
Thyroid hormones and deiodinase activities in plasma and tissues from East Greenland polar bears...
12
ORIGINAL PAPER Thyroid hormones and deiodinase activities in plasma and tissues from East Greenland polar bears (Ursus maritimus) during winter season Kristin Møller Gabrielsen 1 • Julie Stene Krokstad 1 • Maria-Jesus Obregon 2 • Gro Dehli Villanger 1,4 • Christian Sonne 3 • Rune Dietz 3 • Bjørn Munro Jenssen 1 Received: 30 June 2014 / Revised: 6 April 2015 / Accepted: 6 April 2015 Ó Springer-Verlag Berlin Heidelberg 2015 Abstract Thyroid hormones (THs) are essential for metabolism and thermoregulation in arctic animals. Still, there is a lack of deeper basic knowledge regarding the regulation and functioning of THs in the environmental physiology of polar bears (Ursus maritimus). This is nec- essary in order to understand the true impact and conse- quences of the combination of stressors such as depletion of sea ice and endocrine-disrupting contaminants for the polar bear species. As a first step to gain insight into TH physiology in polar bears, TH concentrations in liver, kidney and muscle in East Greenland polar bears sampled February–March 2011 were analysed and their associations with circulating levels of THs were investigated. In addi- tion, type 1 deiodinase (D1) activities in liver, kidney and muscle and type 2 deiodinase (D2) activities in muscle were analysed. Concentrations of 3,5,3 0 ,5 0 -tetraiodothy- ronine (T4) were highest in plasma, followed by liver, kidney and muscle, whereas concentrations of 3,5,3 0 -tri- iodothyronine (T3) were highest in kidney followed by liver, plasma and muscle. D1 activities in the tissues varied in the order liver [ kidney muscle, while D2 activity was only analysed in muscle. There were significant posi- tive relationships between T4 in plasma and liver as well as between T4 in plasma and D1 activity in liver. This implies that liver is an important non-thyroidal organ for deiodi- nation of T4, thus providing T3 to the plasma pool, in polar bears. The polar bears with the lowest body condition also had the lowest concentrations of free T3 in plasma and D2 activities in muscle. Keywords Polar bear Thyroid hormones Deiodinase activity Tissue levels East Greenland Introduction The polar bear (Ursus maritimus) experiences many of the most extreme conditions mammals can be subjected to. Unlike brown bears (U. arctos) and black bears (U. amer- icanus), polar bears do not enter the dormant state referred to as hibernation or winter sleep during the winter season (Stirling 2002; Robbins et al. 2012). The exception is the pregnant female, which enters her den in the fall and stay there until spring, whereas other females and males actively search for food during the winter (Stirling 2002). The long, dark arctic winters with temperatures below -40 °C require adequate thermoregulatory adaptations (Scholander et al. 1950a, b; Øritsland 1970; Best 1982). During the summer periods, ice-free areas result in several months of sparse and unpredictable availability of seals as the major prey, re- sulting in long periods of fasting as a consequence (De- rocher et al. 1990; Stirling 2002). To conserve energy and heat in the Arctic during periods of low temperatures and/or food shortage, the polar bear have evolved fine-tuned bioenergetic adaptations (Welch et al. 2014). The polar bear & Kristin Møller Gabrielsen [email protected] & Bjørn Munro Jenssen [email protected] 1 Department of Biology, Norwegian University of Science and Technology, 7491 Trondheim, Norway 2 Instituto de Investigaciones Biomedicas, Centro Mixto from CSIC-UAM, 28029 Madrid, Spain 3 Department of Bioscience, Arctic Research Centre, Aarhus University, P.O. Box 358, 4000 Roskilde, Denmark 4 Department of Child Development and Mental Health, Norwegian Institute of Public Health, P.O. Box 4404, 0473 Nydalen, Oslo, Norway 123 Polar Biol DOI 10.1007/s00300-015-1694-z
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Thyroid hormones and deiodinase activities in plasma and tissues from East Greenland polar bears...

ORIGINAL PAPER
Thyroid hormones and deiodinase activities in plasma and tissuesfrom East Greenland polar bears (Ursus maritimus) during winterseason
Kristin Møller Gabrielsen1• Julie Stene Krokstad1
• Maria-Jesus Obregon2•
Gro Dehli Villanger1,4• Christian Sonne3
• Rune Dietz3• Bjørn Munro Jenssen1
Received: 30 June 2014 / Revised: 6 April 2015 / Accepted: 6 April 2015
� Springer-Verlag Berlin Heidelberg 2015
Abstract Thyroid hormones (THs) are essential for
metabolism and thermoregulation in arctic animals. Still,
there is a lack of deeper basic knowledge regarding the
regulation and functioning of THs in the environmental
physiology of polar bears (Ursus maritimus). This is nec-
essary in order to understand the true impact and conse-
quences of the combination of stressors such as depletion
of sea ice and endocrine-disrupting contaminants for the
polar bear species. As a first step to gain insight into TH
physiology in polar bears, TH concentrations in liver,
kidney and muscle in East Greenland polar bears sampled
February–March 2011 were analysed and their associations
with circulating levels of THs were investigated. In addi-
tion, type 1 deiodinase (D1) activities in liver, kidney and
muscle and type 2 deiodinase (D2) activities in muscle
were analysed. Concentrations of 3,5,30,50-tetraiodothy-ronine (T4) were highest in plasma, followed by liver,
kidney and muscle, whereas concentrations of 3,5,30-tri-iodothyronine (T3) were highest in kidney followed by
liver, plasma and muscle. D1 activities in the tissues varied
in the order liver[ kidney � muscle, while D2 activity
was only analysed in muscle. There were significant posi-
tive relationships between T4 in plasma and liver as well as
between T4 in plasma and D1 activity in liver. This implies
that liver is an important non-thyroidal organ for deiodi-
nation of T4, thus providing T3 to the plasma pool, in polar
bears. The polar bears with the lowest body condition also
had the lowest concentrations of free T3 in plasma and D2
activities in muscle.
Keywords Polar bear � Thyroid hormones � Deiodinaseactivity � Tissue levels � East Greenland
Introduction
The polar bear (Ursus maritimus) experiences many of the
most extreme conditions mammals can be subjected to.
Unlike brown bears (U. arctos) and black bears (U. amer-
icanus), polar bears do not enter the dormant state referred
to as hibernation or winter sleep during the winter season
(Stirling 2002; Robbins et al. 2012). The exception is the
pregnant female, which enters her den in the fall and stay
there until spring, whereas other females and males actively
search for food during the winter (Stirling 2002). The long,
dark arctic winters with temperatures below-40 �C require
adequate thermoregulatory adaptations (Scholander et al.
1950a, b; Øritsland 1970; Best 1982). During the summer
periods, ice-free areas result in several months of sparse and
unpredictable availability of seals as the major prey, re-
sulting in long periods of fasting as a consequence (De-
rocher et al. 1990; Stirling 2002). To conserve energy and
heat in the Arctic during periods of low temperatures and/or
food shortage, the polar bear have evolved fine-tuned
bioenergetic adaptations (Welch et al. 2014). The polar bear
& Kristin Møller Gabrielsen
& Bjørn Munro Jenssen
1 Department of Biology, Norwegian University of Science
and Technology, 7491 Trondheim, Norway
2 Instituto de Investigaciones Biomedicas, Centro Mixto from
CSIC-UAM, 28029 Madrid, Spain
3 Department of Bioscience, Arctic Research Centre, Aarhus
University, P.O. Box 358, 4000 Roskilde, Denmark
4 Department of Child Development and Mental Health,
Norwegian Institute of Public Health, P.O. Box 4404,
0473 Nydalen, Oslo, Norway
123
Polar Biol
DOI 10.1007/s00300-015-1694-z

is believed to be able to switch between a fasting and a
feeding metabolism, often referred to as walking hiberna-
tion (Derocher et al. 1990). The energy cost during these
periods of fasting is, however, still larger than for hiber-
nating bears, indicating a high net cost of staying active
during cold winters and ice-free conditions (Robbins et al.
2012). Recent evidence suggests changes in annual patterns
of sea ice distribution and subsequent loss of sea ice, the
primary habitat of polar bears, and thus decreased avail-
ability of seals, due to climate warming (Stirling and De-
rocher 2012). For the polar bears, this will inevitably mean
longer periods of fasting and a consequently lower body
condition.
