
The role of holm oak edges in the control of disturbance and conservation of plant diversity in...
13
(This is a sample cover image for this issue. The actual cover is not yet available at this time.) This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
7 -
download
0
Transcript of The role of holm oak edges in the control of disturbance and conservation of plant diversity in...

(This is a sample cover image for this issue. The actual cover is not yet available at this time.)
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
The role of holm oak edges in the control of disturbance and conservationof plant diversity in fire-prone landscapes
J.C. Azevedo a,⇑, A. Possacos b, C.F. Aguiar a, A. Amado a, L. Miguel c, R. Dias a, C. Loureiro d, P.M. Fernandes d
a CIMO-Centro de Investigação de Montanha, Instituto Politécnico de Bragança, Campus de Santa Apolónia, Apartado 1172, 5301-855 Bragança, Portugalb Unidade de Gestão Florestal do Nordeste Transmontano, Instituto da Conservação da Natureza e das Florestas, Parque Florestal, 5300-000 Bragança, Portugalc Autoridade Nacional de Protecção Civil, Comando Distrital de Operações de Socorro de Bragança, 5301-864 Bragança, Portugald CITAB-Centro de Investigação e Tecnologias Agro-ambientais e Biológicas, Universidade de Trás-os-Montes e Alto Douro, 5001-801 Vila Real, Portugal
a r t i c l e i n f o
Article history:Received 25 October 2012Received in revised form 4 February 2013Accepted 8 February 2013
Keywords:EdgesFire behaviorPlant diversityQuercus rotundifoliaMediterranean regionPortugal
a b s t r a c t
Holm oak (Quercus rotundifolia) woodlands have been able to persist in fire-prone landscapes in northernPortugal where they seem to play a relevant role in community and landscape level patterns and pro-cesses, namely in the control of disturbance events and in the maintenance of biodiversity. In thisresearch we tested the following hypotheses: (i) holm oak woodlands affect fire behavior, mainly by dis-rupting and delaying fire spread; (ii) disruption and delaying of fire spread is due to fire extinction at theedge of woodlands; and (iii) plant diversity is favored by frequent low-intensity fires as a result of thecontrol of fire spread by holm oak woodland edges. We sampled herbaceous, shrub and tree structureand herbaceous plant composition across edges of holm oak woodlands with known recent contact withfire. Using BehavePlus we converted structural data into fuel models and simulated fire behavior for thelocations sampled along the exterior–interior gradient of the woodlands. Our results indicate that holmoak woodlands play a relevant role in the control of fire spread and in the maintenance of plant diversity.We found differences in simulated maximum rate of fire spread (ROS) and fireline intensity (FLI) betweenoutside and inside woodlands as well as a 100% probability of fire extinction when fuel moisture was>12% and wind speed at 10 m aboveground was <40 km h�1, supporting the self-extinction hypothesis.We found also that richness and abundance of herbaceous plants varied between the interior and exteriorof the woodland, with higher values outside the woodland, and that plant composition along the interior–exterior gradient had distinct patterns. These variables and the pattern of plant distribution were also sig-nificantly affected by time since last fire event. We conclude that holm oak woodlands affect fire behaviorby interrupting fire spread at the edge zone and that fire-created edges affect herbaceous plant speciesrichness, abundance and distribution by maintaining habitat diversity, which increased plant diversityat patch and landscape levels. Based on our results and other evidence, we developed a conceptual modelof the dynamics of fire, landscape structure, and plant diversity in fire-prone landscapes where holm oakpatches persist.
� 2013 Elsevier B.V. All rights reserved.
1. Introduction
In regions with distinct climates, such as the Mediterranean, fireis an integral component of the dynamics of landscapes and plantcommunities (Allen, 2008), often driving patterns and processes inthese systems (Loepfe et al., 2010; Turner, 1989). In the IberianPeninsula, the interaction of fire and landscape pattern is affectednot just by bioclimatic factors but also by present day and histor-ical landscape and ecosystem management practices (Pausas,
2006). Expansion of agriculture until the late 19th – early 20th cen-tury decreased landscape heterogeneity through dominance ofagriculture and extensive grazing causing fragmentation of naturalhabitats. These activities maintained vegetation fuel loads at lowlevels and a landscape configuration unfavorable to the occurrenceof large and intense fires. Recent abandonment of agriculture hasincreased landscape fire hazard (Azevedo et al., 2011; Loepfeet al., 2010; Viedma et al., 2006) not just because of vegetationencroachment in what was formerly farming or grazing land, butalso due to increasing connectivity of flammable land use/land cov-er (LULC) types in the landscape, namely forests and shrublands(Bielsa et al., 2005). These fast changing landscapes are now recog-nized as providers of valuable ecosystem services and, for that rea-son, many of these areas have received a conservation status.
0378-1127/$ - see front matter � 2013 Elsevier B.V. All rights reserved.http://dx.doi.org/10.1016/j.foreco.2013.02.007
⇑ Corresponding author. Tel.: +351 273 303 341; fax: +351 273 325 405.E-mail addresses: [email protected] (J.C. Azevedo), [email protected] (A. Possacos),
[email protected] (C.F. Aguiar), [email protected] (A. Amado), [email protected] (L. Miguel), [email protected] (R. Dias), [email protected](C. Loureiro), [email protected] (P.M. Fernandes).
Forest Ecology and Management 297 (2013) 37–48
Contents lists available at SciVerse ScienceDirect
Forest Ecology and Management
journal homepage: www.elsevier .com/locate / foreco

Author's personal copy
While forest edges are mainly created by management practices(logging, agriculture and grazing), fire can also be a major edge-shaping agent (Eberhart and Woodard, 1987; Hanson and Stuart,2005; Kremsater and Bunnell, 1999). Weather and terrain (Alexan-der et al., 2006), vegetation (Nunes et al., 2005; Perera et al., 2009;Viedma et al., 2009), and management (Fernandes, 2009) affect firebehavior and the severity of fire effects (Agee, 1998). The combina-tion of these agents often results in heterogeneous landscape pat-terns which include patches of remnant unburned vegetation andlow-severity fire (DeLong and Kessler, 2000; Roman-Cuesta et al.,2009; Fernandes et al., 2010). Low- and high-severity fires createand maintain fine-scale and coarse-scale patterns, respectively.The largest extent of forest edges, however, is observed in moder-ate-severity fire regimes (Agee, 1998).
Although this issue has seldom been explicitly addressed in theliterature, forest edges can have an important role in controllingfire spread. In a mountain landscape in central Spain, the limitsof burned areas were strongly related to differences in LULC com-bustibility prior to the occurrence of fire (Viedma et al., 2009). TheLULC classes that halted fire most frequently were pastures, fol-lowed by shrublands, cropland and deciduous woodlands. Land-scape (LULC) and topographic diversity stopped fire even inextreme weather conditions (Viedma et al., 2009). In Venezuela,fire did not enter forest galleries due to changes in fuel type andfuel moisture in the savanna–forest interface (Biddulph and Kell-man, 1998). Edges with low and medium fuel loadings acted asbarriers for fire spread in a simulated landscape in northern Wis-consin, USA, reducing total and mean burned area (LaCroix et al.,2008). In another simulation study, burned area decreased withincreasing landscape heterogeneity due to the reduction of firespread velocity in edges (Ryu et al., 2007). In Portugal, a significantdecrease in fire severity under extreme weather conditions wasdocumented in wildfires moving from maritime pine (Pinus pinas-ter) stands to adjacent broadleaved (deciduous and evergreen) andshort-needled conifer forest types (Fernandes et al., 2010).
