
Histology of selected immunological organs in polar bear (Ursus maritimus) from East Greenland in...
14
Histology of selected immunological organs in polar bear (Ursus maritimus ) from East Greenland in relation to concentrations of organohalogen contaminants M. Kirkegaard a,b, * , C. Sonne a,c , P.S. Leifsson b , R. Dietz a , E.W. Born d , D.C.G. Muir e , R.J. Letcher f a Department of Arctic Environment, National Environmental Research Institute, Frederiksborgvej 399, 4000 Denmark b Department of Pharmacology and Pathobiology, The Royal Veterinary and Agricultural University, Bu ¨lowsvej 17, 1870 Denmark c Department of Anatomy and Physiology, The Royal Veterinary and Agricultural University, Bu ¨lowsvej 17, 1870 Denmark d Greenland Institute of Natural Resources, P.O. Box 570, DK-3900 Nuuk, Greenland, Denmark e National Water Research Institute, Environment Canada, Burlington, ON Canada L7R 4A6 f Great Lakes Institute for Environmental Research, University of Windsor, Windsor ON Canada N9B 3P4 Received 20 May 2004; accepted 22 September 2004 Abstract Samples of lymph nodes (axillary, n =54 and inguinal, n =45), spleen (n =60), thymus (n =11) and thyroid tissue (n =5) from a total of 82 polar bears (Ursus maritimus ) collected in East Greenland 1999–2002 were examined histologically. The purpose was to relate histology to concentrations of organohalogen contaminants (OHCs) [i.e., sum (A)PCBs, ADDTs, AHCHs, ACHLs, HCB, Dieldrin and APBDEs] determined in adipose tissue, as studies on polar bears have indicated that some OHCs act as immunosuppressive agents. Secondary follicle counts were evaluated in spleen and lymph nodes, and semiquantitively divided into four groups (0: few/absent to 3: high). In the spleen, a high secondary follicle count was found in 21% of the cases (12/60), and this was significantly higher in subadults ( p b0.01) compared to adults of both sexes. Also in the lymph nodes a high secondary follicle count was found in 20% of the cases (20/99), and in the axillary lymph nodes changes were significantly higher in subadults ( p b0.05) compared to adults of both sexes. Significantly weak correlations between concentrations of OHCs and the amount of secondary follicles in lymph nodes was found, but probably occurred as a consequence of other multiple stress factor(s) (all: pb0.05) and also one significant, but modest positive correlation was found between APBDE concentrations and secondary follicle counts ( p b0.01; r =0.41). In spleen, a significant relation between low concentrations of OHCs in adipose tissue and few/absent secondary follicles was found with respect to ACHLs, AHCHs, HCB and Dieldrin. No histopathological observations (e.g., neoplasia) were found in spleen or lymph nodes, nor in thymus or thyroid. In conclusion, the present data suggest that the exposure concentrations of OHCs to polar bears are unlikely to have resulted in adverse effects 0048-9697/$ - see front matter D 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.scitotenv.2004.09.034 * Corresponding author. National Environmental Research Institute, Department of Arctic Environment, Frederiksborgvej 399, DK-4000 Roskilde, Denmark. Tel.: +45 46 30 19 53; fax: +45 46 30 19 53. E-mail address: [email protected] (M. Kirkegaard). Science of the Total Environment 341 (2005) 119– 132 www.elsevier.com/locate/scitotenv
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Histology of selected immunological organs in polar bear (Ursus maritimus) from East Greenland in...

www.elsevier.com/locate/scitotenv
Science of the Total Environm
Histology of selected immunological organs in polar bear
(Ursus maritimus) from East Greenland in relation to
concentrations of organohalogen contaminants
M. Kirkegaarda,b,*, C. Sonnea,c, P.S. Leifssonb, R. Dietza,
E.W. Bornd, D.C.G. Muire, R.J. Letcherf
aDepartment of Arctic Environment, National Environmental Research Institute, Frederiksborgvej 399, 4000 DenmarkbDepartment of Pharmacology and Pathobiology, The Royal Veterinary and Agricultural University, Bulowsvej 17, 1870 Denmark
cDepartment of Anatomy and Physiology, The Royal Veterinary and Agricultural University, Bulowsvej 17, 1870 DenmarkdGreenland Institute of Natural Resources, P.O. Box 570, DK-3900 Nuuk, Greenland, DenmarkeNational Water Research Institute, Environment Canada, Burlington, ON Canada L7R 4A6
fGreat Lakes Institute for Environmental Research, University of Windsor, Windsor ON Canada N9B 3P4
Received 20 May 2004; accepted 22 September 2004
Abstract
Samples of lymph nodes (axillary, n=54 and inguinal, n=45), spleen (n=60), thymus (n=11) and thyroid tissue (n=5) from a
total of 82 polar bears (Ursus maritimus) collected in East Greenland 1999–2002 were examined histologically. The purpose
was to relate histology to concentrations of organohalogen contaminants (OHCs) [i.e., sum (A)PCBs, ADDTs, AHCHs, ACHLs,HCB, Dieldrin and APBDEs] determined in adipose tissue, as studies on polar bears have indicated that some OHCs act as
immunosuppressive agents. Secondary follicle counts were evaluated in spleen and lymph nodes, and semiquantitively divided
into four groups (0: few/absent to 3: high). In the spleen, a high secondary follicle count was found in 21% of the cases (12/60),
and this was significantly higher in subadults ( pb0.01) compared to adults of both sexes. Also in the lymph nodes a high
secondary follicle count was found in 20% of the cases (20/99), and in the axillary lymph nodes changes were significantly
higher in subadults ( pb0.05) compared to adults of both sexes. Significantly weak correlations between concentrations of
OHCs and the amount of secondary follicles in lymph nodes was found, but probably occurred as a consequence of other
multiple stress factor(s) (all: pb0.05) and also one significant, but modest positive correlation was found between APBDEconcentrations and secondary follicle counts ( pb0.01; r=0.41). In spleen, a significant relation between low concentrations of
OHCs in adipose tissue and few/absent secondary follicles was found with respect to ACHLs, AHCHs, HCB and Dieldrin. No
histopathological observations (e.g., neoplasia) were found in spleen or lymph nodes, nor in thymus or thyroid. In conclusion,
the present data suggest that the exposure concentrations of OHCs to polar bears are unlikely to have resulted in adverse effects
0048-9697/$ - s
doi:10.1016/j.sc
* Correspondi
Roskilde, Denm
E-mail addr
ent 341 (2005) 119–132
ee front matter D 2004 Elsevier B.V. All rights reserved.
itotenv.2004.09.034
ng author. National Environmental Research Institute, Department of Arctic Environment, Frederiksborgvej 399, DK-4000
ark. Tel.: +45 46 30 19 53; fax: +45 46 30 19 53.
ess: [email protected] (M. Kirkegaard).

M. Kirkegaard et al. / Science of the Total Environment 341 (2005) 119–132120
on the tissues in question, although ACHLs, AHCHs, HCB and Dieldrin were related to increased secondary follicle counts in
the spleen.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Polar bear; Ursus maritimus; OHCs; Lymph node; Spleen; Thyroid; Thymus
1. Introduction
Generally, organohalogen contaminant (OHC) and
especially polychlorinated biphenyl (PCB) exposure
is believed to negatively affect reproduction and
survival in marine mammals, e.g., in seals from the
Baltic Sea (Bergman and Olsson, 1985; Bergman,
1999) and in polar bears (Ursus maritimus) from
Canadian and Svalbard populations (Letcher et al.,
1996; Wiig et al., 1998; Bernhoft et al., 2000; Lie et
al., 2004a,in press; Skaare et al., 2001a; Haave et al.,
2003; Oskam et al., 2003). Immunosuppressive
effects of OHC exposure may be an environmental
issue in marine species (Vos and Luster, 1989), e.g.,
such effects were found to correlate with concen-
trations of aryl hydrocarbon receptor (AhR)-active
OHCs including non-ortho PCBs [e.g., 3,3V,4,4V-tetrachlorobiphenyl (CB-77) and 3,3V,4,4V,5-penta-chlorobiphenyl (CB-126)] and polychlorinated
dibenzo-p-dioxins/furans (PCDDs/Fs) in harbour
seals (Phoca vitulina) fed Baltic fish under semifield
experiments (Swart et al., 1994; Ross et al., 1995,
1996). Non-ortho PCBs and PCDDs/Fs are also
suspected to cause an increasing prevalence of
intestinal ulcers in Baltic grey seals (Halichoerus
grypus) (Bergmann et al., 1996; Bergman, 1999).
