
THEORETICAL STUDY OF A BIOFILM LIFE CYCLE: GROWTH, NUTRIENT DEPLETION AND DETACHMENT
12
December 20, 2005 12:45 Proceedings Trim Size: 9in x 6in BIOMAT2005 THEORETICAL STUDY OF A BIOFILM LIFE CYCLE: GROWTH, NUTRIENT DEPLETION AND DETACHMENT * G. DOMINGUEZ-ZACAR´ ıAS, E. LUNA AND J. X. VELASCO-HERN ´ ANDEZ Programa de Investigaci´ on en Matem´ aticasAplicadasy Computaci´on Instituto Mexicano del Petr´ oleo, M´ exico, DF 07730 MEXICO Email: [email protected]; [email protected]; [email protected] We present the results of a theoretical investigation, based upon experimental evidence gathered from the existing recent literature, on the interaction of fluid flow and biofilm internal structure, centering on the processes of detachment, spatial structure of microbial communities, and the diffusion and convection of nutrients. The biofilm is viewed as a porous medium with variable porosity, tortuosity and permeability. Our model is a two dimensional approximation to biofilm dynamics and it is capable of reproducing biofilm internal heterogeneities, biofilm surface behavior, nutrient penetration and biofilm critical point of rupture or detachment. 1. Introduction Biofilm detachment is one of the least understood processes in the life cycle of a biofilm. There is a variety of hypothesis 1 formulated to explain it, among them are we can count the existence of matrix-degrading enzymes, appearance of gas bubbles, nutrient levels and microbial growth, quorum sensing among others. To understand biofilm physical and chemical characteristics one must consider mutual influences between biofilm structure and processes. Nu- trient transport, metabolic residue accumulation, and limited diffusion are affected by biofilm structure. However, as reported by Bayebal et al. 2 , this influence is largely hypothetical and awaits elucidation. To this end the development of tools to quantify biofilm structure characteristics and to correlate those with processes is urgently needed. According to several authors 2 biofilm development is associated with : i) structure and function * This work was supported by grant imp d.00330 1
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of THEORETICAL STUDY OF A BIOFILM LIFE CYCLE: GROWTH, NUTRIENT DEPLETION AND DETACHMENT

December 20, 2005 12:45 Proceedings Trim Size: 9in x 6in BIOMAT2005
THEORETICAL STUDY OF A BIOFILM LIFE CYCLE:GROWTH, NUTRIENT DEPLETION AND DETACHMENT∗
G. DOMINGUEZ-ZACARıAS, E. LUNA
AND J. X. VELASCO-HERNANDEZ
Programa de Investigacion en Matematicas Aplicadas y ComputacionInstituto Mexicano del Petroleo,
Mexico, DF 07730 MEXICOEmail: [email protected]; [email protected]; [email protected]
We present the results of a theoretical investigation, based upon experimental
evidence gathered from the existing recent literature, on the interactionof fluid flowand biofilm internal structure, centering on the processes of detachment, spatial
structure of microbial communities, and the diffusion and convection of nutrients.The biofilm is viewed as a porous medium with variable porosity, tortuosity and
permeability. Our model is a two dimensional approximation to biofilm dynamicsand it is capable of reproducing biofilm internal heterogeneities, biofilm surface
behavior, nutrient penetration and biofilm critical point of rupture or detachment.
1. Introduction
Biofilm detachment is one of the least understood processes in the life cycleof a biofilm. There is a variety of hypothesis1 formulated to explain it,among them are we can count the existence of matrix-degrading enzymes,appearance of gas bubbles, nutrient levels and microbial growth, quorumsensing among others.
To understand biofilm physical and chemical characteristics one mustconsider mutual influences between biofilm structure and processes. Nu-trient transport, metabolic residue accumulation, and limited diffusion areaffected by biofilm structure. However, as reported by Bayebal et al.2,this influence is largely hypothetical and awaits elucidation. To this endthe development of tools to quantify biofilm structure characteristics andto correlate those with processes is urgently needed. According to severalauthors2 biofilm development is associated with : i) structure and function
∗This work was supported by grant imp d.00330
1

December 20, 2005 12:45 Proceedings Trim Size: 9in x 6in BIOMAT2005
2
that allows survival on surfaces, ii) efficient transport of nutrients to thewhole biofilm body.
The analysis of biofilm structure fall into several categories: i) com-parative studies, ii) reproducibility of structure, iii) monitoring temporalvariations in structure, iv) testing the effect of substances on structure, v)quantifying effect of environmental factors on structure, and vi) parameterestimation of structure characteristics for use in models.
