
The Role of HIV-Related Chemokine Receptors and Chemokines in Human Erythropoiesis in Vitro
11
The Role of HIV-Related Chemokine Receptors and Chemokines in Human Erythropoiesis in Vitro MARCIN MAJKA, a JANINA RATAJCZAK, a BENHUR LEE, b MAREK HONCZARENKO, b RAY DOUGLAS, b M. ANNA KOWALSKA, c LESLIE SILBERSTEIN, b ALAN M. GEWIRTZ, a MARIUSZ Z. RATAJCZAK a a Department of Pathology & Laboratory Medicine; b Department of Internal Medicine, Division of Hematology/Oncology, University of Pennsylvania School of Medicine; c Department of Pediatrics, Children’s Hospital of Philadelphia, Philadelphia, Pennsylvania, USA Key Words. Hematopoiesis · Chemokines · Chemokine receptors · CXCR4 · CCR5 · Stromal derived factor-1 (SDF-1) · HIV ABSTRACT In order to better define the role of HIV-related chemokines in human erythropoiesis we studied: A) the expression of chemokine receptors, both on human CD34 + cells which include erythroid progenitors and on more mature erythroid cells; B) the functionality of these recep- tors by calcium flux, chemotaxis assay and phosphorylation of mitogen-activated protein kinases (MAPK) p42/44 (ERK1/ERK2) and AKT, and finally C) the influence of chemokines on BFU-E formation. We found that HIV- related chemokine receptor CXCR4, but not CCR5, is detectable on human CD34 + BFU-E cells. CXCR4 surface expression decreased during erythroid maturation, although CXCR4 mRNA was still present in cells isolated from differentiated erythroid colonies. SDF-1, a CXCR4 ligand, induced calcium flux and phosphorylation of MAPK (p42/44) and AKT in CD34 + KIT + bone marrow mononuclear cells which contain BFU-E, as well as chemo- tactic activity of both human CD34 + BFU-E progenitors and erythroid cells isolated from day 2-6 BFU-E colonies. Responsiveness to SDF-1 decreased when the cells differen- tiated to the point of surface expression of the erythroid- specific marker Glycophorin-A. In contrast, the CCR5 ligands (macrophage inflammatory protein-1α [MIP-1α], MIP-1β, and RANTES) did not activate calcium flux, MAPK and AKT phosphorylation or chemotaxis of CD34 + KIT + cells or cells isolated from the BFU-E colonies. Interestingly, none of the chemokines tested in this study had any effect on BFU-E colony formation. In conclusion, only CXCR4 is functional, and its specific ligand SDF-1 may therefore play an important role in the homing and/or retention of early erythroid precursors in the bone marrow environment. Stem Cells 2000;18:128-138 STEM CELLS 2000;18:128-138 www.StemCells.com Correspondence: Mariusz Z. Ratajczak, M.D., Ph.D., Department of Pathology and Laboratory Medicine, University of Pennsylvania, 405A Stellar Chance Labs, 422 Curie Boulevard, Philadelphia, Pennsylvania 19104, USA. e-mail: [email protected] Accepted for publication February 10, 2000. ©AlphaMed Press 1066-5099/00/$5.00/0 INTRODUCTION Patients infected with HIV-1 frequently exhibit a vari- ety of hematological abnormalities, among the most promi- nent is anemia [1-6]. Anemia is very common in HIV-infected individuals, being diagnosed in approxi- mately 10%-20% at initial presentation, and in 70%-80% of patients at some time during the course of their disease [3]. Anemia is likely multifactorial in origin and might result from: A) direct infection of erythroid progenitors by HIV; B) the inhibitory influence of HIV-related proteins, and C) the negative influence of the various inflammatory cytokines and chemokines elaborated in response to the infection itself [2, 3, 5]. The discovery that some chemokine receptors are core- ceptors for HIV entry into cells offers new avenues for increasing our understanding of the mechanisms underlying HIV-1-associated marrow dysfunction [1, 4]. All HIV-1 strains studied to date use CCR5 (R5 strains), CXCR4 (X4) or both receptors (R5X4) to enter cells [1, 4]. It has been found that the CCR5 receptor binds macrophage inflammatory pro- tein-1α (MIP-1α), MIP-1β, and RANTES, whereas the CXCR4 receptor binds stromal cell-derived factor-1 (SDF-1). We previously reported that human erythroid progenitors and erythroblasts cannot be infected by HIV-1 envelope pseudotyped XR and R5 viruses and suggested that other indi- rect mechanisms are responsible for the pathogenesis of HIV-
Transcript of The Role of HIV-Related Chemokine Receptors and Chemokines in Human Erythropoiesis in Vitro

The Role of HIV-Related Chemokine Receptors and Chemokines in Human Erythropoiesis in Vitro
MARCIN MAJKA ,a JANINA RATAJCZAK ,a BENHUR LEE,b MAREK HONCZARENKO ,b RAY DOUGLAS,b
M. ANNA KOWALSKA ,c LESLIE SILBERSTEIN ,b ALAN M. GEWIRTZ ,a MARIUSZ Z. RATAJCZAK a
aDepartment of Pathology & Laboratory Medicine; bDepartment of Internal Medicine, Division of
Hematology/Oncology, University of Pennsylvania School of Medicine; cDepartment of Pediatrics,
Children’s Hospital of Philadelphia, Philadelphia, Pennsylvania, USA
Key Words. Hematopoiesis · Chemokines · Chemokine receptors · CXCR4 · CCR5 · Stromal derived factor-1 (SDF-1) · HIV
ABSTRACT
In order to better define the role of HIV-relatedchemokines in human erythropoiesis we studied: A) theexpression of chemokine receptors, both on human CD34+
cells which include erythroid progenitors and on moremature erythroid cells; B) the functionality of these recep-tors by calcium flux, chemotaxis assay and phosphorylationof mitogen-activated protein kinases (MAPK) p42/44(ERK1/ERK2) and AKT, and finally C) the influence ofchemokines on BFU-E formation. We found that HIV-related chemokine receptor CXCR4, but not CCR5, isdetectable on human CD34+ BFU-E cells. CXCR4 surfaceexpression decreased during erythroid maturation,although CXCR4 mRNA was still present in cells isolatedfrom differentiated erythroid colonies. SDF-1, a CXCR4ligand, induced calcium flux and phosphorylation ofMAPK (p42/44) and AKT in CD34+KIT + bone marrow
mononuclear cells which contain BFU-E, as well as chemo-tactic activity of both human CD34+ BFU-E progenitorsand erythroid cells isolated from day 2-6 BFU-E colonies.Responsiveness to SDF-1 decreased when the cells differen-tiated to the point of surface expression of the erythroid-specific marker Glycophorin-A. In contrast, the CCR5ligands (macrophage inflammatory protein-1α [MIP-1 α],MIP-1β, and RANTES) did not activate calcium flux,MAPK and AKT phosphorylation or chemotaxis ofCD34+KIT + cells or cells isolated from the BFU-E colonies.Interestingly, none of the chemokines tested in this studyhad any effect on BFU-E colony formation. In conclusion,only CXCR4 is functional, and its specific ligand SDF-1may therefore play an important role in the homing and/orretention of early erythroid precursors in the bone marrowenvironment. Stem Cells 2000;18:128-138
STEM CELLS 2000;18:128-138 www.StemCells.com
Correspondence: Mariusz Z. Ratajczak, M.D., Ph.D., Department of Pathology and Laboratory Medicine, University ofPennsylvania, 405A Stellar Chance Labs, 422 Curie Boulevard, Philadelphia, Pennsylvania 19104, USA. e-mail:[email protected] Accepted for publication February 10, 2000. ©AlphaMed Press 1066-5099/00/$5.00/0
INTRODUCTION
Patients infected with HIV-1 frequently exhibit a vari-ety of hematological abnormalities, among the most promi-nent is anemia [1-6]. Anemia is very common inHIV-infected individuals, being diagnosed in approxi-mately 10%-20% at initial presentation, and in 70%-80% ofpatients at some time during the course of their disease [3].Anemia is likely multifactorial in origin and might resultfrom: A) direct infection of erythroid progenitors by HIV;B) the inhibitory influence of HIV-related proteins, and C)the negative influence of the various inflammatorycytokines and chemokines elaborated in response to theinfection itself [2, 3, 5].