In wildlife, histopathological changes in kidneys,
adrenals, thyroids a.o. organs have been correlated to
concentrations of OHCs in several marine mammals
such as harbour seal, Greg seal, ringed seal (Phoca
hispida botnica) and harbour porpoise (Phocoena
phocoena) (Bergman and Olsson, 1985; Schumacher
et al., 1993; Bergman et al., 2001, Siebert et al., 2002) as
well as in fish (bream, Abramis brama) (Koponen et al.,
2001). In harbour porpoise some OHCs have direct
affect on the thymus, causing atrophy, and some affect
the immune system by increasing lymphocyte depletion
from lymphatic organs (Siebert et al., 2002). Immuno-
suppressive effects aremeasured by biomarkers, such as
reduced antibody production when exposed to a foreign
antigen, changes in T-cell populations, decreased
delayed-type hypersensitivity, decreased resistance to
virus infections and decreased natural killer cell activity
(Wong et al., 1992; Tryphonas, 1994).
In polar bear studies from Canada and Svalbard in
1998 and 1999, results show that organochlorines
(OHCs) in combination with specific biological factors
(sex, age, condition, body mass) significantly influence
serum IgG concentrations and antibody production
after immunisation with influenza and reo viruses and
tetanus toxoid. In addition, the results demonstrate in
particular, a negative association between APCBs andthe serum IgG concentrations or production of anti-
bodies to influenza and reo viruses (Lie et al., 2004a).
For lymphocyte proliferation, the results of the
statistical analyses show that high concentrations of
PCBs and/or pesticides were found to decrease specific
lymphocyte function and thus may produce impaired
resistance against infections (Lie et al., in press). The
study demonstrates that high concentrations of APCBsinfluence parts of the humoral immunity in polar bears,
as the ability to produce antibodies following immu-
nisation was impaired (Lie et al., 2004a).
The OHCs classes PCBs, dichlorodiphenyltri-
chloroethanes (DDTs), hexachlorocyclohexanes
(HCHs), chlordanes (CHLs) and PCDDs/Fs that are
present in the tissues of polar bears, have shown to be
transferred transplacentally from mother to foetus in
utero and to cubs via mother’s milk (Bernhoft et al.,
1997; Polischuk et al., 2002). Many of these com-
pounds have demonstrated or have the potential to
function as agonists and/or antagonists with respect to
hormone-dependent processes in endocrine target
organs and tissues (Colborn et al., 1993; De March et
al., 1998; Damstra et al., 2002; AMAP, 2004). OHCs in
exposed organisms can interfere with the main endo-
crine pathway, which may result in disruption of
different endocrine target organs like adrenals, thyroid,
parathyroid, thymus, testicles and ovaries.Mechanisms
resulting in the disruption of endocrine homeostasis
can occur due to modulation/interference of the
concentrations of corticosterone, thyroid hormones

M. Kirkegaard et al. / Science of the Total Environment 341 (2005) 119–132 121
[thyronine (T3) and thyroxine (T4)] and other sex
hormones (Colborn et al., 1993; Feldman, 1995; De
March et al., 1998; Bergman, 1999; Damstra et al.,
2002; AMAP, 2004). OHC exposure can induce
hypothalamus-mediated stress resulting in hyperadre-
nocorticism (adrenocortical hyperplasia), which is a
clinical condition in mammals caused by high concen-
trations of cortisol (glucocorticoids). High glucocorti-
coid concentrations interfere with metabolism in
relation to the immune system leading to anti-inflam-
matory and immune-suppressive responses. In the
Baltic ringed seal and grey seal, hyperadrenocorticism
has been correlated to high concentrations of PCBs and
DDTs (Bergman and Olsson, 1985; Feldman, 1995; De
March et al., 1998; Bergman, 1999; AMAP, 2004).
In order to assess whether relatively high concen-
trations of OHCs in tissues may be linked to
deleterious effects on health of polar bears in East
Greenland, samples of lymph nodes, spleen, thyroid
and thymus tissues were collected 1999–2002 from a
total of 82 polar bears and analysed histologically in
relation to and compared to the OHC residue
concentrations in adipose tissue of these bears.
2. Materials and methods
2.1. Sampling
Fat samples from polar bears were collected by
local subsistence hunters in the Ittoqqortoormiit/Score-
sby Sound area in central East Greenland between
69800VN and 74800VN in 1999–2001. All tissue
samples were taken as soon as possible postmortem
and stored in separate polyethylene (PE) Whirlpak
bags. All samples were kept at outdoor temperature
(�5 to �20 8C) until transferred to a freezer (�10 to
�20 8C). Samples were shipped frozen from Scoresby
Sound to Roskilde, where the portion of fat that had
been in contact with the PE was trimmed off and the
remaining part was transferred to precleaned glass
containers with cleaned aluminium foil in between the
lid and the glass container. Further storage was at�20
8C. Samples from spleen (n=60), lymph nodes
[axillary (n=54) and inguinal (n=45)], thyroid (n=5)
and thymus (n=11) were stored in a solution of
phosphate buffered one part formaldehyde (5% sol-
ution) and nine parts ethanol (96% solution) to prevent
freezing damage. The thymus undergoes involution
after maturation (Dellman and Eurell, 1998), after
which it becomes small, similar in size to the thyroid.
This relatively small lobe size in adults made it
difficult for the Inuit hunters to locate; therefore, only
a few samples of thyroid and thymus were obtained.
2.2. Age determination
The age determination was carried out by counting
the cementum Growth Layer Groups (GLG) of the I3(or P1) tooth using the methods described by Hensel
and Sorensen (1980) and Dietz et al. (1991). Based on
the assigned ages the animals were grouped into adult
females (z5 years), adult males (z6 years) and the
remaining as subadults (Norstrom et al., 1998;
Henriksen et al., 2001).
2.3. Histology
All tissue samples were trimmed, processed con-
ventionally, embedded in paraffin (Sakura Tissue
TekRVIP, Sakura Finetek, USA), and cut into 2–4
Am slices. Sections were stained with haematoxylin-
eosin (HE) for histological examination. All slides
were examined with a Leica DMLB microscope with
50, 100, 200, 400 and 1000� magnification (pictures
were taken and stored by a Leica DC300 digital
camera). Histological findings (secondary follicles)
were assessed semiquantitatively and assigned to four
groups (index) as few/absent (group 0), low (group 1),
moderate (group 2), high (group 3). The index was
calculated as an average of secondary follicles from
repeated counts in four randomly selected low power
fields (50� magnification). In lymph nodes group 0
represented 0–3 secondary follicles, group 1: 4–5
secondary follicles, group 2: 6–8 secondary follicles
and group 3: 9–18 secondary follicles. In spleen
groups were; 0: 0–1 secondary follicles, group 1: 2
secondary follicles, group 2:3 secondary follicles and
group 3: 4–8 secondary follicles.
2.4. Contaminant analyses
2.4.1. PCBs and OHCs
Polar bear blubber samples (n=94)were analysed for
polychlorinatedbiphenyls (PCBs), dichlorodiphenyltri-
chloroethanes (DDTs), hexacyclohexanes (HCHs),
M. Kirkegaard et al. / Science of the Total Environment 341 (2005) 119–132122
CHLordanes (CHLs), hexachlorobenzene (HCB) and
dieldrin according to Sandala et al. (2004) and Dietz et
al. (2004) at theGreat Lakes Institute for Environmental
Research (GLIER), University ofWindsor, Canada. An
external standard quantification approach used for
PCBs and OHCs in the adipose tissues was based on
peak area of the GC-AECD response, which is
described in detail in Dietz et al. (2004). Briefly,
APCBs is the sum (A) of the concentrations of the 51individual or coeluting congeners (if detected): CB #
31/28, 52, 49, 44, 42, 64/71, 74, 70, 66/95, 60, 101/84,
99, 97, 87, 110, 151, 149, 118, 146, 153, 105, 141, 179,
138, 158, 129/178, 182/187, 183, 128, 174, 177, 171/
202/156, 200, 172, 180, 170/190, 201, 203/196, 195,
194, 206. CB congeners are numbered according to the
corrected IUPAC numbering scheme as described by
Guitart et al. (1993). ADDTs is the sum of 4,4V-DDT,4,4V-DDD and 4,4V-DDE. AHCHs is the sum of the a-,
h- and g-hexachlorocyclohexane.ACHLs is the sum of
oxychlordane, trans-chlordane, cis-chlordane, trans-
nonachlor, cis-nonachlor and heptachlor epoxide.
Contaminant fractions were subsequently sent to the
National Water Research Institute [Environment Can-
ada, Burlington, Ontario, Canada L7R 4A6 (NWRI)]
for determination of brominated diphenyl ether (PBDE)
flame retardants.
