
Necessary Condition Analysis (NCA): Logic and Methodology ...

Microbiological Research 166 (2011) 430—436
Available online at www.sciencedirect.com
www.elsevier.de/micres
SHORT COMMUNICATION
The Candida albicans Hwp2 is necessary forproper adhesion, biofilm formation and oxidativestress tolerance
Samer Younesa, Wael Bahnana, Hani I. Dimassib, Roy A. Khalafa,∗
aNatural Sciences Division, Lebanese American University, P.O Box 36, Byblos, LebanonbSchool of Pharmacy, Lebanese American University, P.O Box 36, Byblos, Lebanon
Received 18 November 2009 ; received in revised form 10 August 2010; accepted 14 August 2010
KEYWORDSCandida albicans;Biofilm;Filamentation;Adhesion
SummaryCandida albicans is an important fungal pathogen of humans that is responsible forthe majority of mucosal and systemic candidiasis. The host—pathogen interactionin C. albicans has been the subject of intense investigation as it is the primary stepthat leads to establishment of infection. Hwp2 is a cell wall GPI-anchored cell wallprotein that was previously shown to be necessary for hyphal and invasive growth onsolid media. The purpose of the current study is to further characterize the protein asfar as its role in oxidative stress, sensitivity to cell wall disrupting agents, adhesionto human epithelial and endothelial cells, biofilm formation and chitin content.It appears that Hwp2 is necessary for proper oxidative stress tolerance, adhesion
and biofilm formation as an hwp2 null is more susceptible to increasing doses ofhydrogen peroxide, unable to adhere efficiently to epithelial and endothelial celllines and unable to form wild type biofilm levels.© 2010 Elsevier GmbH. All rights reserved.Introduction
The human opportunistic fungus, Candida albicans
and potentially fatal disseminated disease in itshuman host. C. albicans is the most virulent speciesof the genus Candida (Karkowska-Kuleta et al.
is a diploid microorganism that is capable of achiev-ing filamentous growth, attachment and invasion ofhuman epithelia and is a causative agent of severe
∗Corresponding author. Tel.: +961 9547262x2216;fax: +961 9944851.
E-mail address: [email protected] (R.A. Khalaf).
2h(wosdp
0944-5013/$ – see front matter © 2010 Elsevier GmbH. All rights resedoi:10.1016/j.micres.2010.08.004
009). It usually resides in a benign manner in theost mouth, gut, and vagina without causing diseaseSouthern et al. 2008). At certain disease stateshen a patient is immunocompromised, C. albicans
vercomes host defenses and can causes inva-ive and lethal disease. Physiologic states such asiabetes, pregnancy, neutropenia, and immunosup-ressive therapy predispose individuals to seriousrved.

H on
Cfb
ctrysnaimeecaflfhalsatnwmasrocateTsgoMmIvansliscdra2
anesasFwttfs
M
S
T(hdhhs
M
Tdrwstb(wbtoa3
O
C
HHs
wp2 is necessary for adhesion and biofilm formati
andidiasis. C. albicans is the most common humanungal pathogen, and is occasionally retrieved fromlood cultures (Viudes et al. 2002).
Many factors allow C. albicans to be a suc-essful pathogen. For example C. albicans is ableo interchange between two main morphologies:ound yeast blastospore cells and hyphal cells. Theeast form is necessary for rapid clonal expan-ion while the hyphal form allows tissue invasioneeded for dissemination (Karkowska-Kuleta etl. 2009). Furthermore, C. albicans has the abil-ty to switch between white and opaque colonyorphologies with the opaque form composed of
longated oval cells necessary for mating and forvading the immune system whereas the whiteells are virulent and antigenic (Yang 2003). C.lbicans has evolved a wide set of genes codingor secreted virulence factors such as phospho-ipase, protease, dismutases and others. Theseactors allow invasion, self-defense against theost and other pathogenic processes (Naglik etl. 2003; Brown et al. 2007). Many of these viru-ence factors are located on the cell surface. Asuch the cell wall shows many pathogenic char-cteristics including promoting a good adaptationo the different steps of infection from colo-ization to organ penetration. Furthermore cellall components are responsible for the first andost important process that establishes infection;
dhesion, in addition to morphological switching,tress tolerance, antigenicity and other virulenceelated processes (Masuoka 2004). The structuref the cell wall and its modulable nature is cru-ial for infection. Hyphal cells for example exhibitthicker cell wall with increased chitin deposi-
ion and with different cell wall proteins beingxpressed than do yeast cells (Kapteyn et al. 2000).he C. albicans cell wall is composed of an innertress tolerant chitin layer bound to mainly �-1,3-lucan and some �-1,6-glucan molecules and anuter layer of mannoproteins (Tronchin et al. 1981).ost adhesins characterized such Hwp2, Hwp1, andembers of the Als family attach to the cell wall.