Edges exert a strong influence on the vegetation structure offorest patches and on the spatial patterning of plant diversity atthe patch and landscape scales (Chen et al., 1992). Plant richnesstends to be higher near the edge than inside woodlands (Fraver,1994; Gehlhausen et al., 2000; Marchand and Houle, 2006), mainlydue to changes in radiation, moisture and temperature (Gignac andDale, 2007). Studies of plant diversity in fire-created/maintainededges are not abundant. In Indonesia, woody plant diversity waspositively associated with distance from edge in the burned por-tion of a tropical forest (Slik et al., 2011). In southeastern Australia,vascular plant richness in remnant patches of dry open-forest de-creased with time since fire (Ross et al., 2002). Distance to edgewas not addressed in the study. In Mediterranean systems, plantdiversity patterns across fire-created edges are unknown.
Holm oak (Quercus rotundifolia) woodlands were once dominantelements in many landscapes in the Iberian Peninsula. Today, theyare found mostly in a simplified savanna-like form, particularly inthe south, or as fragmented remnants dominated by edge effects inless productive sites. Q. rotundifolia is a stress tolerator capable ofthriving in rock outcrops and steep slopes. Following marginal landabandonment, reestablished woodlands may become seed sources.In the landscapes of northeastern Portugal, where these fragmentspersist, fire is very frequently used for vegetation/livestock andwildlife management purposes. Between 2001 and 2010, 33.5% offires that occurred in the Bragança municipality (the major munic-ipality in this region) were related to land management activities.
Holm oak typically resprouts vigorously after fire, facilitatingthe maintenance of woodlands in the landscape in locations wherethey existed prior to disturbance (Broncano et al., 2005). Thecapacity of post-fire resprouters to disperse over long distances isvery limited (Buhk et al., 2007) making their persistence difficult
after extreme disturbance events, particularly in fragmented land-scapes. Although frequent fires tend to favor post-fire resprouters(Buhk et al., 2007), holm oak is relatively vulnerable to repeateddisturbances (Bonfil et al., 2004).
In northeastern Portugal many holm oak woodlands survive fire(Azevedo and Caçador, 2000). Observed wildfires during the 1990sand 2000s have actually self-extinguished at the edge of thesewoodlands. Additional evidence of holm oak woodlands acting asfire barriers was obtained by comparing the mapped distributionof holm oak woodlands and burned areas in northeastern Portugal.We found that 25–30% of the existing woodlands have their cur-rent limits coincident with the limits of areas burned in the last15 yrs. We believe that this pattern would not be possible unlessthese woodlands were a barrier to fire spread.
In this research we tested the hypothesis that holm oak wood-lands affect fire behavior, mainly by disrupting and delaying firespread. We further hypothesized that this is due to fires extin-guishing at the edge of the woodlands, which can be explainedby the spatial distribution of fuel structure and load across theboundary of the woodland. Thus, at a broader level, scatteredpatches of holm oak in the landscape should affect the fire regime.Additionally, we hypothesized that in these landscapes plant diver-sity is favored by frequent low-intensity fires as a result of the con-trol of fire spread by holm oak woodland edges. According to thesehypotheses, fire spread control by holm oak woodlands and theiredges is not just a mechanism that increases fine-scale landscapeheterogeneity but also helps to maintain plant diversity at thecommunity and landscape levels.
2. Material and methods
2.1. Study area
This study was conducted in a 10,000 ha region comprising theparishes of Babe, Deilão and S. Julião (center approximately at 41�500 1000N, 6� 340 3000 W), Bragança municipality, northeastern Portu-gal (Fig. 1). This area is located in a 750–900 m elevation plateau ofmetamorphic origin broken by North–South oriented narrow rivervalleys. Soils are mostly leptosols. Climate is Mediterranean withcontinental influence. Mean annual temperature is 10–12.5 �Cand mean precipitation is approximately 800 mm. The landscapeis heterogeneous with agricultural land, forest/woodlands andshrublands in roughly equal amounts. Woodlands of holm oakand Pyrenean oak (Quercus pyrenaica) are the major natural forestsystems. Pine plantations (mainly P. pinaster), established after1958, now form a major component of the landscape. Shrublandsare dominated mainly by Cistus ladanifer and Erica spp. Humanpopulation in the three parishes totals 779 inhabitants (7.7 inhab-itants km�2) working mostly in agriculture. The study area is partof the Montesinho Natural Park and of the Montesinho–NogueiraNatura 2000 site (PTCON0002). In terms of conservation values,the area is particularly relevant for large mammals namely the Ibe-rian wolf (Canis lupus signatus), the red deer (Cervus elaphus) andthe roe deer (Capreolus capreolus).
2.2. Sampling
We mapped holm oak woodlands in the study area by interpret-ing 2005 digital orthophotomaps in a GIS environment. We classi-fied all holm oak woodland patches in general classes based on treedensity, tree height and history of contact with fire derived from anoverlay analysis of GIS coverages of burned areas for 1990–2005and holm oak woodlands.
Among the most developed woodlands in the area, we selectedfive patches with previous contact with fire and adjacent to shrub-
38 J.C. Azevedo et al. / Forest Ecology and Management 297 (2013) 37–48

Author's personal copy
lands of different ages, i.e., burned at different times in the past(year of last fire: 1980, 2000, 2002, and 2005) (Table 1 andFig. 1). A total of 12 sampling transects were placed across the con-tact zones of woodlands and shrublands. Each 60 m transect wasplaced perpendicularly to the boundaries of the patches, 40 m in-side the woodland and 20 m outside (Fig. 2). On the basis of previ-ous work conducted in holm oak edges (Azevedo and Caçador,2000), a depth of 40 m was considered adequate to capture edgeeffects within these woodlands. In each transect we marked sam-pling points at distances of �20, �10, �5, �1, 0, 5, 10, 20 and40 m from the boundary. Negative distances are outside the wood-land and positive distances are inside (Fig. 2).
2.3. Vegetation structure
In each of the sampling points we measured tree density (#ha�1), diameter at breast height (DBH; cm), height (m), and canopycover (%); shrub cover (%), and height (m) by species; and littercomposition, cover (%), and thickness (mm).
To evaluate tree and shrub cover we used the line interceptmethod (Canfield, 1941) based on 10 m lines placed perpendicu-larly to the transects in each of the sampling points. We used thePoint-Centered Quarter Method (Mueller-Dombois and Ellenberg,1974) to evaluate tree density. Litter cover was classified using a4-class scheme: 0–25%, 26–50%, 51–75%, and 76–100%. Litterthickness in each sampling point was averaged from measure-
ments in 10 points placed along the 10 m line used to estimate cov-er. Field data was collected between May 29 and July 25 2007.
2.4. Plant diversity
We conducted inventories of herbaceous vascular plants at thespecies level following the sampling framework described above.We registered interceptions (cm) of all herbaceous plants in twoparallel 10 m lines at a distance of 0.5 m centered in each samplingpoint and positioned perpendicularly to the sampling transect.Two 10 m lines were preferred to one 20 m line to avoid samplingoverlay. This method was adjusted to capture information aboutrare plants by registering species that were within 10 cm of theinterception lines.
Abundance was calculated as the number of records of eachspecies in the 2, 10 m lines. Original data was transformed byassigning a score of 1 to rare species and adding 1 to the abundancevalues of the remaining species. For each sampling point, richnesswas calculated as the total number of plant species.