2.4.2. PBDEs
PBDEs (n=93) were determined by electron
capture negative ion (low resolution) MS using an
external standard. Briefly, APBDEs is the sum (A) ofthe concentrations of the 35 individual or coeluting
IUPAC congeners (if detected): BDE# 10, 7, 11, 8, 12/
13, 15, 30, 32, 28/33, 35, 37, 75, 71, 66, 47, 49, 77,
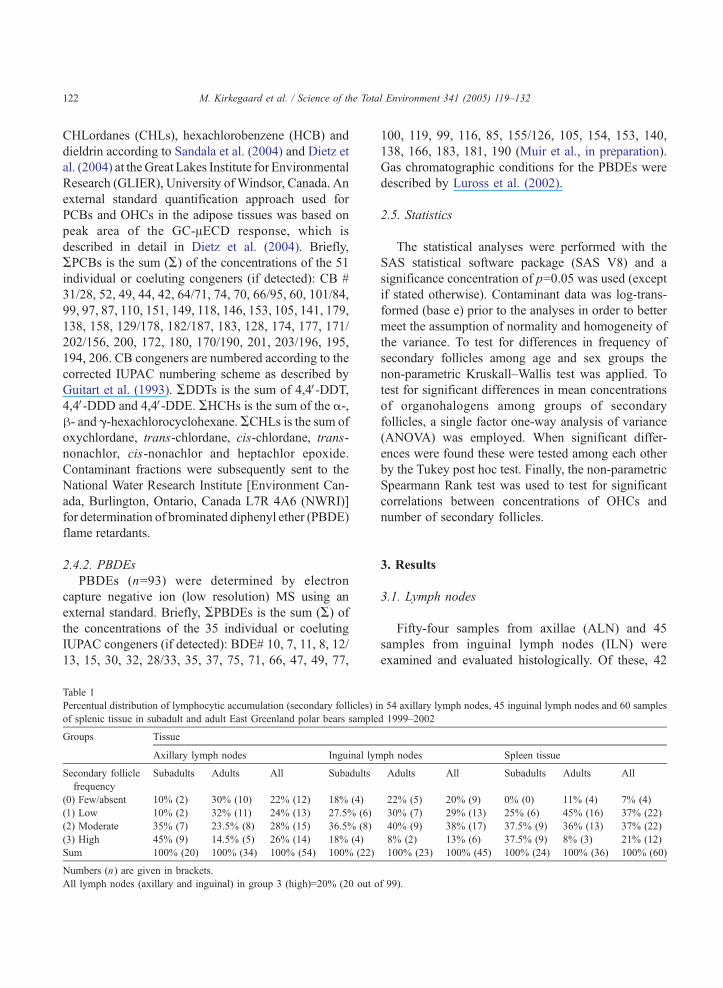
Table 1
Percentual distribution of lymphocytic accumulation (secondary follicles) i
of splenic tissue in subadult and adult East Greenland polar bears sample
Groups Tissue
Axillary lymph nodes Inguinal lym
Secondary follicle
frequency
Subadults Adults All Subadults
(0) Few/absent 10% (2) 30% (10) 22% (12) 18% (4)
(1) Low 10% (2) 32% (11) 24% (13) 27.5% (6)
(2) Moderate 35% (7) 23.5% (8) 28% (15) 36.5% (8)
(3) High 45% (9) 14.5% (5) 26% (14) 18% (4)
Sum 100% (20) 100% (34) 100% (54) 100% (22)
Numbers (n) are given in brackets.
All lymph nodes (axillary and inguinal) in group 3 (high)=20% (20 out o
100, 119, 99, 116, 85, 155/126, 105, 154, 153, 140,
138, 166, 183, 181, 190 (Muir et al., in preparation).
Gas chromatographic conditions for the PBDEs were
described by Luross et al. (2002).
2.5. Statistics
The statistical analyses were performed with the
SAS statistical software package (SAS V8) and a
significance concentration of p=0.05 was used (except
if stated otherwise). Contaminant data was log-trans-
formed (base e) prior to the analyses in order to better
meet the assumption of normality and homogeneity of
the variance. To test for differences in frequency of
secondary follicles among age and sex groups the
non-parametric Kruskall–Wallis test was applied. To
test for significant differences in mean concentrations
of organohalogens among groups of secondary
follicles, a single factor one-way analysis of variance
(ANOVA) was employed. When significant differ-
ences were found these were tested among each other
by the Tukey post hoc test. Finally, the non-parametric
Spearmann Rank test was used to test for significant
correlations between concentrations of OHCs and
number of secondary follicles.
3. Results
3.1. Lymph nodes
Fifty-four samples from axillae (ALN) and 45
samples from inguinal lymph nodes (ILN) were
examined and evaluated histologically. Of these, 42
n 54 axillary lymph nodes, 45 inguinal lymph nodes and 60 samples
d 1999–2002
ph nodes Spleen tissue
Adults All Subadults Adults All
22% (5) 20% (9) 0% (0) 11% (4) 7% (4)
30% (7) 29% (13) 25% (6) 45% (16) 37% (22)
40% (9) 38% (17) 37.5% (9) 36% (13) 37% (22)
8% (2) 13% (6) 37.5% (9) 8% (3) 21% (12)
100% (23) 100% (45) 100% (24) 100% (36) 100% (60)
f 99).

Fig. 1. Histology of a lymph node from a normal 5-year-old East
Greenland male polar bear sampled 1999 (note secondary lymphatic
follicles in the cortex). Trabeculum (T), Mantle zone (M) and
germinal centre (G). (HE, 100�.).
M. Kirkegaard et al. / Science of the Total Environment 341 (2005) 119–132 123
(ALN: 20; ILN; 22) were from subadults, 37 (ALN: 19;
ILN; 18) from adult males and 20 (ALN: 15; ILN; 5)
from adult females (Table 1). Microscopically the
appearance of polar bear lymph nodes resembled that
of the ringed seal (P. hispida) (Dyce et al., 1987; Habel,
1992; Welsch et al., 1997). The nodes were surrounded
Fig. 2. Lymphocytic histological findings in the four distinguished groups
follicles). (B) Group 1 (4–5 secondary follicles). (C) Group 2 (6–8 seconda
germ center. HE (100�: panels B and C), (50�: panels A and D).
by a fibrous capsule from which fibrous trabeculae
(bTQ; Fig. 1) extended into the node to form a
supporting framework, carrying blood vessels and
nerves. The surface of the lymph nodes was penetrated
by a number of afferent lymphatic vessels, which
opened into a subcapsular sinus, forming networks
extending into the hilus. The lymph cortex consisted of
primary and secondary lymphatic follicles separated by
diffuse lymphatic tissue (Dellman and Eurell, 1998).
The primary follicles consisted of naive B cells and
some memory T cells and the secondary lymphoid
follicles consisted mainly of B cells in various stages in
the secondary follicle (bGQ Fig. 1), characterised by
open nuclei and more cytoplasm. The B-cells were also
less densely packed than the smaller naive B cells and
the few Tcells that formed the darker mantle zone (bMQFig. 1) of the follicle (Stevens and Lowe, 1997).
3.1.1. Histology
None of the changes found in the examination of
the lymph nodes could be categorised as severe
histopathological observations (as neoplasia or fib-
rosis). Secondary follicles were found and distributed
as shown in Table 1 and Fig. 2. For group 3, a
. Arrows indicate secondary follicles. (A) Group 0 (0–3 secondary
ry follicles). (D) Group 3 (9–18 secondary follicles). Arrows indicate
M. Kirkegaard et al. / Science of the Total Environment 341 (2005) 119–132124
higher count of secondary follicles in the axillary
lymph nodes was found for subadults compared to
adult males and females ( pb0.05). No similar
significant differences in the inguinal lymph node
were found.
3.1.2. OHCs
Concentrations of tissue residues (blubber) of
OHCs (ng/g lw) of PCBs (51 congeners), DDTs
(4,4V-DDD, 4,4V-DDE, 4,4V-DDT), HCHs (a-HCH, h-HCH, g-HCH), CHLs (oxychlordane, trans-chlor-
dane, cis-chlordane), HCB, PBDEs (35 congeners)
and dieldrin are described in Dietz et al. (2004).