Mai-Prochnow et al.3 explored the role that cell death plays in the struc-ture of biofilms. They show that some biofilm types have a reproduciblepattern of cell death that, in the long term, may play an important rolein biofilm dispersion. These authors support the view that cell death in-side biofilms may be a widespread phenomenon of importance for biofilmdispersal. However little is known about this mechanisms. One possibleexplanation has been put forward by Hunt et al.1 who investigated the roleof nutrient starvation in biofilm detachment. According to this author lo-calized nutrient depletion induces starvation in subsets of cells that, givenenough time, detach from the biofilm main structure.
Of the many modeling approaches4–14 available to model the interactionbetween biofilm structure and the surrounding liquid medium, we haveused a hybrid model where conservation equations are coupled to a cellularautomaton (CA) that simulates the growth of the bacterial consortia15.
A biofilm is an assemblage of surface-associated microbial cells enclosedin an extracellular polymeric matrix16. Biofilms play important roles inmany areas of applied science and technology such as oil enhanced recov-ery and transportation, pharmaceutical research, food processing, medicaldevices, antibiotic resistance, electrochemical corrosion, waste water treat-ment and others17.
2. The mathematical model
Biofilm growth is modeled with a CA that reproduces cell division, spacecompetition and take in account substrate concentrations. The conser-vation equations and CA models are coupled through porosity, interfaceposition (biofilm-fluid interface), and concentration and rate consumptionof nutrients. The numerical and mathematical details of our model can beconsulted in Luna et al.15. Biofilm porosity is incorporated into our modelthrough the partial differential equation, which is given by equation 1
∂
∂t[φρ + (1 − φ) ρb] + ∇ ·
(ρ→u
)= 0 , (1)

December 20, 2005 12:45 Proceedings Trim Size: 9in x 6in BIOMAT2005
3
where ρ and ρb are the fluid and bacterial densities,respectively, and→u =
ux + vy is the velocity vector. We suppose Darcy’s Law governs the rela-tionship between velocities and pressure. The mass nutrient fraction Y isgiven considering the balance equation for variable porosity under the hy-pothesis of low velocities so that dispersion effects can be safely neglected(see equation 2):
φ∂Y
∂t+
→u · ∇Y = ∇ · (φDφ∇Y ) + A . (2)
In the above equation A is the substrate consumption rate, and Dφ
is the effective molecular diffusion coefficient of nutrients in the fluid.Werefer the reader to references to justify the formulas for tortuosity andpermeability18,19,21,15.
Boundary and initial conditions are as follows: initially, there is a givenresource concentration inside the biofilm Y (x, y, t = 0) = Yini . The initialbiofilm distribution Γ (x, t = 0) is given equation 3 as:
Γ (x, t = 0) = Γ0 + Γ0 sin (kΓx) . (3)
where, Γ0 is the profile average value, Γ0 and kΓ represent the amplitudeand wave number of a small sinusoidal perturbation, respectively. Thissinusoidal shape is not that arbitrary as it may appear. Microscopy resultsshow that in biofilms involved in microbiologically induced corrosion orMIC, the bacteria-metal interface, for example, has a periodic characterthat can be approximated by a profile as the one proposed here. The initialporosity φ (x, y, t = 0) is calculated as:
φ (x, y, t = 0) = φmin +N0
NT(1 − φmin) . (4)
where, φmin is the minimum porosity that the bacterial film can reach, N0
and NT are empty and total sites in the arbitrary CA neighborhood usedto compute porosity, respectively.
This hypothesis, namely, that bacterial microcolonies are formed by thegrowth of already attached cells, has been proposed for Pseudomonas sp.and Pseudomonas putida in an experimental setting22.
We briefly describe CA basic setting and refer the reader to Luna etal.15 for further details. Our CA has either of two states at each latticepoint: occupied (1) by a bacterium or empty (0) and also carries infor-mation on the nutrient concentration at that point. Colonization or celldivision occurs with probability NeR
4 where Ne is the number of occupiednodes in a von Neumann neighborhood of each lattice point, and R is the

December 20, 2005 12:45 Proceedings Trim Size: 9in x 6in BIOMAT2005
4
probability of colonization of unoccupied sites. A substratum minimumconcentration Ymin , is required also for successful to colonization. When anode is occupied, there is an increment ∆ab in the substratum consumptionrate at that point. Mortality due to competition occurs with probabilityN0P
4 , where N0 is the number of occupied nodes in a von Neumann neigh-borhood where P is the probability of death. The node becomes empty ifthe amount of substratum at that point is below the minimum. The initialcondition for the CA was set to a random distribution with 80% occupiedlattice points in the area below the fluid bacteria interface Γ (x, t = 0) . Thereader interested in the numerical details of the model implementation isreferred to Luna et al.15 where full information on such issue is provided.