2.5. Fire modeling
We used fire modeling to analyze surface fire behavior along thegradient across the edge of the holm oak woodland by simulatingfire behavior variables in every sampling point along the transects.For each of the 120 sampling points we built a custom fuel model
Fig. 1. Location of the study area in Portugal and of the target patches in the study area.
Table 1Description of holm oak woodland patches sampled in the Braganca municipality, Portugal.
Patch Number of sampling transects Tree density Tree height Fire contact Years since fire contact
Labiados 2 Dense Medium/high Partial 7Petisqueira 4 Very dense Medium/high Partial 2S. Julião - Aldeia 2 Dense Medium/low Partial 5S. Julião - Moinho 2 Very dense Medium/high Partial 5Deilão 2 Very dense Medium/low Partial 27
Tree density and tree height were classified in a previous step to help defining the degree of development of woodland remnants. This was done following a generalclassification assessed from aerial photography and based on relative cover (for tree density) and apparent tree height.
J.C. Azevedo et al. / Forest Ecology and Management 297 (2013) 37–48 39

Author's personal copy
with the BehavePlus Fire Modeling System (Andrews, 2007) usingvegetation and litter data and following standard procedures(Fernandes, 2009). Fuel models are required to implement thesemi-physical fire spread model of Rothermel (1972) in BehavePlus.
Fire behavior simulations with BehavePlus for all the samplingpoints assumed extreme fire weather conditions with wind speed10 m aboveground, dead fuel moisture content and live fuel
moisture content set at 30 km h�1, 3% and 60%, respectively. Ateach sampling point, surface wind speed and dead fuel moisturecontent were adjusted for the effects of canopy structure and phys-iography (slope and aspect) using models by Finney (1998) andRothermel et al. (1986). All woodlands sampled were located inthe lower half of the slope and because fire invariably movesdownwards from upper areas in the study area, we simulated
Fig. 2. Sampling procedure: placement of transects (green) across edges in 4 of the 5 woodland patches considered (top) and schematic of the sampling transects and points(bottom). (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
40 J.C. Azevedo et al. / Forest Ecology and Management 297 (2013) 37–48

Author's personal copy
downslope fire behavior using wind, slope and transect direction.Simulated fire behavior in one location was independent of firebehavior in adjacent locations. Nonetheless, we assumed that var-iation in fire behavior along the exterior–interior gradient repre-sented the spatial variability of fire spreading from the outside(shrubland) in the direction of holm oak patches. This is not thesimulation of a spatial spread process but just the simulation of firebehavior in locations along fuel and environmental gradients.
To test the hypothesis that under less extreme conditions firebecomes extinct when reaching holm oak woodlands, we simu-lated fire intensity in BehavePlus for an average interior holmoak fuel model. We considered dead fuel moisture content levelscorresponding to the fuel moisture medians of five fire danger clas-ses (extreme – 6%, very high – 9%, high – 10%, moderate – 12%, low– 17%) for days with fires larger than 5 ha plus the more extrememoisture level (3%), and 10 m wind speeds of 5, 10, 20 and40 km h�1. In-stand surface wind speeds are 25% of these valuesas determined by characteristics of the average canopy (Finney,1998). For each fuel moisture-wind speed combination we ran 25simulations considering all combinations between fire spreaddirection and wind direction defined by multiples of 45�. When-ever fire intensity was lower than the extinction threshold mea-sured in experimental fires (Fernandes et al., 2000) we assumedit extinguished. Then, we averaged the outcome (n = 25) of eachmoisture-wind combination to calculate probabilities of fireextinction.
2.6. Statistical analyses
We fitted a normal errors mixed model to test for differences invegetation, fire behavior and plant diversity variables inside andoutside the woodland patches and among years since last fireevent. The transect was assigned a random factor and position inthe transect (In–Out = inside vs. outside), time since last fire (YrFire = 2, 6, or 27 yrs) and time since last fire nested within insidevs. outside (Yr Fire(In–Out)) were used as fixed factors. All depen-dent variables were log transformed.
We used Canonical Correspondence Analysis (CCA) to analyzeherbaceous plant patterning across the woodland boundary andtime since last fire event gradients. In this analysis only the firstaxis was constrained to a single explanatory variable; to do thiswe used CANOCO software (ter Braak and Šmilauer, 2002). The ef-fect of each of the environmental variables on species patterningwas tested with Montecarlo permutations. The species data matrixwas built using abundance data. The explanatory variables, usedindividually, were distance (established as distance from a hypo-thetical center of the patch, in meters) and time since last fireevent (Yr Fire), in years.
3. Results and discussion
3.1. General structure of woodlands and edges
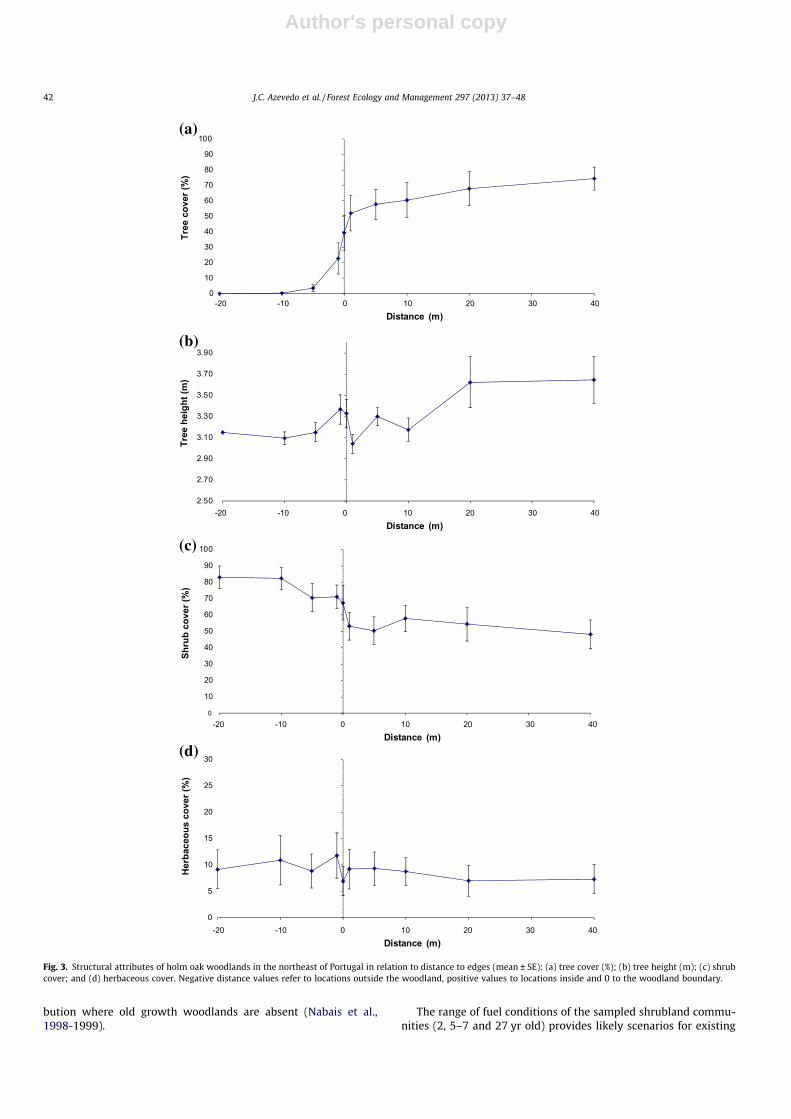
Woodlands sampled in this research were usually low, denseand with a relatively closed canopy (Fig. 3, Table S1). On average,canopy cover was zero 10 m outside and above 50% within (1–40 m) the woodland (max. 74%) (Fig. 3a). It increased from 3.6%to 56.7% within the 10 m transition zone across the boundary ofthe woodland. Outside differed significantly from inside(p = 0.000) (Table 2). Trees inside the woodland were, on average,slightly but significantly (p = 0.000) taller than those outside(3.19 m vs. 3.35 m) (Fig. 3b).
