
Morphometric and histological analysis of the lungs of - NCBI

Glycobiology vol. 18 no. 11 pp. 871–881, 2008doi:10.1093/glycob/cwn074Advance Access publication on August 6, 2008
The structure and proinflammatory activity of the lipopolysaccharidefrom Burkholderia multivorans and the differences between clonal strainscolonizing pre and posttransplanted lungs
Teresa Ierano2, Alba Silipo2, Luisa Sturiale3, DomenicoGarozzo3, Heather Brookes4, C M Anjam Khan5, ClareBryant4, F Kate Gould6,7, Paul A Corris7,8, RosaLanzetta2, Michelangelo Parrilli2, Anthony De Soyza8, andAntonio Molinaro1,2
2Dipartimento di Chimica Organica e Biochimica, Universita di NapoliFederico II, Napoli, Via Cintia 4, I-80126, Italy; 3Istituto per la Chimica e laTecnologia dei Materiali Polimerici – ICTMP – CNR, Catania, Italy; 4Centrefor Veterinary Science, Department of Veterinary Medicine, University ofCambridge, Cambridge, UK; 5Institute for Cell and Molecular Biosciences,The Medical School; 6Department of Medical Microbiology;7Cardiopulmonary Transplantation, Freeman Hospital; and 8AppliedImmunobiology and Transplantation Group, Institute of Cellular Medicine,University of Newcastle, Newcastle, UK
Received on May 30, 2008; revised on July 28, 2008; accepted on July 29, 2008
The Burkholderia cepacia complex is a group of Gram-negative bacteria that are opportunistic pathogens forhumans especially in cystic fibrosis patients. Lipopolysac-charide (LPS) molecules are potent virulence factors ofGram-negative bacteria organisms essential for bacterialsurvival. A complete analysis of the bacterial lipopolysac-charide structure to function relationship is requiredto understand the chemical basis of the inflammatoryprocess. We have therefore investigated the structures oflipopolysaccharides from clonally identical Burkholderiamultivorans strains (genomovar II) isolated pre- andpost-lung transplantation through compositional analysis,mass spectrometry, and 2D NMR spectroscopy. We testedthe LPS proinflammatory activity as a stimulant of humanmyelomonocytic U937 cell cytokine induction and assessedTLR4/MD2 signaling. Marked changes between the pairedstrains were found in the lipid A-inner core region. Suchstructural variations can contribute to the bacterial survivaland persistence of infections despite the loss of a CF milieufollowing lung transplantation.
Keywords: Burkholderia multivorans/cystic fibrosis/lipooligosaccharide/lung transplantation/NMR spectroscopy
Introduction
We report the complete structure of the Lipopolysaccharide(LPS) endotoxin from the Gram-negative Burkholderia multivo-rans clinical isolates for the first time. We have also investigated
1To whom correspondence should be addressed: Tel: +39-081-674123; Fax:+39-081-674393; e-mail: [email protected]
the structural modifications that B. multivorans LPS undergoesafter lung transplantation and the proinflammatory activity ofsuch clinical isolates. B. multivorans is one of the most com-mon and dangerous opportunistic pathogens of the Burkholderiacepacia complex species isolated so far in CF centers.
“Burkholderia cepacia” has long been identified as a plantpathogen where it causes onion rot (Speert 2002). More recentlyit has been recognized as an opportunistic human pathogen forindividuals with cystic fibrosis (CF) and chronic granuloma-tous disease. Previously, infections with “Burkholderia cepa-cia” were attributed to a single species. Indeed, such infectionsare caused by a variety of bacterial species, now grouped intothe “Burkholderia cepacia complex (BCC)” (Mahenthiralingamet al. 2000). Currently this complex comprises 10 related speciesnamed genomovars which are phenotypically similar but geno-typically different. The most prevalent clinical isolates fromCF patients are B. cenocepacia strains (genomovar III) and B.multivorans strains (genomovar II) (De Soyza, Morris, et al.2004; Wellinghausen and Kothe 2006). B. cenocepacia appearsthe most pathogenic with accelerated lung disease reported inpatients with previously well-preserved lung function. Approx-imately 20% of patients develop “cepacia syndrome” a ful-minating necrotizing pneumonitis with associated septicaemia(De Soyza and Corris 2003). B. cenocepacia is the species mostcommonly associated with the “cepacia syndrome” althoughsome cases are due to B. multivorans (Blackburn et al. 2004).BCC genomovars also possess a potentially transmissible na-ture: B. cenocepacia is the most notorious genomovar causingepidemics although B. multivorans epidemics have also beenreported.
Lung transplantation, whilst intensive and aggressive, is theonly treatment that improves both the quantity and quality oflife of CF patients with advanced lung disease. It has beenshown that BCC infections are the second most prevalent pre-transplantation infection. Transplanting such patients remainscontroversial as poor outcomes are common in those patientswith preoperative B. cenocepacia infection. In our experiencerecipients infected with B. cenocepacia died early posttrans-plant in a fashion similar to “cepacia syndrome” (De Soyzaet al. 2001; Jones et al. 2004). In contrast, excellent sur-vival rates in patients with pretransplantation B. multivorans(genomovar II) and B. vietnamiensis (genomovar V) infectionwere noted. Lung transplantation outcomes appear to be depen-dent on a number of factors including preoperative B. cepaciacomplex genomovar status (De Soyza et al. 2001).
The pathogenic mechanisms involved in BCC infections arestill unclear. An investigation of the structure and biologi-cal activity of lipopolysaccharides (LPSs) is required as theyare critically important bacterial virulence factors (Holst 1999;
c© The Author 2008. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: [email protected] 871
by guest on May 29, 2013
http://glycob.oxfordjournals.org/D
ownloaded from

T Ierano et al.
Alexander and Rietschel 2001; Raetz and Whitfield 2002). LPSsare amphiphilic macromolecules indispensable for the growthand the survival of Gram-negative bacteria. LPS has been iden-tified as a pathogen associated molecular pattern (PAMP) andis recognized by host pathogen-recognition receptors to inducea marked inflammatory response via TLR4/MD2-mediated sig-naling pathways (Akira et al. 2006).
We have previously demonstrated that lipid A changes in B.cenocepacia (De Soyza, Ellis, et al. 2004) similar to those re-ported in CF-associated P. aeruginosa strains (Ernst et al. 1999),suggesting an important interaction between the CF microenvi-ronment and infecting pathogen. Deacylation of P. aeruginosaLPS in bacterial strains from CF patients results in reduced ac-tivation of TLR-4/MD2 (Hajjar et al. 2002). Despite the preva-lence of B. multivorans as a CF-related pathogen, limited dataexist regarding LPS in B. multivorans.
