
Katanin p80 Regulates Human Cortical Development by Limiting Centriole and Cilia Number
Upload
khangminh22Category
view
1download
0
THE ROLE OF CEP120 AND SPICE1 IN HUMAN
CENTRIOLE DUPLICATION
By
David Comartin
A thesis submitted in conformity with the requirements
for the degree of Doctor of Philosophy
Department of Molecular Genetics
University of Toronto
© Copyright by David Comartin (2015)

ii
The Role of CEP120 and SPICE1 in Human Centriole Duplication
David Comartin
Doctorate of Philosophy
Department of Molecular Genetics
University of Toronto
2015
ABSTRACT
The centrosome is a collection of proteins scaffolded upon microtubule based structures called
centrioles. Centrosomes play critical roles as the primary microtubule organizing centers in
interphase and mitotic cells, and template the formation of cilia and flagella. A mitotic cell
contains two centrosomes, each with two centrioles, and following mitosis daughter cells inherit
one centrosome. Before the next mitotic division, centrioles are duplicated in a process where
one procentriole forms adjacent to each existing centriole. Mutations in proteins required for
centriole duplication and/or centrosome function cause developmental defects such as primordial
dwarfism, microcephaly and ciliopathies. Additionally many tumor cells contain excess
centrosomes, and extra centrosomes are a driver of cancer progression. Thus, the correct number
and function of centrosomes is critical for human health, and understanding the proteins involved
in centriole assembly will offer insights into important diseases. Here, I describe the functions of
CEP120 and SPICE1 as proteins required for centriole assembly. I show that CEP120 and
SPICE1 cooperate with CPAP in the assembly of centrioles. Within the centriole duplication
pathway, CEP120 and SPICE1 are dependent upon the presence of SASS6, STIL, CPAP and
CEP135, and are in turn required for CEP135, CP110, CEP97 and Centrin recruitment. Further,

iii
CEP120 interacts with CPAP, and they cooperate with SPICE1 in centriole microtubule
assembly and elongation. Following up on this work, I identify a group of proteins with
previously undescribed roles in centriole duplication, including the human nucleosome assembly
protein NAP1L1. These proteins were identified by screening for proteins in the proximity of
CEP120 and SPICE1, and several of them are important for centriole elongation, suggesting a
functional link to CEP120 and SPICE1 in centriole assembly. The results herein represent
important advances in our understanding of how CEP120 and SPICE1 function, and in our
understanding of centriole assembly and elongation.

iv
ACKNOWLEDGEMENTS
I would like to first thank my supervisor Dr. Laurence Pelletier, for his mentorship, scientific
direction, valuable advice, and the resources provided throughout the course of my Ph.D. I have
greatly benefitted from the many lessons learned and opportunities I was given while in the
Pelletier Lab. Thank you for the great experience!
To my committee members, Dr. Frank Sicheri, Dr. David Bazett-Jones and Dr. Ian Scott: Thank
you all for your support, advice, guidance, and the time investment each of you made in
attending meetings and reading reports. I am very fortunate to have had input from such a great
group of scientists.
Many thanks to the groups I have had collaborations with throughout my Ph.D.: Dr. James
Hutchins and Dr. Jan-Michael Peters, Dr. Marco Archinti and Dr. Jens Luders, Dr. Eden Fussner
and Dr. David Bazett-Jones, Doug Holmyard, Dr. Etienne Coyaud and Dr. Brian Raught, and
finally Stine Morthorst and Dr. Lotte Pedersen. Your contributions, and the opportunity to work
with each of you, have been amazing.
To the members of the Pelletier Lab, past and present who made this experience fun, thank you
all! Special thanks to Deborah Pinchev, Sally Cheung, Dr. Steffen Lawo, Dr. Monica Hasegan
and Dr. Gagan Gupta for your work in contribution to the publication of a paper during my Ph.D.
Thank you also to Christina Yeh, Dr. Nicole St.-Denis, Dr. Joao Goncalves, Bahareh Adhami
Mojarad, Dr. Ladan Gheiratmand, Dr. Johnny Tkach, Christine Holly, Dr. Mariana Gomez-
Ferreria, Dr. Mikhail Bashkurov, Andrea Tagliaferro, Rachel Ford, Dr. Yi Luo, Yi Liu, and
Qiazhu-Wu for being great colleagues. Thank you to Dr. Suzanna Prosser, Dr. Monica Hasegan,
Dr. Johnny Tkach, Dr. Gagan Gupta, and soon-to-be-Dr. Christina Yeh for editing this thesis.
Lastly, very special thanks to Dr. Gagan Gupta, Dr. Nicole St.-Denis, and Dr. Steffen Lawo for
frequent advice and guidance on many scientific problems, and for your general wisdom.
Most importantly, a special thank you to my mother, father and brother for encouraging me to
undertake this project, and supporting me throughout the long process.

v
Table of Contents
ABSTRACT....................................................................................................................................i
ACKNOWLEDGEMENTS.........................................................................................................iv
LIST OF TABLES.....................................................................................................................viii
LIST OF FIGURES.....................................................................................................................ix
LIST OF ABBREVIATIONS.....................................................................................................xi
LIST OF PROTEINS DISCUSSED..........................................................................................xii
1. Chapter I: Introduction ........................................................................................................ 1 1.1. Functions of the Centrosome............................................................................................ 2
1.1.1. The Functions of Centrosomes in Interphase ...................................................................................... 2 1.1.2. The Functions of Centrosomes in Mitosis ........................................................................................... 3 1.1.3. The Functions of Centrosomes in Cilia Formation .............................................................................. 5
1.2. The Centrosome in Disease .............................................................................................. 8 1.2.1. Cancer.................................................................................................................................................. 8 1.2.2. Developmental Diseases .................................................................................................................... 10
1.3. The Structure of the Centrosome ................................................................................... 11 1.3.1. Centrioles .......................................................................................................................................... 11 1.3.2. The PCM ........................................................................................................................................... 15
1.4. The Centrosome Cycle ................................................................................................... 17 1.4.1. Disengagement .................................................................................................................................. 17 1.4.2. Centrosome Separation ...................................................................................................................... 18 1.4.3. Centrosome Maturation ..................................................................................................................... 19
1.5. Centriole Duplication ..................................................................................................... 19 1.5.1. Procentriole Formation ...................................................................................................................... 20 1.5.2. Procentriole Elongation and Length Regulation ................................................................................ 25 1.5.3. The Regulation of Centriole Duplication .......................................................................................... 30
1.6. CEP120 and SPICE1 ...................................................................................................... 34
1.7. Rationale of this Thesis .................................................................................................. 35 1.8. Figures ............................................................................................................................ 36
2. Chapter II: CEP120 and SPICE1 Cooperate with CPAP in Centriole Elongation ..... 44 2.1. Statement of Contributions............................................................................................. 45 2.2. Summary ........................................................................................................................ 47
2.3. Introduction .................................................................................................................... 48 2.3.1. Combining Super-Resolution Imaging with PLK4 Induced Centriole Overduplication to Study
Procentriole Assembly ....................................................................................................................................... 48 2.3.2. The Use of Chemical Manipulation of Microtubules to Study Centriole Elongation ........................ 49
2.4. Results. ........................................................................................................................... 50 2.4.1. CEP120 and SPICE1 Interact and are Required for Centriole Duplication in Cycling Cells. ........... 50 2.4.2. CEP120 and SPICE1 are Required for PLK4 Induced Centriole Overduplication ........................... 51 2.4.3. Using 3D SIM to Study Centriole Assembly. ................................................................................... 53 2.4.4. Defining the Roles of CEP120 and SPICE1 in Procentriole Assembly ............................................ 54 2.4.5. Investigating the Role of CEP120 and SPICE1 in Procentriole Structure ......................................... 56 2.4.6. Comparison of the Effects of CEP120 or SPICE1 Depletion versus Microtubule Depolymerization
on PLK4 Induced Centriole Overduplication ..................................................................................................... 58 2.4.7. CEP120 and SPICE1 Cooperate with CPAP in Centriole Elongation .............................................. 60
2.5. Discussion ...................................................................................................................... 62 2.5.1. Placing CEP120 and SPICE1 in the Centriole Assembly Pathway ................................................... 62 2.5.2. CEP120 and SPICE1 Cooperate with CPAP in Centriole Elongation .............................................. 63 2.5.3. CEP120 and SPICE1 are Important for Microtubule Formation during Procentriole Assembly ...... 65 2.5.4. CEP135 Localization Requires CEP120 and SPICE1, and is a Microtubule Dependent Event
During Centriole Duplication ............................................................................................................................. 66

vi
2.5.5. CEP120 and SPICE1 are Required for Tubulin Incorporation into Procentrioles ............................. 67 2.6. Figures ............................................................................................................................ 70 2.7. Materials and Methods ................................................................................................. 103
2.7.1. Cell lines and Tissue Culture ........................................................................................................... 103 2.7.2. RNA Interference ............................................................................................................................ 103 2.7.3. Cloning of CEP120 siRNA Resistant Construct ............................................................................. 104 2.7.4. PLK4 Induced Centriole Overduplication Assays ........................................................................... 104 2.7.5. Taxol Induced Centriole Elongation ................................................................................................ 105 2.7.6. CPAP or CEP120 Induced Centriole Elongation ............................................................................ 105 2.7.7. Immunofluorescence Microscopy (IFM) and 3D-SIM .................................................................... 105 2.7.8. Cloning ............................................................................................................................................ 107 2.7.9. Statistical Methods .......................................................................................................................... 107 2.7.10. Western Blots .................................................................................................................................. 107 2.7.11. Electron Microscopy ....................................................................................................................... 108
3. Chapter III: Identification of CEP120 and SPICE1 Associated Proteins Required for
Centriole Duplication and Elongation .................................................................................... 109 3.1. Statement of Contribution, Rights and Permissions..................................................... 110
3.2. Summary ...................................................................................................................... 111 3.3. Introduction .................................................................................................................. 112
3.3.1. BioID ............................................................................................................................................... 112 3.3.2. Human Nucleosome Assembly Proteins ......................................................................................... 113 3.3.3. The Role of tubulin Glutamylation in Microtubule and Centriole Stability .................................... 114
3.4. Results .......................................................................................................................... 116 3.4.1. Identification of Potential Functional Interactors of CEP120 and SPICE1 by BioID ..................... 116 3.4.2. Characterization of CEP120/SPICE1 Associated Proteins .............................................................. 117 3.4.3. NAP1L1 is a CEP120 and SPICE1 Associated Protein Required for Centriole Duplication .......... 120
3.5. Discussion .................................................................................................................... 124 3.5.1. Identification of CEP120 and SPICE1 Associated Proteins Critical for PLK4 Induced Centriole
Duplication ....................................................................................................................................................... 124 3.5.2. Multiple CEP120 and SPICE1 Associated Proteins Are Required for CEP120 or SPICE1
localization ....................................................................................................................................................... 124 3.5.3. Depletion of CEP120 and SPICE1 Associated Proteins Affects the Microtubule Cytoskeleton ..... 129 3.5.4. NAP1L1 is Required for Centriole Duplication .............................................................................. 131
3.6. Figures .......................................................................................................................... 135 3.7. Materials and Methods ................................................................................................. 180
3.7.1. Cell Culture, RNAi, PLK4 Assay, and Microscopy Sample Preparation and Imaging ................... 180 3.7.2. BioID Analysis ................................................................................................................................ 180 3.7.3. PLK4 Screen ................................................................................................................................... 181 3.7.4. Analyses of CEP120 and SPICE1 localization, Microtubule Glutamylation and Centriole
Elongation following Depletion of CEP120 and SPICE1 Associated Proteins in U-2 OS .............................. 181 3.7.5. Bioinformatic Analysis of NAP1L-family members and Isoforms ................................................. 181 3.7.6. Cloning of Rescue Constructs ......................................................................................................... 182 3.7.7. Quantitative PCR to Detect NAP1L1 and NAP1L4 Transcripts Following treatment with
siRNAs Against NAP1L1 ................................................................................................................................ 182 4. Chapter IV: Conclusion and Future Directions ............................................................. 190
4.1. The Functions of CEP120 and SPICE1 in Procentriole Assembly and Elongation .... 191 4.2. Identification of CEP120 and SPICE1 Associated Proteins with Uncharacterized Roles
in Centriole Assembly ............................................................................................................ 193 4.3. A Role for NAP1L1 in Centriole Duplication ............................................................. 195
5. LITERATURE CITED ..................................................................................................... 197

