
The role of different VEGF isoforms in scar formation after glaucoma filtration surgery
11
The role of different VEGF isoforms in scar formation after glaucoma filtration surgery q Tine Van Bergen a, * , Evelien Vandewalle a, b , Sara Van de Veire a, b, c , Mieke Dewerchin c , Jean-Marie Stassen d , Lieve Moons e , Ingeborg Stalmans a, b, * a Laboratory of Ophthalmology, KU Leuven, Herestraat 49, 3000 Leuven, Belgium b Department of Ophthalmology, University Hospitals Leuven, Kapucijnenvoer 33, 3000 Leuven, Belgium c Vesalius Research Center, VIB e KU Leuven, Herestraat 49, 3000 Leuven, Belgium d ThromboGenics NV, Gaston Geenslaan 1, 3001 Heverlee, Belgium e Department of Biology, KU Leuven, Naamsestraat 61, 3000 Leuven, Belgium article info Article history: Received 23 May 2011 Accepted in revised form 29 August 2011 Available online 1 September 2011 Keywords: vascular endothelial growth factor pegaptanib Tenon fibroblasts human umbilical vein endothelial cells glaucoma filtration surgery wound healing abstract Vascular endothelial growth factor (VEGF) plays an important role in both physiological and pathological angiogenesis. Our previous studies showed a differential role of VEGF isoforms in retinal physiological angiogenesis. We also demonstrated that non-selective inhibition of VEGF by bevacizumab had a bene- ficial effect on surgical outcome after glaucoma filtration surgery by reducing angiogenesis as well as fibrosis. However, the function of the VEGF isoforms in pathological angiogenesis and wound healing in the eye still remains unidentified. This study was designed to elucidate the differential roles of VEGF isoforms in scar formation after trabeculectomy. Furthermore, we also investigated whether pegaptanib (MacugenÔ,Pfizer), an aptamer which specifically blocks VEGF 165 , could improve surgical outcome by reducing postoperative scarring. VEGF-R2 and neuropilin-1 (NRP-1) expression was analyzed in vitro by RT-PCR, and were found to be expressed at higher levels in human umbilical vein endothelial cells (HUVEC) as compared to Tenon fibroblasts (TF). The effect of the different VEGF isoforms (VEGF 121 , VEGF 165 and VEGF 189 ) and pegaptanib on cell proliferation was determined via WST-1 assay. Endothelial cell proliferation was stimulated after addition of VEGF 121 and VEGF 165 , whereas VEGF 121 and VEGF 189 increased fibroblast growth. These effects on proliferation were associated with an activation of the ERK pathway, as revealed using the TransAM c-Myc assay. Inhibition of the ERK pathway, by PD98059 administration, significantly reduced VEGF isoform induced cell growth. A dose-dependent reduction of endothelial cell proliferation was observed after pegaptanib administration, while only the highest dose was able to inhibit fibroblast growth. Next, the in vivo effect of pegaptanib was investigated in a rabbit model of trabeculectomy. The surgical outcome was evaluated by performing clinical investigations (IOP, bleb area, height and survival), as well as histomorphometric analyses of angiogenesis (CD31), inflam- mation (CD45) and fibrosis (Sirius Red). A single postoperative application of pegaptanib had a beneficial impact on surgical outcome, mainly by reducing angiogenesis, but not inflammation or collagen depo- sition. Repeated injections slightly improved surgical outcome, but again solely by reducing angiogen- esis. In summary, our results revealed that the VEGF isoforms play a differential role in ocular wound healing: VEGF 165 and VEGF 121 predominantly affect blood vessel growth, whereas VEGF 189 is rather involved in fibrosis, an important process in wound healing. Ó 2011 Elsevier Ltd. All rights reserved. Abbreviations: VEGF, vascular endothelial growth factor; VEGF-R2, VEGF receptor 2; NRP-1, neuropilin-1; HSPGs, heparin sulphate proteoglycans; HUVEC, human umbilical vein endothelial cells; HTF, human Tenon fibroblasts; RTF, rabbit Tenon fibroblast. q Supported by IWT (Agency for Innovation by Science and Technology in Flanders), FWO (Research Foundation Flanders) and FRO (Funds for Research in Ophthalmology). * Corresponding authors. Laboratory of Ophthalmology, KU Leuven, Herestraat 49, Box 1025, B-3000 Leuven, Belgium. Tel.: þ32 16 33 04 61; fax: þ3216 33 04 67. E-mail addresses: [email protected] (T. Van Bergen), [email protected] (E. Vandewalle), [email protected] (S. Van de Veire), [email protected] (M. Dewerchin), [email protected] (J.-M. Stassen), [email protected] (L. Moons), [email protected] (I. Stalmans). Contents lists available at SciVerse ScienceDirect Experimental Eye Research journal homepage: www.elsevier.com/locate/yexer 0014-4835/$ e see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.exer.2011.08.016 Experimental Eye Research 93 (2011) 689e699
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The role of different VEGF isoforms in scar formation after glaucoma filtration surgery

The role of different VEGF isoforms in scar formation after glaucoma
filtration surgeryq
Tine Van Bergena,*, Evelien Vandewallea,b, Sara Van de Veirea,b,c, Mieke Dewerchinc,Jean-Marie Stassend, Lieve Moonse, Ingeborg Stalmansa,b,*
a Laboratory of Ophthalmology, KU Leuven, Herestraat 49, 3000 Leuven, BelgiumbDepartment of Ophthalmology, University Hospitals Leuven, Kapucijnenvoer 33, 3000 Leuven, BelgiumcVesalius Research Center, VIB e KU Leuven, Herestraat 49, 3000 Leuven, Belgiumd ThromboGenics NV, Gaston Geenslaan 1, 3001 Heverlee, BelgiumeDepartment of Biology, KU Leuven, Naamsestraat 61, 3000 Leuven, Belgium
a r t i c l e i n f o
Article history:
Received 23 May 2011
Accepted in revised form 29 August 2011
Available online 1 September 2011
Keywords:
vascular endothelial growth factor
pegaptanib
Tenon fibroblasts
human umbilical vein endothelial cells
glaucoma filtration surgery
wound healing
a b s t r a c t
Vascular endothelial growth factor (VEGF) plays an important role in both physiological and pathological
angiogenesis. Our previous studies showed a differential role of VEGF isoforms in retinal physiological
angiogenesis. We also demonstrated that non-selective inhibition of VEGF by bevacizumab had a bene-
ficial effect on surgical outcome after glaucoma filtration surgery by reducing angiogenesis as well as
fibrosis. However, the function of the VEGF isoforms in pathological angiogenesis and wound healing in
the eye still remains unidentified. This study was designed to elucidate the differential roles of VEGF
isoforms in scar formation after trabeculectomy. Furthermore, we also investigated whether pegaptanib
(Macugen�, Pfizer), an aptamer which specifically blocks VEGF165, could improve surgical outcome by
reducing postoperative scarring. VEGF-R2 and neuropilin-1 (NRP-1) expression was analyzed in vitro by
RT-PCR, and were found to be expressed at higher levels in human umbilical vein endothelial cells
(HUVEC) as compared to Tenon fibroblasts (TF). The effect of the different VEGF isoforms (VEGF121,
VEGF165 and VEGF189) and pegaptanib on cell proliferation was determined via WST-1 assay. Endothelial
cell proliferation was stimulated after addition of VEGF121 and VEGF165, whereas VEGF121 and VEGF189increased fibroblast growth. These effects on proliferation were associated with an activation of the ERK
pathway, as revealed using the TransAM c-Myc assay. Inhibition of the ERK pathway, by PD98059
administration, significantly reduced VEGF isoform induced cell growth. A dose-dependent reduction of
endothelial cell proliferation was observed after pegaptanib administration, while only the highest dose
was able to inhibit fibroblast growth. Next, the in vivo effect of pegaptanib was investigated in a rabbit
model of trabeculectomy. The surgical outcome was evaluated by performing clinical investigations (IOP,
bleb area, height and survival), as well as histomorphometric analyses of angiogenesis (CD31), inflam-
mation (CD45) and fibrosis (Sirius Red). A single postoperative application of pegaptanib had a beneficial
impact on surgical outcome, mainly by reducing angiogenesis, but not inflammation or collagen depo-
sition. Repeated injections slightly improved surgical outcome, but again solely by reducing angiogen-
esis. In summary, our results revealed that the VEGF isoforms play a differential role in ocular wound
healing: VEGF165 and VEGF121 predominantly affect blood vessel growth, whereas VEGF189 is rather
involved in fibrosis, an important process in wound healing.
� 2011 Elsevier Ltd. All rights reserved.
Abbreviations: VEGF, vascular endothelial growth factor; VEGF-R2, VEGF receptor 2; NRP-1, neuropilin-1; HSPGs, heparin sulphate proteoglycans; HUVEC, human
umbilical vein endothelial cells; HTF, human Tenon fibroblasts; RTF, rabbit Tenon fibroblast.q Supported by IWT (Agency for Innovation by Science and Technology in Flanders), FWO (Research Foundation Flanders) and FRO (Funds for Research in Ophthalmology).
* Corresponding authors. Laboratory of Ophthalmology, KU Leuven, Herestraat 49, Box 1025, B-3000 Leuven, Belgium. Tel.: þ32 16 33 04 61; fax: þ32 16 33 04 67.