The discovery that some chemokine receptors are core-ceptors for HIV entry into cells offers new avenues forincreasing our understanding of the mechanisms underlyingHIV-1-associated marrow dysfunction [1, 4]. All HIV-1strains studied to date use CCR5 (R5 strains), CXCR4 (X4) orboth receptors (R5X4) to enter cells [1, 4]. It has been foundthat the CCR5 receptor binds macrophage inflammatory pro-tein-1α (MIP-1α), MIP-1β, and RANTES, whereas theCXCR4 receptor binds stromal cell-derived factor-1 (SDF-1).
We previously reported that human erythroid progenitorsand erythroblasts cannot be infected by HIV-1 envelopepseudotyped XR and R5 viruses and suggested that other indi-rect mechanisms are responsible for the pathogenesis of HIV-

related anemia [7]. Since many different chemokines are elab-orated in the body during HIV-1 infection [1, 2, 6] and someof these chemokines have been reported to inhibit erythro-poiesis [8-10], we became interested in their potential role inthe pathogenesis of HIV-related anemia. We approached thisquestion from different points of view by: A) looking at theexpression of chemokine receptors both on human CD34+
cells which include erythroid progenitors and on moremature erythroid cells; B) evaluating the functionality ofthe chemokine receptors in chemotaxis, mitogen-activatedprotein kinase (MAPK) p42/44 and AKT phosphorylationand calcium flux studies, and finally by C) studying theeffect of selected chemokines on in vitro proliferation ofhuman BFU-E progenitors.
We found that from all the chemokines tested, only SDF-1 induced calcium flux, MAPK p42/44, and AKT phosphory-lation and chemotaxis in early human erythroid precursorcells. Therefore, as determined by the biological assaysemployed in this study, only CXCR4 appeared to be a func-tional chemokine receptor in cells of this lineage and SDF-1may play an important role in the retention of early erythroidprecursors in the bone marrow (BM) environment [11-17].Finally, we did not observe any influence of chemokines onproliferation of human erythroid progenitor cells.
MATERIALS AND METHODS
CellsLight-density marrow mononuclear cells (MNC) were
obtained from 15 consenting healthy donors and depleted ofadherent cells and T lymphocytes (A–T– MNC) as described[18-21]. A–T– MNC were enriched for CD34+ cells byimmunoaffinity selection with the murine monoclonal anti-body (mAb) HPCA-1 (Becton-Dickinson; San Jose, CA) andmagnetic beads according to the manufacturer’s protocol(Dynal; Oslo, Norway).
Selection of CD34+KIT + A–T– MNC by Fluorescence-ActivatedCell Sorting (FACS)
The c-kit receptor positive (c-kit-R+) subset of CD34+
cells was isolated by FACS. Briefly, 2× 107 human A–T–
MNC were suspended in phosphate-buffered saline (PBS),supplemented with 5% bovine calf serum (BCS), andlabeled for 30 min at 4°C with anti-Kit R mAb (clone#104D2) directly conjugated with Cy5 (Becton Dickinson;20 µl/106 cells) and with an anti-CD34 mAb directly conju-gated with phycoerythrin ([PE] anti-HPCA-2, Becton-Dickinson; 20 µl/106 cells). After the final incubation, cellswere washed 3× in ice-cold PBS supplemented with 5%BCS, and then subjected to FACS using a FACS Star PlusII (Becton Dickinson).
Ex Vivo Expansion of Human Erythroid CellsCD34+ cells were expanded in a serum-free liquid sys-
tem as described [19]. Briefly, CD34+ A–T– MNC wereresuspended in Iscove’s Dulbecco’s modified Eagle’smedium (104/ml; GIBCO BRL; Grand Island, NJ) supple-mented with 25% of artificial serum containing 1% delipi-dated, deionized, and charcoal-treated bovine serum albumin(BSA), 270 µg/ml iron-saturated transferrin, insulin (20µg/ml), and 2 mmol/l L-glutamine (all from Sigma; St. Louis,MO). BFU-E growth was stimulated with recombinanthuman (rHu) erythropoietin (EPO) (2 U/ml), and rHu kitligand (KL) (10 ng/ml). Cytokines were from R&DSystems, Inc., Minneapolis, MN. Cultures were incubatedat 37°C in a fully humidified atmosphere supplementedwith 5% CO2. Under these conditions, approximately 100%of expanded cells were glycophorin A (GPA-A) positive,CD33, and gpIIa/IIIb-negative after 14 days [19]. In our stud-ies we employed erythroid cells isolated from these cultureswhich were expanded from 2 to 14 days.
Cell CulturesBriefly, CD34+ A–T– MNC cells (1 × 104) were cloned
in 1 ml of Iscove’s modified Dulbecco’s medium (IMDM)(GIBCO BRL) containing 0.8% methylcellulose (MethocelMC; Fluka, Switzerland) and supplemented with 1% delipi-dated, deionized, charcoal-absorbed BSA (Sigma), 270µg/ml iron-saturated transferrin (Sigma), 5.6 µg/ml choles-terol (Sigma), and 2 mmol/l L-glutamine as described [18].rHu growth factors appropriate for growth of the colonieswere added to the mixture which was then transferred to 3.5-cm plastic petri dishes and incubated (37°C, 95% air, 5%CO2 humidified atmosphere) for the appropriate times.Growth factors employed and utilized concentrations wereas follows: EPO (5 U/ml) + KL (100 ng/ml) for optimal stim-ulation and EPO (1 U/ml) + KL (40 ng/ml) for suboptimalstimulation of BFU-E.
The various chemokines (SDF-1, MIP-1α, MIP-1β,RANTES, MIP-3α, interleukin (IL)-8, Gro-α, Gro-β, Gro-γ, NAP-2, ENA-78) in a dose of 100 ng/ml (R&D SystemsInc. and PeproTech Inc.; Rocky Hill, NY) were added ascostimulators to some culture dishes. In all the experimentsBFU-E colonies were counted with the aid of an invertedmicroscope on day 14.