In this work we define the complete primary structure andproinflammatory activity of the LPS extracts from B. multivo-rans. Lung transplantation offers interesting insights into assess-ing possible host–pathogen interactions as following transplan-tation any bacterial interaction will be with a lung allograft thatdoes not bear cystic fibrosis epithelium or CFTR defects. We in-vestigated the structure of the LPS from clinical isolates prior toand shortly after lung transplantation. Such data are essential toelucidate the molecular modifications involved in the inflamma-tory process that occur pre- and postsurgery. Understanding howbacteria modify and adapt their LPS under new physiologicalconditions may allow greater understanding of LPS-associatedsignaling. These data may also allow new therapeutic targets tobe identified including the design of antimicrobial compoundsagainst chemically different targets.
Results
Strain selection and SDS electrophoresis analysis of B.multivorans LOSsWe identified three patients who had paired pre- and posttrans-plantation strains of BCC that demonstrated differences in roughLPS (lipo-oligosaccharide; LOS) migration patterns upon silverstaining of 16% SDS gels (LOSs from B. multivorans strains areshown in Figure 1A). This change was not seen in the remainderof the transplant cohort analyzed. Of these three patients, onepatient had B. multivorans infection, one had B. vietnamensisinfection, and one had B. cenocepacia infection. The datareported will be limited to the patient with B. multivorans in-fection. The pretransplantation strain was isolated immediatelypretransplant and the posttransplantation strain was isolatedat 1 week after transplantation (Figure 1B). Sodium dodecylsulfate–polyacrylamide gel electrophoresis (SDS–PAGE) re-vealed that the extracted B. multivorans lipopolysaccharide wasa rough-type LPS or lipooligosaccharide (LOS) (Figure 1A).
Isolation and compositional analysis of LOS, Fr1, and Fr2from B. multivorans pre- and posttransplantationLPS fractions from both clinical isolates were extracted andanalyzed by SDS–PAGE. Both fractions consisted of rough LPSchemotype (lipo-oligosaccharide, LOS) as suggested by the runat the bottom of the gel (Figure 1A).
Fig. 1. Evaluation LPS from B. multivorans pre- and posttransplantation.(A) SDS gel of extracted Lipo-oligosaccharide from clonal strains suggestedmigration differences when comparing pre-transplant (lane 1) to posttransplant(lane 2). (B) Typical pulsed field gel electophoretogram of paired pre- andposttransplant strain after SpeI enzyme digestion. The index case was knownto have B. multivorans infection pretransplant. Lane 1 represents molecularweight markers. PFGE analysis demonstrated that the pretransplant (lane 2)and posttransplant strain (lane 3) were clonally identical (adapted from DeSoyza, Morris, et al. 2004).
Monosaccharide and fatty acid analyses of LOS isolated fromB. multivorans pre- and posttransplantation showed the samecontent in sugar and fatty acids but the presence of an additional8-substituted D-glycero-D-talo-oct-2-ulosonic acid (Ko) residuewas detected only in the posttransplantation strain. The fullcompositional analysis of all fractions is reported in the text andTable I of Supplementary Data.
Structural characterization of Fr1 product from the B.multivorans pretransplantation strainThe usual mild acid treatment on pretransplantation strain LPSyielded two fractions, Fr1 and Fr2. The monosaccharide anal-ysis of both fractions is shown in the text and Table I availablein Supplementary Data.
In order to assign all the spin systems of Fr1 and to de-fine the monosaccharide sequence, a combination of homo-and heteronuclear 2D NMR experiment (DQF-COSY, TOCSY,ROESY, NOESY, 1H-13C HSQC, 1H-13C HSQC-TOCSY and1H-13C HMBC, see supporting Figure 3A–F) was undertaken.In the anomeric region of the 1H-NMR spectrum (Figure 2A),12 anomeric signals were identified (A–N, Table II available inSupplementary Data). Furthermore, the signals at 1.94/2.05 ppmwere identified as the H-3 methylene protons of the Kdo residue(K) whereas residue J of α-Ko was assigned starting from itsoxymethine H-3 signal. The relative intensities of anomeric
872
by guest on May 29, 2013
http://glycob.oxfordjournals.org/D
ownloaded from

Lipopolysaccharide structure from Burkholderia multivorans
Fig. 2. NMR structural investigation of Fr1 product. (A) 1H NMR spectrum of Fr1 product with a zoom of the anomeric region. (B) Zoom of the ROESY (red) andTOCSY (black) spectra of oligosaccharide Fr1. Monosaccharide labels are indicated in Supplementary Data Table II whereas complete NMR experimentalconditions are given in Supplementary Data.
signals suggested the existence of a mixture of oligosaccharideswith different length of the carbohydrate chain.
In accordance with the chemical analysis, spin systems A, B,E, G were all identified as L-glycero-D-manno-heptose whereasspin systems C and I were identified as 2-deoxy-2-amino-
galactose, both acetylated at C-2 position. Spin systems Mand N were identified as glucose residues while D, H, andH′ were recognized as α-rhamnose residues. Residue L (H-1 at4.68 ppm) was recognized as a β-quinovosamine acetylated atC-2 position.
873
by guest on May 29, 2013
http://glycob.oxfordjournals.org/D
ownloaded from

T Ierano et al.
Fig. 3. Negative-ion MALDI mass spectrum of Fr1 and Fr2 oligosaccharides.(A) Species X, X-QuiNAc, and Y, matching with NMR analysis, are the mainpeaks. Ions identified as lipid A species (indicates with asterisks) are alsovisible. (B) High-resolution negative MALDI mass spectrum of Fr2oligosaccharide acquired in reflectron mode. Peaks R and S are related tooligosaccharides X and Y lacking the Ko moiety, respectively. Ions due tospecies missing QuiNAc and Hep residues are also present.