vii
LIST OF TABLES
Table 3.1 CEP120 Associated Proteins Detected by BioID 137
Table 3.2 SPICE1 Associated Proteins Detected by BioID 140
Table 3.3 Summary of the Phenotypes of CEP120 and SPICE1 Associated Proteins in
Secondary Assays
158
Table 3.4 Summary of the NAP1L1 siRNAs and Their Associated Phenotypes 179
Table 3.5 Primary Antibodies Used in this Work 184
Table 3.6 Secondary Antibodies Used for Microscopy in this Work 185
Table 3.7 Sequences of Small Interfering RNA Sequences Used in this Work 186
Table 3.8 Primers for esiRNA Used in this Study 188
Table 3.9 Plasmids Used in this Study 189

viii
LIST OF FIGURES
Figure 1.1 ...................................................................................................................................... 36
Figure 1.2 ...................................................................................................................................... 38
Figure 1.3 ...................................................................................................................................... 40
Figure 1.4 ...................................................................................................................................... 42
Figure 2.1.......................................................................................................................................70
Figure 2.2 ...................................................................................................................................... 71
Figure 2.3.. .................................................................................................................................... 73
Figure 2.4 ...................................................................................................................................... 74
Figure 2.5 ...................................................................................................................................... 76
Figure 2.6 ...................................................................................................................................... 78
Figure 2.7 ...................................................................................................................................... 80
Figure 2.8 ...................................................................................................................................... 82
Figure 2.9 ...................................................................................................................................... 84
Figure 2.10 .................................................................................................................................... 86
Figure 2.11 .................................................................................................................................... 88
Figure 2.12. ................................................................................................................................... 90
Figure 2.13 .................................................................................................................................... 92
Figure 2.14 .................................................................................................................................... 94
Figure 2.15 .................................................................................................................................... 96
Figure 2.16 .................................................................................................................................... 98
Figure 2.17 .................................................................................................................................... 99
Figure 2.18 .................................................................................................................................. 101
Figure 3.1 .................................................................................................................................... 135
Figure 3.2 .................................................................................................................................... 144
Figure 3.3 .................................................................................................................................... 145
Figure 3.4 .................................................................................................................................... 146
Figure 3.5 .................................................................................................................................... 148
Figure 3.6 .................................................................................................................................... 150
Figure 3.7 .................................................................................................................................... 152
Figure 3.8 .................................................................................................................................... 154

ix
Figure 3.9 .................................................................................................................................... 156
Figure 3.10 .................................................................................................................................. 159
Figure 3.11 .................................................................................................................................. 160
Figure 3.12 .................................................................................................................................. 162
Figure 3.13 .................................................................................................................................. 164
Figure 3.14 .................................................................................................................................. 166
Figure 3.15 .................................................................................................................................. 168
Figure 3.16 .................................................................................................................................. 170
Figure 3.17 .................................................................................................................................. 172
Figure 3.18 .................................................................................................................................. 174
Figure 3.19 .................................................................................................................................. 175
Figure 3.20 .................................................................................................................................. 176
Figure 3.21 .................................................................................................................................. 178

x
LIST OF ABBREVIATIONS
3D-SIM Three-dimensional structured illumination microscopy
APC Anaphase promoting complex
AP-MS Affinity purification followed by mass spectrometry
Co-IP Co-immunoprecipitation
CDK Cyclin-Dependent-kinase
CDKI Cyclin-Dependent-kinase Inhibitor
CTD Carboxy-terminal domain of a protein
DAPI 4', 6-diamidino-2-phenylindole
DNA Deoxyribonucleic acid
EM Electron microscopy
esiRNA Endoribonuclease-prepared siRNA
FSG Fish skin gelatin
G1 Gap 1 phase of the cell cycle
G2 Gap 2 phase of the cell cycle
HU Hydroxyurea
IP Immunoprecipitation
kDa kilo-Daltons
M Mitosis/mitotic-phase of the cell cycle
MAP Microtubule-associated protein
Min Minute
MS Mass spectrometry
MT Microtubule
MTOC Microtubule-organizing center
NA Numerical Aperture
NTD Amino-terminal domain of a protein
PAGE Polyacrylamide gel electrophoresis
PALM Photoactivated Localization Microscopy
PBS Phosphate buffered saline
PCM Pericentriolar material
RNA Ribonucleic acid
RNAi RNA interference
SCF Skp1, Cullin, F-box E3 ubiquitin ligase complex
SDS Sodium dodecyl sulfate
SEM Standard error of the mean
siRNA Small interfering RNA
S-phase Synthesis phase of the cell cycle
STED Stimulated Emission Depletion Microscopy
STORM Stochastic Optical Reconstruction Microscopy
TBST Tris buffered saline containing Tween-20
TEM Transmission electron microscopy
.tif Tagged image file (.tif)
γ-TuRC Gamma-tubulin Ring Complex

xi
LIST OF PROTEINS DISCUSSED
HUGO Gene Symbol/Protein Name SYNONYM USED
ANK2/Ankyrin2 ANK2
AURKA/Aurora-kinase A Aurora-A
BirA Biotin Ligase BirA
BTF3/Basic Transcription Factor 3 BTF
BTRCP/-transducin repeat containing E3 ub. Prot. ligase TRCP
CEP350/Centrosomal protein 350 kDa CAP350
CCDC77/coiled-coil domain containing 77 CCDC77
CCDC138/coiled-coil domain containing 138 CCDC138
CCNA2/cyclin A2 Cyclin-A
CCNE1/cyclin E1 Cyclin-E
CCNF/cyclin-F, FBXO1 FBXO1
CDC20/cell division cycle 20 CDC20
CDK1/Cyclin-Depedent-Kinase-1 CDK1
CDK2/Cyclin-Depedent-Kinase-2 CDK2
CDK5RAP2/CDK5 regulatory subunit assoc. prot. 2 CDK5RAP2
CDKN1A/cyclin-dependent-kinase-inhibitor 1A, p21,cip1 p21
CENPJ/Centromere protein J CPAP
CEP83/CCDC41 CEP83
CEP89/CCDC123 CEP89
CEP97/Centrosomal protein 97 kDa CEP97
CEP120/ Centrosomal protein 120 kDa CEP120
CEP131/Azi1 CEP131
CEP135/Centrosomal protein 135 kDa CEP135
CEP152/Centrosomal protein 152 kDa CEP152
CEP164/Centrosomal protein 164 kDa CEP164
CEP170/Centrosomal protein 170 kDa CEP170
CEP192/Centrosomal protein 192 kDa CEP192
CEP250/Centrosomal protein 250 kDa C-NAP1
CETN2/Centrin 2 CETN2/Centrin
CDH3/chromodomain helicase DNA binding protein 3 CDH3
CNTROB/Centrosomal BRCA2 interacting protein Centrobin
CP110/centriolar coiled-coil protein 110kDa CP110
CROCC/cilia rootlet coiled-coil Rootletin
CSPP1/Centrosome and spindle pole assoc. protein 1 CSPP1
CUL1/Cullin 1 CUL1
DCTN2/dynactin-2, p50 Dynactin Dynactin
DCTN4/dynactin-4, p62 dynactin DCTN4
ESPL1/extra spindle pole bodies homolog 1 Separase
FBF1/Fas binding factor 1 FBF1
FBXW5/F-box and WD repeat domain containing 5 FBXW5
FOP/ FGFR1 oncogene partner FOP
FZR1/fizzy/cell division cycle 20 related 1 Cdh1
HIST3H2BB/ histone cluster 3, H2bb HIST3H2BB

xii
LIST OF PROTEINS DISCUSSED
HUGO GENE SYMBOL/NAME SYNONYM USED H2AFY/H2A histone family, member Y H2AFY
IPO(4,5,7,8)/Importin(4,5,7,8) IPO4, IPO5, IPO7, IPO8
KIAA0586 KIAA0586
KIAA1731 KIAA1731
KIAA0753 KIAA0753
KIF24/kinesin-family-member 24 KIF24
KIF11/kinesin-family-member 11, Eg5 Eg5
LRRC45/leucine rich repeat containing 45 LRRC45
MAP7/microtubule assoc. protein 7 MAP7
MAP9/microtubule assoc. protein 9 MAP9
MAP7D3/MAP7 domain containing 3 MAP7D3
MIB1/Mindbomb E3 ubiquitin protein ligase 1 MIB1
MTOR/Mechanistic Target of Rapamycin mTOR
MTUS1/microtubule assoc. tumour suppressor prot. 1 MTUS1
NAP1L1/nucleosome assembly prot.1-like 1 NAP1L1
NAP1L1/nucleosome assembly prot.1-like 2 NAP1L2
NAP1L1/nucleosome assembly prot.1-like 3 NAP1L3
NAP1L1/nucleosome assembly prot.1-like 4 NAP1L4
NAP1L1/nucleosome assembly prot.1-like 5 NAP1L5
NEDD1/neural precursor cell expressed, developmentally
down-regulated 1
NEDD1
NEK2/Nima-related kinase 2 NEK2
NIN/Ninein Ninein
NPM1/Nucleophosmin Nucleophosmin
OFD1/orofacial-digital syndrome 1 OFD1
OFD14/orofacial-digital syndrome 14 OFD14, C2cd3
PCNT/Pericentrin Pericentrin
PLK1/Polo-like kinase 1 PLK1
PLK2/Polo-like kinase 2 PLK2
PLK4/Polo-like kinase 4 PLK4
POC1A/POC centriolar protein A POC1 or POC1A
POC1B/POC centriolar protein B POC1 or POC1B
hPOC5/POC centriolar protein 5 hPOC5
POLH/Polymerase (DNA Directed) eta POLH
PP1CA/prot. phosphatase 1, catalytic subunit, isozyme PP1
RAD21/SCC1, Sister Chromatid Cohesion 1 SCC1
SASS6/spindle assembly protein 6 homolog (C.elegans) SASS6
SAV1/Salvador Homolog 1 hSAV1
SCLT1/sodium channel and clathrin linker 1 SCLT1
SPICE1/spindle and centrosome assoc. Prot.1 SPICE1
STIL/SCL/TAL1 interrupting locus STIL
STK3/MST2, serine/threonine kinase 3 MST2
TOP2(A,B)Topoisomerase-2(A,B) TOP2A,TOP2B
TP53/tumour protein p53 p53

xiii
LIST OF PROTEINS DISCUSSED
HUGO GENE SYMBOL/NAME SYNONYM USED TP54BP2/tumour protein p53 binding protein 2 p53BP2
TTLL/ tubulin tyrosine ligase-like family member (1-13) TTLL1, TTLL2, etc.
TUB1A/tubulin, 1a -tubulin
TUBB/tubulin, polypeptide -tubulin
TUBG1/tubulin, polypeptide tubulin
TUBGPC (2, 6)/tubulin, complex assoc. prot. (2,6) GCP2, GCP6
USP33/ ubiquitin specific peptidase 33 USP33
USP37/ ubiquitin specific peptidase 37 USP37
ZC2HC1A/Zinc-finger, C2HC-type containing 1A ZC2HC1A

1
1. Chapter I: Introduction

2
1.1. Functions of the Centrosome
My thesis describes the function of two proteins critical for centriole, and accordingly
centrosome, duplication. To understand why centrosomes and centrioles are worth studying, I
briefly discuss their cellular functions in the first part of this introduction. In the second part of
the introduction, I describe some of the diseases that can arise from, or be driven by, defects in
centrosome function and centriole assembly. Together, these sections of the introduction
highlight the importance of research aimed at furthering our understanding of these organelles.
1.1.1. The Functions of Centrosomes in Interphase
The human centrosome is a combination of tubulin based structures called centrioles and a
surrounding matrix of proteins called the pericentriolar material (PCM). The centrosome has
over a hundred associated proteins that perform a variety of cellular functions (Andersen et al.,
2003; Jakobsen et al., 2011). In interphase, the PCM that surrounds centrioles nucleates and
anchors the majority of the cellular microtubules, leading to the term ‘microtubule organizing
center’ (MTOC) (Gould and Borisy, 1977; Nigg and Raff, 2009; Woodruff et al., 2014). By
anchoring the majority of cellular microtubules in interphase, centrosomes play an important role
in cellular organization, shape, motility and intracellular signaling (Arquint et al., 2014; Fu et al.,
2015). For example, the presence of an intact microtubule network organized by the centrosome
is required for the correct positioning of the Golgi apparatus around the centrosome (Rios, 2014).
Golgi apparatus positioning around the centrosome is important in polarized cells. During cell
migration, for example into a wound in a wound-healing assay, centrosomes and the Golgi
become localized between the nucleus and the edge of the cell facing the wound in preparation
for directed cell migration (Yadav and Linstedt, 2011; Yadav et al., 2009). Disruption of Golgi
apparatus structure, or of the connection between the Golgi and the centrosome, prevents this
polarization of the cell and directed migration (Hurtado et al., 2011; Yadav et al., 2009).
Polarization of the centrosome and Golgi apparatus is also important for determining the position
of neuronal axon formation, and in T-cells is important for immunological synapse formation
and directed vesicular secretion (de Anda et al., 2005; Stinchcombe and Griffiths, 2014;
Stinchcombe et al., 2006; Yadav and Linstedt, 2011).