FACS Analysis of Chemokine Receptor ExpressionThe expression of CCR1, CCR2, CCR3, CCR4, CCR5,
CCR6, CCR8, CXCR2, CXCR3, and CXCR4 on expanded exvivo erythroid cells from CD34+ BMMC under serum-freeconditions and mature peripheral blood erythrocytes wereevaluated by FACS as described [7]. mAbs against CCR1(Clone #53504), CCR2 (Clone #48607), CCR3 (rat mAbs
129 Chemokines and Erythropoiesis

Clones #61856, 61832, 61837, 61834), CCR5 (Clone #45529),CCR6 (Clone #53113), CXCR2 (Clone #48301), CXCR3(Clone #49801), and CXCR4 (Clone #44701) were obtainedfrom R&D Systems. The CCR4 mAb was a kind gift fromDr. Patrick Gray(ICOS Corp.; Seattle, WA) and the rabbitpolyclonal against CCR8 was a kind gift from Dr. RichardHoruk (Berlex; San Rafael, CA) to Dr. Robert Doms(University of Pennsylvania; Philadelphia, PA). Before weperformed our studies, the specificity of these antibodiesagainst their cognate receptors was confirmed on transfected293T cells. Flow cytometric staining and analysis of thereceptors were performed as described [7]. Briefly, the cellswere stained in PBS (Ca and Mg-free) supplemented with 5%BCS (Hyclone; Logan, Utah). Primary mAbs were detectedwith secondary phycoerythrin or fluorescein isothiocyanate(FITC)-conjugated goat antimouse mAbs (Sigma) (1:100) orantirat antibodies. After the final wash, cells were fixed in 1%paraformaldehyde prior to FACS analysis using FACscan(Becton Dickinson). Similarly, we have stained expanded exvivo human erythroid cells under serum-free conditions [19]and mature erythrocytes isolated from peripheral blood. Inaddition, we have also been looking for an expression ofchemokine receptors on CD34+ BMMC. In brief, BMMCwere double stained with chemokine receptors, and specificmAbs were detected with PE and anti-CD34 mAb directlyconjugated with FITC anti-HPCA-1, (20 µl/106 cells; BectonDickinson) as described [7]. Data analysis was performedusing the Cell Quest (Becton Dickinson).
Chemotaxis StudiesAll experiments were performed in triplicate on puri-
fied CD34+ or serum-free expanded erythroid cells. Briefly,after isolation the cells were resuspended in serum-freemedium (106/ml) and equilibrated for 10 min at 37°C. In themeantime 600 µl/point of prewarmed serum-free mediumcontaining appropriate ligand was added to the lowerchamber of the Costar Transwell 24-well plate, 6.5 mmdiameter, 5 µM pore filter (Costar Corning Co.; Cambridge,MA). Subsequently, 100 µl aliquots of the cell suspensionwere distributed to the upper chambers, and cultures wereincubated at 37°C, 95% humidity, 5% CO2 for 4 h. After 4h of incubation, the plates were evaluated under an invertedmicroscope and subsequently, cells from the lower cham-bers were collected and cell number was scored by FACscan(Becton Dickinson) as described [13]. Briefly, the cellswere gated according to their forward scatter (FSC) andsideward scatter (SSC) parameters and counted during a 20-sec acquisition. Data are demonstrated as a percentage ofthe input number of the cells. If required, samples of cellsfrom the lower chambers have been plated in vitro andstimulated to growth erythroid colonies as described above.
Calcium Flux Studies by FACSSorted by FACS CD34+ KIT+, isolated by magnetic beads
CD34+ and expanded ex vivo under serum-free conditions,erythroid cells were employed for Ca2+ flux studies. Cellswere loaded with Indo-1AM (Molecular Probes; Eugene, OR)as described [22]. Subsequently, 2 × 105 sorted cells or 5 × 105
expanded cells were spun down at room temperature andresuspended in 1 ml of prewarmed loading medium (1×Hank’s balanced salt solution, 1 mM MgCl2, 1 mM CaCl2, 1%fetal bovine serum [FBS]), and 40 µl of 100 mM Probenecid +10 µl of Indo-1AM mixture (50 µg of Indo-1 AM, 25 µl ofPluronic acid 127, 113 µl of FBS) were added. Cells wereincubated for 30-45 min at 30°C and were then gently mixedevery 10 min. After incubation, the cells were spun down atroom temperature, supernatant was discarded and cells wereresuspended in 300-500 µl of fresh loading medium. The cellswere then equilibrated for 10 min at 37°C and analyzed byFACS StarPlus as described [22].
Calcium Flux Studies by SpectrophotofluorimeterCalcium flux studies on expanded ex vivo CD34+ cells
were also performed by a spectrophotofluorimeter, as previ-ously described [21]. Briefly, the CD34+ cells were loadedwith Fura-2/AM (Molecular Probes) for 30 min at 37°C.Fluorescence was recorded with an Aminco-Bowman Series-2 Luminescence Spectrophotofluorimeter (SLM Instruments,Inc.; Urbana IL). Cells (1 ml aliquots) were stirred continu-ously in a warmed holder during the period of changes in flu-orescence recording. Fluorescence was monitored at 340 nmand 380 nm for excitation and 510 nm for emission. The datawere recorded as the relative ratio of fluorescence excited at340 nm and 380 nm.
Detection of MAPK and AKT Phosphorylation1 × 106 of purified CD34+KIT + cells were put in RPMI
medium containing 0.5% BCS over night in order to makethe cells quiescent. After 12 h, cells were transferred to thefresh RPMI without serum for an additional 2 h. After thattime, cells were divided and stimulated with appropriateligands for 1 min at 37°C, and subsequently they werespun down in order to stop the reaction. Supernatants wereremoved immediately and the cells were lysed in the lysingbuffer (150 mM NaCl/50 mM Tris HCl/1% Triton-X-100plus protease and phosphatase inhibitors) for 5 min on ice.Subsequently, the equal amounts of 2× loading buffer wereadded and the lysates were boiled for 3 min. The extractedproteins were separated by 15% SDS-PAGE gel and trans-ferred to the nitrocellulose membrane—Hybond ECL(Amersham Life Sciences; Little Chalfont, England).Phosphorylation of 44/42 MAPK and AKT was detectedby protein immunoblotting using monoclonal 44/42
Majka, Ratajczak, Lee et al. 130

phospho-specific MAPK or rabbit polyclonal AKT phos-pho-specific AKT antibodies (New England Biolabs;Beverly, MA) with horseradish peroxidase-conjugatedgoat anti-mouse IgG as secondary antibody (Santa CruzBiotech; Santa Cruz, CA) as described [23]. The mem-branes were developed with ECL reagent (Amersham LifeSciences) and subsequently dried and exposed to film.Equal loading in the lanes was evaluated by stripping theblot and reprobing with an anti-MAPK antibody clone#9102 and an anti-AKT antibody clone #9272 (NewEngland Biolabs).
