
The disruption of JEN1 from Candida albicans impairs the transport of lactate

The SUN41 and SUN42 genes are essential for cellseparation in Candida albicans
Arnaud Firon,1 Sylvie Aubert,1† Ismaïl Iraqui,2‡
Stéphanie Guadagnini,3 Sophie Goyard,1
Marie-Christine Prévost,3 Guilhem Janbon2 andChristophe d’Enfert1*1Unité Biologie et Pathogénicité Fongiques, INRAUSC2019, Institut Pasteur, Paris, France.2Unité de Mycologie Moléculaire, CNRS URA3012,Institut Pasteur, Paris, France.3Plate-forme de Microscopie Ultrastructurale, InstitutPasteur, Paris, France.
Summary
Completion of the yeast cell cycle involves extensiveremodelling of the cell wall upon separation of motherand daughter cells. We have studied two members ofthe ascomycete-specific SUN gene family in Candidaalbicans. Inactivation of SUN41 yields defects in cellseparation and hyphal elongation while inactivationof SUN42 results in minor phenotypic alterations.Simultaneous inactivation of SUN41 and SUN42 issynthetically lethal due to lysis of mother cells afterseptation. Electronic microscopy reveals cell walldefects mainly localized in the region surrounding thesepta. This phenotype is osmoremediable and theconditional double mutants show increased sensitiv-ity to cell wall or cell membrane perturbing agents.The essential function shared by Sun41p and Sun42pis conserved among yeasts because UTH1, a Saccha-romyces cerevisiae SUN gene, suppresses thelethality of SUN41 and SUN42 conditional mutants.Investigation of functional genomic data obtained inS. cerevisiae reveals links between members of theSUN gene family and the RAM pathway regulating cellwall-degrading enzymes specifically involved duringcell separation. Thus, the main function of ascomyc-etous Sun proteins appears linked to cell wall remod-elling, with a probable role in counter-balancing cellwall degradation to avoid cell lysis upon cellseparation.
Introduction
The last step of the yeast cell cycle is the separation ofmother and daughter cells by the degradation of theprimary septum. This event is tightly controlled andrequires extensive remodelling of the cell wall. In particu-lar, studies performed using Saccharomyces cerevisiaehave evidenced a complex regulatory system thatensures proper expression of cell wall-remodellingenzymes such as the Cts1p chitinase and Scw11p gluca-nase involved in degrading the primary septum (Cabibet al., 2001; Yeong, 2005). Central to this regulatorysystem is the essential RAM–Ace2p signalling pathwaythat controls daughter-specific gene expression of asubset of genes including CTS1 and SCW11 (Colman-Lerner et al., 2001; Nelson et al., 2003). Components ofthe RAM pathway – the Cbk1p kinase and associatedproteins (Mob2p, Tao3p, Hym1p, Sog2p, Kic1p) – controlthe nuclear sequestration of the Ace2p transcription factorin the daughter cell nucleus during mitotic exit as well asan Ace2p-independent polarization process (Bidlingmaieret al., 2001; Colman-Lerner et al., 2001; Weiss et al.,2002; Nelson et al., 2003; Kurischko et al., 2005). Degra-dation of the primary septum by the Cts1p chitinase andthe Scw11p glucanase is counter-balanced by the activityof the Chs1p chitin synthase allowing completion of cellseparation without cell lysis (Bulawa et al., 1986; Cabibet al., 1992; Yeong, 2005). However, details are stilllacking on how the opposite activities of chitinase andchitin synthase are balanced independently of anyco-ordinated regulation (Selvaggini et al., 2004).
As will be discussed below several lines of evidencelink the S. cerevisiae proteins of the Sun protein family tocell separation and to the RAM–Ace2p pathway. The Sunprotein family was originally recognized following theidentification of four paralogous genes in the buddingyeast S. cerevisiae, namely SIM1, UTH1, NCA3 andSUN4 (Austriaco, 1996). Conserved features among theSun proteins (Fig. 1A) include a signal peptide and ahomologous carboxy-terminal domain, referred to as theSUN domain (Pfam domain PF03856; Finn et al., 2006).The SUN domain is characterized by a C-X5-C-X3-C-X24-Cmotif predicted to bind iron based on similarities to Fe-Sproteins (Bandara et al., 1998). S. cerevisiae harboursa fifth protein with a putative SUN domain, namelyYmr244w. Yet, this protein differs from the four
Accepted 15 October, 2007. *For correspondence. E-mail [email protected]; Tel. (+33) 1 40 61 32 57; Fax (+33) 1 45 68 89 38. Presentaddresses: †Groupe à 5 ans de Génétique des Nématodes, InstitutPasteur, Paris, France; ‡CNRS UMR2027, Institut Curie, Bâtiment110, 91405 Orsay, France.
Molecular Microbiology (2007) 66(5), 1256–1275 doi:10.1111/j.1365-2958.2007.06011.x
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd

prototypical Sun proteins by a shorter N-terminal region, alow complexity region at the C-terminal end and multipleinsertions in the conserved SUN domain including a3-amino-acid insertion in the putative iron-binding motifleading to a spacing of 27 amino acids instead of 24between the third and fourth cysteines (Fig. 1A).
Since the initial definition of the Sun protein family, othermembers have been identified through inspection ofgenome sequences. Interestingly, the Sun protein familyappears restricted to ascomycetes because an iterativeBLAST (PSI-BLAST) analysis fails to detect homologues ofthe Sun proteins in fully sequenced organisms from otherphyla. As shown in Fig. 1B, Sun proteins identified in fully
sequenced genomes can be assigned to two groups.Group I includes the four prototypical Sun proteins ofS. cerevisiae and related proteins. Group II includes pro-teins related to the S. cerevisiae Ymr244w protein. It isnotable that euascomycetes (and Yarrowia lipolytica)have one group I protein and that hemiascomycetes havetwo group I proteins, except those species that haveexperienced the whole genome duplication (Dietrichet al., 2004; Kellis et al., 2004; Dujon, 2006). Therefore,the two paralogous pairs in S. cerevisiae (SIM1/SUN4and UTH1/NCA3) are likely to descend, respectively, fromtwo unique ancestors.
The function of each of the Sun proteins remainsunclear. Many of these proteins have been annotatedunduly as having a beta-glucosidase activity on the basisof the annotation available in GenBank for a Candidawickerhamii Sun protein, BglA (Skory and Freer, 1995).The bglA and bglB genes were identified in a screen forC. wickerhamii genes encoding proteins that immunore-act with polyclonal antibodies directed towards a purifiedextracellular beta-glucosidase. Yet, beta-glucosidaseactivity was demonstrated for the BglB protein only (Skoryand Freer, 1995). Thus, Sun proteins cannot be consid-ered beta-glucosidases because no experimental evi-dences are available at this date (Omi et al., 1999; deGroot et al., 2007).
Fig. 1. Phylogenetic analysis of the Sun protein family.A. Schematic representation of the typical organization ofSun proteins. The global architecture is in three parts: anamino-terminal signal peptide (SP), a low complexity region(LCR) rich in serine and threonine residues and a conservedcarboxy-terminal SUN domain containing a four-cysteine motifpredicted to bind iron.B. Phylogeny of Sun proteins identified in yeast and filamentousascomycetes. The sequences of known and putative Sun proteinsidentified in the deduced proteomes of S. cerevisiae (Sc), Candidaglabrata (Cg), Kluyveromyces lactis (Kl), Ashbya (Eremothecium)gossypii (Eg), C. albicans (Ca; dark grey background),Debaryomyces hansenii (Dh), Yarrowia lipolytica (Yl),Schizosaccharomyces pombe (Sp), Aspergillus (Emericella)nidulans (En), A. fumigatus (Af), A. oryzae (Ao), Neurospora crassa(Nc), Giberella zeae (Gz) and Coccidioides immitis (Ci) werealigned and an unrooted tree was generated using CLUSTALX.Branches with a bootstrap value above 85% (1000 repetitions) areindicated with a black dot. Two main groups were defined: Group Icontains typical Sun proteins including S. cerevisiae Sim1p, Uth1p,Nac3p and Sun4p while Group II contains atypical Sun proteinswith a degenerate SUN domain and a carboxy-terminal lowcomplexity domain. Proteins from hemiascomycetes that haveexperienced the whole genome duplication have a blackbackground. Proteins from euascomycetes have a light greybackground.C. Similarity of S. cerevisiae (S. c) and C. albicans (C. a) Sunproteins. Sun proteins and their corresponding SUN domains havebeen compared using BLASTP. E-values obtained for proteincomparisons are indicated with a white background and thepercentages of amino acid identity in the SUN domain are shownwith a grey background. The size in amino acids for each proteinand for each corresponding SUN domain as well as the position ofthe SUN domain on the protein sequence (in parenthesis) areindicated.
SUN genes of C. albicans 1257
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

