
Cyclin D1 Is a Selective Modifier of Androgen-dependent Signaling and Androgen Receptor Function

Original article 1
Androgen receptor gene CAG repeat length as a modifier ofthe association between persistent organohalogen pollutantexposure markers and semen characteristicsAleksander Giwercmana, Lars Rylanderb, Anna Rignell-Hydbomb, Bo A.G.Jonssonb, Henning S. Pedersenc, Jan K. Ludwickif, Vladimir Lesovoyg, ValentynaZvyezdayh, Marcello Spanoi, Gian-Carlo Manicardij, Davide Bizzarok, Eva C.Bonefeld-Jørgensend, Gunnar Tofte, Jens Peter Bondee, Charlotte Giwercmana,Tarmo Tiidoa, Yvonne Lundberg Giwercmana and INUENDOl
Objectives Exposure to persistent organohalogen pollu-
tants was suggested to impair male reproductive function.
A gene–environment interaction has been proposed. No
genes modifying the effect of persistent organohalogen
pollutants on reproductive organs have yet been identified.
We aimed to investigate whether the CAG and GGN
polymorphisms in the androgen receptor gene modify the
effect of persistent organohalogen pollutant exposure on
human sperm characteristics.
Methods Semen and blood from 680 men [mean (SD) age
34 (10) years] from Greenland, Sweden, Warsaw (Poland)
and Kharkiv (Ukraine) were collected. Persistent organo-
halogen pollutant exposure was assessed by measuring
serum levels of 2,20, 4,40,5,50-hexachlorobiphenyl (CB-153)
and dichlorodiphenyl dichloroethene (p,p0-DDE). Semen
characteristics (volume, sperm concentration, total count,
proportion of progressively motile and morphology) and
DNA fragmentation index (DFI) were determined. CAG and
GGN repeat lengths were determined by direct sequencing
of leukocyte DNA.
Results A statistically significant interaction was found
between the CB-153 group and CAG repeat category in
relation to sperm concentration and total sperm count
(P = 0.03 and 0.01, respectively). For p,p0-DDE, in the
European cohorts a significant interaction was found in
relation to DFI (P = 0.01). For CAG < 20, sperm concentra-
tion and total sperm count were 35 and 42% lower,
respectively, when the group with CB-153 exposure above
median was compared with that below the median. DFI was
40% higher in the high p,p0-DDE exposure group for
CAGr21.
Conclusions This study indicated that the androgen re-
ceptor CAG repeat length might modify the susceptibility of
an individual to the adverse effects of persistent organo-
halogen pollutant exposure on semen quality. Other
studies regarding this matter are warranted. Pharmaco-
genetics and Genomics 00:000–000 �c 2006 Lippincott
Williams & Wilkins.
Pharmacogenetics and Genomics 2006, 00:000–000
Keywords: androgen receptor, CAG repeats, gene–environment interaction,persistent organohalogen pollutants, semen quality
aMolecular Reproductive Medicine Research Unit, Department of ClinicalSciences, Fertility Centre, Malmo University Hospital, Lund University, Malmo,bDivision of Occupational and Environmental Medicine and PsychiatricEpidemiology, Lund University, Lund University Hospital, Lund, Sweden, cCentrefor Arctic Environmental Medicine, Nuuk, Greenland, dInstitute of Public Health,Department of Occupational and Environmental Medicine, University of Aarhus,eDepartment of Occupational Medicine, Aarhus University Hospital, Aarhus,Denmark, fDepartment of Environmental Toxicology, National Institute of Hygiene,Warsaw, Poland, gRegional Clinical Center of Urology and Nephrology,hLaboratory of Human Reproduction, Kharkiv State Medical University, Kharkiv,Ukraine, iSection of Toxicology and Biomedical Sciences, BIOTEC-MED, ENEACasaccia Research Centre, Rome, jUniversity of Modena and Reggio Emilia,Reggio Emilia, kIstituto di Biologia e Genetica Universita Politecnica delle Marche,Ancona, Italy and lwww.inuendo.dk.
Correspondence and requests for reprints to Professor Aleksander Giwercman,Fertility Centre, Malmo University Hospital, SE 205 02 Malmo, SwedenTel: + 46 40 33 79 14; fax: + 46 40 33 82 66;e-mail: [email protected]
The INUENDO Project: Biopersistent organochlorines in diet and human fertility.Epidemiological studies of time to pregnancy and semen quality in Inuit andEuropean populations, is supported by The European Commission to the 5thFramework Programme Quality of Life and Management of living resources, Keyaction four on environment and health, (Contract no. QLK4-CT-2001–00202),running 01.01.02–30.06.05. www.inuendo.dk. The Ukrainian part of the studywas supported by a grant from INTAS (Contract no. 2001 2205). The Swedishpart of the study has also been funded by the Swedish Medical Research Council(Grant No: 521-2004-6072) and the Swedish Research Council for Environment,Agricultural sciences and Spatial Planning.
Received 28 April 2006 Accepted 18 July 2006
IntroductionThe alarming reports on a time-related impairment of
male reproductive function, together with an increased
incidence of testicular cancer, cryptorchidism and hypos-
padias and a possible decline in sperm counts, have drawn
attention to the effects of ‘endocrine disrupters’ [1]. The
endocrine disrupters include a long list of chemicals with
agonistic or antagonistic properties with respect to the
action of endogenous hormones. Among the endocrine
disrupters, persistent organohalogen pollutants (POPs)
have been considered as strong candidates for interfering
with the male reproductive system.
CE: ayesha ED: raji Op: vp FPC: lww_fpc_200712
1744-6872 �c 2006 Lippincott Williams & Wilkins

POPs, for example, polychlorinated dibenzofurans
(PCDFs), polychlorinated dibenzo-p-dioxins (PCDD),
polychlorinated biphenyls (PCBs), dichlorodiphenyl tri-
chloroethane (DDT), and dichlorodiphenyl dichlor-
oethene (p,p0-DDE), the most stable metabolite of
DDT, are ubiquitous environmental contaminants. As a
result of the hydrophobic and lipophilic nature of these
chemicals and their long half-times, these compounds are
highly persistent, with a tendency to bioaccumulate and
biomagnify in the food chain. Studies have shown
measurable levels of PCBs and p,p0-DDE in a large
proportion of the general population almost everywhere
[2]. Some of these POPs can disrupt multiple endocrine
pathways and induce a wide range of toxic responses [1].
A number of studies have demonstrated their estrogenic,
anti-estrogenic, dioxin-like and androgen competing
properties [3–6].
A finding of significant ethnical and geographical
differences in semen quality and in the incidence of
testicular cancer and of congenital malformations raises
the question of an interaction between genetic and
environmental/lifestyle-related factors, certain genotypes
making individuals more susceptible to adverse exogen-
ous exposures [7,8]. Studying such interactions has
biological, epidemiological and public health-related
implications. It could help us in understanding the
background for the defects in the male reproductive
organs, facilitate proper design of epidemiological studies
and add to identifying individuals susceptible to certain
environmental and lifestyle-related hazards. Such ‘sus-
ceptibility genes’ need to be identified, those involved in
the action of sex steroids being strong candidates.
Serum concentrations of 2,20,4,40,5,50-hexachlorobiphenyl
(CB-153) and p,p0-DDE are suitable biomarkers for the
assessment of POP exposure. The PCB congener CB-153
is highly correlated with total PCB concentration [9,10],
the estimated 2,3,7,8-tetrachlorodibenzo-p-dioxin equiva-
lent (TEQ) from PCB, and the total POP-derived TEQ
[11], respectively. Likewise, the major DDT metabolite
p,p0-DDE, which is an antiandrogenic compound [4], is
another good indicator of exposure. Previous studies
indicate that the exposure levels for p,p0-DDE still are
considerably high in many countries [11,12]. Recently, as
a part of a study supported by the European Union
(www.inuendo.dk), the impact of POP exposure on
semen characteristics in four populations was investi-
gated: Swedish fishermen and spouses to pregnant
women from Greenland, (Warsaw) Poland and (Kharkiv)
Ukraine [13,14]. The major findings of this study were
negative association between serum CB-153 and sperm
motility and sperm DNA integrity, respectively [13,15–
17].