Average shrub cover was significantly (p = 0.000) higher outside(76.8%) than inside (52.8%) the woodland (Fig. 3c). There was anabrupt decrease within 1 m either side of the boundary. Herba-
ceous cover was slightly but significantly (p = 0.002) higher outside(10.2%) than inside (8.3%) and variability among sampling tran-sects was relatively high (Fig. 3d). Herbaceous plant cover de-creased sharply from �1 to 0 m. Given that the highest spatialvariability of all attributes considered was recorded near the 0 mmark, defining the woodland boundary by the location of the out-ermost tree stem in the patches seemed appropriate (Fig. 3).
The distance from the boundary in which a patch is affected byan adjacent area (or where edge effects are noticeable) is referredto as depth of edge penetration (Fraver, 1994), depth of edge influ-ence or edge width (Chen et al., 1992) or distance-of-edge influ-ence (LaCroix et al., 2008). Depending on the parameter underconsideration, this distance may range from 2 to 137 m (Fraver,1994). Based on the woodland characteristics we considered, edgewidth in holm oak woodlands is very narrow. Such abrupt (hard)edges are usually associated to other types of disturbances suchas clearcutting (Larrivée et al., 2008).
3.2. Fire behavior
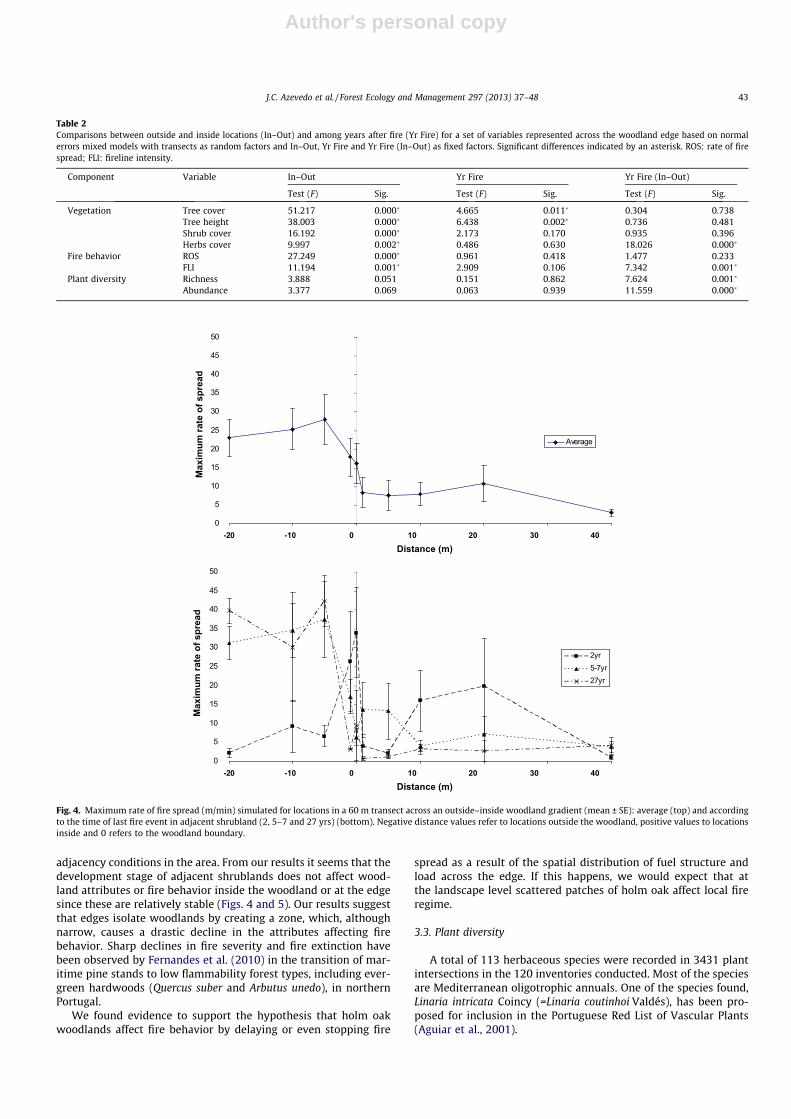
Simulated maximum rate of fire spread (ROS) and fireline inten-sity (FLI) were higher outside the woodland than inside (Figs. 4 and5, Table S1). The output variables showed abrupt transitions at theedge region of the woodland (–1 to +1 m). Differences betweenoutside and inside were statistically significant for both variables(p = 0.000 and p = 0.001, respectively) (Table 2). Based on firebehavior variables edge width was also very small, usually around1 m.
Fire behavior outside the woodland differed depending on timesince last fire occurrence (Figs. 4 and 5). For young (2 yr) and lessflammable adjacent shrubland areas, fire variables were similar in-side and outside the woodland, although they peaked at the wood-land boundary. For older shrublands (5–7 and 27 yr) fire intensityand spread rates outside were much higher than those inside thewoodland. Differences in ROS or FLI among years since last firewere not statistically significant but in the model containing YrFire nested within the inside–outside variable differences were sig-nificant for FLI (Table 2).
Inside the woodland, ROS ranged usually from 7.5 to10 m min�1 and FLI was often >2000 kW m�1, implying full canopyscorch and partial crown consumption (Byram, 1959) given thattrees in the woodlands were not very tall. However, FLI>4000 kW m�1, which generally corresponds to a fully developedcrown fire (Hirsch and Martell, 1996) was very rarely predictedto occur within the woodland. The simulations suggest that atmost study sites trees would be top-killed by fire (Catry et al.,2010).
Simulations above were performed for extreme, worst-caseweather conditions under which fire growth can be insensitive tochanges in LULC (Viedma et al., 2009) and to small scale disconti-nuities such as woodland edges. In less extreme conditions, firewould eventually be sensitive to this level of variation and becomeextinct when reaching the woodland. Additionally, while the sim-ulations indicated the maximum fire potential at the head of thefire, fire behavior in reality will vary significantly depending onwind direction in relation to slope. The results of the test of thefire-extinction hypothesis in holm oak woodlands (Fig. 6) indicatedthat when fuel moisture was >12% and 10 m wind speed was<40 km h�1, the probability of fire extinction is 100%. Even forthe driest scenario (moisture content of 3%) the probability ofextinction is near 50% when wind speed is very low (5 km h�1).Considering that woodlands occur in relatively sheltered topo-graphical positions and that moisture levels above 9% are frequent,the possibility of fire self-extinction in holm oak woodlands is real.On the other hand, it is also likely that woodlands in fact burnwhich is consistent with the current holm oak woodland age distri-
J.C. Azevedo et al. / Forest Ecology and Management 297 (2013) 37–48 41
Author's personal copy
bution where old growth woodlands are absent (Nabais et al.,1998-1999).
