioning based on lipid solubility of the OCs and the composition of compartment lipids, as described by Matthews and Dedrick (1984). Subcutaneous lipid is composed mainly of non-polar triacylglycerols, while the plasma lipids consist mainly of lipoproteins with the non-polar lipids cholesterol esters and triacylglycerols, and the more polar phospholipids and apoproteins. In total, the plasma is a somewhat polar compartment which may explain the lower level of the most lipophilic OCs in plasma lipid than in subcutaneous lipid. Thus, our finding that the relative levels in plasma lipid com- pared to subcutaneous lipid are lower for DDE and PCBs than for chlordanes, HCB and HCHs which show similar levels in the lipids of plasma and subcutis, are consistent with the lipophilicity of the compounds esti- mated as n-octanol/water partition coefficients (Geyer et al., 1987). Addison and Brodie (1987) determined PCBs and DDTs in grey seal (Halichoerus grypus) and found levels in blood serum lipid below 50% of blubber lipid levels. The lipophilicity of PCBs generally increases with increasing number of chlorines (Erickson, 1986). How- ever, in the present study, similar relative levels were found for all PCB congeners in blood plasma and sub- cutaneous tissue. In grey seal, the PCB patterns in blubber and blood serum were different with decreasing levels of high chlorinated PCBs in serum (Addison & Brodie, 1987). Similar PCB patterns in subcutaneous tissue and plasma in polar bear may be due to a parti- cularly high level of cholesterol esters in polar bear plasma compared to other species (Long, 1961) indicat- ing a less polar plasma compartment of polar bears compared to other species.
Significant correlations of all OCs in blood cell and plasma lipids and similar levels of individual PCBs and DDE suggest rapid equilibrium of the compounds between blood cells and plasma. A ready exchange of halogenated biphenyls between plasma and the cells of rat blood has been reported by Matthews et al. (1984). The significantly higher levels of the least lipophilic OCs, chlordanes, HCB and HCHs in blood cell than in plasma lipids of the polar bear indicate higher affinity to the blood cell lipids. This is most probably caused by differences in polarities of lipids between plasma and cellular components. Partition of individual OCs between blood cells and plasma varies by species (Yang et al., 1975; Matthews et al., 1984; Mes et al., 1992) and seems most likely to be due to great species differences in patterns of lipids of both blood cells and plasma (Long, 1961). The significant correlation of OCs in lipids of subcutaneous tissue, blood cells and plasma of

172 A. Bernhoft et al.
the bears make blood cell and plasma lipids usable as well as depot lipid of subcutaneous tissue for comparing OC levels between groups of polar bears. However, comparison of OCs between species should be based on levels in depot lipid as OC patterns in blood may devi- ate from that of depot lipid in various species due to the species differences in blood lipid patterns.
The OC patterns in milk lipid of the polar bear have similarities to that of plasma lipids with similar levels of HCB, HCHs and chlordanes and lower levels of PCBs and DDE in milk and plasma lipids compared to sub- cutaneous lipid. However, as higher chlorinated PCBs show relatively lower levels in milk, the most lipophilic PCBs seem less transferable to milk lipids than to cir- culatory lipids. This may be due to the complex bio- synthesis of milk lipid. As in subcutaneous depot lipid the main milk lipid is triacylglycerol. However, milk triacylglycerols are based on both fatty acids produced in the mammary gland from carbohydrate sources in addition to fatty acids from circulating triacylglycerol (Collier, 1985). Thus, the incomplete transfer of lipid from the circulatory system to milk may reduce the effi- ciency of the OC transfer, particularly of the most lipo- philic compounds. This is supported by the low correlations between plasma and milk lipids for most OCs, and particularly for the higher chlorinated PCBs and DDE.