The oligosaccharide sequence reported below (species X)was established identifying the interresidual NOE contacts(Figure 2B) and the long-range correlations present in theHMBC spectrum:
An alternative glycoform of the α-rhamnose D was found,namely residue F, identified as a 3-α-Rha residue. This wasglycosylated by residue I of β-GalNAc that was in turn sub-stituted at O-3 by the α-GalNAc C. Additionally, residue Cwas glycosylated at position 3 by the terminal β-Glc N. Thelinkage between these residues was attested by NOE contacts(Figure 2B) and confirmed by scalar correlations in the HMBCspectrum. Thus, the oligosaccharide sequence (species Y) re-ported below differed from the previous one for the presence ofthe additional terminal trisaccharide:
MALDI-MS characterization of Fr1 and Fr2 products from B.multivorans pretransplantationMatrix-assisted laser desorption ionisation-mass spectrometry(MALDI-MS) confirmed the oligosaccharide structures hypoth-esized above. The negative mass spectrum (Figure 3A) showedtwo peaks, at m/z 1883.2 and at m/z 2264.1, matching with theoligosaccharide X and Y (�m/z ∼= 568 Th from 1696.3) as-signed by NMR analysis and a further peak at m/z 1696.3 thatdiffered from X by a β-QuiNAc residue. Likewise, Fr2 frac-tion underwent mass spectrometric investigation. The negative-ion MALDI mass spectrum of the oligosaccharide mixture Fr2(Figure 3B) showed ion peaks related to Fr1 oligosaccharidesbut lacked the Ko residue (�m/z ∼= 236 Th).
Collectively, these results yielded the full oligosaccharide se-quence of the B. multivorans pretransplantation LPS as depictedbelow:
MALDI-MS characterization of oligosaccharide product fromthe B. multivorans posttransplantation strainAs compared to the pretransplantation strain, the LOS extractedfrom the B. multivorans posttransplantation strain underwentthe same treatment and one oligosaccharide fraction, namelyFr, was isolated by gel-permeation chromatography. The com-positional analysis of the fractions is reported in Table I avail-able in Supplementary Data. Both the NMR and the MS datagave evidence of a saccharide sequence identical to the Fr2fraction found in the pretransplantation strain (data not shown).The matrix-assisted laser desorption ionization-time of flight(MALDI-TOF) spectra of Fr fraction are reported in Supple-mentary Data Figure 2.
874
by guest on May 29, 2013
http://glycob.oxfordjournals.org/D
ownloaded from
Lipopolysaccharide structure from Burkholderia multivorans
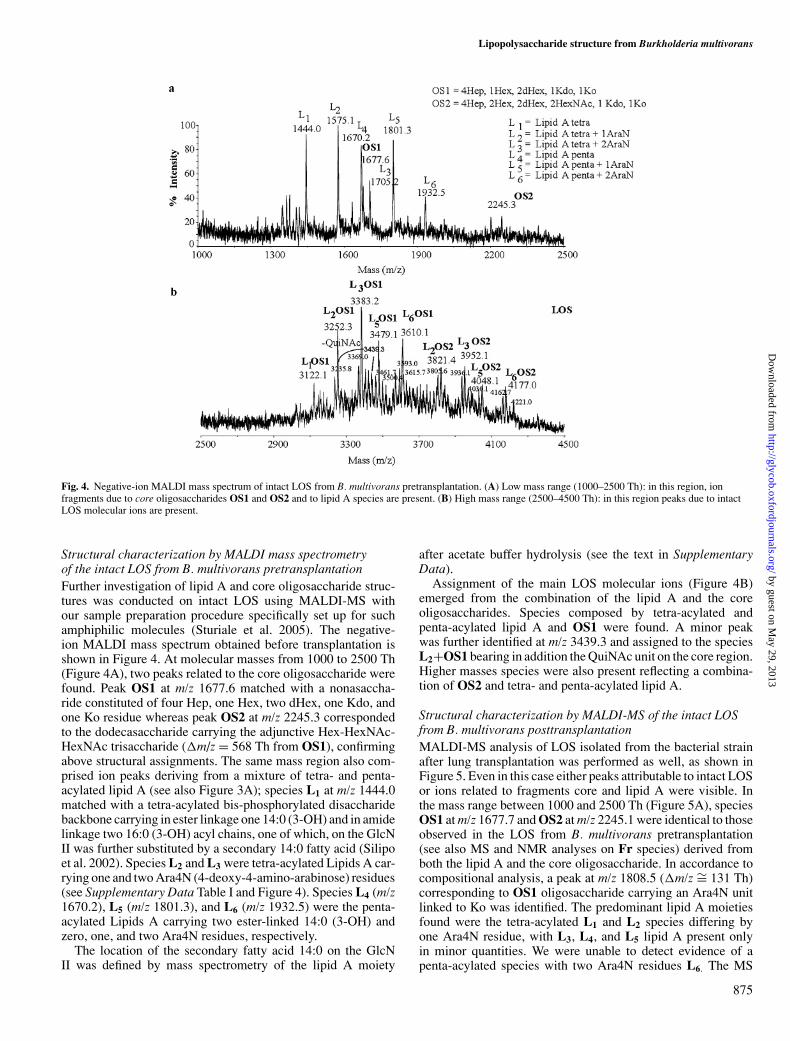
Fig. 4. Negative-ion MALDI mass spectrum of intact LOS from B. multivorans pretransplantation. (A) Low mass range (1000–2500 Th): in this region, ionfragments due to core oligosaccharides OS1 and OS2 and to lipid A species are present. (B) High mass range (2500–4500 Th): in this region peaks due to intactLOS molecular ions are present.
Structural characterization by MALDI mass spectrometryof the intact LOS from B. multivorans pretransplantationFurther investigation of lipid A and core oligosaccharide struc-tures was conducted on intact LOS using MALDI-MS withour sample preparation procedure specifically set up for suchamphiphilic molecules (Sturiale et al. 2005). The negative-ion MALDI mass spectrum obtained before transplantation isshown in Figure 4. At molecular masses from 1000 to 2500 Th(Figure 4A), two peaks related to the core oligosaccharide werefound. Peak OS1 at m/z 1677.6 matched with a nonasaccha-ride constituted of four Hep, one Hex, two dHex, one Kdo, andone Ko residue whereas peak OS2 at m/z 2245.3 correspondedto the dodecasaccharide carrying the adjunctive Hex-HexNAc-HexNAc trisaccharide (�m/z = 568 Th from OS1), confirmingabove structural assignments. The same mass region also com-prised ion peaks deriving from a mixture of tetra- and penta-acylated lipid A (see also Figure 3A); species L1 at m/z 1444.0matched with a tetra-acylated bis-phosphorylated disaccharidebackbone carrying in ester linkage one 14:0 (3-OH) and in amidelinkage two 16:0 (3-OH) acyl chains, one of which, on the GlcNII was further substituted by a secondary 14:0 fatty acid (Silipoet al. 2002). Species L2 and L3 were tetra-acylated Lipids A car-rying one and two Ara4N (4-deoxy-4-amino-arabinose) residues(see Supplementary Data Table I and Figure 4). Species L4 (m/z1670.2), L5 (m/z 1801.3), and L6 (m/z 1932.5) were the penta-acylated Lipids A carrying two ester-linked 14:0 (3-OH) andzero, one, and two Ara4N residues, respectively.