The range of fuel conditions of the sampled shrubland commu-nities (2, 5–7 and 27 yr old) provides likely scenarios for existing
0
10
20
30
40
50
60
70
80
90
100
-20 -10 0 10 20 30 40
Tree
cov
er (%
)
Distance (m)
-20 -10 0 10 20 30 40Distance (m)
-20 -10 0 10 20 30 40Distance (m)
-20 -10 0 10 20 30 40Distance (m)
(a)
(b)
(c)
(d)
2.50
2.70
2.90
3.10
3.30
3.50
3.70
3.90
Tree
hei
ght (
m)
0
10
20
30
40
50
60
70
80
90
100
Shru
b co
ver (
%)
0
5
10
15
20
25
30
Her
bace
ous
cove
r (%
)
Fig. 3. Structural attributes of holm oak woodlands in the northeast of Portugal in relation to distance to edges (mean ± SE): (a) tree cover (%); (b) tree height (m); (c) shrubcover; and (d) herbaceous cover. Negative distance values refer to locations outside the woodland, positive values to locations inside and 0 to the woodland boundary.
42 J.C. Azevedo et al. / Forest Ecology and Management 297 (2013) 37–48
Author's personal copy
adjacency conditions in the area. From our results it seems that thedevelopment stage of adjacent shrublands does not affect wood-land attributes or fire behavior inside the woodland or at the edgesince these are relatively stable (Figs. 4 and 5). Our results suggestthat edges isolate woodlands by creating a zone, which, althoughnarrow, causes a drastic decline in the attributes affecting firebehavior. Sharp declines in fire severity and fire extinction havebeen observed by Fernandes et al. (2010) in the transition of mar-itime pine stands to low flammability forest types, including ever-green hardwoods (Quercus suber and Arbutus unedo), in northernPortugal.
We found evidence to support the hypothesis that holm oakwoodlands affect fire behavior by delaying or even stopping fire
spread as a result of the spatial distribution of fuel structure andload across the edge. If this happens, we would expect that atthe landscape level scattered patches of holm oak affect local fireregime.
3.3. Plant diversity
A total of 113 herbaceous species were recorded in 3431 plantintersections in the 120 inventories conducted. Most of the speciesare Mediterranean oligotrophic annuals. One of the species found,Linaria intricata Coincy (=Linaria coutinhoi Valdés), has been pro-posed for inclusion in the Portuguese Red List of Vascular Plants(Aguiar et al., 2001).
Table 2Comparisons between outside and inside locations (In–Out) and among years after fire (Yr Fire) for a set of variables represented across the woodland edge based on normalerrors mixed models with transects as random factors and In–Out, Yr Fire and Yr Fire (In–Out) as fixed factors. Significant differences indicated by an asterisk. ROS: rate of firespread; FLI: fireline intensity.
Component Variable In–Out Yr Fire Yr Fire (In–Out)
Test (F) Sig. Test (F) Sig. Test (F) Sig.
Vegetation Tree cover 51.217 0.000� 4.665 0.011� 0.304 0.738Tree height 38.003 0.000� 6.438 0.002� 0.736 0.481Shrub cover 16.192 0.000� 2.173 0.170 0.935 0.396Herbs cover 9.997 0.002� 0.486 0.630 18.026 0.000�
Fire behavior ROS 27.249 0.000� 0.961 0.418 1.477 0.233FLI 11.194 0.001� 2.909 0.106 7.342 0.001�
Plant diversity Richness 3.888 0.051 0.151 0.862 7.624 0.001�
Abundance 3.377 0.069 0.063 0.939 11.559 0.000�
0
5
10
15
20
25
30
35
40
45
50
-20 -10 0 10 20 30 40Distance (m)
-20 -10 0 10 20 30 40Distance (m)
Max
imum
rate
of s
prea
d M
axim
um ra
te o
f spr
ead
Average
0
5
10
15
20
25
30
35
40
45
50
2yr5-7yr27yr
Fig. 4. Maximum rate of fire spread (m/min) simulated for locations in a 60 m transect across an outside–inside woodland gradient (mean ± SE): average (top) and accordingto the time of last fire event in adjacent shrubland (2, 5–7 and 27 yrs) (bottom). Negative distance values refer to locations outside the woodland, positive values to locationsinside and 0 refers to the woodland boundary.
J.C. Azevedo et al. / Forest Ecology and Management 297 (2013) 37–48 43

Author's personal copy
There were differences, although not significant (p = 0.051), innumber of species recorded inside and outside the woodland(Fig. 7; Tables S1 and 2). Richness peaked just outside the wood-land boundary (�1 m) and was at a minimum just inside theboundary (1 m). Inside the woodland, richness was relatively con-stant (approximately nine species, on average). Abundance, mea-sured as the total number of interceptions recorded along thetransects, was higher outside than inside the woodland. However,this difference was not statistically significant (p = 0.069; Table 2).
Furthermore, there was a 2 m-wide transition zone (�1 m to 1 m)where variability was high (Fig. 7).
When results were discriminated based on time since last fire inthe shrubland adjacent to the woodland, differences between in-side and outside were higher. Outside the woodland, richnesswas higher when shrubland was younger (2 yr) than when itburned 5–7 or 27 yrs before (Fig. 7). The oldest shrublands(27 yr) had the lowest richness values and also the highest contrastbetween inside and outside. Differences in richness among Yr Fireclasses were statistically significant (p = 0.001) only when timesince last fire was nested within the In–Out variable (Table 2).For the same conditions, changes were observed mainly within2 m (�1 m to 1 m) of the woodland boundary. A peak was regis-tered at the �1 m distance in the three curves although more pro-nounced for the 2 yr series. Abundance in the 2 yr series outsidethe woodland was higher than that at 5–7 or 27 yrs (Fig. 7;Table S1). In the 2 yr series, abundance was much higher outsidethan inside the woodland whereas in the remaining series it washigher inside. Differences in abundance among time since last firewere statistically significant only when Yr Fire was nested withinIn–Out (Table 2).
Plant richness and abundance are expected to be higher inedges than within woodlands (Fraver, 1994; Gehlhausen et al.,2000). Although observed in Pyrenean oak woodlands in this re-gion (Rocha et al., 2006), this pattern was not observed in thisstudy. In holm oak woodlands, richness and abundance peaked justoutside (�1 m) the woodland boundary.
0
5000
10000
15000
20000
25000
-20 -10 0 10 20 30 40
Distance (m)
-20 -10 0 10 20 30 40
Distance (m)
Fire
line
Inte
nsity
(kW
/m)
Fire
line
Inte
nsity
(kW
/m)
Average
0
5000
10000
15000
20000
25000
2yr5-7yr27yr
Fig. 5. Fireline intensity (kW/min) simulated for locations in a 60 m transect across an outside–inside woodland gradient (mean ± SE): average (top) and according to thetime of last fire event in adjacent shrubland (2, 5–7 and 27 yrs) (bottom). Negative distance values refer to locations outside the woodland, positive values to locations insideand 0 refers to the woodland boundary.
36
910
1217
510
2040
0
0.2
0.4
0.6
0.8
1
Probability of Fire Extinction
Fuel Moisture Content (%)Windspeed (km/hr)
0.8-10.6-0.80.4-0.60.2-0.40-0.2
Fig. 6. Fire extinction probability as a function of 10-m wind speed and dead fuelmoisture content. Results based on 25 simulations per wind speed/moisturecontent combination and fire extinction thresholds for fire in the region.