3
Progression through the cell cycle also requires a functional centrosome. In G1 there is a
centrosome-dependent checkpoint that arrests cells if centrosomes are not functioning properly
(Mikule et al., 2007; Pihan, 2013). The checkpoint can be activated by depletion of multiple
centrosomal proteins, before or during the G1 phase of the cell cycle, and is independent of
mitotic defects (Mikule et al., 2007; Pihan, 2013). In this case p53 is activated by p38
phosphorylation, which occurs at the centrosome, and p53 accumulates in the nucleus and drives
transcription of the CDK2 inhibitor p21, causing a block of cell cycle progression into S-phase
(Besson et al., 2008; El-Deiry, 1993; Kishi et al., 2001; Mikule et al., 2007; Sherr and Roberts,
1999). A distinct p53 induced G1 arrest occurs when centrosomes are lost due to prolonged
chemical inhibition of centriole duplication in human cell lines with functional p53 (Wong et al.,
2015). The same inhibitor, when applied to cancer cell lines lacking normal p53, does not stop
them from progressing through the cell cycle (Wong et al., 2015). Upon washout of the
chemical inhibitor, cancer cell lines initiate de novo assembly of centrioles, whereas G1 arrested
cells with functional p53 are not able to enter the cell cycle or assemble centrioles (Wong et al.,
2015). Importantly, the p53-dependent G1 arrest upon chemically induced loss of centrioles
does not involve p38 activation, and is thus different from that observed when centrosomal
proteins are disrupted (Mikule et al., 2007; Wong et al., 2015). These results are consistent with
two distinct p53-dependent checkpoints monitoring cells for the presence and function of
centrosomes as a requisite for progress through the cell cycle. p53 activation in response DNA
damage occurs through phosphorylation by the ATM kinase, but ATM is not involved in either
of the centrosome related G1 arrests (Ciciarello, 2001; Mikule et al., 2007; Shiloh, 2001;
Tritarelli et al., 2004; Wong et al., 2015). Finally, prolonged mitosis is also a known trigger for
p53 activation, through another pathway as discussed in the next section (Chavali et al., 2014;
Insolera et al., 2014; Mikule et al., 2007; Mogensen et al., 2000; Pihan, 2013; Uetake and Sluder,
2010).
1.1.2. The Functions of Centrosomes in Mitosis
In mitosis, each centrosome contains two tightly connected centrioles surrounded by PCM that
nucleates and anchors microtubules to segregate DNA (Gould and Borisy, 1977; Kuriyama and
Borisy, 1981). Centrosomes are localized to the mitotic spindle pole, to which the mitotic
spindle microtubules are anchored. Despite their normal localization at the spindle poles, mitosis

4
has been observed in the absence of centrioles. When centrioles are disassembled in HeLa cells,
mitosis proceeds with spindle poles lacking centrioles (Bobinnec et al., 1998a; Debec et al.,
2010). Similarly, in monkey fibroblast cells, laser-ablation of one or both centrosomes does not
prevent bipolar spindle assembly (Khodjakov et al., 2000). In the context of whole organisms,
results have been more variable. For example, in Drosophila that lack centrioles, asymmetric
cell division is abnormal and spindle formation is slowed during development (Basto et al., 2006;
Debec et al., 2010). These flies, however, die due to failure of cilia formation rather than mitotic
defects (Basto et al., 2006; Debec et al., 2010). Recently, analogous mouse models have been
generated that lack a protein essential for centriole assembly (CPAP/SAS-4), and unlike
Drosophila the mouse embryos die at a stage earlier than mouse embryos that cannot make cilia
(Bazzi and Anderson, 2014). In these acentriolar embryos, mitosis is delayed by the lack of
centrosomes, and widespread p53-dependent apoptosis occurs leading to lethality (Bazzi and
Anderson, 2014). Similar effects were seen when microtubule poisons were used to prolong
mitosis in cultured embryonic cells, indicating that in the context of mouse embryos one or more
checkpoints activate p53-mediated programmed cell death in cases where the mitotic spindle is
slow to form (Bazzi and Anderson, 2014; Fridman and Lowe, 2003). Mammalian female
meiotic cell divisions occur without centrosomes, and spindle assembly in this case is achieved
by a distinct pathway (Manandhar, 2005; Ohkura, 2015). Therefore, in some contexts
centrosomes ensure timely mitotic divisions that are essential during development, but in
cultured somatic cells where centrosomes are ablated, spindle formation is still achieved.
Asymmetric cell divisions produce two distinct daughter cells, for example a progenitor (stem)
cell and a differentiated cell (Morrison and Kimble, 2006). In both Drosophila and mouse
development, a requirement has been demonstrated for centrosomes in regulating the
maintenance of progenitor pools through ensuring asymmetric cell division (Wang et al., 2009;
Yamashita et al., 2007). This is achieved in some contexts through centrosome-based control of
mitotic spindle orientation to ensure retention of only one daughter cell within a stem-cell niche
(Yamashita and Fuller, 2008). For example, in Drosophila male germline stem cells, the older
centrosome maintains close contact with the apical cortex throughout the cell cycle, setting up
spindle orientation to ensure division that leads to a daughter cell being excluded from the niche
(Yamashita and Fuller, 2008; Yamashita et al., 2003, 2007). In the case of Drosophila
neuroblast stem cells, a similar microtubule-based anchoring of one centrosome to the cortex

5
near the niche is also observed, and in this context the younger (daughter) centrosome is retained
(Conduit and Raff, 2010; Januschke et al., 2013). In Drosophila embryos engineered to lose
centrioles early in development, asymmetric cell division still succeeds 70% of the time in larval
neuroblasts, so evidently mechanisms exist that can achieve asymmetric divisions in the absence
of centrioles (Basto et al., 2006). In embryonic mice, the basal epidermal cells undergo
asymmetric cell division with specified spindle orientation, where one daughter cell is retained in
the original niche and the other daughter cell divides perpendicular to the basal layer and
becomes differentiated (Lechler and Fuchs, 2005). Centrosomes may also carry cell-fate
determinants. For example, in the brains of developing mouse embryos, radial glia stem cells
inherit the older centrosome in each asymmetric division (Wang et al., 2009). When the
expression of the mother-centriole subdistal appendage protein Ninein is knocked down,
centrosome inheritance becomes random with subsequent depletion of radial glia progenitor
cells, and this depletion of progenitors is independent of spindle orientation (Bouckson-Castaing
et al., 1996; Mogensen et al., 2000; Wang et al., 2009). Thus centrosome inheritance,
presumably through differential retention of proteins to the mother centrosome, can influence
cell differentiation (Wang et al., 2009). Overall there is good evidence that centrosomes are
required for both timely cell division during development and the maintenance of stem cell
niches through asymmetric cell divisions.
1.1.3. The Functions of Centrosomes in Cilia Formation
Outside of mitotic cell division, the centrosome has an important role in many tissues as a basal
body, the structure from which cilia and flagella are built (Bettencourt-Dias et al., 2011;
Ishikawa and Marshall, 2011; Nigg and Raff, 2009). Cilia and flagella are structures that are
formed by the fusion of the mother centriole with the cell membrane, followed by the elongation
of the distal portion of the mother centriole to form a ciliary axoneme (Bettencourt-Dias et al.,
2011; Goetz and Anderson, 2010; Ishikawa and Marshall, 2011; Nigg and Raff, 2009). The
ciliary axoneme is isolated from the cytoplasm by a barrier (transition zone) but remains
surrounded by the cell membrane along its length (Bettencourt-Dias et al., 2011; Goetz and
Anderson, 2010; Ishikawa and Marshall, 2011; Nigg and Raff, 2009). Cilia can be motile or
non-motile, and cells can contain one (primary) cilium or many (motile) cilia (Bettencourt-Dias
et al., 2011; Ishikawa and Marshall, 2011; Nigg and Raff, 2009). Primary (non-motile) cilia

6
allow cells to receive and internalize extracellular signals and activate critical signaling pathways
(Goetz and Anderson, 2010). Motile cilia and flagella rely on the sliding of microtubule motors
between microtubule doublets to allow them to ‘beat’, i.e. bend and deform in a coordinated
effort to generate fluid movement in the extracellular space, or cell propulsion in the case of
flagella (Lindemann and Lesich, 2010; Roberts et al., 2013). The importance of these organelles
are made clear by the variety of developmental defects that can occur as a result of failure of cilia
formation, collectively called the ‘ciliopathies’ (Goetz and Anderson, 2010; Nigg and Raff,
2009; Sharma et al., 2008; Tobin and Beales, 2009; Waters and Beales, 2011). These include
patterning defects such as polydactyly or situs inversus/heterotaxy, orofacialdigital syndrome
and Jeune asphyxiating thoracic dystrophy, as well as defects in kidneys, retina and brain
development (Pennekamp et al., 2015; Sharma et al., 2008; Waters and Beales, 2011). Both
motile cilia and primary cilia have been shown to be important for proper development (Sharma
et al., 2008; Tobin and Beales, 2009). Mouse embryos that lack cilia and centrioles due to the
absence of the centriole assembly protein STIL show developmental failures consistent with loss
of Sonic Hedgehog (Shh) signaling (David et al., 2014; Huangfu et al., 2003; Izraeli et al., 1999).
On the other hand, induction of excess centriole formation in cultured mouse embryonic
fibroblasts leads to the formation of multiple primary-cilia per cell upon serum starvation
(Mahjoub and Stearns, 2012). The formation of excess cilia diffuses the localization of
Smoothened (Smo) to the ciliary membrane and leads to a reduction of downstream
transcriptional activation following addition of Shh protein to the cells (Mahjoub and Stearns,
2012). Hence control of centriole number is important for ensuring formation of a single
primary cilium capable of activating cellular responses to external signals (Mahjoub and Stearns,
2012).
The breaking of left-right (LR) symmetry during development requires the function of cilia
within a structure called the ‘node’ (Blum et al., 2014; Lee and Anderson, 2008). The nodal cilia
exhibit a distinct rotational movement that establishes a directed flow in the extracellular fluid
towards the left side of the embryo (called leftward-flow), and this flow is both necessary and
sufficient to break the symmetry of the developing embryo (Blum et al., 2014; Lee and
Anderson, 2008; Nonaka et al., 1998, 2002). The fluid current is in turn detected by sensory cilia
on cells to the left of the node, which then initiate a signaling cascade that is unique to one side
of the embryo (Blum and Vick, 2015; Blum et al., 2014; Lee and Anderson, 2008). Thus cilia