n many instances attachment to the surface occursia a preformed GPI (glycosylphosphatidylinositol)nchor added postranslationally to the C termi-us. Most adhesins studied to date have commontructural features, with an N terminal globular Igike domain, serine threonine rich region contain-ng conserved repeats and a C terminal glycosylatedtalk region that extends the active site from theell wall surface (Otoo et al. 2008). As antigenic
eterminants, deletion of many of these proteinsendered the mutant cell significantly defective indhesion and virulence (Yang 2003; Hayek et al.010).h(Hp
431
Here we present our attempt to further char-cterize Hwp2 which we found previously to beecessary for invasive growth on solid media (Hayekt al. 2010) by assessing whether the lack of inva-ive growth in the mutant is mimicked as a lack ofdhesion to human cells and whether a lack of adhe-ion is mirrored as a decrease in biofilm formation.urthermore we were interested in determininghether the deletion of Hwp2 — a cell wall pro-
ein — affects chitin deposition and in turn affecthe susceptibility of the mutant to various cell sur-ace disrupting agents, and tolerance to oxidativetress.
aterials and methods
trains utilized
he C. albicans wild type strain RM1000 + pABSK2ura3�::�imm434/ura3�::�imm434his1::hisG/is1::hisG) (Negredo et al. 1997) histidine and uri-ine auxotroph, HWP2/hpw2::URA3 heterozygote,wp2:URA3/hwp2::HIS1 null, and ura3::HWP2/wp2::HIS1 revertant strains were used in thistudy (Hayek et al. 2010).
edia preparation and culture conditions
he C. albicans strains were grown on rich potatoextrose agar (PDA) medium (Hi Media, India) foroutine cultures. PDA plates were supplementedith uridine and histidine in all experiments of
potting, adhesion, biofilm and oxidative stressolerance. For the biofilm assay, yeast nitrogenase (YNB) synthetic medium (Fluka, Switzerland)Kaiser et al. 1994) was used and was supplementedith uridine and histidine. Rich potato dextroseroth (PDB) liquid media (Hi Media, India) was usedo grow the strains until exponential phase forxidative stress, cell surface disrupting media anddhesion experiments. Strains were incubated at0 ◦C under aerobic conditions.
Colony images were captured by the use of anlympus E330-ADU-1.2× stereomicroscope.
ell culture
uman colon adenocarcinoma epithelial cell lineT-29 ATCC number HTB-38 was utilized in thistudy for epithelial adhesion experiments. The
uman cells were cultured in RPMI 1640 mediumBiowhittaker, Belgium) which contains 25 mMEPES and l-glutamine. The medium was sup-lemented with 10% FBS (fetal bovine serum,
aiwu3wcawAfdbmw
C
Ctewctwtlf0f(cmtsput
S
Dpsvcp
R
432
Biowhittaker), 100 U/ml of penicillin, 100 �g/ml ofstreptomycin, and the culture flasks were incu-bated at 37 ◦C in 5% CO2. Human endothelial cellline ECV304 originating from human bladder carci-noma with a perfect endothelial phenotype (ATCCnumber CRL 1998) was utilized in this study forendothelial adhesion experiments. Culture condi-tions utilized were identical to that of the epithelialadenocarcinoma cell line.
Oxidative stress tolerance
The wild type, heterozygote, revertant and nullmutant were assayed for their ability to withstanda potentially lethal dose of hydrogen peroxide.Strains were incubated with 10, 25 and 50 mMconcentration of hydrogen peroxide for 1 h duringtheir exponential growth phase (between OD600 0.2and 0.8). After incubation, 5 �l of a serial dilutionwas spotted on PDA plates (Pedreno et al. 2007).
Cell surface disrupting agents
Strains were grown until exponential phase andserially diluted. 5 �l of the serial dilutions werespotted on PDA plates treated with 0.2—0.03%SDS (Sigma—Aldrich, Germany), 50—300 �g/ml cal-cofluor white (Sigma—Aldrich, Germany), and100—300 �g/ml Congo red (Sigma—Aldrich, Ger-many). A control plate lacking cell surfacedisrupting agents was spotted with the exact samevolume of the three strains. The plates were incu-bated at 30 ◦C for 3—4 days (Plaine et al. 2008).