Characterization of deletion mutants in S. cerevisiaehas uncovered numerous, often pleiotropic, phenotypeslinking the SUN genes to several cellular processes: DNAreplication for SIM1 (Dahmann et al., 1995), ageing, oxi-dative stress, mitochondrial biogenesis and autophagyfor UTH1 (Kennedy et al., 1995; Bandara et al., 1998;Camougrand et al., 2004; Kissova et al., 2004), mitochon-drial biogenesis for NCA3 (Pelissier et al., 1995), andseptation for SUN4 (Mouassite et al., 2000). Simulta-neous inactivation of SUN4 and UTH1 aggravates the cellseparation defect associated to SUN4 deletion, suggest-ing that the UTH1 gene product might also be involvedin cell separation (Mouassite et al., 2000). Yet, a compre-hensive analysis of double, triple and quadruple deletionmutants of SUN genes in S. cerevisiae that would takeinto account their phylogenetic relationship (see aboveand Fig. 1B) is still lacking. Interestingly, deletion of aSchizosaccharomyces pombe group I SUN gene, namelypsu1 +, is lethal and associated with cell wall defectsduring cell separation (Omi et al., 1999).
Consistent with an involvement in cell separation ofSun4p (Mouassite et al., 2000) and possibly other Sunproteins such as Uth1p (Mouassite et al., 2000) andS. pombe Psu1p (Omi et al., 1999), several members ofthe S. cerevisiae SUN gene family are linked to the RAM–Ace2p signalling pathway: (i) inactivation of SUN4 altersthe S. cerevisiae transcriptome in a manner similar to thatobserved when genes specifically expressed in daughtercells are inactivated as revealed by genome-wide hierar-chical clustering of mutant transcriptomes (Colman-Lerner et al., 2001; Nelson et al., 2003), (ii) SIM1 is adosage suppressor of the lethality associated to inactiva-tion of components of the RAM pathway (Du and Novick,2002; Kurischko et al., 2005) and (iii) inactivation of CBK1is associated with upregulation of NCA3 along with theCHS1 chitin synthase gene and downregulation of CTS1and SCW11 (Bidlingmaier et al., 2001). Furthermore,psu1 + is one of seven genes that are positively regulatedby Ace2p in S. pombe (Alonso-Nunez et al., 2005).
Candida albicans is currently responsible for a signifi-cant fraction of fungal infections in otherwise healthy indi-viduals and for the majority of fungal infections contractedin the hospital environment (Wisplinghoff et al., 2004). Inthis species, three SUN genes have been identified:SUN41 (orf19.3642) and SUN42 (orf19.5032) encodeGroup I Sun proteins while orf19.5896 encodes a Group IIprotein with significant divergence at the amino acid levelwhen compared with Sun41p and Sun42p (Fig. 1C).Nothing is known about the function of these genes inC. albicans. However, transcript profiling studies haveindicated that SUN41 is upregulated upon induction ofhyphal growth (Sohn et al., 2003) and that SUN41 andSUN42 are overexpressed in biofilm relative to planktoniccells (Garcia-Sanchez et al., 2004) and under hypoxia
(Setiadi et al., 2006). Because SUN41 and SUN42 areupregulated under conditions that are relevant to thepathogenesis of C. albicans – hyphal morphogenesis,hypoxia, biofilm formation – and encode secretory pro-teins, they may play important roles in the morphogenesisand pathobiology of C. albicans. This prompted us tostudy the function of SUN41 and SUN42 in C. albicans.Here, we present the characterization of C. albicanssun41 and sun42 single and double mutants and showthat SUN41 is necessary for proper cell separation andhyphal differentiation. Moreover, SUN41 and SUN42 aresynthetically lethal and their simultaneous inactivationresults in a cell wall defect evidenced after cell separation.Because S. cerevisiae UTH1 can substitute the essentialfunction of SUN41 and SUN42, we propose that membersof the Sun protein family have a conserved role in cell wallremodelling necessary to maintain cell integrity upon cellseparation in yeasts.
Results
The C. albicans SUN41 gene is required for cellseparation
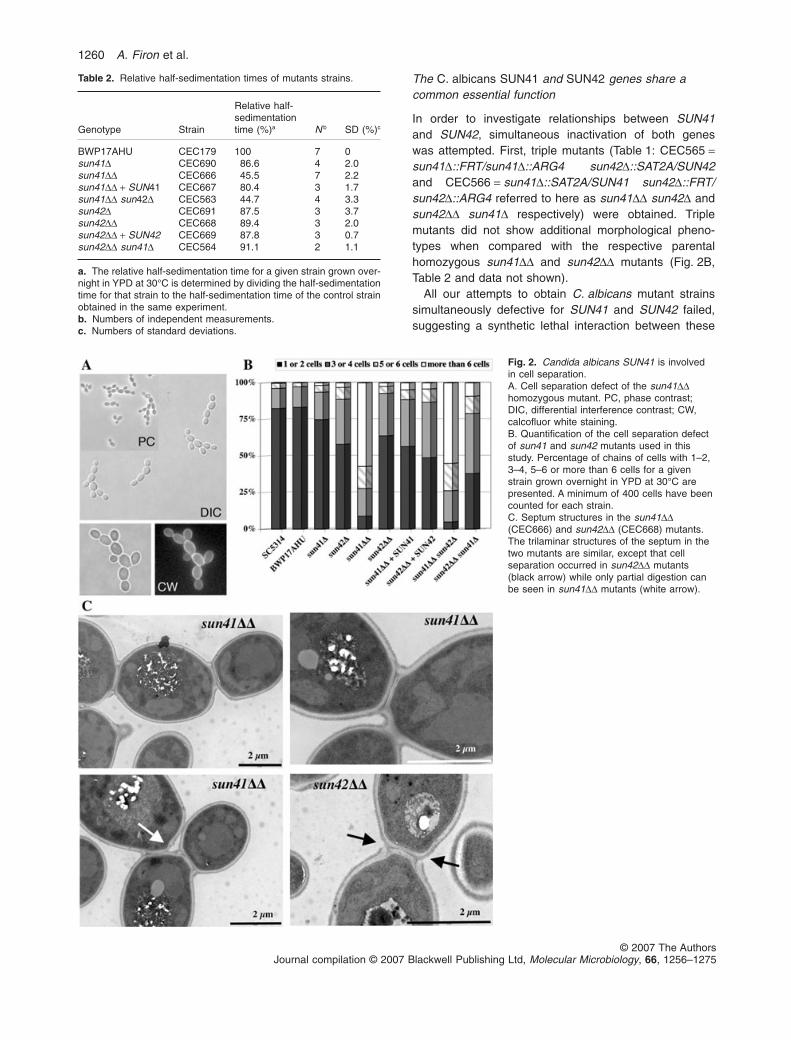
Candida albicans mutant strains deleted for oneallele (Table 1: CEC690 = sun41D::SAT2A/SUN41 andCEC691 = sun42D::SAT2A/SUN42 referred to belowas sun41D and sun42D for clarity) or two alleles(Table 1: CEC666 = sun41D::SAT2A/sun41D::ARG4 andCEC668 = sun42D::SAT2A/sun42D::ARG4 referred tobelow as sun41DD and sun42DD for clarity) of the SUN41and SUN42 genes were constructed (see Experimentalprocedures). When grown on YPD or synthetic dextrose(SD) solid media at 30°C, the mutant strains did not showany apparent growth defect relative to an isogenic wild-type strain (data not shown). In contrast, when mutantcells were grown in YPD or SD liquid media at 30°C,accelerated sedimentation was observed for cells of thesun41DD mutant strain relative to cells from the wild-typeand other mutant strains. As shown in Table 2, cells of thesun41DD mutant grown in YPD medium had a rela-tive half-sedimentation time of 45.5% � 2.2% (n = 7;wild-type = 100%). The sedimentation phenotype of thesun41DD mutant was not abolished by treatments such asvortexing, sonication or chelation of Ca2+ by addition of upto 0.4 M EDTA (data not shown), suggesting that thisphenotype is not linked to defects in cell–cell adhesionprocesses (Verstrepen and Klis, 2006).
Microscopic observation of a liquid culture of thesun41DD mutant strain revealed cell separation defects(Fig. 2A): sun41DD mutant cells had an apparently normalmorphology but formed chains of yeast cells connected bya septum as indicated by calcofluor white staining of chitin(Fig. 2A). Relative half-sedimentation times and cell sepa-ration defects were correlated and the reintroduction of a
1258 A. Firon et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

wild-type copy of SUN41 in the sun41DD mutant restoredsedimentation and cell separation to a level similar to thatobserved for the heterozygous sun41D mutant strain(Fig. 2B and Table 2). As shown in Table 2, the sun41Dmutant showed slightly accelerated sedimentation(86.6% � 2%, n = 4) suggesting a haplo-insufficiencyphenotype. A slight decrease in the relative half-sedimentation time was also observed for the heterozy-gous sun42D mutant (Table 2: 87.5% � 3.7%, n = 3). Yet,this phenotype was not aggravated in a homozygoussun42DD mutant (Table 2: 89.4% � 2%, n = 3) that doesnot form chains of connected cells (Fig. 2B).
Structure of the septum in sun41DD and sun42DDmutants was investigated by transmission electronic
microscopy (TEM) (Fig. 2C). In both mutants, septaappeared normal with a three laminar structure consistingof a primary septum between two secondary septa (oneon the mother cell side, the other on the daughter side).However, while sun42DD mutant cells showed normaldegradation of the primary septum upon cell separation,sun41DD mutant cells remained physically connected bytheir septum, even though initiation of septum degrada-tion could be observed in some instances. Overall, thestructure of the cell wall in sun41DD and sun42DD mutantswas similar to that in the control strain. Thus, the separa-tion defect of the sun41DD mutant appears confined to aninability to degrade the primary septum at the end of thecell cycle.
Table 1. Strains used in this study.
Strain Genotype Auxotrophy Reference/abbreviation
Reference strainSC5314 Clinical isolate Wilson et al. (1999)BWP17 ura3D::limm434/ura3D::limm434 Ura Wilson et al. (1999)
his1::hisG/his1::hisG Hisarg4::hisG/arg4::hisG Arg
Strains constructed during this studyBWP17AHU ura3D::limm434/URA3 BWP17AHU
his1::hisG/HIS1arg4::hisG/ARG4
CEC690 sun41D::SAT2A/SUN41 sun41DCEC691 sun42D::SAT2A/SUN42 sun42DCEC666 sun41D::SAT2A/sun41D::ARG4 sun41DDCEC668 sun42D::SAT2A/sun42D::ARG4 sun42DDCEC667 sun41D::SAT2A/sun41D::ARG4; CIp-SUN41 sun41DD + SUN41CEC669 sun42D::SAT2A/sun42D::ARG4; CIp-SUN42 sun42DD + SUN42CEC849 sun41D::SAT2A/sun41D::ARG4; HIS1-PTEF1-SUN42/SUN42 sun41DD + PTEF1-SUN42CEC563 sun41D::FRT/sun41D::ARG4; sun42D::HIS1/SUN42 sun41DD sun42DCEC564 sun41D::HIS1/SUN41; sun42D::FRT/sun42D::ARG4 sun42DD sun41DCEC565 sun41D::FRT/sun41D::ARG4; Ura sun41DD sun42D
sun42D::SAT2A/SUN42 HisCEC566 sun41D::SAT2A/SUN41; Ura sun42DD sun41D
sun42D::FRT/sun42D::ARG4 HisCEC606 sun41D::SAT2A/HIS1-PMET3-SUN41;
sun42D::FRT/sun42D::ARG4sun42DD sun41D PMET3-SUN41
CEC608 sun41D::SAT2A/HIS1-PMET3-SUN41;sun42D::FRT/sun42D::ARG4; CIp-SUN42
sun42DD sun41D PMET3-SUN41 + SUN42
CEC610 sun41D::SAT2A/HIS1-PMET3-SUN41;sun42D::FRT/sun42D::ARG4; CIp-SUN41
sun42DD sun41D PMET3-SUN41 + SUN41
CEC612 sun41D::SAT2A/HIS1-PMET3-SUN41;sun42D::FRT/sun42D::ARG4; CIp-SUN4
sun42DD sun41D PMET3-SUN41 + SUN4
CEC613 sun41D::SAT2A/HIS1-PMET3-SUN41;sun42D::FRT/sun42D::ARG4; CIp-SIM1
sun42DD sun41D PMET3-SUN41 + SIM1
CEC614 sun41D::SAT2A/HIS1-PMET3-SUN41;sun42D::FRT/sun42D::ARG4; CIp-UTH1
sun42DD sun41D PMET3-SUN41 + UTH1
CEC615 sun41D::SAT2A/HIS1-PMET3-SUN41;sun42D::FRT/sun42D::ARG4; CIp-NCA3
sun42DD sun41D PMET3-SUN41 + NCA3
CEC601 sun41D::FRT/sun41D::ARG4;sun42D::SAT2A/HIS1-PMET3-SUN42
sun41DD sun42D PMET3-SUN42
CEC603 sun41D::FRT/sun41D::ARG4;sun42D::SAT2A/HIS1-PMET3-SUN42; CIp-SUN42
sun41DD sun42D PMET3-SUN42 + SUN42
CEC616 sun41D::FRT/sun41D::ARG4;sun42D::SAT2A/HIS1-PMET3-SUN42; CIp-SUN41
sun41DD sun42D PMET3-SUN42 + SUN41
CEC618 sun41D::FRT/sun41D::ARG4;sun42D::SAT2A/HIS1-PMET3-SUN42; CIp-UTH1
sun41DD sun42D PMET3-SUN42 + UTH1
CEC785 sun41D::PSUN41-GFP-HIS1/SUN41 Arg PSUN41–GFPCEC786 sun42D::PSUN42-GFP-HIS1/SUN42 Arg PSUN42–GFP
SUN genes of C. albicans 1259
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275
The C. albicans SUN41 and SUN42 genes share acommon essential function
In order to investigate relationships between SUN41and SUN42, simultaneous inactivation of both geneswas attempted. First, triple mutants (Table 1: CEC565 =sun41D::FRT/sun41D::ARG4 sun42D::SAT2A/SUN42and CEC566 = sun41D::SAT2A/SUN41 sun42D::FRT/sun42D::ARG4 referred to here as sun41DD sun42D andsun42DD sun41D respectively) were obtained. Triplemutants did not show additional morphological pheno-types when compared with the respective parentalhomozygous sun41DD and sun42DD mutants (Fig. 2B,Table 2 and data not shown).
All our attempts to obtain C. albicans mutant strainssimultaneously defective for SUN41 and SUN42 failed,suggesting a synthetic lethal interaction between these
Table 2. Relative half-sedimentation times of mutants strains.
Genotype Strain
Relative half-sedimentationtime (%)a N b SD (%)c
BWP17AHU CEC179 100 7 0sun41D CEC690 86.6 4 2.0sun41DD CEC666 45.5 7 2.2sun41DD + SUN41 CEC667 80.4 3 1.7sun41DD sun42D CEC563 44.7 4 3.3sun42D CEC691 87.5 3 3.7sun42DD CEC668 89.4 3 2.0sun42DD + SUN42 CEC669 87.8 3 0.7sun42DD sun41D CEC564 91.1 2 1.1
a. The relative half-sedimentation time for a given strain grown over-night in YPD at 30°C is determined by dividing the half-sedimentationtime for that strain to the half-sedimentation time of the control strainobtained in the same experiment.b. Numbers of independent measurements.c. Numbers of standard deviations.
Fig. 2. Candida albicans SUN41 is involvedin cell separation.A. Cell separation defect of the sun41DDhomozygous mutant. PC, phase contrast;DIC, differential interference contrast; CW,calcofluor white staining.B. Quantification of the cell separation defectof sun41 and sun42 mutants used in thisstudy. Percentage of chains of cells with 1–2,3–4, 5–6 or more than 6 cells for a givenstrain grown overnight in YPD at 30°C arepresented. A minimum of 400 cells have beencounted for each strain.C. Septum structures in the sun41DD(CEC666) and sun42DD (CEC668) mutants.The trilaminar structures of the septum in thetwo mutants are similar, except that cellseparation occurred in sun42DD mutants(black arrow) while only partial digestion canbe seen in sun41DD mutants (white arrow).
1260 A. Firon et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