Thyroid hormones (THs) are essential in numerous
physiological processes in mammals, including metabolism
and thermoregulation (McNabb 1992; Hulbert 2000; Silva
2006). Basal metabolic rate (BMR) and daily energy ex-
penditure in humans (Homo sapiens) have both been linked
to THs (Hulbert 2000). Studies have also shown that
hibernation or winter sleep is associated with decreased
plasma concentrations of THs in American black bears and
European brown bears (Azizi et al. 1979; Hissa et al. 1994;
Tomasi et al. 1998). Furthermore, reduced levels of THs
have been reported in fasting mammals, such as the Arctic
fox (Vulpes lagopus) (Fuglei et al. 2000; Nieminen et al.
2004). The lowered concentration of circulating THs dur-
ing hibernation and fasting is thought to conserve energy
due to a decrease in metabolic rate and increased protein
conservation (Carter et al. 1975; Cox et al. 1984; Hulbert
2000). The THs are highly conserved among vertebrates,
and their production is regulated via a negative feedback
system controlled by the hypothalamus–pituitary–thyroid
(HTP) axis (Zoeller et al. 2007). The thyroid gland syn-
thesises, stores and secretes both 3,5,30,50-tetraiodothy-ronine (thyroxine [T4]) and 3,5,30-triiodothyronine (T3)
(Hulbert 2000; Zoeller et al. 2007). Since T3 has the
highest affinity towards the nuclear TH receptors (TRs), it
is regarded as the active hormone (Cheng et al. 2010).
However, TH action can also be mediated through non-
genomic mechanisms.
Approximately 80 % of the circulating T3 is produced
by deiodination enzymes in extrathyroidal tissues, although
the percentage varies among species and physiological
conditions (Kohrle 2000; Orozco et al. 2012). The
deiodination of T4 to T3 occurs via outer (50) ring
deiodination performed by type 1 (D1) and type 2 (D2)
deiodinases (Kohrle 2000; Orozco et al. 2012). In mam-
mals, liver, kidney and thyroid gland are the primary tis-
sues where D1 is expressed and has high activity, while D2
functions in, e.g., the brain, skeletal muscle, brown adipose
tissue (BAT) and heart. Both D1 and D2 can inactivate T4
or T3 by producing, e.g., reverse T3 (rT3) or other
iodothyronines, although the major inactivator of T3 is type
3 (D3) deiodinase (Bianco et al. 2002). D1 activity in all
tissues, except for in the thyroid gland, is increased by
excessive levels of THs (hyperthyroidism) and decreased
by insufficient levels of THs (hypothyroidism), while the
opposite is true for D2 activity (Kohrle 1999). Altogether,
the deiodinases are very important for regulating plasma
and intracellular levels of THs and especially levels of T3,
thus subsequently regulating the biological action of THs
(Kohrle 1999, 2000; Bianco and Kim 2006).
In plasma, T4 and T3 are transported bound to transport
proteins and\0.01 % of T4 and 0.1 % of T3 exist as free
hormones (McNabb 1992; Zoeller et al. 2007). Transport of
THs from plasma into tissues occurs through carrier-me-
diated membrane transport systems (Hennemann et al.
2001). The transport and uptake of THs as well as available
intracellular binding sites all contribute to regulate intra-
cellular levels of THs together with deiodinases and other
enzymes involved in TH metabolism (Kohrle 1999; Hul-
bert 2000; Hennemann et al. 2001). The deiodinase en-
zymes, involved in activating and deactivating THs, have
the last few years received renewed scientific attention
(Bianco et al. 2002; Gereben et al. 2008; Arrojo e Drigo
et al. 2013; Dentice et al. 2013). This is due to their ability
to regulate tissue levels of THs in response to tissue-
specific needs, without necessarily altering circulating
levels.
Concentrations of THs in vertebrates will vary with life
history events such as development, growth and repro-
duction (McNabb 1992; Hulbert 2000; Santisteban and
Bernal 2005) and seasonally with annual events such as
moulting, hibernation and periods of low food availability
(Hissa et al. 1994; Boily 1996; Tomasi et al. 1998; Ebling
and Barrett 2008). The diversity of adaptations evolved for
life strategies in wildlife inhabiting extreme environments
gives reasons to believe that TH responses, functioning and
regulation might differ between species. In pinnipeds,
where the TH system has been widely studied, TH re-
sponses to fasting, in addition to varying widely with sex,
life history stage and breeding status as well as individual
needs for energy expenditure (Crocker et al. 2012; Kelso
et al. 2012), also show atypical response to fasting during
development. Martinez et al. (2013) described an atypical
TH response to fasting in fasting-adapted elephant seal
pups (Mirounga angustirostris) as compared to, e.g., hu-
mans and rodents. There are, thus, reasons to believe that
there is a lack of knowledge regarding the functioning and
regulation of the TH system in wildlife adapted to extreme
environmental conditions. Despite the suggested involve-
ment of THs in explaining the seasonal rhythms of food
intake and energy expenditure in mammals experiencing
alternating external environments (Ebling and Barrett
2008), the knowledge concerning the role of THs in arctic
mammals, such as the polar bear, is scarce.
Polar Biol
123

Although levels of T4 and T3 have been reported in
polar bears previously, these measurements have been
constrained to circulating levels (Leatherland and Ronald
1981; Cattet 2000; Skaare et al. 2001; Braathen et al. 2004;
Villanger et al. 2011a; Bytingsvik 2012). However, circu-
lating levels of THs are thought not to reflect intracellular
TH status and do therefore not give information about TH
status in tissues (Bianco 2013; Dentice et al. 2013). Levels
of THs and deiodinase activities in various tissues have
been reported in studies using experimental animals
(McCann et al. 1984; Morreale de Escobar et al. 1985; van
Doorn et al. 1985; Raasmaja et al. 1996; Viluksela et al.
2004; Kunisue et al. 2011; Lavado-Autric et al. 2013), but
there are to our knowledge no reports of levels of THs or
deiodinase activities in tissues of free-ranging polar bears
or other arctic wildlife species.
The polar bear species is facing numerous threats. In
addition to loss of suitable habitat for obtaining prey and
reproduction due to climate warming, polar bears also
carry high amounts of persistent organic pollutants (POPs)
(Letcher et al. 2010; McKinney et al. 2011; Stirling and
Derocher 2012; Jenssen et al. 2015). Several health effects
have been associated with high levels of these anthro-
pogenic contaminants in polar bears, including effects on
the TH concentrations (Sonne 2010; Villanger et al.
2011a). Lipophilic POPs accumulate in lipid-rich tissues
such as adipose tissue, and mobilisation of lipid stores
during periods of fasting can result in redistribution of
POPs from adipose tissue to blood and other organs that
are targets of POP-mediated toxicity (Lydersen et al.
2002; Polischuk et al. 2002). There does, however, seem
to be a lack of deeper basic knowledge regarding the
regulation and functioning of THs in the environmental
physiology of polar bears. This is necessary in order to
understand the true impact and consequences of the
combination of stressors climate warming and con-
taminant threats for the polar bear species. Thus, as a first
step to gain insight into TH physiology in polar bears, we
analysed levels of T4 and T3 in liver, kidney and muscle
in polar bears from East Greenland sampled during winter
conditions and investigated their associations with circu-
lating levels of THs measured in plasma. In addition, D1
activities in liver, kidney and muscle and D2 activity in
muscle were analysed, and associations between levels of
THs and D1 and D2 activities were investigated.
Depending on the polar bears hunting successes, the polar
bears may have experienced long periods of food depri-
vation during winter. Body condition is an important
measure for individual animal health and nutritional sta-
tus, so associations between body condition of the polar
bears and the thyroid hormone levels and deiodinase ac-
tivities were therefore also assessed.
Materials and methods
Sampling
The polar bears (N = 7) investigated in the present study
were legally harvested by local Inuit hunters in February to
March 2011(*4 weeks) as part of the annual subsistence
quota of 35 polar bears in the Scoresbysund (Ittoqqor-
toormiit) area, 70�290N 21�580W, East Greenland (Fig. 1).
The map of the sampling area was produced using ArcGIS
(ArcGIS 10.1, ERSI, Redlands, CA, USA). The two males
and five females were 2–4 years of age and defined as
sexually immature and hence classified as sub-adults
(Rosing-Asvid et al. 2002). The present study was part of
the AURORAE programme led by Prof. DSc Rune Dietz at
Aarhus University.
The animals were as quickly as possible subjected to
post-mortem necropsy and sampling (\1–3 h after death).