Adhesion assay
Adhesion ability of the strains was assayedagainst human epithelial cell line HT-29 and humanendothelial cell line ECV304 as previously described(Tsuchimori et al. 2000). Briefly, around 100 cellsof the three strains were incubated with humancell lines in 6-well microtiter plates for either 90 or180 min followed by washing with PBS and overlaywith molten PDA agar. Colonies were countedafter overnight growth and were compared with acontrol plated inoculated with the same number ofC. albicans cells. The results of this experiment areexpressed as percentage adhesion of the controlplate.
Biofilm formation
The ability of the strains to form biofilms onpolystyrene microtiter wells was assayed accordingto a previously described protocol (Peeters et
O
Ta
S. Younes et al.
l. 2008). Briefly, 5 × 106 cells were incubatedn a flat bottomed microtiter well pretreatedith 5% serum overnight. Strains along with anninoculated control well, were left to incubate at7 ◦C in a shaking incubator at 75 rpm for 2 h. Wellsere rinsed with PBS to remove non-adherentells followed by addition of YNB and incubationt 37 ◦C for 48 h. After incubation plates wereashed again, fixed with methanol and air dried.0.2% crystal violet solution was added and left
or 20 min. Excess crystal violet was washed withistilled water and the crystal violet bound toiofilm cells was released by acetic acid treat-ent. Absorbance of the released crystal violetas measured spectrophotometrically at 590 nm.
hitin quantification
ell wall chitin content was measured accordingo a modified protocol described previously (Munrot al. 2003; Kapteyn et al. 2000). Briefly, 6N HClas used to hydrolyze 50 mg wet weight purifiedell walls at 100 ◦C overnight. After centrifugationhe pellet was reconstituted in 1 ml of distilledater. A 0.1 ml aliquot of this sample was added
o 0.1 ml of solution A (1.5N Na2CO3 in 4% acety-acetone). The mixture was incubated at 100 ◦Cor 20 min and after cooling to room temperature,.7 ml of 96% ethanol was added to the mixtureollowed by addition of a 0.1 ml of solution B1.6 g of p-dimethyl-aminobenzaldehyde in 30 ml ofoncentrated HCl and 30 ml of 96% ethanol). Theixture was incubated for 1 h at room tempera-
ure and absorbance of the samples was measuredpectrophotometrically at 520 nm. The results werelotted against a standard curve generated by these of known glucosamine standards taken throughhe same procedure as our samples.
tatistical analysis
ata was coded and entered into the SPSS 17rogram (SPSS Inc., USA) for analysis. Mean andtandard deviations were calculated. Analysis ofariance (ANOVA) was carried out with multipleomparison technique to control for alpha inflation.-Value less than 5% was considered significant.
Experiments were performed in triplicates.
esults
xidative stress response
he C. albicans strains were grown until station-ry phase and treated with various concentrations

Hwp2 is necessary for adhesion and biofilm formation 433
Figure 1. Oxidative stress response: The wild type, heterozygote, revertant and hwp2 null mutant were incubatedw e form pero
oic
C
TpiswNhtC
A
Ticahtatd1e
B
TiTmi
C
Tabad
D
Tcfawha
rnwosiaHiPtaae
ith 10, 25 and 50 mM concentration of hydrogen peroxidutant clearly shows increase susceptibility to hydrogen
f hydrogen peroxide. Our mutant strain showedncreased sensitivity to hydrogen peroxide whenompared to the wild type strain (Fig. 1).
ell surface disrupting agents
he C. albicans strains were grown until stationaryhase and then spotted on PDA plates contain-ng various cell wall disrupting agents. The mutanttrain showed a very slight sensitivity to SDS,hen compared to the wild type strain (Fig. 2D).o discrepancy was seen between the mutant,eterozygote, and wild type strains on plates con-aining calcofluor white or Congo red (Fig. 2B and).
dhesion assay
he C. albicans strains were assayed for the abil-ty to adhere to human epithelial and endothelialell lines. After 90 min the mutant strain showedsignificant (75%) decrease in ability to adhere to
uman epithelial cells; in comparison to the wildype strain (p < 0.001). The null mutant also showed
significant (70%) decrease in ability to adhereo human endothelial cells (p < 0.001, Fig. 3). Thisiscrepancy was not overcome after prolonged80-min incubation as no additional adhesion wasvident.