The location of the secondary fatty acid 14:0 on the GlcNII was defined by mass spectrometry of the lipid A moiety
after acetate buffer hydrolysis (see the text in SupplementaryData).
Assignment of the main LOS molecular ions (Figure 4B)emerged from the combination of the lipid A and the coreoligosaccharides. Species composed by tetra-acylated andpenta-acylated lipid A and OS1 were found. A minor peakwas further identified at m/z 3439.3 and assigned to the speciesL2+OS1 bearing in addition the QuiNAc unit on the core region.Higher masses species were also present reflecting a combina-tion of OS2 and tetra- and penta-acylated lipid A.
Structural characterization by MALDI-MS of the intact LOSfrom B. multivorans posttransplantationMALDI-MS analysis of LOS isolated from the bacterial strainafter lung transplantation was performed as well, as shown inFigure 5. Even in this case either peaks attributable to intact LOSor ions related to fragments core and lipid A were visible. Inthe mass range between 1000 and 2500 Th (Figure 5A), speciesOS1 at m/z 1677.7 and OS2 at m/z 2245.1 were identical to thoseobserved in the LOS from B. multivorans pretransplantation(see also MS and NMR analyses on Fr species) derived fromboth the lipid A and the core oligosaccharide. In accordance tocompositional analysis, a peak at m/z 1808.5 (�m/z ∼= 131 Th)corresponding to OS1 oligosaccharide carrying an Ara4N unitlinked to Ko was identified. The predominant lipid A moietiesfound were the tetra-acylated L1 and L2 species differing byone Ara4N residue, with L3, L4, and L5 lipid A present onlyin minor quantities. We were unable to detect evidence of apenta-acylated species with two Ara4N residues L6. The MS
875
by guest on May 29, 2013
http://glycob.oxfordjournals.org/D
ownloaded from

T Ierano et al.
Fig. 5. Negative-ion MALDI mass spectrum of intact LOS from B. multivorans posttransplantation. (A) In the low mass range, the presence of oligosaccharidespecie OS1 carrying an additional Ara4N residue is noteworthy. (B) High mass region of the same spectrum: the majority of peaks originate from multiplyassignments due to nonstoichiometric linkage of a Ara4N residue with the core portion or the lipid A species.
profile related to LOS molecular ions (Figure 5B) showed peaksreflecting quantitatively the relative amount of lipid A species.The main ion peaks were given by the combination of OS1 withtetra-acylated lipid A (L1 + OS1 at m/z 3122.2, L2 + OS1 at m/z3253.1, and L3 + OS1 at m/z 3384.1). The possibility of havingan Ara4N residue nonstoichiometrically linked to the lipid A orto the core region led to multiple assignments, as indicated inFigure 5B. An ion peak at m/z 3439.9 was present, reflecting thenonstoichiometric amount of terminal QuiNAc residue.
The above MS results on intact LOSs are consistent withthe previous structural hypotheses allowing a precise molecularcharacterization. It is also noteworthy that such careful massspectrometry approaches disclose new avenues on the analy-sis on the intact LPS/glycolipid-like biomolecules excludingany structural alteration owing to the chemical manipulationof samples. Thus, the lipo-oligosaccharide from B. multivoransstrains isolated pre- and post-lung transplantation are reportedin Figure 6.
Biological activity of B. multivorans LOSsThe LOS molecules isolated pre- and post-lung transplanta-tion were tested for their proinflammatory activity of elicitingTNF-alpha induction from human myelomonocytic U937 cells(Figure 7A).
These findings are in agreement with our prior data andshowed that B. cenocepacia ET-12 LMG 12614 epidemic strainand E. coli 055 were more potent TNF-alpha inducers than anyof the B. multivorans strains tested (LMG 14273 and the pairedpre- and posttransplant strains).
Assessment of lipid A and LOS activity using transienttransfection of TLR 4/MD2/CD 14 into HEK cellsThere was, in general, a greater biological activity at inductionof NF-KB observed for the LOS as compared to the extractedlipid A (Figure 7B and C). This implies a biological role for thecore region. All stimulants had relatively potent proinflamma-tory activity attaining similar levels of NF-KB at ng concentra-tions in similar to that of the positive control E. coli O55 LPS(10 ng/mL). The NF-KB induction activity of the B. multivo-rans strains was however less potent than that achieved by theB. cenocepacia ET-12 strain (not shown).
The B. multivorans lipid A isolated prior to transplantationinduced greater activation in the NF-KB reporter assay than af-ter transplantation. Using the NF-KB luciferase assay the proin-flammatory activity of extracted lipid A from the B. multivoranspretransplantation strain (A pre 10) was significantly higher thanthat of lipid A from the posttransplant strain (A post 10), pair-wise t-test comparison of data with pooled standard deviations,p < 0.05. This was also significantly tested at both concentra-tions from the B. multivorans strain. This confirms the signifi-cant differences observed in the cytokine output in the U937 as-say that were likely to reflect signaling through the TLR4/MD2complex.
Discussion
Infection with the B. cepacia complex (BCC) is problematic inpatients with cystic fibrosis reflecting a remarkable antibiotic re-sistance and the potential for epidemic spread. B. multivorans,
876
by guest on May 29, 2013
http://glycob.oxfordjournals.org/D
ownloaded from

Lipopolysaccharide structure from Burkholderia multivorans
Fig. 6. The full structure of R-type LPS from B. multivorans pre- andposttransplantation strains. Dotted lines indicate the nonstoichiometricsubstitutions which are present in very different amounts between LOSs fromB. multivorans pre- and posttransplantation (see text, figures, andSupplementary Data). The Ara4N moiety in the circle was found exclusivelyin LOS isolated from the posttransplantation strain while the one in therectangle is prevalently present in pretransplantation lipid A moiety.
a member of the BCC, is prevalent in European CF centers.Interestingly, this organism appears to be associated with lesssevere clinical outcomes than those seen in patients infectedwith B. cenocepacia. Prior in vitro data have suggested thatB. multivorans genomovar strains have a reduced capacity toelicit proinflammatory cytokines as compared to B. cenocepacia(De Soyza, Ellis, et al. 2004). Recent studies have demonstrateddivergent proinflammatory signaling pathways for these endo-toxins. In an in vitro model with macrophages as target cells,B. multivorans LPS promoted a My-D88 independent inflam-matory pathway whilst that elicited by B. cenocepacia LPS wasMy-D88 dependent (Bamford et al. 2007). This qualitative di-vergence in signaling pathways could be a possible explanationof our prior data. Previously, we noted qualitative differences incytokine induction patterns: B. multivorans LOS induced IL-6production from U937 macrophages peaking at 24–48 h whilstB. cenocepacia LOS induced production continued to increaseup to 60 h poststimulation (De Soyza, Ellis, et al. 2004). Thesecombined observations in conjunction offer possible explana-tions for the different clinical outcomes observed in patientsinfected with B. cenocepacia and B. multivorans (Bamfordet al. 2007) (De Soyza, Ellis, et al. 2004).