Sex, standard length, zoological length and circumference
were noted. Blood samples were collected primarily from
the heart using a 50-mL syringe, transferred into Vacuet-
te� heparinised tubes and then centrifuged in order to
collect plasma. The plasma samples were kept frozen at
-10 to -20 �C during the field period (*4 weeks). Tissue
samples of liver, kidney and muscle were sampled and then
stored in two different manners. Samples of 20–40 g were
taken and stored frozen at -10 to -20 �C during the field
period (*4 weeks) and then transferred to and stored at
-20 �C. Plasma and these tissue samples were shipped
frozen and stored at -20 �C until analysis. Additional
tissue samples (2–5 g) were snap-frozen during the field
period and subsequently stored and shipped in liquid ni-
trogen (-196 �C). These samples were then transferred to
and stored at -70 �C until analysis. The nitrogen-stored
tissues were the preferred tissues for analysing levels of TH
and deiodinase activity in tissues. However, snap-frozen
samples were not available for all specimens, and there-
fore, analyses of regular frozen tissue from liver/kidney
and muscle were conducted on two and three specimens,
respectively.
Analysis of thyroid hormones in plasma
Analyses of total T3 and T4 (i.e. free and protein-bound)
and free T4 and T3 in plasma (T4P, fT4P, T3P, fT3P, re-
spectively) were conducted at the Norwegian University of
Science and Technology, Trondheim, Norway, using
commercially available solid-phase 125I radioimmune as-
say (RIA) kits (Coat-A-Count Free T3; Coat-A-Count Free
T4; Coat-A-Count Total T3; Coat-A-Count Total T4, Sie-
mens Medical Solutions, CA, USA) without modifications.
Although developed for human use, commercial kits have
Polar Biol
123
been validated to be applicable to polar bear (Braathen
et al. 2004; Bytingsvik 2012) as well as other marine
mammals’ plasma (Gabrielsen et al. 2011; Villanger et al.
2011b) by running a parallelism assay of diluted samples
with the standards curve. This is due to the fact that THs
are highly conserved among vertebrate species (Zoeller
et al. 2007). The procedure was similar to that reported for
TH analysis by Gabrielsen et al. (2011). The bound ra-
dioactive antigen was quantified using a Cobra Auto-
Gamma gamma counter (Packard Instruments Company,
Dowers Grove, IL, USA). Calibration curves from analysed
calibrators were generated from the gamma counter soft-
ware (Spectra Works Spectrum Analysis Software,
Meriden, USA) and used to calculate the TH concentra-
tions in the samples. The samples were analysed in du-
plicates (T3P and fT3P) and triplicates (T4P and fT4P), and
the means were reported. Instrumental detection limits
were given for each RIA run and were 0.016 and
2.048 nmol/L for T3P and T4P, respectively, and 0.012 and
0.425 pmol/L for fT3P and fT4P, respectively. All samples
were above detection limits.
The coefficients of variation (CVs) were acceptable
(B15 %) except for two samples with CV values of 51.5 and
51.7 %. As the intra-assay control parameters were all ac-
ceptable, the high variation in these two samples was most
likely caused by the very low hormone concentrations. These
Fig. 1 Map showing the
location of the study area where
the polar bears (Ursus
maritimus) included in the study
(n = 7) were captured
Polar Biol
123

CV values were nevertheless accepted as the fT3 levels were
within physiological fT3 values previously reported in polar
bears (Braathen et al. 2004). All hormone assays were single
runs except for fT3, where results from two runs were used.
Standard reference material (SRM, Lypohchek� Im-
munoassay Plus Control, Levels 2, BIO-RAD, CA, USA),
bovine plasma and/or repeated measures of individual
samples were used to test intra- and interspecific variation.
Intra-assay variation ranged from 2.2 to 13.2 %, while inter-
assay variation ranged from 3.8 to 14.1 % (fT3 only). TH
concentrations in plasma were converted to per gram (1 mL
plasma = 1.025 g plasma) and reported in pmol/g (total
hormone) and fmol/g (free hormone) in order to compare
levels across the investigated tissues.
Analysis of thyroid hormones and deiodinase
activity in liver, kidney and muscle
Analyses of TH concentrations and D1 and D2 activities in
liver, kidney and muscle were performed at Instituto de
Investigaciones Biomedicas (IIBM), Madrid, Spain. The
high specific activity ([3000 mCi/mg) labelled iodothy-
ronines ([125I]-T4, [131I]-T4, [125I]-T3, [125I]-rT3) used in
the analyses were synthesised at the laboratory using
chloramine-T and T3, 3,5-T2 and 30,3-T2 as substrates,
respectively, as described by Weeke and Orskov (1973).
[125I]-T3 and [125I]-T4 were used for the determination of
TH concentrations in RIAs, while [131I]-T4 and [125I]-T3
were used as recovery tracers during the extraction. Fur-
ther, [125I]-rT3 was used as substrate for the determination
of D1 and D2 activities.
Thyroid hormone concentrations in liver, muscle
and kidney
Total T4 and T3 in liver (T4L, T3L), kidney (T4K, T3K) and
muscle (T4M, T3M) samples were analysed by RIAs after
extraction and purification of the tissue samples, as
originally described in Morreale de Escobar et al. (1985)
and later in Lavado-Autric et al. (2013). In short, frozen
tissue samples were homogenised in methanol, and tracer
amounts of [131I]-T4 and [125I]-T3 were added to each
sample. The extraction of the iodothyronines was done
twice using chloroform–methanol (2:1) before back-ex-
traction into an aqueous phase (Cl2Ca at 0.05 %). Bio-Rad
AG1x2 resin columns (Bio-Rad Laboratories, Hercules,
CA) were then used to purify the aqueous extracts. The
acetic extracts obtained in the last elution were evaporated
to dryness and dissolved in RIA buffer. Recovery of
[131I]-T4 and [125I]-T3 was determined in each sample.
Aliquots of the samples were then analysed by sensitive
RIAs to determine T4 and T3 concentrations. Limits of
sensitivity were 2.5 pg T4 and 0.75 pg T3 per tube.
Samples were analysed in duplicate. Calculations of T4 and
T3 concentrations were done using the amount of hor-
mones detected by RIAs, the individual recovery of the
added tracers and the weight of the extracted tissue sample.
Corrections for the amounts of THs in the blood trapped in
the tissues were not performed. Results were given as pg/g
and recalculated to pmol/g.
Determination of iodothyronine deiodinase activities
in liver, kidney and muscle tissues
The analyses of D1 and D2 activities were performed as
described in Lavado-Autric et al. (2013). Tissue ho-
mogenates were prepared in a buffer [1:30 wt/vol] con-
sisting of 0.32 M sucrose, 10 mm HEPES buffer and
dithiothreitol (DTT; 1 mM for D1 and 10 mM for D2), pH
7.0. The DTT was added on the day of the assay. All
samples were analysed in duplicate. Before the assays were
carried out, the [125I]-rT3 and/or [125I]-T4 were purified to
separate the labelled hormones from the contaminating
iodide, so that blanks were\1 % of the total radioactivity
used. This was done using paper electrophoresis on a
Whatman 3 MM paper strip in ammonium acetate (50 mM,
pH 6.8) at 500 V.
For analyses of D1 activity in liver (D1L activity) and
kidney (D1K activity), tissue homogenates were diluted
1:400 and 20–30 lg protein per tube was used for analysis.
[125I]-rT3 (100.000 cpm/tube) plus 400 nM rT3 were used
as the substrate, and DTT was added to have a final con-
centration of 2 mM DDT and a total volume of 100 lL.The reaction was run for 10 min at 37 �C before stopped
by adding a plasma/propylthiouracil (PTU)-mix (1:1),
placing the vials in an ice bath and adding 10 % tri-
chloroacetic acid (TCA). After centrifugation, the super-
natant was collected for the determination of the released
[125I]- during the deiodination reaction. The iodine was
separated using Dowex 50 W-2X columns. The iodine was
eluted with 10 % acetic acid and the eluates counted.
Equimolar amounts of iodine and 3,30,2-T2 should be
produced. In liver and kidney, most of the deiodinase ac-
tivity is PTU-sensitive (i.e. D1).
For the analysis of D1 and D2 activities in muscle (D1Mand D2M, respectively), [
125I]-rT3 (100.000 cpm/tube) plus
2 nM rT3 were used as substrates together with ±1 mM
PTU and 20 mM DTT at 37 �C for 1 h. Approximately
200 lg of protein per tube was used. The reactions were
stopped and the released iodine separated and counted in
the same manner as with D1 in liver and kidney. D1 ac-
tivity was found by subtracting the D2 activity (insensitive
to PTU, assayed by adding PTU) from the total 50-deiodinase activity (no PTU). When using [125I]-rT3 as
substrate for the deiodinase activity in muscle, D1 activity
ranged between 22 and 49 %, whereas D2 activity ranged
Polar Biol
123

between 51 and 78 % of the total 50- deiodinase activity.