iofilm formation
he C. albicans strains were assayed for their abil-
ty to form biofilms on polystyrene plastic surfaces.he mutant strain showed a 30% decrease in biofilmass (p = 0.008) with the revertant exhibiting anntermediate phenotype (Fig. 4).
nI(o
1 h. (A) 50 mM concentration of hydrogen peroxide. Thexide when compared to the wild type. (B) Control plate.
hitin content
he amount of cell wall chitin was determined bycid hydrolysis for all the C. albicans strains. As cane seen in Fig. 5 a slight yet significant discrep-ncy was observed between the wild type and theeletion strains.
iscussion
his work has further characterized the C. albi-ans cell wall protein Hwp2. We have previouslyound it to be implicated in C. albicans virulencend filamentation (Hayek et al. 2010). In this work,e show that Hwp2 is essential for adhesion touman epithelia and endothelia, biofilm formationnd hydrogen peroxide stress response.
Our hwp2 null mutant did not exhibit a defectegarding cell wall integrity and brittleness sinceo increase in susceptibility upon exposure to cellall disrupting agents such as SDS, Congo redr calcofluor white was found. Furthermore noignificant decrease in chitin content was foundndicating that the rigidity of the cell wall was notffected. This should not come as a surprise sincewp2 as a �-1,6-glucan linked adhesin is not an
ntegral cell wall component as is the case withir1a �-1,3-glucan linked integral proteins. Dele-ion of Pir1 dramatically affects cell wall integritynd decreases resistance to cell surface disruptinggents (Martínez et al. 2004). Interestingly how-ver, resistance to hydrogen peroxide stress did
ot follow that pattern and decreased dramatically.n C. albicans, a family of superoxide dismutasesSods) is responsible for counteracting the effectsf oxidative stress. Sod5 in particular was shown to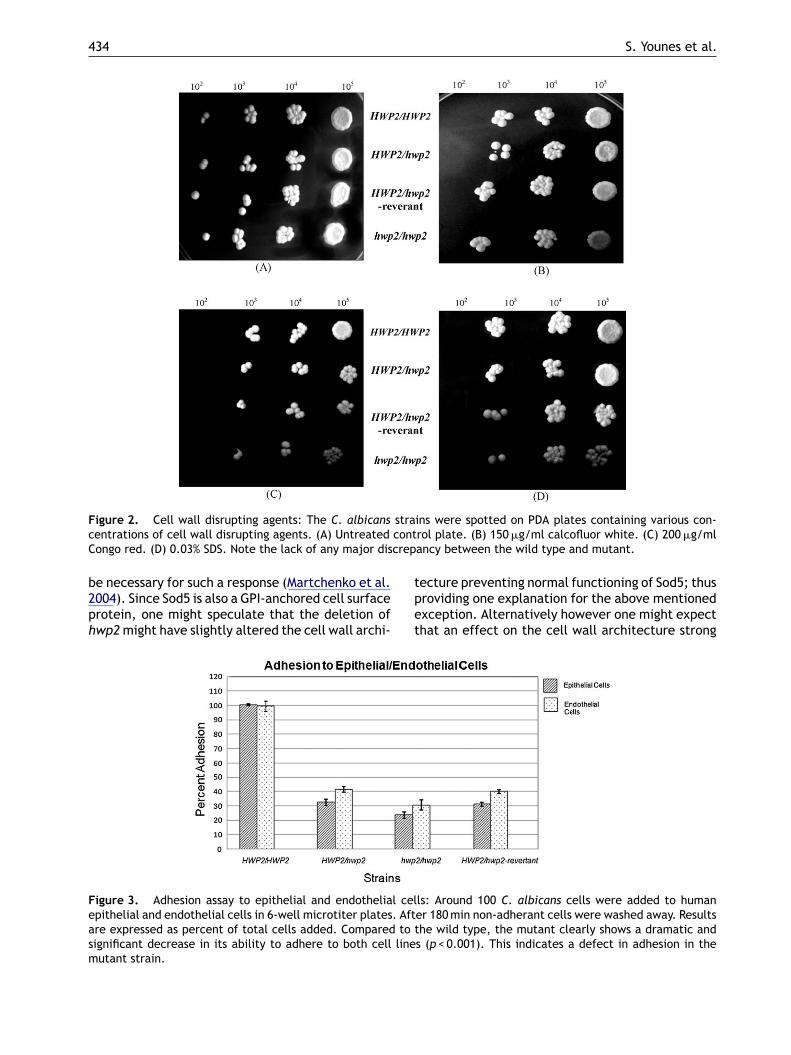