E-mail addresses: [email protected] (T. Van Bergen), [email protected] (E. Vandewalle), [email protected] (S. Van de Veire),
[email protected] (M. Dewerchin), [email protected] (J.-M. Stassen), [email protected] (L. Moons), [email protected]
(I. Stalmans).
Contents lists available at SciVerse ScienceDirect
Experimental Eye Research
journal homepage: www.elsevier .com/locate/yexer
0014-4835/$ e see front matter � 2011 Elsevier Ltd. All rights reserved.
doi:10.1016/j.exer.2011.08.016
Experimental Eye Research 93 (2011) 689e699

1. Introduction
Vascular endothelial growth factor (VEGF) is best known as an
endothelial and permeability factor and plays a role in both normal
(e.g. embryonic development) (Carmeliet et al.,1996) and pathological
angiogenesis (e.g. in tumour growth, inflammation, wound healing
and various ocular diseases) (Dvorak et al., 1999; Kowanetz and
Ferrara, 2006; Wilgus et al., 2008). VEGF is known to stimulate
endothelial cells (Rini and Small, 2005) and many non-vascular cells,
such as Tenon fibroblasts (Li et al., 2009). It acts primarily via three
high-affinity VEGF tyrosine kinase receptors: Flt-1 (VEGF-R1), KDR
(VEGF-R2) and Flt-4 (VEGF-R3) (Ferrara et al., 2003; Li et al., 2009).
VEGF-R2 mediates most biologically relevant VEGF responses,
including cell migration and proliferation (Zachary, 2003). VEGF
binding to this receptor mediates the activation of the mitogen-
activated protein kinase kinase (MEK)/extracellular signal-regulated
kinase (ERK) cascade, known to be a major mediator of VEGF-
induced proliferation, both in endothelial cells (Takahashi et al.,
2001) and fibroblasts (Wu et al., 2004). Upon activation, ERK phos-
phorylates a variety of intracellular targets, one of which is c-Myc,
a short-lived transcription factor involved in multiple aspects of
growth control (Seth et al., 1992) and proliferation (Pap et al., 2004;
Pintus et al., 2002).
Alternative splicing of a single VEGF-gene results in multiple
isoforms, such as VEGF121, VEGF165 and VEGF189. These isoforms
are classified by the number of amino acids (Zhang et al., 2008)
and differ in molecular mass, solubility and (co-)receptor binding
properties (Stalmans et al., 2002). Although it is well-known that
the different isoforms bind VEGF-R2 with equal affinity (Yamazaki
and Morita, 2006; Zhang et al., 2000), they do differ in their
affinity for VEGF-R co-receptors, such as neuropilin-1 (NRP-1) and
heparin sulphate proteoglycans (HSPGs). The presence of
sequences encoded by exon 6 or 7 regulates interactions with
these co-receptors (Olsson et al., 2006; Robinson and Stringer,
2001). VEGF121 lacks these exons and therefore also lacks the
HSPGs- and NRP-1-binding domains. NRP-1, recognizing the exon-
7-encoded domain of VEGF, acts as a co-receptor for VEGF165 and
VEGF189 and enhances their binding to VEGF-R2 and their bioac-
tivity (Soker et al., 2002). HSPGs, known to be expressed on the
cell surface of almost all mammalian cells, interact with many
proteins. VEGF165, lacking exon 6, has a moderate affinity for
HSPGs, while VEGF189 contains an additional sequence encoded by
exon 6a and therefore strongly binds HSPGs (Olsson et al., 2006;
Robinson and Stringer, 2001).
The differential role of the VEGF isoforms in vascular patterning
and arterial development in the retina has been described
(Stalmans et al., 2002). However, the function of these isoforms in
pathological angiogenesis and wound healing in the eye still
remains unidentified. Recently, we showed that VEGF plays an
important role in scar formation after filtration surgery. VEGF was
expressed in samples of aqueous humor from glaucoma patients
and rabbits that had undergone filtration surgery. Furthermore,
VEGF stimulated the proliferation of Tenon fibroblasts in vitro and
different isoforms (VEGF121, VEGF165, VEGF189) were expressed by
these cells (Li et al., 2009). Recently, Zhang et al. (Zhang et al., 2008)
reported different vascular responses to VEGF121 and VEGF165.
While VEGF165 significantly stimulated the signalling pathway in
human umbilical vein endothelial cells (HUVEC), VEGF121 had little
effect. These data suggest that VEGF isoforms might play distinct
roles by activating different signalling pathways.
Pegaptanib sodium (Macugen�, Pfizer, New York), an aptamer
that prevents the binding of VEGF165 to its receptor, has been
approved by the Food and Drugs Administration for the treatment
of neovascular age-relatedmacular degeneration (AMD). The phase
III VEGF Inhibition Study in Ocular Neovascularization (VISION)
clinical trials showed that intravitreal injections of pegaptanib slow
visual loss in neovascular AMD (Ciulla and Rosenfeld, 2009).
The aim of this study was to elucidate the role of the VEGF
isoforms in scar formation after glaucoma filtration surgery.
Different in vitro and in vivo models were used to study angio-
genesis, as well as inflammatory and fibrotic processes. Secondly,
we evaluated the therapeutic potential of selective VEGF inhibition
(pegaptanib) on ocular wound healing, using a rabbit model of
glaucoma filtration surgery.
2. Materials and methods
All animal procedures were performed in accordance with the
ARVO Statement for the Use of Animals in Ophthalmic and Vision
Research and the EC Directive 86/609/EEC for animal experiments,
using the guidelines of our own institution.
2.1. Cells and culture conditions
Human umbilical vein endothelial cells (HUVEC) (Lonza, Wal-
kersville, Maryland) and Tenon fibroblasts (TF) were used. Human
Tenon fibroblasts (HTF) were obtained during strabismus surgery
and rabbit Tenon fibroblasts (RTF) were harvested during filtration
surgery, as previously described (Li et al., 2009).
HUVEC and TF were maintained at 37 �C in a humidified
atmosphere of 5% CO2 in respectively EGM-2 and DMEM complete
medium, as previously described (Li et al., 2009). EGM-2 medium
was supplemented with 10% foetal bovine serum (FBS), 0.1% human
recombinant epidermal growth factor, 0.4% recombinant human
fibroblast growth factor, 0.1% VEGF, 0.1% recombinant lung insulin-
like growth factor, 0.04% hydrocortisone, 0.1% ascorbic acid, 0.1%
heparin, 0.1% gentamicin sulphate amphotericin-B (EGM-2 Bullet-
Kit, Lonza, Walkersville, Maryland). HUVEC were cultured in 0.1%
gelatine (Invitrogen, Carlsbad, California) coated dishes. In all
experiments cells between the 3rd and 6th passage were used.
2.2. Real-time RT-PCR
To investigate the expression of VEGF-R2 and NRP-1 at the
mRNA level, RNA from HTF and HUVEC was isolated using the
RNeasy Minikit (Qiagen, Valencia, California), as described previ-
ously (Carmeliet et al., 2001). Expression was normalized to that
of the housekeeping gene b-actin. Primers and probes, designed
using Primer Express� 10 software, were described previously
(Brusselmans et al., 2005; Li et al., 2009).
2.3. Proliferation assay
To investigate the effect of the different VEGF isoforms and
pegaptanib, HUVEC, HTF and RTF were seeded in (0.1% gelatine
coated) 96-well plates (Falcon, Becton Dickinson Labware, Franklin
Lakes, New Jersey) at an initial density of 5 � 103 cells per well in
100 ml complete medium. In one series of experiments (6 h after
seeding), the cells were serum starved (medium supplementedwith
0.1% FBS) overnight. The medium was changed to fresh serum-free
medium containing 10, 25 and 100 ng/ml or 0.25, 1, 2, 5, 10 nM
recombinant human VEGF121 (4644-VS; R&D Systems, Minneapolis,
Minnesota), VEGF165 (293-VE; R&D Systems, Minneapolis, Minne-
sota) or VEGF189 (CRV114B, Cell Sciences, Canton, Massachusetts).
Forty-eight hours after administration, cell proliferation was
assessed using the WST-1 Cell Proliferation Assay System (Roche,
Mannheim, Germany). To examine whether inhibition of the ERK
pathway would exert any influence on proliferation of HTF and
HUVEC, cellswere pretreatedwith 20 mMPD98059 (an ERK pathway
inhibitor) 2 h prior to VEGF-treatments (25 ng/ml). Three hours after
T. Van Bergen et al. / Experimental Eye Research 93 (2011) 689e699690

VEGFadministration, cell proliferationwas assessed using theWST-1
Cell Proliferation Assay. Different time periods and concentrations
were chosen based on a study by Zhang et al. (Zhang et al., 2008),
shown to exert inhibitory effects on HUVEC proliferation.
In a second series, the medium was replaced by complete
medium, pre-incubated with different concentrations of pegapta-
nib (0, 0.03, 0.3, 1 and 2 mg/ml) or 10 mg/ml of bevacizumab, an
anti-VEGF antibody, known to exert pronounced inhibitory effects
on TF proliferation (Li et al., 2009). Of note, the pegaptanib
concentrations were based on those that are routinely used in
clinical practice for intravitreal injections, the highest concentra-
tion being the one that could be obtained by minimal dilution with
the required culture medium. Forty-eight hours after administra-
tion, cell proliferation was assessed using the WST-1 Cell Prolifer-
ation Assay. Complete or serum-free medium served as control.