APCBs and ACHLs constituted the highest concen-
trations followed by ADDTs (4,4V-DDE constituted
the most), dieldrin and AHCHs, while HCB and
APBDEs carried the lowest concentrations. AMeSO2-
PCB and 3-MeSO2-4,4V-DDE concentrations were not
included in these comparisons due to covariance, i.e.,
A-MeSO2-PCB to A-PCBs and 3-MeSO2-4,4V-DDEto 4,4V-DDE ratios for the present Greenland bears
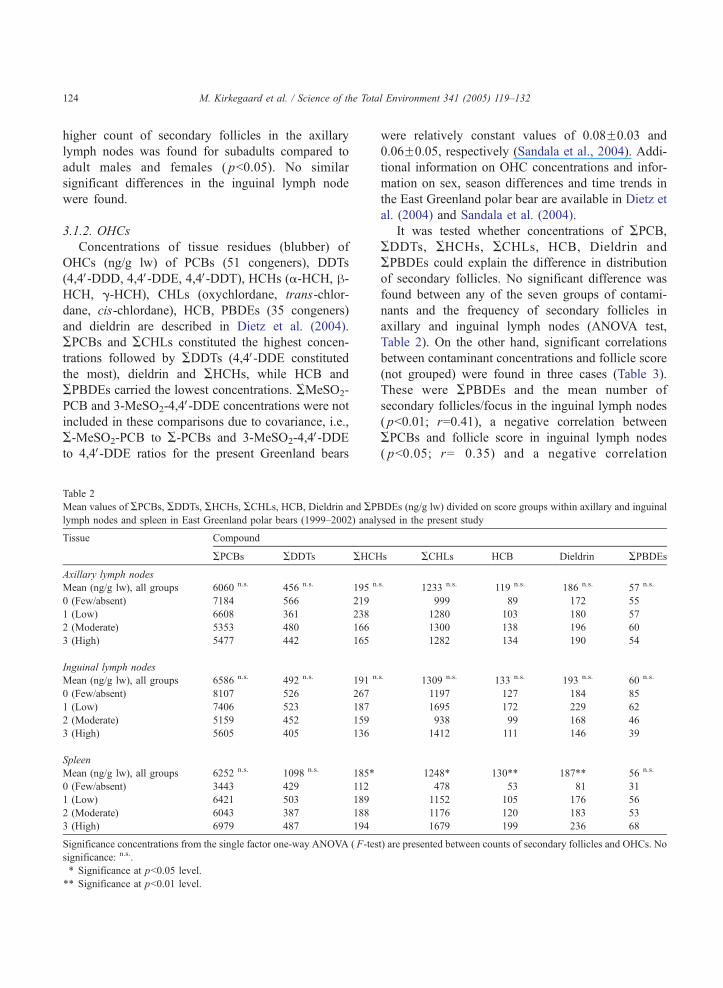
Table 2
Mean values of APCBs, ADDTs, AHCHs, ACHLs, HCB, Dieldrin and APlymph nodes and spleen in East Greenland polar bears (1999–2002) anal
Tissue Compound
APCBs ADDTs AHCH
Axillary lymph nodes
Mean (ng/g lw), all groups 6060 n.s. 456 n.s. 195 n
0 (Few/absent) 7184 566 219
1 (Low) 6608 361 238
2 (Moderate) 5353 480 166
3 (High) 5477 442 165
Inguinal lymph nodes
Mean (ng/g lw), all groups 6586 n.s. 492 n.s. 191 n
0 (Few/absent) 8107 526 267
1 (Low) 7406 523 187
2 (Moderate) 5159 452 159
3 (High) 5605 405 136
Spleen
Mean (ng/g lw), all groups 6252 n.s. 1098 n.s. 185*
0 (Few/absent) 3443 429 112
1 (Low) 6421 503 189
2 (Moderate) 6043 387 188
3 (High) 6979 487 194
Significance concentrations from the single factor one-way ANOVA ( F-tes
significance: n.s..
* Significance at pb0.05 level.
** Significance at pb0.01 level.
were relatively constant values of 0.08F0.03 and
0.06F0.05, respectively (Sandala et al., 2004). Addi-
tional information on OHC concentrations and infor-
mation on sex, season differences and time trends in
the East Greenland polar bear are available in Dietz et
al. (2004) and Sandala et al. (2004).
It was tested whether concentrations of APCB,ADDTs, AHCHs, ACHLs, HCB, Dieldrin and
APBDEs could explain the difference in distribution
of secondary follicles. No significant difference was
found between any of the seven groups of contami-
nants and the frequency of secondary follicles in
axillary and inguinal lymph nodes (ANOVA test,
Table 2). On the other hand, significant correlations
between contaminant concentrations and follicle score
(not grouped) were found in three cases (Table 3).
These were APBDEs and the mean number of
secondary follicles/focus in the inguinal lymph nodes
( pb0.01; r=0.41), a negative correlation between
APCBs and follicle score in inguinal lymph nodes
( pb0.05; r=�0.35) and a negative correlation
BDEs (ng/g lw) divided on score groups within axillary and inguinal
ysed in the present study
s ACHLs HCB Dieldrin APBDEs
.s. 1233 n.s. 119 n.s. 186 n.s. 57 n.s.
999 89 172 55
1280 103 180 57
1300 138 196 60
1282 134 190 54
.s. 1309 n.s. 133 n.s. 193 n.s. 60 n.s.
1197 127 184 85
1695 172 229 62
938 99 168 46
1412 111 146 39
1248* 130** 187** 56 n.s.
478 53 81 31
1152 105 176 56
1176 120 183 53
1679 199 236 68
t) are presented between counts of secondary follicles and OHCs. No
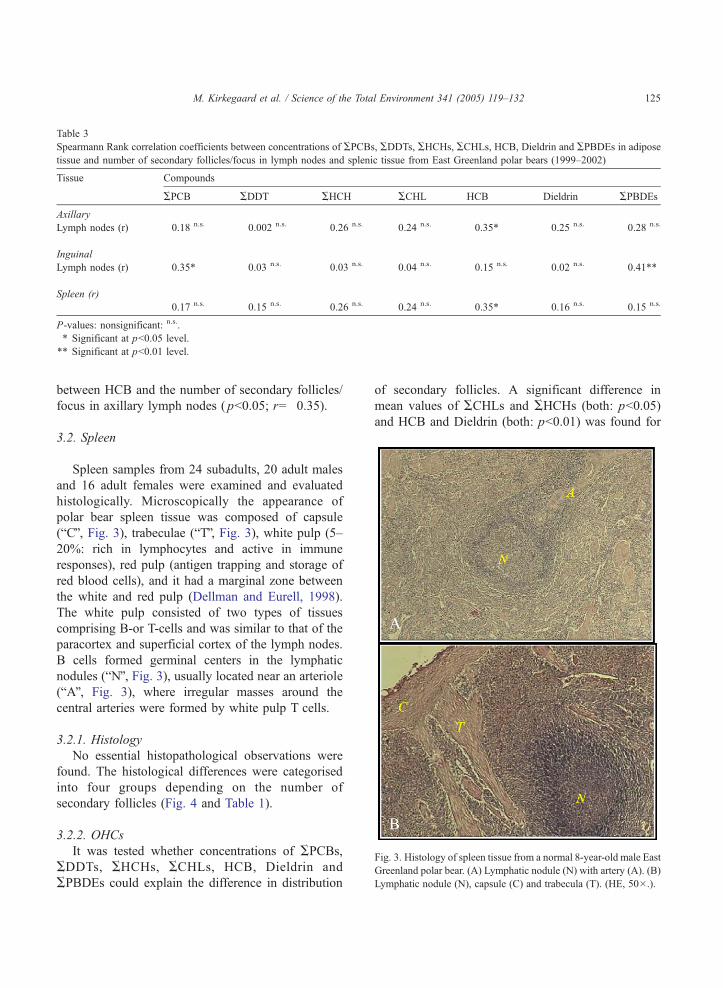
Table 3
Spearmann Rank correlation coefficients between concentrations of APCBs, ADDTs, AHCHs, ACHLs, HCB, Dieldrin and APBDEs in adipose
tissue and number of secondary follicles/focus in lymph nodes and splenic tissue from East Greenland polar bears (1999–2002)
Tissue Compounds
APCB ADDT AHCH ACHL HCB Dieldrin APBDEs
Axillary
Lymph nodes (r) �0.18 n.s. �0.002 n.s. 0.26 n.s. 0.24 n.s. �0.35* 0.25 n.s. 0.28 n.s.
Inguinal
Lymph nodes (r) �0.35* 0.03 n.s. �0.03 n.s. 0.04 n.s. 0.15 n.s. 0.02 n.s. 0.41**
Spleen (r)
0.17 n.s. �0.15 n.s. 0.26 n.s. 0.24 n.s. �0.35* �0.16 n.s. �0.15 n.s.
P-values: nonsignificant: n.s..
* Significant at pb0.05 level.
** Significant at pb0.01 level.
Fig. 3. Histology of spleen tissue from a normal 8-year-old male Eas
Greenland polar bear. (A) Lymphatic nodule (N) with artery (A). (B
Lymphatic nodule (N), capsule (C) and trabecula (T). (HE, 50�.).
M. Kirkegaard et al. / Science of the Total Environment 341 (2005) 119–132 125
between HCB and the number of secondary follicles/
focus in axillary lymph nodes ( pb0.05; r=�0.35).
3.2. Spleen
Spleen samples from 24 subadults, 20 adult males
and 16 adult females were examined and evaluated
histologically. Microscopically the appearance of
polar bear spleen tissue was composed of capsule
(bCQ, Fig. 3), trabeculae (bTQ, Fig. 3), white pulp (5–
20%: rich in lymphocytes and active in immune
responses), red pulp (antigen trapping and storage of
red blood cells), and it had a marginal zone between
the white and red pulp (Dellman and Eurell, 1998).
The white pulp consisted of two types of tissues
comprising B-or T-cells and was similar to that of the
paracortex and superficial cortex of the lymph nodes.