The OC pattern in subcutaneous lipid of yearlings reflects the corresponding pattern found in maternal milk. Lower transfer of higher chlorinated PCBs from mother to offspring are also reported in harp seal (Phoca groenlandica) and hooded seal (Cystophora cris- tata) (Espeland et al., 1997). Except for the highest chlorinated PCBs, most of the OCs are present at higher levels in yearling bears than in their mothers, and demonstrate the importance of lactation for transfering OCs from one generation to the next. Yearling bears consume substantial amounts of milk; however, the intake is much lower and the milk has lower percentage fat than milk available for cubs of the year (Arnould & Ramsay, 1994; Derocher et al., 1993). Seal blubber that gradually becomes more important as food for the young polar bears contains much lower OC levels (Daelemans et al., 1993) than the maternal milk. Thus Polischuk et al. (1995) found higher levels of the lipo- philic persistent compounds in polar bear cubs of the year than in yearlings. Generally lower metabolic capa- city for xenobiotica in young animals than in adults (Timbrell, 1991) may also influence the OC level and pattern in the young bears. The levels of PCBs nos. 105 and 118 in yearlings are lower than expected relative to the levels in milk, which may suggest a faster metabo- lism of these congeners than other PCBs in young bears.
The findings of lower OC levels in females with nur- sing yearlings than females with 2-year-olds or without young and females with cubs of the year are indications of an especially high OC excretion during the first year of lactation. Females with yearlings have been lactating for about 1 1/4 years. According to Arnould and Ram- say (1994) the significant part of the total milk lipid
mobilised during the whole lactation period seems to be excreted within the first year. This suggests similar excretion pattern of the OC body burden. For females with 2-year-old young the milk excretion is low and has probably an insignificant influence on OC burden. Thus, the results indicate that OC levels in females lac- tating for about 2 1/4 years may have started to increase. Females of 7 years or more without young are grouped together with females with 2-year-old young, as those without young may recently have been naturally separated from their offspring (Oehme et al., 1995). However, the adult females without young may also include females out of reproduction and any females that may have lost their offspring or fetus. Thus, the interpretation of OC levels in the group of females with 2-year-olds or without young should be cautious. The tendency of higher OC levels in females with cubs of the year than in those with yearlings may be due to recent start of lactation and hence short time of OC excretion. As females with cubs of the year at sampling time had just left their dens after months of fasting, any decrease of adipose reserves may have contributed to concentrate the OC residues. Polischuk et al. (1995) found increased levels of persistent OCs in polar bears during fasting period due to decreased adipose reserves. The lack of significant group differences for DDE may be explained by the high metabolising capacity in polar bear for this compound.
OC levels in individual bears followed from year to year The survey of OC levels in bears followed from year to year supports the impression of particularly low OC levels in females with yearlings as discussed above. In addition, considerable variation in OC levels may be found from one year to the next also in the subadult female and the old male studied, which could be due to differences in nutritional status according to Polischuk et al. (1995).
Time trends The results demonstrate a great variability of OC levels in polar bears without any significant trend in levels with time. Many factors like diet, age, sex, metabolising capacity, nutritive and reproductive status are known to affect the variability in OC levels (Bignert et al., 1994). In this presentation, the influences of age, sex and reproductive status are standardized as only subadult bears are compared. The 5 year period from 1990-94 is, however, too short to reveal temporal trends. So far, no well-defined temporal trends in OC levels have been found in polar bear studies (Muir et al., 1992; Norstrom & Muir, 1994; Norstrom et al., 1996).
Effects on reproduction and offspring We found no difference in OC content between avail- able females that became pregnant and those that did not become pregnant. However, the sample sizes for this comparison are very small. The reproductive success of adult female polar bears at Svalbard is about 0.75 which

Organochlorines in polar bears 173
is similar to values found in other polar bear popula- tions (Wiig, 1996). However, Wiig (1996) found a very high mortality of young polar bears at Svalbard. High intake of OC pollutants occurs at a crucial period of growth and development of the young polar bears. This could adversely influence the early development of cubs and lead to a higher mortality. Negative correlations of plasma retinol and thyroid hormone with sum PCB levels in polar bears at Svalbard are found by Skaare et al. (1994). However, the biological significance of the findings is not elucidated.