2.4. Transcription factor activation assay
To clarify the differential effects elicited by VEGF121, VEGF165 and
VEGF189 on HUVEC and HTF, cells were serum starved overnight and
treated with the different isoforms (25 ng/ml) for 10 min, a concen-
tration and time period shown to exert the highest increase in acti-
vation of the MEK/ERK pathway in HUVEC (Zhang et al., 2008).
Serum-freemedium served as control. Nuclear protein extracts were
obtained from both cell types using a Nuclear Extract Kit (40010;
Active Motif, Rixensart, Belgium), as previously described (Akiyama
et al., 2004). Shortly, the cells were collected in ice-cold PBS in the
presence of phosphatase inhibitors. After resuspension in hypotonic
buffer, detergent was added to collect the cytoplasmic fraction.
Nuclei were lysed and nuclear proteins were solubilised in lysis
buffer. The protein concentration of the nuclear extract was
measured by Pierce bicinchoninic acid (BCA) protein assay kit
(23225, Thermo Scientific, Rockford, Illinois). ERK DNA binding
activity was assessed by the TransAM c-Myc assay, according the
manufacturer’s instructions (43396; Active Motif, Rixensart, Bel-
gium). The nuclear extract (5 mg) was subjected to binding of c-Myc
using an immobilized consensus sequence in a 96-well plate, and
primary and secondary antibodies were added. The colorimetric
reaction was analyzed using a spectrophotometer and DNA binding
specificitywas assessedusingmutated orwild-type oligonucleotides
(20 pmol/well). The wild-type consensus oligonucleotide prevented
c-Myc binding to the probe immobilized on the plate. Conversely, the
mutated consensus oligonucleotide had no effect on c-Myc binding.
2.5. Rabbit model of filtration surgery
Female New Zealand rabbits, 12e14 weeks of age and weighing
2e3 kg, were used. Before surgery, the intra-ocular pressure (IOP)
wasmeasured in both eyes with a tonometer (Tono-Pen;Medtronic
Solan, Jacksonville, Florida) under 4 mg/ml topical anaesthesia
(Unicain, Thea Pharma, Schaffhausen, Switzerland). Three record-
ings per eye were averaged. General anaesthesia was induced by
intramuscular injection of 50 mg/ml ketamine (Ketalar, Pfizer, Ann
Arbor, Michigan) and 2% sedative (Rompun, Bayer Health Care,
Pittsburgh, Pennsylvania). Surgery was performed on both eyes,
using a technique as previously described (Khaw et al., 1993).
In a group of 20 rabbits, one eye was injected with pegaptanib
(0.3 mg/90 ml; 3.3 mg/ml) immediately after surgery; the other eye
was used as a control and received an injection of 0.9% NaCl (Braun
Medical, Bethlehem, Pennsylvania). For each eye, 90 ml were
injected into the anterior chamber and 90 ml were injected sub-
conjunctivally into the filtration bleb. Rabbits were sacrificed at 3, 8,
14 and 30 days after surgery (n ¼ 5 per time point).
In another group of rabbits (n ¼ 12), repeated injections of
pegaptanib (0.3 mg/90 ml; 3.3 mg/ml) were given every 2 days
subconjunctivally until day 14 and animals were sacrificed 15 and
30 days after surgery (n ¼ 6 per time point).
2.6. Clinical examination and immunohistochemistry
All rabbits were clinically examined and the bleb vascularity
was graded according to the Moorfield bleb grading system
(Wells et al., 2004) on day 1 after surgery and then every 2 days
until the animals were sacrificed. The IOP, the bleb area (width
and length) and height were analyzed under topical anaesthesia
using callipers. Bleb survival was taken as the end-point of the
study; bleb failure was defined as the appearance of a scarred and
flat bleb.
On days 3, 8,14,15 and 30 after surgery, rabbits were killed using
a lethal intravenous injection of sedative. Both eyes were enucle-
ated, fixed overnight in 4% paraformaldehyde (PFA), dehydrated
and embedded in paraffin. Serial sections (7 mm) were immuno-
stained using a murine antibody against human CD31 (M0823;
DakoCytomation A/S, Copenhagen, Denmark) to check the density
of blood vessels and a murine antibody against rabbit CD45
(MC808G; AbD Serotec, Oxford, UK) to analyze inflammation, as
previously described (Li et al., 2009). Deposition of collagen was
analyzed by Sirius Red staining.
Images were obtained using a microscope (Leica Microsystems,
Wetzlar, Germany), equipped with a digital camera (Axiocam
MrC5; Carl Zeiss, Oberkochen, Germany), at a magnification of 20�
and a resolution of 2584 � 1936 pixels. Morphometric analyses
were performed using commercial software (KS300; Zeiss, Ober-
kochen, Germany). The density of blood vessels was determined by
calculating the CD31-positive area as a proportion of the total
wound area. The density of leukocytes was quantified by counting
the CD45-positive cells, expressed in number/mm2 of the total area
of wound healing. Deposition of collagen was determined by
measuring the percentage of collagen fibres in the total wound area
under polarized light.
2.7. Statistical analysis
All in vitro and immunomorphometric data were analyzed using
the Student’s t-test for independent samples. Data at individual
time points were analyzed usingmixedmodel analysis for repeated
measures (using SAS 9.2). KaplaneMeier survival analysis was
performed for bleb failure using the logrank test. P � 0.05 were
considered to be statistically significant. Data are represented as
mean � SEM.
3. Results
3.1. Expression of VEGF-R2 and NRP-1 on HTF and HUVEC
Quantitative real-time RT-PCR on HTF and HUVEC showed
expression of VEGF-R2 and NRP-1 on both cell types. The expres-
sion of VEGF-R2 mRNA in HUVEC was 20 times higher as compared
to HTF (P ¼ 0.02). On the other hand, a 1.3 fold higher expression of
NRP-1 was found on HUVEC as compared to HTF (P¼ 0.04; data not
shown).
3.2. Effect of VEGF isoform administration on proliferation
of HUVEC and TF
To analyze the effect of the VEGF isoforms on proliferation of
HUVEC (as a measure of an angiogenic response) and of HTF and
RTF (as a measure of a fibrotic response), cells were grown in
serum-free medium in the presence of increasing concentrations
of the isoforms. Administration of VEGF121 induced a significant
T. Van Bergen et al. / Experimental Eye Research 93 (2011) 689e699 691
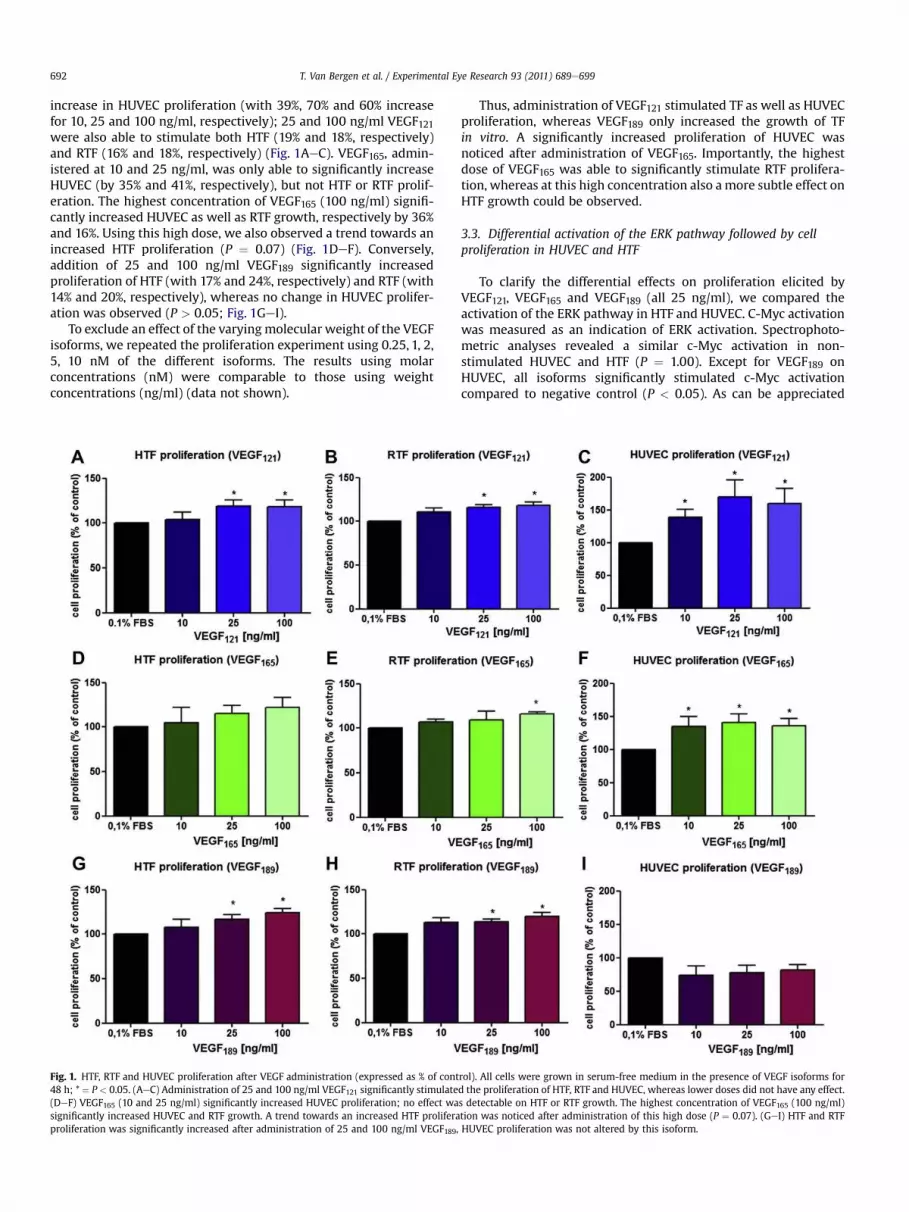
increase in HUVEC proliferation (with 39%, 70% and 60% increase
for 10, 25 and 100 ng/ml, respectively); 25 and 100 ng/ml VEGF121were also able to stimulate both HTF (19% and 18%, respectively)
and RTF (16% and 18%, respectively) (Fig. 1AeC). VEGF165, admin-
istered at 10 and 25 ng/ml, was only able to significantly increase
HUVEC (by 35% and 41%, respectively), but not HTF or RTF prolif-
eration. The highest concentration of VEGF165 (100 ng/ml) signifi-
cantly increased HUVEC as well as RTF growth, respectively by 36%
and 16%. Using this high dose, we also observed a trend towards an
increased HTF proliferation (P ¼ 0.07) (Fig. 1DeF). Conversely,
addition of 25 and 100 ng/ml VEGF189 significantly increased
proliferation of HTF (with 17% and 24%, respectively) and RTF (with
14% and 20%, respectively), whereas no change in HUVEC prolifer-
ation was observed (P > 0.05; Fig. 1GeI).