434 S. Younes et al.
Figure 2. Cell wall disrupting agents: The C. albicans strains were spotted on PDA plates containing various con-contcrep
centrations of cell wall disrupting agents. (A) UntreatedCongo red. (D) 0.03% SDS. Note the lack of any major dis
be necessary for such a response (Martchenko et al.2004). Since Sod5 is also a GPI-anchored cell surfaceprotein, one might speculate that the deletion ofhwp2 might have slightly altered the cell wall archi-
tpet
Figure 3. Adhesion assay to epithelial and endothelial celepithelial and endothelial cells in 6-well microtiter plates. Aftare expressed as percent of total cells added. Compared to tsignificant decrease in its ability to adhere to both cell linemutant strain.
rol plate. (B) 150 �g/ml calcofluor white. (C) 200 �g/mlancy between the wild type and mutant.
ecture preventing normal functioning of Sod5; thusroviding one explanation for the above mentionedxception. Alternatively however one might expecthat an effect on the cell wall architecture strong
ls: Around 100 C. albicans cells were added to humaner 180 min non-adherant cells were washed away. Resultshe wild type, the mutant clearly shows a dramatic and
s (p < 0.001). This indicates a defect in adhesion in the

Hwp2 is necessary for adhesion and biofilm formation
Figure 4. Biofilm formation: Biofilm formation onpolystyrene plastic microtiter plates was performed asdescribed in the materials and methods. Note the veryslight decrease in crystal violet absorbance at 590 nmcorresponding to a decrease in biofilm mass in themutant strain as opposed to the wild type (p < 0.008). Therp
eeimrn
oitctcfnta
Fwhcc
fmabclamafa
A
ISch
R
B
H
H
K
evertant HWP2/hwp2 strain exhibited an intermediatehenotype.
nough to altered Sod5 function would have anffect on numerous other cell wall proteins involvedn cell wall biosynthesis consequently the hwp2 nullutant would have been affected by cell wall dis-
upting agents. More such experimental evidence isecessary.
On the other hand the most striking defectbserved in our mutant is the dramatic decreasen adhesion and biofilm formation. In that respecthe hwp2 null phenotype is similar to that of manyharacterized GPI-anchored adhesins such as Eap1hat has been shown to be necessary for properell-cell adhesion and adhesion to polystyrene sur-aces (Li and Palecek 2003; Li et al. 2007). Another
otable adhesin family is the Als family. Deple-ion of Als2 has been shown to dramatically affectdhesion and result in abnormal levels of biofilmigure 5. Chitin content: The amount of cell wall chitinas determined spectrophotometrically following acidydrolysis. Data represents percent chitin content asompared to the wild type. Note the slight decrease inhitin content in the mutant strain.
K
K
L
L
M
M
435
ormation (Hoyer 2001). Interestingly, the aboveentioned genes are exclusively cell wall proteins
nd contain a serine threonine rich repeat that haseen shown both bionformatically and biochemi-ally to be necessary for aggregation into amyloidike fibers, a precursor step to adhesion (Otoo etl. 2008; P. Lipke, personal communication). Nullutants of these proteins also show defects in
dhesion, invasive growth and filamentation addingurther evidence of the role such repeats play inggregation and adhesion (Yang 2003).
cknowledgements
would like to express my gratitude to Dr. M. El-ibai, S. El Sitt and B. Khalil for their help in cellulturing. I would also like to thank S. Youhanna forer help in statistical analysis of data.
eferences
rown A, Odds F, Gow N. Infection-related geneexpression in Candida albicans. Curr Opin Microbiol2007;10:307—13.
ayek P, Yazbek P, Beyrouthy B, Khalaf RA. Char-acterization of Hwp2, a Candida albicans putativeGPI-anchored cell wall protein necessary for invasivegrowth. Microbiol Res 2010;165:250—8.
oyer LL. The ALS gene family of Candida albicans.Trends Microbiol 2001;9:176—80.
aiser C, Michaelis S, Mitchell A. Methods in yeast genet-ics. New York: Cold Spring Harbor Laboratory Press;1994.