As androgens play a crucial role in the development and
function of male reproductive organs, polymorphisms in
the androgen receptor gene (AR) might modify the effect
of POP on semen characteristics. The fine-tuning of the
AR function is regulated by two polymorphic sequences
in the transactivating part of the receptor; that is,
polyglutamine encoding CAG repeats and polyglycine
encoding GGN triplets [18,19]. Previous studies have
indicated that the CAG and the GGN number can
influence the functional status of the AR. In Caucasian
men, the CAG number is normally distributed with a
median length of 22 and normal range between 12 and 30
[20]. Men with more than 40 CAGs suffer from
Kennedy’s disease characterized by neuromuscular symp-
toms and hypogonadism [21]. In-vitro [19] and in-vivo
[22,23] studies, however, have indicated that even within
the normal CAG length range, there is an inverse
correlation between CAG number and receptor function,
including lower sperm counts in those with long CAGs.
Less information is available regarding the physiological
role of the GGN triplets. In a Caucasian population, the
lengths of 23 and 24 encounters for 50 and 35% of all
genotypes, respectively. In-vivo data indicate that the
length of 23 is associated with superior receptor activity,
with shorter and longer GGNs giving less well functioning
AR [24–26].
Apart from regulating the sensitivity to endogenous
androgens, however, AR polymorphisms might influence
the effect of endocrine disrupters acting through this
receptor. Our main objective was, therefore, to investi-
gate whether the length of these repeats modified the
effect of POP exposure on semen characteristics. We also
used the data set to assess whether there were any
associations between CAG and GGN repeat lengths and
semen characteristics.
Study participants and methodsThe details of the study populations, methods applied
and the data on association between exposure and semen
characteristics have been published previously [14,17].
Study populations
The present study is part of a European study on fertility
(INUENDO) using a uniform protocol for data collection
in Greenland, Sweden, Kharkiv in Ukraine and Warsaw in
Poland [14]. The participation rates ranged from 7% in
the Swedish fishermen, 29% in Warsaw, 33% in Kharkiv to
79% in Greenland.
The populations were selected to represent different
exposure scenarios of relatively high POP exposure in the
populations from Greenland and Sweden, with high
consumption of POP-contaminated seafood. The Ukrai-
nian population was expected to be exposed to POPs
2 Pharmacogenetics and Genomics 2006, Vol 00 No 00
mainly from previous agricultural use of organochlorine
pesticides, whereas the exposure level in Poland was
expected to be in the lower range and similar to other
European populations without specific exposure sources.
Men were recruited for the study as partners of pregnant
women who were consecutively listed when a pregnancy
was recorded at the participating hospitals, except for the
Swedish men who were recruited from a previously
established population of fishermen, independent of
fertility. Seventy-nine percent of the Swedish fishermen,
however, had fathered a child. In all populations, the men
were consecutively asked to donate a semen sample until
approximately 200 men had agreed to participate. All
included men were 18 years or more at the time of
enrolment.
The men gave their informed consent and were asked to
deliver a semen sample and a blood sample for CB-153
and p,p0-DDE analyses as well as for extraction of
leukocyte DNA.
In total, 798 men delivered a semen sample. Among
those, CB-153 and p,p0-DDE data were lacking in 35. Of
the remaining 763 men, DNA for genotyping of the AR
was available in 680 who, thus, comprised the study
population. Semen characteristics included volume,
concentration, total sperm count, proportion progres-
sively motile and percentage morphologically normal;
sperm chromatin characteristics did not differ between
the 680 men included in the study and the 118 excluded
owing to lack of genetic and/or exposure data (data not
shown). Similarly, there were no differences in CB-153 or
p,p0-DDE levels between the 680 men included and the
83 in whom POP levels, but not genotype, were assessed
(data not shown).
Local ethical committees approved the study.
The background characteristics of the present study
population are given in Table 1.
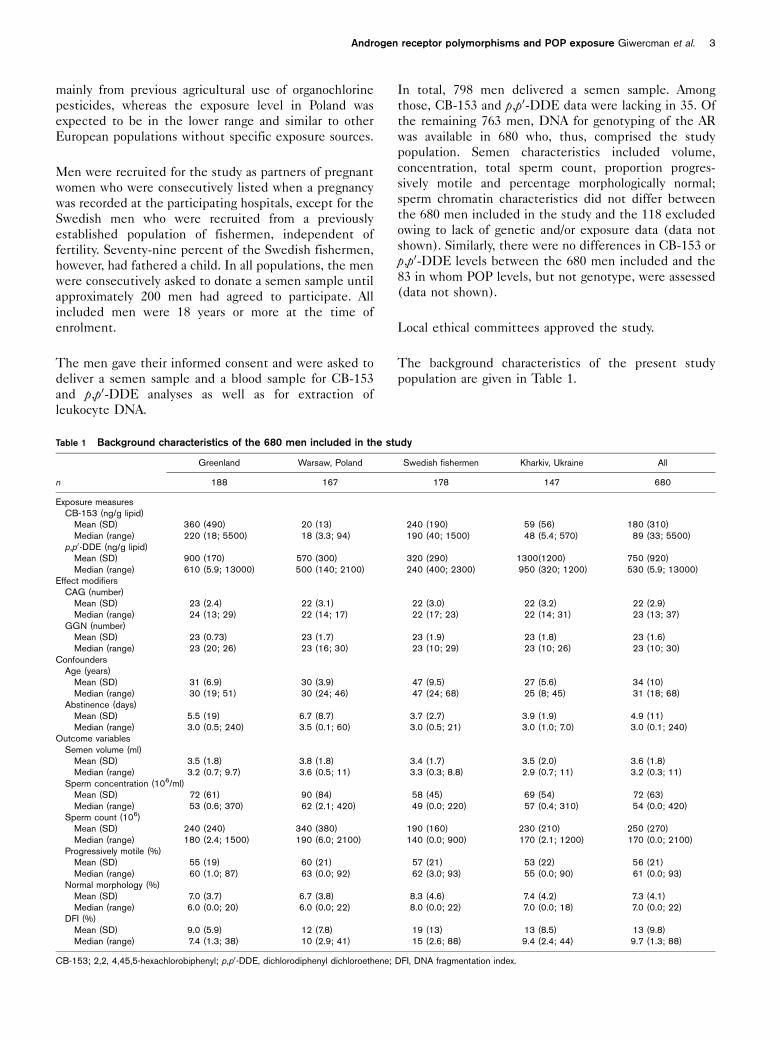
Table 1 Background characteristics of the 680 men included in the study
Greenland Warsaw, Poland Swedish fishermen Kharkiv, Ukraine All
n 188 167 178 147 680
Exposure measuresCB-153 (ng/g lipid)
Mean (SD) 360 (490) 20 (13) 240 (190) 59 (56) 180 (310)Median (range) 220 (18; 5500) 18 (3.3; 94) 190 (40; 1500) 48 (5.4; 570) 89 (33; 5500)
p,p0-DDE (ng/g lipid)Mean (SD) 900 (170) 570 (300) 320 (290) 1300(1200) 750 (920)Median (range) 610 (5.9; 13000) 500 (140; 2100) 240 (400; 2300) 950 (320; 1200) 530 (5.9; 13000)
Effect modifiersCAG (number)
Mean (SD) 23 (2.4) 22 (3.1) 22 (3.0) 22 (3.2) 22 (2.9)Median (range) 24 (13; 29) 22 (14; 17) 22 (17; 23) 22 (14; 31) 23 (13; 37)
GGN (number)Mean (SD) 23 (0.73) 23 (1.7) 23 (1.9) 23 (1.8) 23 (1.6)Median (range) 23 (20; 26) 23 (16; 30) 23 (10; 29) 23 (10; 26) 23 (10; 30)
ConfoundersAge (years)
Mean (SD) 31 (6.9) 30 (3.9) 47 (9.5) 27 (5.6) 34 (10)Median (range) 30 (19; 51) 30 (24; 46) 47 (24; 68) 25 (8; 45) 31 (18; 68)
Abstinence (days)Mean (SD) 5.5 (19) 6.7 (8.7) 3.7 (2.7) 3.9 (1.9) 4.9 (11)Median (range) 3.0 (0.5; 240) 3.5 (0.1; 60) 3.0 (0.5; 21) 3.0 (1.0; 7.0) 3.0 (0.1; 240)
Outcome variablesSemen volume (ml)
Mean (SD) 3.5 (1.8) 3.8 (1.8) 3.4 (1.7) 3.5 (2.0) 3.6 (1.8)Median (range) 3.2 (0.7; 9.7) 3.6 (0.5; 11) 3.3 (0.3; 8.8) 2.9 (0.7; 11) 3.2 (0.3; 11)
Sperm concentration (106/ml)Mean (SD) 72 (61) 90 (84) 58 (45) 69 (54) 72 (63)Median (range) 53 (0.6; 370) 62 (2.1; 420) 49 (0.0; 220) 57 (0.4; 310) 54 (0.0; 420)