44 J.C. Azevedo et al. / Forest Ecology and Management 297 (2013) 37–48

Author's personal copy
Canonical analyses indicated significant patterning in the distri-bution of plant species along the interior to exterior and the timesince last fire gradients (p = 0.002 for both cases). One analysiswas able to discriminate among interior (right side of biplot inFig. 8 top), edge (center of the biplot), and heliophillous oligro-trophic annual species (left of the biplot). Edge species are mostlyeutrophic, some of which are perennial grasses (e.g., Dactylis lusita-nica and Sedum forsterianum), suggesting the existence of higherfertility at the woodland edge. The interior of the woodland isthe habitat of annual shade-tolerant nitrophilous species such asGeranium purpureum. Differences were also found in plant speciesdistribution based on time since last fire (Fig. 8 bottom). Annualoligotrophic species, among them the rare Linaria intricata, wereassociated with open habitats (recently burned). Older shrublandsappeared more suitable to perennial species such as S. forsterianumand Conopodium marizianum. A group of plant species associatedwith intermediate years after fire was also evident (Fig. 8 bottom,center of biplot).
Thus, we found evidence that plant diversity is favored by fre-quent fire outside and at the edge of holm oak woodlands. We ob-served higher richness and abundance outside the woodland butalso in adjacent recently burned shrublands. The barrier effect ofholm oak woodlands discussed above (Section 3.2) is responsiblefor these patterns through the establishment and maintenance ofhabitat diversity (open/edge/interior) along the fire-created gradi-ent, to which assemblages of plants species respond. As a result ofthis process, fire creates fine-scale landscape heterogeneity favor-ing plant diversity (richness and abundance) at both local andlandscape scales. This pattern is, however, compatible only withhigh frequency of fire occurrence but low intensity fire regimes.
3.4. A conceptual model of the dynamics of fire, landscape structure,and plant diversity
Based on the results of this research, and other empirical evi-dence collected over the years in the region or reported in the lit-erature, we propose a conceptual model for the dynamics of fireand community, landscape and plant diversity. This is a hypothesisto be tested in future research at multiple scales. The model in-cludes the following components/processes:
– Control of fire spread by holm oak woodlands . Fire is controlledat the narrow boundary between holm oak woodlands andadjacent shrublands where changes in vegetation structurecause abrupt decreases in fire spread rate and intensity.Under non-extreme weather conditions, fire self-extinguishesat the woodland edge. This hypothesis is supported by ourresearch.– Local-scale dynamics:� Maintenance of shrubland adjacent to woodland in the ini-
tial development stages by relatively frequent fire events.Control of fire spread potentially affects fire regime,favoring frequent fires that keep shrublands adjacent towoodlands in the initial development stage. This hypoth-esis has not been tested. Usually, fire recurrence in thenortheastern of Portugal is strongly dependent on shrub-land age and the median fire-return interval is 19 yrs(Fernandes et al., 2012). This estimate, however, is basedon summer fire statistics, which do not include theregion’s frequent winter fires and do not account forproximity to holm oak woodlands.
0
2
4
6
8
10
12
14
16
18
-20 -10 0 10 20 30 40
Ric
hnes
s (n
umbe
r of s
peci
es)
Distance (m)
2yr
5-7yr
27yr
Average
0
20
40
60
80
100
120
-20 -10 0 10 20 30 40
Abu
ndan
ce (n
umbe
r)
Distance (m)
2yr
5-7yr
27yr
Average
Fig. 7. Herbaceous plant richness (top) and abundance (bottom) for locations in a 60 m transect across an outside–inside woodland gradient (mean ± SE). Average (n = 12)and data discriminated according to age of adjacent shrub communities: 2 yr- burned 2 yrs before (n = 4); 5–7 yr- burned 5–7 yrs before (n = 6); 27 yr- burned 27 yrs before(n = 2).
J.C. Azevedo et al. / Forest Ecology and Management 297 (2013) 37–48 45

Author's personal copy
� Promotion of plant diversity. Maintenance of herbaceousrichness, including edge, open area and interior species,across the boundary between woodlands and shrublandsunder the high-frequency/low-severity fire regime, thuscontributing to conservation of plant diversity. Thishypothesis is supported by our study.
– Landscape-scale dynamics:� Control of fire size by means of fire growth control by holm
oak woodlands. The barrier effect of holm oak woodlands,reinforced by their topographical location, decreases firesize thus increasing patchiness in the landscape. Togetherwith the variability in size, shape and configuration of
Fig. 8. CCA biplots showing the distribution of the species with the highest fit in relation to a single explanatory variable: distance (top) and years after fire (bottom). In thetop biplot, interior plant species are shown on the right side, heliophillous oligrotrophic annual species on the left side, and edge species in the center. In the bottom biplot,plant species in recently burned shrublands are shown on the left side whereas plants in undisturbed shrublands are showed on the right side. Ab = Anarrhinum bellidifolium,As = Allium sphaerocephalum, Al = Anthyllis lusitanica, Aq = Asplenium quadrivalens, Cs = Cerastium semidecandrum, Cm = Conopodium marizianum, Cf = Corynephorus fasciculatus,Ca = Crucianella angustifolia, Dl = Dactylis lusitanica, Dd = Daucus durieua, Fp = Filago pyramidata, Gp = Geranium purpureum, Hh = Hispidella hispânica, Lc = Lactuca chondrilliflora,Ll = Leontodon longirostris, Li = Linaria intricate, Mp = Micropyrum patens, Oc = Ornithopus compressus, Sf = Sedum forsterianum, Sj = Senecio jacobaea, Ta = Trifolium arvense,Tc = Trifolium campestre, Vs = Vicia sativa.
46 J.C. Azevedo et al. / Forest Ecology and Management 297 (2013) 37–48

Author's personal copy
vegetation patches created by fire, this process increaseslandscape heterogeneity. This component is inferred fromgeneral landscape ecology and fire theory based onempirical evidence from the study area. This hypothesisis supported by simulation research (LaCroix et al.,2008; Ryu et al., 2007).
� Promotion of plant diversity. Maintenance of plant rich-ness and abundance at the landscape scale by means ofthe landscape heterogeneity above.
– Disturbance of mature holm oak woodlands by infrequent high-intensity fire events. Infrequent high-intensity fires disturbmature holm oak woodlands. The frequency of these fires isunknown but they probably have a periodicity of 40–50 yrs,which is the maximum estimated age of woodlands in the studyarea (Nabais et al., 1998–1999). Given its vulnerability torepeated disturbances, higher frequency of such extreme fireevents would eventually remove holm oak from the landscape(Bonfil et al., 2004). Fire is followed by strong regenerationand recovery of the woodland (Broncano et al., 2005). Holmoak grows faster and taller (causing quicker canopy closure)at lower than upper slope positions (Gracia and Retana, 2004),which can in part explain the presence of woodlands at the bot-tom of slopes. This hypothesis is supported by the current agedistribution of holm oak woodlands in the region (Nabaiset al., 1998–1999) and the structure (height, density, and cover)of current woodlands surveyed in this research.
3.5. Implications for management and restoration
Our results provide information on the structure and function-ing of communities and landscapes where holm oak is distributed,and can contribute to the understanding of the dynamics of thesesystems, which may be useful for planning and management at dif-ferent scales. The fact that holm oak woodlands can act as fire bar-riers offers promising possibilities for landscape planning andmanagement. Wildfires in Europe, including Portugal and thestudy region, are a major social, political and economic issue andtheir prevention and suppression have not proven sufficientlyeffective to mitigate their frequency and severity over the years(Birot, 2009). As suggested by previous research (Fernandes et al.,2010; González et al., 2006; Moreira et al., 2009) less fire-pronecover types are a rational option for fuel-breaks. Considering theresults of this study, holm oak woodlands in this part of the coun-try and elsewhere in the Mediterranean, seem to be a strong can-didate for such approaches. The expansion of holm oakwoodlands, both artificially by planting and seedling, or by pro-moting the existing natural regeneration, could contribute signifi-cantly to this purpose. Furthermore, appropriate design of currentand future woodlands could contribute to fire hazard reduction atthe landscape level. Currently, holm oak woodlands represent lessthan 3% of the Montesinho/Nogueira Natura 2000 Site(PTCON0002) where the study area is located.