arkowska-Kuleta J, Rapala-Kosik M, Kozik A. Fungipathogenic to humans: molecular basis of viru-lence in Candida albicans, Cryptococcus neofor-mans, and Aspergillus fumigatus. Acta Biochim Pol2009;56:211—24.
apteyn JC, Hoyer LL, Hecht JE, Müller WH, Andel A,Verkleij AJ, et al. The cell wall architecture of Can-dida albicans wild-type cells and cell wall-defectivemutants. Mol Microbiol 2000;35:601—11.
i F, Palecek SP. EAP1, a Candida albicans gene involvedin binding human epithelial cells. Eukaryot Cell2003;2:1266—73.
i F, Svarovsky MJ, Karlsson AJ, Wagner JP, Marchillo K,Oshel P, et al. Eap1p, an adhesin that mediates Can-dida albicans biofilm formation in vitro and in vivo.Eukaryot Cell 2007;6:931—9.
artchenko M, Alarco AM, Harkus D, Whiteway M.Superoxide dismutases in Candida albicans: tran-scriptional regulation and functional characterization
of the hyphal-induced SOD5 gene. Mol Biol Cell2004;15:456—67.artínez AI, Castillo L, Garcerá A, Elorza MV, ValentínE, Sentandreu R. Role of Pir1 in the construc-

P
P
S
T
T
V
436
tion of the Candida albicans cell wall. Microbiology2004;150:3151—61.
Masuoka J. Surface glycans of Candida albicans andother pathogenic fungi: physiological roles, clinicaluses, and experimental challenges. Clin Microbiol Rev2004;17:281—310.
Munro CA, Whitton RK, Hughes HB, Rella M, Selvaggini S,Gow NA. CHS8-a fourth chitin synthase gene of Can-dida albicans contributes to in vitro chitin synthaseactivity, but is dispensable for growth. Fungal GenetBiol 2003;40:146—58.
Naglik J, Challacombe S, Hube B. Candida albicanssecreted aspartyl proteinases in virulence and patho-genesis. Microbiol Mol Biol Rev 2003;67:400—28.
Negredo A, Monteoliva L, Gil C, Pla J, Nombela C. Cloning,analysis and one-step disruption of the ARG5,6gene of Candida albicans. Microbiology 1997;143:297—302.
Otoo HN, Lee KG, Qiu W, Lipke PN. Candida albicans Alsadhesins have conserved amyloid-forming sequences.Eukaryot Cell 2008;7:776—82.
Pedreno Y, González-Párraga P, Martínez-Esparza M, Sen-tandreu R, Valentín E, Argüelles JC. Disruption of the
Candida albicans ATC1 gene encoding a cell-linkedacid trehalase decreases hypha formation and infec-tivity without affecting resistance to oxidative stress.Microbiology 2007;153:1372—81.Y
S. Younes et al.
eeters E, Nelis HJ, Coenye T. Comparison of multi-ple methods for quantification of microbial biofilmsgrown in microtiter plates. J Microbiol Methods2008;72:157—65.
laine A, Walker L, Da Costa G, Mora-Montes HM, McKin-non A, Gow NA, et al. Functional analysis of Candidaalbicans GPI-anchored proteins: roles in cell wallintegrity and caspofungin sensitivity. Fungal GenetBiol 2008;45:1404—14.
outhern P, Horbul J, Maher D, Davis D. C. albicanscolonization of human mucosal surfaces. PLoS One2008;3:e2067.
ronchin G, Poulain D, Herbaut J, Biguet J. Localization ofchitin in the cell wall of Candida albicans by means ofwheat germ agglutinin. Fluorescence and ultrastruc-tural studies. Eur J Cell Biol 1981;26:121—8.
suchimori N, Sharkey LL, Fonzi WA, French SW, EdwardsJr JE, Filler SG. Reduced virulence of HWP1-deficientmutants of Candida albicans and their interactionswith host cells. Infect Immun 2000;68:1997—2002.
iudes A, Peman J, Canton E, López-Ribot J, GobernadoM. Candidemia at a tertiary-care hospital: epidemiol-ogy, treatment, clinical outcome and risk factors for
death. Eur J Clin Microbiol Infect Dis 2002;21:767—74.ang Y. Virulence factors of Candida species. J MicrobiolImmunol Infect 2003;36:223—8.
Copyright © 2022 FDOKUMEN




