two genes. Thus, C. albicans strains where only one alleleof one of the two genes was maintained and placed underthe control of the methionine/cysteine-repressible pro-moter of the C. albicans MET3 gene (PMET3) (Care et al.,1999) were produced (Table 1: CEC601 = sun41D::FRT/sun41D::ARG4 sun42D::SAT2A/HIS1-PMET3-SUN42and CEC606 = sun41D::SAT2A/HIS1-PMET3-SUN41sun42D::FRT/sun42D::ARG4 referred to here as sun41DDsun42D PMET3-SUN42 and sun42DD sun41D PMET3-SUN41respectively). Data presented in Fig. 3A show that thesun41DD sun42D PMET3-SUN42 and sun42DD sun41DPMET3-SUN41 conditional strains failed to grow on syn-thetic medium under PMET3-repressing conditions. Simi-larly, when the conditional strains were grown in liquidmedium, we observed growth inhibition under PMET3-repressing conditions (data not shown) and increaseddoubling times under non-repressing conditions (wild-type: 90 min; sun42DD sun41D PMET3-SUN41: 120 min;sun41DD sun42D PMET3-SUN42: 145 min).
The growth defect associated with PMET3 -dependentrepression of SUN41 or SUN42 was complemented tothe same extent by ectopic integration of either one copyof SUN41 or SUN42 under the control of their own pro-moter (Fig. 3A). Yet, ectopic integration of one copy ofSUN42 in the sun42DD sun41D PMET3-SUN41 strain did
not suppress the cell separation phenotype associatedwith the inactivation of SUN41 (data not shown). This isin agreement with the morphology of the parental triplemutant sun41DD sun42D that harbours one wild-typecopy of SUN42 and shows a cell separation defect(Fig. 2B). Thus, SUN41 and SUN42 share a commonessential function for C. albicans growth even thoughSUN41 may perform additional functions that are notshared with SUN42.
C. albicans SUN genes are simultaneously required forviability of mother cells
Microscopic observation of the conditional sun41DDsun42D PMET3-SUN42 and sun42DD sun41D PMET3-SUN41 mutants after 16 h in PMET3-repressing conditionshowed clumps of mostly ghost cells (Fig. 4A). Themother and daughter cells remained physically con-nected, even after cell lysis, a cell separation defectreminiscent of the phenotype of the homozygoussun41DD mutant. Calcofluor white staining showed vari-able content and distribution of chitin in these clumps(Fig. 4A). Although most cells were ghosts, some viablecells remained at the periphery of the clumps (Fig. 4A).The presence of viable cells was confirmed by transfer-
Fig. 3. C. albicans SUN41 and SUN42 synthetic lethality is suppressed by expression of the S. cerevisiae UTH1 gene and is linked to cellwall defects. Overnight culture in SD medium at 30°C of strains BWP17AHU (SUN41/SUN41 SUN42/SUN42), conditional strain sun42DDsun41D PMET3-SUN41 (CEC606) with one copy of C. albicans SUN41 (CEC610), SUN42 (CEC608) or S. cerevisiae UTH1 (CEC614), andconditional strain sun41DD sun42D PMET3-SUN42 (CEC601) with one copy of C. albicans SUN41 (CEC616), SUN42 (CEC603) or S. cerevisiaeUTH1 (CEC618), were adjusted to OD600 = 0.4 (~5 ¥ 106 cells ml-1) and serial dilutions (5¥) were spotted on (A) SD agar medium (SD), SDagar medium supplemented with 5 mM methionine and 0.25 mM cysteine (SD + M + C, PMET3 repressing conditions) and (B) on YPD and YPDcontaining 1 M sorbitol. Plates were incubated at 30°C for 2 days for the sun42DD sun41D PMET3-SUN41 series and 3 days for the sun41DDsun42D PMET3-SUN42 series. (C) Drug sensitivities of strains BWP17AHU (SUN41/SUN41 SUN42/SUN42), homozygous sun41DD (CEC666)and sun42DD (CEC668), triple sun41DD sun42D (CEC565) and sun42DD sun41D (CEC566), and conditional sun42DD sun41D PMET3-SUN41(CEC606) and sun41DD sun42D PMET3-SUN42 (CEC601). Overnight cultures were adjusted to OD600 = 0.4 (~5 ¥ 106 cells ml-1) and serialdilutions (5¥) were spotted (3 ml) on SD agar medium (SD), YPD and YPD + drugs (CW, calcofluor white; Nikko Z, nikkomycin Z). Photos weretaken after 3 days at 30°C.
SUN genes of C. albicans 1261
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275
ring overnight cultures of the sun41DD sun42D PMET3-SUN42 and sun42DD sun41D PMET3-SUN41 conditionalmutants grown in PMET3-repressing conditions in freshPMET3 non-repressing medium. Under these conditions,growth resumed after a long lag phase (~8 h comparedwith 2 h for a wild-type strain, data not shown).
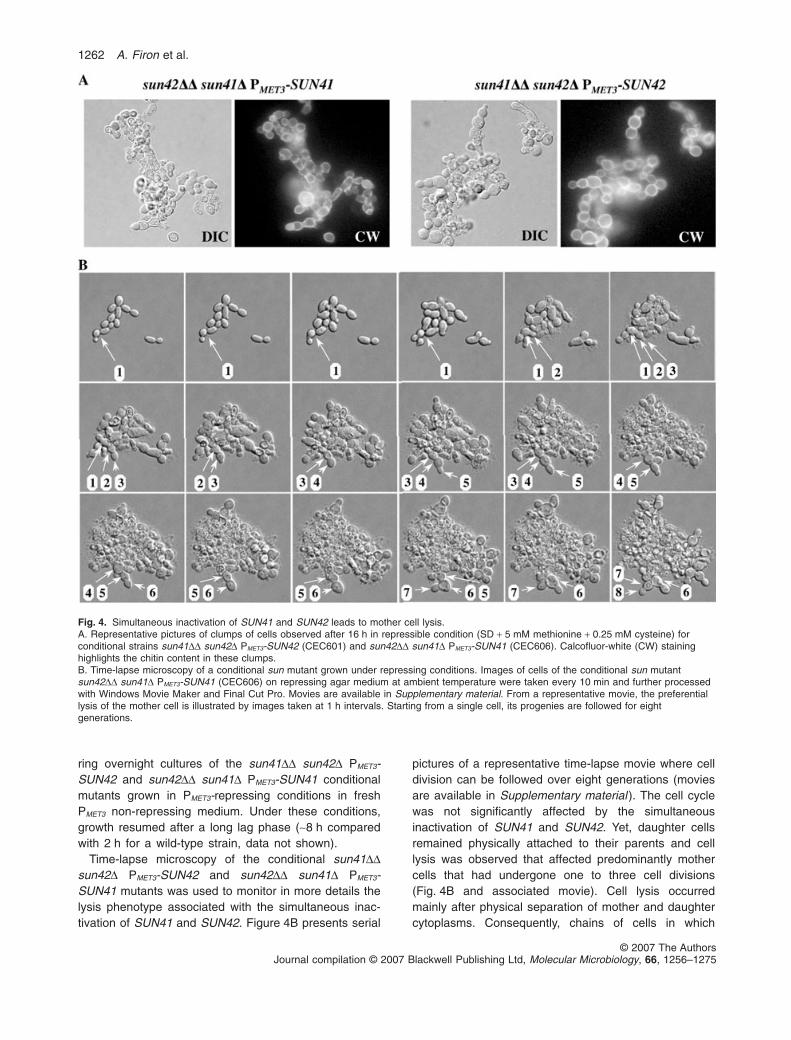
Time-lapse microscopy of the conditional sun41DDsun42D PMET3-SUN42 and sun42DD sun41D PMET3-SUN41 mutants was used to monitor in more details thelysis phenotype associated with the simultaneous inac-tivation of SUN41 and SUN42. Figure 4B presents serial
pictures of a representative time-lapse movie where celldivision can be followed over eight generations (moviesare available in Supplementary material ). The cell cyclewas not significantly affected by the simultaneousinactivation of SUN41 and SUN42. Yet, daughter cellsremained physically attached to their parents and celllysis was observed that affected predominantly mothercells that had undergone one to three cell divisions(Fig. 4B and associated movie). Cell lysis occurredmainly after physical separation of mother and daughtercytoplasms. Consequently, chains of cells in which
Fig. 4. Simultaneous inactivation of SUN41 and SUN42 leads to mother cell lysis.A. Representative pictures of clumps of cells observed after 16 h in repressible condition (SD + 5 mM methionine + 0.25 mM cysteine) forconditional strains sun41DD sun42D PMET3-SUN42 (CEC601) and sun42DD sun41D PMET3-SUN41 (CEC606). Calcofluor-white (CW) staininghighlights the chitin content in these clumps.B. Time-lapse microscopy of a conditional sun mutant grown under repressing conditions. Images of cells of the conditional sun mutantsun42DD sun41D PMET3-SUN41 (CEC606) on repressing agar medium at ambient temperature were taken every 10 min and further processedwith Windows Movie Maker and Final Cut Pro. Movies are available in Supplementary material. From a representative movie, the preferentiallysis of the mother cell is illustrated by images taken at 1 h intervals. Starting from a single cell, its progenies are followed for eightgenerations.
1262 A. Firon et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