To exclude an effect of the varyingmolecular weight of the VEGF
isoforms, we repeated the proliferation experiment using 0.25, 1, 2,
5, 10 nM of the different isoforms. The results using molar
concentrations (nM) were comparable to those using weight
concentrations (ng/ml) (data not shown).
Thus, administration of VEGF121 stimulated TF as well as HUVEC
proliferation, whereas VEGF189 only increased the growth of TF
in vitro. A significantly increased proliferation of HUVEC was
noticed after administration of VEGF165. Importantly, the highest
dose of VEGF165 was able to significantly stimulate RTF prolifera-
tion, whereas at this high concentration also amore subtle effect on
HTF growth could be observed.
3.3. Differential activation of the ERK pathway followed by cell
proliferation in HUVEC and HTF
To clarify the differential effects on proliferation elicited by
VEGF121, VEGF165 and VEGF189 (all 25 ng/ml), we compared the
activation of the ERK pathway in HTF and HUVEC. C-Myc activation
was measured as an indication of ERK activation. Spectrophoto-
metric analyses revealed a similar c-Myc activation in non-
stimulated HUVEC and HTF (P ¼ 1.00). Except for VEGF189 on
HUVEC, all isoforms significantly stimulated c-Myc activation
compared to negative control (P < 0.05). As can be appreciated
Fig. 1. HTF, RTF and HUVEC proliferation after VEGF administration (expressed as % of control). All cells were grown in serum-free medium in the presence of VEGF isoforms for
48 h; * ¼ P < 0.05. (AeC) Administration of 25 and 100 ng/ml VEGF121 significantly stimulated the proliferation of HTF, RTF and HUVEC, whereas lower doses did not have any effect.
(DeF) VEGF165 (10 and 25 ng/ml) significantly increased HUVEC proliferation; no effect was detectable on HTF or RTF growth. The highest concentration of VEGF165 (100 ng/ml)
significantly increased HUVEC and RTF growth. A trend towards an increased HTF proliferation was noticed after administration of this high dose (P ¼ 0.07). (GeI) HTF and RTF
proliferation was significantly increased after administration of 25 and 100 ng/ml VEGF189, HUVEC proliferation was not altered by this isoform.
T. Van Bergen et al. / Experimental Eye Research 93 (2011) 689e699692

from Table 1, VEGF121 activated c-Myc (and ERK pathway) in HTF
and HUVEC in a similar way (P ¼ 0.37). In contrast, administration
of VEGF165 resulted in a significantly higher c-Myc activation level
in HUVEC, as compared to HTF (P ¼ 0.02), whereas c-Myc was
significantly more activated in HTF as compared to HUVEC after
VEGF189 administration (P ¼ 0.01; Table 1).
Thus, VEGF121 increased the activation of the ERK pathway in
both HTF and HUVEC, whereas VEGF165 and VEGF189 mainly
induced ERK activation in, respectively HUVEC and HTF, at least as
shown via activation of c-Myc.
3.4. Effect of ERK pathway inhibition (PD98059) on VEGF-induced
cell proliferation
To study the effect of ERK-inhibition on VEGF isoform induced
proliferation, cells were pretreated with PD98059 2 h prior to
VEGF-treatments. Three hours after administration, VEGF121 and
VEGF189 stimulated HTF growth, compared to control (with 3% and
21%, respectively), which was significantly reduced by PD98059
(with 22% and 25%, respectively; data not shown). On the other
hand, HUVEC proliferationwas significantly increased after VEGF121and VEGF165 administration (with 8% and 20%, respectively), which
was significantly inhibited after addition of the ERK-inhibitor (with
21% and 24%, respectively; data not shown).
Thus, VEGF121 and VEGF189 induced HTF proliferation was
significantly reduced by PD98059, whereas the ERK pathway
inhibitor significantly inhibited HUVEC proliferation, induced by
VEGF121 and VEGF165.
3.5. Effect of pegaptanib administration on proliferation
of HUVEC and TF
To evaluate the effect of specific inhibition of VEGF165,
increasing concentrations of pegaptanib (0, 0.03, 0.3, 1 and 2 mg/
ml) were added to HUVEC, HTF and RTF, all growing in complete
medium. A dose-dependent reduction of HUVEC proliferation was
seen 48 h after pegaptanib administration, as compared to
complete medium (P < 0.05 for all concentrations from 0.03 mg/ml
onwards; Fig. 2A). A concentration of 2 mg/ml was necessary to
inhibit the proliferation of HTF (P ¼ 0.009). Comparable to HTF,
2 mg/ml pegaptanib significantly inhibited RTF proliferation
(P ¼ 0.001). Also 1 mg/ml of the aptamer was able to reduce the
growth of RTF (P¼ 0.002), but not of HTF (Fig. 2B, C). In all cases, the
highest dose of pegaptanib (2 mg/ml) inhibited HUVEC, HTF and
RTF to a similar extent as 10 mg/ml bevacizumab (Fig. 2).
Thus, proliferation of HUVEC was significantly inhibited by
pegaptanib in a dose-dependent manner, whereas HTF growth was
only significantly reduced at the highest dose (2 mg/ml). Prolifer-
ation of RTF was reduced already after administration of 1 mg/ml of
the aptamer.
3.6. Effects of pegaptanib in a rabbit trabeculectomy model
To verify the role and relative contribution of VEGF165 in scar
formation after filtration surgery and to determine whether the
VEGF165 selective inhibitor, pegaptanib, is able to improve glau-
coma surgery outcome in vivo, one group of rabbits received one
single injection immediately after surgery. Another group was
treated with repeated injections of pegaptanib every 2 days sub-
conjunctivally until day 14. IOP, bleb area, height and survival were
analyzed as a measure of filtration surgery outcome. A single
injection or repeated injections of pegaptanib did not influence the
IOP; in both groups the pressure was similar in the treated and
control eyes over 30 days (Fig. 4AeE).