3. Detachment
Studies centered in biofilm disintegration and detachment consider the ef-fect of fluid flow over the biofilm surface23,24,25,14. We assume that micro-bial death within the biofilm weakens attachment strength and mechanicalresistance properties, especially at the biofilm-plate interface. Pseudomonasfluorescens biofilms form very quickly in well-oxygenated environments butas oxygen is depleted due to growth, disintegration develops26 may be dueto the denaturalization of the extrapolyscacharide substance matrix (EPS).In our model the maximum nutrient concentration exists, for each x, ina neighborhood of Γ (x); below this interface, nutrient concentration de-creases with depth. If the biofilm grows enough, it increases thickness andtherefore, nutrient concentration reaches a critical value at the bottom ofthe biofilm, near the attachment interface, where bacteria can no longerreproduce or survive. Cell death correlation with nutrient depletion andbiofilm growth is a reasonable approximation to the process. As discussedby Lewis27, biofilm destruction because of nutrient limitation other thanoxygen has not been conclusively demonstrated but it is a reasonable mech-anism to induce biofilm destruction. Therefore, under this assumption,namely, that nutrient depletion is associated with biofilm destruction, wederive a very simple formula to predict the breaking point of the bacterialfilm. Biofilm breakage requires that the forces per unit length due to fluiddrag (Fd) be greater than a critical value that depends on the thicknessand the internal structure of the biofilm
|Fd| > σaΓ (xa) . (5)
where Γ is the biofilm thickness and σa represents the maximum stress thatthe biofilm can resist. Full details of this derivation can be consulted in 15.
December 20, 2005 12:45 Proceedings Trim Size: 9in x 6in BIOMAT2005
5
The cellular automaton describes colonization extinction processthrough competition where R is the probability of colonization of unoc-cupied sites (related to bacterial replication), and P is the probability ofdeath due to competition for space and resources. The simulations per-formed were done for a range of values of P and R , with α = 0.57,δ = 1 × 10−2 , Ω = 1 × 10−4 , β = 1 × 10−4 , ε = 0.01 , Π0 = 0.11 (σ obtained from 14), ∆ab = 0.07 , ∆ad = 0.04 , Ymin
Yf= 0.5.
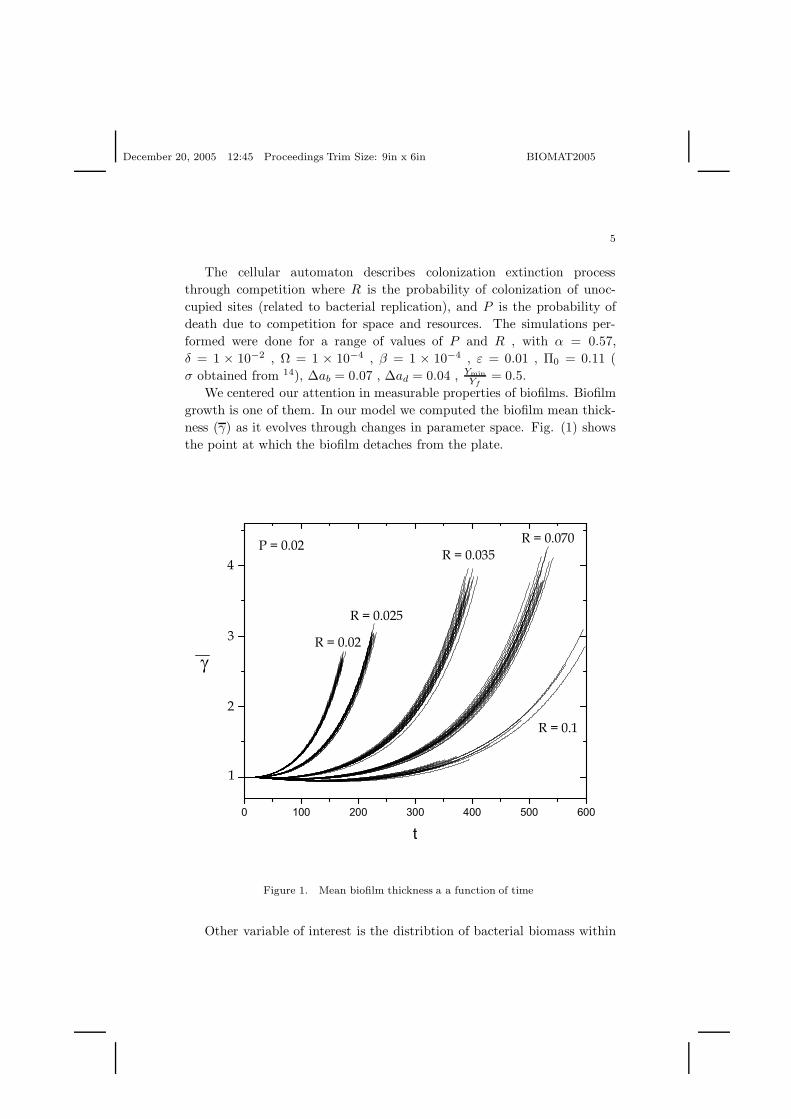
We centered our attention in measurable properties of biofilms. Biofilmgrowth is one of them. In our model we computed the biofilm mean thick-ness (γ) as it evolves through changes in parameter space. Fig. (1) showsthe point at which the biofilm detaches from the plate.