B cells formed germinal centers in the lymphatic
nodules (bNQ, Fig. 3), usually located near an arteriole
(bAQ, Fig. 3), where irregular masses around the
central arteries were formed by white pulp T cells.
3.2.1. Histology
No essential histopathological observations were
found. The histological differences were categorised
into four groups depending on the number of
secondary follicles (Fig. 4 and Table 1).
3.2.2. OHCs
It was tested whether concentrations of APCBs,ADDTs, AHCHs, ACHLs, HCB, Dieldrin and
APBDEs could explain the difference in distribution
of secondary follicles. A significant difference in
mean values of ACHLs and AHCHs (both: pb0.05)
and HCB and Dieldrin (both: pb0.01) was found for
t
)

Fig. 4. Histological findings in spleen tissue in the four distinguished groups. (A) Group 0 (0–1 germ centers). (B) Group 1 (2 germ centers).
(C) Group 2 (3 germ centers). (D) Group 3 (4–8 germ centers), -more germ centers are visible in the actual focus of the microscope. Arrows
indicate germ center. (HE, 50�.).
Fig. 5. Thyroid tissue from a normal 4-year-old East Greenland
male polar bear. Thyroid follicle (F) and lumen filled with colloid
(C). (HE, 50�.).
M. Kirkegaard et al. / Science of the Total Environment 341 (2005) 119–132126
all groups of follicle score (Table 2). The Tukey post
hoc test of these four contaminants showed that group
0 had significantly lower contaminant concentrations
compared to groups 1–3 ( pb0.05) (exception was
HCB, where group 0 was significantly different only
from group 3). No significant correlations were found
between APCBs, ADDTs, AHCHs, ACHLs, Dieldrinand APBDEs, and secondary follicle score; however,
a negative correlation was found between HCB and
follicle score (Table 3).
3.3. Thyroid and thymus
The thyroid gland of the polar bear was divided
into irregular lobes and each thyroid follicle (bFQ, Fig.5) comprised an epithelium composed of secretory
thyroid follicular cells secreting the stained colloid
(bCQ, Fig. 5) rich in thyroglubulin (storage form of
thyroxine) (Dellman and Eurell, 1998). The thymus
was a lympheaepithelial organ, composed of thymic
lobules (bLQ, Fig. 6). Controlled T-cell production was
located in the cortex (bCQ, Fig. 6) and maturation of
T-cells were indicated in the medulla (bMQ, Fig. 6).All the examined samples of the thyroid (n=5) and
thymus (n=11) tissue were found to be similar to
normal mammalian tissues and no histopathological
changes were detected (Figs. 5 and 6). The contam-
inant concentrations of these few animals for the
thyroid (ng/g lw) were APCBs=2341–8124 (n=5),
ADDTs=86–205 (n=5) and APBDEs=26–69 (n=3);
for the thymus, APCBs=2466–8372 (n=8),
ADDTs=101–249 (n=8) and APBDEs=25–36 (n=3).

Fig. 6. Normal thymus tissue from a 4-year-old East Greenland
female polar bear. Thymic lobules (L), medulla (M), and cortex.(C).
(HE, 50�.).
M. Kirkegaard et al. / Science of the Total Environment 341 (2005) 119–132 127
3.4. Hydroxylated (HO) metabolites
Modulation effects on endocrine systems in wild-
life may also occur as a result of PBDE and PCB
exposure and or subsequent hydroxylated (HO) PBDE
and HO-PCB, respectively, metabolite formation
(Letcher et al., 2002; Darnerud, 2003; Hakk and
Letcher, 2003; Legler and Brouwer, 2003). For
example, HO-PBDE metabolites have been shown
in vitro to competitively interact with human trans-
thyretin (TTR, thyroid hormone transport protein)
(Meerts et al., 2000). HO-PCBs have also demon-
strated thyroid-related activity in humans and wildlife,
including possible activity in Canadian polar bear
(Brouwer et al., 1998; Sandau et al., 2000b; Fisk et al.,
submitted for publication). The present East Green-
land polar bears have been found to contain among
the highest blood-plasma concentrations of HO-PCB
than that of any other species or population (Sandala
et al., 2004).
4. Discussion
Studies on IgG concentrations have indicated that
some OHCs function as immunosuppressive sub-
stances in polar bears (Bernhoft et al., 2000; Skaare
et al., 2001b; Lie et al., 2002, 2004a,in press; Fisk
et al., submitted for publication). In a normal
functioning immune system, a suppression of
immunity can lead to decreased host resistance to
infection. This could possibly be seen as increased
infection rates, expressed by high secondary follicle
counts in lymph nodes and spleen, although this has
not been proved. In a suppressed immune system,
decreased activity in the immune organs could be
expected, but this depends on the mechanism of the
xenobiotic on the immunity, and for many of the
OHCs this is not clear and suppression is dose/
exposure dependent. Furthermore, the effects of
synergy among OHCs and the lack of controlled
studies evaluating histopathological changes in
relation to OHCs in wild animals, makes the
evaluation about cause and effect problematic.
Currently, reference material on histopathological
effects of contaminants in wild animals is sparse
compared to controlled laboratory studies, and more
research on the issue is needed.
4.1. Lymph nodes
Our present results showed significant correla-
tions between the frequency of secondary follicles in
the lymph nodes and concentrations of OHCs in
adipose tissue, but as these correlations showed both
weak negative and positive relationships, OHC
exposure is not likely linked to modulation of the
frequency of secondary follicles, and other possible
multiple stress factors probably explain any modu-
lation effects. However, a modest, positive correla-
tion was found in relation to APBDE concentrations.
Evidence is accumulating that exposure to very high
concentrations of PBDEs causes immunomodulatory
effects in laboratory animals (McDonald, 2002;
Darnerud, 2003; AMAP, 2004). Exposure of mice
to BDE-47 or Bromkal 70-5DE suppressed the
proliferation of lymphocytes and the production of
antibodies (Darnerud and Thuvander, 1998;
Thuvander and Darnerud, 1999). Thymotoxic effects
occurred in mice exposed to BDE-71 (Fowles et al.,
1994). High secondary follicle counts (group 3) were
found in 20% of all lymph nodes and this
accumulation of secondary follicles indicated a
response to antigens. The significantly higher num-
ber of secondary follicles in the axillary lymph
nodes of subadults compared to adults showed a
higher immune response among the young animals,
similar to the inguinal lymph nodes. Germinal

M. Kirkegaard et al. / Science of the Total Environment 341 (2005) 119–132128
centers in secondary follicles are not present at birth
(Nielsen, 1977), so the difference was evaluated to
possibly be a natural age related immune adaptation/
maturation to antigens in subadult polar bears
(Grundmann and Vollmer, 1990).
4.2. Spleen
In 20% of the spleen samples, the frequencies of
secondary follicles were categorised as high (group
3). Again a significantly higher number of secondary
follicles in the spleen of subadults (37.5%) indicated
an activity in immune response not found in adults
(8%), as is described in humans by Stevens and
Lowe (1997). ACHLs, AHCHs, HCB and Dieldrin
showed a significant difference in mean values
among groups 0–3 in the spleen and the post hoc
analysis showed that only the group with few or
absent secondary follicles (group 0) was significantly
lower in contaminant concentrations compared to
groups 1–3. This could possibly have indicated the
immunological reaction against foreign antigens
circulating in the blood and thereby the possible
increase of the IgG response. Siebert et al. (2002)
found a relation between elevated concentrations of
DDE and spleen depletion in harbour porpoise (P.
phocoena), although these concentrations (adults
from North Sea, Baltic Sea, Norway and Island)
were much higher (range: 62–14,000 ng/g lw, n=30)
than found for all polar bears in the present study
(range: 74–1152 ng/g lw, n=92).
4.3. Thyroid and thymus
In general, no severe histopathological observa-
tions (atrophy, fibrosis and neoplasia) could be found
in either of the investigated tissues. In laboratory
studies of effects of PCB on the thyroid gland, in
utero, dietary and/or intra peritoneal (ip) exposure to
rats have shown histopathological changes like
decreases in serum T4, increased follicular cell height
and decreased colloid area (Ness et al., 1993; Saeed
and Hansen, 1997; Hood et al., 1999; Khan et al.,
2002). In Table 4, the concentrations used in the
investigations of relations between organohalogens
and thyroid changes in laboratory mammals (rats),
harbour seals, grey seals and harbour porpoise are
compared to the concentrations found in the adipose
tissue (blubber), as well as the estimated daily intake
by the East Greenland polar bears in the present
study.
According to Born and Bocher (2001), an baverageQpolar bear (weight: 200 kg) consumes app. 1000 kg
blubber per year. Based on concentrations in ringed
seal blubber from East Greenland sampled 1999–2002
(Riget, unpublished data) the average intake of a 200
kg baverageQ polar bear of APCBs is 13 Ag/kg APCBs/day as seen in Table 4. This is, of course, a rough
estimate but makes it comparable with concentrations
used in other investigations. Based on Riget et al.