The typical appearance of the blebs after a single injection of
pegaptanib on different time points after surgery is shown in
Fig. 3A. One single pegaptanib-injection was associated with an
elevated bleb in the initial days after surgery (bottom panels)
compared to a flat and scarred bleb in the control group (top
panels). A single injection was able to improve the outcome after
surgery by increasing bleb area until day 9 (n ¼ 20; P ¼ 0.0009;
Fig. 4B) and by increasing the bleb height immediately after surgery
(n ¼ 20; P ¼ 0.09; Fig. 4C). No significant differences were seen in
bleb survival (n ¼ 20; P ¼ 0.83; Fig. 4D). (Immuno-)histological
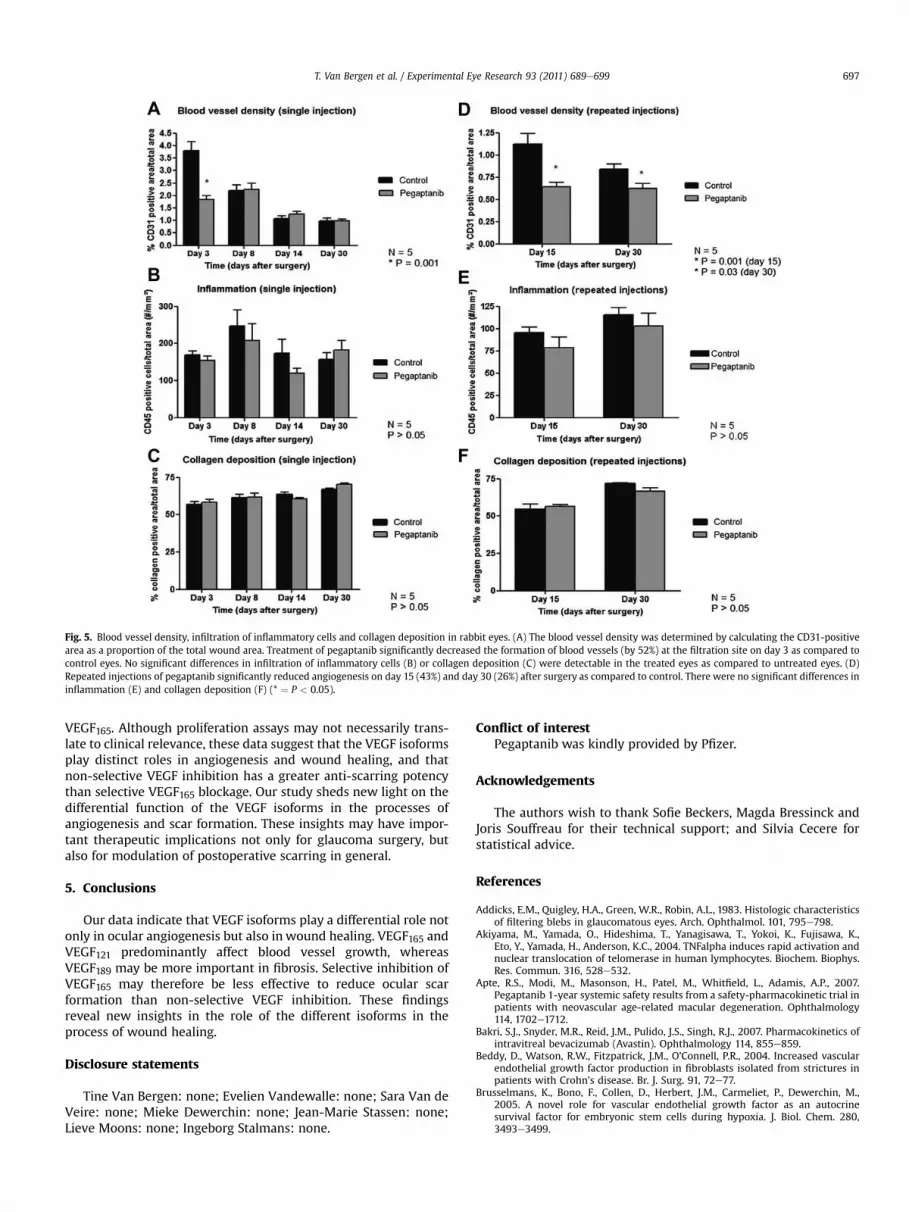
stainings showed a significant reduction of 52% in blood vessel
density in the treated eyes compared to controls, at 3 days after
surgery (n ¼ 5; P ¼ 0.001; Fig. 5A). These results were in keeping
with the Moorfields’ scoring of the blebs during follow-up. After
scoring, a significant reduction in vascularity was seen as well at
postoperative day 3, but not at day 30 (data not shown). Therewere
no significant differences in inflammatory responses (number of
infiltrating leukocytes) and collagen deposition (n ¼ 5; Fig. 5B, C).
In the group with repeated injections, two rabbits died unex-
pectedly during the experiment (one on day 9 and another on day
19 after surgery). It was not clear whether this occurred due to the
treatment or the anaesthesia. In the remaining 10 rabbits, repeated
injections were able to significantly improve the bleb area up until
15 days after surgery (n ¼ 10; P ¼ 0.0001; Fig. 4F). Bleb height was
increased a few days after surgery (n ¼ 10; P ¼ 0.10; Fig. 4G).
Repeated injections significantly prolonged bleb survival after
filtration surgery, as shown in the KaplaneMeier survival curve
(n¼ 10; P¼ 0.05; Fig. 4H). Fig. 3B shows the appearance of the blebs
after repeated pegaptanib-injections (bottom panels) compared to
control eyes (top panels) at different time points. Angiogenesis was
significantly reduced by 43% on day 15 (n ¼ 5; P ¼ 0.001) and by
26% on day 30 (n ¼ 5; P ¼ 0.03) after surgery, as compared to
control conditions (Fig. 5D). Therewere no significant differences in
inflammatory responses and collagen deposition (n ¼ 5; Fig. 5E, F).
Thus, a single administration of pegaptanib at the time of tra-
beculectomy reduced postoperative angiogenesis during the initial
phase, but not inflammation or collagen deposition. Repeated
injections of pegaptanib improved surgery outcome by improving
bleb survival and by reducing angiogenesis up to at least 30 days
after surgery.
4. Discussion
Filtering surgery is the most effective treatment for glaucoma,
and is therefore a crucial procedure in the management of this
blinding disease (Lama and Fechtner, 2003). Unfortunately, exces-
sive postoperative wound healing with subsequent scarring
frequently leads to surgical failure (Addicks et al., 1983). Perioper-
ative anti-mitotics, such as mitomycin-C (MMC) and 5-Fluorouracil
(5-FU) can improve surgical outcome (Katz et al., 1995), but these
antimetabolites carry a risk of vision-threatening complications,
such as scleral thinning and infections (Gillies and Su, 1991;
Greenfield et al., 1998; Hitchings and Grierson, 1983; Jampel
et al., 1988; Katz et al., 1995; Lama and Fechtner, 2003; Skuta and
Parrish, 1987). Furthermore, blocking TGF-b, which seemed prom-
ising in animal models (Cordeiro et al., 2003), was not efficient in
Table 1
Activation of c-Myc in HTF and HUVEC, as an indication of ERK activation. Data are
represented as mean � SEM.
C-Myc activation
(OD at 450 nm)
HTF HUVEC P-value
(HTF vs HUVEC)
0.1% FBS 0.32 � 0.04 0.32 � 0.01 1.00
VEGF121 0.44 � 0.03 0.51 � 0.08 0.37
VEGF165 0.44 � 0.03 0.67 � 0.11 0.02
VEGF189 0.47 � 0.03 0.30 � 0.01 0.01
T. Van Bergen et al. / Experimental Eye Research 93 (2011) 689e699 693

a clinical study (Khaw et al., 2007). Therefore, there is still a need
for alternative strategies to prevent filtration failure.
Wound healing occurs through a combination of four processes:
clot formation, angiogenesis, inflammation and collagen deposition
(Addicks et al., 1983; Gillies and Su, 1991; Lama and Fechtner, 2003;
Lee et al., 1995). VEGF is a well-known key player in angiogenesis
(Carmeliet et al., 1996; Dvorak et al., 1999; Kowanetz and Ferrara,
2006). An additional role in inflammation and fibrosis has been
suggested, but has been studied much less extensively (Beddy et al.,
2004;Murakami et al., 2006;Wilgus et al., 2008). AsVEGFmight play
a role in several processes of wound healing (Li and Li, 2003; Nissen
et al., 1998; Wilgus et al., 2008), we hypothesized that this growth
factor might be a good target candidate for anti-fibrotic adjunctive
strategies in glaucoma surgery. Indeed, we recently published
encouraging in vitro and in vivo data indicating that VEGF as well as
its receptors are expressed by Tenon fibroblasts. Moreover, we
showed that non-selective VEGF inhibition by bevacizumab blocks
fibroblast proliferation in vitro, aswell as postoperative angiogenesis
and fibrosis after filtering surgery in vivo (Li et al., 2009). In addition,
Memerzadeh et al. showed that repeated injections of bevacizumab
were able to reduce both vascularity andfibrosis in a rabbitmodel for
trabeculectomy (Memarzadeh et al., 2009). These experiments were
performed with the non-isoform selective anti-VEGF antibody bev-
acizumab. However, other anti-VEGF treatments are used in the
ophthalmological practice, some of which have more selective
properties. Importantly, we previously reported that various VEGF
isoforms exert a different function during physiological angiogenesis
(Stalmans et al., 2003, 2002). Their differential role in pathological
angiogenesis has also been studied, but unfortunately still remains
rather controversial (Ishida et al., 2003b; Usui et al., 2004).
Furthermore, the specific role of the VEGF isoforms in inflammation
(Ishida et al., 2003a) and fibrosis (Roberts et al., 2007) remains
largely unknown.
Using in vitro experiments, the current study revealed that
VEGF121 and VEGF165 significantly stimulate proliferation of
HUVEC, by activating the ERK pathway. Moreover, PD98059, an
inhibitor of the ERK pathway, significantly inhibited VEGF121 and
VEGF165-induced HUVEC proliferation. These data are in keeping
with the observations by Zhang et al. (Zhang et al., 2008), who
recently reported that VEGF165, and to a lesser extent VEGF121,
stimulated HUVEC proliferation. These authors showed that
VEGF165 significantly increased the level of phosphorylation in the
MEK/ERK pathway, whereas VEGF121 had little effect. For the first
time, we report here the effect of the different isoforms on TF
growth. We showed that TF proliferation was mainly induced by
VEGF121 and VEGF189, associated with an activation of the ERK
pathway. These proliferative effects were significantly reduced by
the ERK pathway inhibitor PD98059. Also at the highest concen-
tration, VEGF165 was able to significantly increase RTF prolifera-
tion, whereas a subtle effect of high VEGF165 levels on HTF could
not be excluded. Our results indicate that VEGF165 is the most
important isoform in inducing HUVEC proliferation, while VEGF189is the most prominent in inducing TF proliferation. VEGF121 clearly
affects both cell types. Additionally, we demonstrated that HUVEC
were effectively inhibited by pegaptanib (a selective VEGF165-
inhibitor) in a dose-dependent way. HTF growth was only signif-
icantly reduced using the highest dose of this selective inhibitor.