Figure 1. Mean biofilm thickness a a function of time
Other variable of interest is the distribtion of bacterial biomass within
December 20, 2005 12:45 Proceedings Trim Size: 9in x 6in BIOMAT2005
6
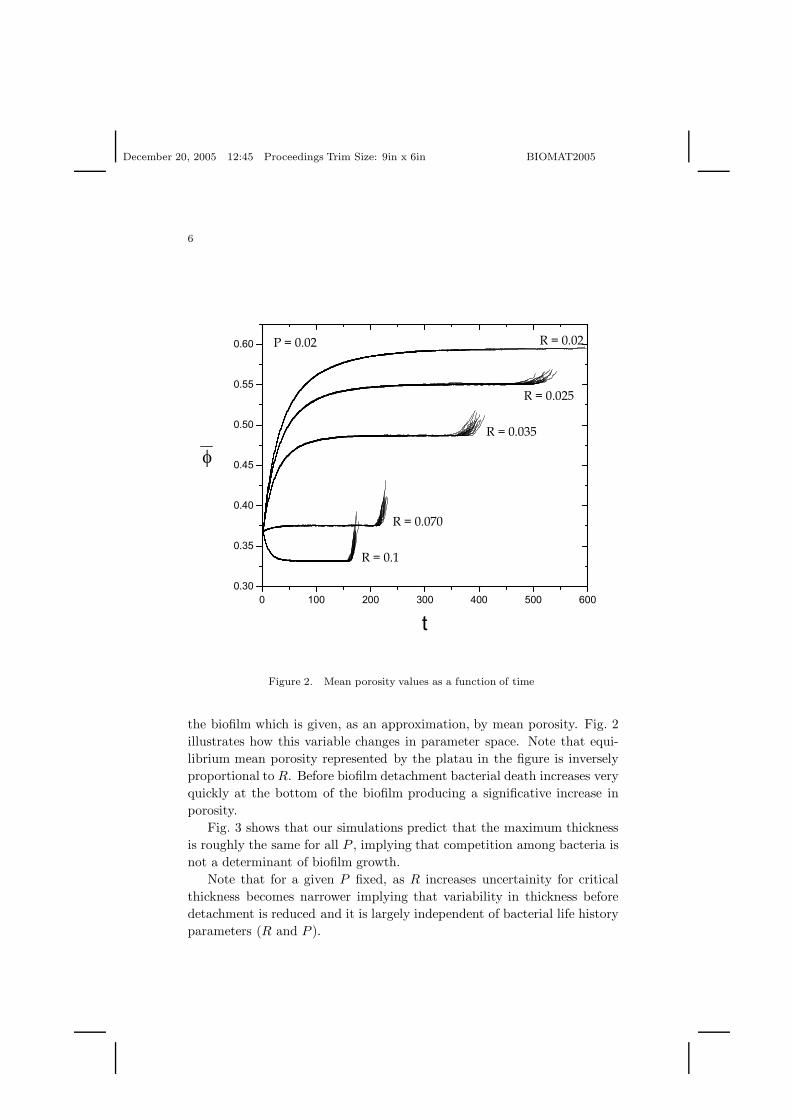
Figure 2. Mean porosity values as a function of time
the biofilm which is given, as an approximation, by mean porosity. Fig. 2illustrates how this variable changes in parameter space. Note that equi-librium mean porosity represented by the platau in the figure is inverselyproportional to R. Before biofilm detachment bacterial death increases veryquickly at the bottom of the biofilm producing a significative increase inporosity.