In spite of high sum PCB levels, concentrations of the non-ortho coplanar congeners which are regarded as the most toxic PCBs (Safe, 1990; Ahlborg et al., 1994) seem to be present at low concentrations in polar bears (Oehme et al., 1995; Letcher et al., 1996). Planar PCBs may elicit body weight loss and reproductive, immuno- and hepatotoxic effects comparable to those reported for 2,3,7,8-tetrachlorodibenzo-p-dioxin (2,3,7,8-TCDD) through a proposed common mechanism of action (A_h-receptor) (Safe, 1990; Ahlborg et al., 1994). Further- more, the concentrations of polychlorinated dibenzo- p-dioxins (PCDDs) and dibenzofurans (PCDFs) expressed as 2,3,7,8-TCDD equivalents (TEQs) in polar bears do not tend to bioaccumulate in polar bears (Nor- strom et al., 1990; Oehme et al., 1995; Letcher et al., 1996). The mono-ortho chlorinated analogues of the toxic coplanar congeners, PCBs 105, 118, 156 and 157, determined in the present polar bear samples, have similar but weaker biological effects than the non-ortho chlorinated congeners. The two former seem easily metabolisable in polar bears and are present at low concentrations. PCBs 156 and 157, however, are bio- magnified from ringed seal (Daelemans et al., 1993). Letcher et al. (1996) found PCBs 156 and 157 to be the predominant TEQ contributors in polar bears. They found a strong positive correlation between hepatic TEQ level and content of the protein probably respon- sible for metabolism of dioxin-like compounds, cyto- chrome P450 1A. Thus, as the polar bear is able to metabolise several of the OC compounds regarded as particularly toxic, it may handle the threat of high OC exposure better than most other species.
CONCLUSIONS
In this study, extremely high levels of PCBs are found in polar bears at Svalbard. The various OC compounds determined differ in toxicokinetics, and higher chlori- nated PCBs accumulate with age, particularly in males. In females, considerable amounts of most OCs except the highest chlorinated PCBs are excreted into milk. No relation between OC level in females and their ability to get pregnant is found. The capability to metabolise sev- eral toxic OCs may protect the polar bear against toxic action. However, possible effects of the age-accumula- ted concentrations of PCBs and of the high OC expo- sure with milk at a period of growth and development should be further investigated in polar bears.
ACKNOWLEDGEMENTS
The authors would like to thank The Norwegian Research Council for fundings. Appreciation is expressed to Erna Stai and Siri Foreid for technical assistance, and to Dag Marcus Eide for statistical advice, all employed at Norwegian College of Veterinary Medi- cine. We are grateful to Andy Derocher, Norwegian Polar Institute, for constructive criticism of the manu- script.
REFERENCES
Addison, R. F. & Brodie, P. F. (1987). Transfer of organo- chlorine residues from blubber through the circulatory system to milk in the lactating grey seal Halichoerus grypus. Can. J. Fish. Aquat. Sci., 44, 782-786.
Aguilar, A. & Borrell, A. (1988). Age- and sex-related changes in organochlorine compound levels in fin whales (Balaenop- tera physalus) from the Eastern North Atlantic. Mar. Environ. Res., 25, 195-211.
Ahlborg, U. G., Hanberg, A. & Kenne, K. (1992). Risk assessment of polychlorinated biphenyls (PCBs). Nord, 26, 1-99.
Ahlborg, U. G., Becking, G. C., Birnbaum, L. S., Brouwer, A., Derks, H. J. G. M., Feeley, M., Golor, G., Hanberg, A., Lar- sen, J. C., Liem, A. K. D., Safe, S. H., Schlatter, C., W~ern, F.Younes, M. & Yrj/inheikki, E. (1994). Toxic equivalency factors for dioxin-like PCBs. Chemosphere, 28, 1049-1067.