viability decreased with each completed division couldbe observed.
Simultaneous inactivation of the SUN genes isassociated with defects at the septum
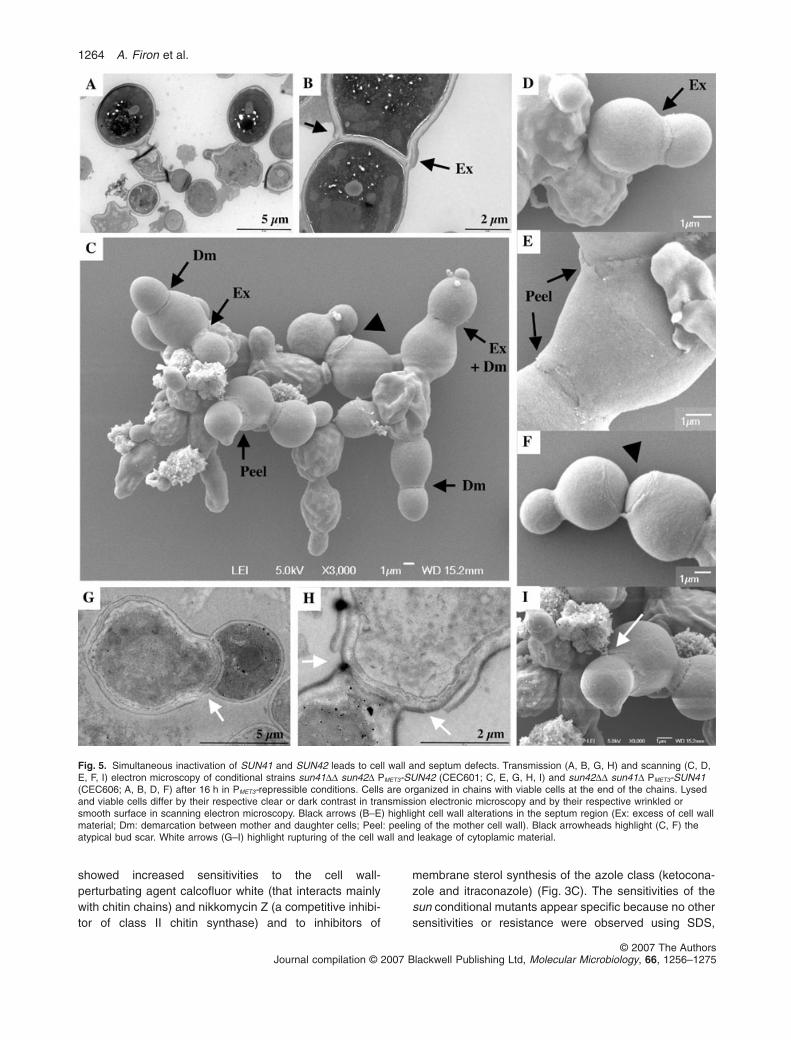
Cell wall and septum structures were investigated by elec-tronic microscopy in the conditional sun41DD sun42DPMET3-SUN42 and sun42DD sun41D PMET3-SUN41 mutantsafter incubation in PMET3-repressing condition (Fig. 5). Asobserved previously by optical microscopy, lysed cellswere organized in chains with some viable cells at the endof the chains. Lysed and viable cells differed by theirrespective clear or dark contrast in TEM and by theirrespective wrinkled or smooth surface in scanning elec-tron microscopy (SEM).
Although high morphological heterogeneity was ob-served, viable and lysed cells remained interconnectedthrough their septa (Fig. 5). TEM revealed that septumstructures were heterogeneous with the marked absenceof a clear trilaminar structure and examples of very largesepta (Fig. 5A). Yet, near wild-type septum structureswere observed between viable cells (Fig. 5B). However, inthese cases, a wider cell wall was often detected in themother cells with lateral expansions that projected beyondthe septum (Fig. 5B). These observations suggested thata compensatory cell wall salvage pathway is activated inmother cells to strengthen the cell wall and the septum onthe mother cell side.
Observation by SEM of the SUN conditional mutantsrevealed (i) an excess of cell wall material originating fromthe mother cell side and surrounding the septum (Fig. 5Cand D), (ii) a demarcation between mother and daughtercells indicative of the initiation of the cell separationprocess (Fig. 5C) and (iii) ‘peeling’ of the mother cell wallin the area surrounding the septum (Fig. 5C and E).Notably, atypical bud scars were visible, with a largedome-shaped structure readily different of that normallyseen in wild-type cells and the absence of the typicalchitin ring (Fig. 5C and F).
Electronic microscopy did not allow to unambiguouslyuncover the site(s) associated with cell lysis. However,rupturing of the cell wall was observed by TEM predomi-nantly at the junction between the mother cell wall and theseptum (Fig. 5G and H) at sites that may correspond tothose where cell wall ‘peeling’ was observed by SEM.These might represent an enfeebled area on the cell wallwhere leakage of cytoplasmic material can occur as seenin Fig. 5I. Other instances of cell wall damage wereobserved at the junction between mother and daughtercell wall before septum synthesis and at the new bud tips(data not shown). Taken together, our observations sug-gested that inactivation of the C. albicans SUN genes wasassociated with a defect in cell wall biogenesis in the
vicinity of the septum and with activation of a cell wallsalvage pathway that could not prevent their lysis uponinitiation of a new cell cycle (see also time-lapse movies inSupplementary material ).
Conditional lethality of the SUN genesis osmoremediable
A cell wall defect associated with the simultaneous inac-tivation of the C. albicans SUN genes was suggested bythe cell lysis phenotypes, aberrant chitin distribution in theconditional mutant strains and the structure of the cell walland the septum in mother cells (Figs 4 and 5). Pheno-types of C. albicans mutants with defects in the cell wallare usually suppressed by increasing the osmolarity of thegrowth medium. Addition of 1 M sorbitol to the mediumcould not restore growth of the sun41DD sun42D PMET3-SUN42 and sun42DD sun41D PMET3-SUN41 conditionalmutants grown in minimal medium under PMET3-repressingcondition (SD + 5 mM methionine + 0.25 mM cysteine,data not shown). In contrast, growth restoration by addi-tion of 1 M sorbitol was observed when these mutantswere grown in rich medium (YPD) that results in partialgrowth inhibition (Fig. 3B). Under this condition of partialrepression of the PMET3 promoter, suppression of thegrowth defect by osmostabilisation is consistent with adefective cell wall in strains where SUN41 and SUN42genes are simultaneously inactivated.
Drug sensitivities of sun mutants
The sun41 and sun42 heterozygous and homozygousmutants were screened for sensitivity and resistance to aset of chemical agents: oxidative stress reagents (H2O2,tert-butyl, cumene, menadione), cell wall-perturbatingagents (calcofluor white, SDS, polyoxin D, nikkomycin Z),antifungal drugs (amphotericin B, ketoconazole, itracona-zole), a protein synthesis inhibitor (cycloheximide) andcell signalling inhibitors (FK506, rapamycin, cyclosporinA, staurosporine). No significant sensitivity or resistancewere detected except a slight but reproducible sensitivityof the sun42DD strain for calcofluor white and rapamycin(data not shown). Triple mutants (sun41DD sun42D andsun42DD sun41D) did not show additional phenotypesexcept for the sun42DD sun41D strain that exhibitedincreased sensitivity to rapamycin relative to the homozy-gous sun42DD mutant (data not shown). This was consis-tent with a genetic link between SUN41 and SUN42 andan additional support for the haploinsufficiency due to theinactivation of one allele of SUN41.
Taking advantage of the partial growth defect observedin rich media, sensitivities of the conditional sun41DDsun42D PMET3-SUN42 and sun42DD sun41D PMET3-SUN41mutants to different drugs were tested. Both mutants
SUN genes of C. albicans 1263
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275
showed increased sensitivities to the cell wall-perturbating agent calcofluor white (that interacts mainlywith chitin chains) and nikkomycin Z (a competitive inhibi-tor of class II chitin synthase) and to inhibitors of
membrane sterol synthesis of the azole class (ketocona-zole and itraconazole) (Fig. 3C). The sensitivities of thesun conditional mutants appear specific because no othersensitivities or resistance were observed using SDS,
Fig. 5. Simultaneous inactivation of SUN41 and SUN42 leads to cell wall and septum defects. Transmission (A, B, G, H) and scanning (C, D,E, F, I) electron microscopy of conditional strains sun41DD sun42D PMET3-SUN42 (CEC601; C, E, G, H, I) and sun42DD sun41D PMET3-SUN41(CEC606; A, B, D, F) after 16 h in PMET3-repressible conditions. Cells are organized in chains with viable cells at the end of the chains. Lysedand viable cells differ by their respective clear or dark contrast in transmission electronic microscopy and by their respective wrinkled orsmooth surface in scanning electron microscopy. Black arrows (B–E) highlight cell wall alterations in the septum region (Ex: excess of cell wallmaterial; Dm: demarcation between mother and daughter cells; Peel: peeling of the mother cell wall). Black arrowheads highlight (C, F) theatypical bud scar. White arrows (G–I) highlight rupturing of the cell wall and leakage of cytoplamic material.
1264 A. Firon et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

H2O2, FK506 and rapamycin when comparing conditionalmutants with the control strain, the homozygous mutantsand with their parental triple mutants (data not shown).These drug sensitivities suggested that chitin biosynthe-sis or remodelling was probably necessary to compensatecell wall or septum defects associated with inactivation ofSUN genes.
The essential function of the Sun proteins is conservedamong yeasts
Data presented above show that Sun proteins share acommon function that is essential for C. albicans growth.In order to test whether this function is conserved amongyeasts, the four S. cerevisiae SUN genes (SIM1, UTH1,NCA3 and SUN4) were cloned into a C. albicans integrat-ing vector (CIp10S) and introduced into the genome ofthe C. albicans conditional sun mutants. Under PMET3-repressing conditions, the presence of one copy of UTH1expressed under the control of its own promoter restoredviability of the conditional sun mutants (Fig. 3A and B).Complementation by UTH1 was highest in the sun42DDsun41D PMET3-SUN41 strain (Fig. 3A and B). Thesedata demonstrated the functional conservation of SUNgenes among yeasts. Introduction of the three otherS. cerevisiae SUN genes (NCA3, SIM1 and SUN4) in theC. albicans conditional sun mutants barely complementedthe lethality associated with the conditional inactivation ofthe C. albicans SUN genes (data not shown). However,this resulted from very low expression as revealed byquantitative RT-PCR (data not shown).
SUN genes are also essential for hyphal growth inC. albicans
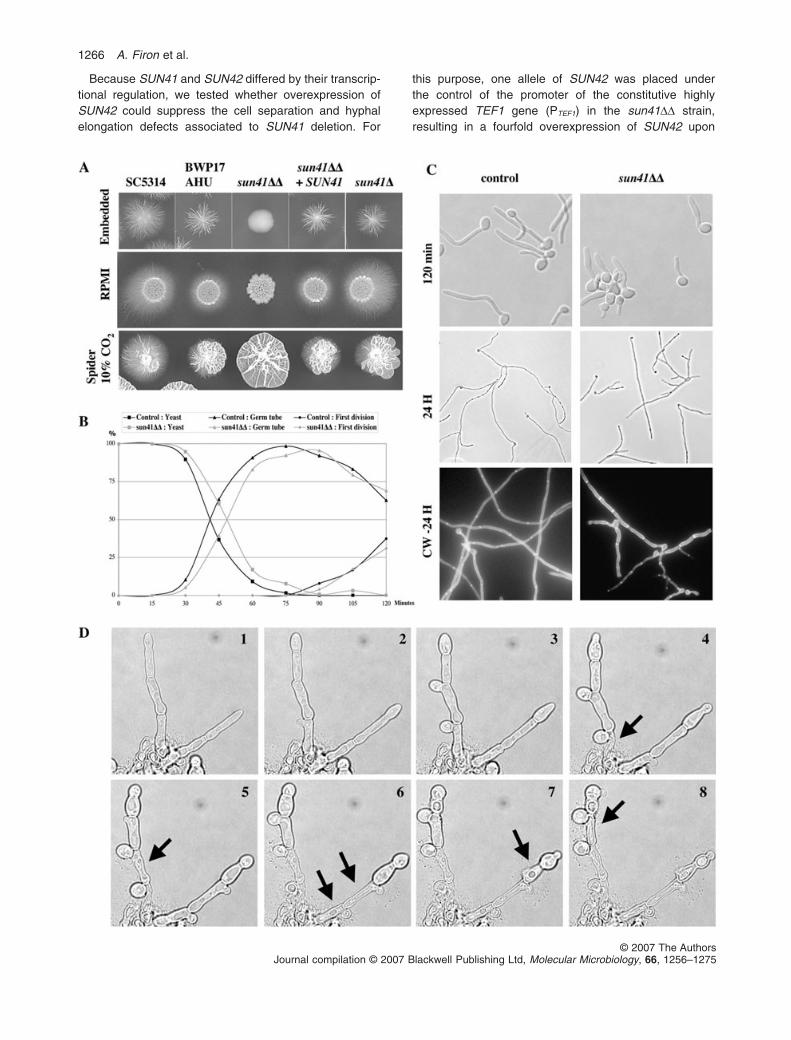
The ability of the C. albicans heterozygous and homo-zygous sun41 and sun42 mutants to switch from theyeast to hyphal forms was evaluated on different solidmedia (RPMI 0.5¥, YPD + 10–20% serum and Spider at37°C � 10% CO2, and embedded condition at roomtemperature). In all conditions tested, the sun41DD strainwas defective for filamentation (Fig. 6A). This filamenta-tion defect resulted from the inactivation of SUN41because reintroduction of a wild-type allele of SUN41in sun41DD mutants restored wild-type filamentation(Fig. 6A). The other mutant strains (sun41D, sun42D andsun42DD) were all able to form filaments similar to thecontrol strain (Fig. 6A and data not shown).
Microscopic observations of the sun41DD strain under-going the yeast-to-hypha switch in liquid medium revealedthat while germ tube emergence was not significantlyaffected by inactivation of the SUN41 gene (Fig. 6B),germ tube elongation was impaired. As shown in Fig. 6B,100% of the wild-type and sun41DD yeast cells had pro-duced a germ tube 75–90 min following transfer of yeasts
in hypha-inducing medium. Defects in filamentation of thesun41DD mutant arose shortly after completion of the firstcell cycle and the appearance of the first septum (Fig. 6C,t = 120 min). At this stage, hyphae of the sun41DD mutantbecame more heterogeneous than those of the controlstrain (Fig. 6C). These defects lead to heterogeneoushyphae characterized by irregular length, diameter andbranching (Fig. 6C). A likely consequence of this filamen-tation defect was the inability of the sun41DD mutant toform a biofilm in microfermentor and microtiter platemodels (data not shown).
Time-lapse microscopy of conditional sun mutantspresented above (Fig. 4B) was conducted at ambienttemperature and between a layer of solid medium and acoverslip. These represent a hypoxy-like microenviron-ment (similar to that in embedded growth) that promotesfilamentation. Thus, filament-like growth was observed insome microscope fields. As shown in Fig. 6D (and inassociated movie in Supplementary material ), simulta-neous inactivation of SUN41 and SUN42 resulted inhyphal lysis. Apical hyphae maintained growth despitelysis of the subapical compartment (Fig. 6D). Lysis wasoften observed after the emergence of a branching budclose to the apex-proximal septum (Fig. 6D). Therefore,SUN41 and SUN42 are also required for cellular integrityin subapical compartments, a role that is only revealedwhen these compartments initiate a new cell cycle.
Functional redundancy of SUN41 and SUN42
Results presented above show that inactivation of SUN41is associated to specific defects in cell separation andhyphal morphogenesis, suggesting that Sun41 mayperform additional functions that are not shared withSun42. We first investigated this hypothesis throughanalysis of SUN41 and SUN42 expression upon yeastand hyphal growth. Quantification of transcript levels byquantitative RT-PCR showed that SUN41 and SUN42 areexpressed at similar levels upon yeast growth (Fig. 7A).Upon hyphal growth, SUN41 is upregulated 4.9-fold andSUN42 is downregulated 25-fold (Fig. 7A), consistent withpreviously published data (Sohn et al., 2003). These re-sults were confirmed through examination of C. albicansstrains where the coding region of one allele of SUN41or SUN42 had been replaced by the green fluorescentprotein (GFP)-coding region. PSUN41- and PSUN42-drivenexpression of GFP was observed in yeast cells with noapparent restriction to the mother or daughter cells(data not shown). Upon hyphal growth, PSUN42-drivenexpression of GFP was restricted to the basal cells ofhyphae for up to two cell cycles while PSUN41-drivenexpression of GFP was detected predominantly in theactively growing apical and subapical compartments ofhyphae (Fig. S1).
SUN genes of C. albicans 1265
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275
Because SUN41 and SUN42 differed by their transcrip-tional regulation, we tested whether overexpression ofSUN42 could suppress the cell separation and hyphalelongation defects associated to SUN41 deletion. For
this purpose, one allele of SUN42 was placed underthe control of the promoter of the constitutive highlyexpressed TEF1 gene (PTEF1) in the sun41DD strain,resulting in a fourfold overexpression of SUN42 upon
1266 A. Firon et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