Sperm count (106)Mean (SD) 240 (240) 340 (380) 190 (160) 230 (210) 250 (270)Median (range) 180 (2.4; 1500) 190 (6.0; 2100) 140 (0.0; 900) 170 (2.1; 1200) 170 (0.0; 2100)
Progressively motile (%)Mean (SD) 55 (19) 60 (21) 57 (21) 53 (22) 56 (21)Median (range) 60 (1.0; 87) 63 (0.0; 92) 62 (3.0; 93) 55 (0.0; 90) 61 (0.0; 93)
Normal morphology (%)Mean (SD) 7.0 (3.7) 6.7 (3.8) 8.3 (4.6) 7.4 (4.2) 7.3 (4.1)Median (range) 6.0 (0.0; 20) 6.0 (0.0; 22) 8.0 (0.0; 22) 7.0 (0.0; 18) 7.0 (0.0; 22)
DFI (%)Mean (SD) 9.0 (5.9) 12 (7.8) 19 (13) 13 (8.5) 13 (9.8)Median (range) 7.4 (1.3; 38) 10 (2.9; 41) 15 (2.6; 88) 9.4 (2.4; 44) 9.7 (1.3; 88)
CB-153; 2,2, 4,45,5-hexachlorobiphenyl; p,p0-DDE, dichlorodiphenyl dichloroethene; DFI, DNA fragmentation index.
Androgen receptor polymorphisms and POP exposure Giwercman et al. 3

Semen samples
All semen samples were collected by masturbation at the
residence or in privacy in a room at the hospital. The
participants were asked to abstain from sexual activities
for at least 2 days before collecting the samples, and to
report the actual abstinence time. The samples were
analysed for concentration, motility and morphology
according to a manual for the project based on the latest
version of the World Health Organization manual for basic
semen analysis [27]. Semen samples from Warsaw and
Kharkiv were analysed for concentration and motility at
one central hospital in each region, whereas the samples
from Greenland were analysed at the local hospital or
nursing station in nine municipalities and one settlement
scattered all over the country. The Swedish samples were
collected at the residence of the participants and the
basic semen analyses were performed in a mobile
laboratory. Briefly, the sperm concentration was deter-
mined in duplicate using an improved Neubauer
haemocytometer (Glaswarenfabrik Karl Hecht KG, Sond-
heim, Germany). The sperm motility was determined by
counting the proportion of (a) fast progressive sperms,
(b) progressive sperms, (c) nonprogressive motile sperms
and (d) immotile sperms among 100 spermatozoa within
each of two fresh drops of semen. One person performed
the semen analysis in each of the four regions. These four
persons were previously trained in a quality control
program set up specifically for this study. The median
coefficient of variation among investigators in estimating
sperm concentration and the proportion of progressive
motile sperm was 8 and 11%, respectively [28].
Smears for sperm cell morphology analysis were sent to
the Malmo University Hospital for centralized staining
and evaluation. The morphology was evaluated in at least
200 sperms per sample by two technicians from a
reference laboratory for the NAFA-ESHRE (Nordic
Association for Andrology–European Society of Human
Reproduction and Embryology) external quality control
program.
Sperm DNA integrity was assessed by the Sperm
Chromatin Structure Assay at the Section of Toxicology
and Biomedical Sciences at ENEA Casaccia Research
Centre, Italy. The coded frozen semen samples (200 ml at
– 801C) were shipped on dry ice. Briefly, 1–2� 106
thawed cells were treated with an acid–detergent
solution (0.08 N HCl, 0.15 mol/l NaCl, 0.1% Triton-X
100; pH 1.2) for 30 s and then stained with 6 mg/l of
purified Acridine Orange (Molecular Probes, Eugene,
Oregon, USA) in a phosphate-citrate buffer, pH 6.0. Cells
were analysed by a FACScan (Becton Dickinson, San
Jose, California, USA) equipped with an air-cooled argon
ion laser and standard optical filters to detect green and
red fluorescence. The percentage of abnormal sperm with
detectable DNA fragmentation index (DFI) was calcu-
lated from the DFI frequency histogram obtained from
the ratio between red and total (red + green) fluores-
cence intensity [29].
In an external quality control study that was carried out
within the framework of this project, based on blind
analysis of three aliquots from seven donors not selected
among the participants in current study, the median
intersample variability for DFI, expressed as coefficient
of variation, was 1.5%.
Determination of CB-153 and p,p0-DDE
All analyses of CB-153 and p,p0-DDE in serum were
performed at the Division of Occupational and Environ-
mental Medicine and Psychiatric Epidemiology in Lund,
Sweden, applying solid phase extraction using on-column
degradation of the lipids and analysis by gas chromato-
graphy mass spectrometry as previously described. Levels
of detection, coefficients of variation and participation in
quality control programs have been described in detail
elsewhere [30].
Determination of lipids by enzymatic methods
Serum concentrations of triglycerides and cholesterol
were determined by enzymatic methods as described
elsewhere [30]. Total lipid concentration in serum (g/l)
was calculated by the following equation:
total = 0.96 + 1.28� (triglycerides + cholesterol) [31].
Genetic analyses
Assessment of CAG and GGN repeat number was
performed as previously reported [20].
Statistical analysis
In linear regression models, we evaluated whether CAG
and GGN repeat length modified the impact of CB-153
and p,p0-DDE exposure on semen variables listed in Table
1. CAG repeat numbers were categorized into five groups
of almost equal size: < 20, 20/21, 22/23, 24 and > 24.
GGN lengths were divided into three categories: the
most common length of 23, < 23 and > 23. The
exposure levels were categorized as low and high by
dividing at the medians for the whole INUENDO male
cohort: 83 ng/g lipid for CB-153 and 569 ng/g lipid for p,p0-DDE, respectively.
The distribution of participants in the different genotype
groups by country is given in Table 2.
Model assumptions, assessed by residual analysis, were
better fulfilled for sperm concentration and total sperm
count when these variables were log transformed. In all
analyses, based on former studies in these, population
[13,17] age (continuous) and abstinence time (categor-
ized: r 2, 2 to r 4, 4 to r 6 and > 6 days) were
4 Pharmacogenetics and Genomics 2006, Vol 00 No 00

included as covariates owing to their potential confound-
ing effect.