Altman, D. G. (1991). Describing data. In Practical Statistics for Medical Research, ed. D. G. Altman. Chapman and Hall, London, UK, pp. 19-47.
Arnould, J. P. Y. & Ramsay, M. A. (1994). Milk production and milk consumption in polar bears during the ice-free per- iod in western Hudson Bay. Can. J. Zool., 72, 1365-1370.
Ballschmiter, K. & Zell, M. (1980). Analysis of polychlorina- ted biphenyls (PCB) by glass capillary gas chromatography. Fresenius Z. Anal. Chem., 302, 20-31.
Barrie, L. A., Gregor, D., Hargrave, B., Lake, R., Muir, D., Shearer, R., Tracey, B. & Bidleman, T. (1992). Arctic con- taminants: sources, occurrence and pathways. Sci. Tot. Environ., 122, 1-74.
Bergman, A. & Olsson, M. (1985). Pathology of Baltic grey seal and ringed seal females with special reference to adre- nocortical hyperplasia: Is environmental pollution the cause of a widely distributed disease syndrome? Finnish Game Res., 44, 47-62.
Bernhoft, A. & Skaare, J. U. (1994). Levels of selected indivi- dual polychlorinated biphenyls in different tissues of har- bour seals (Phoca vitulina) from the southern coast of Norway. Environ. Pollut., 86, 99-107.
Bignert, A., Olsson, A., de Wit, C., Litzrn, K., Rappe, C. & Reuterg~rdh, L. (1994). Biological variation --an important factor to consider in ecotoxicological studies based on envir- onmental samples. Fresenius J. Anal. Chem., 348, 76-85.
Boon, J. P., Oostingh, I., van der Meer, J. & Hillebrand, M. T. J. (1994). A model for the bioaccumulation of chlorobiphenyl congeners in marine mammals. Eur. J. Pharmacol. Environ. Toxicol. Pharmacol. Sec., 270, 237-251.
Bowes, G. W. & Jonkel, C. J. (1975). Presence and distribution of polychlorinated biphenyls (PCB) in arctic and subarctic marine food chains. J. Fish Res. Board Can., 32, 2111-2123.
Brevik, E. M. (1978). Gas chromatographic method for the determination of organochlorine pesticides in human milk. Bull. Environ. Contain. ToxicoL, 19, 281-286.
Clausen, J. & Berg, O. (1976). The content of polychlorinated hydrocarbons in Arctic ecosystems. Pure Appl. Chem., 42, 223-232.

174 A. Bernhoft et al.
Collier, R. J. (1985). Nutritional, metabolic, and enviromental aspects of lactation. In Lactation, ed. B. L. Larson. The Iowa State University Press, Ames, Iowa, USA, pp. 80-128.
Daelemans, F. F., Mehlum, F., Lydersen, C. & Schepens, P. J. C. (1993). Mono-ortho and non-ortho substituted PCBs in Arctic ringed seal (Phoca hispida) from the Svalbard area: Analyses and determination of their toxic threat. Chemo- sphere, 27, 429-437.
Derocher, A. E. (1991). Population dynamics and ecology of polar bears in Western Hudson Bay. Ph.D. thesis, Univer- sity of Alberta, Edmonton.
Derocher, A. E., Andriashek, D. & Arnould, J. P. Y. (1993). Aspects of milk composition and lactation in polar bears. Can. J. Zool., 71, 561-567.
Ecobichon, D. J. (1996). Toxic effects of pesticides. In Casarett and Doulls Toxicology. The Basic Science of Poisons, ed. C. D. Klaassen. McGraw-Hill, New York, USA, pp. 643-689.
Erickson, M. D. (1986). Physical, chemical, commercial, environmental, and biological properties of PCBs. In Ana- lytical Chemistry ofPCBs, ed. M. D. Erickson. Butterworth Publishers, Boston, MA, USA, pp. 5-40.