Fig. 7. Biological assays of pre- and post- transplantation isolated LOS andlipid A moieties. (A) TNF induction from U937 cells elicited by standard LPSand paired clonal strains. Key: histograms represent TNF induction (ng) at24 h. U937 cells were stimulated with 100 ng of purified LPS including E. coliO55 as positive control. US represents unstimulated cells (negative control).Purified LPS from strains LMG 14273 (B. multivorans) and LMG 12614 (B.cenocepacia ET-12 strain) as previously reported (De Soyza et al. 2004) wasalso used as comparators. “Pre” represents the TNF induction elicited by thepretransplantation clonal B. multivorans strain. “Post” represents thebiological activity of 100 ng of extracted LOS from the posttransplantationstrain which was significantly lower than the pretransplant strain (p < 0.001).(B, C) In these figures, the NF-KB induction elicited by extracted lipid A andLPS/LOS in transfected TLR 4/MD2/CD 14 HEK cell lines is reported. Infigures, pre- and post- represent a B. multivorans strain pre- andposttransplantation, respectively, at doses of 10 ng/mL (vertical striped bar) or100 ng/mL (checkerboard bar). Negative control (culture media only) isrepresented by C (open bar) and E. coli (closed bar) represents the biologicalactivity of 10 ng/mL of the positive control E. coli 055 LPS. The reporter assayis measured in relative light units (RLU).
The full primary structure of lipopolysaccharides from BCCgenomovars and LPS structure/function relationships are there-fore clinically relevant. Such studies represent a contributivestep in the understanding of molecular mechanisms involved inthe inflammatory process. We report for the first time the com-plete structure of the LOS from B. multivorans clinical isolates
877
by guest on May 29, 2013
http://glycob.oxfordjournals.org/D
ownloaded from

T Ierano et al.
and also report the structural modifications that B. multivoransLOS undergoes after a change in host microenvironment as seenfollowing lung transplantation.
The CF airway is known to be associated with inducing spe-cific changes in the lipid A of both Pseudomonas and Burkholde-ria species (Ernst et al. 1999; De Soyza, Ellis, et al. 2004). Itis not, however, known if these changes are reversible nor overwhat timescale any reversibility occurs. Studying strains thathave coexisted in vivo with CF airway epithelia and that aresubsequently required to interact with non-CF (normal) airwayepithelial following transplantation may offer powerful insightsinto host–pathogen relationships.
The analysis of core oligosaccharide from B. multivoranspre- and post-lung transplantation clinical isolates revealed anovel oligosaccharide structure. The Burkholderia genus has ahighly conserved LPS inner core structure with several char-acteristics replicated in the current study. The main feature isthe presence of the [3,4-α-L,D-Hep-(1→5)-α-D-Kdo-(4→2)-α-D-Ko] trisaccharide in the inner core. Notably, within the genusthe Ko residue is frequently not terminal but further links anAra4N residue that, in the present case, was only present inthe posttransplantation strain. Usually the heptose residue bearsanother heptose linked at C-3 and glycosylated at C-4 by a β-D-Glc; this latter can be optionally glycosylated either by terminalα-Glc (Molinaro et al. 2002; Silipo et al. 2007) or as in thepresent case by a terminal α-Hep, composing a Hep-(1→6)-Glcdisaccharide. Moreover, in the outer core region, the presenceof a nonstoichiometric [β-D-Glc-(1→3)-β-D-GalNAc-(1→3)-α-D-GalNAc] trisaccharide sitting on a rhamnose residue isnoteworthy as it is likely preassembled and then attached.
The main structural changes between the clonal strains werefound in the different distribution of the lipid A species. Thelipid A moiety from both species possessed a carbohydratebackbone characterized by a [P→4-β-D-GlcpN-(1→6)-α-D-GlcpN-1→P→Ara4N] sequence, already found in other BCCgenomovars such as B. cenocepacia and B. pyrrocinia (Silipoet al. 2006, 2007). The lipid A from B. multivorans pretrans-plantation isolate comprised a mixture of penta- and tetra-acylated species in similar proportions carrying up to two Ara4Nresidues. The posttransplantation B. multivorans strains’ LPSdemonstrated marked changes in the distribution and composi-tion of lipid A species with an overall lower level of acylationand substitution by Ara4N. In particular, there was a negligibleamount of penta-acylated species with a predominance of tetra-acylated lipid A. Furthermore, each species carried no more thanone Ara4N residue.
The core oligosaccharide sequence of the B. multivoransspecies studied was distinct from that of B. cenocepacia (Silipoet al. 2007). It has been postulated that B. cenocepacia biologicalactivity and survival in vivo is in part dependent on the presenceof a complete LOS as a B. cenocepacia mutant strain lacking acomplete LPS core oligosaccharide is sensitive to antimicrobialpeptides and polymyxin B (Loutet et al. 2006). Further evidencefor an important biological activity of the core oligosaccharidewas suggested by the higher levels of NF- induction KB bythe complete LOS as compared to lipid A (Figure 7B and C).The differences in the core oligosaccharide between the twogenomovars could contribute to clinical phenotypes (inflamma-tory syndromes).
The core portion in B. multivorans is likely to contribute tobacterial resistance to antimicrobial compounds. The net charge
surface on the external membrane of Gram-negative bacteria isfelt to play a key role in pathogenesis. LPSs from the Burkholde-ria genus are frequently positively charged or in an isoelectricstate. This is closely related to the abundance of aminoarabinose(Ara4N) residues present in the lipid A-inner core and confersresistance to antibiotic compounds and host cationic antimicro-bial peptides. Under physiological conditions, Ara4N moietiesare positively charged and reduce the net charge surface onthe external membrane weakening the ionic attraction for res-piratory tract antimicrobial defensins. The inherent resistanceof these microorganisms to polymyxin B, a cyclic polycationicantibiotic peptide that has high affinity for negatively chargedbacterial LPS, is likely to be Ara4N dependent. These residuesprevent the antimicrobial action of polymyxin B, increasingpermeability in a bacterial outer membrane (Shimomura et al.2003).