Primarily, in order to investigate whether the data from
different centres could be pooled, a statistical test for
heterogeneity of risk, including significance test for
interaction terms between centre, AR polymorphism
and exposure level in relation to semen characteristics,
was performed in multiple linear regression models.
The heterogeneity between centres with regard to the
modifying effect of AR polymorphism on the impact of
POP exposure on semen characteristics was only statis-
tically significant for the interaction centre� CAG
category� p,p0DDE low/high (P = 0.03) with DFI as
outcome. The same was true for the interaction centre�GGN category�CB153 low/high (P = 0.048) with semen
volume as outcome. These heterogeneities, however,
were no longer statistically significant when the Inuit
cohort was excluded. Therefore, for the assessment of the
impact of CAG number on the effect of p,p0-DDE
exposure on DFI and of the GGN number on the effect
of CB153 exposure on semen volume, the analysis was
performed separately for the European cohorts and for
the Inuit cohort.
Thereafter, the potential of CAG or GGN number as
modifiers of the effect of exposure was investigated by
including interaction terms in the regression models (i.e.
CAG category� CB-153 low/high, CAG category� p,p0-DDE low/high, GGN category� CB-153 low/high and
GGN category� p,p0-DDE low/high, respectively). For
this analysis, ‘centre’ was included as a potential
confounder.
Subsequently, for each genotype subgroup the level of
particular semen parameter in the low and high exposure
groups were compared, separately for CB-153 and for p,p0-
DDE. The low and high exposure groups were compared
using linear regression and the differences were ex-
pressed both in absolute (Tables 3–6) and in relative
terms (in the Results section). As the Inuit population
differed from the other cohorts with respect to genotype
distribution and exposure level, for those associations in
which significant interaction was found, a separate
analysis was performed for the Caucasian cohorts only.
Similarly, as the inclusion criteria for Swedish fishermen
differed from the other groups, in case of significant
interaction an analysis for the partners of pregnant
women only was also performed. These calculations were
done as supplements to the above-mentioned interaction
analyses in order to exclude the heterogeneity between
the centres as the cause of statistically significant
exposure–genotype interactions.
Finally, in linear regression models, we evaluated the
effect of CAG and GGN repeat length on the semen
outcome variables.
All analyses were carried out by SPSS software (SPSS for
Windows 11.0; SPSS Inc., Chicago, Illinois, USA).
ResultsA statistically significant interaction was found between
CB-153 and CAG repeat category in relation to sperm
concentration and total sperm count (P = 0.03 and 0.01,
respectively; Table 3). When ‘centre’ was included as a
covariate, these interactions were still statistically sig-
nificant (P = 0.049 and 0.03, respectively). For
CAG < 20, the ‘high CB-153’ group presented with a
sperm concentration being 35% [95% confidence interval
(CI): 4.4; 79%] lower and the total sperm count being
42% (95% CI: 8.9; 91%) lower than in the ‘low CB-153’
category (Fig. 1).
Table 2 Distribution of study participants according to the levels of CB-153 and p,p0-DDE, androgen receptor genotype and place of origin
n (%)
Greenland Warsaw, Poland Swedish fishermen Kharkiv, Ukraine All
CB-153Low 20 (11) 165 (99) 18 (10) 121 (82) 324 (48)High 168 (89) 2 (1.0) 160 (90) 26 (18) 356 (52)
p,p0-DDELow 87 (46) 103 (62) 156 (88) 23 (16) 369 (54)High 101 (54) 64 (38) 22 (11) 124 (84) 311 (46)
CAG number< 20 17 (9) 35 (21) 34 (19) 26 (18) 112 (17)20/21 21 (11) 39 (23) 48 (27) 37 (25) 145 (21)22/23 38 (20) 45 (27) 40 (23) 38 (26) 161 (24)24 77 (41) 12 (7.0) 13 (7.0) 8 (5.0) 110 (16)> 24 35 (19) 36 (22) 43 (24) 38 (26) 152 (22)
GGN number< 23 8 (4.0) 13 (8.0) 25 (14) 16 (11) 62 (9.0)23 161 (86) 82 (49) 95 (53) 86 (59) 424 (62)> 23 19 (10) 72 (43) 58 (33) 45 (31) 194 (29)
CB-153; 2,2, 4,45,5-hexachlorobiphenyl; p,p0-DDE, dichlorodiphenyl dichloroethene.
Androgen receptor polymorphisms and POP exposure Giwercman et al. 5

When separate analyses were performed for the Cauca-
sian cohorts only, the ‘low’ and ‘high’ CB-153 exposure
groups still differed with respect to total sperm counts for
CAG < 20 (relative mean difference: 41%; 95% CI: 5.3;
62%) as well as sperm concentration (relative mean
difference: 36%; 95% CI: 1.0; 58%). For the cohorts of
partners of pregnant women (except Sweden), the
corresponding differences were statistically significant
for total sperm number (relative mean difference: 47%;
95% CI: 10; 68%), whereas the difference with regard to
sperm concentration was borderline statistically signifi-
cant (relative mean difference: 36%; 95% CI: – 3.6%;
61%).
For p,p0-DDE, among the Caucasians, a significant
interaction with CAG number was found in relation to
DFI (P = 0.01 without and P = 0.006 with ‘centre’ as a
covariate) (Table 4). The ‘high p,p0-DDE’ exposure
group, as compared with the ‘low p,p0-DDE’ group,
presented with 43% (95% CI: 9.6; 78%) higher DFI for
CAG < 20 and 38% (95% CI: 7.6; 68%) higher DFI for
CAG 20/21, respectively. In the Inuit cohort, the
interaction between CAG category and the effect of
p,p0-DDE level on DFI was not statistically significant
(P = 0.64).
After exclusion of the Swedish fishermen in the CAG
category 20/21, the DFI was significantly higher for ‘high’
p,p0-DDE than for ‘low’ p,p0-DDE (relative mean
difference: 48%; 95% CI: 16; 79%). A similar trend,
without reaching the level of statistical significance
(relative mean difference: 17%; 95% CI: – 13%; 47%),
was seen for CAG < 20.
Furthermore, despite the lack of a statistically significant
interaction between exposure and genotype, a signifi-
cantly lower percentage of progressively motile sperms
was found in ‘high CB-153’ for CAG < 20 (mean
difference 11%; 95% CI: 2.6; 19%) and in ‘high p,p0-
Table 3 Interaction between CB-153 levels (low/high) and categorized CAG repeat length in relation to semen characteristics
CAG repeat length
CB-153 < 20 20/21 22/23 24 > 24 P value for interaction
Semen volume (ml)Low 3.6 3.8 3.4 4.1 3.7 0.67High 3.5 3.9 3.5 3.6 3.5Median (95% CI)a 0.17 ( – 0.55; 0.88) – 0.070 ( – 0.69; 0.55) – 0.094 ( – 0.69; 0.50) 0.48 ( – 0.37; 1.3) 0.24 ( – 0.34; 0.83)
Sperm concentration (106/ml)Low 60b 55 47 47 54 0.03High 38b 60 61 49 62Median (95% CI)a 21 (4.0; 73) – 5.1 ( – 18; 14) – 14 ( – 20; 1.8) – 2.4 ( – 18; 22) – 7.9 ( – 19; 9.6)Sperm number (106)Low 190b 190 140 170 170 0.01High 110b 200 180 160 180Median (95% CI)a 79 (29; 300) – 14 ( – 65; 60) – 42 ( – 64; 10) 14 ( – 56; 130) – 18 ( – 58; 43)Progressively motile (%)Low 60b 59 56 53 57 0.15High 49b 60 55 55 57Median (95% CI)a 11 (2.6; 19) – 0.93 ( – 8.3; 6.4) 0.40 ( – 6.7; 7.4) – 2.2 (12; 7.7) – 0.038 ( – 7.0; 6.9)Normal morphology (%)Low 7.6 7.4 6.8 7.2 7.2 0.05High 6.1 8.9 7.7 6.5 7.9Median (95% CI)a 1.5 ( – 0.15; 3.1) – 1,4 ( – 2.8; 0.036) – 0.82 ( – 2.2; 0.56) 0.67 ( – 1.3; 2.7) – 0.62 ( – 2.0; 0.74)DFI (%)Low 16 14 13 13 15 0.89High 15 14 14 12 14Median (95% CI)a 0.91 ( – 2.9; 4.7) 0.41 ( – 2.9; 3.7) – 0.90 ( – 4.1; 2.3) 0.42 ( – 4.1; 4.9) 1.2 ( – 1.9; 4.4)
CB-153; 2,2, 4,45,5-hexachlorobiphenyl; CI, confidence interval; DFI, DNA fragmentation index.aMean difference (low – high) and 95% CI of this difference. As the values for concentration and total sperm count are back transformed from log10 values, thecalculations of mean difference (95% CI) are based on the ratios between low and high.bP < 0.05 for difference between low and high.The results, adjusted for the effect of age and abstinence time, are expressed as means and the difference between the means (95% CI).