Isolation of mRNA from BFU-ECells from expanded ex vivo BFU-E colonies growing
in serum-free methylcellulose cultures were analyzed forCXCR4, CCR5, and β-actin expression by reverse tran-scriptase-polymerase chain reaction (RT-PCR). Briefly,colonies were visualized with the aid of an inverted micro-scope and plucked from serum-free cultures on day 6through day 9 using a Pasteur pipette. Randomly selectedcolonies (~20) from each culture dish were utilized for eachassay. Colonies were resuspended in 10 ml of IMDM andincubated for 1 h at 37°C to dissolve the methylcellulose.The cells were washed 2× in PBS after which their mRNAwas extracted as detailed below.
RT-PCRCells from 20 colonies were lysed in 200 µl of RNAzol
(Biotecx Labs; Houston, TX) + 22 µl of chloroform (Sigma).The aqueous phase was collected and mixed with 1 volumeof isopropanol (Sigma). RNA was precipitated overnight at–20°C. The RNA pellet was washed in 75% ethanol andresuspended in 3×-autoclaved H2O. RT-PCR was carried outas described [7, 18, 20]. Briefly, mRNA (0.5 µg) wasreverse-transcribed with 500 U of Moloney murineleukemia virus reverse transcriptase and 50 pmol of anoligodeoxynucleotide (ODN) primer complementary to the3′ end of the following sequence of CXCR4 (5′-CAA GGAAGC TGT TGG CTG AAA-3′) or CCR5 (5′-GAA AATGAG AGC TGC AGG TGT-3′), according to the reportedcDNA sequences as described [7]. The resulting cDNAfragments were amplified using 5 U of Thermus aquaticus(Taq) polymerase and primers specific for the 5′ end ofCXCR-4 (5′-CGA GGC AAG TGA CGC CGA GGG GCTG-3′) or for the 5′ end CCR5 (5′-GCT GTC CAC ATC TCGTTC TCG-3′). β-actin mRNAs were amplified simultane-ously using specific primers as described respectively [18].Amplified products (10 µl) were electrophoresed on a 2%agarose gel, and transferred to a nylon filter and documentedphotographically. Specificity of the amplified products wasfurther confirmed by Southern blotting as described [18].
Statistical AnalysisArithmetic means and standard deviations were calculated
on a MacIntosh computer using Instat 1.14 (GraphPad; SanDiego, CA) software. Experiments were performed on cellsobtained from 15 healthy donors. Cell cultures were carriedout in quadruplicate. Data were analyzed using the Student’s t-test for unpaired samples. Statistical significance was definedas p < 0.05.
RESULTS
Expression of Chemokine Receptors on Human CD34+ CellsWhich Include Erythroid Progenitors and on More MatureErythroid Cells
The earliest erythroid progenitors (BFU-E) are foundamong CD34+ cells [24]. We therefore examined these cellsfor the presence of chemokine receptors. In our previous stud-ies, we demonstrated that ~50% of CD34+ cells express theCXCR4 protein by FACS [4, 7]. At the same time, the CCR5protein was undetectable on the surface [4, 7]; however, at thesame time we demonstrated in these cells a presence of CCR5intracellularly [25].
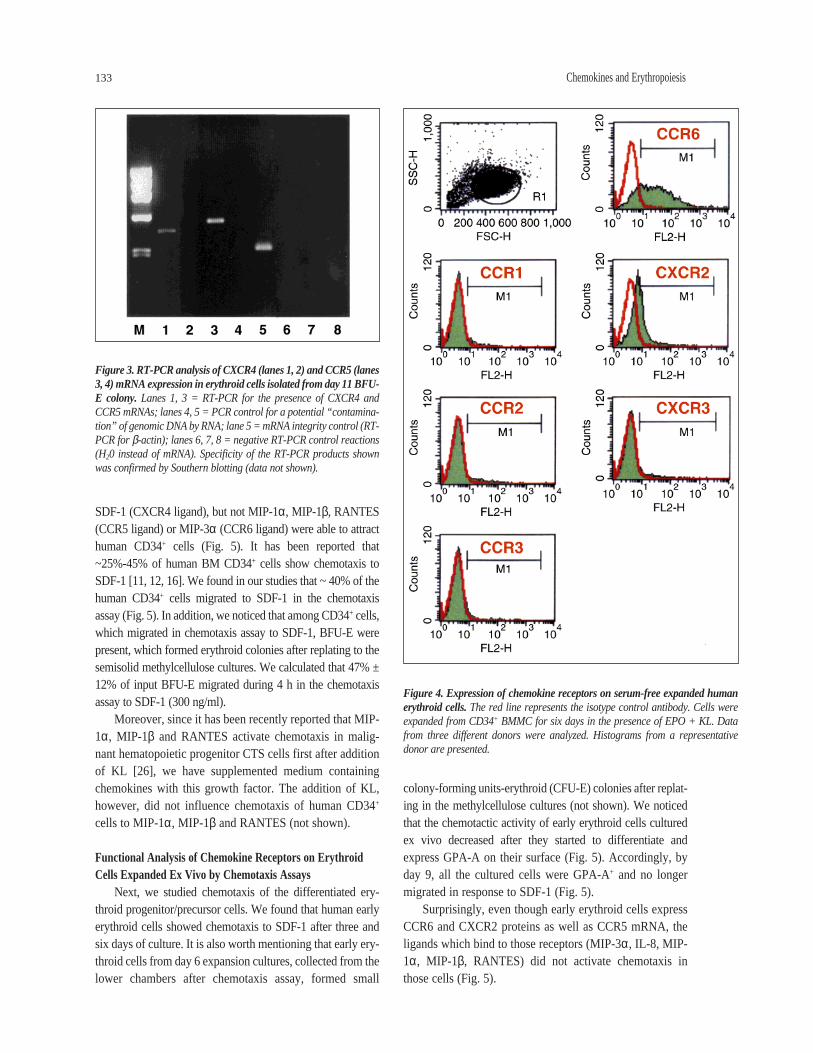
In this paper, we extended these observations by lookingfor the presence of eight other chemokine receptor proteinson human CD34+ cells. We found that in addition toCXCR4, a small percentage of CD34+ cells (~5%-10%)express the CCR6 receptor. At the same time, we wereunable to demonstrate the presence of CCR1, CCR2, CCR3,CCR4, CCR5, CCR8, CXCR2, or CXCR3 receptors on thesame cells using flow cytometry. Figure 1 demonstrates thecoexpression of selected chemokine receptors (CCR1,CCR3, CCR5, CCR6, and CXCR4) and CD34 antigen onhuman BMMC in the lymphocyte gate. We then phenotypedmore differentiated erythroid cells for expression of thesechemokine receptors. First, we looked for the presence ofmajor HIV coreceptors (CXCR4, CCR5) on human ery-throid cells grown in serum-free liquid cultures, (Fig. 2A-D). Figure 2B shows that the CXCR4 protein was stilldetectable on ~15% of human erythroid cells cultured exvivo for four days and became undetectable on more differ-entiated erythroid cells which were cultured for six days(Fig. 2D). By employing a more sensitive RT-PCR assay,we were still able to detect the presence of CXCR4 mRNAin cells isolated from day 11 erythroid colonies (Fig. 3).Similarly, while the CCR5 protein was not detected byFACS on human CD34+ cells (Fig. 1D), or cultured ery-throid cells (Fig. 2A, C), CCR5 mRNA was easily detectedby RT-PCR (Fig. 3). Therefore, we postulated that theapparent lack of expression of major chemokine receptorson the differentiated erythroid cells was the result of limitedFACS detection sensitivity.