Peritransplantation processes that promote increased bacte-rial defensin resistance are suggested by our observation of afurther Ara4N residue on the inner core portion of LOS fromB. multivorans posttransplantation. There are many possible ex-planations for this observation including the effect of antibioticsadministered during the transplant operation or an as yet, un-described effect of immunosuppressant agents. However, it isinteresting to speculate that, posttransplantation, the replace-ment of CF epithelium with a normal airway epithelium wouldincrease defensin activity by normalizing high airway sodiumcontent (a known inhibitor of defensin activity).
Differences in SDS gel migration patterns of LOSs were notobserved in every clonal-paired strain from our transplant cohort(data not shown). It was observed in one of the four recipientswith pretransplant B. multivorans infection. This suggests thatthe phenomenon may be strain specific and was not a uniformeffect as may be expected if peritransplant immunosuppressivedrug therapy or antimicrobials were a cause.
Lipid A contributes to the majority of endotoxic activity ofLPS. A number of factors influence the lipid A biological activ-ity including the number and the distribution of acyl chains, thephosphorylation pattern, and the presence of charged groups onthe polar heads. The correlation between increasing acylationof lipid A and a greater cytokine induction capacity has pre-viously been reported in Pseudomonas (Ernst et al. 1999) andBurkholderia (De Soyza, Ellis, et al. 2004). The lipid A fromB. multivorans pretransplantation had a comparable amount ofpenta- and tetra-acylated lipid A species and a notable abun-dance of Ara4N moieties. These data in part confirm our previ-ous report comparing B. multivorans and B. cenocepacia withhigher levels of lipid A acylation noted in B. cenocepacia(De Soyza, Ellis, et al. 2004). Interestingly, the lipid A from B.multivorans posttransplantation was less toxic in our U937 celland HEK TLR4/MD2 bioreporter assays consistent with theobserved relative reduction in penta-acylated lipid A species.These data confirm that the lipid A changes are recognized atthe human TLR-4 receptor. A reduction in cytokine inductionmay allow bacteria to survive in a more hostile environment(Munford and Varley 2006; Reife et al. 2006; Raetz et al. 2007).
Other novel findings presented suggest that expressed lipidA species are relatively rapidly altered to reflect changes inhost–pathogen interfaces (Munford and Varley 2006; Raetzet al. 2007). The mechanisms responsible for controlling lipidA biosynthesis remain unclear though recent reports have elu-cidated the pathways for aminoarabinose biosynthesis (Ortega
878
by guest on May 29, 2013
http://glycob.oxfordjournals.org/D
ownloaded from

Lipopolysaccharide structure from Burkholderia multivorans
et al. 2007). A critical role of airway adaptation in cystic fibrosis-related Pseudomonas aeruginosa strains has been seen (Ernstet al. 1999; Hajjar et al. 2002). Interestingly, these studiesdemonstrated that a specific penta-acylated LPS was found inmild/moderate CF lung disease and hepta-acylated species werefound in severe CF disease. Such data in conjunction with ourobservations suggest that increasing acylation of lipid A in cys-tic fibrosis may be a slow adaptation to the CF environment(reflecting disease progression) that can be more rapidly down-regulated once selective pressures are removed (e.g., removalof the CF microenvironment). Notably, the finding of species-specific recognition of the CF Pseudomonas adaptation to theCF airway suggests that bacterial adaptation to the airway re-flects the host innate immune system (Hajjar et al. 2002). In thenew microenvironment post transplantation, B. multivorans de-creases the immunostimulant activity of lipid A by decreasingits acylation pattern and reducing the consequent host immuneresponse. A further adaptation that increases the antimicrobialresistance is the addition of an Ara4N residue to the outer coreKo residue. Further elucidation of controllers of acylation statusmay offer the development of therapeutic strategies and newtreatments for the management of Gram-negative infections incystic fibrosis.
In conclusion, in this work we have chemically and biolog-ically defined for the first time the LPS endotoxin from B.multivorans. We have also investigated the molecular modifi-cations that B. multivorans LOS undergoes after lung transplan-tation. This study improves the understanding of the endotoxinstructure–activity relationship, which is of pivotal importancein the comprehension of the overall process of pathogenesis ofsuch important microorganisms.
Material and methods
Bacterial growth and LPS extractionBacterial strains isolated from recipients were stored on mi-crobeads. These were streaked on to LB-agar. Standard hotphenol/water LPS extraction was undertaken as previously de-scribed (Westphal and Jahn 1965). Large volume extractionswere carried out as described and the complete experimentaldetails are given in Supplementary Data. LPS fractions wereanalyzed by SDS–PAGE on 16% gels, which were stained withsilver nitrate.
Patient selectionCystic fibrosis patients were listed for transplantation accordingto international guidelines (Orens et al. 2006). Patients who hadpaired B. cepacia complex isolates pre- and posttransplantationthat were recoverable from storage repositories were consideredeligible for study selection.
Strain preparation and selectionSputum was collected from patients immediately before surgery.Posttransplant lower airway (bronchoalveolar) lavages were col-lected from recipients at day 7. Presumed B. cepacia complexbacteria were isolated by culture. For this study we restrictedinvestigations to B. multivorans strains.
Pulsed-field gel electrophoresisConfirmation of the clonal nature of paired pre- and posttrans-plant strains has previously been reported (De Soyza, Morris,et al. 2004). B. cepacia complex strains were blinded to theinvestigators and genotyped by macrorestriction of whole ge-nomic DNA with the restriction enzyme SpeI (New EnglandBiolabs, UK), followed by separation of the fragments by pulsefield gel electrophoresis (PFGE) (CHEF DRII system; Bio-Rad,CA).
SDS electrophoresis analysis and biological activity of B.multivorans LOSsExtracted LPS or LOS were subjected to SDS–PAGE on 16%SDS gels and silver stained as previously (De Soyza et al. 2004).B. multivorans LOS was tested for its proinflammatory activityfor TNF-alpha induction in human myelomonocytic U937 cellsas described (De Soyza, Morris, et al. 2004). Detailed method-ology is available in Supplementary Data.