Another potential application of our findings is in plant conser-vation. The fact that holm oak woodlands promote abundance andrichness of herbaceous species, including species with conserva-tion value, is of extreme relevance in terms of conservation plan-ning. Although relatively poor, these woodlands are able tocontribute to the conservation of interior species but also edgeand open system species through their role in fire regime. Theirmajor contribution, however, should be in terms of enhancementand maintenance of landscape heterogeneity, which favors plantdiversity at the landscape scale. This is particularly importantwhere sources of disturbance (traditional agriculture, grazing andpastoral burning), responsible for the high-frequency/low-inten-sity disturbance regimes in the past, are currently absent. This
has driven landscapes to structural homogeneity favoring theoccurrence of high intensity wildfires (Azevedo et al., 2011; Loepfeet al., 2010). Fire is therefore the only viable process of maintaininglandscape heterogeneity and species diversity in the region.
Additional research is, however, still required not only to testthe proposed conceptual model but also to improve the under-standing of the dynamics of these communities and landscapesand their relationship with fire regimes. Furthermore, in terms oflandscape planning and management, additional information is re-quired to better address the use of holm oak woodlands in pro-grams of conservation and landscape fire hazard mitigation,including optimal patch size, shape, and orientation and optimalproportion for holm oak woodlands and configuration in thelandscape.
4. Conclusions
Holm oak woodlands have a role in the control of fire spreadand the maintenance of plant diversity in landscapes where theseforest patches persist. Differences in simulated maximum rate offire spread (ROS) and fireline intensity (FLI) between the interiorand exterior of these woodlands were statistically significant. In-side the woodland, fire was of lower intensity and spread moreslowly than it did outside. Outside the woodland, fire behavior,particularly FLI, was affected by time since last fire event. Patternsof simulated fire behavior across edges of holm oak woodlandssupport the hypothesis of self-extinction of fire at these transitionzones. The probability of fire extinction is 100% when fuel moistureis >12% and 10 m aboveground wind speed is <40 km h�1 and closeto 50% when moisture content is 3% as long as wind speed remainsvery low (5 km h�1). We conclude, therefore, that holm oak wood-lands affect fire behavior by interrupting fire spread at their edges.Number of herbaceous plant species and their abundance werelower inside than outside the woodlands but differences werenot statistically significant. Additionally, there was significant pat-terning of plant composition along the interior–exterior gradient.Time since last fire event significantly affected the richness, abun-dance and distribution of individual species. We conclude that fire-created edges affect herbaceous species richness, abundance anddistribution. Under conditions of frequent low intensity fires, plantdiversity is favored by maintaining habitat diversity, which in turnincreases plant diversity at the patch and landscape levels. The re-sults of this study support in part a conceptual model of thedynamics of fire, landscape structure, and plant diversity in fire-prone landscapes.
Acknowledgments
The authors would like to thank Parque Natural de Montesinhoand Câmara Municipal de Bragança for provision of geographicalinformation of the study area, Daniel Saraiva Morais for help inthe field data collection, Miguel Vaz Pinto for assistance with thegraphics, and M. Alice Pinto and two anonymous reviewers fortheir helpful and constructive comments and suggestions on themanuscript. Special thanks go to Manuel Rainha who first drewour attention to the fire barrier effect of holm oak woodlandsand challenged us to explain it. This study was funded by the ‘‘Fun-do Florestal Permanente’’ (Project 2006.09.001200.3), PortugueseGovernment.
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.foreco.2013.02.007.
J.C. Azevedo et al. / Forest Ecology and Management 297 (2013) 37–48 47

Author's personal copy
References
Agee, J.K., 1998. The landscape ecology of western forest fire regimes. NorthwestSci. 72, 24–34.
Aguiar, C., Honrado, J.J., Alves P., Caldas, F.B., Nepomuceno, H., Jansen, J., Sequeira,M., 2001. Terceira aproximação à lista da flora rara e a proteger no Norte dePortugal Continental: II Gimnospérmicas e Angiospérmicas. 2� CongressoNacional de Conservação da Natureza. Lisboa.
Alexander, J.D., Seavy, N.E., Ralph, C.J., Hogoboom, B., 2006. Vegetation andtopographical correlates of fire severity from two fires in the Klamath–Siskiyou region of Oregon and California. Int. J. Wildland Fire 15, 237–245.
Allen, H.D., 2008. Fire: plant functional types and patch mosaic burning in fire-prone ecosystems. Prog. Phys. Geogr. 32, 421–437.
Andrews, P.L., 2007. BehavePlus fire modeling system: past, present, and future. In:Proceedings of the Seventh Symposium on Fire and Forest MeteorologicalSociety. American Meteorological Society, Boston, pp. 13.
Azevedo, J., Caçador, F., 2000. Bordaduras de bosques de Quercus rotundifolia Lam.no Parque Natural de Montesinho. Quercetea 1, 126–137.
Azevedo, J.C., Moreira, C., Castro, J.P., Loureiro, C., 2011. Agriculture abandonment,land-use change and fire hazard in mountain landscapes in NortheasternPortugal. In: Li, C., Lafortezza, R., Chen, J. (Eds.), Landscape Ecology in ForestManagement and Conservation: Challenges and Solutions for Global Change.HEP-Springer, Beijing, pp. 329–351.
Biddulph, J., Kellman, M., 1998. Fuels and fire at savanna gallery forest boundaries insoutheastern Venezuela. J. Trop. Ecol. 14, 445–461.
Bielsa, I., Xavier, P., Bunce, B., 2005. Agricultural abandonment in the North EasternIberian Peninsula: the use of basic landscape metrics to support planning. J.Environ. Plann. Man. 48, 85–102.
Birot, Y. (Ed.), 2009. Living with Wildfires. What Science Can Tell Us. EuropeanForest Institute Discussion Paper 15. EFI, Joensuu, 86 p.
Bonfil, C., Cortes, P., Espelta, J.M., Retana, J., 2004. The role of disturbance in the co-existence of the evergreen Quercus ilex and the deciduous Quercus cerrioides. J.Veg. Sci. 15, 423–430.
Broncano, M.J., Retana, J., Rodrigo, A., 2005. Predicting the recovery of Pinushalepensis and Quercus ilex forests after a large wildfire in northeastern Spain.Plant Ecol. 180, 47–56.
Buhk, C., Meyn, A., Jentsch, A., 2007. The challenge of plant regeneration after fire inthe Mediterranean Basin: scientific gaps in our knowledge on plant strategiesand evolution of traits. Plant Ecol. 192, 1–19.
Byram, G.M., 1959. Combustion of forest fuels. In: Davis, K.P. (Ed.), Forest Fire:Control and Use. McGraw-Hill, New York, pp. 61–89.
Canfield, R.H., 1941. Application of the line interception method in sampling rangevegetation. J. Forest. 39, 388–394.
Catry, F., Rego, F., Moreira, F., Fernandes, P.M., Pausas, J., 2010. Post-fire tree mortalityin mixed forests of central Portugal. Forest Ecol. Manage. 260, 1184–1192.
Chen, J.Q., Franklin, J.F., Spies, T.A., 1992. Vegetation Responses to edgeenvironments in old-growth Douglas-fir forests. Ecol. Appl. 2, 387–396.