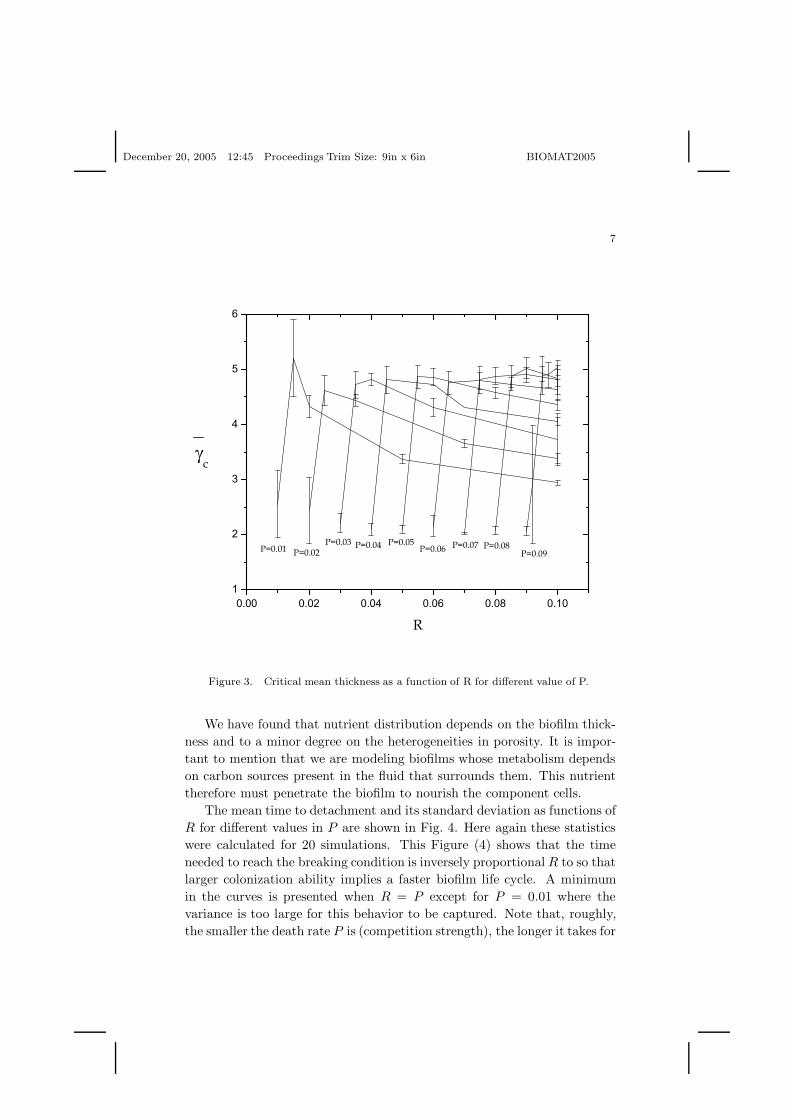
Fig. 3 shows that our simulations predict that the maximum thicknessis roughly the same for all P , implying that competition among bacteria isnot a determinant of biofilm growth.
Note that for a given P fixed, as R increases uncertainity for criticalthickness becomes narrower implying that variability in thickness beforedetachment is reduced and it is largely independent of bacterial life historyparameters (R and P ).
December 20, 2005 12:45 Proceedings Trim Size: 9in x 6in BIOMAT2005
7
Figure 3. Critical mean thickness as a function of R for different value of P.
We have found that nutrient distribution depends on the biofilm thick-ness and to a minor degree on the heterogeneities in porosity. It is impor-tant to mention that we are modeling biofilms whose metabolism dependson carbon sources present in the fluid that surrounds them. This nutrienttherefore must penetrate the biofilm to nourish the component cells.