7
also play a central role in the breaking of embryonic symmetry and correct establishment of the
vertebrate body plan. The many important roles of cilia during development all depend on
properly functioning centrosomes capable of acting as basal bodies to facilitate their assembly.
The presence of centrioles correlates strongly with the ability to form cilia or flagella throughout
various organisms, and both centrioles and cilia/flagella are thought to have been present in the
last eukaryotic common ancestor (LECA) (Azimzadeh, 2014; Carvalho-Santos et al., 2011;
Debec et al., 2010). Mitotic spindle pole formation and accurate DNA segregation had been
widely thought to be the important role of centrioles, however as discussed, spindle formation
can be achieved without centrioles (Debec et al., 2010; Dumont and Desai, 2012). In organisms
that do not make cilia or flagella, such as yeasts or certain amoebas (Dictyostelium discoidum),
their microtubule organizing centers (called spindle pole bodies and nuclear associated bodies,
respectively) do not contain centrioles and are structurally distinct from centrosomes
(Azimzadeh, 2014; Debec et al., 2010). Further, in planarians only their multiciliated cells
assemble centrioles, and planarians lack the genes that encode certain conserved PCM proteins
required to generate functional centrosomes (Azimzadeh, 2014; Azimzadeh et al., 2012).
Accordingly, it has been proposed that the essential role of a centriole that results in its
maintenance through evolution may be the ability to template cilia/flagella, and that centrioles
exist as centrosomes at mitotic spindle poles primarily to ensure daughter cells inherit the
capacity to make cilia/flagella in some species (Azimzadeh, 2014; Carvalho-Santos et al., 2011;
Debec et al., 2010; Friedländer and Wahrman, 1970). Indeed, in human cells, when de novo
assembled centrioles are not tethered to parental centrioles within PCM-nucleating centrosomes,
they are randomly segregated between daughter cells (Wang et al., 2011). Overall, in some
eukaryotes centrioles have been lost along with cilia/flagella and other microtubule organizing
centers evolved, whereas in others the formation of cilia and flagella is achieved through an on-
demand de novo assembly of centrioles, and in species such as humans centrioles act as
centrosomes in mitosis at least partly to ensure the centrioles are passed on equally to every cell.
Aside from the ancestral role of centrosomes in centriole distribution, additional essential
functions of centrosomes have since evolved in complex multicellular organisms (Debec et al.,
2010). For example, as discussed in the previous sections, there are essential roles for
centrosomes in mitosis and interphase. Further, centrioles are absent from female oocytes that
retain PCM proteins, and in some species including humans the sperm donates centrioles without

8
PCM during fertilization to activate embryogenesis (Debec et al., 2010; Dumont and Desai,
2012; Schatten and Sun, 2010). Thus centrioles may have been co-opted to help regulate the
onset of embryonic development in some sexually reproducing species (Debec et al., 2010;
Dumont and Desai, 2012; Schatten and Sun, 2010).
1.2. The Centrosome in Disease
1.2.1. Cancer
In 1914 Theodore Boveri published a book describing the centrosome as a duplicating cellular
structure required for cell division, including the observation that abnormal numbers of
centrosomes gave abnormal cell divisions that gave rise to aneuploidy and the hypothesis that
aneuploidy could lead to cancer (recently translated in Boveri, 2008; Hansford and Huntsman,
2014; Scheer, 2014). Indeed, aneuploidy and extra centrosomes are both now known as
hallmarks of cancer (Chan, 2011; Godinho and Pellman, 2014). In mice lacking the tumour
suppressor gene p53, centrosome amplification is observed and mice develop tumors
(Donehower et al., 1992; Fukasawa, 2005, 2008; Fukasawa et al., 1997). There is debate as to
whether centrosome numerical abnormalities initiate cancer, although they are thought to drive it
(Nigg, 2002; Nigg and Raff, 2009). Extra centrosomes are known to lead to multipolar cell
divisions which are observed in cancers (Godinho and Pellman, 2014; Nigg, 2002). Experiments
using cancer cell lines have revealed that multipolar cell divisions normally result in cells that
eventually die, suggesting that this is not a common initial cause of cancer (Ganem et al., 2009).
Cancer cells with excess centrosomes can form bipolar spindles through a process called
centrosome clustering, to avoid potentially fatal multipolar divisions (Ganem et al., 2009;
Quintyne et al., 2005). These bipolar spindles form after a transient multipolar state during
mitosis, and during this temporary multi-polar state misaligned centrosomes can form
attachments to either kinetochore through spindle microtubules (Ganem et al., 2009). Later in
mitosis, centrosomes from the ‘extra’ poles are pulled together into a bipolar formation, and this
results in kinetochores with attachments to both spindle poles (merotelic) (Ganem et al., 2009;
Godinho and Pellman, 2014; Gregan et al., 2011; Quintyne et al., 2005; Ring et al., 1982). As a
result of merotelic attachment, chromosomes are incorrectly segregated leading to aneuploidy, or

9
lag behind during pole-ward movement in anaphase, resulting in DNA damage (Ganem et al.,
2009; Janssen et al., 2011). Recently, experiments have tested the impact of extra centrosomes
on developing organisms. When embryonic mice are engineered to have extra centrosomes
within their neural cells, multipolar mitosis leads to cell death and microcephaly, rather than
over-proliferation (Marthiens et al., 2013). In Drosophila, neural stem cells generated to have
excess centrosomes initiate tumour formation upon transplantation to an adult Drosophila (Basto
et al., 2008). While this suggests tumorigenic potential in cells with too many centrosomes, the
caveat here is that the flies that have excess centrosomes are not themselves prone to genomic
instability or other obvious defects (Basto et al., 2008). Thus in many models tested so far,
centrosome amplification alone appears unlikely to initiate cancer directly (Godinho and
Pellman, 2014; Nigg, 2002). In a fly model where asymmetric stem-cell division is disrupted by
targeting cell polarity regulators, these stem-cells generate tumors upon transplantation to an
adult fly (Caussinus and Gonzalez, 2005). Despite initially being genetically normal with two
centrosomes, a fraction of the tumors that later developed from these cells obtained both extra
centrosomes and aneuploidy (Caussinus and Gonzalez, 2005). Based on this finding, coupled
with the common occurrence of superfluous centrosomes in cancers, the hypothesis that extra
centrosomes drive aneuploidy in cancer but don’t commonly initiate it, has been proposed
(Godinho and Pellman, 2014; Nigg, 2002).
The role of centrosome amplification in tumour aggressiveness has been linked to an effect of
interphase centrosomes on cellular migration. PLK4 is a kinase required for centriole
duplication, and when overexpressed PLK4 causes the formation of multiple centrioles around a
parental centriole (Bettencourt-Dias et al., 2005; Habedanck et al., 2005; Kleylein-Sohn et al.,
2007). In 3D MCF10A mammary epithelia cell cultures cells that have extra centrioles due to
PLK4 overexpression have increased migration, decreased cell-cell adhesion, and form dynamic
cellular protrusions that promote invasiveness in culture (Godinho et al., 2014). The extra
centrosomes in these cells cluster in interphase, and accordingly nucleate greater numbers of
microtubules, leading to Rac1 activation and RhoA deactivation (Godinho et al., 2014). Rac1
activation leads to lamellipodia and cell migration through stimulation of actin filament
formation at the leading edge of cells (Burridge and Wennerberg, 2004; Lawson and Burridge,
2014; Sadok and Marshall, 2014). The invasive phenotype, and formation of cellular protrusions
were reversed by depletion of CEP192, a protein required for centriole duplication and

10
microtubule nucleation within the PCM (Godinho et al., 2014; Gomez-Ferreria et al., 2007;
Joukov et al., 2010, 2014; Zhu et al., 2008). CEP192 depletion reduced the activation of Rac1,
presumably through reducing-tubulin and microtubule nucleation at centrosomes (Godinho et
al., 2014; Zhu et al., 2008). Engineered aneuploidy in MCF10A cells in the absence of
centrosome amplification did not lead to the formation of invasive cellular protrusions (Godinho
et al., 2014). Thus the amplification of centrosomes can likely drive metastasis through
changing the interphase microtubule landscape, providing a mechanism for centrosome
involvement in aggressive cancers beyond aneuploidy (Godinho et al., 2014).
1.2.2. Developmental Diseases
Aside from ciliopathies, several developmental diseases have been linked to proteins of the
centrosome. Notably, these diseases share the characteristic of decreasing the size of patient
tissues, as in the case of primordial dwarfism or microcephaly wherein the brain is under-sized
(Barbelanne and Tsang, 2014; Chavali et al., 2014; Faheem et al., 2015; Nigg et al., 2014).
Autosomal recessive primary microcephaly (MCPH) has been linked to 13 genes to date
(MCPH1-13) and many of these are key centriole duplication proteins (Barbelanne and Tsang,
2014; Chavali et al., 2014; Faheem et al., 2015; Nigg et al., 2014). MCPH6, 7, 8, 9 and 13 have
been identified as the essential centriole duplication proteins CPAP, STIL, CEP135, CEP152 and
CEP63 respectively (Barbelanne and Tsang, 2014; Nigg et al., 2014). The other MCPH genes
can be broadly described as genes that affect mitosis or proliferation (Barbelanne and Tsang,
2014; Chavali et al., 2014; Faheem et al., 2015; Nigg et al., 2014).
How do defects in proteins affecting centriole duplication or centrosome function lead to these
developmental effects? There are two major models for how centrosome defects contribute to
microcephaly; one is disruption of the asymmetric versus symmetric cell division balance, and
the other is a reduction in cell viability as a result of mitotic defects (Chavali et al., 2014).
Mouse models have shown that loss of mother-centrosome functionality leads to stem cell
depletion presumably by disrupting asymmetric cell division (Wang et al., 2009; Yamashita et
al., 2007). Conversely, in mouse models that lack centrosomes, delays in mitosis cause cell
death and microcephaly (Bazzi and Anderson, 2014; Insolera et al., 2014). When p53 is also
removed in such mice to block apoptosis, microcephaly is suppressed without restoration of
correct spindle orientation (Chavali et al., 2014; Insolera et al., 2014). This suggests that in

11
developing mammalian brains, the predominant effect of loss of centrosome function may be
apoptosis driven by inefficient mitosis (Insolera et al., 2014). Consistent with this idea,
depletion of an MCPH protein (WDR62/MCPH2) causes prolonged mitosis with fragmentation
of the PCM after bipolar spindle formation (Barbelanne and Tsang, 2014; Bogoyevitch et al.,
2012). Mice engineered to have reduced WDR62 expression display dwarfism and
microcephaly, with widespread mitotic spindle checkpoint arrest and programmed cell death
leading to a reduction in the progenitor pools in their brains (Chen et al., 2014). Extra centrioles
may also cause developmental microcephaly. Mice which constitutively over-express PLK4
form extra centrioles, and are born smaller than normal (dwarfism) and with microcephaly
(Marthiens et al., 2013). This result is attributed to programmed cell death that occurs in
response to aneuploidy stemming from multipolar mitoses (Marthiens et al., 2013). Consistent
with this idea, several of the MCPH patient mutations to centrosomal genes can cause centriole
amplification (Nigg et al., 2014). Overall, abnormalities of centriole number or centrosome
function are causative in human developmental diseases.
1.3. The Structure of the Centrosome
To understand the results described in the data chapters of this thesis, it is important to have an
understanding of the anatomy of the centrioles and centrosomes. This section of the introduction
will describe the general anatomy of a centriole, and make clear the distinction between a
centriole and a centrosome. The structures of both the centrosome and centriole(s) change
through the cell cycle, and these changes are the topic of the subsequent section of the
introduction.
1.3.1. Centrioles
The structure of the centrosome is maintained by, and organized around the centrioles (Abal et
al., 2005; Bobinnec et al., 1998a; Bornens et al., 1987; Kuriyama and Borisy, 1981). The human
centriole is a barrel-shaped structure with a nine-fold symmetrically organized array of
microtubule structures individually referred to as ‘microtubule triplets’ (Bernhard and De
Harven, 1956; Winey and O’Toole, 2014). and tubulin hetero-dimers can form polymers
called ‘protofilaments’, and microtubules are comprised of a number of protofilaments arranged

12
side-by-side to form a tube (Amos and Klug, 1974; Meurer-Grob et al., 2001; Nogales et al.,
1999). The exact number of protofilaments in a microtubule can vary between cell types both
within and between organisms, but the most often observed microtubules in eukaryotes are made
of 13 protofilaments, having a diameter of ~25 nm, and with the tubulin dimers having a
periodicity of 8 nm along the long-axis (Amos and Klug, 1974; Choi et al., 2009; Meurer-Grob
et al., 2001; Nogales et al., 1999). The structure of the centriolar microtubule triplets from the
model organisms Chlamydomonas and Trichonympha have been studied in sub-nanometer
resolution using cryo-electron tomography (Guichard et al., 2013; Li et al., 2012; Winey and
O’Toole, 2014). The centriole microtubule triplets in these organisms have a single complete
microtubule with 13 protofilaments, connected to two partial microtubules with 10
protofilaments each (Guichard et al., 2013; Li et al., 2012; Winey and O’Toole, 2014). These
microtubules are called the “A”, “B” and “C” microtubules, respectively (Guichard et al., 2013;
Li et al., 2012; Winey and O’Toole, 2014). In mature centrioles the C microtubule extends only
partway up the centriole, and the distal portion is a doublet made up of only the A and B
microtubules (Bornens et al., 1987; Ibrahim et al., 2009; Paintrand et al., 1992).
In most organisms that have centrioles, the nine microtubule triplets of the centriole are initially
organized around a central structure referred to as the ‘cartwheel’ based on its structure having a
central ‘hub’ (ring) from which emanates 9 ‘spokes’ at 40° angles that make contact with the
triplets (Figure 1.1) (van Breugel et al., 2011; Kitagawa et al., 2011; Winey and O’Toole, 2014).
In Caenorhabditis elegans, centrioles differ from the triplet and cartwheel organization of human
centrioles, as their centrioles contain nine single microtubules arranged around a central tube
(Pelletier et al., 2006; Sharp et al., 1999; Winey and O’Toole, 2014). The diameter of the human
centriole is approximately 250 nm, and this diameter is flexible (Ibrahim et al., 2009; Paintrand
et al., 1992; Winey and O’Toole, 2014). Each human centriole cartwheel is composed of 9
dimers of SASS6, where the carboxy-terminal domains (CTDs) of SASS6 point outwards as the
‘spokes’ of the cartwheel towards the microtubule triplets, and facilitate SASS6 dimerization
(van Breugel et al., 2011; Kitagawa et al., 2011). The amino-terminal domains (NTDs) of
SASS6 comprise the ‘central hub’, and their interactions facilitate formation of the 9-dimer hub
(van Breugel et al., 2011; Kitagawa et al., 2011). In other model organisms, including
Drosophila and Chlamydomonas, SASS6 homologs likewise are critical for correct 9-fold
symmetry and formation of the central cartwheel (Nakazawa et al., 2007; Rodrigues-Martins et