Fig. 1
CAG number
0
50
100
150
200
250
<20 20/21 22/23 24 >24
Tota
l spe
rm c
ount
(×10
6 )
P<0.05
Mean total sperm counts in low (white bars) and high (black bars) 2,2,4,45,5-hexachlorobiphenyl (CB-153) exposure groups in relation toCAG repeat length. Statistically significant differences for low and highexposure are indicated in the figure.
6 Pharmacogenetics and Genomics 2006, Vol 00 No 00

DDE’ for CAG 20/21 (mean difference 7.8%; 95% CI:
0.71; 14%) and 21/22 (mean difference 12%; 95% CI: 5.1;
19%; Table 3).
No significant interaction between POP levels and GGN
category in relation to sperm characteristics was found
(Tables 5 and 6). For GGN = 23, however, significantly
Table 4 Interaction between p,p0-DDE levels (low/high) and categorized CAG repeat length in relation to semen characteristics
CAG repeat length
p,p0-DDE < 20 20/21 22/23 24 > 24 P value for interaction
Semen volume (ml)Low 3.5 4.0 3.4 4.0 3.7 0.60High 3.6 3.7 3.6 3.5 3.5Median (95% CI)a – 0.19 ( – 0.89; 0.51) 0.26 ( – 0.35; 0.86) – 0.16 ( – 0.73; 0.42) 0.46 ( – 1.1; 0.23) 0.16 ( – 0.42; 0.73)
Sperm concentration (106/ml)Low 49 55 56 54 55 0.70High 47 59 51 45 62Median (95% CI)a 2.2 ( – 13; 24) – 4.4 ( – 18; 15) 5.8 ( – 9.6; 28) 8.6 ( – 8.8; 37) – 6.2 ( – 18; 11)
Sperm number (106)Low 150 200 170 200 170 0.63High 140 190 160 140 180Median (95% CI)a 8.7 ( – 42; 85) 8.6 ( – 51; 94) 8.7 ( – 39; 77) 60 ( – 5.1; 220) – 5.7 ( – 52; 58)
Progressively motile (%)Low 54 63b 61b 58 57 0.62High 54 55b 49b 51 56Median (95% CI)a – 0.23 ( – 8.37.9) 7.8 (0.71; 14) 12 (5.1; 19) 6.3 ( – 1.7; 14) 1.2 ( – 5.5; 7.9)
Normal morphology (%)Low 6.8 8.3 7.8 6.7 7.4 0.11High 6.7 8.0 6.6 6.5 7.7Median (95% CI)a 0.075 ( – 1.5; 1.7) 0.32 ( – 1.1; 1.7) 1.1 ( – 0.19; 2.5) 0.26 ( – 1.3; 1.9) – 0.37 ( – 1.7; 0.96)
DFI (%)c
Low 13b 13b 15 16 18 0.01High 19b 18b 14 15 15Median (95% CI)b – 5.8 ( – 10; – 1.2) – 4.8 ( – 8.7; – 0.98) 1.9 ( – 2.1; 6.0) 0.98 ( – 6.9; 8.8) 2.3 ( – 1.6; 6.2)
p,p0-DDE, dichlorodiphenyl dichloroethene; CI, confidence interval; DFI, DNA fragmentation index.aMean difference (low – high) and 95% CI of this difference. As the values for concentration and total sperm count are back transformed from log10 values, thecalculations of mean difference (95% CI) are based on the ratios between low and high.bP < 0.05 for difference between low and high.cBased on the three Caucasian cohorts only.
Table 5 Interaction between CB-153 levels (low/high) and categorized GGN repeat length in relation to semen characteristics
GGN repeat length
CB-153 23 < 23 > 23 P value for interaction
Semen volume (ml)c
Low 3.7 3.9 3.3 0.81High 3.8 3.9 3.6Median (95% CI)a,b – 0.074 ( – 0. 65; 0.50) 0.0095 ( – 0.99; 1.0) – 0.23 ( – 0.90; 0.44)
Sperm concentration (106/ml)Low 52 53 54 0.73High 53 62 54Median (95% CI)a – 1.6 ( – 11; 29) – 8.8 ( – 29; 23) – 0.090 ( – 14; 18)
Sperm number (106)Low 170 190 160 0.69High 160 210 160Median (95% CI)a 3.4 ( – 28; 43) – 24 ( – 100; 103) – 0.083 ( – 45; 63)
Progressively motile (%)Low 55 64 58 0.61High 55 60 55Median (95% CI)a 0.33 ( – 4.1; 4.7) 3.8 ( – 7.0; 15) 3.5 ( – 3.2; 10)
Normal morphology (%)Low 7.1 8.6 7.0 0.31High 7.1 8.1 8.2Median (95% CI)a 0.068 ( – 0.80; 0.93) 0.50 ( – 1.6; 2.6) – 1.2 ( – 2.6; 0.12)
DFI (%)Low 15 15 13 0.66High 14 14 13Median (95% CI)a 1.3 ( – 0.65; 3.3) 1.2 ( – 3.6; 6.1) – 0.26 ( – 3.3; 2.8)
CB-153; 2,2, 4,45,5-hexachlorobiphenyl; CI, confidence interval; DFI, DNA fragmentation index.aMean difference (low – high) and 95% confidence interval (CI) of this difference. As the values for concentration and total sperm count are back transformed from log10
values, the calculations of mean difference (95% CI) are based on the ratios between low and high.bP < 0.05 for difference between low and high.cBased on the three Caucasian cohorts only.The results, adjusted for the effect of age and abstinence time, are expressed as means and the difference between the means (95% CI).
Androgen receptor polymorphisms and POP exposure Giwercman et al. 7
higher (mean difference 6.7%; 95% CI: 2.6; 11%)
percentage progressively motile sperms was found for
‘low’ vs. ‘high’ CB-153 exposure group.
No statistically significant association was found between
the categorized CAG repeat number and any of the
semen parameters tested, except for the proportion of
morphologically normal cells (P = 0.02). This association
was due to a significantly higher percentage of normal
sperms for the CAG length of 20/21 (mean: 8.2%) than
for the CAG length of < 20 (mean difference: 1.4%; 95%
CI: 0.32; 2.7%) and CAG length of 24 (mean difference:
1.7%; 95% CI: 0.62; 2.7%), respectively.