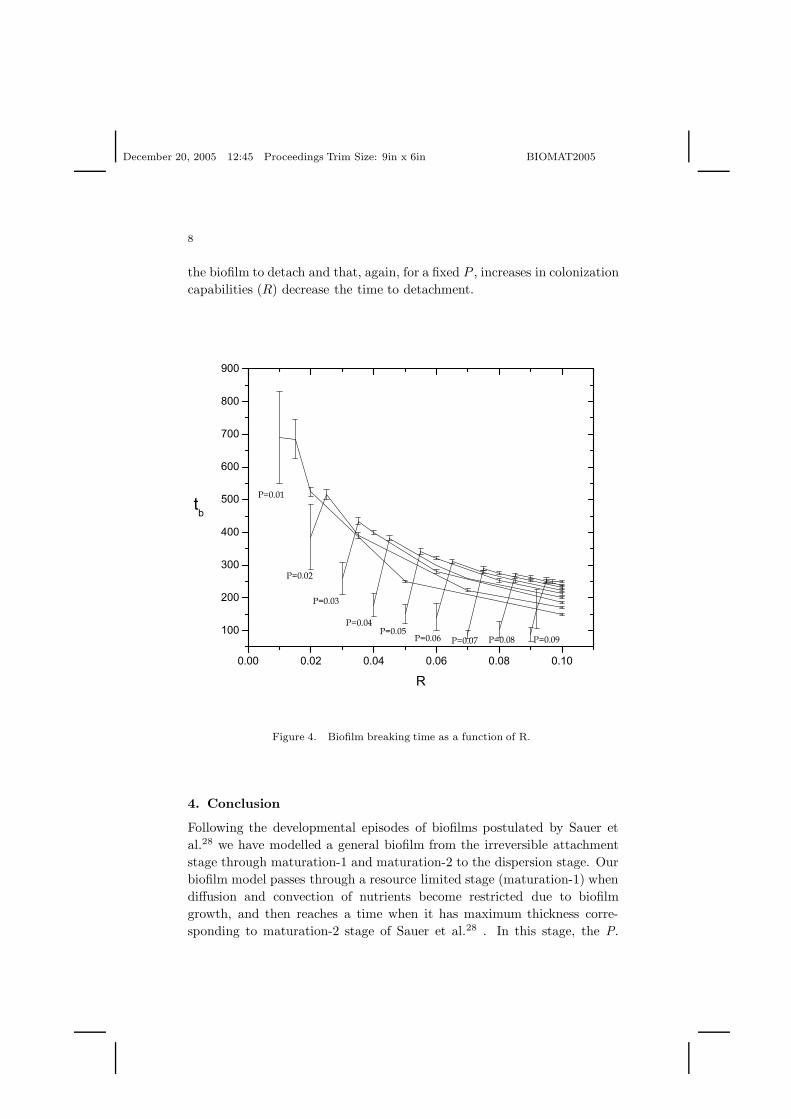
The mean time to detachment and its standard deviation as functions ofR for different values in P are shown in Fig. 4. Here again these statisticswere calculated for 20 simulations. This Figure (4) shows that the timeneeded to reach the breaking condition is inversely proportional R to so thatlarger colonization ability implies a faster biofilm life cycle. A minimumin the curves is presented when R = P except for P = 0.01 where thevariance is too large for this behavior to be captured. Note that, roughly,the smaller the death rate P is (competition strength), the longer it takes for
December 20, 2005 12:45 Proceedings Trim Size: 9in x 6in BIOMAT2005
8
the biofilm to detach and that, again, for a fixed P , increases in colonizationcapabilities (R) decrease the time to detachment.
Figure 4. Biofilm breaking time as a function of R.
4. Conclusion
Following the developmental episodes of biofilms postulated by Sauer etal.28 we have modelled a general biofilm from the irreversible attachmentstage through maturation-1 and maturation-2 to the dispersion stage. Ourbiofilm model passes through a resource limited stage (maturation-1) whendiffusion and convection of nutrients become restricted due to biofilmgrowth, and then reaches a time when it has maximum thickness corre-sponding to maturation-2 stage of Sauer et al.28 . In this stage, the P.

December 20, 2005 12:45 Proceedings Trim Size: 9in x 6in BIOMAT2005
9
aeruginosa biofilm studied by the cited authors, experiences anaerobic orreduced oxygen conditions. In our model this resource limitation occurstoo and induces cell death and an increased porosity that, according toour model, leads to the final detachment or dispersion stage. Accordingto Sauer et al.28 characterize this last stage by the observation of bacterialclusters that move away from the biofilm core leaving hollow structures orvoids. Although our model was not constructed to emulate this experimen-tal system it is surprising to see that it predicts exactly this phenomenonright before detachment takes place. Webb et al.29 report that cell deathinside biofilms plays a role in the differentiation and dispersal of subpop-ulations of surviving biofilm cells but point out that the mechanisms bywhich voids are produced within the biofilm is unclear.