yeast growth and a 1000-fold overexpression upon hyphalgrowth (Fig. 7A). PTEF1-driven expression of SUN42 re-stored filamentation in liquid and on solid media and cellseparation to almost wild-type levels (Fig. 7B and data notshown). Thus, when overexpressed, Sun42 is able to fulfilall of the functions of Sun41. Yet, while low expression ofSUN42 upon hyphal growth may account for the observedfilamentation defect of the sun41DD mutant, this does notappear to explain the separation defect of this mutantwhen grown under the yeast form.
Discussion
Results presented in this study indicate that the Sun41pand Sun42p proteins play a critical role during the sepa-ration of cellular compartments in C. albicans. Inactivationof SUN41 results in cell separation and hyphal elongationdefects. These defects appear following septation duringboth yeast and hyphal growth, resulting in the formationof yeast chains or hyphae with shorter and enlargedcompartments. Simultaneous inactivation of SUN41 andSUN42 results in similar defects that are exacerbated bythe localized lysis of yeast mother cells and subapicalcompartments in hyphae. Because SUN41 and SUN42share an interchangeable essential function and becauseSUN42 can fulfil all the functions of SUN41 when overex-pressed and is expressed predominantly in yeast cells incontrast to SUN41 that is expressed in yeast and upregu-lated in actively growing hyphal cells, we believe that bothgene products play a similar function.
We would like to propose a model whereby Sun pro-teins are involved in the remodelling of the cell wall in theseptum area. Septum formation is a timely process involv-ing the following steps: (i) the formation of a chitin ring atthe emerging bud site, (ii) the synthesis of a chitin discphysically separating the mother and the daughter cellsduring cytokinesis, (iii) the formation of secondary septa inboth cells and (iv) the degradation in yeast cells only ofthe primary septum by the combined action of specific
chitinases and glucanases (Cabib, 2004; Yeong, 2005). Inyeast cells, this last stage of the cell separation processinvolves a balance between the action of chitinases andglucanases and chitin synthases to complete cell separa-tion without cell lysis. In hyphae, the dynamics of septumformation and maturation are less documented but thekey difference is the absence of septum degradation.Several lines of evidence concur to support a directinvolvement of Sun proteins in a localized strengtheningof the cell wall in the region surrounding the septum on themother cell side to avoid cell lysis during the next cellcycle that necessitates cell wall remodelling.
Sun proteins are involved in cell separation
A role in cell wall biogenesis and cell separation has alsobeen suggested for other members of the Sun family.Mouassite et al. (2000) have shown that the S. cerevisiaeSun4 protein is involved in cell separation. Electronmicroscopy of sun4D mutant cells showed that daughtercells remained anchored to mother cells by the primarychitin ring of the septum, a phenotype reminiscent of thatof the C. albicans sun41 mutant (Mouassite et al., 2000).Moreover, a uth1 null mutation aggravates the septationphenotype of a sun4 mutant (Mouassite et al., 2000).Here, we have shown that Uth1p can substitute for theessential function shared by Sun41p and Sun42p inC. albicans, suggesting that Uth1p performs a similarfunction in S. cerevisiae. Similarly, deletion of theS. pombe psu1 + gene results in cell wall and cell separa-tion defects resulting in cell lysis (Omi et al., 1999). Thus,a subset of the ascomycetous Sun protein, if not all giventheir phylogenetic link, may share an essential function inthe remodelling of the cell wall that occurs upon septation.
Sun proteins are necessary for viability of mother cells
A striking observation made in this study is that inactiva-tion of C. albicans SUN41 and SUN42 results in prefer-
Fig. 6. Candida albicans SUN41 is involved in hyphal elongation and SUN41 and SUN42 are necessary during branching.A. Filamentation defect of the DDsun41 homozygous mutant on solid media. Cells of control strains SC5314 (SUN41/SUN41) and BWP17AHU(SUN41/SUN41), homozygous mutant sun41DD (CEC666), complemented mutant sun41DD + SUN41 (CEC667) and heterozygous mutantsun41D (CEC690) were embedded in YPS agar medium, spotted on RPMI agar medium and on Spider agar medium. Photos were taken after6 (Spider) or 7 days (Embedded, RPMI) of incubation at room temperature (Embedded) or at 37°C (RPMI, Spider) in the presence of 10%CO2 when indicated.B. Kinetics of germ tube emergence and nuclear division of C. albicans wild-type and DDsun41 strains. Strain BWP17AHU (SUN41/SUN41)and homozygous mutant sun41DD (CEC666) were grown 48 h in YPD at 30°C and diluted in RPMI medium at 37°C. At different timeintervals, the number of yeast cells, of cells with a germ tube and of cells having completed one division (i.e. harbouring two nuclei) wasrecorded. An average of 100 cells are counted for each time point and the graph is a mean of two independent experiments.C. Representative microscopic pictures of C. albicans wild-type and sun41DD strains. Representative pictures at T = 120 min and after 24 h ofthe experiment described in B are shown. CW, calcofluor white. The sun41DD mutant showed more heterogenous morphology compared withthe control strain starting at T = 120 min corresponding to the completion of the first cell cycle.D. Time-lapse microscopy of a conditional sun mutant grown under repressing conditions. Filamentation phenotype were observed in somemicroscopic field during time-lapse movie done as in Fig. 4B. Representative serial images of cells of the conditional sun mutant sun42DDsun41D PMET3-SUN41 (CEC606) on repressing agar medium were taken at 1 h intervals. Sequential lysis of hyphal compartment followingbranching is marked by black arrows. Movies are available in Supplementary material.
SUN genes of C. albicans 1267
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

ential lysis of yeast mother cells or subapical hyphalcompartments following separation from the yeast daugh-ter cell or compartmentalization of the apical hyphae. Thissuggests that the cell wall of the primary cells is specifi-cally weakened. Entry of these primary cells into a newcell cycle may in particular favour cell lysis. That lysis ofthe conditional sun mutants is osmoremediable on YPDalso argues for a role of the Sun proteins in cell wallbiogenesis because osmoremediation is frequentlyobserved for essential genes involved in cell wallbiogenesis.
Mother cells are characterized by a septum that is sen-sitive to degradation by enzymes synthesized in thedaughter cell and a chitin ring that identifies the bud scaronce separation is achieved. Electron microscopy of sunconditional mutants revealed that cell wall defects aremainly restricted to mother cells and appear mainly aftersynthesis of the septum because daughter cells have fewobservable defects. Rupturing of the cell wall was local-ized mainly at its junction with the septum in an enfeebledarea on the mother cell side. Moreover, the ‘bud scar’ insun conditional mutants adopted an atypical dome-shaped structure in the few cases of near complete cellseparation. These defects are likely to result from amodification of the chitin ring that becomes accessible todegrading enzymes and/or an uncontrolled activity of thecell wall-degrading enzymes, leading to a fragile area inthe mother cell wall at the level of the septum. Initiation ofa new cell cycle and cell wall expansion would result inlysis at this site. The same hypotheses may account forthe lysis phenotype observed at subapical hyphalcompartments. Indeed, cell wall-degrading enzymes arelikely to act in the vicinity of the septum upon branchingand may either exacerbate a pre-existing cell wall defector degrade a defective septum resulting in lysis.
The SUN genes are linked to the RAM–Ace2p pathwaycontrolling cell wall-degrading enzymes specificallyinvolved in septum degradation
As already introduced, the S. cerevisiae and S. pombeSUN genes are linked to the RAM–Ace2p signallingpathway that regulates the expression specifically indaughter cells of genes encoding cell wall-degradingenzymes involved in septum degradation along with anAce2p-independent polarization process. Interestingly, intwo independent studies, S. cerevisiae SIM1 has beenidentified as a dosage suppressor of the lethality associ-ated to inactivation of components of the RAM pathway(Du and Novick, 2002; Kurischko et al., 2005). BecauseSIM1 and other dosage suppressors identified in thesescreens rescue the cell lysis phenotype by altering cellwall composition but do not restore daughter-specificlocalization of Ace2p nor suppress the associated cell
Fig. 7. SUN41 and SUN42 are functionally redundant.A. Relative quantification of SUN41 and SUN42 mRNA duringyeast growth (4 h in YPD at 30°C) and hyphal growth (4 h in RPMIat 37°C). Transcript levels in the control strain BWP17AHU(SUN41/SUN41 SUN42/SUN42), the sun41DD mutant (CEC666),the sun42DD mutant (CEC668) and a sun41DD mutantoverexpressing SUN42 (sun41DD PTEF1-SUN42, CEC849) weredetermined by two-step quantitative RT-PCR. Results arerepresented as a relative quantity of transcript compared with 1000arbitrary units of the EFT3 transcript used as the control for eachsample. Each sample was analysed in triplicate for four dilutionsand the mean of at least 9-values at three different dilutions areindicated.B. Overexpression of SUN42 suppresses the specific defects of ahomozygous sun41DD mutant. Comparison of the filamentationdefect on solid medium (RPMI 0.5x), and the cell separation defect,quantified by the relative half-sedimentation time in YPD, ofsun41DD mutant (CEC666) and a sun41DD mutant overexpressingSUN42 (CEC849). Similar results were obtained for twoindependent overexpression strains confirming that suppression ofthe sun41DD mutant phenotypes was the result of SUN42overexpression.
1268 A. Firon et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