For GGN categories, statistically significant differences
were found for morphology (P = 0.05) and proportion of
progressively motile sperms (P = 0.03). In both cases, the
GGN < 23 group diverged from GGN = 23 by presenting
with more sperms with normal morphology: (mean: 8.4%
vs. 7.0%; 95% CI of difference: 0.23; 2.4%) and
progressive motility (mean: 62 vs. 55%; 95% CI of
difference: 1.7; 12%).
DiscussionThe main objective of this study was to investigate
whether polymorphisms in the AR gene may modify the
effect of POP exposure on semen characteristics. Our
results indicate that the variation in the length CAG
repeats may play a role in the individual sensitivity to the
adverse effects of POP exposure on the male reproduc-
tive system. We found a statistically significant interac-
tion between CAG number and CB-153 levels in relation
to sperm concentration and total sperm number and for
p,p0-DDE levels in relation to sperm DNA integrity. This
is, to our knowledge, the first study showing a gene–
environment interaction in relation to semen character-
istics.
The most consistent finding of the study was that the
difference in semen characteristics between low and high
POP exposure groups was most pronounced in men with
short CAG tracts. In a recently published study, based on
the same populations, no significant associations between
CB-153 or p,p0-DDE levels and sperm concentration and
total sperm count were found [17]. The proportion of
motile sperms, however, was found to decrease with
increasing CB-153 levels. In the current work, we found
that men with CAG < 20 had a 35% reduction in sperm
concentration and 42% reduction in total sperm number
when the group with CB-153 concentration above the
median was compared with those with CB-153 below the
50th percentile. In this CAG group, high CB-153
exposure was also associated with a 11% reduction in
the proportion of progressively motile sperms.
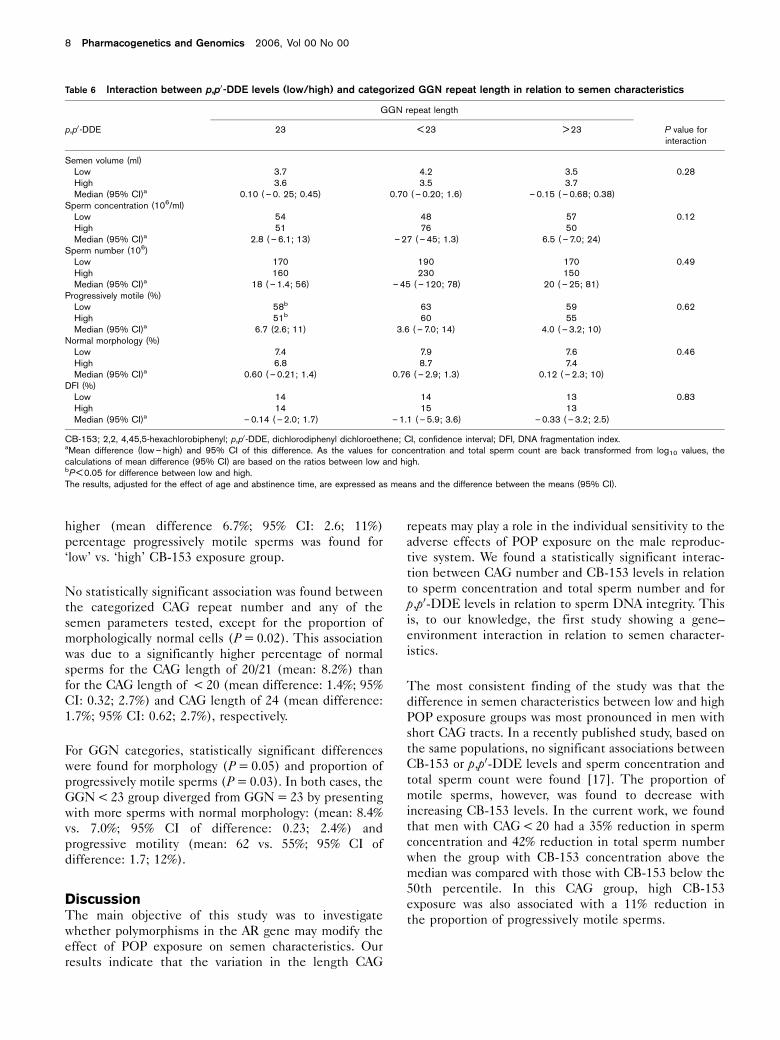
Table 6 Interaction between p,p0-DDE levels (low/high) and categorized GGN repeat length in relation to semen characteristics
GGN repeat length
p,p0-DDE 23 < 23 > 23 P value forinteraction
Semen volume (ml)Low 3.7 4.2 3.5 0.28High 3.6 3.5 3.7Median (95% CI)a 0.10 ( – 0. 25; 0.45) 0.70 ( – 0.20; 1.6) – 0.15 ( – 0.68; 0.38)
Sperm concentration (106/ml)Low 54 48 57 0.12High 51 76 50Median (95% CI)a 2.8 ( – 6.1; 13) – 27 ( – 45; 1.3) 6.5 ( – 7.0; 24)
Sperm number (106)Low 170 190 170 0.49High 160 230 150Median (95% CI)a 18 ( – 1.4; 56) – 45 ( – 120; 78) 20 ( – 25; 81)
Progressively motile (%)Low 58b 63 59 0.62High 51b 60 55Median (95% CI)a 6.7 (2.6; 11) 3.6 ( – 7.0; 14) 4.0 ( – 3.2; 10)
Normal morphology (%)Low 7.4 7.9 7.6 0.46High 6.8 8.7 7.4Median (95% CI)a 0.60 ( – 0.21; 1.4) 0.76 ( – 2.9; 1.3) 0.12 ( – 2.3; 10)
DFI (%)Low 14 14 13 0.83High 14 15 13Median (95% CI)a – 0.14 ( – 2.0; 1.7) – 1.1 ( – 5.9; 3.6) – 0.33 ( – 3.2; 2.5)
CB-153; 2,2, 4,45,5-hexachlorobiphenyl; p,p0-DDE, dichlorodiphenyl dichloroethene; CI, confidence interval; DFI, DNA fragmentation index.aMean difference (low – high) and 95% CI of this difference. As the values for concentration and total sperm count are back transformed from log10 values, thecalculations of mean difference (95% CI) are based on the ratios between low and high.bP < 0.05 for difference between low and high.The results, adjusted for the effect of age and abstinence time, are expressed as means and the difference between the means (95% CI).
8 Pharmacogenetics and Genomics 2006, Vol 00 No 00

No statistically significant interactions were found in
relation to the other AR polymorphism – the GGN
repeats. Although the proportion of progressively motile
spermatozoa was significantly higher for the ‘low’ CB-163
group than for the ‘high’ CB-163 group when GGN was
23, this finding is probably due to the fact that GGN = 23
is the most common genotype for this repetitive
sequence and generally a negative association between
CB-153 levels and sperm motility was found in the
INUENDO population [17]
Spermatogenesis is a highly androgen-dependent process
[32], the concentration of testosterone being more than
100 times higher in the testis than in serum [33]. An anti-
androgenic effect of p,p0-DDE [4] and of CB-153 has
been reported [4,34]. In a reporter in-vitro assay, the
highest anti-androgenic activity was found in the Kharkiv
cohort presenting with high p,p0-DDE levels (T. Kruger,
P.S. Hjelmborg, B.A.G. Jonsson, L. Hagmar, A. Giwerc-
man, G.-C. Manicardi et al., in preparation). This study
does not allow a clarification of the mechanism by which
the CAG repeat length might modify the effect of POP
on semen characteristics. As the three-dimensional
structure of the receptor is affected by the length of
the CAG stretch, however, one could hypothesize that
the strength of the POP binding, or of any necessary co-
factor, is regulated by CAG number. Although we
measured levels of CB-153 and p,p0-DDE, the effect
observed might also be due to the interaction of other
POPs with the AR. CB-153 concentration was reported to
be highly correlated with the total PCB concentration
[9,35], the estimated TEQ from PCB and the total POP-
derived TEQ [11], respectively. As some of these
compounds are known to bind to the arylhydrocarbon
receptor, short CAG lengths might also modify the
interaction between the arylhydrocarbon receptor and
the AR, rather than directly affect the binding of POP to
the latter receptor [36].