Espeland, O., Kleivane, L., Haugen, S. & Skaare, J. U. (1997). Organochlorines in mother and pup pairs in two Arctic seal species, harp seal (Phoca groenlandica) and hooded seal (Cystophora cristata). Mar. Environ. Res., in press.
Eubank, R. L. (1988). Smoothing splines. In Spline Smoothing and Nonparametric Regression, cd. R. J. Eubank. Marcel Dekker Inc., New York, USA, pp. 189-273.
Fancy, S. G., Pank, L. F., Douglas, D. C., Curby, G. W., Garner, G. W., Amstrup, S. C. & Regelin, W. L. (1988). Satellite telemetry: a new tool for wildlife research and management. US Fish Wildl. Serv., Resour. Publ. 172.
Gabrielsen, G. W., Skaare, J. U., Polder, A. & Bakken, V. (1995). Chlorinated hydrocarbons in glaucous gulls (Larus hyberboreus) in the southern part of Svalbard. Sci. Tot. Environ., 160/161, 337-346.
Geyer, H. J., Scheunert, I. & Korte, F. (1987). Correlation between the bioconcentration potential of organic environ- mental chemicals in humans and their n-octanol/water par- tition coefficients. Chemosphere, 16, 239-252.
Grue, H. & Jensen, B. (1979). Review of the formation of incremental lines in tooth cementum of terrestrial mammals. Danish Rev. Game Biol., II, 1-48.
Hargrave, B. T., Harding, G. C., Vass, W. P., Erickson, P. E., Fowler, B. R. & Scott, V. (1992). Organochlorine pesticides and polychlorinated biphenyls in the Arctic ocean food web. Arch. Environ. Contam. Toxicol., 22, 41-54.
Harris, R. B., Fancy, S. G., Douglas, D. C., Garner, G. W., Amstrup, S. C., McCabe, T. R. & Pank, L. F. (1990). Tracking wildlife by satellite: current systems and perfor- mance. US Star. Dept. Inter. Fish. Wildl. Serv., Fish Wildl. Tech. Rep., 30, 1-52.
Hayes, W. J. (1982a). Chlorinated hydrocarbon insecticides. In Pesticides Studied in Man, ed. W. J. Hayes. Williams & Wiikins, Baltimore, USA, pp. 172-260.
Hayes, W. J. (1982b). Fungicides and related compounds. In Pesticides Studied in Man, ed. W. J. Hayes. Williams & Wilkins, Baltimore, USA, pp. 578-614.
Helle, E., Olsson, M. & Jensen, S. (1976). DDT and PCB levels and reproduction in ringed seal from the Bothnian Bay. Ambio, 5, 188-189.
Iwata, H., Tanabe, S., Sakai, N. & Tatsukawa, R. (1993). Distribution of persistent organochlorines in the oceanic air and surface seawater and the role of ocean on their global transport and fate. Environ. Sci. Technol., 27, 1080-1098.
Jensen, J. (1991). Report on organochlorines. In The State of the Arctic Environment Reports, Attic Centre Publications 2. ISBN 951-634-231-0, University of Lapland, Rovaniemi, Finland, pp. 325-384.
Kleivane, L., Skaare, J. U., Bj~rge, A., de Ruiter, E. & Reijn- ders, P. J. H. (1995). Organochlorine pesticide residue and
PCBs in harbour porpoise (Phocoena phocoena) incidentally caught in Scandinavian waters. Environ. Pollut., 89, 137- 146.
Letcher, R. J., Norstrom, R. J., Lin, S., Ramsay, M. A. & Bandiera, S. M. (1996). Immunoquantitation and microso- mal monooxygenase activities of hepatic cytochromes P4501A and P4502B and chlorinated hydrocarbon con- taminant levels in polar bear (Ursus maritimus). Toxicol. Appl. Pharmacol., 137, 127-140.
Long, C. (1961). Composition of blood. In Biochemists' Handbook, ed. C. Long. Spon Ltd., London, UK, pp. 839- 885.