separation defects, it has been proposed that they actdownstream of the RAM–Ace2p signalling pathway in acell wall-dependent mechanism (Kurischko et al., 2005).Transcript profiling data link the transcription factor Ace2pto the expression of S. cerevisiae SUN4 (Colman-Lerneret al., 2001; Nelson et al., 2003), NCA3 (Bidlingmaieret al., 2001), S. pombe psu1 + (Alonso-Nunez et al., 2005)and C. albicans SUN41 (Mulhern et al., 2006). Yet, adirect regulation of the SUN genes by Ace2p remains tobe demonstrated.
Taken together, these genetic and transcriptionalobservations are in favour of the SUN genes acting down-stream of the RAM–Ace2p signalling pathway. As RAM–Ace2p signalling pathway controls daughter-specificexpression of genes encoding enzymes that degrade theseptum on the mother cell side and C. albicans SUNgenes are necessary to maintain mother cell wall integrityfollowing cell separation, it is possible that the Sunproteins might be involved in counteracting the actionof the chitinase (ScCts1p or CaCht3p) and glucanase(ScScw11p or CaEng1p) through strengthening of the cellwall at the level of the septum.
Sun proteins are localized at the cell surface
A common feature of Sun proteins is the presence of asignal peptide at their amino terminal end suggesting thatthese proteins are targeted to the secretory pathway andmay reach the cell wall (Fig. 1A). A cell wall localizationof several Sun proteins has been demonstrated inS. cerevisiae and S. pombe. Biotinylation of intactS. cerevisiae cells followed by DTT treatment identifiedseven mannosylated proteins non-covalently bound to thecell wall, including Sun4p (referred as Scw3p in this study)(Cappellaro et al., 1998). Using fusion proteins, Sun4p,Uth1p and Sim1p have been shown to be released fromthe cell wall by DTT treatment although a mitochondriallocation was also proposed for Sun4p and Sim1p (Velourset al., 2002). Sim1p has also been independentlydescribed as a secreted protein following its detectionmainly in culture supernatants (Du and Novick, 2002). InS. pombe, Psu1p was recently identified in the cell wall bymass spectrometric analysis and proposed to becovalently bound to cell wall glucans via mild alkali sen-sitive linkages (de Groot et al., 2007). Despite thesebiochemical evidences for a cell wall localization ofS. cerevisiae and S. pombe Sun proteins, attempts tolocalize them in vivo using GFP fusion proteins has beenunsuccessful (Velours et al., 2002; Huh et al., 2003;Alonso-Nunez et al., 2005).
In C. albicans, the signal peptide of Sun42p is func-tional according to a large-scale screen for exportedproteins (Monteoliva et al., 2002). Despite repeatedattempts, we have been unsuccessful at visualizing
Sun41p-GFP and Sun42p-GFP fusion proteins or detect-ing TAP-tagged Sun41p and Sun42p fusion proteins byimmunofluorescence (data not shown). However, theTAP-tagged Sun41p fusion protein is functional and hasa molecular weight higher than anticipated (data notshown). This fusion protein is insensitive to treatment withN-glycosidase, suggesting that it is O-glycosylated, pos-sibly in a Ser/Thr-rich region located after the signalpeptide cleavage site. The three S. cerevisiae Sun4p,Uth1p and Sim1p proteins are also suspected to be gly-cosylated based on their affinity to concanavalin A or onimmunodetection experiments (Cappellaro et al., 1998;Velours et al., 2002). The presence of a signal peptide,the probable glycosylation of the proteins and the inde-pendent identification of Sun proteins in the cell wall,argue for a cell wall localization of Sun proteins.
Chitin synthesis compensates defects of sun mutants
Drug sensitivity of the C. albicans strains with residualexpression of only one of the two SUN genes revealed anincreased sensitivity to cell wall-perturbing agents suchas calcofluor white and nikkomycin Z, two compoundsrelated to chitin biosynthesis. As modulation of theexpression of an essential gene often leads to hypersen-sitivity to chemical compounds acting on the cognategene product (Roemer et al., 2003; Lum et al., 2004;Mnaimneh et al., 2004), it seems likely that inactivation ofthe SUN genes is linked to chitin biosynthesis. Yet, thislink might be indirect because changes in the cell wall dueto the lack of the Sun proteins may result in altered chitinbiosynthesis triggered by a cell wall salvage pathway(Levin, 2005) or by a compensatory mechanism. Elec-tronic microscopy of conditional C. albicans sun mutantssuggests that cell wall biogenesis is stimulated in yeastmother cells after septum synthesis and the beginning ofthe cell separation process possibly as a means to coun-teract defects in the septum and the surrounding area.Such induction of a cell wall salvage pathway followinginactivation of the SUN41 and SUN42 genes may alsocontribute to the increased sensitivity of the conditionalsun mutants to the azole class of drugs. It should be notedthat we could not test whether strains with conditionallyinactivated SUN genes showed altered sensitivities toamphotericin B, an antifungal related to sterol synthesis,and caspofungin, a b-(1,3)-glucan synthase inhibitor.Indeed, the MET3 gene is upregulated upon exposure ofC. albicans to these two antifungals (Liu et al., 2005), thuspreventing repression of the PMET3-SUN41 and PMET3-SUN42 gene fusions.
Interestingly, a link between S. cerevisiae SUN genesand chitin biosynthesis has also been establishedbecause uth1 and chs1 null mutations are syntheticallylethal (Lesage et al., 2005). CHS1 encodes the chitin
SUN genes of C. albicans 1269
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

synthase that is specifically required for repairing the cellwall during cell separation. Deletion of CHS1 led to thepreferential lysis of the daughter cell (Bulawa et al., 1986),a phenotype suppressed by inactivation of the chitinaseCTS1 encoding gene (Cabib et al., 1992). Thus, UTH1seems to cooperate with CHS1, or act in a parallelpathway, to strengthen the septum during cell separationand counteract the action of chitinases and glucanases.Noteworthy, C. albicans has two orthologues of ScCHS1,the class I chitin synthase encoding genes CaCHS2 andCaCHS8, none of them showing the bud lysis phenotypewhen deleted (Munro et al., 2003).
Toward the function of Sun proteins
Saccharomyces cerevisiae Sun proteins have been pro-posed to participate in other processes such as oxidativestress response, mitochondrial biogenesis and autophagy(Camougrand et al., 2003; 2004; Kissova et al., 2004).Yet, the combined evidences presented above warrant aprimary role of the Sun proteins in strengthening the cellwall of yeast mother cells or subapical hyphal compart-ments in the area surrounding the septum. The precisefunction of Sun41 and Sun42 in modifying cell wall poly-mers remains to be investigated. The purification of func-tional tagged Sun41 and Sun42 proteins will enable us totest whether these proteins directly catalyse cell wallpolymer biosynthesis or cross-linking or are involved inthe activation of the chitin synthase that is specificallyrequired to repair the septum during cell separation.
Our study provides new insights on the function of theSun proteins in C. albicans and other ascomycetes. Manyquestions remain open in order to establish the biochemi-cal role of the Sun proteins. Our experiments usingPSUN41–GFP and PSUN42–GFP gene fusions have notshown daughter-specific expression of the SUN41 orSUN42 genes and it remains to be established whetherthe link between these genes and the RAM–Ace2ppathway might lie in regulated secretion of the Sun pro-teins rather than daughter-specific expression (Kurischkoet al., 2005). This is likely because Sun41p, as well asseveral other members of the Sun protein family, hasfeatures of preproproteins that are processed by theKex2p protease (Newport et al., 2003). In this regard, acareful study of the localization of the Sun proteins duringthe cell cycle and hyphal growth should provide furtherinsights on their common or specific functions and theirregulation. Finally, because the Sun proteins appear cellwall located, essential for sustained growth of C. albicansand ascomycete-specific, they may constitute relevanttargets for the development of novel antifungals that couldbe used individually or in combined therapy with azoles orchitin synthesis inhibitors. Although the occurrence of twoSun proteins in C. albicans may be seen as a limitation for
such a development, the example of caspofungin target-ing b(1,3)-glucan synthase shows that drugs can be usedas antifungal despite the presence of two isoenzymes(Fks1p and Fks2p in S. cerevisiae).
When this manuscript was under revision, two articlesreporting the characterization of C. albicans sun41D/sun41D mutants have been accepted for publication(Hiller et al., 2007; Norice et al., 2007). Consistent withour observations, these reports show that SUN41 isrequired for separation of yeast mother and daughtercells, hyphal differentiation and biofilm formation. Interest-ingly, these authors have shown that inactivation ofSUN41 results in hypersensitivity to caspofungin andcongo red, two drugs that we had not included in ourpanel (see above), and is associated with altered expres-sion of several cell wall damage response genes, support-ing our conclusion that the main function of the Sunproteins is in cell wall biogenesis. Finally, Norice et al.(2007) have shown that SUN41 is necessary for virulenceof C. albicans in mouse models of oropharyngeal anddisseminated candidiasis, reinforcing the potential ofSun proteins as targets for the development of novelantifungals.
Experimental procedures
Strains, media and growth conditions
C. albicans strains used in this study are listed in Table 1. Allstrains were constructed in the BWP17 background, an aux-otrophic derivative of the clinical isolate SC5314 (Wilson et al.,1999). Yeast growth was routinely done at 30°C in YPD (1%yeast extract, 2% peptone, 2% glucose) or in SD minimalmedium (0.67% yeast nitrogen base without amino acids, 2%glucose, pH 5.4) supplemented with uridine (80 mg ml-1), argi-nine (50 mg ml-1) and/or histidine (20 mg ml-1) when needed.Repression of the PMET3 promoter was achieved with 5 mMmethionine and 0.25 mM cysteine in SD. Filamentation inliquid was assayed in RPMI medium (Gibco) buffered with50 mM HEPES pH 7.3 at 37°C with 150–200 r.p.m. shaking ona rotary shaker. Filamentation on plate were assayed onSpider medium (Liu et al., 1994), on YPD supplemented with10–20% fetal bovine serum (Sigma) and on 0.5¥ RPMI (bymixing an equal volume of buffered RPMI medium with 3%agar solution) at 37°C under atmospheric or 10% CO2 concen-tration. Filamentation in embedded condition was done byembedding yeast cells between two layers of YPS medium(1% yeast extract, 2% peptone, 2% saccharose, 1% agar).
Plasmids for PCR-based gene replacementsand fusions
To generate transformation cassettes for gene replacementand fusion, pFA plasmids were used as previously described(Gola et al., 2003). This PCR-based system relies on thegeneration of transformation cassettes amplified from pFAplasmids with long oligonucleotides (synthesized by Biopoly-
1270 A. Firon et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

mers) of 100 nucleotides of the target sequence followed bya short (20–28 nucleotides) sequence homologous to theplasmid template (Table S1). PCR products were ethanolprecipitated, concentrated 10 times and 10–20 ml was rou-tinely used for yeast transformation by the acetate lithium/PEG method with a 15 min 44°C heat shock as describedpreviously (Walther and Wendland, 2003). Gene replacementby a fungal marker (ARG4 or HIS1) or by a GFP open readingframe (ORF), promoter exchange (PMET3) and 3′ end GFPtagging have been made with the already published plasmids(Gola et al., 2003) using long oligonucleotides listed inTable S1.
In the course of this study, we constructed an additional setof pFA vectors for overexpression purpose. The PmeI-SpeIfragment containing the MET3 promoter of pFA-ARG4-MET3p and pFA-HIS1-MET3p (Gola et al., 2003) wasreplaced by a PmeI-SpeI PCR-generated fragment (primersTEFpPme and TEFpSpe; all primers used for cloning arelisted in Table S2) containing the strong and constitutiveTEF1 promoter of C. albicans. These pFA-ARG4-TEF1p andpFA-HIS1-TEF1p plasmids allow the generation of PCR cas-settes for promoter replacement with the use of long oligo-nucleotides S2-Prom TEF1 (Tables S1 and S2) as previouslydescribed (Gola et al., 2003).
Integrative plasmids for mutant complementation
The integrative vector CIp10 (Murad et al., 2000) was modi-fied by the introduction of an adaptator bearing an I-SceIcleavage recognition site at the unique NcoI site by usingthe complementary primers Nco-5Sce and Nco-3Sce. Theresulting CIp10S plasmid can be linearized prior to yeasttransformation by the rare-cutting endonuclease I-SceI(New England BioLabs) regardless of insert sequence clonedin this vector.
Complementing plasmids were obtained by cloning PCRproducts generated with high-fidelity Taq polymerase (Triple-Master Eppendorf). BWP17 genomic DNA was used as tem-plate with primers 5′CIP41 + 3′CIP41 and 5′CIP42 + 3′CIP42to clone SUN41 (with 2381 bp upstream of the start codonand 351 bp downstream of the stop codon) and SUN42 (with800 bp upstream of the start codon and 449 bp downstreamof the stop codon) respectively. PCR fragments were clonedas KpnI-HindIII sites in CIp10S (for SUN41) or CIp10 (forSUN42) to give CIp-SUN41 and CIp-SUN42.
The four S. cerevisiae SUN genes were PCR amplifiedfrom BY4743 genomic DNA as template. SIM1 (from 751 bpupstream of the start codon to 262 bp downstream of the stopcodon, primers Cip-5′sim + Cip-3′sim, 2442 bp fragment),UTH1 (from 723 bp upstream of the start codon to 467 bpdownstream of the stop codon, primers Cip-5′uth + Cip-3′uthand 5′uth-R5 + 3′uth-R5, 2442 bp fragments), NCA3 (from526 bp upstream of the start codon to 355 bp downstream ofthe stop codon, primers Cip-5′nca + Cip-3′nca, 1896 bp frag-ment) and SUN4 (from 629 bp upstream of the start codonto 235 bp downstream of the stop codon, primers Cip-5′sun4 + Cip-3′sun4, 2138 bp fragment) were cloned inCIp10S as HindIII (SIM1, NCA3, SUN4) or ClaI or EcoRV (forUTH1) fragments to give CIp-SIM1, CIp-UTH1, CIp-NCA3and CIp-SUN4. As already observed, plasmids with clonedUTH1 are toxic and unstable in Escherichia coli at 37°C
(Bandara et al., 1998). Therefore, bacterial cultures weredone at room temperature prior to CIp-UTH1 plasmidpreparation. The plasmids were carefully checked by PCR forabsence of rearrangement (data not shown).
C. albicans homozygous and conditionalmutant construction
sun mutants were obtained by coupling two techniques: thefirst allele was deleted using a SAT-flipper cassette contain-ing a recyclable nourseothricine-resistance marker (Reusset al., 2004) and the second allele using a PCR-based dele-tion cassette with an auxotrophic gene (ARG4 or HIS1) astransformation marker (Gola et al., 2003). This strategy waschosen in order to construct further isogenic double mutantsfor SUN41 and SUN42. Correct integration of deletion cas-settes and presence or absence of wild-type copy of SUNgenes were checked by analytical PCR (data not shown). Toavoid phenotypic variation due to auxotrophy, mutants wererendered prototroph for amino-acid and nucleoside prior tophenotypic characterization. In particular, uracil prototrophywas restored by the introduction of a copy of the URA3 geneat the RPS10 loci with the CIp10 or CIp10S plasmids (Muradet al., 2000). Several independent homozygous mutantswere generated and no phenotypic discrepancy wereobserved between them. Therefore, only one homozygousmutant for each SUN gene was fully characterized pheno-typically and presented in this study.
The SAT-flipper cassette used to make targeted deletionof SUN41 and SUN42 were constructed by a two-stepapproach. First, SUN41 and SUN42 ORFs were PCRamplified (primers ApaI-5′sun41 + SacI-3′sun41 and ApaI-5′sun42 + SacI-3′sun42) from BWP17 genomic DNA withadditional 557 bp and 605 bp at the 5′ end of the ORF and549 bp and 505 bp at the 3′ end respectively, and cloned asa ApaI-SacI fragment into pBluescript KS to give pB-SUN41and pB-SUN42 respectively. Inverse PCR was performedon pB-SUN41 and pB-SUN42 with primers IPCR41-SacII + IPCR41-XhoI (located at 25 bp and 36 bp of the startand stop codon respectively, and oriented in the oppositedirection of the SUN41 ORF) and IPCR42-SacII + IPCR42-XhoI (located at 60 bp and 23 bp of the start and stop codon,respectively, of the SUN42 ORF) respectively. The IPCRproducts were digested by SacII and XhoI and cloned with aSacII-XhoI fragment of pSFS2A (Reuss et al., 2004) con-taining the nourseothricine-flipper cassette. The resultingpD41::SAT2A and pD42::SAT2A plasmids were digested withSacI and ApaI to released the SAT flipper cassette flanked bySUN genes homologous sequences (557 and 605 bp ofupstream sequence and 649 and 505 bp of downstreamsequence for SUN41 and SUN42 respectively). SacI-ApaIfragments were gel purified and concentrated for yeasttransformation. After heat shock, C. albicans cells wereplated on YPD, incubated overnight at 30°C and nourseothri-cine (Werner Bioagent) resistant colonies were selected byreplica-plating on YPD + 150 mg nourseothricine ml-1. Whenneeded, excision of the SAT-flipper cassette by recombina-tion at the flanking FRT sites was selected by picking smallcolonies on YPD + 20–40 mg nourseothricine ml-1 (Reusset al., 2004). Loss of the SAT cassette was checked by thephenotypic recoveree of nourseothricine sensitivity and
SUN genes of C. albicans 1271
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