Short CAG repeats also seemed to modify the association
between p,p0-DDE and sperm DNA integrity. In the
groups with CAG lengths shorter than the median, DFI
was 40% higher in highly exposed men than in those with
low p,p0-DDE exposure. Although sperm DNA strand
breaks can be caused by post-testicular events, topoi-
somerase II, which is an androgen-dependent DNA
reparation enzyme, also plays an important role for
ensuring sperm DNA integrity [37]. Therefore, it seems
plausible that chemicals interfering with AR function may
also have an impact on sperm number and DNA integrity.
In a recent study on the association between POP
exposure and DFI [13], this association was restricted to
the Caucasian men. The Inuit population was found to
differ significantly from the European men with respect
to sperm DNA integrity, possibly owing to differences in
the genetic background and/or in dietary habits.
Developmental and reproductive disorders are critical
toxic effects caused by POP and most of these effects are
believed to be the result of impaired steroidogenesis and/
or steroid action [1]. The finding of a genetic modifier of
the environmental effects on male reproductive organs
has important implications, not only from a biological
point of view but also from a public health point of view.
In order to prevent any further increases in disorders of
male reproductive organs, it is important to develop tools
for identifying individuals susceptible to adverse effects
of environmental and lifestyle-related factors.
Finally, taking genetically determined susceptibility into
account is also important when planning epidemiological
studies. With respect to this, the INUENDO study failed
to show any negative impact of POP exposure on sperm
numbers [17], whereas inclusion of the AR genotype
allowed the identification of such an effect in a subset of
the same study population.
When evaluating the results of this study, the issue of
potential selection bias needs to be considered. Except
for the Inuit population, the participation rate in the
semen study, in similarity with other similar surveys, was
rather low. With respect to the Swedish fishermen, the
age distributions and the mean number of children were
very similar among the participants and the non-
participants [16]. In the three remaining cohorts, it was
found that time to pregnancy did not differ between
those who delivered semen for analysis and those who did
not [14]. Furthermore, it is difficult to imagine how the
interaction between genotype and environmental expo-
sure should affect the participation rate. Therefore, we
do not consider that selection bias is of major concern.
As multiple comparisons have been performed, in order to
minimize the risk of mass significance, we have focused
on the associations in which a statistically significant
interaction between exposure levels and CAG length was
found. In total, the P value was below 0.05 for three of 12
such tests, or for two of 10, if sperm concentration and
total sperm count were considered as markers of the same
biological mechanism. Although these proportions were
higher than expected if our findings had been due to
chance, mass significance as an explanation of our results
cannot be completely ruled out.
It could be asked whether the results might be
confounded by significant differences in distribution of
AR genotypes and exposure profiles, mainly between the
Inuit cohort and the Caucasians. Significant interaction
between genotype and exposure reported here were not
affected by inclusion of centre as potential confounder.
Additionally, for CAG < 20; the differences between the
groups with ‘high’ and ‘low’ CB-153, with respect to
sperm concentration and total number, persisted even
Androgen receptor polymorphisms and POP exposure Giwercman et al. 9

when the Inuits were excluded from the analysis. The
same was true when the Swedish fishermen, who differed
from the other cohorts as these men were not included as
partners of pregnant women, were not included in the
analysis. Furthermore, no significant interaction between
centre, AR genotype group and exposure level was found
for any of the reproductive endpoints, except for DFI in
relation to p,p0-DDE exposure. As, in the latter case, a
separate analysis was performed for the Caucasians and
the Inuit population, we believe that merging of all
groups was justified in the remaining cases and the
interactions we have observed are due to the modifying
effect of the CAG repeat length on the impact of POP
exposure on semen parameters.
The main focus of this study was to assess the impact of
AR polymorphisms as modifiers of the effect of POP on
semen parameters. It is notable that CAG or GGN
number did not, per se, seem to have any strong
associations with sperm concentration or total number.
Previous studies showed conflicting results regarding the
link between CAG length and sperm number [10,22].
Provided the gene–environment interaction found by us,
one could speculate whether the interpopulation differ-
ences in POP exposure levels might contribute to the
diverging results of studies on CAG repeats and
reproductive characteristics. Thus, if short CAG repeat
renders an individual more susceptible to the effect of at
least some POPs, in highly exposed populations this
effect might counteract the per se beneficial effect of low
CAG number on sperm production.
We found a slightly higher percentage of morphologically
normal spermatozoa for CAG = 20 or 21 and for
GGN < 23. The latter group also presented with a higher
percentage of progressively motile sperms. The biological
significance of these findings, however, is difficult to
interpret as they do not fit with previous data indicating
superior AR activity for the shortest CAG categories and
for GGN = 23 as compared with shorter and longer GGNs
[25,26,38].
In conclusion, our data might indicate that the AR CAG
repeat length influences the impact of POP on sperm
characteristics as sperm number and DNA integrity. More
studies, however, are needed in order to establish
whether the AR or other genetic polymorphisms modify
the effect of endocrine disrupters on male reproductive
function and to enlighten possible mechanisms behind
such gene–environment interaction.
AcknowledgementsProfessor Lars Hagmar, Division of Occupational and
Environmental Medicine and Psychiatric Epidemiology,
Lund University, Lund University Hospital, was one of
the key persons behind the INUENDO project and
contributed significantly to the current manuscript. Lars
Hagmar, however, passed away during the time when the
manuscript was under the review process.
References1 Toppari J, Larsen JC, Christiansen P, Giwercman A, Grandjean P, Guillette LJ
Jr, et al. Male reproductive health and environmental xenoestrogens. EnvironHealth Perspect 1996; 104:741–803.
2 Longnecker MP, Rogan WJ, Lucier G. The human health effects of DDT(dichlorodiphenyltrichloroethane) and PCBS (polychlorinated biphenyls) andan overview of organochlorines in public health. Annu Rev Public Health1997; 18:211–244.
3 Andersen HR, Vinggaard AM, Rasmussen TH, Gjermandsen IM, Bonefeld-Jorgensen EC. Effects of currently used pesticides in assays forestrogenicity, androgenicity, and aromatase activity in vitro. Toxicol ApplPharmacol 2002; 179:1–12.
4 Bonefeld-Jorgensen EC, Andersen HR, Rasmussen TH, Vinggaard AM.Effect of highly bioaccumulated polychlorinated biphenyl congeners onestrogen and androgen receptor activity. Toxicology 2001; 158:141–153.
5 Danzo BJ. Environmental xenobiotics may disrupt normal endocrine functionby interfering with the binding of physiological ligands to steroid receptorsand binding proteins. Environ Health Perspect 1997; 105:294–301.
6 Kelce WR, Stone CR, Laws SC, Gray LE, Kemppainen JA, Wilson EM.Persistent DDT metabolite p,p0-DDE is a potent androgen receptorantagonist. Nature 1995; 375:581–585.
7 Giwercman YL, Kleist KE, Giwercman A, Giwercman C, Toft G, Bonde JP, etal. Remarkably low incidence of hypospadias in Greenland despite highexposure to endocrine disupters; possible effect of androgen receptorgenotype. Pharmacogenet Genomics 2006; 16:375–377.
8 Skakkebck NE, Rajpert-De Meyts E, Main KM. Testicular dysgenesissyndrome: an increasingly common developmental disorder withenvironmental aspects. Hum Reprod 2001; 16:972–978.
9 Glynn AW, Wolk A, Aune M, Atuma S, Zettermark S, Maehle-Schmid M, et al.Serum concentrations of organochlorines in men: a search for markers ofexposure. Sci Total Environ 2000; 263:197–208.