Our explanation for the process of biofilm development is the formationof cavities or pores that ultimately determine the detachment process. Wehave also shown that nutrient distribution depends importantly on biofilmthickness and, secondly, on heterogeneities in porosity, tortuosity and per-meability, all this parameters related to bacterial distribution within thebiofilm. We report that biofilm thickness is inversely proportional to nu-trient concentration since transport phenomena are inefficient in providingnutrients to an ever increasing bacterial colony that consumes them. Wealso have found that there exists a critical thickness for which the nutrientconcentration at the bottom interface in not enough to maintain the bacte-ria alive. At this point, the biofilm begins to lose its mechanical resistancethat combined with the fluid drag forces generates the detachment/ruptureprocesses.
We must point out that our model does not consider the effect on biofilmdynamics of the extracellular polymeric substance matrix (EPS) that en-closes biofilms. EPS is important in certain types of antibiotic neutraliza-tion and that its mechanical and physical characteristics should play animportant role in biofilm constitution and transport of cell signaling, nu-trient and biocide diffusion. However, it is also known that27, at least inPseudomonas aeruginosa, the expression of the quorum sensing factor HSLalthough required for the conformation of a biofilm with typical architecture(channels, mushroom structures, and so forth) is not associated with theirability to resist killing by antibiotics, that is, typical biofilm architecturedoes not matter that much in this case. Moreover, while studying the prop-erties of antimicrobial resistance in biofilms and detached emboli, Fux etal.30 report that in large emboli or in stationary state biofilms starvation-induced dormancy is the common underlying mechanism for protection

December 20, 2005 12:45 Proceedings Trim Size: 9in x 6in BIOMAT2005
10
against biocides. These authors conclude that the mechanical disruption ofbiofilms, with the concomitant disruption of diffusion barriers, is correlatedwith antibiotic resistance. So although we are aware that not introducingEPS into our model may make it less general, as far as our objectives areconcern, it is a reasonable assumption.
In a S. aureus biofilm30 bacteria either detach from the surface whereit forms leaving bare spots, or from the main biofilm body (other bacte-ria) leaving on the surface an attached layer of cells. These same authorsreport that there is growing evidence that the detachment of biofilms isnot only due to the interaction with hydrodynamic forces, a process that iscalled passive detachment, but actively also in response to population den-sity, changes in substrate concentration and exposure to antimicrobials30.Hunt et al.1 have reported, in a very interesting study, supporting resultson the conjecture that nutrient starvation is a cue for detachment, findingsaligned in the direction of the conjecture generated with our model. Theseauthors use a mathematical model to explore their hypothesis. The mainpoint here is that biofilm life cycle and the determination of the mechanismsfor biofilm detachment require the application of modeling techniques andmethodologies. Other recent work in this direction, namely the study ofbiofilm detachment and associated processes using models has been pub-lished by Laspidou and Rittmann31,32.
Donlan16 cites work on the physical causes of biofilm detachment wherethree processes appear to be the most important: erosion or shearing,sloughing, and abrasion. The model presented in this paper addresses theissue of sloughing due to nutrient depletion. As stated by Lewis27 genes con-trolling biofilm destruction can be a very important factor for eradicationof biofilms. Our model, although in theoretical terms, is able to reproducebreaking/detachment, what amounts to biofilm destruction, under the sim-ple and reasonable hypothesis of postulating a correlation between biofilmdestruction and nutrient depletion. We are aware of results that indicatethat the mechanisms for detachment modeled here are not universal. Forexample, Kaplan et al.33 report that in Actinobacillus actinomycetemcomi-tans, the mechanisms for cell detachment involve the release of indivudualscells within the biofilm. Nevertheless, there is ample experimental evidencesuporting that, at least in vitro, resource limitation and detachment are as-sociated. Telman et al.34 have published experimental measurements of theeffect of increasing liquid shear on the stability and detachment of biofilms.Their results indicate that detachment is a combination of sloughing, theprocess modeled in this work, and erosion (which we do not consider here).

December 20, 2005 12:45 Proceedings Trim Size: 9in x 6in BIOMAT2005
11
Also, an important result for our approach is that biofilm life history isimportant for the detachment process and in this respect the results of Tel-man et al.34 refer to the impact of life history in biofilm morphology andsloughing. We have incorporated life history parameters in our bacteriapopulation through parameters R and P . Figures 1, 2, 3 and 4 illustratehow changing life history affects biofilm thickness, biofilm porosity and timeto detachment. Finally, we certainly share the point of view that more ex-perimental and computational studies are needed to understand biofilm lifecycles, in particular the final stage represented by detachment1.