Protein determination was done following the method of
Lowry et al. (1951) after precipitation of the homogenates
with 10 % TCA (to avoid interference from DTT). The
results are given as pmol/min/mg protein (D1 in liver and
kidney) and fmol/h/mg protein (D1 and D2 in muscle).
Age and body condition
Age determination was carried out by counting the ce-
mentum growth layer groups (GLG) of the third incisor
tooth (I3) or alternatively the first premolar tooth (P1) using
the methods described in Hensel and Sorensen (1980) and
Dietz et al. (1991). Body condition was calculated based on
the body condition index (BCI) by Cattet et al. (2002)
using body mass (BM; calculated according to Derocher
and Wiig 2002) and straight-line body length.
Data treatment and statistics
The present sub-adult female and male polar bears were
treated as one group. Univariate statistics were performed
using SPSS (PASW Statistics 18.0, SPSS Inc., Chicago,
USA, 2009). Differences in means between TH-related
parameters in tissues were tested with Student’s paired
samples t test when comparing between two tissues or one-
way repeated measures ANOVA if comparing across more
than two tissues. Relationships between biological vari-
ables and TH variables were explored using Pearson cor-
relation analysis. To meet the requirements of parametric
tests, all hormone and enzyme data were log10-transformed
to approximate normal distribution. The BCI data were
normal distributed and used untransformed in the correla-
tion analyses. Significance level for all tests was p B 0.05
(two-tailed).
Results
The concentrations of the TH hormones varied across
plasma and tissues (Table 1). The order of concentrations
for T4 was plasma � liver & kidney[muscle, whereas
the order of tissues with respect to T3 concentrations was
kidney & liver � plasma & muscle. Concentrations of
fT4 and fT3 were measured in plasma, with fT4P being
almost seven times higher than fT3P (Table 1). D1 ac-
tivities in the tissues varied in the order liver[ kid-
ney � muscle, while D2 activity was only analysed in
muscle (Table 1). D2 activity in muscle was approximately
two times higher than D1 activity in muscle. Significant
differences of TH variables in the matrices are shown in
Table 1.
There were significant relationships between TH vari-
ables within and across the tissues. The concentrations of
Table 1 Thyroid hormone concentrations and deiodinase activities in plasma (T4P, T3P, fT4P, fT3P), liver (T4L, T3L, D1L), muscle (T4M, T3M,
D1M, D2M, and kidney (T4K, T3K, D1K) in East Greenland sub-adult polar bears (Ursus maritimus) sampled in February–March 2011 (N = 7)
Plasma Liver Kidney Muscle
T4 (pmol/g) Mean ± SD 13.0 ± 3.6a 3.62 ± 1.64b 3.27 ± 0.28b 0.99 ± 0.18c
Min–Max 10.1–20.2 1.18–6.03 2.83–3.56 0.75–1.15
T3 (pmol/g) Mean ± SD 1.86 ± 0.50a 7.43 ± 1.72b 8.02 ± 1.6b 1.58 ± 0.33a
Min–Max 1.28–2.58 5.02–9.25 5.29–10.4 1.29–2.14
fT4 (fmol/g) Mean ± SD 7.22 ± 1.78* NA NA NA
Min–Max 4.20–9.23 NA NA NA
fT3 (fmol/g) Mean ± SD 1.10 ± 0.98* NA NA NA
Min–Max 0.18–2.78 NA NA NA
D1-act�) (pmol/min/mgprot) Mean ± SD NA 66.9 ± 23.7** 30.2 ± 15.5** 15.9 ± 9.56***
Min–Max NA 35–95.8 5.92–43.5 5.04–32.3
D2-act (fmol/h/mgprot) Mean ± SD NA NA NA 29.3 ± 18***
Min–Max NA NA NA 16.3–67.9
NA not analysed
Different letters (a–c) denote significant differences for T4 and T3, respectively, between tissues (p B 0.05, ANOVA repeated measures test,
log10-data, df = 6)
* Significant differences in fT4 and fT3 (p B 0.05, Students’ paired t test)
** Significant differences in D1 activity between liver and kidney (p B 0.05, Students’ paired t test)
*** Significant differences between D1 and D2 activity in muscle (p B 0.05, Students’ paired t test)�) D1M is expressed as fmols/h/mgprot
Polar Biol
123
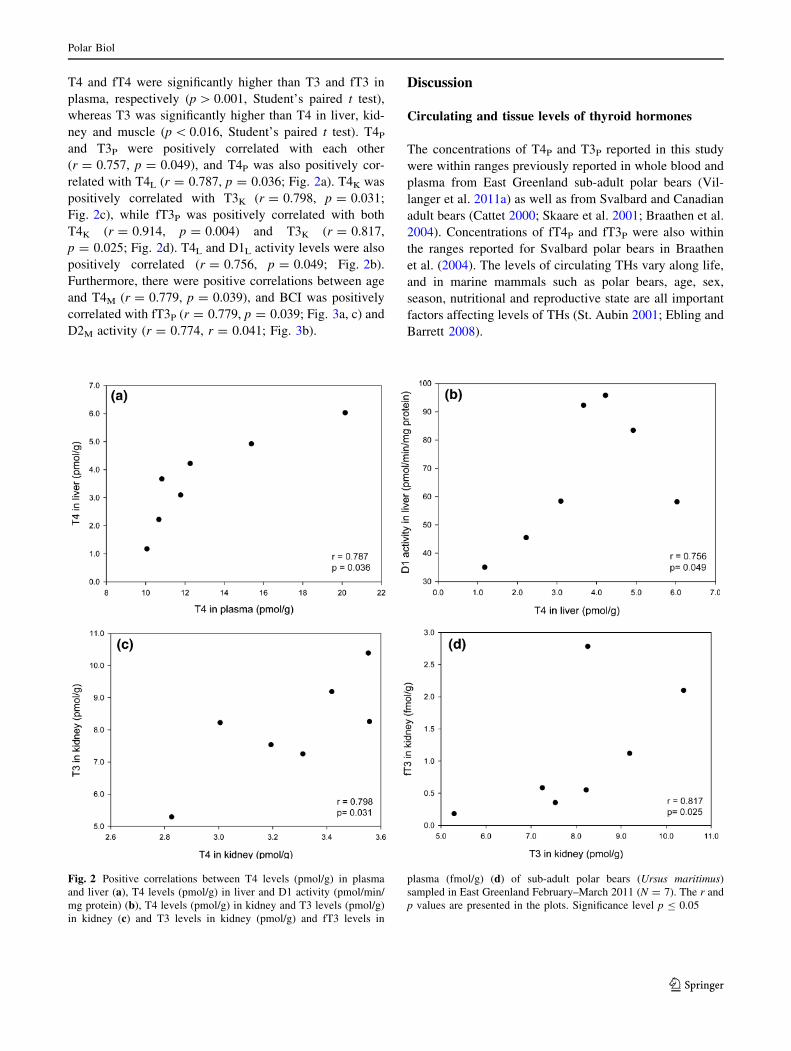
T4 and fT4 were significantly higher than T3 and fT3 in
plasma, respectively (p[ 0.001, Student’s paired t test),
whereas T3 was significantly higher than T4 in liver, kid-
ney and muscle (p\ 0.016, Student’s paired t test). T4Pand T3P were positively correlated with each other
(r = 0.757, p = 0.049), and T4P was also positively cor-
related with T4L (r = 0.787, p = 0.036; Fig. 2a). T4K was
positively correlated with T3K (r = 0.798, p = 0.031;
Fig. 2c), while fT3P was positively correlated with both
T4K (r = 0.914, p = 0.004) and T3K (r = 0.817,
p = 0.025; Fig. 2d). T4L and D1L activity levels were also
positively correlated (r = 0.756, p = 0.049; Fig. 2b).