131 Chemokines and Erythropoiesis

We also examined erythroid cells for expression of eightother chemokine receptors after 6, 9, and 11 days of ex vivoexpansion cultures. We noticed that at day 6 these cellsexpress CCR6 and CXCR2, but not CCR1, CCR2, CCR3,CCR4, CCR8, and CXCR3 proteins (Fig. 4). We also foundthat CCR6 was still present on ~20% erythroid cells,expanded for 11 days (not shown).
Finally, in order to complete the phenotype of the entirespectrum of erythroid cells, we examined human peripheralblood erythrocytes for expression of chemokine receptors, butnone were found (not shown).
Functional Studies on Chemokine Receptor-Chemokine AxesFlow cytometric detection of chemokine receptors on cell
surfaces is sensitive to the level of ~200-500 molecules. If aprotein is expressed at a lower level, a definitive statement
about expression cannot be made. Further, the absence of aparticular protein as assessed by FACS does not allow us toconclude that this receptor is not present on the cell surface. In fact, mRNAs for many chemokine receptors havebeen found in early human erythroid cells [9, 10]. Weassumed that various chemokine receptors that were notdetectable by FACS might still be functional even thoughthey are expressed at low levels on the cell surface. Toexamine this possibility we performed functional studies,looking specifically for the ability of selected chemokines toactivate chemotaxis in CD34+ progenitors and differentiat-ing erythroid cells, induce calcium flux and phosphorylateMAPK p42/44 (ERK-1, ERK-2) and AKT.
Functional Analysis of Chemokine Receptors on CD34+ Cellsby Chemotaxis Assay
As demonstrated above, ~50% of human CD34+ cellscoexpress X4 HIV-binding CXCR4 coreceptor, while ~5%-10% coexpress the CCR6 protein. In addition, human CD34+
cells express at least CCR5 mRNA for the R5 HIV cell surfacecoreceptor [4]. The biological significance of CXCR4, CCR6expression and potential presence of CCR5 receptor wasinvestigated by employing various functional studies. First,we asked whether any of the chemokines show a chemotac-tic activity against human CD34+ cells. We found that only
Majka, Ratajczak, Lee et al. 132
Figure 1. Expression of chemokine receptors on BM CD34+ BMMC. BMMCwere isolated from BM aspirates of healthy donors by Ficoll-gradient cen-trifugation and stained with mAbs against CD34 antigen (FITC) andchemokine receptors (PE). (A) Forward and side scatter analysis of BMMC.Lymphocyte region is defined by R1. (B-F) Analysis of cells dual-labeled withFITC-anti-CD34 mAb and (B) PE-anti-CCR1 mAb; (C) PE-anti-CCR3 mAb;(D) PE-anti-CCR5 mAb; (E) PE-anti-CCR6 mAb, and (F) PE-anti-CXCR4mAb. Data from at least three different donors were analyzed with similarresults. Data from a representative donor is presented.
Figure 2. FACS analysis of CCR5 (A, C) and CXCR4 (B, D) expression inerythroid cells expanded from CD34+ cells under serum-free conditions forfour days (upper panel) and six days (lower panel). The isotype negative con-trols are overlaid (bold line), and M1 represents a positive population. Datafrom four different donors were analyzed. Histograms from a representativedonor are presented.
SDF-1 (CXCR4 ligand), but not MIP-1α, MIP-1β, RANTES(CCR5 ligand) or MIP-3α (CCR6 ligand) were able to attracthuman CD34+ cells (Fig. 5). It has been reported that ~25%-45% of human BM CD34+ cells show chemotaxis toSDF-1 [11, 12, 16]. We found in our studies that ~ 40% of thehuman CD34+ cells migrated to SDF-1 in the chemotaxisassay (Fig. 5). In addition, we noticed that among CD34+ cells,which migrated in chemotaxis assay to SDF-1, BFU-E werepresent, which formed erythroid colonies after replating to thesemisolid methylcellulose cultures. We calculated that 47% ±12% of input BFU-E migrated during 4 h in the chemotaxisassay to SDF-1 (300 ng/ml).
Moreover, since it has been recently reported that MIP-1α, MIP-1β and RANTES activate chemotaxis in malig-nant hematopoietic progenitor CTS cells first after additionof KL [26], we have supplemented medium containingchemokines with this growth factor. The addition of KL,however, did not influence chemotaxis of human CD34+
cells to MIP-1α, MIP-1β and RANTES (not shown).
Functional Analysis of Chemokine Receptors on ErythroidCells Expanded Ex Vivo by Chemotaxis Assays
Next, we studied chemotaxis of the differentiated ery-throid progenitor/precursor cells. We found that human earlyerythroid cells showed chemotaxis to SDF-1 after three andsix days of culture. It is also worth mentioning that early ery-throid cells from day 6 expansion cultures, collected from thelower chambers after chemotaxis assay, formed small
colony-forming units-erythroid (CFU-E) colonies after replat-ing in the methylcellulose cultures (not shown). We noticedthat the chemotactic activity of early erythroid cells culturedex vivo decreased after they started to differentiate andexpress GPA-A on their surface (Fig. 5). Accordingly, byday 9, all the cultured cells were GPA-A+ and no longermigrated in response to SDF-1 (Fig. 5).
Surprisingly, even though early erythroid cells expressCCR6 and CXCR2 proteins as well as CCR5 mRNA, theligands which bind to those receptors (MIP-3α, IL-8, MIP-1α, MIP-1β, RANTES) did not activate chemotaxis inthose cells (Fig. 5).
133 Chemokines and Erythropoiesis
Figure 3. RT-PCR analysis of CXCR4 (lanes 1, 2) and CCR5 (lanes3, 4) mRNA expression in erythroid cells isolated from day 11 BFU-E colony. Lanes 1, 3 = RT-PCR for the presence of CXCR4 andCCR5 mRNAs; lanes 4, 5 = PCR control for a potential “contamina-tion” of genomic DNA by RNA; lane 5 = mRNA integrity control (RT-PCR for β-actin); lanes 6, 7, 8 = negative RT-PCR control reactions(H20 instead of mRNA). Specificity of the RT-PCR products shownwas confirmed by Southern blotting (data not shown).
Figure 4. Expression of chemokine receptors on serum-free expanded humanerythroid cells. The red line represents the isotype control antibody. Cells wereexpanded from CD34+ BMMC for six days in the presence of EPO + KL. Datafrom three different donors were analyzed. Histograms from a representativedonor are presented.