13
al., 2007). Similar to the human cartwheel, the central tube in C.elegans requires SAS-6 and
SAS-5, the homologs of SASS6 and STIL respectively (Arquint et al., 2012; Leidel et al., 2005;
Pelletier et al., 2006). Thus despite the variations in cartwheel structure, many centriole
assembly proteins are conserved across species, as is the nine-fold symmetry of centrioles. In
some cases, non-nine-fold centrioles are observed, as in male germ line cells of the gnat Sciara
where massive centrioles are observed with over 20 centriolar microtubules (Gönczy, 2012;
Phillips, 1967). The lumen of these giant centrioles does not appear to contain a cartwheel
(Phillips, 1967). How these centrioles are assembled has not been studied.
The proximal end of a newly formed centriole (procentriole) contains multiple cartwheels.
Recently, the application of cryo-electron tomography in the model organism Trichonympha has
revealed the structure of the cartwheel stacks and their connections to the microtubule triplets in
basal bodies (Figure 1.1) (Guichard et al., 2012, 2013). Trichonympha was used in this work
because its basal bodies have substantially longer centrioles with more cartwheels than humans
or other model organisms (Guichard et al., 2012, 2013; Hirono, 2014). Trichonympha
cartwheels are stacked as pairs where the cartwheel spoke tips of each pair of cartwheels make
contact at a structured dubbed the ‘spoke junction’ (SP-J), which is joined to the spoke-tip (SP-
T) which in turn connects to the A-tubule of the triplet through a structure called the ‘pinhead’
(Figure 1.1) (Guichard et al., 2012, 2013; Hirono, 2014). These observations have yet to be
repeated outside Trichonmypha. Similar spacing of cartwheels is observed within the centriole
lumen in Chlamydomonas, so inter-cartwheel organization may be a conserved feature among
organisms where centrioles contain cartwheels (Hirono, 2014).
There are discs of unknown composition and function within the distal lumen of human
centrioles, making connections with the triplet and doublet microtubules (Ibrahim et al., 2009;
Winey and O’Toole, 2014). These disks are tilted with respect to the longitudinal axis of the
centriole, unlike the cartwheels, and their composition and function is unknown (Ibrahim et al.,
2009; Winey and O’Toole, 2014). The distal lumen of the human centriole contains proteins that
include Centrin and an interacting partner protein hPOC5 (Azimzadeh et al., 2009; Kleylein-
Sohn et al., 2007; Paoletti et al., 1996). The structure and function of the distal lumen of human
centrioles is much less studied than the cartwheel structures of the proximal lumen.

14
At full maturity, the distal portion of a human centriole is decorated with a set of nine distal and
nine sub-distal appendages (Bornens et al., 1987; Brito et al., 2012; Jana et al., 2014; Paintrand et
al., 1992; Winey and O’Toole, 2014). There are several proteins associated with subdistal
appendages. Ninein is a subdistal appendage protein, and it is required for anchoring of
microtubules to the centrosome (Brito et al., 2012; Delgehyr, 2005; Mogensen et al., 2000;
Shinohara et al., 2013; Winey and O’Toole, 2014). Likewise, EB1 is another protein of the
subdistal appendages that plays a role in microtubule anchoring (Askham et al., 2002; Brito et
al., 2012; Louie, 2004). EB1 recruitment to centrosomes requires an interacting protein called
FOP, and FOP in turn requires an interacting protein called CAP350 for its centrosome
localization (Brito et al., 2012; Yan et al., 2006). Depletion of any of these three proteins leads
to loss of microtubule organization by the interphase centrosomes, and FOP and CAP350 are
dispensable for microtubule nucleation but required for microtubule anchoring at the centrosome
following nocodazole washout (Louie, 2004; Yan et al., 2006). CEP170 is another subdistal
appendage protein that localizes to microtubules when overexpressed, and its depletion alters the
microtubule cytoskeleton (Brito et al., 2012; Guarguaglini et al., 2005). The subdistal appendage
protein Centriolin/CEP110 is required along with ODF2 to recruit several proteins important for
recycling endosome function to the mother centriole, and Centriolin is required to target SNARE
and exocyst complexes to the midbody in a process essential for abscission during cytokinesis
(Gromley, 2003; Gromley et al., 2005; Hehnly et al., 2012). Lastly, the -tubulin protein is
localized to subdistal appendages, and its depletion blocks the formation of microtubule asters
and centriole duplication following sperm centrosome addition in Xenopus egg extracts (Chang
et al., 2003). In summary, the functions of subdistal appendages include anchoring microtubules
to the centrosome, and the localization of proteins important for recycling endosome formation.
Mature centrioles are also the templates from which cilia and flagella are assembled. The distal
centriole appendages are required for ciliogenesis, as evident by the fact that disruption of distal
appendage proteins inhibits this process. ODF2 is required for cilia formation, and assembly of
both distal and subdistal appendages (Ishikawa et al., 2005). The function of ODF2 in distal
appendage assembly and cilia formation requires a specific isoform of ODF2, now called
Cenexin1 (Chang et al., 2013). C2CD3/OFD14 is critical for the localization of five distal
appendage proteins; CEP164, SCLT1, CCDC41/CEP83, CCDC123/CEP89, and FBF1 (Ye et al.,

15
2014). CEP164, SCLT1, CEP83, CEP89, FBF1 are all proteins required for cilia formation
(Graser et al., 2007; Tanos et al., 2013). OFD14 is also required for the assembly of the distal
portion of centrioles, so the failed distal appendage formation is likely an indirect effect rather
than a direct recruitment of the five proteins by OFD14 (Thauvin-Robinet et al., 2014; Ye et al.,
2014). OFD1, an interacting protein of OFD14, is also required for distal appendage assembly
and cilia formation (Singla et al., 2010). This is probably also due to a role in regulation of
centriole length, as depletion of OFD1 leads to elongation of the distal portion of the centriole
and failure of distal appendages to form (Singla et al., 2010). These two proteins highlight an
important role for regulators of centriole length in proper cilia formation (Thauvin-Robinet et al.,
2014). CEP83 is required for recruitment of the CEP164, SCLT1, CEP89 and FBF1, while
CEP164 and FBF1 were also dependent on SCLT1 for localization (Tanos et al., 2013).
Disruption of the distal appendage proteins by CEP83 depletion inhibits membrane docking of
the mother centriole during ciliogenesis, consistent with distal appendages playing a critical role
in that step of ciliogenesis (Tanos et al., 2013).
1.3.2. The PCM
The pericentriolar material is organized by centrioles and plays a critical role in nucleating
microtubules at the centrosome (Gould and Borisy, 1977). This function of the PCM is achieved
through localization and regulation of -tubulin ring complexes (-TuRC) (Kollman et al., 2010,
2011). The -TuRC is a conserved multi-subunit complex including -tubulin that acts as a
template to seed the assembly of microtubules (Kollman et al., 2010, 2011). The PCM itself was
until recently viewed as a disorganized mass of proteins, due to limitations in both the resolution
of immunofluorescence microscopy and the capabilities of immuno-electron microscopy to
differentiate between multiple antigens in the same sample (Mennella et al., 2014). Innovations
in fluorescence microscopy have provided the additional resolution required to for examination
of the PCM structure, and microscopy in human and Drosophila cells shows that individual
proteins of the PCM occupy distinct regions surrounding the centrioles (Fu and Glover, 2012;
Lawo et al., 2012; Mennella et al., 2012, 2014; Sonnen et al., 2012). Pericentrin and
CDK5RAP2 are two large scaffold proteins of the PCM, and Pericentrin can be co-
immunoprecipitated with CDK5RAP2 (Kraemer et al., 2011; Wang et al., 2010). Whereas most
PCM proteins studied to date are organized within distinct regions around the centrioles,

16
Pericentrin and CDK5RAP2 appear to span larger regions of the PCM and are polarized with
regard to the centriole, having their CTDs closer to the centrioles (Lawo et al., 2012).
Overexpression of Pericentrin or CDK5RAP2 in interphase results in an excess of PCM around
centrioles, reminiscent of the PCM expansion normally observed in mitosis, with both proteins
and -tubulin recruited to the enlarged PCM (Lawo et al., 2012). CDK5RAP2 is critical for
interphase -TuRC recruitment to the centrosomes and depletion of CDK5RAP2 reduces -
tubulin localization and microtubule organization (Fong et al., 2008). In Drosophila, there is
evidence that the homolog of CDK5RAP2 (CNN) controls the size of the PCM by regulating the
rate of its incorporation. In flies lacking CNN expression, the amount of GFP-CNN expressed is
proportional to how fast GFP-CNN becomes incorporated into the PCM around centrioles, and to
the total amount of CNN that gets incorporated (Conduit et al., 2010). The incorporation of
CNN into the PCM occurs first with CNN appearing at the wall of the centriole, then migrating
outwards into the peripheral PCM (Conduit et al., 2010). In C. elegans, SAS-4 depletion leads to
shorter centrioles, and along with shorter centrioles a reduction in the amount of PCM observed
at the centrosomes (Kirkham et al., 2003; Schmidt et al., 2009; Tang et al., 2009a). When
centrioles are abnormally elongated, as in the case of SAS-4 overexpression, they accumulate
PCM along their lengths and this additional PCM is capable of nucleating microtubules
(Kohlmaier et al., 2009). In human cells, destruction of centrioles by antibody injection leads to
fragmentation of the PCM (Bobinnec et al., 1998a). Only mature centrioles are able to nucleate
PCM, and only after they have passed through mitosis successfully (Wang et al., 2011). When
de novo centriole assembly is induced, newly formed procentrioles are unable to nucleate
microtubule regrowth in interphase, and in mitosis are randomly segregated between daughter
cells when they lack attachment to a mature centriole (Wang et al., 2011). When procentriole
assembly is inhibited, the amount of PCM organized at each mitotic spindle pole is comparable
to that of a spindle pole with a centriole pair (Wang et al., 2011). Newly formed centrioles gain
the ability to nucleate PCM only after passage through mitosis and entry to G1 in the cell cycle,
and this requires the activity of PLK1 around the G2/M transition, but is independent of centriole
engagement (Wang et al., 2011). In sum, mature centrioles are critical to the organization of
PCM, and their length is also proportional to how much PCM they can recruit and ultimately
how they function as centrosomes.