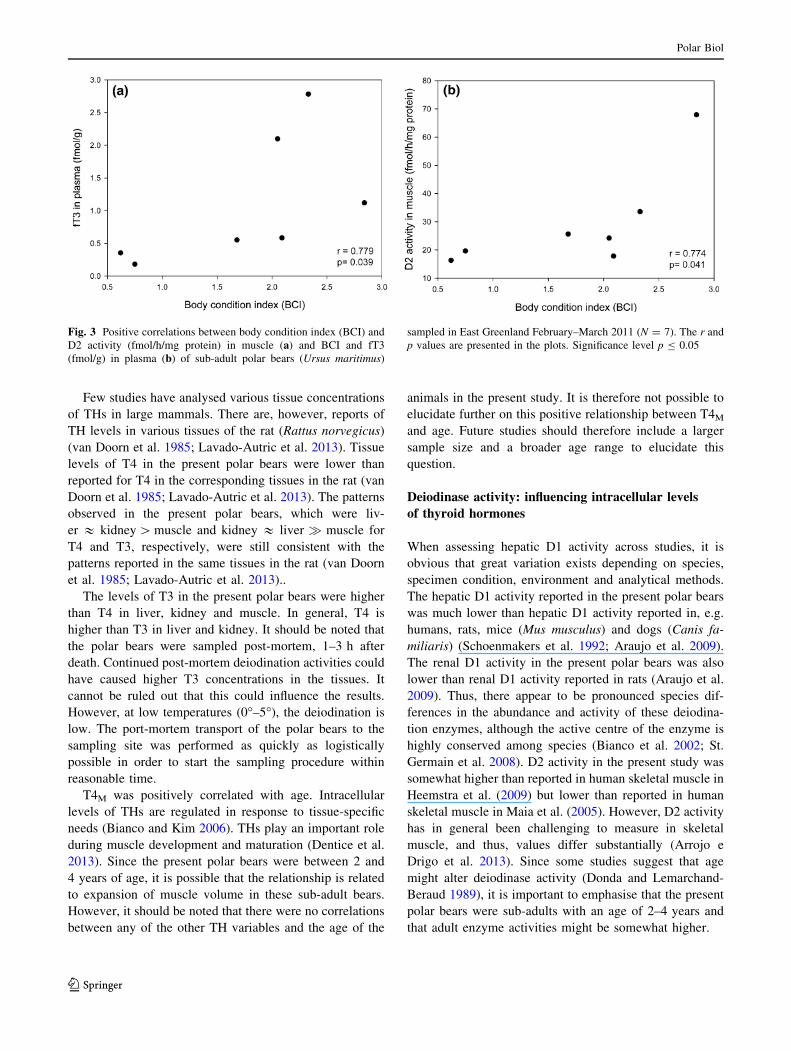
Furthermore, there were positive correlations between age
and T4M (r = 0.779, p = 0.039), and BCI was positively
correlated with fT3P (r = 0.779, p = 0.039; Fig. 3a, c) and
D2M activity (r = 0.774, r = 0.041; Fig. 3b).
Discussion
Circulating and tissue levels of thyroid hormones
The concentrations of T4P and T3P reported in this study
were within ranges previously reported in whole blood and
plasma from East Greenland sub-adult polar bears (Vil-
langer et al. 2011a) as well as from Svalbard and Canadian
adult bears (Cattet 2000; Skaare et al. 2001; Braathen et al.
2004). Concentrations of fT4P and fT3P were also within
the ranges reported for Svalbard polar bears in Braathen
et al. (2004). The levels of circulating THs vary along life,
and in marine mammals such as polar bears, age, sex,
season, nutritional and reproductive state are all important
factors affecting levels of THs (St. Aubin 2001; Ebling and
Barrett 2008).
Fig. 2 Positive correlations between T4 levels (pmol/g) in plasma
and liver (a), T4 levels (pmol/g) in liver and D1 activity (pmol/min/
mg protein) (b), T4 levels (pmol/g) in kidney and T3 levels (pmol/g)
in kidney (c) and T3 levels in kidney (pmol/g) and fT3 levels in
plasma (fmol/g) (d) of sub-adult polar bears (Ursus maritimus)
sampled in East Greenland February–March 2011 (N = 7). The r and
p values are presented in the plots. Significance level p B 0.05
Polar Biol
123
Few studies have analysed various tissue concentrations
of THs in large mammals. There are, however, reports of
TH levels in various tissues of the rat (Rattus norvegicus)
(van Doorn et al. 1985; Lavado-Autric et al. 2013). Tissue
levels of T4 in the present polar bears were lower than
reported for T4 in the corresponding tissues in the rat (van
Doorn et al. 1985; Lavado-Autric et al. 2013). The patterns
observed in the present polar bears, which were liv-
er & kidney[muscle and kidney & liver � muscle for
T4 and T3, respectively, were still consistent with the
patterns reported in the same tissues in the rat (van Doorn
et al. 1985; Lavado-Autric et al. 2013)..
The levels of T3 in the present polar bears were higher
than T4 in liver, kidney and muscle. In general, T4 is
higher than T3 in liver and kidney. It should be noted that
the polar bears were sampled post-mortem, 1–3 h after
death. Continued post-mortem deiodination activities could
have caused higher T3 concentrations in the tissues. It
cannot be ruled out that this could influence the results.
However, at low temperatures (0�–5�), the deiodination is
low. The port-mortem transport of the polar bears to the
sampling site was performed as quickly as logistically
possible in order to start the sampling procedure within
reasonable time.
T4M was positively correlated with age. Intracellular
levels of THs are regulated in response to tissue-specific
needs (Bianco and Kim 2006). THs play an important role
during muscle development and maturation (Dentice et al.
2013). Since the present polar bears were between 2 and
4 years of age, it is possible that the relationship is related
to expansion of muscle volume in these sub-adult bears.
However, it should be noted that there were no correlations
between any of the other TH variables and the age of the
animals in the present study. It is therefore not possible to
elucidate further on this positive relationship between T4Mand age. Future studies should therefore include a larger
sample size and a broader age range to elucidate this
question.
Deiodinase activity: influencing intracellular levels
of thyroid hormones
When assessing hepatic D1 activity across studies, it is
obvious that great variation exists depending on species,
specimen condition, environment and analytical methods.
The hepatic D1 activity reported in the present polar bears
was much lower than hepatic D1 activity reported in, e.g.
humans, rats, mice (Mus musculus) and dogs (Canis fa-
miliaris) (Schoenmakers et al. 1992; Araujo et al. 2009).
The renal D1 activity in the present polar bears was also
lower than renal D1 activity reported in rats (Araujo et al.
2009). Thus, there appear to be pronounced species dif-
ferences in the abundance and activity of these deiodina-
tion enzymes, although the active centre of the enzyme is
highly conserved among species (Bianco et al. 2002; St.
Germain et al. 2008). D2 activity in the present study was
somewhat higher than reported in human skeletal muscle in
Heemstra et al. (2009) but lower than reported in human
skeletal muscle in Maia et al. (2005). However, D2 activity
has in general been challenging to measure in skeletal
muscle, and thus, values differ substantially (Arrojo e
Drigo et al. 2013). Since some studies suggest that age
might alter deiodinase activity (Donda and Lemarchand-
Beraud 1989), it is important to emphasise that the present
polar bears were sub-adults with an age of 2–4 years and
that adult enzyme activities might be somewhat higher.
Fig. 3 Positive correlations between body condition index (BCI) and
D2 activity (fmol/h/mg protein) in muscle (a) and BCI and fT3
(fmol/g) in plasma (b) of sub-adult polar bears (Ursus maritimus)
sampled in East Greenland February–March 2011 (N = 7). The r and
p values are presented in the plots. Significance level p B 0.05
Polar Biol
123

Deiodinase activities are also rapidly altered in response
to endogenous factors, one of the most important ones
being the THs themselves (Kohrle, 1999). Thus, TH status
in the individuals and organs investigated might influence
the results. Recently, the deiodinases have been proposed
to be the major regulators of TH concentrations and con-
sequently TH action (Bianco et al. 2002; Bianco and Kim
2006; Arrojo e Drigo et al. 2013; Dentice et al. 2013). In
the present study, D1L activity was approximately two
times higher than D1K activity, whereas D1M had consid-
erably lower activity. That liver was the tissue with the
highest D1 activity in the present polar bears is in accor-
dance with literature on D1 activity in mammals (McNabb
1992; Bianco et al. 2002; Zoeller et al. 2007; Gereben et al.
2008). The positive correlation between T4L and D1L ac-
tivity in the present polar bears may reflect the ability of T4
to induce D1 activity (Kohrle 1999; Bianco et al. 2002).
The positive correlation reported between T4P and T4Lprobably demonstrates that liver is a primary receiver of
circulating T4, as it is an organ that receives a high blood
input and is a key organ for D1-mediated T3 production.
Thus, as in other mammals, the liver in polar bears is most
likely the primary site for D1 activity and deiodination of
T4–T3.
The relatively high D1K activity in the present study
demonstrates that the kidney is also an important organ for
D1-mediated conversion of T4 in polar bears, as it is in
other mammals (McNabb 1992; Bianco et al. 2002; Zoeller
et al. 2007; Gereben et al. 2008). The low D1M activity in
the polar bears is general in accordance with previous re-
ports of low D1 activity in skeletal muscle (Lavado-Autric
et al. 2013). D2 activity was only analysed in skeletal
muscle in the present polar bears, and it was approximately
two times higher than D1M activity. This coincides with the
less important role skeletal muscle and other D2-rich tis-
sues have in producing T3 for the plasma pool (Larsen
2009; Arrojo e Drigo et al. 2013). Due to its intracellular
position close to the nucleus, the endoplasmic reticulum
(ER)-bound D2 is thought to primarily contribute to the
production of T3 to the local nuclear pool (Bianco et al.