Acknowledgments
JXVH completed his part of this work while an International Fellow of theSanta Fe Institute.We thank C.P. Ferreira to help to CA
References
1. S. M. Hunt ,E. Werner, B. Huang ,MA. Hamilton, PS. Stewart, Applied andEnvironmental Microbiology 7418 (2004).
2. H. Beyenal, C. Donovan, et al., Journal of Microbiological methods 59(3), 395(2004).
3. A. Mai-Prochnow, F. Evans, D. Dalisay-Saludes, S. Stelzer, et al., Applied andEnvironmental Microbiology 70, 3232 (2004).
4. S. Pilyugin and P. Waltman, SIAM J.Appl.Math. 59, 1552 (1999).5. D. A. Jones, H. Smith, SIAM J.Appl.Math. 60, 1576 (2000).6. E. D. Stemmons, HL. Smith, SIAM J.Appl.Math. 61, 567 (2000).7. J. Dockery, I. Klapper, SIAM J.Appl. Math. 62(3), 853 (2001).8. D. Jones, H. V. Kojouharov, D. Le, HL. Smith, J.Math.Biol. DOI.
10.1007/s00285-003-0202-I, (2003).9. C. Picioreanu, M. C. M. van Loosdrecht, JJ. Heijnen, Biotechnol. Bioeng. 57,
718 (1998).10. C. Picioreanu, MCM. van Loosdrecht, JJ. Heijnen, Biotechnol. Bioeng.
58(1), 101 (1998).11. C. Picioreanu, M. C. M. van Loosdrecht, JJ. Heijnen, Water Sci. Technol.
39(7), 115 (1999).12. C. Picioreanu, MCM. van Loosdrecht, JJ. Heijnen, Biotechnol. Bioeng. 68,
355 (2000).13. C. Picioreanu, MCM. van Loosdrecht, JJ. Heijnen, Biotechnol. Bioeng. 69,
504 (2000).14. C. Picioreanu, MCM. van Loosdrecht, JJ. Heijnen, Biotechnol. Bioeng. 72,
205 (2001).15. E. Luna, G. Dominguez-Zacarias, C. Pio Ferreira, JX. Velasco-Hernandez,
Physical Review E 70, 061909 (2004).16. R. M. Donlan, Biofilms 8, 881 (2002).

December 20, 2005 12:45 Proceedings Trim Size: 9in x 6in BIOMAT2005
12
17. P. S. Stewart, MA. Hamilton, B. Goldstein, BT. Schneider, Biotech-nol.Bioeng. 49, 445 (1996).
18. R. Islas-Juarez, M.I. Thesis, Univerisad Nacional Autonoma de Mexico(2003).
19. E. L. Cussler, Cambridge University Press., New York, US (1997).20. C. N. Davies, Proc. Inst.Mech. Eng. 1B, 185 (1952).21. P. M. Adler and J-F. Thover, Appl. Mech. Rev. 51(9), 537 (1998).22. T. Tolker-Nielsen, UC. Brinch, PC. Ragas, JB. Andersen, CS. Jacobsen, S.
Molin, Journal of Bacteriology 182, 6482 (2000).23. H. Horn, H. Reiff, E. Morgenroth, Biotechnol. Bioeng. 81, 607 (2003).24. L. Mascari, P. Ymele-Leki, CD. Eggleton, P. Speziale, JM. Ross, Biotechnol.
Bioeng. 83, 65 (2003).25. I. Klapper, C. J. Rupp, R. Cargo, B. Purvedorj, P. Stoodley, Biotechnol.
Bioeng. 80, 289 (2002).26. D. G. Allison, B. Ruiz, C. San Jose, A. Jaspe, p. Gilbert, FEMS Micro-
biol.Lett. 167, 179 (1998).27. K. Lewis, Antimicrobial agents and chemotherapy 95, 999 (2001).28. K. Sauer, A. K. Camper, GD. Ehrlich, JW. Costerton, DG. Davies, Journal
of Bacteriology 184, 1140 (2002).29. J. S. Webb, L. S. Thompson, S. James, T. Charlton, T. Tolker-Nielsen, B.
Koch, M. Givskov, S. Kjelleberg, Journal of Bacteriology 185, 4585 (2003).30. C. A. Fux, S. Wilson, P. Stoodley, Journal of Bacteriology 186, 4486 (2004).31. C. Laspidou, B. E. Rittmann, Water Research 38, 3349 (2004).32. C. Laspidou , B. E. Rittmann, Water Research 38 3362 (2004).33. J. Kaplan, M. F. Meyenhofer, D. H. Fine, Journal of Bacteriology 185, 1399
(2003).34. U. Telmann, H. Horn, E. Mongenroth, Water Research 38, 3671 (2004).




