Assessment of lipid A and LOS activity using transienttransfection of TLR 4/MD2/CD 14 into HEK cellsHEK293 cells were maintained in Dulbecco’s modified Eagle’smedium (DMEM) supplemented with 10% fetal calf serum,2 mM L-glutamine, 100 U/mL penicillin, and 100 µg/mL strep-tomycin. Cells were seeded at 3 × 104/well of a 96-well plateand transiently transfected 2 days later. Expression vectors con-taining cDNA encoding TLR4, MD-2, and CD14 (1 ng/well ofeach), a NFκB transcription reporter vector encoding firefly lu-ciferase (5 ng/well pNFκB-luc, Clontech), and a constitutivelyactive reporter vector encoding Renilla luciferase (5 ng/wellphRG-TK, Promega), together with empty vector to ensure anoptimal amount of DNA were mixed with JetPEI (Polyplustransfection) according to the manufacturer’s instructions. After48 h cells were stimulated with LOS and lipid A at two con-centrations 10 ng and 100 ng of stimulant for 6 h and comparedto control (media only) and compared to a positive control oftransfected cells stimulated with 10 ng of E. coli 055 strain LPS(Sigma, UK). The effect of stimulation was assessed using a NF-KB luciferase reporter assay (Dual Luciferase assay, Promega,Southampton, UK). The luciferase results were normalized tothe constitutively active renilla transfection control activity andresults were expressed as a percentage of the positive control.Experiments were repeated in quadruplicate.
Structural characterization of oligosaccharides Isolationof Oligosaccharide Fr1, Fr2, and FrThe lipid A fractions and core oligosaccharides (Fr1, Fr2, andFr) from B. multivorans pre- and posttransplantation LPS wereobtained by acetate buffer hydrolysis at 100◦C for 3 h as de-scribed (Silipo et al. 2007).
General and analytical methodsDetermination of sugars residues, their absolute configuration,methylation analysis, fatty acid, and their absolute configura-tions were all determined as described previously and com-pletely described in Supplementary Data (Hakomori 1964;Rietschel 1976; Leontein and Lonngren 1978; Silipo et al. 2007).
879
by guest on May 29, 2013
http://glycob.oxfordjournals.org/D
ownloaded from

T Ierano et al.
NMR spectroscopyIn all structural assignments, 1D and 2D 1H-NMR spectra wererecorded with a solution of 0.5–1 mg in 0.5 mL of D2O, at300 K, at pD 7 (uncorrected value) on Bruker 600 DRX equippedwith a cryo probe. Spectra were calibrated with internal acetone(δH 2.225, δC 31.45). 31P NMR experiments were carried out us-ing a Bruker DRX-400 spectrometer, aqueous 85% phosphoricacid was used as external reference (0.00 ppm). The full struc-tural determination and NMR experiments used are reported inSupplementary Data.
MALDI-TOF mass spectrometryMALDI-TOF mass spectra were recorded in negative or in pos-itive polarity on a Perseptive (Framingham, MA) Voyager STRinstrument equipped with delayed extraction technology. High-resolution mass spectra were acquired in reflector mode on a4800 Proteomics analyzer MALDI-TOF/TOF mass spectrome-ter (Applied Biosystems, Framingham, MA). Sample prepara-tions and details of mass spectra executed are fully described inSupplementary Data.
Supplementary Data
Supplementary data for this article is available online athttp://glycob.oxfordjournals.org/.
Funding
Italian Cystic Fibrosis Research Foundation (grant FFC 11/2006to A.M.)
Acknowledgements
A.D.S was supported by the Special Trustees of The Newcastleupon Tyne Teaching Hospitals and a Higher Education FundingCouncil for England (HEFCE) Senior lecturership. We are grate-ful to Professor John Govan, University of Edinburgh, UK, forthe pulsed field gel electrophoresis experiments as previouslypublished, and Christine Aldridge for technical support. We arealso grateful to Dr T. J. McKinely, Department of VeterinaryMedicine, University of Cambridge, for statistical advice.
Conflict of interest statement
None declared.
Abbreviations
Ara4N, 4-deoxy-4-amino-L-arabinose; BCC, Burkholderiacepacia complex; CF, cystic fibrosis; DQF-COSY, double quan-tum filtered correlation spectroscopy; HMBC, heteronuclearmultiple bond correlation; HSQC, heteronuclear single quantumcoherence; Kdo, 3-deoxy-D-manno-oct-2-ulosonic acid; Ko, D-glycero-D-talo-oct-2-ulosonic acid; LOS, lipo-oligosaccharideLPS, lipopolysaccharides; MALDI-MS, matrix-assisted laserdesorption ionisation-mass spectrometry; NOE, nuclear Over-hauser effect; NOESY, nuclear Overhauser enhancementspectroscopy; PAMP, pathogen-associated molecular pattern;
ROESY, rotating frame Overhauser enhancement spectroscopy;SDS–PAGE, sodium dodecyl sulphate–polyacrylamide gel elec-trophoresis; TOCSY, total correlation spectroscopy; TOF, time-of-flight.
References
Akira S, Uematsu S, Osamu T. 2006. Pathogen recognition and innate immunity.Cell. 124:783–801.
Alexander C, Rietschel ET. 2001. Bacterial lipopolysaccharides and innateimmunity. J Endotoxin Res. 7:167–202.
Bamford S, Ryley H, Jackson SK. 2007. Highly purified lipopolysaccharidesfrom Burkholderia cepacia complex clinical isolates induce inflammatorycytokine responses via TLR4-mediated MAPK signalling pathways andactivation of NFkappaB. Cell Microbiol. 9:532–543.
Blackburn L, Brownlee K, Conway S, Denton M. 2004. ‘Cepacia syndrome’with Burkholderia multivorans, 9 years after initial colonization. J CystFibros 3:133–134.
De Soyza A, Corris PA. 2003. Lung transplantation and the Burkholderia cepa-cia complex. J Heart Lung Transplant. 22:954–958.
De Soyza A, Ellis CD, Khan CMA, Corris PA, Demarco de Hormaeche R.2004. Burkholderia cenocepacia lipopolysaccharide, lipid A, and proin-flammatory activity. Am J Respir Crit Care Med. 170:70–77.
De Soyza A, McDowell A, Archer L, Dark JH, Elborn SJ, Mahenthiralingam E,Gould K, Corris PA. 2001. Burkholderia cepacia complex genomovars andpulmonary transplantation outcomes in patients with cystic fibrosis. Lancet.358:1780–1781.
De Soyza A, Morris K, McDowell A, Doherty C, Archer L, Perry J, GovanJRW, Corris PA, Gould K. 2004. Prevalence and clonality of Burkholderiacepacia complex genomovars in UK patients with cystic fibrosis referredfor lung transplantation. Thorax. 59:526–528.