17
1.4. The Centrosome Cycle
The duplication of centrioles and centrosomes is regulated throughout the cell cycle, with distinct
events predictably occurring in specific phases (Figure 1.2) (Vorobjev and YuS, 1982; Winey
and O’Toole, 2014). Following mitosis, a cell contains a pair of loosely connected (disengaged)
centrioles, capable of acting as sites for the formation of new centrioles (procentrioles) (Chrétien
et al., 1997; Firat-Karalar and Stearns, 2014). In S-phase, the procentrioles are formed and begin
to elongate (Chrétien et al., 1997; Kuriyama and Borisy, 1981). By G2, the cell has two parental
centrioles, each with a single tightly associated procentriole. These centrioles must separate to
become distinct centrosomes, and accumulate additional PCM, in preparation for the next mitotic
division. The steps of the centrosome cycle are described below.
1.4.1. Disengagement
When procentrioles assemble, they form in close association with the wall of the parental
centriole, and remain tightly anchored there throughout S phase and G2. Late in mitosis, each
parental centriole loses tight association with its procentriole, in a process known as
disengagement. Disengagement is regulated by PLK1 and the protease Separase (Firat-Karalar
and Stearns, 2014; Tsou and Stearns, 2006; Tsou et al., 2009). Separase is known to target the
Scc1 subunit of the Cohesin protein complex, and the cleavage of Cohesin by Separase is a key
step in mitosis that allows separation of sister chromatids (Firat-Karalar and Stearns, 2014; Hauf
et al., 2001). Separase cleavage of the cohesin subunit SCC1 is critical for centriole
disengagement in vitro using purified centrosomes, however, in vivo expression of a non-
cleavable SCC1 does not block centriole disengagement (Schöckel et al., 2011; Tsou et al.,
2009). Further, in the absence of Separase, disengagement can eventually occur during the
subsequent interphase (Tsou et al., 2009). Chemical inhibition of PLK1 in G2, but not later in
mitosis, inhibits centriole disengagement (Tsou et al., 2009). A combination of Separase
depletion and PLK1 inhibition in G2/early-mitosis strongly inhibited centriole disengagement
throughout S-phase of the following cell cycle, demonstrating that a combination of PLK1
activity early in mitosis with Separase activity are important for disengagement (Tsou et al.,
2009). The critical substrate of Separase in disengagement may in fact be Pericentrin, cleavage
of which by Separase is essential for disengagement (Lee and Rhee, 2012; Matsuo et al., 2012).
Pericentrin cleavage may facilitate removal of the PCM protein CDK5RAP2, as evident from the

18
fact that depletion of CDK5RAP2 allows disengagement in the presence of a non-cleavable
Pericentrin construct (Lee and Rhee, 2012; Matsuo et al., 2012; Pagan et al., 2014). Consistent
with this, loss of CDK5RAP2 function leads to premature disengagement and re-duplication of
centrioles in mouse embryonic fibroblast cells (MEFs) (Barrera et al., 2010). Thus centriole
engagement is maintained the PCM components Pericentrin and CDK5RAP2, and
disengagement requires Separase cleavage of Pericentrin along with the activity of PLK1 in early
mitosis.
1.4.2. Centrosome Separation
During S and G2 phase, the two parental centrioles remain connected by a linker that formed in
the preceding G1. In order for bipolar mitosis to occur, the connected parental centrioles must
separate to form two distinct centrosomes. Rootletin and LRRC45 form the linker and are
connected through their interacting protein C-Nap1 to CEP135 at the centrioles (Agircan et al.,
2014; Bahe, 2005; Fry et al., 1998; He et al., 2013; Kim et al., 2008; Mayor et al., 2000). This
linker is dissolved at the onset of mitosis to allow centrosome separation (or disjunction), after
which the two centrosomes can participate in bipolar spindle formation (Agircan et al., 2014).
The dissolution of the linker is regulated by NEK2 kinase, which phosphorylates the linker
proteins C-Nap1, as well as Rootletin and LRRC45, leading to their loss from the centrosomes
and centrosome separation (Agircan et al., 2014; Bahe, 2005; Fry et al., 1998; He et al., 2013).
The activity of NEK2 is regulated by Protein Phosphatase 1 (PP1). The PP1subunit interacts
with NEK2, and PP1 can de-phosphorylate NEK2 reducing its activity (Helps et al., 2000;
Meraldi and Nigg, 2001). NEK2 undergoes an activating auto-phosphorylation, and also
regulates PP1 through a deactivating phosphorylation (Helps et al., 2000; Meraldi and Nigg,
2001). PP1 can also reverse phosphorylation of C-Nap1 by NEK2, indicating that the complex
of NEK2 and PP1 works to regulate C-Nap1 phosphorylation (Helps et al., 2000; Meraldi and
Nigg, 2001). NEK2 localization to centrosomes and subsequent centrosome separation also
requires hSav1 and the kinase MST2, which phosphorylates NEK2 promoting its centrosomal
localization (Agircan et al., 2014; Mardin et al., 2010). The activity of PLK1 also regulates
NEK2 activity, where phosphorylation of MST2 by PLK1 prevents association of PP1 with the
complex of NEK2 and MST2, allowing NEK2 to promote centrosome separation at the onset of
mitosis (Agircan et al., 2014; Mardin et al., 2010, 2011). The physical separation of centrosomes

19
once the linker is dissolved is orchestrated by microtubule motor proteins, including Eg5 and
dynein (Agircan et al., 2014).
1.4.3. Centrosome Maturation
In preparation for mitosis, the PCM undergoes an increase in size, with the additional
recruitment of key PCM proteins important for microtubule nucleation and/or anchoring (Piehl et
al., 2004; Woodruff et al., 2014). The most important regulator of PCM maturation is the kinase
PLK1, which phosphorylates multiple PCM proteins and is required for PCM maturation (Haren
et al., 2009; Lane and Nigg, 1996; Woodruff et al., 2014). One of the key targets of PLK1 for
this process is Pericentrin, as its phosphorylation by PLK1 is critical for recruitment of PCM
proteins involved in microtubule nucleation including -tubulin, Nedd1, CEP192, Aurora-kinase-
A and PLK1 itself (Haren et al., 2009; Joukov et al., 2014; Lee and Rhee, 2011). In mitosis,
Pericentrin acts as a scaffold to which CEP192 is localized, and CEP192 in turn is a scaffold for
PLK1 and Aurora-A that facilitates PLK1 phosphorylation and activation by Aurora-A (Joukov
et al., 2014; Zhu et al., 2008). CEP192 is a substrate of PLK1 in this pathway, and its ability to
bind -tubulin and function in microtubule organization requires phosphorylation by PLK1
(Joukov et al., 2014). CEP192 is a critical component of the mitotic PCM, and is required for
Nedd1 and -tubulin recruitment and bipolar spindle formation (Gomez-Ferreria et al., 2007;
Joukov et al., 2014; Zhu et al., 2008). Nedd1 interacts with the -TuRC subunit GCP2 and -
tubulin, and is likewise required for mitotic spindle formation and centrosomal microtubule
nucleation (Haren, 2006; Lüders et al., 2006). Centrosome maturation is reversed by a reduction
of PCM protein concentrations at the centrosomes after metaphase, in a process thought to
involve phosphatases and microtubule motor driven fragmentation (Woodruff et al., 2014).
1.5. Centriole Duplication
Proteins required for the process of centriole duplication are the focus of this thesis. The
following section provides an in-depth view of the current understanding of the process of
centriole assembly and the conserved proteins involved. How centriole length is regulated is
also a central theme of this thesis, and thus positive and negative regulators of centriole length

20
are discussed. Finally, the regulatory mechanisms that control centriole duplication are covered
to provide the reader with a complete picture of the process of centriole assembly from which to
understand the results described in this thesis.
1.5.1. Procentriole Formation
The process of centriole assembly requires a core set of conserved proteins found in a variety of
organisms that have centrioles (Carvalho-Santos et al., 2010). In the model organism C. elegans,
a set of 5 core proteins was described as being critical for this process: Spd-2, Zyg-1, SAS-4,
SAS-5, SAS-6 (Delattre et al., 2004, 2006; Kemp et al., 2004; Kirkham et al., 2003; Leidel and
Gönczy, 2003; Leidel et al., 2005; O’Connell et al., 2001; Pelletier et al., 2006). The structural
details of this assembly pathway were described using electron microscopy following
fertilization in C. elegans oocytes (Pelletier et al., 2006). Procentriole assembly begins with the
recruitment of Spd-2 (Cep192 homolog) and Zyg-1 kinase to the sperm-donated centrioles
(Pelletier et al., 2006). Spd-2 localization is required for (but not dependent upon) recruitment of
Zyg-1, and both Spd-2 and Zyg-1 are required for recruitment of the SAS proteins (Pelletier et
al., 2006). The assembly of the procentrioles begins with the formation of a central tube, which
then elongates and increases in diameter before gaining “hooks” at sites where single
microtubules eventually assembled (Pelletier et al., 2006). Central tube formation was blocked
by depletion of SAS-5 or SAS-6 in this assay, whereas SAS-4 depletion allowed central tube
formation and elongation, but did not permit central-tube widening or the formation of hooks or
single microtubules around the central tube (Pelletier et al., 2006). Despite the unique structure
of C. elegans centrioles, the centriole assembly factors described in C. elegans have functional
homologs conserved in higher organisms (Carvalho-Santos et al., 2012).
The Zyg-1 kinase in C. elegans is essential for initiation of centriole duplication, though the
critical substrate is not known (Lettman et al., 2013; O’Connell et al., 2001; Pearson and Winey,
2010). The functional homolog of Zyg-1 in other species is the kinase SAK/PLK4 (Bettencourt-
Dias et al., 2005; Habedanck et al., 2005; Pearson and Winey, 2010). PLK4 localization to
centrioles requires CEP192 and another protein CEP152 (Cizmecioglu et al., 2010; Hatch et al.,
2010; Sonnen et al., 2013). Both CEP192 and CEP152 interact with PLK4 and are together
important for its recruitment to the mother centriole (Cizmecioglu et al., 2010; Hatch et al., 2010;
Sonnen et al., 2013). Human CEP152 and the Drosophila homolog Asterless (ASL) have both

21
been shown to interact with PLK4/SAK and CPAP/SAS-4, and to be critical for centriole
duplication (Blachon et al., 2008; Cizmecioglu et al., 2010; Dzhindzhev et al., 2010; Hatch et al.,
2010; Sonnen et al., 2013). CEP152 in turn depends on CEP192 and another protein required for
centriole duplication, CEP63, for its localization to centrioles (Brown et al., 2013; Kim et al.,
2013; Sonnen et al., 2013). CEP192, like Spd-2, is essential for centriole duplication (Delattre et
al., 2006; Kemp et al., 2004; Pelletier et al., 2004; Zhu et al., 2008). Both CEP192 and CEP152
interact with the cryptic polo-box-domain of PLK4 through a conserved sequence in their NTDs,
and the binding appears to be competitive and mutually exclusive (Cizmecioglu et al., 2010;
Hatch et al., 2010; Kim et al., 2013; Sonnen et al., 2013). The nature of the proposed
cooperation of CEP192 and CEP152 was initially obscure, as CEP152 depletion alone increased
PLK4 recruitment to mother centrioles, whereas CEP192 depletion decreased PLK4 recruitment,
but simultaneous depletion of both CEP152 and CEP192 was more effective than CEP192
depletion alone at preventing PLK4 recruitment (Kim et al., 2013; Sonnen et al., 2013). An
understanding of the regulation of PLK4 localization was only recently achieved through a
combination of crystallography and three-dimensional structured illumination microscopy (3D-
SIM) to study sub-resolution centrosomal domains and localizations (Park et al., 2014). PLK4 is
initially recruited to CEP192 in early G1 and forms a ring of similar diameter to CEP192 around
the centrioles, but when CEP152 is recruited later in G1/S, the PLK4 ring expands in diameter
(Park et al., 2014). When CEP152 is absent, the PLK4 ring stays smaller, indicating this
transition requires CEP152 (Park et al., 2014). Indeed, co-crystal-structures of the binding
domains of CEP192 and CEP152 with PLK4 show that CEP192-PLK4 and CEP152-PLK4
interactions are mutually exclusive, and CEP152-PLK4 binding is several-fold tighter than
CEP192-PLK4 binding (Park et al., 2014). Overall, CEP192 recruits and sequesters PLK4
before CEP152 arrives at the centriole and ‘steals’ PLK4 as a pre-requisite for initiation of
centriole duplication (Park et al., 2014).
PLK4 has multiple known substrates important for centriole duplication. STIL, the human
homolog of Ana2 and SAS-5, is a critical substrate of PLK4 in centriole duplication (Ohta et al.,
2014; Stevens et al., 2010; Vulprecht et al., 2012). As with SAS-5 and SAS-6 in C. elegans,
STIL and SASS6 are co-dependent for targeting to procentriole assembly sites, and STIL is
essential for centriole duplication in human cells (Ohta et al., 2014; Tang et al., 2011; Vulprecht
et al., 2012). STIL interacts with SASS6 through its STAN motif, and a PLK4 phosphorylation