Proliferation of RTF was already reduced after administration of
lower doses of the aptamer. Our findings thus indicate that HTF
might be somewhat less sensitive compared to RTF as more
VEGF165/pegaptanib is required for a significant HTF stimulation/
inhibition.
Fig. 2. HUVEC, HTF and RTF proliferation after pegaptanib administration (expressed as % of control). All cells were grown in complete medium in the presence of pegaptanib for
48 h; * ¼ P < 0.05. (A) Pegaptanib significantly inhibited the proliferation of HUVEC in a dose-dependent manner, at a minimal dose of 0.03 mg/ml. (B) A concentration of 2 mg/ml
was necessary to inhibit the proliferation of HTF (P ¼ 0.009). (C) Comparable to HTF, 2 mg/ml pegaptanib inhibited RTF proliferation significantly (P ¼ 0.001). Furthermore, even
1 mg/ml of the aptamer was able to reduce the growth of RTF (P ¼ 0.002). In all cases, the highest dose of pegaptanib (2 mg/ml) inhibited HUVEC, HTF and RTF to a similar extent as
10 mg/ml bevacizumab.
T. Van Bergen et al. / Experimental Eye Research 93 (2011) 689e699694

The functional difference of the various isoforms on HTF and
HUVEC proliferation can be attributed to different expression
levels of VEGF-R2 and/or VEGF-R co-receptors on HTF and
HUVEC. Both HTF (Li et al., 2009) and HUVEC (Castro-Rivera et al.,
2004) are known to express VEGF-R2 but, of note, a lower
expression of this receptor on synovial fibroblasts as compared to
HUVEC has been reported (Kim et al., 2006). Our real-time RT-
PCR analyses indeed revealed that the expression of VEGF-R2
mRNA on HUVEC was 20 times higher as compared to HTF.
Additionally, HUVEC (Castro-Rivera et al., 2004; Deroanne et al.,
2002; Kokolakis et al., 2006; Soker et al., 1998; Tirand et al.,
2006) and certain types of fibroblasts (Haag et al., 2010; Kim
et al., 2006) were also reported to express NRP-1, but whether
NRP-1 is expressed on Tenon fibroblasts remained unknown. Our
RT-PCR data showed that HTF indeed express NRP-1, but a lower
level as compared to HUVEC.
Our expression data may explain the difference in activation of
both cell types by the various VEGF isoforms. Indeed, VEGF121,
lacking the VEGF-R co-receptor binding domains (Robinson and
Stringer, 2001), exerts proliferative effects on both cell types by
binding VEGF-R2. However, similar concentrations of VEGF121 seem
to stimulate HUVEC more than TF, probably because of the lower
expression of VEGF-R2 on HTF. VEGF165 binds VEGF-R2 but also
strongly binds NRP-1. As such, the reduced expression of VEGF-R2,
and possibly of NRP-1, on HTF may explain why VEGF165 seems to
be more potent in inducing proliferation of HUVEC as compared to
HTF. In comparison to VEGF121 and VEGF165, VEGF189 strongly binds
HSPGs and is believed to constitute a reservoir of growth factor,
which can be slowly released by exposure to urokinase plasmin-
ogen activator (uPA) (Robinson and Stringer, 2001). Indeed, protein
folding of this larger isoform obscures regions responsible for
receptor binding and therefore native VEGF189 is unable to bind
VEGF-R2 and NRP-1 (Plouet et al., 1997). Cleavage of VEGF189 by uPA
is required for its receptor binding and biological actions. Impor-
tantly, zymographic analyses did reveal the presence of uPA in cell
lysates of conjunctival fibroblasts (Chao et al., 2011), but not of
HUVEC (Herve et al., 2005). These findings may explain our
observation that VEGF189 could stimulate HTF, but not HUVEC,
however, further research is required. Moreover, we cannot exclude
that the observed VEGF isoform specific effects on the two cell
types may also be explained by a direct or indirect activation of
different signal transduction pathways upon binding VEGF-R2.
Based on the in vitro results we hypothesized that, whereas we
previously published that non-selective VEGF inhibition is anti-
angiogenic as well as anti-fibrotic (Li et al., 2009), selective
VEGF165 blockage might mainly affect angiogenesis. Our in vivo
surgery experiments indeed confirmed that a single administration
of pegaptanib significantly impaired angiogenesis, but did not
reduce collagen deposition. Of note, neither selective nor non-
selective VEGF inhibition could reduce the amount of post-
operative inflammatory responses inour trabeculectomymodel (this
study and Li et al., 2009). Besides a difference in isoform selectivity,
the greater potency of bevacizumab compared to pegaptanib to
inhibit postoperative scarring after a single peroperative adminis-
trationmight be attributed to a difference in time of action between
both inhibitors. Indeed, the half-life of intra-ocular bevacizumab
Fig. 3. Macroscopic postoperative photographs of eyes after surgery. Arrows: edges of the blebs. (A) Macroscopic photographs of eyes after a single pegaptanib-injection show
surviving blebs that remain diffusely elevated (bottom) compared to control (top) until day 8. At day 14 the blebs were comparable. (B) Repeated injections of pegaptanib were
associated with an elevated bleb until day 15 after surgery (bottom) compared to a flat and scarred bleb in the control group (top).
T. Van Bergen et al. / Experimental Eye Research 93 (2011) 689e699 695

(4.88 days) (Bakri et al., 2007) was shown to be significantly longer
than that of pegaptanib (3.91 days) (Apte et al., 2007). To further
elucidate whether the observed difference in anti-fibrotic efficacy
was caused by a difference in half-life, we extended the treatment by
administering additional repeated postoperative injections of
pegaptanib after the initial peroperative treatment. Although the
effect on postoperative outcome slightly improved, this effect was
still solely attributable to the anti-angiogenic properties of the
aptamer and not to its effects in collagen deposition. In both groups
(single and repeated injections of pegaptanib) no significant changes
in IOP compared to placebo were observed. However, this is in
keeping with the previous literature describing this model for sub-
conjunctival scarring (Cordeiro et al., 2003; Mead et al., 2003;
Memarzadeh et al., 2009), and can be explained by the fact that
the surgery is performed on non-glaucomatous rabbit eyes with
normal IOPs at baseline.
In summary, our in vitro and in vivo data indicate that selective
VEGF165 inhibition by pegaptanib is less effective than non-
selective VEGF inhibition by bevacizumab (Li et al., 2009) in
reducing scarring after glaucoma surgery, presumably due to
retained action of VEGF121 and VEGF189 isoforms, which have
a more pronounced effect on the Tenon fibroblasts as compared to
Fig. 4. Clinical investigation of eyes after surgery and pegaptanib administration. (A) A single injection or (E) repeated injections of pegaptanib did not influence the IOP; in both
groups the pressure was similar in the treated and the control eyes over 30 days. (B) A single injection of pegaptanib significantly improved the outcome of the surgery by increasing
the bleb area until day 9 (P ¼ 0.0009) and (C) by increasing the bleb height immediately after surgery (P ¼ 0.09). (D) No significant differences were seen in bleb survival (P ¼ 0.83).
(F) Repeated pegaptanib-injections significantly improved the bleb area up until 15 days after surgery but not thereafter (P ¼ 0.0001). (G) Bleb height was increased a few days after
surgery (P ¼ 0.10). (H) As shown in the KaplaneMeier survival curve, repeated injections significantly prolonged bleb survival after filtration surgery, compared to control (P ¼ 0.05).
T. Van Bergen et al. / Experimental Eye Research 93 (2011) 689e699696
VEGF165. Although proliferation assays may not necessarily trans-
late to clinical relevance, these data suggest that the VEGF isoforms
play distinct roles in angiogenesis and wound healing, and that
non-selective VEGF inhibition has a greater anti-scarring potency
than selective VEGF165 blockage. Our study sheds new light on the
differential function of the VEGF isoforms in the processes of
angiogenesis and scar formation. These insights may have impor-
tant therapeutic implications not only for glaucoma surgery, but
also for modulation of postoperative scarring in general.
5. Conclusions
Our data indicate that VEGF isoforms play a differential role not
only in ocular angiogenesis but also in wound healing. VEGF165 and
VEGF121 predominantly affect blood vessel growth, whereas
VEGF189 may be more important in fibrosis. Selective inhibition of
VEGF165 may therefore be less effective to reduce ocular scar
formation than non-selective VEGF inhibition. These findings
reveal new insights in the role of the different isoforms in the
process of wound healing.
Disclosure statements
Tine Van Bergen: none; Evelien Vandewalle: none; Sara Van de
Veire: none; Mieke Dewerchin: none; Jean-Marie Stassen: none;
Lieve Moons: none; Ingeborg Stalmans: none.
Conflict of interest
Pegaptanib was kindly provided by Pfizer.
Acknowledgements
The authors wish to thank Sofie Beckers, Magda Bressinck and
Joris Souffreau for their technical support; and Silvia Cecere for
statistical advice.
References
Addicks, E.M., Quigley, H.A., Green, W.R., Robin, A.L., 1983. Histologic characteristicsof filtering blebs in glaucomatous eyes. Arch. Ophthalmol. 101, 795e798.
Akiyama, M., Yamada, O., Hideshima, T., Yanagisawa, T., Yokoi, K., Fujisawa, K.,Eto, Y., Yamada, H., Anderson, K.C., 2004. TNFalpha induces rapid activation andnuclear translocation of telomerase in human lymphocytes. Biochem. Biophys.Res. Commun. 316, 528e532.