Functional Analysis of Chemokine Receptors on CD34+ Cellsby Calcium Flux Studies and Phosphorylation of MAPK(ERK1 and ERK2) and AKT
In order to learn more about the biological role ofchemokine receptor expression on human CD34+ cells, weperformed calcium flux studies. Since CD34 antigen marksearly lymphoid cells as well as myeloid cells, we FACSsorted a population of CD34+KIT+ cells, which is enriched inBFU-E [26], to identify responding cells by lineage.Accordingly, CD34+KIT+ cells were loaded with Indo-1AM,and calcium flux was evaluated by FACS. We found thatSDF-1 (Fig. 6A) but not MIP-1α, MIP-1β (Fig. 6B), orRANTES and MIP-3α (not shown) strongly induced calciumflux in this population of early stem/progenitor cells. Asdemonstrated in Figure 6A, SDF-1 induced calcium flux inalmost 100% of CD34+KIT+ cells. Similarly, we found thatSDF-1, but not MIP-1α, MIP-1β, MIP-3α, or MCP-3, wereable to phosphorylate ERK1 and ERK2 and AKT in humanCD34+KIT+ cells (Fig. 7). This strongly supports our obser-vation that, in contrast to CCR5, CCR6 and CCR2, CXCR4receptor protein is not only detectable but functional onhuman CD34+ KIT+ cells.
Functional Analysis of the Chemokine Receptors on ErythroidCells Expanded Ex Vivo by the Calcium Flux Studies
The ability of different chemokines to induce calciumflux, both in the CD34+ cells stimulated for two days withKL + EPO for erythropoietic expansion (Fig. 8), and inmore differentiated erythroid cells that had been expandedex vivo for three to six days, was also evaluated (Fig. 9).
Majka, Ratajczak, Lee et al. 134
MIP-1β Glycophorin A
45%
40%
35%
30%
25%
20%
15%
10%
5%
0%
100%
90%
80%
70%
60%
50%
40%
30%
20%
10%
0%
Ch
emo
taxi
s
Gly
cop
ho
rin
A
Day 0 Day 3 Day 6 Day 9
SDF-1MIP-3α
MIP-1αControl
Figure 5. Activation of chemotaxis by various chemokines in the human erythroid cells, expanded under serum-free conditions from CD34+
BMMC. Chemotaxis was evaluated at days 3, 6, and 9 (x axis). The percentage of the cells which responded by chemotaxis to different chemokinesis shown on y axis (to the left), and the percentage of GPA-A+ cells is shown on y axis (to the right). The data are pooled from the three inde-pendent experiments performed on cells from the three different donors.
Figure 6. FACS study on calcium flux in CD34+KIT+ cells. Cells were loadedwith Indo-1AM and subsequently stimulated by SDF-1 (A) and MIP-1α, MIP-1β,and RANTES (B). A representative experiment is shown of three independentrepeats with similar results.

Accordingly, CD34+ cells were isolated by magnetic beadsand cultured serum-free for two days with KL + EPO (Fig. 8),and calcium flux was first evaluated by spectrophotofluo-rimeter. We found that SDF-1 (Fig. 8B) but not MIP-1α(Fig. 8A), MIP-1β, RANTES, and MIP-3α (not shown)were able to induce calcium flux in this population of cells.
Subsequently, calcium flux was examined in culturedhuman erythroid cells by FACS (Fig. 9). We chose this tech-nique because FACS, in contrast to spectrophotofluorimetry,
not only allows for detection of a calcium flux itself but alsoallows for an estimate of the percentage of the cells whichrespond in this assay. We noticed that a subpopulation of ery-throid cells, which was expanded ex vivo for three (Fig. 9A)and four days (Fig. 9B) responded to stimulation with SDF-1.To our surprise ex vivo-expanded erythroid cells did notshow calcium flux after stimulation with MIP-1α, MIP-1β,RANTES, IL-8, and MIP-3α (not shown) even though thecorresponding chemokine receptors were expressed.Simultaneously, we did not observe any influence of those
135 Chemokines and Erythropoiesis
Figure 7. Western blot data on phosphorylation of MAPK (ERK1 - p44 andERK2 - p42) (panel A) and AKT (panel B) in CD34+KIT+ BMMC cells. Sortedby FACS, human CD34+KIT+ BMMC were starved for 24 h in serum-depletedmedium, and subsequently nonstimulated (lane 1) or stimulated for 1 min withMIP-1α (lane 2), MIP-1β (lane 3), MIP-3α (lane 4), MCP-1 (lane 5), and SDF-1 (lane 6). Equal loading in the lanes was evaluated by stripping the blot andreprobing with an anti-MAPK antibody (ERK1/ERK2) and an anti-AKT antibody(lower parts of panels A and B, respectively). A representative experiment isshown of three independent repeats with similar results.
Figure 8. Representative spectrophotofluorimetrical calcium flux study inhuman CD34+ cells expanded serum-free for two days toward erythroid lineage.CD34+ cells were isolated by magnetic beads, loaded with Fura-2/AM and sub-sequently stimulated by MIP-1α (A) and SDF-1 (B). Three different experimentswere performed with similar results.
Figure 9. Calcium flux in human day-3 (A), day-4 (B) and day-6 (C) serum-free expanded erythropoietic cells. Cells were loaded with Indo-1AM and subsequently stimulated by SDF-1. Calcium flux was evaluated by FACS. A representative experiment is shown of three independentrepeats with similar results.

chemokines on MAPK and AKT phosphorylation in thesame population of cells (not shown).
Influence of HIV-Related and Other Chemokines on BFU-EGrowth Under Serum-Free Conditions
Data from the literature on the influence of chemokineson human hematopoiesis are inconsistent [6, 8-10, 27-32] andin our hands neither MIP-1α nor IL-8 inhibited human ery-thropoiesis in vitro [33]. We therefore reexamined theseobservations focusing on those chemokines, for which thereceptors are expressed at a high level on the BFU-E(CXCR4) or on more differentiated erythroid cells (CCR6,CXCR2). Human CD34+ cells were isolated by magneticbeads and cultured serum-free in the presence or absence ofdifferent chemokines (Table 1). Transforming growth factor-β1, a potent known inhibitor of the erythropoietic colony formation, was added to some cultures [34].
As shown in Table 1, SDF-1 (CXCR4 ligand), MIP-3α(CCR6 ligand) and IL-8, Gro-α, -β, -γ, NAP-2, ENA-78(CXCR2 ligands) all failed to affect human BFU-E growthin vitro. Similarly, MIP-1α, MIP-1β, and RANTES (CCR5ligands) were without apparent effect. After addition ofchemokines, the number of colonies, their hemoglobiniza-tion, and expression of GPA-A remain unchanged. At thesame time, however, the growth of the erythroid colonieswas strongly inhibited after an addition of TGF-β1 to thecultures. Similar data also were obtained when BFU-Egrowth was stimulated with a suboptimal dose of EPO + KL(not shown).
DISCUSSION
The role of HIV-related chemokines in human erythro-poiesis is important for two reasons. First, HIV-relatedchemokines are elaborated in the serum of AIDS patients andthus may influence human erythropoiesis [1, 5]. Second, thetherapeutic targeting of HIV-related chemokine receptor-chemokine axes, e.g., by employing receptor agonists orblocking mAbs [1], could potentially affect the proliferationof human erythroid progenitors in HIV-infected patients.