2002; Gereben et al. 2008; Arrojo e Drigo et al. 2013). In
certain tissues, e.g. the brain, D2 can be responsible for up
to 75 % of the nuclear pool of T3. In comparison, in the
liver only approximately 5 % of the nuclear T3 is derived
from local conversion of T4 (Hennemann et al. 2001).
Thus, D2-produced T3 is thought to be more important
locally in tissues where D2 is abundant (Bianco et al. 2002;
Zoeller et al. 2007; Gereben et al. 2008). In this manner,
TH levels can be regulated locally in response to, e.g.,
specific development periods (Gereben et al. 2008), local
tissue damage (Mourouzis et al. 2013) or metabolic and
thermogenic needs (Bianco and Kim 2006; Arrojo e Drigo
et al. 2013).
Relationships among plasma and tissue levels
of thyroid hormones
The study identified some significant relationships between
plasma TH concentrations and tissue concentrations of
THs. There were significant relationships between T4P and
T4L, and between fT3P and both T3K and T4K. As previ-
ously suggested, the positive relationship between T4P and
T4L is probably related to the importance of the hepatic
tissue in producing T3 to the plasma pool in mammals
(Gereben et al. 2008). The liver is shown to extract 5–10 %
of T4 available in plasma during just one single passage,
which is a much higher amount than can be attributed to by
fT4 alone (Mendel et al. 1988; Malik and Hodgson 2002).
Thus, a substantial amount of protein-bound T4 is sug-
gested to be available for uptake by the liver, and this is
probably related to the important role the liver plays in the
extrathyroidal tissue conversion of T4–T3.
It is also possible that the positive relationships between
T3K and fT3K and between T4K and fT3P reflect that kidney
has an important role in supplying T3 to the plasma pool in
polar bears. In general, it has become clear that tissue levels
of THs are regulated independently of plasma levels (Bianco
et al. 2002; Gereben et al. 2008; Arrojo e Drigo et al. 2013;
Dentice et al. 2013). This suggests that the plasma levels of
THs in polar bears may not directly give information re-
garding TH status in tissues. Thus, it might be that TH status
in the tissues is sufficient even if plasma levels are low, or
opposite, that tissues might show hypo- or hyperthyroid
characteristics while plasma levels are within the normal,
euthyroid range (Castillo 2011; McAninch and Bianco
2014). It does therefore appear to be important to investigate
processes that occur at the organ and tissue level in addition
to circulating levels in the future.
Relationships with body condition
In the present polar bears, fT3P and D2M were both
positively associated with BCI. BCI is a measure of indi-
vidual health and nutritional status (Cattet et al. 2002;
Robbins et al. 2004). Since BCI is related to skeletal
muscle mass and total body fat in polar bears, which will
be decreased upon fasting (Polischuk et al. 2002), one
might hypothesise that bears with a low BCI have experi-
enced a reduced availability of food and thus be in a fasting
state. In mammals, it is believed that fasting in general
results in a down-regulation of metabolic rate through a
suppression of the thyroid axis (Boelen et al. 2008). Re-
duced levels of fT3 and/or T3 in plasma have been asso-
ciated with food restriction in ursids and arctic foxes
previously (Azizi et al. 1979; Tomasi et al. 1998; Fuglei
et al. 2000; Nieminen et al. 2004). The reduction of T3 in
combination with food restriction has been suggested to be
Polar Biol
123

associated with an energy-saving mechanism to ensure
protein conservation (Carter et al. 1975; Cox et al. 1984).
As deiodinase activities also are reduced upon fasting
(Maia et al. 2011), the reduced levels of circulating T3 in
combination with fasting have been thought to be caused
by a decreased deiodination of T4–T3 by D1, as D1 is the
main producer of T3 to the plasma pool (Hulbert 2000).
However, whether the decreased D1 or D2 activities are
responsible for or secondary to the reduction on T3 levels
associated with food restriction is debated (Bianco et al.
2002; Araujo et al. 2009).
In the present polar bears, a positive relationship be-
tween BCI and D2M activity was present, indicating that
polar bears with low body condition also had low D2Mactivity. There were no relationships between BCI and D1
activity in any of the tissues. Skeletal muscle is one of the
primary target tissues of THs, as it is a tissue involved in
energy consumption as well as heat production (Dentice
et al. 2013). Thus, in muscle tissue, D2-produced T3 is
important for energy expenditure (Bianco and Kim 2006;
Arrojo e Drigo et al. 2013; Dentice et al. 2013). Fasting has
been shown to cause a down-regulation of D2 mRNA gene
expression in human skeletal muscle (Heemstra et al. 2009)
and adult male elephant seals (Lee et al. 2014), while D2
activity in BAT was decreased in food restricted rats
(Araujo et al. 2009). According to Bianco et al. (2002), it is
more likely that a fasting-associated reduction in T3 is a
consequence of D2 deficiency rather than a reduced D1-
mediated T3 production. This is because D2 have an ex-
tremely short half-life as compared to both D1 and D3.
Thus, production of D2 must be persistent in order to
maintain normal levels, and this might not be the case
during nutritional stress. Thus, it might be that the lower
levels of fT3P and reduced D2M activity in the polar bears
with a low BCI in the present study reflect energy con-
servation and/or a fasting state. This is in contrast to the
results found by Martinez et al. (2013), where D2 mRNA
gene expression and protein content in muscle were in-
creased despite prolonged fasting in developing elephant
seal pups. This indicates that there may be age- and de-
velopmental-specific responses linked to fasting, in addi-
tion to differences in responses across species. The
duration of the fasting period may also play a role, and
especially prolonged fasting, resulting in extreme ema-
ciation, may alter the responses as compared to responses
caused by fasting periods of shorter duration.
The available data in the present study do not allow for
any conclusions, as the relationships between BCI, fasting
and TH homoeostasis need further investigation. In addition,
this study has some limitations which should be addressed in
future studies. The health of the animals should be more
investigated in addition to estimating their condition, and
their actual fasting state should be determined by, e.g., using
urea/creatinine ratios. Future studies should also try to in-
crease the sample size and include additional age groups.
Nevertheless, as periods of fasting are expected to be pro-
longed for polar bears in the prospect of climatewarming and
loss of sea ice habitat, subsequently resulting in decreasing
polar bear body condition (Stirling andDerocher 1993, 2012;
Stirling et al. 1999), the role of THs in adaptations to long
periods of fasting and low body condition should be further
investigated in polar bears.
In addition, other stressors also might affect the TH ho-
moeostasis. Subsequent lipid mobilisation upon fasting
might increase concentrations of lipophilic contaminants
(Polischuk et al. 2002), another threat for the polar bears. In
addition, climate warming-mediated diet change is reported
to increase concentrations of certain legacy contaminants
and alter exposure scenarios (McKinney et al. 2009). Future
studies should therefore try to gain further insight into
regulation of tissue levels of THs in response to energy-
conserving adaptations and other TH-dependent mechan-
isms in mammals and to do so in relation to stressors such as
climate warming and contaminant exposures.
Conclusion
The liver and kidney are important extrathyroidal tissues
for deiodination of T3–T4 and supplying the plasma pool
with T3 in polar bears as it is for other mammals. The
lower fT3p concentrations and D2M activities might reflect
food restriction and energy conservation in polar bears with
a relatively low BCI. Future studies are necessary to shed
light on the involvement of THs and deiodinase enzymes in
relation to energy conservation and adaptation to prolonged
periods of fasting resulting from a warmer Arctic.
Acknowledgments The field work for this study was funded by the
Danish Co-operation for Environment in the Arctic (DANCEA). The
analyses of thyroid hormones and deiodinase activities in tissues were
supported by research Grants SAF2012-32491 from MINECO and
S2010/BMD-2423 from CAM, Spain. The authors thank all those
who contributed to the field work in the Scoresby Sound area in East
Greenland, including the local hunters who provided the polar bears
for this study through their aboriginal hunting quotas. The authors
would also like to thank Sigga Joensen and Rune Dietz at Aarhus
University for performing the ageing of the polar bears and Grethe
Stavik Eggen at NTNU for help with the thyroid hormone analysis in
plasma. The study was funded by the Norwegian University of Sci-
ence and Technology (NTNU).