DeLong, S.C., Kessler, W.B., 2000. Ecological characteristics of mature forestremnants left by wildfire. Forest Ecol. Manage. 131, 93–106.
Eberhart, K.E., Woodard, P.M., 1987. Distribution of residual vegetation associatedwith large fires in Alberta. Can. J. Forest Res. 17, 1207–1212.
Fernandes, P.M., 2009. Combining forest structure data and fuel modelling toclassify fire hazard in Portugal. Ann. Forest Sci. 66, 415.
Fernandes, P.M., Catchpole, W.R., Rego, F.C., 2000. Shrubland fire behaviourmodelling with microplot data. Can. J. Forest Res. 30, 889–899.
Fernandes, P.M., Luz, A., Loureiro, C., 2010. Changes in wildfire severity frommaritime pine woodland to contiguous forest types in the mountains ofnorthwestern Portugal. Forest Ecol. Manage. 260, 883–892.
Fernandes, P.M., Loureiro, C., Magalhães, M., Ferreira, P., Fernandes, M., 2012. Fuelage, weather and burn probability in Portugal. Int. J. Wildland Fire 21, 380–384.
Finney, M., 1998. FARSITE: fire Area Simulator – model development andevaluation. USDA Forest Service, Ogden, 47 p.
Fraver, S., 1994. Vegetation responses along edge-to-interior gradients in the mixedhardwood forests of the Roanoke river basin, North-Carolina. Conserv. Biol. 8,822–832.
Gehlhausen, S.M., Schwartz, M.W., Augspurger, C.K., 2000. Vegetation andmicroclimatic edge effects in two mixed-mesophytic forest fragments. PlantEcol. 147, 21–35.
Gignac, L.D., Dale, M.R.T., 2007. Effects of size, shape, and edge on vegetation inremnants of the upland boreal mixed-wood forest in agro-environments ofAlberta, Canada. Can. J. Bot. -Rev. Can. Bot. 85, 273–284.
González, J., Palahi, M., Trasobares, A., Pukalla, T., 2006. A fire probability model forforest stands in Catalonia (north-east Spain). Ann. Forest Sci. 63, 169–176.
Gracia, M., Retana, J., 2004. Effect of site quality and shading on sprouting patternsof holm oak coppices. Forest Ecol. Manage. 188, 39–49.
Hanson, J.J., Stuart, J.D., 2005. Vegetation responses to natural and salvage loggedfire edges in Douglas-fir/hardwood forests. Forest Ecol. Manage. 214, 266–278.
Hirsch, K.G., Martell, D.L., 1996. A review of initial attack fire crew productivity andeffectiveness. Int. J. Wildland Fire 6, 199–215.
Kremsater, L., Bunnell, F.L., 1999. Edge effects: theory, evidence and implications tomanagement of western North American Forests. In: Rochelle, J.A., Lehmann,L.A., Wisniewski, J. (Eds.), Forest Fragmentation: Wildlife and ManagementImplications. Brill, Leiden, The Netherlands, pp. 117–153.
LaCroix, J.J., Li, Q.L., Chen, J.Q., Henderson, R., John, R., 2008. Edge effects on firespread in a disturbed Northern Wisconsin landscape. Landscape Ecol. 23, 1081–1092.
Larrivée, M., Drapeau, P., Fahrig, L., 2008. Edge effects created by wildfire and clear-cutting on boreal forest ground-dwelling spiders. Forest Ecol. Manage. 255,1434–1445.
Loepfe, L., Martinez-Vilalta, J., Oliveres, J., Pinol, J., Lloret, F., 2010. Feedbacksbetween fuel reduction and landscape homogenisation determine fire regimesin three Mediterranean areas. Forest Ecol. Manage. 259, 2366–2374.
Marchand, P., Houle, G., 2006. Spatial patterns of plant species richness along aforest edge: what are their determinants? Forest Ecol. Manage. 223, 113–124.
Moreira, F., Vaz, P., Catry, F., Silva, J., 2009. Regional variations in wildfiresusceptibility of land-cover types in Portugal: implications for landscapemanagement to minimize fire hazard. Int. J. Wildland Fire 18, 563–574.
Mueller-Dombois, D., Ellenberg, H., 1974. Aims and Methods of Vegetation Ecology.Wiley, New York, 547 p.
Nabais, C., Freitas, H., Hagemeyer, J., 1998-1999. Tree-rings to climate relationshipsof Quercus ilex L. in the NE-Portugal. Dendrochronologia 16-17, pp. 37–44.
Nunes, M.C.S., Vasconcelos, M.J., Pereira, J.M.C., Dasgupta, N., Alldredge, R.J., 2005.Land cover type and fire in Portugal: do fires burn land cover selectively?Landscape Ecol. 20, 661–673.
Pausas, J.G., 2006. Simulating Mediterranean landscape pattern and vegetationdynamics under different fire regimes. Plant Ecol. 187, 249–259.
Perera, A.H., Dalziel, B.D., Buse, L.J., Routledge, R.G., 2009. Spatial variability ofstand-scale residuals in Ontario’s boreal forest fires. Can. J. Forest Res. -Rev. Can.Rech. Forest 39, 945–961.
Rocha, S., Aguiar, C., Azevedo, J.C., 2006. Ecologia da vegetação de orla de bosque deCarvalho-negral na Serra da Nogueira, Bragança, Portugal. VI Encontro ALFA deFitossociologia. ALFA, Angra do Heroísmo, Portugal, pp. 79.
Roman-Cuesta, R.M., Gracia, M., Retana, J., 2009. Factors influencing the formationof unburned forest islands within the perimeter of a large forest fire. Forest Ecol.Manage. 258, 71–80.
Ross, K.A., Fox, B.J., Fox, M.D., 2002. Changes to plant species richness in forestfragments: fragment age, disturbance and fire history may be as important asarea. J. Biogeogr. 29, 749–765.
Rothermel, R.C., 1972. A mathematical model for predicting fire spread in wildlandfuels. USDA Forest Service, Ogden, 40 p.
Rothermel, R.C., Wilson, R.A., Morris, G.A., Sackett, S.S., 1986. Modeling moisturecontent of fine dead wildland fuels. USDA Forest Service, Ogden, 61 p.
Ryu, S.-R., Chen, J., Zheng, D., Lacroix, J.J., 2007. Relating surface fire spread tolandscape structure: an application of FARSITE in a managed forest landscape.Landscape Urban Plann. 83, 275–283.
Slik, J.W.F., van Beek, M., Bernard, C., Bongers, F., Breman, F.C., Cannon, C.H.,Sidiyasa, K., 2011. Limited edge effects along a burned–unburned borneanforest boundary seven years after disturbance. Biotropica 43, 288–298.
Ter Braak, C.J.F., Šmilauer, P., 2002. CANOCO Reference Manual and CanoDraw forWindows User’s Guide: Software for Canonical Community Ordination (version4.5). Microcomputer Power, Ithaca NY, USA.
Turner, M.G., 1989. Landscape ecology – the effect of pattern on process. Annu. Rev.Ecol. Syst. 20, 171–197.
Viedma, O., Angeler, D.G., Moreno, J.M., 2009. Landscape structural features controlfire size in a Mediterranean forested area of central Spain. Int. J. Wildland Fire18, 575–583.
Viedma, O., Moreno, J.M., Rieiro, I., 2006. Interactions between land use/land coverchange, forest fires and landscape structure in Sierra de Gredos (Central Spain).Environ. Conserv. 33, 212–222.
48 J.C. Azevedo et al. / Forest Ecology and Management 297 (2013) 37–48




