Martineau, D., B61and, P., Desjardins, C. & Lagac6, A. (1987). Levels of organochlorine chemicals in tissues of beluga whales (Delphinapterus leucas) from the St. Lawrence Estuary, Qu6bec, Canada. Arch. Environ. Contain. Toxicol., 16, 137-147.
Matthews, H. B. & Dedrick, R. L. (1984). Pharmacokinetics of PCBs. Ann. Rev. Pharmacol. Toxicol., 24, 85-103.
Matthews, H. B., Surles, J. R., Carver, J. G. & Anderson, M. W. (1984). Halogenated biphenyl transport by blood com- ponents. Fundam. Appl. Toxicol., 4, 420-428.
Mehlum, F. & Daelemans, F. F. (1995). PCBs in Arctic sea- birds from the Svalbard region. Sci. Tot. Environ., 160]161, 441-446.
Mes, J., Marchand, L., Tuton, D. & Lau, P.-Y. (1992). The determination of polychlorinated biphenyl congeners and other chlorinated hydrocarbon residues in human blood, serum and plasma. A comparative study. Intern. J. Environ. Anal. Chem., 48, 175-186.
Muir, D. C. G., Norstrom, R. J. & Simon, M. 0988). Orga- nochlorine contaminants in Arctic marine food chains: Accumulation of specific polychlorinated biphenyls and chlordane-related compounds. Environ. Sci. Technol., 22, 1071-1079.
Muir, D. C. G., Wagemann, R., Hargrave, B. T., Thomas, D. J., Peacall, D. B. & Norstrom, R. J. (1992). Arctic marine ecosystem contamination. Sci. Tot. Environ., 122, 75-134.
Norheim, G., Skaare, J. U. & Wiig, 13. (1992). Some heavy metals, essential, elements, and chlorinated hydrocarbons in polar bear (Ursus maritimus) at Svalbard. Environ. Pollut., 77, 51-57.
Norstrom, R. J. & Muir, D. C. G. (1994). Chlorinated hydro- carbon contaminants in arctic marine mammals. Sci. Tot. Environ., 154, 107-128.
Norstrom, R. J., Simon, M., Muir, D. C. G. & Schweinsburg, R. E. (1988). Organochlorine contaminants in arctic marine food chains: Identification, geographical distribution, and temporal trends in polar bears. Environ. Sci. Technol., 22, 1063-1071.
Norstrom, R. J., Simon, M. & Muir, D. C. G. (1990). Poly- chlorinated dibenzo-p-dioxins and dibenzofurans in marine mammals in the Canadian North. Environ. Pollut., 66, 1-19.
Norstrom, R. J., Malone, B., Born, E. W., Garner, G. W., Olpinski, S., Ramsay, M. A., Schliebe, S., Stifling, I., Taylor, M. K. & Wiig, 0. (1996). Chlorinated hydrocarbon contaminants in polar bears from Eastern Russia, North America, Greenland and Svalbard: Biomonitoring of hemispheric pollution. Manuscript.
Oehme, M. (1991). Dispersion and transport paths of toxic persistent organochlorines to the Arctic Levels and conse- quences. Sci. Tot. Environ., 106, 43-53.
Oehme, M., Fiirst, P., Krfiger, C., Meemken, H. A. & Groe- bei, W. (1988). Presence of polychlorinated dibenzo-p-diox- ins, dibenzofurans and pesticides in Arctic seal from Spitzbergen. Chemosphere, 17, 1291-1300.
Oehme, M., Biseth, A., Schlabach, M. & Wiig, I~. (1995). Concentrations of polychlorinated dibenzo-p-dioxins, dibenzofurans and non-ortho substituted bipbenyls in polar bear milk from Svalbard (Norway). Environ. Pollut., 90, 401-407.