Ernst RK, Guo EC, Yi L, Lim KB, Burns JL, Hackett M, Miller SI. 1999.Specific lipopolysaccharide found in cystic fibrosis airway Pseudomonasaeruginosa. Science. 286:1561–1565.
Hajjar AM, Ernst RK, Tsai JH, Wilson CB, Miller S. 2002. Human toll-likereceptor 4 recognizes host-specific LPS modifications. Nature Immunol.3:354–359.
Hakomori S. 1964. A rapid permethylation of glycolipid, and polysaccharidecatalyzed by methylsulfinyl carbanion in dimethyl sulfoxide. J Biochem(Tokyo). 55:205–208.
Holst O. 1999. Chemical structure of the core region of lipopolysaccharides. In:Brade H, Morrsion DC, Opal S, Vogel S, editors. Endotoxin in Health andDisease. New York: Dekker. p. 115–154.
Jones AM, Dodd ME, Govan JRW, Barcus VA, Doherty C, Morris J, Webb AK.2004. Burkholderia cenocepacia and Burkholderia multivorans: Influenceon survival in cystic fibrosis. Thorax. 59:948–951.
Leontein K, Lonngren J. 1978. Determination of the absolute configuration ofsugars by gas-liquid chromatography of their acetylated 2-octyl glycosides.Methods Carbohydr Chem. 62:359–362.
Loutet SA, Flannagan RS, Kooi C, Sokol PA, Valvano MA. 2006. A completelipopolysaccharide inner core oligosaccharide is required for resistance ofBurkholderia cenocepacia to antimicrobial peptides and bacterial survivalin vivo. J Bacteriol. 188:2073–2080.
Mahenthiralingam E, Coenye T, Chung JW, Speert DP, Govan JRW,Taylor P, Vandamme P. 2000. Diagnostically and experimentally usefulpanel of strains from the Burkholderia cepacia complex. J Clin Microbiol.38:910–913.
Molinaro A, De Castro C, Lanzetta R, Evidente A, Parrilli M, Holst O. 2002.Lipopolysaccharides possessing two L-glycero-D-manno-heptopyranosyl-alpha-(1→5)-3-deoxy-D-manno-oct-2-ulopyranosonic acid moieties in thecore region. The structure of the core region of the lipopolysaccharides fromBurkholderia caryophylli. J Biol Chem. 277:10058–10063.
Munford RS, Varley AW. 2006. Shield as signal: Lipopolysaccharides and theevolution of immunity to gram-negative bacteria. PLOS Pathog. 2:467–471.
Orens JB, Estenne M, Arcasoy S, Conte JV, Corris P, Egan JJ, Egan T, KeshavjeeS, Knoop C, Kotloff R, et al. 2006. International guidelines for the selectionof lung transplant candidates: 2006 Update—A consensus report from thePulmonary Scientific Council of the International Society for Heart andLung Transplantation. J Heart Lung Transplant. 25:745–755.
Ortega XP, Cardona ST, Brown AR, Loutet SA, Flannagan RS, Campopiano DJ,Govan JR, Valvano MA. 2007. A putative gene cluster for aminoarabinosebiosynthesis is essential for Burkholderia cenocepacia viability. J Bacteriol.189:3639–3644.
880
by guest on May 29, 2013
http://glycob.oxfordjournals.org/D
ownloaded from

Lipopolysaccharide structure from Burkholderia multivorans
Raetz CR, Reynolds CM, Trent MS, Bishop RE. 2007. Lipid A modificationsystems in gram-negative bacteria. Annu Rev Biochem. 76:295–329.
Raetz CR, Whitfield C. 2002. Lipopolysaccharide endotoxins. Annu RevBiochem. 71:635–700.
Reife RA, Coats SR, Al-Qutub M, Dixon DM, Braham PA, Billharz RJ, HowaldWN, Darveau RP. 2006. Porphyromonas gingivalis lipopolysaccharide lipidA heterogeneity: Differential activities of tetra- and penta-acylated lipid Astructures on E-selectin expression and TLR4 recognition. Cell Microbiol.8:857–868.
Rietschel ET. 1976. Absolute configuration of 3-hydroxy fatty acids presentin lipopolysaccharides from various bacterial groups. Eur J Biochem.64:423–428.
Seltmann G, Holst O. 2001. The Bacterial Cell Wall. Heidelberg: Springer.Shimomura H, Matsuura M, Saito S, Hirai Y, Isshiki Y, Kawahara K. 2003. Un-
usual interaction of a lipopolysaccharide isolated from Burkholderia cepaciawith polymyxin B. Infect Immun. 71:5225–5230.
Silipo A, Lanzetta R, Amoresano A, Parrilli M, Molinaro A. 2002. Am-monium hydroxide hydrolysis: A valuable support in the MALDI-TOFmass spectrometry analysis of lipid A fatty acid distribution. J Lipid Res.43:2188–2195.
Silipo A, Molinaro A, Comegna D, Sturiale L, Cescutti P, Garozzo D, ParrilliM, Lanzetta R. 2006. Full structural characterisation of the lipooligosaccha-ride of a Burkholderia pyrrocinia clinical isolate. Eur J Org Chem. 4874–4883.
Silipo A, Molinaro A, Ierano T, De Soyza A, Sturiale L, Garozzo D, AldridgeC, Corris PA, Khan CMA, Lanzetta R, et al. 2007. The complete struc-ture and pro-inflammatory activity of the lipooligosaccharide of the highlyepidemic and virulent gram-negative bacterium Burkholderia cenocepaciaET-12 (strain J2315). Chemistry. 13:3501–3511.
Speert DP. 2002. Advances in Burkholderia cepacia complex. Paediatr RespirRev. 3:230–235.
Sturiale L, Garozzo D, Silipo A, Lanzetta R, Parrilli M, Molinaro A. 2005. Newconditions for matrix-assisted laser desorption/ionization mass spectrom-etry of native bacterial R-type lipopolysaccharides. Rapid Commun MassSpectrom. 19:1829–1834.
Wellinghausen N, Kothe J. 2006. Evidence of coinfection with distinct strains ofBurkholderia multivorans in a cystic fibrosis patient. Infection. 34:289–291.
Westphal O, Jahn K. 1965. Bacterial lipopolysaccharides: Extraction withphenol-water and further applications of the procedure. Methods Carbo-hydr Chem. 5:83–91.
881
by guest on May 29, 2013
http://glycob.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