We found that CXCR4 is highly expressed on humanCD34+ cells enriched for BFU-E. In this study, we also phe-notyped human CD34+ cells for the expression of eightother chemokine receptors using FACS. We found that asmall population of CD34+ cells (5%-10%) expresses theCCR6 protein, which binds MIP-3α. Other receptors wereundetectable. Hence, CXCR4 remains the most highlyexpressed member of this receptor family on human CD34+
cells. This observation supports other reports demonstratingthat CXCR4 plays an important and privileged function inhoming and trafficking of human CD34+ progenitor/stemcells [11, 13, 15, 17, 35]. Our failure to detect CCR1, CCR3,
and CCR5 proteins onCD34+ cells is intrigu-ing because all thesereceptors bind chemo-kines which interferewith R5 HIV-1 entryinto the cells (MIP-1α,MIP-1β, RANTES) [10].In addition, CCR1 andCCR5 bind MIP-1α,which has been foundto inhibit proliferationof CD34+ cells [8].
To validate theseobservations on a popu-lation of pure erythroidcells, we phenotypedhuman erythroid cellsexpanded ex vivo aswell as mature erythro-cytes. We found that theexpression of CXCR4varied with the state oferythroid maturationand could not bedetected on BFU-E-derived cells expanded for six days. Nevertheless, these moredifferentiated erythroid cells still contained CXCR4 mRNA.In contrast, human erythroid cells expressed CCR6 protein,and to a lesser degree CXCR2, after six days of culture.Expression of CCR6 and CXCR2 decreased as the erythroidcells matured. Finally, we looked for the presence ofchemokine receptors on mature human peripheral blood ery-throcytes but were unable to detect any of the chemokinereceptors for which we tested.
In functional assays, we found that SDF-1 (CXCR4 lig-and), but not MIP-1α, MIP-1β, RANTES (CCR5 ligand),IL-8 (CXCR2 ligand) or MIP-3α (CCR6 ligand) inducedcalcium flux and phosphorylated MAPK ERK-1 and ERK-2 and AKT in a FACS-purified CD34+KIT+ population.Moreover, only SDF-1-induced calcium flux in human ery-throid cells expanded for two to four days. The response toSDF-1 disappeared when the cells began to differentiateand express GPA-A. In contrast, even though more differ-entiated erythroid cells expressed CCR6 and CXCR2receptors, their corresponding ligands (MIP-3α and IL-8)did not activate calcium flux in those cells. We also con-firmed that of all the chemokines tested, only SDF-1 is astrong chemotactic factor for human CD34+ cells [11, 13]as well as for human CD34+ BFU-E and for the more dif-ferentiated CD34– CFU-E progenitor cells. Of note, none of
Majka, Ratajczak, Lee et al. 136
Table 1. The effect of various chemokines (100 ng/ml) and TGF-β1 (5 ng/ml) on BFU-Eformation by human CD34+ cells
BFU-E growth n BFU-E coloniescostimulated with: (±SD)/104 cells plated
(—) 219 ± 34
SDF-1 222 ± 51
MIP-1α 215 ± 49
MIP-1β 243 ± 66
RANTES 219 ± 57
MIP-3α 211 ± 43
IL-8 228 ± 39
Gro-α 217 ± 52
Gro-β 220 ± 37
Gro-γ 208 ± 28
NAP-2 231 ± 53
ENA-78 214 ± 28
TGF-β1 43 ± 22*
Isolated by magnetic beads CD34+ cells wereplated serum-free and stimulated with EPO (5 U/ml) + KL (100 ng/ml). Each data entryconstitutes four independent clonogenic assaysfrom six different donors.
*p < 0.0001 in comparison to (—).

the chemokines evaluated in this study affected cloningefficiency or maturation of human BFU-E. These data arein apparent conflict with other reports which found thatMIP-1α inhibits human erythroid colony formation [8, 10].
The contradictory responses to MIP-1α reported heremight be explained by differences in MIP-1α protein purifi-cation, differences in the cloning system itself and in theformation of inactive dimers [29]. This latter possibility isunlikely because the same preparations of MIP-1α-acti-vated calcium flux in our study and chemotaxis in certainhuman hematopoietic cell lines (not shown). This suggeststhat the source of MIP-1α we utilized in our experimentswas biologically active. In addition, the reported inhibitoryeffects of MIP-1α on hematopoiesis are often unclear [6].Mice with MIP-1α knock-out do not show any hematopoi-etic defects [28]. In some reports MIP-1α was found to beonly inhibitory if added at a very low dose [29], whereas inothers was reported to inhibit cell growth if added at a veryhigh dose and repetitively added doses [31].
Finally, after we recently demonstrated that human BMCD34+ cells secrete MIP-1α endogenously [25], we concludedthat the role of this chemokine in regulating humanhematopoiesis is more complex than we initially thought [6, 8, 29]. In light of our recent data we conclude that humanBM CD34+ cells are exposed in vivo to low levels of secretedMIP-1α and other chemokines [25].
Based on the assays carried out in this study (chemotaxis,calcium flux, phosphorylation of MAPK and AKT), we con-cluded that only the CXCR4 receptor is functional on the ear-liest human erythroid cells. It appears to play an important rolein regulating chemotaxis and perhaps in homing and retentionof erythroid progenitors in the BM microenvironment.Therefore, the perturbation of the SDF-1/CXCR4 axis whichoccurs during HIV infection or when induced by therapeuticagents designed to block HIV cell entry (e.g., ALX40) [1, 36]may affect the homing and development of erythroid cells.Since we did not observe any effect of chemokines on ery-throid colony formation by human CD34+ cells, we concludethat the influence of chemokines on human hematopoiesis ismore complicated than initially thought. Further studies areneeded to elucidate their role in human hematopoiesis, includ-ing examining their autocrine effect on individual hematopoi-etic lineages [25] as well as their indirect influence as mediatedby interactions with hematopoietic accessory cells.
ACKNOWLEDGMENTS
We would like to thank R&D Systems for providingsome of the chemokine receptor antibodies used in this study.This work was supported by a grant NIH R01 HL61796-01to Dr. MZRand NIH DK52558-A2 to Dr. AMG.
137 Chemokines and Erythropoiesis
REFERENCES
1 Cairns JS, D’Souza MP. Chemokines and HIV-1 second recep-tors: the therapeutic connection. Nat Med 1998;4:563-568.
2 Fauci AS. Host factors and the pathogenesis of HIV-induceddisease. Nature 1996;384:529-534.
3 Hoxie JA. Hematologic manifestations of AIDS. In Hoffman R,Benz EJ, Shatill SJ et al., eds. Hematology Basic Principles andPractice. New York: Churchill Livingstone, 1995;2171-2200.
4 Lee B, Doranz BJ, Ratajczak MZ et al. An intricate web:chemokine receptors, HIV-1 and hematopoiesis. STEM CELLS1998;16:79-88.
5 Moses A, Nelson J, Bagby GC. The influence of human immun-odeficiency virus-1 on hematopoiesis. Blood 1998;91:1479-1495.
6 Rollins BJ. Chemokines. Blood 1997;90:909-928.
7 Lee B, Ratajczak J, Doms RW et al. Coreceptor/chemokinereceptor expression on human hematopoietic cells: biologicalimplications for HIV-1 infection. Blood 1999;93:1145-1156.