(2003), the concentrations in East Greenland are a
factor 2–4 higher compared to West Greenland. These
data were used in evaluating the intake in East
Greenland polar bears (Table 4). It is seen that the
concentrations found and the daily intake calculated
are significantly lower in the bears compared to the
laboratory mammals (rats) which, as well as a low
sample size, could explain that we did not find a
relation between thyroid histology (atrophy and
fibrosis) and concentrations of PCBs.
4.4. Wildlife
Earlier studies of wildlife have shown that histo-
pathological changes in kidney, adrenals, uterus,
thyroids a.o. were related to concentrations of organo-
halogens in marine mammals such as harbour seal (P.
vitulina), grey seal (H. grypus), ringed seal (P. hispida
botnica) and harbour porpoise (P. phocoena) (Table
4) (Bergman and Olsson, 1985; Schumacher et al.,
1993; Bergman et al., 2001; Siebert et al., 2002).
Siebert et al. (2002) found a significant relation
between the degree of depletion of lymphoid organs
of harbour porpoise (P. phocoena) and elevated
concentrations of PBDEs and PCBs. In the present
study of polar bears, all thyroid and thymus samples
examined showed normal tissue and also much lower
concentrations of PBDE than that of the harbour
porpoises (P. phocoena) investigated by Siebert et al.
(2002) (Table 4).
Bernhoft et al. (2000) investigated possible
immunotoxic effects of OHCs in polar bears from
Svalbard sampled during 1991–1994, and found
blood plasma concentrations of IgG negatively
correlated with APCBs (mean ranges in different
age groups: 11200–21,700 ng/g lw, Bernhoft et al.,
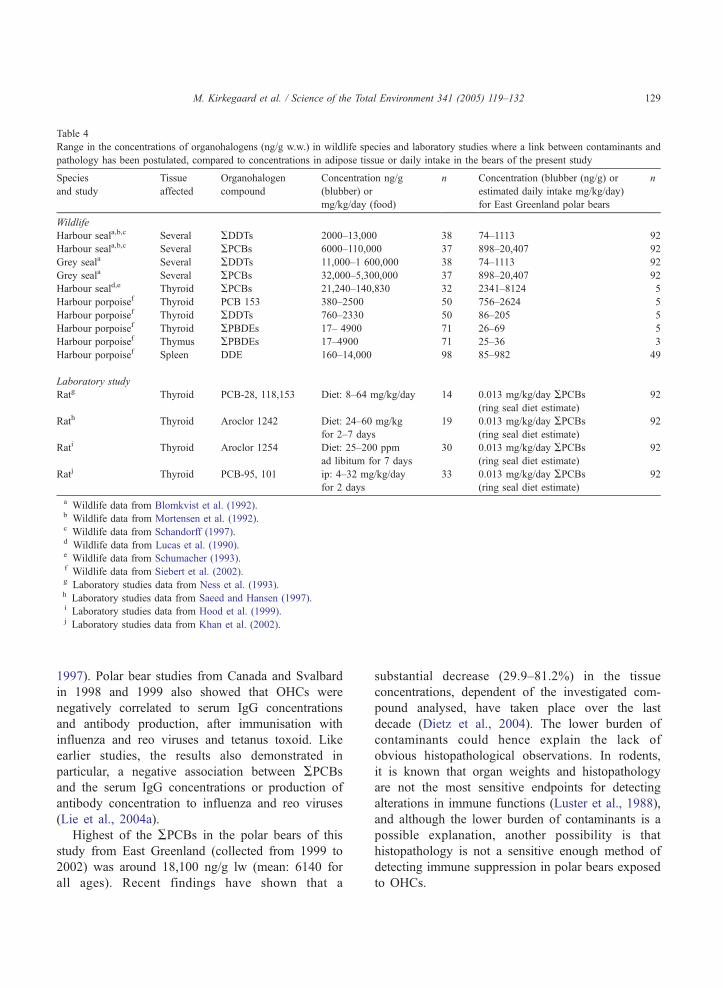
Table 4
Range in the concentrations of organohalogens (ng/g w.w.) in wildlife species and laboratory studies where a link between contaminants and
pathology has been postulated, compared to concentrations in adipose tissue or daily intake in the bears of the present study
Species
and study
Tissue
affected
Organohalogen
compound
Concentration ng/g
(blubber) or
mg/kg/day (food)
n Concentration (blubber (ng/g) or
estimated daily intake mg/kg/day)
for East Greenland polar bears
n
Wildlife
Harbour seala,b,c Several ADDTs 2000–13,000 38 74–1113 92
Harbour seala,b,c Several APCBs 6000–110,000 37 898–20,407 92
Grey seala Several ADDTs 11,000–1 600,000 38 74–1113 92
Grey seala Several APCBs 32,000–5,300,000 37 898–20,407 92
Harbour seald,e Thyroid APCBs 21,240–140,830 32 2341–8124 5
Harbour porpoisef Thyroid PCB 153 380–2500 50 756–2624 5
Harbour porpoisef Thyroid ADDTs 760–2330 50 86–205 5
Harbour porpoisef Thyroid APBDEs 17– 4900 71 26–69 5
Harbour porpoisef Thymus APBDEs 17–4900 71 25–36 3
Harbour porpoisef Spleen DDE 160–14,000 98 85–982 49
Laboratory study
Ratg Thyroid PCB-28, 118,153 Diet: 8–64 mg/kg/day 14 0.013 mg/kg/day APCBs(ring seal diet estimate)
92
Rath Thyroid Aroclor 1242 Diet: 24–60 mg/kg
for 2–7 days
19 0.013 mg/kg/day APCBs(ring seal diet estimate)
92
Rati Thyroid Aroclor 1254 Diet: 25–200 ppm
ad libitum for 7 days
30 0.013 mg/kg/day APCBs(ring seal diet estimate)
92
Ratj Thyroid PCB-95, 101 ip: 4–32 mg/kg/day
for 2 days
33 0.013 mg/kg/day APCBs(ring seal diet estimate)
92
a Wildlife data from Blomkvist et al. (1992).b Wildlife data from Mortensen et al. (1992).c Wildlife data from Schandorff (1997).d Wildlife data from Lucas et al. (1990).e Wildlife data from Schumacher (1993).f Wildlife data from Siebert et al. (2002).g Laboratory studies data from Ness et al. (1993).h Laboratory studies data from Saeed and Hansen (1997).i Laboratory studies data from Hood et al. (1999).j Laboratory studies data from Khan et al. (2002).
M. Kirkegaard et al. / Science of the Total Environment 341 (2005) 119–132 129
1997). Polar bear studies from Canada and Svalbard
in 1998 and 1999 also showed that OHCs were
negatively correlated to serum IgG concentrations
and antibody production, after immunisation with
influenza and reo viruses and tetanus toxoid. Like
earlier studies, the results also demonstrated in
particular, a negative association between APCBsand the serum IgG concentrations or production of
antibody concentration to influenza and reo viruses
(Lie et al., 2004a).
Highest of the APCBs in the polar bears of this
study from East Greenland (collected from 1999 to
2002) was around 18,100 ng/g lw (mean: 6140 for
all ages). Recent findings have shown that a
substantial decrease (29.9–81.2%) in the tissue
concentrations, dependent of the investigated com-
pound analysed, have taken place over the last
decade (Dietz et al., 2004). The lower burden of
contaminants could hence explain the lack of
obvious histopathological observations. In rodents,
it is known that organ weights and histopathology
are not the most sensitive endpoints for detecting
alterations in immune functions (Luster et al., 1988),
and although the lower burden of contaminants is a
possible explanation, another possibility is that
histopathology is not a sensitive enough method of
detecting immune suppression in polar bears exposed
to OHCs.

M. Kirkegaard et al. / Science of the Total Environment 341 (2005) 119–132130
5. Conclusion
It was investigated whether OHCs have contrib-
uted to histopathological observations in lymph
nodes, spleen, thymus and thyroid glands of the
Greenland Sea polar bear collected 1999–2002. No
essential histopathological observations were found in
any of the tissues examined. No significant relations
between concentrations of organohalogens and the
amount of secondary follicles were found in lymph
nodes although significant correlations were found for
APBDEs (positive), APCBs (negative) and HCB
(negative) and follicle score. Significantly lower
concentrations of organohalogens in spleen samples
categorized in group 0 (few/absent secondary fol-
licles) were found, which indicated a positive relation-
ship between the OHCs, ACHLs, AHCHs, HCB and
Dieldrin, and the number of secondary follicles in the
spleen. The histopathological evaluation of the few
samples of thymus and thyroid revealed no change
either. In conclusion the data suggest that the exposure
concentrations of contaminants to polar bears in East
Greenland today are unlikely to result in adverse
effects on tissues of lymph nodes, spleen, thyroid and
thymus (histopathology) although an effect, described
as increased secondary follicle count, was observed in
splenic tissue for the contaminants ACHLs, AHCHs,HCB and Dieldrin.