References
Araujo R, Andrade B, da Silva M, Ferreira A, Carvalho D (2009)
Tissue-specific deiodinase regulation during food restriction and
low replacement dose of leptin in rats. Am J Physiol Endocrinol
Metab 296:E1157–E1163
Polar Biol
123

Arrojo e Drigo R, Fonseca TL, Werneck-de-Castro JPS, Bianco AC
(2013) Role of the type 2 iodothyronine deiodinase (D2) in the
control of thyroid hormone signaling. Biochim Biophys Acta
Gen Subj 1830:3956–3964
Azizi F, Mannix JE, Howard D, Nelson RA (1979) Effect of winter
sleep on pituitary–thyroid-axis in american black bear. Am J
Physiol 237:E227–E230
Best R (1982) Thermoregulation in resting and active polar bears.
J Comp Physiol 146:63–73
Bianco AC (2013) Cracking the code for thyroid hormone signaling.
Tran Am Clin Climatol Assoc 124:26–35
Bianco AC, Kim BW (2006) Deiodinases: implications of the local
control of thyroid hormone action. J Clin Investig
116:2571–2579
Bianco AC, Salvatore D, Gereben B, Berry MJ, Larsen PR (2002)
Biochemistry, cellular and molecular biology, and physiological
roles of the iodothyronine selenodeiodinases. Endocr Rev
23:38–89
Boelen A, Wiersinga WM, Fliers E (2008) Fasting-induced changes
in the hypothalamus–pituitary–thyroid axis. Thyroid 18:123–129
Boily P (1996) Metabolic and hormonal changes during the molt of
captive gray seals (Halichoerus grypus). Am J Physiol 270:R1051–
R1058
Braathen M, Derocher A, Wiig O, Sormo E, Lie E, Skaare J, Jenssen
B (2004) Relationships between PCBs and thyroid hormones and
retinol in female and male polar bears. Environ Health Perspect
112:826–833
Bytingsvik J (2012) Organohalogenated contaminants (OHCs) in
polar bear mother-cub pairs from Svalbard, Norway. Disserta-
tion, Norwegian University of Science and Technology
Carter WJ, Shakir KM, Hodges S, Faas FH, Wynn JO (1975) Effect of
thyroid hormone on metabolic adaptation to fasting. Metabolism
24:1177–1183
Castillo V (2011) Canine hypothyroidism. Veterinary Focus 21:2–8
Cattet MRL (2000) Biochemical and physiological aspects of obesity,
high fat diet, and prolonged fasting in free-ranging polar bears.
Dissertation, University of Saskatchewan
Cattet MRL, Caulkett NA, Obbard ME, Stenhouse GB (2002) A
body-condition index for ursids. Can J Zool 80:1156–1161
Cheng S-Y, Leonard JL, Davis PJ (2010) Molecular aspects of thyroid
hormone actions. Endocr Rev 31:139–170
Cox MD, Dalal SS, Heard CRC, Millward DJ (1984) Metabolic rate
and thyroid status in rats fed diets of different protein-energy
value: the importance of free T3. J Nutr 114:1609–1616
Crocker DE, Ortiz RM, Houser DS, Webb PM, Costa DP (2012)
Hormone and metabolite changes associated with extended
breeding fasts in male northern elephant seals (Mirounga
angustirostris). Comp Biochem Physiol A 161:388–394
Dentice M, Marsili A, Zavacki A, Larsen PR (2013) The deiodinases
and the control of intracellular thyroid hormone signaling during
cellular differentiation. Biochim Biophys Acta Gen Subj
1830:3937–3945
Derocher AE, Wiig Ø (2002) Postnatal growth in body length and
mass of polar bears (Ursus maritimus) at Svalbard. J Zool
256:343–349
Derocher AE, Nelson RA, Stirling I, Ramsay MA (1990) Effects of
fasting and feeding on serum urea and serum creatinine levels in
polar bears. Mar Mamm Sci 6:196–203
Dietz R, Heide-Jørgensen MP, Harkonen T, Teilmann J, Valentin N
(1991) Age determination of european harbour seal, Phoca
vitulina L. Sarsia 76:17–21
Donda A, Lemarchand-Beraud T (1989) Aging alters the activity of
50-deiodinase in the adenohypophysis, thyroid gland, and liver of
the male rat. Endocrinology 124:1305–1309
Ebling FJP, Barrett P (2008) The regulation of seasonal changes in
food intake and body weight. J Neuroendocrinol 20:827–833
Fuglei E, Aanestad M, Berg JP (2000) Hormones and metabolites of
arctic foxes (Alopex lagopus) in response to season, starvation
and re-feeding. Comp Biochem Physiol A 126:287–294
Gabrielsen KM, Villanger GD, Lie E, Karimi M, Lydersen C, Kovacs
KM, Jenssen BM (2011) Levels and patterns of hydroxylated
polychlorinated biphenyls (OH-PCBs) and their associations
with thyroid hormones in hooded seal (Cystophora cristata)
mother–pup pairs. Aquat Toxicol 105:482–491
Gereben B, Zavacki AM, Ribich S, Kim BW, Huang SA, Simonides
WS, Zeold A, Bianco AC (2008) Cellular and molecular basis of
deiodinase-regulated thyroid hormone signaling. Endocr Rev
29:898–938
Heemstra KA, Soeters MR, Fliers E, Serlie MJ, Burggraaf J, van
Doorn MB, van Der Klaauw AA, Romijn JA, Smit JW, Corssmit
EP, Visser TJ (2009) Type 2 iodothyronine deiodinase in skeletal
muscle: effects of hypothyroidism and fasting. J Clin Endocrinol
Metab 94:2144–2150
Hennemann G, Docter R, Friesema ECH, de Jong M, Krenning EP,
Visser TJ (2001) Plasma membrane transport of thyroid
hormones and its role in thyroid hormone metabolism and
bioavailability. Endocr Rev 22:451–476
Hensel RJ, Sorensen FE (1980) Age determination of live polar bears.
Int Conf Bear Res Manag 4:93–100
Hissa R, Siekkinen J, Hohtola E, Saarela S, Hakala A, Pudas J (1994)
Seasonal patterns in the physiology of the european brown bear
(Ursus arctos arctos) in Finland. Comp Biochem Physiol A
109:781–791
Hulbert AJ (2000) Thyroid hormones and their effects: a new
perspective. Biol Rev 75:519–631
Jenssen BM, Villanger GD, Gabrielsen KM, Bytingsvik J, Bechshoft
T, Ciesielski TM, Sonne C, Dietz R (2015) Anthropogenic flank
attack on polar bears: interacting consequences of climate
warming and pollutant exposure. Front Ecol Evol 3:16
Kelso EJ, Champagne CD, Tift MS, Houser DS, Crocker DE (2012) Sex
differences in fuel use and metabolism during development in
fasting juvenile northern elephant seals. J Exp Biol 215:2637–2645
Kohrle J (1999) Local activation and inactivation of thyroid hormones:
the deiodinase family. Mol Cell Endocrinol 151:103–119
Kohrle J (2000) The deiodinase family: selenoenzymes regulating
thyroid hormone availability and action. Cell Mol Life Sci
57:1853–1863
Kunisue T, Fisher JW, Kannan K (2011) Determination of six thyroid
hormones in the brain and thyroid gland using isotope-dilution
liquid chromatography/tandem mass spectrometry. Anal Chem
83:417–424
Larsen PR (2009) Type 2 iodothyronine deiodinase in human skeletal
muscle: new insights into its physiological role and regulation.
J Clin Endocrinol Metab 94:1893–1895
Lavado-Autric R, Calvo RM, de Mena RM, Morreale de Escobar G,
Obregon MJ (2013) Deiodinase activities in thyroids and tissues
of iodine-deficient female rats. Endocrinology 154:529–536
Leatherland JF, Ronald K (1981) Plasma concentrations of thyroid
hormones in a captive and feral polar bear (Ursus maritimus.
Comp Biochem Physiol A 70:575–577
Lee D, Martinez B, Crocker D, Ortiz R (2014) Thyroid hormone
changes associated with prolonged food deprivation in adult
male northern elephant seals. FASEB J 28:1101.6
Letcher R, Bustnes J, Dietz R, Jenssen B, Jorgensen E, Sonne C,
Verreault J, Vijayan M, Gabrielsen G (2010) Exposure and
effects assessment of persistent organohalogen contaminants in
arctic wildlife and fish. Sci Total Environ 408:2995–3043
Lowry O, Rosebrough N, Farr A, Randall R (1951) Protein
measurement with the folin phenol reagent. J Biol Chem 193:265
Lydersen C, Wolkers H, Severinsen T, Kleivane L, Nordoy E, Skaare
J (2002) Blood is a poor substrate for monitoring pollution
burdens in phocid seals. Sci Total Environ 292:193–203
Polar Biol
123

Maia AL, Kim BW, Huang SA, Harney JW, Larsen PR (2005) Type 2
iodothyronine deiodinase is the major source of plasma T3 in
euthyroid humans. J Clin Investig 115:2524–2533
Maia AL, Goemann IM, Meyer ELS, Wajner SM (2011) Type 1
iodothyronine deiodinase in human physiology and disease:
deiodinases: the balance of thyroid hormone. J Endocrinol 209:283
Malik R, Hodgson H (2002) The relationship between the thyroid
gland and the liver. QJM 95:559–569
Martinez B, Sonanez-Organis JG, Vazquez-Medina JP, Viscarra JA,
MacKenzie DS, Crocker DE, Ortiz RM (2013) Prolonged food
deprivation increases mRNA expression of deiodinase 1 and 2,
and thyroid hormone receptor b-1 in a fasting-adapted mammal.