8 Broxmeyer HE, Sherry B, Cooper S et al. Comparative analy-sis of the human macrophage inflammatory protein family ofcytokines (chemokines) on proliferation of human myeloidprogenitor cells. J Immunol 1993;150:3448-3458.
9 Su S, Mukaida N, Wang J et al. Inhibition of immature ery-throid progenitor cell proliferation by macrophage inflamma-tory protein-1α by interacting mainly with a C-C chemokinereceptor, CCR1. Blood 1997;90:605-611.
10 de Wynter EA, Durig J, Cross MA et al. Differential responseof CD34+ cells isolated from cord blood and bone marrow toMIP-1α and the expression of MIP-1α receptors on theseimmature cells. STEM CELLS 1998;16:349-356.
11 Aiuti A, Webb IJ, Bleul C et al. The chemokine SDF-1 is achemoattractant for human CD34+ hematopoietic progenitorcells and provides a new mechanism to explain the mobiliza-tion of CD34+ progenitors to peripheral blood. J Exp Med1997;185:111-120.
12 Bleul CC, Fuhlbridge RC, Casanovas JM et al. A highly effi-cacious lymphocyte chemoattractant, stromal cell-derivedfactor 1 (SDF-1). J Exp Med 1996;184:1101-1109.
13 Kim CH, Broxmeyer HE. In vitro behavior of hematopoieticprogenitor cells under the influence of chemoattractants: stro-mal cell-derived factor-1, steel factor, and the bone marrowenvironment. Blood 1998;91:100-110.
14 Ma Q, Jones D, Borghesani PR et al. Impaired B-lymphopoiesis,myelopoiesis, and derailed cerebellar neuron migration inCXCR4 and SDF-1 deficient mice. Proc Natl Acad Sci USA1998;95:9448-9453.
15 Mohle R, Bautz F, Rafii S et al. The chemokine receptor CXCR4is expressed on CD34+ hematopoietic progenitors and leukemiccells and mediates transendothelial migration induced by stromalcell-derived factor-1. Blood 1998;91:4523-4530.

16 Nagasawa T, Hirota S, Tachibana K et al. Defects of B-celllymphopoiesis and bone-marrow myelopoiesis in mice lackingthe CXC chemokine PBSF/SDF-1. Nature 1998;382:635-638.
17 Zou Y, Kottmann AH, Kurod M et al. Function of thechemokine receptor CXCR4 in haematopoiesis and in cerebellardevelopment. Nature 1998;393:595-599.
18 Ratajczak J, Zhang Q, Pertusini E et al. The role of insulin(INS) and insulin like growth factor-I (IGF-I) in regulatinghuman erythropoiesis. Studies in vitro under serum free con-ditions—comparison to other cytokines and growth factors.Leukemia 1998;12:371-381.
19 Ratajczak J, Marlicz W, Machalinski B et al. An improvedserum free system for cloning human “pure” erythroid colonies.The role of the different growth factors and cytokines on BFU-E formation by the bone marrow and cord blood CD34+ cells.Folia Histochem Cytobiol 1998;36:55-60.
20 Ratajczak MZ, Plecher ChH, Marlicz W et al. CD34+, Kit+,Rhodamine123low phenotype identifies a marrow cell popula-tion highly enriched for human hematopoietic stem cells.Leukemia 1998;12:942-950.
21 Kowalska MA, Ratajczak J, Hoxie J et al. Platelet andmegakaryocytes express the HIV co-receptor CXCR4 on theirsurface but do not respond to stromal derived factor (SDF)-1. BrJ Haematol 1999;104:220-229.
22 Abe R, Vandenberghe P, Craighead N et al. Distinct signaltransduction in mouse CD4+ and CD8+ splenic T cells afterCD28 receptor ligation. J Immunol 1995;154:985-997.
23 Rocca JDG, Biesen T, Daaka Y et al. Ras-dependent mito-gen-activated protein kinase activation by G protein-coupledreceptors. Convergence of Gi- and Gq- mediated pathways oncalcium/calmodulin, Pyk2, and Src kinase. J Biol Chem1998;272:19125-19132.
24 De Jong MO, Wagemaker G, Wognum AW. Separation ofmyeloid and erythroid progenitors based on expression ofCD34 and c-kit. Blood 1995;86:4076-4085.
25 Majka M, Rozmyslowicz T, Lee B et al. Bone marrow CD34+
cells and megakaryoblasts secrete β-chemokines; implica-tions for infectability by M-tropic human immunodeficiencyvirus (R5 HIV). J Clin Invest 1999;104:1739-1749.
26 Dutt P, Wang JF, Groopman JE. Stromal cell-derived factor-1αand stem cell factor/kit ligand share signaling pathways in
hemopoietic progenitors: a potential mechanism for cooperativeinduction of chemotaxis. J Immunol 1998;161:3652-3658.
27 Cashman JD, Eaves CJ, Sarris AH et al. MCP-1, not MIP-1α,is the endogenous chemokine that cooperates with TGF-β toinhibit the cycling of primitive normal but not leukemic(CML) progenitors in long-term human marrow cultures.Blood 1998;92:2338-2344.
28 Cook DN, Beck MA, Coffman TM et al. Requirement ofMIP-1α for an inflammatory response to viral infection.Science 1995;269:1583-1585.
29 Mantel C, Kim YJ, Cooper S et al. Polymerization of murinemacrophage inflammatory protein 1α inactivates its myelo-suppressive effects in vitro: the active form is monomer. ProcNatl Acad Sci USA 1993;90:2232-2236.
30 Miller JS, McCullar V, Verfaille CM. Ex vivo culture ofCD34+/Lin–/DR– cells in stroma-derived soluble factors, inter-leukin-3, and macrophage inflammatory protein-1α maintainsnot only myeloid but also lymphoid progenitors in a novelswitch culture assay. Blood 1998;91:4516-4522.
31 Patel VP, Kreider BL, Li Y et al. Molecular and functionalchracterization of two novel human C-C chemokines asinhibitors of two distinct classes of myeloid progenitors. J ExpMed 1997;185:1163-1172.
32 Sanchez X, Cousins-Hodges B, Aguilar T et al. Activation ofHIV-1 coreceptor (CXCR4) mediates myelosuppression. J BiolChem 1997;272:27529-27531.
33 Gewirtz AM, Zhang J, Ratajczak J et al. Chemokine regulationof human megakaryocytopoiesis. Blood 1995;86:2559-2567.
34 Zauli G, Vitale M, Gibellini D et al. Inhibition of purifiedCD34+ hematopoietic progenitor cells by human immunodefi-ciency virus 1 or gp 120 mediated by endogenous transforminggrowth factor β1. J Exp Med 1996;183:99-108.
35 Wang JF, Liu ZY, Groopman JF. The β-chemokine receptorCXCR4 is expressed on the megakaryocytic lineage fromprogenitor to platelets and modulates migration and adhesion.Blood 1998;92:756-764.
36 Doranz BJ, Govit-Ferbas K, Sharron MP et al. A small mol-ecule inhibitor directed against the chemokine receptorCXCR4 prevents its use as an HIV-1 coreceptor. J Exp Med1997;186:1395-1400.
Majka, Ratajczak, Lee et al. 138




