Acknowledgements
Danish Cooperation for Environment in the Arctic
and The Commission for Scientific Research in
Greenland are acknowledged for financial support,
Jonas Brbnlund who gathered the samples through
local hunters, Hanne Tuborg Sandell and Birger
Sandell who helped with local contacts to hunters.
Steen Andersen (Foxtrot) made the instruction video
for the polar bear hunters, for which Lars 2by also
supplied footage. The laboratory technicians at
National Water Research Institute and Great Lakes
Institute for Environmental Research (Mr. Greg Sandla
and Ms. Rodica Lazar) are acknowledged for con-
ducting the chemical analysis. The laboratory techni-
cians at the Laboratory of Pathology for conducting
the histology slides, and finally the reviewers are
acknowledged for valuable comments to this article.
References
AMAP. Amap Assessment 2002: Persistent organic pollutants in the
arctic. Oslo (Norway)7 Arctic Monitoring and Assessment
Programme (AMAP); 2004. xvi+310 pp.
Bergman A. Health condition of the Baltic grey seal (Halichoerus
grypus) during two decades. Apmis 1999;107:270–82.
Bergman A, Olsson M. Pathology of Baltic grey seal and ringed seal
females with special reference to adrenocortical hyperplasia: is
environmental pollution the cause of a widely distributed
disease syndrome? Finn Game Res 1985;44:47–62.
Bergman A, Bergstrand A, Bignert A. Renal lesions in Baltic grey
seals (Halichoerus grypus) and ringed seals (Phoca hispida
botnica). Ambio 2001;30(7):397–409.
Bergmann A, Bignert A, Helander B, Olsson M. Miljfgifta och
marina toppkonsumenter. Ostersjf ’95: 2rsrapport fr3n den
marina miljfovervakningen, Stockholms Marina Forskningscen-
trum. Sweden7 Stockholm University; 1996. p. 43–8.
Bernhoft A, Wiig O, Skaare JU. Organohalogens in polar bears
(Ursus maritimus) at Svalbard. Environ Pollut 1997;96:159–75.
Bernhoft A, Skaare JU, Wiig O, Derocher AE, Larsen HJS. Possible
immunotoxic effects of organochlorines in polar bears (Ursus
maritimus) at Svalbard. J Toxicol Environ Health A 2000;
59(7):561–74.
Blomkvist G, Roos A, Jensen S, Bignert A, Olsson M. Concen-
trations of sDDT and PCB in seals from Swedish and Scottish
waters. AMBIO 1992;21(8):539–45.
Born EW, Bfcher J. The ecology of Greenland. Nuuk7 Ministry of
Environment and Natural Resources; 2001.
Brouwer A, Morse DC, Lans MC, Schuur AG, Murk AJ, Klassen-
Wehler E, et al. Interactions of persistent environmental
organohalogens with the thyroid hormone system: mechanism
and possible consequences for animal. J Toxicol Ind Health
1998;14:59–84.
Colborn T, Vom Saal FS, Soto AM. Developmental effects of
endocrine-disrupting chemicals in wildlife and humans. Environ
Health Perspect 1993;101:378–84.
Damstra T, Barlow S, Bergman A, Kavlock R, Kraak GVD. Global
assessment of the state-of-the-science of endocrine disruptors-
WHO;2002. 180 pp.
Darnerud PO. Toxic effects of brominated flame retardants in man
and in wildlife. Environ Int 2003;29:841–51.
Darnerud PO, Thuvander A. Studies on immunological effects of
polybrominated diphenyl ethers (PBDE) and polychlorinated
biphenyls (PCB) exposure in rats and mice. Organohalog
Compd 1998;35:415–8.
Dellman HD, Eurell J, 1998. Textbook of veterinary histology, fifth
ed. Oslo (Norway)7 Lippincott Williams and Wilkins;1998. p.
128–303.
de March BGE, de Wit C, Muir DGC, Braune B, Gregor DJ,
Norstrom RJ, et al. Chapter 6: persistent organic pollutants.
AMAP assessment report: arctic pollution issues. Oslo
(Norway)7 Arctic Monitoring and Assessment Programme;
1998. p. 183–372.
Dietz R, Heide-Jbrgensen MP, H7rkfnen T, Teilmann J, Valentin N.
Age determination of European harbour seal (Phoca vitulina L).
Sarsia 1991;76:17–21.

M. Kirkegaard et al. / Science of the Total Environment 341 (2005) 119–132 131
Dietz R, Riget F, Sonne-Hansen C, Letcher R, Born EW, Muir
DCG. Polychlorinated biphenyls and organochlorine pesticides
in East Greenland polar bears (Ursus maritimus), 1990–2001.
Sci Total Environ 2004;331:107–24.
Dyce KM, Sack WO, Wensing CJG, 1995. Veterinary anatomy, 3rd
ed. Saunders;1987. p. 417–35.
Feldman EC. Hyperadrenocorticism. In: Ettinger SJ, Feldman EC,
editors. Textbook of veterinary internal medicine, vol. II.
Philadelphia7 W.B. Saunders; 1995. p. 1538–78.
Fisk AT, de Wit C, Wayland M, Kuzyk ZZ, Burgess N, Letcher R,
et al. An assessment of the toxicological significance of
anthropogenic contaminants in Canadian arctic wildlife. Sci
Total Environ;2004 [submitted for publication].
Fowles JR, Fairbrother A, Baecher-Steppan L, Kerkvliet N.
Immunologic and endocrine effects of the flame-retardant
pentabromondiphenyl ether (DE-71) in C57BL/6J mice. Tox-
icology 1994;86:49–61.
Grundmann E, Vollmer E. Cell types and functions. Reaction
patterns of the lymph nodeCurrent topics in pathology, vol. 84.
Berlin7 Springer; 1990. p. 288 part 1.
Guitart R, Puig P, Gomeezcacatalan J. Requirements for a stand-
ardidized nomenclature criterium for PCBs-computer-assisted
assignment of correct congener denomination and numbering.
Chemosphere 1993;27(8):1451–9.
Haave M, Ropstad E, Derocher AE, Lie E, Dahl E, Wiig a., et al.Polychlorinated biphenyls and reproductive hormones in female
polar bears at Svalbard. Environ Health Perspect 2003;111(4):
431–6.
Habel RE. Splanchnologia. In: Schaller Oscar, editor. Illustrated
veterinary anatomical nomenclature. Stuttgard (Germany)7 Enke
Verlag; 1992. p. 194–7.
Hakk H, Letcher RJ. Metabolism in the toxicokinetics and fate of
brominated flame retardants—a review. Environ Int 2003;29:
829–39.
Henriksen EO, Wiig O, Skaare JU, Gabrielsen GW, Derocher AE.
Monitoring PCBs in polar bears: lessons learned from Svalbard.
J Environ Monit 2001;3:493–8.
Hensel RJ, Sorensen FE. Age determination of live polar bears. Int
Conf Bear Res Manage 1980;4:95–100.
Hood A, Hashmi R, Klaassen CD. Effects of microsomal enzyme
inducers on the thyroid-follicular cell proliferation, hyper-
plasia, and hypertrophy. Toxicol Appl Pharmacol 1999;160(2):
163–70.
Khan MA, Lichtensteiger CA, Faroon O, Mumtaz M, Schaeffer DJ,
Hansen LG, 2002. Toxicol Sci 2002;65(1):52–61.
Koponen K, Myers MS, Ritola O, Huuskonen SE, Seppa-Lindstrom
P. Histopathology of feral fish from a PCB-contaminated
freshwater lake. Ambio 2001;30(3):122–6.
Legler J, Brouwer A. Are brominated flame retardants endocrine
disruptors? Environ Int 2003;29:879–85.
Letcher RJ, Nordstrom RJ, Lin S, Ramsay MA, Bandiera SM.
Immunoquantitation and microsomal monooxygenase activities
of hepatic cytochromes P450 1A and P450 2B and chlorinated
hydrocarbon contaminant levels in polar bear (Ursus mariti-
mus). Toxicol Appl Pharmacol 1996;137:127–40.
Letcher RJ, Lemmen JG, van der Burg B, Brouwer A, Bergman A,
Giesy JP, et al. In vitro antiestrogenic effects of aryl methyl
sulfone metabolites of polychlorinated biphenyls and 2,2-bis(4-
chlorophenyl)-1,1-dichloroethene on 17 beta-estradiol-induced
gene expression in several bioassay systems. Toxicol Sci 2002;
69(2):362–72.
Lie E, Larsen HJS, Derocher AE, Lunn N, Norstrom R,Wiig O, et al.
Do POPs impair the infection resistance of polar bears (Ursus
maritimus). AMAP conference abstract; 2002.