22
appears to be critical for this interaction (Ohta et al., 2014). When the residue is substituted to
alanine, SASS6 is not recruited to procentriole assembly sites, whereas STIL harboring a
phosphorylation-mimicking residue at S1061 can interact in vitro with SASS6 and facilitate its
delivery to procentriole assembly sites in vivo (Ohta et al., 2014). Endogenous PLK4 initially
appears as a ring around centrioles in G1, but later (G1/S) becomes focused at a single spot
where a procentriole will assemble (Ohta et al., 2014; Park et al., 2014). When STIL or SASS6
are depleted, or the proteasome is inhibited by MG132, PLK4 remains ring-like and fails to focus
to a single spot (Ohta et al., 2014). Overexpression of STIL, or STIL that lacks the SASS6-
interaction domain, can also protect PLK4 rings around the mother centriole, leading to the
model that STIL can prevent PLK4 degradation (Ohta et al., 2014). Consistent with this,
overexpression of STIL inhibits PLK4 ubiquitination in vivo (Ohta et al., 2014). Similarly, in
Drosophila, SAK phosphorylation of Ana2 is required for SAS-6 localization, the SAS-6/Ana2
interaction, and centriole duplication (Dzhindzhev et al., 2014). Thus one conserved critical
substrate for PLK4 in centriole duplication is STIL, and a PLK4-phosphorylation dependent
interaction of STIL with SASS6 plays a critical role in limiting PLK4 localization and centriole
duplication to a single procentriole assembly site (Dzhindzhev et al., 2014; Ohta et al., 2014). In
C. elegans, the interaction of SAS-6 and SAS-5 has been studied in vitro, and the association of
SAS-5 prevents abnormal tetramers of SAS-6 from forming through its coiled-coil domains
(Qiao et al., 2012). Further, in vitro SAS-5 and SAS-6 complexes form partial rings that are of a
diameter consistent with the C. elegans central tube, consistent with SAS-5 and SAS-6
cooperating to form the tube (Pelletier et al., 2006; Qiao et al., 2012).
PLK4 also phosphorylates the-TuRC component GCP6, and the -TuRC is required for
centriole duplication (Bahtz et al., 2012; Kleylein-Sohn et al., 2007). When HeLa cells are
depleted of GCP6 the PCM has reduced amounts of -tubulin, and -TuRC structure, mitotic
spindle formation and centriole duplication are impaired (Bahtz et al., 2012). PLK4 can be used
to co-IP GCP6, and PLK4 interacts with and phosphorylates GCP6 in vitro (Bahtz et al., 2012).
Twenty residues were identified in GCP6 that were phosphorylated by PLK4 in vitro, and
mutants of GCP6 that have all 20 serine residues substituted for alanines are unable to rescue
centriole duplication defects (Bahtz et al., 2012). The 20-alanine mutant GCP6 localizes to the
centrosome and rescues the localization of -tubulin to the centrosome, so PLK4 phosphorylation

23
of GCP6 specifically regulates -TuRC function, but not -TuRC assembly (Bahtz et al., 2012).
Therefore -TuRC regulation is another important aspect of how PLK4 regulates centriole
duplication.
The conserved protein SASS6 is the structural component of the cartwheel hub of centrioles,
conferring nine-fold symmetry to centrioles through formation of a nonamer of dimers (van
Breugel et al., 2011; Kitagawa et al., 2011). Disengagement of two centrioles in late mitosis or
early G1 is a requisite for centriole duplication in the subsequent S-phase (Kuriyama and Borisy,
1981; Tsou and Stearns, 2006). In early S-phase, there is a small but detectable population of
SASS6 initially recruited to the proximal lumen of the existing disengaged centrioles (Fong et
al., 2014). In super-resolution microscopy experiments, this population of SASS6 appears to be
ring-like and polarized from N to C termini, suggesting the strong likelihood of SASS6
cartwheels forming anew in the lumen of the parental centrioles in S-phase (Fong et al., 2014).
Critically, this population of SASS6 is lost in S-phase, when SASS6 accumulates outside the
mother centrioles where the procentrioles are assembling (Fong et al., 2014). The transition of
SASS6 from the parental lumen to the procentriole site requires the presence of both PLK4 and
STIL, and when either are absent the SASS6 population within the parental lumen persists
throughout S-phase (Fong et al., 2014). The de novo re-formation of cartwheels of SASS6
within the lumen of mother centrioles is possible only when centrioles are disengaged, and this is
hypothesized to be part of the reason engagement of centrioles inhibits re-duplication during the
cell cycle (Fong et al., 2014). Taken together with the evidence that PLK4 initially forms a ring
around parental centrioles in G1 before a STIL-and-SASS6 dependent transition to a single focus
at the site of procentriole assembly in S-phase, this work provides an appealing model for how a
single procentriole is formed adjacent to the mother centriole per cell cycle (Fong et al., 2014;
Ohta et al., 2014). New questions arise, such as whether the SASS6 cartwheel is directly moved
from the lumen to the side wall without disassembly, and how can such a transition be
experimentally captured in real-time (Fong et al., 2014)? How many cartwheels are assembled
within the parental centriole lumen, and what is the ‘trigger’ mechanism for their release from
the parental centrioles (Fong et al., 2014)? These questions are challenging but important to
address in pursuit of a complete understanding of centriole duplication.

24
Human CEP135 is a largely conserved protein required for human centriole duplication
downstream of SASS6, although there is no homolog in C. elegans (Carvalho-Santos et al.,
2010; Kleylein-Sohn et al., 2007; Lin et al., 2013a). When CEP135 is depleted, microtubule
organization by interphase centrosomes is disrupted, and monopolar or abnormal mitotic spindles
are observed (Ohta, 2002). In Chlamydomonas and Tetrahymena thermophila, the CEP135
homolog Bld10 forms part of the connection between the cartwheel spokes and the microtubule
triplets (Bayless et al., 2012; Hiraki et al., 2007; Matsuura, 2004). In Chlamydomonas, when
truncated Bld10 constructs are expressed, shorter cartwheel spokes and loss of connection
between spokes and microtubule triplets are observed (Hiraki et al., 2007). Likewise,
Drosophila Bld10 localizes to the cartwheel spokes, and mutant flies lacking CEP135/Bld10
have defective cartwheel attachments to microtubule triplets, although centrioles can form
(Roque et al., 2012). In humans, CEP135 interacts with SASS6 and CPAP/SAS-4 and has a
microtubule binding domain (Lin et al., 2013a). CEP135 localizes along the centriolar
microtubules, and within the proximal and distal portions of the lumen of centrioles and
procentrioles (Kleylein-Sohn et al., 2007; Lin et al., 2013a). The localization of CEP135 is thus
consistent with the hypothesis that it forms part of the cartwheel spoke tip, but may have
additional function in the stability of the centriole.
Subsequent to central tube formation, the assembly of microtubules in C. elegans procentrioles
requires SAS-4 (Pelletier et al., 2006). Likewise, the SAS-4 homolog CPAP is also required for
procentriole formation in humans (Kleylein-Sohn et al., 2007). CPAP interacts directly with
STIL in vivo and in vitro, and the interaction involves the CTD of CPAP (residues 895-1338)
and the NTD of STIL (residues 281-781) (Tang et al., 2011). Both SASS6 and STIL are
required for CPAP localization to procentrioles during S-phase, but CPAP is not required for
SASS6 or STIL recruitment (Cottee et al., 2013; Kleylein-Sohn et al., 2007; Tang et al., 2011).
CPAP also directly binds CEP135, and depletion of CEP135 appears to prevent the localization
of CPAP when CPAP is overexpressed (Lin et al., 2013a). The same region of CPAP appears to
be required for the CEP135 and STIL interactions (residues 895-1338), and the NTD of CEP135
(residues 50-460) is important for its interaction with CPAP (Lin et al., 2013a). CPAP
localization to both mature centrioles and procentrioles also requires Centrobin, another protein
essential to human centriole duplication (Gudi et al., 2014; Zou et al., 2005). Thus, CPAP
recruitment during centriole duplication is downstream of CEP152, PLK4 SASS6, STIL, and

25
Centrobin, while its localization to centrosomes when overexpressed also appears to depend on
CEP135 (Cizmecioglu et al., 2010; Hatch et al., 2010; Kleylein-Sohn et al., 2007; Lin et al.,
2013a). The interactions of CPAP suggest that along with CEP135 and STIL it occupies a
region connecting SASS6 cartwheel spokes to the microtubule triplets (Lin et al., 2013a). This
idea is consistent with the recent evidence that SASS6 recruitment to the parental centriole
lumen in G1 requires CPAP (Fong et al., 2014).
Lastly, CP110 was identified as a CDK2 substrate required for centriole duplication, and further
characterized as one of the proteins required for procentriole formation following PLK4
overexpression in human cells (Chen et al., 2002; Kleylein-Sohn et al., 2007). In the human
centriole duplication pathway, CP110 is not recruited when CPAP or CEP135 are absent, placing
it downstream in terms of recruitment dependencies (Kleylein-Sohn et al., 2007). CP110 is
localized to the distal portion of centrioles and procentrioles, and it acts like a “cap” on the
microtubules under which procentriole microtubules elongate (Kleylein-Sohn et al., 2007). The
process of procentriole assembly is summarized in Figure 1.3. There are numerous other
proteins required for centriole duplication, and the discovery and characterization of novel
proteins required for this process is an area of intensive research.
1.5.2. Procentriole Elongation and Length Regulation
Following formation in S-phase, procentrioles continue to elongate during in G2, and only reach
full length in mitosis (Azimzadeh and Marshall, 2010; Brito et al., 2012; Chrétien et al., 1997).
The human centriole is typically described as reaching a length of ~500 nm at maturity, with an
outer diameter of ~250 nm (Gönczy, 2012; Jana et al., 2014). In reality, “~500 nm” represents
an average of a range of lengths, as mature single centrioles isolated from KE37 human
lymphoblasts measure 512 ± 53 nm in length (n=165) (Chrétien et al., 1997). Overall though,
this shows a remarkable conservation of length. Several proteins are known to be required for
centriole elongation, and these are counter-balanced by other proteins that restrict the length of
the centriole.
CPAP plays a pivotal role in promoting centriole elongation, and excess CPAP leads to centriole
elongation beyond the normal length (Jana et al., 2014; Kohlmaier et al., 2009; Schmidt et al.,
2009; Tang et al., 2009a). This activity is partly regulated by PLK2 phosphorylation at two

26
specific serine residues, as neither overexpression of a mutated CPAP where these residues are
replaced with alanine residues nor depletion of PLK2 followed by overexpression of CPAP can
drive centriole elongation (Chang et al., 2010). Centrobin is also required for CPAP driven
centriole elongation (Gudi et al., 2011). Similar to CPAP, Centrobin can drive abnormal
centriole elongation when overexpressed (Gudi et al., 2015). Given that Centrobin is required
for CPAP centriolar-localization, and when overexpressed raises both overall CPAP levels and
the concentration of CPAP at centrosomes, it is probable that Centrobin affects centriole length
partly through regulating CPAP levels (Gudi et al., 2014, 2015). Alternatively, Centrobin has a
tubulin-dimer binding domain and could play a direct role in centriole stabilization and/or
tubulin incorporation during elongation (Gudi et al., 2011). The relative importance of
Centrobin tubulin binding activity versus CPAP regulation in centriole elongation is not yet
clear. CPAP also has two tubulin binding domains, called PN2-3 and A5N (Hsu et al., 2008).
The A5N domain binds to polymerized microtubules, whereas the PN2-3 domain binds tubulin
dimers and microtubule plus-ends, prevents microtubule polymerization, and can destabilizing
pre-formed taxol-stabilized microtubules in vitro (Hsu et al., 2008; Hung et al., 2004). High-
levels of full length CPAP overexpression can sequester tubulin leading to disruption of the
microtubule skeleton (Hsu et al., 2008). CPAP carrying a mutation in the PN2-3 that inhibits
tubulin binding does not drive abnormal centriole elongation, and is not functional in centriole
duplication (Tang et al., 2009a). Thus the PN2-3 domain may not be acting as a microtubule
destabilizing domain in the context of centriole elongation, or its destabilizing effects may be
required for elongation through some unknown mechanism. In Drosophila, SAS-4 has been
shown to bind PCM components and help incorporate them into centrosomes, in a process
requiring the PN2-3 domain and regulated by tubulin binding (Gopalakrishnan et al., 2011,
2012). In C. elegans, SAS-4 was initially described as regulating PCM size at centrosomes, but
this observation was probably the result of shorter centrioles following SAS-4 depletion
(Kirkham et al., 2003). The exact mechanisms by which CPAP and Centrobin cooperate with
each other and utilize their respective tubulin interactions to promote centriole elongation
remains unclear (Gönczy, 2012). Whether CPAP is involved in PCM recruitment or PCM
anchoring to centrioles in organisms other than Drosophila has not been investigated, though it
has been reported that a population of CPAP exists within the outer-regions of the PCM of