Apte, R.S., Modi, M., Masonson, H., Patel, M., Whitfield, L., Adamis, A.P., 2007.Pegaptanib 1-year systemic safety results from a safety-pharmacokinetic trial inpatients with neovascular age-related macular degeneration. Ophthalmology114, 1702e1712.
Bakri, S.J., Snyder, M.R., Reid, J.M., Pulido, J.S., Singh, R.J., 2007. Pharmacokinetics ofintravitreal bevacizumab (Avastin). Ophthalmology 114, 855e859.
Beddy, D., Watson, R.W., Fitzpatrick, J.M., O’Connell, P.R., 2004. Increased vascularendothelial growth factor production in fibroblasts isolated from strictures inpatients with Crohn’s disease. Br. J. Surg. 91, 72e77.
Brusselmans, K., Bono, F., Collen, D., Herbert, J.M., Carmeliet, P., Dewerchin, M.,2005. A novel role for vascular endothelial growth factor as an autocrinesurvival factor for embryonic stem cells during hypoxia. J. Biol. Chem. 280,3493e3499.
Fig. 5. Blood vessel density, infiltration of inflammatory cells and collagen deposition in rabbit eyes. (A) The blood vessel density was determined by calculating the CD31-positive
area as a proportion of the total wound area. Treatment of pegaptanib significantly decreased the formation of blood vessels (by 52%) at the filtration site on day 3 as compared to
control eyes. No significant differences in infiltration of inflammatory cells (B) or collagen deposition (C) were detectable in the treated eyes as compared to untreated eyes. (D)
Repeated injections of pegaptanib significantly reduced angiogenesis on day 15 (43%) and day 30 (26%) after surgery as compared to control. There were no significant differences in
inflammation (E) and collagen deposition (F) (* ¼ P < 0.05).
T. Van Bergen et al. / Experimental Eye Research 93 (2011) 689e699 697

Carmeliet, P., Ferreira, V., Breier, G., Pollefeyt, S., Kieckens, L., Gertsenstein, M.,Fahrig, M., Vandenhoeck, A., Harpal, K., Eberhardt, C., Declercq, C., Pawling, J.,Moons, L., Collen, D., Risau, W., Nagy, A., 1996. Abnormal blood vessel devel-opment and lethality in embryos lacking a single VEGF allele. Nature 380,435e439.
Carmeliet, P., Moons, L., Luttun, A., Vincenti, V., Compernolle, V., De Mol, M., Wu, Y.,Bono, F., Devy, L., Beck, H., Scholz, D., Acker, T., DiPalma, T., Dewerchin,M., Noel, A.,Stalmans, I., Barra, A., Blacher, S., Vandendriessche, T., Ponten, A., Eriksson, U.,Plate, K.H., Foidart, J.M., Schaper, W., Charnock-Jones, D.S., Hicklin, D.J.,Herbert, J.M., Collen, D., Persico, M.G., 2001. Synergism between vascular endo-thelial growth factor and placental growth factor contributes to angiogenesis andplasma extravasation in pathological conditions. Nat. Med. 7, 575e583.
Castro-Rivera, E., Ran, S., Thorpe, P., Minna, J.D., 2004. Semaphorin 3B (SEMA3B)induces apoptosis in lung and breast cancer, whereas VEGF165 antagonizes thiseffect. Proc. Natl. Acad. Sci. U. S. A. 101, 11432e11437.
Chao, S.C., Hu, D.N., Yang, P.Y., Lin, C.Y., Yang, S.F., 2011. Overexpression of urokinase-type plasminogen activator in pterygia and pterygium fibroblasts. Mol. Vis. 17,23e31.
Ciulla, T.A., Rosenfeld, P.J., 2009. Antivascular endothelial growth factor therapy forneovascularage-relatedmaculardegeneration.Curr.Opin.Ophthalmol.20,158e165.
Cordeiro, M.F., Mead, A., Ali, R.R., Alexander, R.A., Murray, S., Chen, C., York-Defalco, C., Dean, N.M., Schultz, G.S., Khaw, P.T., 2003. Novel antisense oligo-nucleotides targeting TGF-beta inhibit in vivo scarring and improve surgicaloutcome. Gene Ther. 10, 59e71.
Deroanne, C.F., Bonjean, K., Servotte, S., Devy, L., Colige, A., Clausse, N., Blacher, S.,Verdin, E., Foidart, J.M., Nusgens, B.V., Castronovo, V., 2002. Histone deacety-lases inhibitors as anti-angiogenic agents altering vascular endothelial growthfactor signaling. Oncogene 21, 427e436.
Dvorak, H.F., Nagy, J.A., Feng, D., Brown, L.F., Dvorak, A.M., 1999. Vascular perme-ability factor/vascular endothelial growth factor and the significance ofmicrovascular hyperpermeability in angiogenesis. Curr. Top. Microbiol. Immu-nol. 237, 97e132.
Ferrara, N., Gerber, H.P., LeCouter, J., 2003. The biology of VEGF and its receptors.Nat. Med. 9, 669e676.
Gillies, M.C., Su, T., 1991. Cytokines, fibrosis and the failure of glaucoma filtrationsurgery. Aust. N. Z. J. Ophthalmol. 19, 299e304.
Greenfield, D.S., Liebmann, J.M., Jee, J., Ritch, R., 1998. Late-onset bleb leaks afterglaucoma filtering surgery. Arch. Ophthalmol. 116, 443e447.
Haag, M., Van Linthout, S., Schroder, S.E., Freymann, U., Ringe, J., Tschope, C.,Sittinger, M., 2010. Endomyocardial biopsy derived adherent proliferatingcells e a potential cell source for cardiac tissue engineering. J. Cell Biochem. 109,564e575.
Herve, M.A., Buteau-Lozano, H., Mourah, S., Calvo, F., Perrot-Applanat, M., 2005.VEGF189 stimulates endothelial cells proliferation and migration in vitro andup-regulates the expression of Flk-1/KDR mRNA. Exp. Cell Res. 309, 24e31.
Hitchings, R.A., Grierson, I., 1983. Clinico pathological correlation in eyes with failedfistulizing surgery. Trans. Ophthalmol. Soc. U. K. 103 (Pt 1), 84e88.
Ishida, S., Usui, T., Yamashiro, K., Kaji, Y., Ahmed, E., Carrasquillo, K.G., Amano, S.,Hida, T., Oguchi, Y., Adamis, A.P., 2003a. VEGF164 is proinflammatory in thediabetic retina. Invest. Ophthalmol. Vis. Sci. 44, 2155e2162.
Ishida, S., Usui, T., Yamashiro, K., Kaji, Y., Amano, S., Ogura, Y., Hida, T., Oguchi, Y.,Ambati, J., Miller, J.W., Gragoudas, E.S., Ng, Y.S., D’Amore, P.A., Shima, D.T.,Adamis, A.P., 2003b. VEGF164-mediated inflammation is required for patho-logical, but not physiological, ischemia-induced retinal neovascularization.J. Exp. Med. 198, 483e489.
Jampel, H.D., McGuigan, L.J., Dunkelberger, G.R., L’Hernault, N.L., Quigley, H.A., 1988.Cellular proliferation after experimental glaucoma filtration surgery. Arch.Ophthalmol. 106, 89e94.
Katz, G.J., Higginbotham, E.J., Lichter, P.R., Skuta, G.L., Musch, D.C., Bergstrom, T.J.,Johnson, A.T., 1995. Mitomycin C versus 5-fluorouracil in high-risk glaucomafiltering surgery. Extended follow-up. Ophthalmology 102, 1263e1269.
Khaw, P., Grehn, F., Hollo, G., Overton, B., Wilson, R., Vogel, R., Smith, Z., 2007.A phase III study of subconjunctival human anti-transforming growth factorbeta(2) monoclonal antibody (CAT-152) to prevent scarring after first-timetrabeculectomy. Ophthalmology 114, 1822e1830.
Khaw, P.T., Doyle, J.W., Sherwood, M.B., Grierson, I., Schultz, G., McGorray, S., 1993.Prolonged localized tissue effects from 5-minute exposures to fluorouracil andmitomycin C. Arch. Ophthalmol. 111, 263e267.
Kim, W.U., Kang, S.S., Yoo, S.A., Hong, K.H., Bae, D.G., Lee, M.S., Hong, S.W.,Chae, C.B., Cho, C.S., 2006. Interaction of vascular endothelial growth factor165 with neuropilin-1 protects rheumatoid synoviocytes from apoptoticdeath by regulating Bcl-2 expression and Bax translocation. J. Immunol. 177,5727e5735.
Kokolakis, G., Mikelis, C., Papadimitriou, E., Courty, J., Karetsou, E., Katsoris, P., 2006.Effect of heparin affin regulatory peptide on the expression of vascular endo-thelial growth factor receptors in endothelial cells. In Vivo 20, 629e635.
Kowanetz, M., Ferrara, N., 2006. Vascular endothelial growth factor signalingpathways: therapeutic perspective. Clin. Cancer Res. 12, 5018e5022.
Lama, P.J., Fechtner, R.D., 2003. Antifibrotics and wound healing in glaucomasurgery. Surv. Ophthalmol. 48, 314e346.