Organochlorines in polar bears 175
Olsson, M., Andersson, 6., Bergman, ,A., Blomkvist, G., Frank, A. & Rappe, C. (1992). Contaminants and diseases in seals from Swedish Waters. Ambio, 21, 561-562.
Peterson, R. E., Theobald, H. M. & Kimmel, G. L. (1993). Developmental and reproductive toxicity of dioxins and related compounds: Cross-species comparisons, Crit. Rev. Toxicol., 23, 283-335.
Polischuk, S. C., Letcher, R. J., Norstrom, R. J. & Ramsay, M. A. (1995). Preliminary results of fasting on the kinetics of organochlorines in polar bears (Ursus maritimus). Sci. Tot. Environ., 160/161, 465-472.
Ramsay, M. A. & Stifling, I. (1988). Reproductive biology and ecology of female polar bears (Ursus maritimus). J. Zool. Lond., 214, 601-634.
Safe, S. (1984). Polychlorinated biphenyls (PCBs) and poly- brominated biphenyls (PBBs): Biochemistry, toxicology, and mechanism of action. Crit. Rev. Toxicol., 13, 319-395.
Safe, S. (1990). Polychlorinated biphenyls (PCBs), dibenzo-p- dioxins (PCDDs), dibenzofurans (PCDFs), and related compounds: Environmental and mechanistic conciderations which support the development of toxic equivalency factors (TEFs). Crit. Rev. Toxicol., 21, 51-88.
Sandell, M. (1990). The evolution of seasonal delayed implantation. Quart. Rev. Biol., 65, 2342.
Skaare, J. U., Wiig, 13. & Bernhoft, A. (1994). Klorerte orga- niske miljogifter; NivAer og effekter pA isbj~rn. Norsk Polarinstitutt, rapport, 86, 1-23.
Stifling, I. & Archibald, W. R. (1977). Aspects of predation of seals by polar bears. J. Fish. Res. Board Can., 34, 1126- 1129.
Takagi, Y., Aburada, S., Hashimoto, K. & Kitaura, T. (1986). Transfer and distribution of accumulated (14C) polychlori- nated biphenyls from maternal to fetal and suckling rats. Arch. Environ. Contam. Toxicol., 15, 709-715.
Tanabe, S., Iwata, H. & Tatsukawa, R. (1994). Global con- tamination by persistent organochlorines and their ecotoxi- cological impact on marine mammals. Sci. Tot. Environ., 154, 163-177.
Tiison, H. A., Jacobson, J. L. & Rogan, W. J. (1990). Polychlor- inated biphenyls and the developing nervous system: Cross- species comparisons. Neurotoxicol. Teratol., 12, 239-248.
Timbreil, J. A. (1991). Factors affecting metabolism and dis- position. In Principles of biochemical toxicology, ed. J. A. Timbrell. Taylor and Francis, London, UK, pp. 125-191.
Wang-Andersen, G., Skaare, J, U., Prestrud, P. & Steinnes, E. (1993). Levels and congener pattern of PCBs in Arctic fox, Alopex lagopus, in Svalbard. Environ. Pollut., 82, 269-275.
Wiig, 121. (1995). Distribution of polar bears (Ursus maritimus) in the Svalbard area. J. Zool., Lond., 237, 515-529.
Wiig, I~. (1996). Population biology of polar bears from Sval- bard. Int. Conf. Bear Res. Manage., submitted.
Wiig, gl., Gjertz, I., Hansson, R. & Thomassen, J. (1992). Breeding of polar bears in Hornsund, Svalbard. Polar Rec., 28, 157-159.
Yang, R. S. H., Coulston, F. & Goldberg, L. (1975). Binding of hexachlorobenzene to erythrocytes: Species variation. Life Sci., 17, 545-550.
Zhu, J. & Norstrom, R. J. (1993). Identification of polychlor- ocamphenes (PCCs) in the polar bear (Ursus maritimus) food chain. Chemosphere, 27, 1923-1936.




