replacement of SUN ORFs by FRT sequences only wasconfirmed by PCR (data not shown).
Triple mutants were constructed by deleting the third allelewith the corresponding SAT-flipper cassette. The promoter ofthe last SUN genes in triple mutant backgrounds werereplaced by the methionine/cysteine-repressible promoterPMET3 (Care et al., 1999) by using a PCR-generated cassetteusing pFA-HIS-MET3p as template (Table S1) to give theC. albicans conditional sun mutants sun41DD sun42D PMET3-SUN42 and sun42DD sun41D PMET3-SUN41. Some indepen-dent sun41DD sun42D PMET3-SUN42 transformants differ bythe observed strength of growth inhibition under PMET3-repressing condition (data not shown), probably due to abasal activity of PMET3 under repressing condition (Care et al.,1999). Only a conditional mutant with the strongest growthdefect is presented in this study.
Quantification of sedimentation
To quantify the sedimentation phenotype, a protocol for deter-mination of the sedimentation half-time by spectrophotometricanalysis was adapted from Voth et al. (2005). Stationaryphase cultures (20 h in YPD at 30°C) were vortexed and 500 mlplaced in a microcuvette. Optical density (OD) at 660 nm wasrecorded every 150 s in a Beckman Coulter DU800 spectro-photometer for six independent samples per run. Data werenormalized (initial upper plateau OD = 100% and final lowerplateau OD = 0%) and the sedimentation half-time determinedas the time ODnormalized = 0.5 is reached. Reproducibility wasobtained when comparing the relative time of half-sedimentation of mutants with a control strain (BWP17AHU)done in the same run (relative half-sedimentation time = half-sedimentation time mutant ¥ 100/half-sedimentation time control).The average relative half-sedimentation time of mutants wascalculated from independent cultures and independent runs ofspectrophotometric measurements.
Microscopic observations
All microscopic observations were done on a Nikon EclipseE600 and images acquired with a Nikon Digital CameraDXM1200F. When needed, cell fixation was performed by a30 min room temperature incubation with 3.7% formaldehydefollowed by 30 min immobilization on polylysine microscopicslides and three washes in PBS buffer. Cell permeabilizationwas done by -20°C ethanol washing and three PBS washesbefore calcofluor white staining (10 mg ml-1 for 10 minfollowed by three PBS washes) or DAPI staining(VectaShield + DAPI).
For quantification of the cell separation defect, overnightcultures in liquid YPD at 30°C were diluted 100 times andmore than 400 cells were counted for each strain. Each groupof physically connected cells (chain of cells) were counted asa unit, and the proportions of chains of 1 + 2, 3 + 4, 5 + 6 andmore than 6 cells were determined.
To follow the kinetic of germination, synchronization ofC. albicans strains was achieved by pre-cultivating cells for48 h in rich medium (YPD at 30°C) and inoculation (d = 100¥)in prewarmed filamentation-inducing medium (buffered RPMIat 37°C). Every 15 min, 150 ml of cells was fixed, spotted on
a polylysine microscopic slide and stained with DAPI. Anaverage of 100 cells were counted for each time point. Thecompletion of the first cell cycle was defined by the presenceof two nuclei in cells.
For time-lapse movies, cells were placed on a micro-scope slide between a layer of solid SD medium(SD + methionine + cysteine) and a coverslip. Mediumexcess was cut and sealing was done with a 1:1:1 mixture ofvaseline, lanolin and paraffin wax. Images were capturedevery 10 min with a 60¥ immersion objective at roomtemperature. For movies, images were imported in WindowsMovie Maker (Microsoft) software and further processed withFinal Cut Pro (Apple). Each image resting time was set at0.125 s, resulting in a 4800¥ time acceleration.
Electronic microscopy
Overnight cultures in minimal medium at 30°C were diluted infresh minimal medium supplemented with methionine andcysteine and incubated for 16 h at 30°C. Cells were collectedby centrifugation and washed twice in PBS buffer beforefixation 1 h at room temperature in 2.5% glutaraldehyde in0.1 M cacodylate buffer (pH 7.2). For TEM, fixed sampleswere rinsed three times with 0.1 M cacodylate buffer, post-fixed with 2% osmium tetraoxide, 1% potassium ferricyanidein 0.1 M cacodylate buffer for 1 h at room temperature, andwashed again with 0.1 M cacodylate buffer and water. Thesamples were dehydrated through a graded series of ethanolbath and overnight in a mixture of Epon 812/ethanol 100%at room temperature. After being embedded in Epon 812,samples were polymerized for 48 h at 60°C. Ultrathin sec-tions (70–80 nm) were cut with a diamond knife, stained withuranyl acetate and Reynolds lead citrate, and viewed in aJEM 1010 TEM (Jeol, Tokyo, Japan) at 80 kV by using anEloïse Mega View III camera and AnalySIS Pro Softwareversion 3.1 (comEloïse SARL, Roissy, France). For SEM,fixed cells were washed three times for 5 min in 0.2 Mcacodylate buffer, postfixed for 1 h in 1% (wt/vol) osmiumtetroxide in 0.2 M cacodylate buffer, and then rinsed withdistilled water. Cells were dehydrated through a gradedseries of 25%, 50%, 75% and 95% ethanol solution. Sampleswere then dehydrated for 10 min in 100% ethanol followed bycritical point drying with CO2. Dried specimens were sput-tered with 10 nm gold palladium, with a GATAN Ion BeamCoater and were examined and photographed with a JEOLJSM 6700F field emission scanning electron microscopeoperating at 5 kV. Images were acquired with the upper SEdetector (SEI) and the lower secondary detector (LEI).
Drug sensitivities
Overnight cultures were diluted to OD600 0.4 and drug sensi-tivities were assayed by serial dilution (5¥) spotting (3 ml)on rich media (YPD). Chemical agents were purchasedfrom Sigma unless otherwise indicated and concentra-tions used in spotting assay are indicated: H2O2 (2–8 mM),tert-butyl hydroperoxide (1–4 mM), cumene (0.05–0.2 mM),menadione (0.05–0.2 mM), calcofluor white (15–40 mg ml-1),SDS (Q-BIOgene, 0.01–0.15%), polyoxin D (Calbiochem,10 mM), nikkomycin Z (2–10 mg ml-1), amphotericin B
1272 A. Firon et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