Lee, P.F., Myers, K.S., Hsieh, M.M., Wood, E.F., Hotaling, D.M., 1995. Treatment offailing glaucoma filtering cystic blebs with tissue plasminogen activator (tPA).J. Ocul. Pharmacol. Ther. 11, 227e232.
Li, W.W., Li, V.W., 2003. Therapeutic angiogenesis for wound healing. Wounds e ACompend. Clin. Res. Pract. 15, 3Se12S.
Li, Z., Van Bergen, T., Van de Veire, S., Van de Vel, I., Moreau, H., Dewerchin, M.,Maudgal, P.C., Zeyen, T., Spileers, W., Moons, L., Stalmans, I., 2009. Inhibition ofvascular endothelial growth factor reduces scar formation after glaucomafiltration surgery. Invest. Ophthalmol. Vis. Sci. 50, 5217e5225.
Mead, A.L., Wong, T.T., Cordeiro, M.F., Anderson, I.K., Khaw, P.T., 2003. Evaluation ofanti-TGF-beta2 antibody as a new postoperative anti-scarring agent in glau-coma surgery. Invest. Ophthalmol. Vis. Sci. 44, 3394e3401.
Memarzadeh, F., Varma, R., Lin, L.T., Parikh, J.G., Dustin, L., Alcaraz, A., Eliott, D.,2009. Postoperative use of bevacizumab as an antifibrotic agent in glaucomafiltration surgery in the rabbit. Invest. Ophthalmol. Vis. Sci. 50, 3233e3237.
Murakami, M., Iwai, S., Hiratsuka, S., Yamauchi, M., Nakamura, K., Iwakura, Y.,Shibuya, M., 2006. Signaling of vascular endothelial growth factor receptor-1tyrosine kinase promotes rheumatoid arthritis through activation of mono-cytes/macrophages. Blood 108, 1849e1856.
Nissen, N.N., Polverini, P.J., Koch, A.E., Volin, M.V., Gamelli, R.L., DiPietro, L.A., 1998.Vascular endothelial growth factor mediates angiogenic activity during theproliferative phase of wound healing. Am. J. Pathol. 152, 1445e1452.
Olsson, A.K., Dimberg, A., Kreuger, J., Claesson-Welsh, L., 2006. VEGF receptor sig-nalling e in control of vascular function. Nat. Rev. Mol. Cell Biol. 7, 359e371.
Pap, T., Nawrath, M., Heinrich, J., Bosse, M., Baier, A., Hummel, K.M., Petrow, P.,Kuchen, S., Michel, B.A., Gay, R.E., Muller-Ladner, U., Moelling, K., Gay, S., 2004.Cooperation of Ras- and c-Myc-dependent pathways in regulating the growthand invasiveness of synovial fibroblasts in rheumatoid arthritis. ArthritisRheum. 50, 2794e2802.
Pintus, G., Tadolini, B., Posadino, A.M., Sanna, B., Debidda, M., Bennardini, F., Sava, G.,Ventura, C., 2002. Inhibition of the MEK/ERK signaling pathway by the novelantimetastatic agent NAMI-A down regulates c-myc gene expression andendothelial cell proliferation. Eur. J. Biochem. 269, 5861e5870.
Plouet, J., Moro, F., Bertagnolli, S., Coldeboeuf, N., Mazarguil, H., Clamens, S.,Bayard, F., 1997. Extracellular cleavage of the vascular endothelial growth factor189-amino acid form by urokinase is required for its mitogenic effect. J. Biol.Chem. 272, 13390e13396.
Rini, B.I., Small, E.J., 2005. Biology and clinical development of vascular endothelialgrowth factor-targeted therapy in renal cell carcinoma. J. Clin. Oncol. 23,1028e1043.
Roberts, J.R., Perkins, G.D., Fujisawa, T., Pettigrew, K.A., Gao, F., Ahmed, A.,Thickett, D.R., 2007. Vascular endothelial growth factor promotes physicalwound repair and is anti-apoptotic in primary distal lung epithelial and A549cells. Crit. Care Med. 35, 2164e2170.
Robinson, C.J., Stringer, S.E., 2001. The splice variants of vascular endothelial growthfactor (VEGF) and their receptors. J. Cell Sci. 114, 853e865.
Seth, A., Gonzalez, F.A., Gupta, S., Raden, D.L., Davis, R.J., 1992. Signal transductionwithin the nucleus by mitogen-activated protein kinase. J. Biol. Chem. 267,24796e24804.
Skuta, G.L., Parrish 2nd, R.K., 1987. Wound healing in glaucoma filtering surgery.Surv. Ophthalmol. 32, 149e170.
Soker, S., Miao, H.Q., Nomi, M., Takashima, S., Klagsbrun, M., 2002. VEGF165mediates formation of complexes containing VEGFR-2 and neuropilin-1 thatenhance VEGF165-receptor binding. J. Cell Biochem. 85, 357e368.
Soker, S., Takashima, S., Miao, H.Q., Neufeld, G., Klagsbrun, M., 1998. Neuropilin-1 isexpressed by endothelial and tumor cells as an isoform-specific receptor forvascular endothelial growth factor. Cell 92, 735e745.
Stalmans, I., Lambrechts, D., De Smet, F., Jansen, S., Wang, J., Maity, S., Kneer, P., vonder Ohe, M., Swillen, A., Maes, C., Gewillig, M., Molin, D.G., Hellings, P., Boetel, T.,Haardt, M., Compernolle, V., Dewerchin, M., Plaisance, S., Vlietinck, R.,Emanuel, B., Gittenberger-de Groot, A.C., Scambler, P., Morrow, B., Driscol, D.A.,Moons, L., Esguerra, C.V., Carmeliet, G., Behn-Krappa, A., Devriendt, K.,Collen, D., Conway, S.J., Carmeliet, P., 2003. VEGF: a modifier of the del22q11(DiGeorge) syndrome? Nat. Med. 9, 173e182.
Stalmans, I., Ng, Y.S., Rohan, R., Fruttiger, M., Bouche, A., Yuce, A., Fujisawa, H.,Hermans, B., Shani, M., Jansen, S., Hicklin, D., Anderson, D.J., Gardiner, T.,Hammes, H.P., Moons, L., Dewerchin, M., Collen, D., Carmeliet, P., D’Amore, P.A.,2002. Arteriolar and venular patterning in retinas of mice selectively expressingVEGF isoforms. J. Clin. Invest. 109, 327e336.
Takahashi, T., Yamaguchi, S., Chida, K., Shibuya, M., 2001. A single autophosphor-ylation site on KDR/Flk-1 is essential for VEGF-A-dependent activation of PLC-gamma and DNA synthesis in vascular endothelial cells. EMBO J. 20, 2768e2778.
Tirand, L., Frochot, C., Vanderesse, R., Thomas, N., Trinquet, E., Pinel, S., Viriot, M.L.,Guillemin, F., Barberi-Heyob, M., 2006. A peptide competing with VEGF165binding on neuropilin-1 mediates targeting of a chlorin-type photosensitizerand potentiates its photodynamic activity in human endothelial cells. J. Control.Release 111, 153e164.
Usui, T., Ishida, S., Yamashiro, K., Kaji, Y., Poulaki, V., Moore, J., Moore, T., Amano, S.,Horikawa, Y., Dartt, D., Golding, M., Shima, D.T., Adamis, A.P., 2004.VEGF164(165) as the pathological isoform: differential leukocyte and endo-thelial responses through VEGFR1 and VEGFR2. Invest. Ophthalmol. Vis. Sci. 45,368e374.
Wells, A.P., Crowston, J.G., Marks, J., Kirwan, J.F., Smith, G., Clarke, J.C., Shah, R.,Vieira, J., Bunce, C., Murdoch, I., Khaw, P.T., 2004. A pilot study of a system forgrading of drainage blebs after glaucoma surgery. J. Glaucoma 13, 454e460.
Wilgus, T.A., Ferreira, A.M., Oberyszyn, T.M., Bergdall, V.K., Dipietro, L.A., 2008.Regulation of scar formation by vascular endothelial growth factor. Lab. Invest.88, 579e590.
Wu, Y., Zhang, Q., Ann, D.K., Akhondzadeh, A., Duong, H.S., Messadi, D.V., Le, A.D.,2004. Increased vascular endothelial growth factor may account for elevated
T. Van Bergen et al. / Experimental Eye Research 93 (2011) 689e699698

level of plasminogen activator inhibitor-1 via activating ERK1/2 in keloidfibroblasts. Am. J. Physiol. Cell Physiol. 286, C905eC912.
Yamazaki, Y., Morita, T., 2006. Molecular and functional diversity of vascularendothelial growth factors. Mol. Divers. 10, 515e527.
Zachary, I., 2003. VEGF signalling: integration and multi-tasking in endothelial cellbiology. Biochem. Soc. Trans. 31, 1171e1177.
Zhang, H.T., Scott, P.A., Morbidelli, L., Peak, S., Moore, J., Turley, H., Harris, A.L.,Ziche, M., Bicknell, R., 2000. The 121 amino acid isoform of vascular endothelialgrowth factor is more strongly tumorigenic than other splice variants in vivo.Br. J. Cancer 83, 63e68.
Zhang, Y., Furumura,M., Morita, E., 2008. Distinct signaling pathways confer differentvascular responses to VEGF 121 and VEGF 165. Growth Factors 26, 125e131.
T. Van Bergen et al. / Experimental Eye Research 93 (2011) 689e699 699




