J Exp Biol 216:4647–4654
McAninch EA, Bianco AC (2014) Thyroid hormone signaling in
energy homeostasis and energy metabolism. Ann N Y Acad Sci
1311:77–87
McCannUD, Shav EA,KaplanMM (1984) Iodothyronine deiodination
reaction types in several rat tissues: effects of age, thyroid status,
and glucocorticoid treatment. Endocrinology 114:1513–1521
McKinney MA, Peacock E, Letcher RJ (2009) Sea ice-associated diet
change increases the levels of chlorinated and brominated
contaminants in polar bears. Environ Sci Technol 43:4334–4339
McKinneyMA,LetcherRJ,Aars J,BornEW,BraniganM,DietzR,Evans
TJ,GabrielsenGW,PeacockE, SonneC (2011)Flame retardants and
legacy contaminants in polar bears from Alaska, Canada, East
Greenland and Svalbard, 2005–2008. Environ Int 37:365–374
McNabb FMA (1992) Thyroid hormones. Prentice Hall, New Jersey
Mendel CM, Cavalieri RR, Weisiger RA (1988) Uptake of thyroxine
by the perfused rat-liver—implications for the free hormone
hypothesis. Am J Physiol 255:E110–E119
Morreale de Escobar G, Pastor R, Obregon MJ, Rey FED (1985)
Effects of maternal hypothyroidism on the weight and thyroid
hormone content of rat embryonic tissues, before and after onset
of fetal thyroid function. Endocrinology 117:1890–1900
Mourouzis I, Politi E, Pantos C (2013) Thyroid hormone and tissue
repair: new tricks for an old hormone? J Thyroid Res 2013:5
Nieminen P, Pyykonen T, Asikainen J, Mononen J, Mustonen A-M
(2004) Effects of fasting and exogenous melatonin on annual
rhythms in the blue fox (Alopex lagopus). Comp Biochem
Physiol A 139:183–197
Øritsland NA (1970) Temperature regulation of the polar bear
(Thalarctos maritimus). Comp Biochem Physiol 37:225–233
Orozco A, Valverde-R C, Olvera A, Garcıa-G C (2012) Iodothyronine
deiodinases: a functional and evolutionary perspective. J En-
docrinol 215:207–219
Polischuk SC, Norstrom RJ, Ramsay MA (2002) Body burdens and
tissue concentrations of organochlorines in polar bears (Ursus
maritimus) vary during seasonal fasts. Environ Pollut 118:29–39
Raasmaja A, Viluksela M, Rozman KK (1996) Decreased liver type I
50-deiodinase and increased brown adipose tissue type II 50-deiodinase activity in 2,3,7,8-tetrachlorodibenzo-p-dioxin
(TCDD)-treated long-evans rats. Toxicology 114:199–205
Robbins CT, Schwartz CC, Felicetti LA (2004) Nutritional ecology of
ursids: a review of newer methods and management implica-
tions. Ursus 15:161–171
Robbins CT, Lopez-Alfaro C, Rode KD, Toien O, Nelson OL (2012)
Hibernation and seasonal fasting in bears: the energetic costs and
consequences for polar bears. J Mammal 93:1493–1503
Rosing-Asvid A, Born E, Kingsley M (2002) Age at sexual maturity
of males and timing of the mating season of polar bears (Ursus
maritimus) in Greenland. Polar Biol 25:878–883
Santisteban P, Bernal J (2005) Thyroid development and effect on the
nervous system. Rev Endocr Metab Disord 6:217–228
Schoenmakers CHH, Pigmans IGAJ, Visser TJ (1992) Species
differences in liver type I iodothyronine deiodinase. Biochim
Biophys Acta Protein Struct Mol Enzymol 1121:160–166
Scholander P, Hock R, Walters V, Irving L (1950a) Adaptation to
cold in arctic and tropical mammals and birds in relation to body
temperature, insulation, and basal metabolic rate. Biol Bull
99:259–271
Scholander P, Hock R, Walters V, Johnson F, Irving L (1950b) Heat
regulation in some arctic and tropical mammals and birds. Biol
Bull 99:237–258
Silva J (2006) Thermogenic mechanisms and their hormonal regula-
tion. Physiol Rev 86:435–464
Skaare JU, Bernhoft A, Wiig O, Norum KR, Haug E, Eide DM,
Derocher AE (2001) Relationships between plasma levels of
organochlorines, retinol and thyroid hormones from polar bears
(Ursus maritimus) at Svalbard. J Toxicol Environ Health Part A
62:227–241
Sonne C (2010) Health effects from long-range transported con-
taminants in arctic top predators: an integrated review based on
studies of polar bears and relevant model species. Environ Int
36:461–491
St. Aubin DS (2001) Chapter 10. Endocrinology. In: La D, Gulland F
(eds) CRC handbook of marine mammal medicine: health,
disease, and rehabilitation, 2nd edn. CRC Press, Boca Raton
St. Germain D, Galton V, Hernandez A (2008) Minireview: defining
the roles of the iodothyronine deiodinases: current concepts and
challenges. Endocrinology 150:1097–1107
Stirling I (2002) Polar bear Ursus maritimus. In: Perrin WF, Bernd W,
Thewissen JGM (eds) Encyclopedia of marine mammals.
Academic Press, San Diego
Stirling I, Derocher AE (1993) Possible impacts of climatic warming
on polar bears. Arctic 46:240–245
Stirling I,DerocherAE (2012)Effects of climatewarmingonpolar bears:
a review of the evidence. Glob Change Biol 18:2694–2706
Stirling I, Lunn NJ, Iacozza J (1999) Long-term trends in the
population ecology of polar bears in western hudson bay in
relation to climatic change. Arctic 52:294–306
Tomasi TE, Hellgren EC, Tucker TJ (1998) Thyroid hormone
concentrations in black bears (Ursus americanus): hibernation
and pregnancy effects. Gen Comp Endocrinol 109:192–199
van Doorn J, Roelfsema F, Heide DVD (1985) Concentrations of
thyroxine and 3,5,30-triiodothyronine at 34 different sites in
euthyroid rats as determined by an isotopic equilibrium
technique. Endocrinology 117:1201–1208
Villanger GD, Jenssen BM, Fjeldberg RR, Letcher RJ, Muir DCG,
Kirkegaard M, Sonne C, Dietz R (2011a) Exposure to mixtures of
organohalogen contaminants and associative interactions with
thyroid hormones in East Greenland polar bears (Ursus mar-
itimus). Environ Int 37:694–708
Villanger GD, Lydersen C, Kovacs KM, Lie E, Skaare JU, Jenssen BM
(2011b) Disruptive effects of persistent organohalogen con-
taminants on thyroid function in white whales (Delphinapterus
leucas) from Svalbard. Sci Total Environ 409:2511–2524. doi:10.
1016/j.scitotenv.2011.03.014
Viluksela M, Raasmaja A, Lebofsky M, Stahl BU, Rozman KK
(2004) Tissue-specific effects of 2,3,7,8-tetrachlorodibenzo-p-
dioxin (TCDD) on the activity of 50-deiodinases I and II in rats.
Toxicol Lett 147:133–142
Weeke J, Orskov H (1973) Synthesis of l25I monolabeled 3,5,30-triiodo-thyronine and thyroxine of maximum specific activity for
radioimmunoassay. Scand J Clin Lab Investig 32:357
Welch AJ, Bedoya-Reina OC, Carretero-Paulet L, Miller W, Rode
KD, Lindqvist C (2014) Polar bears exhibit genome-wide
signatures of bioenergetic adaptation to life in the arctic
environment. Gen Biol Evol 6:433–450
Zoeller RT, Tan S, Tyl R (2007) General background on the hypotha-
lamic–pituitary–thyroid (HTP) axis. Crit Rev Toxicol 37:11–53
Polar Biol
123




