(0.25–1 mg ml-1), ketoconazole (1–10 mg ml-1), itraconazole(1–10 mg ml-1), cycloheximide (100–500 mg ml-1), FK506(2 mg ml-1), rapamycin (10–100 nM), cyclosporin A (20 mM),staurosporine (2 mM).
Quantitative RT-PCR
Overnight cultures at 30°C in YPD were diluted to OD600 0.2in YPD or buffered RPMI and grown for 4 h at 30°C (YPD)or 37°C (RPMI). Cells were collected from 50 ml cultures bycentrifugation and frozen at -80°C (YPD) or by filtering andfreezing in liquid nitrogen (RPMI). Total RNA were preparedwith the RNeasy mini kit (Qiagen) with an on-column DNasetreatment. A second DNase treatment (Turbo kit, Ambion)was done on purified RNA before RT-PCR (Superscript III,Invitrogen) with 1 mg of total RNA and a mix of gene-specificreverse primers. Quantitative PCR were done in triplicatefor each dilution with the SybrGreen detection system(Power SYBR Green master mix and Abi Prism 7000,Applied Biosystem) using primers designed to give specificamplicons of 100–150 bp (Table S2). Difference of lessthan 0.5 Ct between the triplicates are used for validation.Relative quantification by the DCt method were done withEFT3 as the control gene. For representation, relativequantification of specific genes were compared with afixed 1000 arbitrary units of EFT3 transcript (RNAgene X =(1/2(Ct gene X - Ct EFT3)) ¥ 1000).
Acknowledgements
We thank J. Wendland and J. Morschhäuser for pFA andSAT-flipper plasmids. We are grateful to members of A. Jac-quier (especially A. Thierry), B. Dujon (especially C.Saveanu) and J.P. Latgé (especially V. Kumar) laboratoriesfor S. cerevisiae strains and for stimulating discussions. A.Firon is the recipient of a postdoctoral fellowship from InstitutPasteur (Bourse Roux).
References
Alonso-Nunez, M.L., An, H., Martin-Cuadrado, A.B., Mehta,S., Petit, C., Sipiczki, M., et al. (2005) Ace2p controls theexpression of genes required for cell separation inSchizosaccharomyces pombe. Mol Biol Cell 16: 2003–2017.
Austriaco, N.R., Jr (1996) Review: to bud until death: thegenetics of ageing in the yeast, Saccharomyces. Yeast12: 623–630.
Bandara, P.D., Flattery-O’Brien, J.A., Grant, C.M., andDawes, I.W. (1998) Involvement of the Saccharomycescerevisiae UTH1 gene in the oxidative-stress response.Curr Genet 34: 259–268.
Bidlingmaier, S., Weiss, E.L., Seidel, C., Drubin, D.G., andSnyder, M. (2001) The Cbk1p pathway is important forpolarized cell growth and cell separation in Saccharomy-ces cerevisiae. Mol Cell Biol 21: 2449–2462.
Bulawa, C.E., Slater, M., Cabib, E., Au-Young, J., Sburlati, A.,Adair, W.L., Jr, and Robbins, P.W. (1986) TheS. cerevisiae structural gene for chitin synthase is notrequired for chitin synthesis in vivo. Cell 46: 213–225.
Cabib, E. (2004) The septation apparatus, a chitin-requiringmachine in budding yeast. Arch Biochem Biophys 426:201–207.
Cabib, E., Silverman, S.J., and Shaw, J.A. (1992) Chitinaseand chitin synthase 1: counterbalancing activities in cellseparation of Saccharomyces cerevisiae. J Gen Microbiol138: 97–102.
Cabib, E., Roh, D.H., Schmidt, M., Crotti, L.B., and Varma, A.(2001) The yeast cell wall and septum as paradigms of cellgrowth and morphogenesis. J Biol Chem 276: 19679–19682.
Camougrand, N., Grelaud-Coq, A., Marza, E., Priault, M.,Bessoule, J.J., and Manon, S. (2003) The product of theUTH1 gene, required for Bax-induced cell death in yeast, isinvolved in the response to rapamycin. Mol Microbiol47: 495–506.
Camougrand, N., Kissova, I., Velours, G., and Manon, S.(2004) Uth1p: a yeast mitochondrial protein at the cross-roads of stress, degradation and cell death. FEMS YeastRes 5: 133–140.
Cappellaro, C., Mrsa, V., and Tanner, W. (1998) New poten-tial cell wall glucanases of Saccharomyces cerevisiae andtheir involvement in mating. J Bacteriol 180: 5030–5037.
Care, R.S., Trevethick, J., Binley, K.M., and Sudbery, P.E.(1999) The MET3 promoter: a new tool for Candida albi-cans molecular genetics. Mol Microbiol 34: 792–798.
Colman-Lerner, A., Chin, T.E., and Brent, R. (2001) YeastCbk1 and Mob2 activate daughter-specific genetic pro-grams to induce asymmetric cell fates. Cell 107: 739–750.
Dahmann, C., Diffley, J.F., and Nasmyth, K.A. (1995)S-phase-promoting cyclin-dependent kinases preventre-replication by inhibiting the transition of replicationorigins to a pre-replicative state. Curr Biol 5: 1257–1269.
Dietrich, F.S., Voegeli, S., Brachat, S., Lerch, A., Gates, K.,Steiner, S., et al. (2004) The Ashbya gossypii genome as atool for mapping the ancient Saccharomyces cerevisiaegenome. Science 304: 304–307.
Du, L.L., and Novick, P. (2002) Pag1p, a novel protein asso-ciated with protein kinase Cbk1p, is required for cell mor-phogenesis and proliferation in Saccharomyces cerevisiae.Mol Biol Cell 13: 503–514.
Dujon, B. (2006) Yeasts illustrate the molecular mechanismsof eukaryotic genome evolution. Trends Genet 22: 375–387.
Finn, R.D., Mistry, J., Schuster-Bockler, B., Griffiths-Jones, S.,Hollich, V., Lassmann, T., et al. (2006) Pfam: clans, webtools and services. Nucleic Acids Res 34: D247–D251.
Garcia-Sanchez, S., Aubert, S., Iraqui, I., Janbon, G., Ghigo,J.M., and d’Enfert, C. (2004) Candida albicans biofilms: adevelopmental state associated with specific and stablegene expression patterns. Eukaryot Cell 3: 536–545.
Gola, S., Martin, R., Walther, A., Dunkler, A., and Wendland,J. (2003) New modules for PCR-based gene targeting inCandida albicans: rapid and efficient gene targeting using100 bp of flanking homology region. Yeast 20: 1339–1347.
de Groot, P.W., Yin, Q.Y., Weig, M., Sosinska, G.J., Klis,F.M., and de Koster, C.G. (2007) Mass spectrometricidentification of covalently bound cell wall proteins fromthe fission yeast Schizosaccharomyces pombe. Yeast 24:267–278.
Hiller, E., Heine, S., Brunner, H., and Rupp, S. (2007)Candida albicans SUN41 a putative glycosidase is
SUN genes of C. albicans 1273
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

involved in morphogenesis, cell wall biogenesis and bio-film formation. Eukaryot Cell 10.1128/EC.00285-00207(in press). doi:10.1128/EC.00285-07
Huh, W.K., Falvo, J.V., Gerke, L.C., Carroll, A.S., Howson,R.W., Weissman, J.S., and O’Shea, E.K. (2003) Globalanalysis of protein localization in budding yeast. Nature425: 686–691.
Kellis, M., Birren, B.W., and Lander, E.S. (2004) Proof andevolutionary analysis of ancient genome duplication in theyeast Saccharomyces cerevisiae. Nature 428: 617–624.
Kennedy, B.K., Austriaco, N.R., Jr, Zhang, J., and Guarente,L. (1995) Mutation in the silencing gene SIR4 can delayaging in S. cerevisiae. Cell 80: 485–496.
Kissova, I., Deffieu, M., Manon, S., and Camougrand, N.(2004) Uth1p is involved in the autophagic degradation ofmitochondria. J Biol Chem 279: 39068–39074.
Kurischko, C., Weiss, G., Ottey, M., and Luca, F.C. (2005) Arole for the Saccharomyces cerevisiae regulation of Ace2and polarized morphogenesis signaling network in cellintegrity. Genetics 171: 443–455.
Lesage, G., Shapiro, J., Specht, C.A., Sdicu, A.M., Menard,P., Hussein, S., et al. (2005) An interactional network ofgenes involved in chitin synthesis in Saccharomycescerevisiae. BMC Genet 6: 8.
Levin, D.E. (2005) Cell wall integrity signaling in Saccharo-myces cerevisiae. Microbiol Mol Biol Rev 69: 262–291.
Liu, H., Kohler, J., and Fink, G.R. (1994) Suppression ofhyphal formation in Candida albicans by mutation of aSTE12 homolog. Science 266: 1723–1726.
Liu, T.T., Lee, R.E., Barker, K.S., Wei, L., Homayouni, R., andRogers, P.D. (2005) Genome-wide expression profiling ofthe response to azole, polyene, echinocandin, and pyrimi-dine antifungal agents in Candida albicans. AntimicrobAgents Chemother 49: 2226–2236.
Lum, P.Y., Armour, C.D., Stepaniants, S.B., Cavet, G., Wolf,M.K., Butler, J.S., et al. (2004) Discovering modes of actionfor therapeutic compounds using a genome-wide screen ofyeast heterozygotes. Cell 116: 121–137.
Mnaimneh, S., Davierwala, A.P., Haynes, J., Moffat, J., Peng,W.T., Zhang, W., et al. (2004) Exploration of essential genefunctions via titratable promoter alleles. Cell 118: 31–44.
Monteoliva, L., Matas, M.L., Gil, C., Nombela, C., and Pla, J.(2002) Large-scale identification of putative exported pro-teins in Candida albicans by genetic selection. EukaryotCell 1: 514–525.
Mouassite, M., Camougrand, N., Schwob, E., Demaison, G.,Laclau, M., and Guerin, M. (2000) The ‘SUN’ family: yeastSUN4/SCW3 is involved in cell septation. Yeast 16: 905–919.
Mulhern, S.M., Logue, M.E., and Butler, G. (2006) Candidaalbicans transcription factor Ace2 regulates metabolismand is required for filamentation in hypoxic conditions.Eukaryot Cell 5: 2001–2013.
Munro, C.A., Whitton, R.K., Hughes, H.B., Rella, M., Selvag-gini, S., and Gow, N.A. (2003) CHS8-a fourth chitin syn-thase gene of Candida albicans contributes to in vitro chitinsynthase activity, but is dispensable for growth. FungalGenet Biol 40: 146–158.
Murad, A.M., Lee, P.R., Broadbent, I.D., Barelle, C.J., andBrown, A.J. (2000) CIp10, an efficient and convenient inte-grating vector for Candida albicans. Yeast 16: 325–327.
Nelson, B., Kurischko, C., Horecka, J., Mody, M., Nair, P.,Pratt, L., et al. (2003) RAM: a conserved signaling networkthat regulates Ace2p transcriptional activity and polarizedmorphogenesis. Mol Biol Cell 14: 3782–3803.
Newport, G., Kuo, A., Flattery, A., Gill, C., Blake, J.J., Kurtz,M.B., et al. (2003) Inactivation of Kex2p diminishes thevirulence of Candida albicans. J Biol Chem 278: 1713–1720.
Norice, C.T., Smith, F.J., Jr, Solis, N., Filler, S.G., and Mitch-ell, A.P. (2007) Requirement for C. albicans Sun41 inbiofilm formation and virulence. Eukaryot Cell doi: 10.1128/EC.00314-00307 (in press). doi:10.1128/EC.00314-07
Omi, K., Sonoda, H., Nagata, K., and Sugita, K. (1999)Cloning and characterization of psu1(+), a new essentialfission yeast gene involved in cell wall synthesis. BiochemBiophys Res Commun 262: 368–374.
Pelissier, P., Camougrand, N., Velours, G., and Guerin, M.(1995) NCA3, a nuclear gene involved in the mitochondrialexpression of subunits 6 and 8 of the Fo-F1 ATP synthaseof S. cerevisiae. Curr Genet 27: 409–416.
Reuss, O., Vik, A., Kolter, R., and Morschhauser, J. (2004)The SAT1 flipper, an optimized tool for gene disruption inCandida albicans. Gene 341: 119–127.
Roemer, T., Jiang, B., Davison, J., Ketela, T., Veillette, K.,Breton, A., et al. (2003) Large-scale essential gene identi-fication in Candida albicans and applications to antifungaldrug discovery. Mol Microbiol 50: 167–181.
Selvaggini, S., Munro, C.A., Paschoud, S., Sanglard, D., andGow, N.A. (2004) Independent regulation of chitin syn-thase and chitinase activity in Candida albicans and Sac-charomyces cerevisiae. Microbiology 150: 921–928.
Setiadi, E.R., Doedt, T., Cottier, F., Noffz, C., and Ernst, J.F.(2006) Transcriptional response of Candida albicans tohypoxia: linkage of oxygen sensing and Efg1p-regulatorynetworks. J Mol Biol 361: 399–411.
Skory, C.D., and Freer, S.N. (1995) Cloning and character-ization of a gene encoding a cell-bound, extracellularbeta-glucosidase in the yeast Candida wickerhamii.Appl Environ Microbiol 61: 518–525.
Sohn, K., Urban, C., Brunner, H., and Rupp, S. (2003) EFG1is a major regulator of cell wall dynamics in Candida albi-cans as revealed by DNA microarrays. Mol Microbiol 47:89–102.
Velours, G., Boucheron, C., Manon, S., and Camougrand, N.(2002) Dual cell wall/mitochondria localization of the ‘SUN’family proteins. FEMS Microbiol Lett 207: 165–172.
Verstrepen, K.J., and Klis, F.M. (2006) Flocculation, adhesionand biofilm formation in yeasts. Mol Microbiol 60: 5–15.
Voth, W.P., Olsen, A.E., Sbia, M., Freedman, K.H., andStillman, D.J. (2005) ACE2, CBK1, and BUD4 in buddingand cell separation. Eukaryot Cell 4: 1018–1028.
Walther, A., and Wendland, J. (2003) An improved transfor-mation protocol for the human fungal pathogen Candidaalbicans. Curr Genet 42: 339–343.
Weiss, E.L., Kurischko, C., Zhang, C., Shokat, K., Drubin,D.G., and Luca, F.C. (2002) The Saccharomyces cerevi-siae Mob2p-Cbk1p kinase complex promotes polarizedgrowth and acts with the mitotic exit network to facilitatedaughter cell-specific localization of Ace2p transcriptionfactor. J Cell Biol 158: 885–900.
Wilson, R.B., Davis, D., and Mitchell, A.P. (1999) Rapid
1274 A. Firon et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275

hypothesis testing with Candida albicans through genedisruption with short homology regions. J Bacteriol 181:1868–1874.
Wisplinghoff, H., Bischoff, T., Tallent, S.M., Seifert, H.,Wenzel, R.P., and Edmond, M.B. (2004) Nosocomialbloodstream infections in US hospitals: analysis of 24,179cases from a prospective nationwide surveillance study.Clin Infect Dis 39: 309–317.
Yeong, F.M. (2005) Severing all ties between mother anddaughter: cell separation in budding yeast. Mol Microbiol55: 1325–1331.
Supplementary material
This material is available as part of the online article from:http://www.blackwell-synergy.com/doi/abs/10.1111/j.1365-2958.2007.06011.x(This link will take you to the article abstract).
Please note: Blackwell Publishing is not responsible for thecontent or functionality of any supplementary materials sup-plied by the authors. Any queries (other than missing mate-rial) should be directed to the corresponding author for thearticle.
SUN genes of C. albicans 1275
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1256–1275
Copyright © 2022 FDOKUMEN




