10 Rajpert-De Meyts E, Leffers H, Petersen JH, Andersen A-G, Carlsen E,Jørgensen N, et al. CAG repeat length in androgen receptor gene andreproductive variables in fertile and infertile men. Lancet 2002; 359:44–46.
11 Gladen BC, Longnecker MP, Schecter AJ. Correlations amongpolychlorinated biphenyls, dioxins, and furans in humans. Am J Ind Med1999; 35:15–20.
12 Sjodin A, Hagmar L, Klasson-Wehler E, Bjork J, Bergman A. Influence of theconsumption of fatty Baltic Sea fish on plasma levels of halogenatedenvironmental contaminants in Latvian and Swedish men. Environ HealthPerspect 2002; 108:1035–1041.
13 Spano M, Toft G, Hagmar L, Eleuteri P, Rescia M, Rignell-Hydbom A, et al.Exposure to PCB and p,p0-DDE in European and Inuit populations: impacton human sperm chromatin integrity. Hum Reprod 2005; 20:3488–3499.
14 Toft G, Axmon A, Giwercman A, Thulstrup AM, Rignell-Hydbom A, PedersenHS, et al. Fertility in four regions spanning large contrasts in serum levels ofwidespread persistent organochlorines: a cross-sectional study. EnvironHealth 2005; 4:26.
15 Rignell-Hydbom A, Rylander L, Giwercman A, Jonsson BA, Lindh C, EleuteriP, et al. Exposure to PCBs and p,p0-DDE and human sperm chromatinintegrity. Environ Health Perspect 2005; 113:175–179.
16 Rignell-Hydbom A, Rylander L, Giwercman A, Jonsson BA, Nilsson-Ehle P,Hagmar L. Exposure to CB-153 and p,p0-DDE and male reproductivefunction. Hum Reprod 2004; 19:2066–2075.
17 Toft G, Rignell-Hydbom A, Tyrkiel E, Shvets M, Giwercman A, Lindh CH, etal. Semen quality and exposure to persistent organochlorine pollutants in anInuit and three European cohorts. Epidemiology 2006; 17:450–458.
18 Gao T, Marcelli M, McPhaul MJ. Transcriptional activation and transientexpression of the human androgen receptor. J Steroid Biochem Mol Biol1996; 59:9–20.
19 Tut TG, Ghadessy FJ, Trifiro MA, Pinsky L, Young EL. Long polyglutaminetracts in the androgen receptor are associated with reduced trans-activation,impaired sperm production, and male infertility. J Clin Endocrinol Metab1997; 82:3777–3782.
20 Lundin KB, Giwercman A, Richthoff J, Abrahamsson P-A, Giwercman YL. Noassociation between mutations in the human androgen receptor GGNrepeat and inter-sex conditions. Mol Hum Reprod 2003; 9:375–379.
21 MacLean HE, Warne GL, Zajac JD. Spinal and bulbar atrophy: androgenreceptor dysfunction caused by a trinucleotide repeat expansion. J NeurolSci 1996; 135:149–157.
10 Pharmacogenetics and Genomics 2006, Vol 00 No 00

22 von Eckardstein S, Syska A, Gromoll J, Kamischke A, Simoni M, NieschlagE. Inverse correlation between sperm concentration and number ofandrogen receptor CAG repeats in normal men. J Clin Endocrinol Metab2001; 86:2585–2590.
23 Zitzmann M, Brune M, Kornmann B, Gromoll J, Junker R, Nieschlag E. TheCAG repeat polymorphism in the androgen receptor gene affects bonedensity and bone metabolism in healthy males. Clin Endocrinol 2001;55:649–657.
24 Aschim EL, Nordenskjold A, Giwercman A, Lundin KB, Ruhayel Y, HaugenTB, et al. Linkage between cryptorchidism, hypospadias, and GGN repeatlength in the androgen receptor gene. J Clin Endocrinol Metab 2004;89:5105–5109.
25 Hillmer AM, Hanneken S, Ritzmann S, Becker T, Freudenberg J,Brockschmidt FF, et al. Genetic variation in the human androgen receptorgene is the major determinant of common early-onset androgeneticalopecia. Am J Hum Genet 2005; 77:140–148.
26 Ruhayel Y, Lundin K, Giwercman Y, Hallden C, Willen M, Giwercman A.Androgen receptor gene GGN and CAG polymorphisms among severelyoligozoospermic and azoospermic Swedish men. Hum Reprod 2004;19:2076–2083.
27 World Health Organization. WHO laboratory manual for the examination ofhuman semen and sperm-cervical mucus interaction. 3rd ed. Cambridge:Cambridge University Press; 1999.
28 Toft G, Rignell-Hydbom A, Tyrkiel E, Shvets M, Giwercman A. Quality controlworkshops in standardization of sperm concentration and motilityassessment in multicentre studies. Int J Androl 2005; 28:144–149.
29 Evenson DP, Larson KL, Jost LK. Sperm chromatin structure assay: itsclinical use for detecting sperm DNA fragmentation in male infertility andcomparisons with other techniques. J Androl 2002; 23:25–43.
30 Richthoff J, Rylander L, Jonsson BA, Akesson H, Hagmar L, Nilsson-Ehle P,et al. Serum levels of 2,20 ,4,40 ,5,50-hexachlorobiphenyl (CB-153) in relationto markers of reproductive function in young males from the generalSwedish population. Environ Health Perspect 2003; 111:409–413.
31 Rylander L, Nilsson-Ehle P, Hagmar L. A simplified precise method foradjusting serum levels of persistent organohalogen pollutants to total serumlipids. Chemosphere 2006; 62:333–336.
32 ElZanaty S, Giwercman YL, Giwercman A. Significant impact of 5alpha-reductase type 2 polymorphisms on sperm concentration and motility. Int JAndrol 2006; 29:414–420.
33 Maddocks S, Hargreave TB, Reddie K, Fraser HM, Kerr JB, Sharpe RM.Intratesticular hormone levels and the route of secretion of hormones fromthe testis of the rat, guinea pig, monkey and human. Int J Androl 1993;16:272–278.
34 Schrader TJ, Cooke GM. Effects of Aroclors and individual PCB congenerson activation of the human androgen receptor in vitro. Reprod Toxicol 2003;17:15–23.
35 Grimvall E, Rylander L, Nilsson-Ehle P, Nilsson U, Stromberg U, Hagmar L, etal. Monitoring of polychlorinated biphenyls in human blood plasma:methodological developments and influence of age lactation and fishconsumption. Arch Environ Contam Toxicol 1997; 32:329–336.
36 Morrow D, Qin C, Smith R 3rd, Safe S. Aryl hydrocarbon receptor-mediatedinhibition of LNCaP prostate cancer cell growth and hormone-inducedtransactivation. J Steroid Biochem Mol Biol 2004; 88:27–36.
37 Bakshi RP, Galande S, Bali P, Dighe R, Muniyappa K. Developmental andhormonal regulation of type II DNA topoisomerase in rat testis. J MolEndocrinol 2001; 26:193–206.
38 Lundin KB, Giwercman YL, Rylander L, Hagmar L, Giwercman A. Androgenreceptor gene GGN repeat length and reproductive characteristics in youngSwedish men. Eur J Endocrinol 2006; 155:347–354.
Androgen receptor polymorphisms and POP exposure Giwercman et al. 11

AUTHOR QUERY FORM
LIPPINCOTTWILLIAMS and WILKINS
QUERIES AND / OR REMARKS
JOURNAL NAME: FPCARTICLE NO: 200712
QUERY NO Details Required Author's Response
.No Queries
Copyright © 2022 FDOKUMEN




