Lie E, Jbrgen H, Larsen S, Johansen GM, Derocher AE, Lunn NJ,
et al. Does high organochlorine (OC) exposure impair the
resistance to infection in polar bears (Ursus maritimus)?: Part I
Effect of OCs on the humoral immunity. J Toxicol Environ
Health 2004a;67:555–82.
Lie E, Jbrgen H, Larsen S, Johansen GM, Derocher AE, Lunn NJ,
et al. Does high organochlorine (OC) exposure impair the
resistance to infection in polar bears (Ursus maritimus)?: Part I
Effect of OCs on mitogen and antigen induced lymphocyte
profileration. J Toxicol Environ Health 2004b [in press].
Lucas B, Vetler W, Fischer P, Heidemann G, Plotz J. Characteristic
chlorinated-hydrocarbon patterns in the blubber of seals from
different marine regions. Chemosphere 1990;21(1–2):13–9.
Luross JM, Alaee M, Sergeant DB, Cannon CM, Whittle DM,
Solomon KR, et al. Spatial distribution of polybrominated
diphenyl ethers and polybrominated biphenyls in lake trout from
the Laurentian Great Lakes. Chemosphere 2002;46:665–72.
Luster MI, Munson AE, Thomas PT, Holsapple MP, Fenters JD,
White Jr KL, et al. Development of a testing battery to assess
chemical-induced immunotoxicity: National Toxicology Pro-
gram’s guidelines for immunotoxicity evaluation in mice.
Fundam Appl Toxicol 1988;10:2–19.
McDonald TA. A perspective on the potential health risks of
PBDEs. Chemosphere 2002;46:745–55.
Meerts IATM, van Zanden JJ, Luijks EA, van Leeuwen-Bol I,
Marsh G, Jakobsson E, et al. Potent competitive interactions of
some brominated flame retardants and related compounds with
human transthyretin in vitro. Toxicol Sci 2000;56:95–104.
Mortensen P, Bergmann A, Bignert A, Hansen H, Harkonen T,
Olsson M. Prevalence of skull lesions in harbour seals (Phoca
vitulina) in Swedish and Danish museum collections 1835–
1988. AMBIO 1992;21(8):520–4.
Muir, DCG, Dietz, R, Riget, FF, Sonne, C, Letcher, RJ, Born, EW,
Polybrominated diphenylethers in East Greenland polar bears
(Ursus maritimus) 1990–2001. [in preparation].
Ness DK, Schantz SL, Moshtaghian J, Hansen LG. Effects of
perinatal exposure to specific PCB congeners on the thyroid-
hormone concentrations and thyroid histology in rat. Toxicol
Lett 1993;68(3):311–23.
Nielsen EH, 1977. Kompendium i speciel histologi, 3.ed. Denmark7
F.A.D.L.s Forlag; 1977. p. 11–162.
Norstrom RJ, Belikov SE, Born EW, Garner GW, Malone B,
Olpinski S, et al. Chlorinated hydrocarbon contaminants in polar
bears from eastern Russia, North America, Greenland, and
Svalbard: biomonitoring of arctic pollution. Arch Environ
Contam Toxicol 1998;35:354–67.
Oskam IC, Ropstad E, Dahl E, Lie E, Derocher AE, Wiig a, et al.Organochlorines affect the major androgenic hormone, testos-
terone, in male polar bears (Ursus maritimus) at Svalbard. J
Toxicol Environ Health A 2003;66(22):2119–39.

M. Kirkegaard et al. / Science of the Total Environment 341 (2005) 119–132132
Polischuk S, Ramsay M, Norstrom RJ. Body burdens and tissue
concentrations of organohalogens in polar bears (Ursus mariti-
mus) vary during seasonal fasts. Environ Pollut 2002;118:29–39.
Riget FF, Johansen P, Glasius M, Vorkamp K, Dahlgaard H, Muir D,
et al. Chapter 4 Marine environment. In: Riget FF, Christensen J,
Johansen P, editors. AMAP Greenland and the Faroe Islands
1997–2001. Denmark7 Ministry of Environment; 2003.
Riget, FF. Senior scientist, National Environmental Research
Institute, PO Box 358, Frederiksborgvej 399, DK-4000 Ros-
kilde, Denmark; Unpublished data. Ph: (+45) 46301948; e-mail:
Ross PS, de Swart RL, Reijnders PJH, Van Loveren H, Vos JGV,
Osterhaus ADME. Contaminant related suppression of delayed-
type hypersensitivity and antibody response in harbour seals fed
herring from the Baltic Sea. Environ Health Perspect 1995;103:
162–7.
Ross PS, de Swart RL, Timmermann HH, Reijnders PJH, Vos JGV,
Van Loveren H, et al. Suppression of natural killer cell activity
in harbour seals (Phoca vitulina) fed Baltic Sea herring. Aquat
Toxicol 1996;34:71–84.
Saeed A, Hansen LG. Morphometric changes in the prepubertal
femal rat thyroid gland following acute exposure to 2,2V44V-tetrachlorobiphenyl and aroclor 1242. J Toxicol Environ Health
1997;51(5):503–13.
Sandala GM, Sonne-Hansen C, Dietz R, Muir DCG, Valters K,
Bennett ER, et al. Methyl sulfone and hydroxylated PCB
metabolites in adipose and whole blood of polar bear (Ursus
maritimus) from Scoresby Sound, Greenland. Sci Total Environ
2004;331:125–41.
Sandau CD, Meerts IATM, Letcher RJ, McAlees AJ, Chittim B,
Brouwer A, et al. Identification of 4-hydroxyheptachlorostyrene
in polar bear plasma and its binding affinity to transthyretin: a
metabolite of octachlorostyrene? Environ Sci Technol 2000;34:
3871–7.
Schandorff S. Developmental stability and skull lesions in the
harbour seal (Phoca vitulina) in the 19th and 20th centuries. Ann
Zool Fennici 1997;34:151–66.
Schumacher U, Zahler S, Horny HP, Heidemann G, Skirnisson K,
Welsch U. Histological investigations on the thyroids of marine
mammals (Phoca vitulina , Phocoena phocoena) and the
possible implications of marine pollution. J Wildl Dis 1993;
29(1):103–8.
Siebert U, Vossen A, Baumg7rtner W, Mqller G, Beineke A,
McLachlan M, et al. Investigations of the influence of Pollutants
on the endocrinium and immune system of harbour porpoise in
the German North and Baltic Seas. UBA F+E Vorhaben FKZ 2
99 65 211/01 2002:1–308.
Skaare JU, Bernhoft A,Wiiga., NorumKR, Haug E, Eide DM, et al.
Relationship between plasma levels of organochlorines, retinol
and thyroid hormones from polar bears (Ursus maritimus) at
Svalbard. J Toxicol Environ Health A 2001;62:227–41.
Skaare JU, Hansen HJ, Lie E, Ropstad E, Bernhoft A, Derocher AE,
et al. Relationships between high loads of organic contaminants
and health effects in arctic mammals—immune response and
chlorinated environmental pollutants in polar bear. Pharmacol
Toxicol 2001;88(Suppl. 1):14.
Stevens A, Lowe J, 1994. Human histology, 2nd ed. New
York7 Mosby;1997. p. 117–258.
Swart RD, Ross PS, Vedder LJ, Timmerman HH, Heisterkamp S,
Loveren HV, et al. Impairment of immune function in harbour
seals (Phoca vitulina) feeding on fish from polluted waters.
Ambio 1994;23:155–9.
Thuvander A, Darnerud PO. Effects of polybrominated diphenyl
ether (PBDE) and polychlorinated biphenyl (PCB) on some
immunological parameters after oral exposure in rats and mice.
Toxicol Environ Chem 1999;70:229–42.
Tryphonas H. Immunotoxicity of polychlorinated biphenyls: present
status and further considerations. Exp Clin Immunogenet 1994;
11:149–62.
Vos JG, Luster MI. Immune alterations. In: Kimborough RD, Jensen
S, editors. Halogenated biphenyls, terphenyls, naphthalenes,
dibenzodioxins and related products. New York7 Elseviere
Science Publishers; 1989. p. 295–322.
Welsch U, Schwertfirm S, Skirnisson K, Shumacher U. Histolog-
ical, histochemical and fine structural observations on the lymph
node of the common seal (Phoca vitulina) and the grey seal
(Halichoerus grypus). Anat Rec 1997;247:225–42.
Wiig a, Derocher AE, Cronin MM, Skaare JU. Female pseudo-
hermaphrodite polar bears at Svalbard. J Wildl Dis 1998;34(4):
792–6.
Wong S, Fournier M, Coderre D, Banska W, Krzystyniak K.
Environmental immunotoxicology. In: Peakall DB, editor.
Animal biomarkers as pollution indicators. London7 Chapman
and Hall; 1992. p. 8–189.




