27
human centrosomes in addition to the population within the centriole lumen (Sonnen et al.,
2012).
In addition to cartwheel-microtubule interactions, CEP135 plays a role in microtubule
stabilization. The microtubule binding domain of Drosophila Bld10, which is within the NTD of
Bld10 as it is in Homo sapiens CEP135, can stabilize microtubules when expressed alone as a
truncated peptide (Carvalho-Santos et al., 2012; Lin et al., 2013a). Conversely, loss of Bld10
disrupts spindle formation and results in loss of flagella formation with abnormal basal bodies in
Chlamydomonas (Bayless et al., 2012; Matsuura, 2004). CEP135 localizes to both the proximal
and distal portions of the lumen in centrioles and T. thermophila basal bodies (Bayless et al.,
2012; Kleylein-Sohn et al., 2007). Drosophila mutants lacking Bld10 have shorter centrioles and
spermatid basal bodies, and are incapable of forming the ‘central pair’ of microtubules normally
found in motile cilia and flagella (Carvalho-Santos et al., 2012; Mottier-Pavie and Megraw,
2009). Consistent with a role in microtubule stability, depletion of Bld10 in T. thermophila leads
to loss of already-assembled basal bodies over time due to the mechanical stress of ciliary
beating (Bayless et al., 2012). Depletion of CEP135 appears to prevent the localization of CPAP
and centriole elongation when CPAP is overexpressed (Lin et al., 2013a). In summary,
Bld10/CEP135 is required for cilia/flagella and centriole microtubule stability, and plays a role
in centriole elongation (Bayless et al., 2012; Carvalho-Santos et al., 2012; Hiraki et al., 2007;
Kleylein-Sohn et al., 2007; Matsuura, 2004; Mottier-Pavie and Megraw, 2009; Roque et al.,
2012).
Poc1 is essential for centriole duplication, and its overexpression drives centriole elongation in
S-phase, in contrast to CPAP which is G2 specific (Keller et al., 2009). Two lines of evidence
suggest that the role for Poc1 in centriole length may relate to stabilizing of centriolar or ciliary
microtubules. First, it has been demonstrated that depletion of Poc1 in T. thermophila leads to
formation of unstable basal bodies (Pearson et al., 2009). Second, depletion of Poc1 in human
cells leads to immature procentrioles that lack acetylation (Venoux et al., 2013). There are two
Poc1 paralogs, called Poc1A and Poc1B (Keller et al., 2009). Each of these proteins is
independently capable of centrosome localization and functioning in centriole assembly, and
only co-depletion of both blocks this process (Venoux et al., 2013).

28
hPOC5 is one of two proteins that appear to drive centriole elongation specifically within distal
portion of the procentriole, and it is recruited to nascent centrioles in G2 (Azimzadeh et al.,
2009). hPOC5 interacts with Centrin, and co-localizes with Centrin within the distal portion of
the centriole (Azimzadeh et al., 2009). In the absence of hPOC5, cells do form procentrioles that
lack hPOC5 but contain Centrin and recruit PCM (Azimzadeh et al., 2009). In HeLa cells
depletion of hPOC5 leads to an S-phase arrest, but the procentrioles formed are shorter than
procentrioles in otherwise S-phase arrested controls and are made up of only doublet
microtubules (Azimzadeh et al., 2009). The authors offer two explanations for how hPOC5
affects procentrioles in S-phase despite not being detected on S-phase procentrioles; it is possible
that hPOC5 is present in S-phase at undetectable levels, or hPOC5 participates in some extra-
centriolar regulation of procentriole assembly related to its essential role in cell-cycle
progression (Azimzadeh et al., 2009). Overall, hPOC5 is an essential protein that is required for
procentriole microtubule assembly beginning in S-phase, and it appears to be critical for the
formation of the C-microtubule and the distal part of the procentriole (Azimzadeh et al., 2009).
OFD14 (formerly C2CD3) was identified as mutated in oral-facial-digital/orofacial-digital
syndrome patients, and is the other protein known to drive abnormal elongation of the distal
portion of the centrioles when overexpressed (Thauvin-Robinet et al., 2014). OFD14 is critical
for cilia formation, being required for both distal and subdistal appendage formation on mature
centrioles (Hoover et al., 2008; Thauvin-Robinet et al., 2014). OFD14 depletion results in loss
of Centrin and OFD1 from centrioles, and the centrioles remain shorter (~300 nm) (Thauvin-
Robinet et al., 2014). Consistent with a role in distal centriole assembly, OFD14 localizes in the
region near the distal appendage marker CEP164, where it interacts with OFD1 (Thauvin-
Robinet et al., 2014).
Prolonged chemical inhibition of proteolysis within a cell has been shown to lead to abnormal
centriole elongation, and this discovery was leveraged in an assay to look for proteins required
for centriole elongation (Korzeniewski et al., 2010). This approach identified CAP350, FOP,
CEP170, C-Nap1 and SASS6 as being required for this to occur (Korzeniewski et al., 2010).
While the presence of CEP170, C-Nap1, Ninein and SASS6 as hits in this screen suggest that
some of the proteins found are only indirectly required for centriole elongation, CAP350 has
since been shown to be important for centriole microtubule stability (Le Clech, 2008;

29
Korzeniewski et al., 2010). It seems likely that other proteins will be identified that are required
for centriole elongation, and these will be important for furthering our understanding of the
intricate regulation of centriole elongation that produces consistent centriole lengths in each
organism.
CP110, and its interacting partner CEP97, are critical for restricting the length of centrioles
(Spektor et al., 2007). CP110 and CEP97 are co-dependent for centriole localization, and
depletion of either leads to elongation of the centrioles (Spektor et al., 2007). CP110 decorates
the distal tips of centrioles, and even elongated centrioles when CPAP is overexpressed
(Kleylein-Sohn et al., 2007; Schmidt et al., 2009). Initially it was reported that cilia formation
occurred upon depletion of CP110 or CEP97, however, the elongated structures observed upon
CP110 depletion are actually abnormal centrioles (Schmidt et al., 2009). The abnormal
elongation of centrioles upon CEP97 depletion can be inhibited by simultaneous overexpression
of KIF24, a microtubule de-polymerizing kinesin that acts specifically on centriolar microtubules
(Kobayashi et al., 2011). Consistent with their role in restricting centriole length, it is clear that
CP110 and CEP97 are absent from basal bodies, and when localized there by overexpression
they will suppress cilia formation (Spektor et al., 2007). How exactly CP110 and CEP97
cooperate to regulate tubulin incorporation, or how the cell ‘measures’ the length of a centriole to
ensure uniformity, remain important open questions.
Like CP110/CEP97, depletion of OFD1 gives abnormal centriole elongation in mouse embryonic
stem cells (Singla et al., 2010). OFD1 localizes to both mature centrioles and procentrioles, in
the distal portion of each, near CP110 and hPOC5 (Singla et al., 2010). Depletion of OFD1 only
affects the lengths of the parental centrioles (not procentrioles), leading to elongation of the
distal portion of the centrioles in G2 specifically, as indicated by an expanded region of hPOC5
localization (Singla et al., 2010). The distal structure of the mother centriole is also affected in
that subdistal appendage proteins localize along an unusually long portion of the centriole, while
distal appendages do not form at all (Singla et al., 2010). Remarkably, the distal centriole
elongation observed upon overexpression of OFD14 is reversible by simultaneous
overexpression of OFD1, suggesting that these two proteins are antagonistically striking a
balance to establish the correct distal centriole length and facilitate appendage formation
(Thauvin-Robinet et al., 2014). The loss of OFD1 correlates with a loss of centriolar

30
microtubule glutamylation, which is unexpected because glutamylation is associated with
microtubule/centriole stability (Bobinnec et al., 1998a; Janke and Bulinski, 2011). The fact that
centrioles without OFD1 elongate abnormally in their distal regions but are not glutamylated
may be explained by a lag between elongation and stabilization, or a requirement for OFD1 in
recruiting glutamylation enzymes. Resolving this contradiction will be important for furthering
our understanding of centriole length regulation and stabilization by glutamylation.
In summary, a variety of proteins positively and negatively regulate centriole elongation (Figure
1.3), and likely more remain to be discovered. These proteins can act on distinct domains within
the centriole (proximal, distal), and imbalances between them can have dramatic effects on
centriole length, structure, mitotic function and capability in cilia formation. Several of these
proteins, such as CPAP (MCPH6), OFD1 and OFD14 are affected in diseases (Barbelanne and
Tsang, 2014; Gul et al., 2006; Singla et al., 2010; Thauvin-Robinet et al., 2014). Thus,
understanding how key proteins function in centriole length regulation may provide important
insights into human developmental diseases.
1.5.3. The Regulation of Centriole Duplication
The levels of a number of centriole duplication proteins have been shown to be cell cycle
regulated. This is achieved largely through proteolysis ensuring their presence at appropriate
levels at during the proper cell cycle stage. Careful regulation of their levels is critical to
ensuring formation of one and only one procentriole per parental centriole in a cell cycle. The
degradation of these proteins is mediated by ubiquitin-dependent proteolysis. Ubiquitin ligases
catalyze the transfer of ubiquitin molecules to specific substrate proteins within the cell, and
ubiquitination can make these proteins substrates for degradation by the 26S proteasome (Finley,
2009; Voges et al., 1999). The anaphase promoting (APC) and Skp, Cullin,-F-box (SCF)
complexes are ubiquitin ligase complexes that regulate a variety of key cellular proteins, and
play critical roles in the cell cycle (Cardozo and Pagano, 2004; Silverman et al., 2012). The SCF
E3 ubiquitin ligase complexes are formed on a scaffold of CUL1, which brings RBX1 and an E2
(ubiquitin conjugating enzyme) together with Skp1 and an adaptor protein of the F-box family
(Cardozo and Pagano, 2004; Silverman et al., 2012). F-box proteins act as the substrate
specificity factors, and so each one has specific protein targets, including key regulators of the
cell cycle and centriole duplication (Cardozo and Pagano, 2004; Silverman et al., 2012; Skaar et

31
al., 2013). The APC consists of 19 subunits at its core, which bring together an E2 with one of
two adaptor molecules, CDC20 or Cdh1 (Peters, 2006; Sivakumar and Gorbsky, 2015). CDC20
and Cdh1 have both been implicated in recognition and regulation of centriole duplication
proteins (Arquint and Nigg, 2014; Arquint et al., 2012). Both the APC and SCF E3 ubiquitin
ligase complexes can regulate centriole duplication through controlling the stability of core
centriole assembly proteins.
SASS6 is normally degraded during late-mitosis/early-G1 following ubiquitination by the APC-
Cdh1 E3 ubiquitin ligase complex, and is undetectable in early G1 at centrosomes (Arquint and
Nigg, 2014; Puklowski et al., 2011; Strnad et al., 2007). A SASS6 mutant construct lacking its
KEN domain, the target for Cdh1, drives excess centriole formation when overexpressed (Strnad
et al., 2007). Conversely, SASS6 becomes ubiquitinated when Cdh1 is overexpressed
(Puklowski et al., 2011; Strnad et al., 2007). Levels of STIL also decrease in late-mitosis and
G1, and this is likewise achieved by the activity of the APC and critical for limiting centriole
duplication (Arquint and Nigg, 2014; Arquint et al., 2012). SASS6 is also a substrate of the SCF
E3 ubiquitin ligase complex through its interaction with the SCF adaptor FBXW5 (Puklowski et
al., 2011). Overexpression of FBXW5 causes increased ubiquitination of SASS6, leading to
degradation of SASS6 and inhibition of centriole duplication (Puklowski et al., 2011). FBXW5
itself is regulated by APC dependent ubiquitination (Puklowski et al., 2011). To ensure FBXW5
levels are low during G1, the APC-Cdh1 and APC-CDC20 complexes ubiquitinates FBXW5 to
promote its degradation (Puklowski et al., 2011).
PLK4 induces centriole duplication partly by negatively regulating FBXW5 (Puklowski et al.,
2011). FBXW5 is kept at low levels in mitosis and G1 through APC regulation, but at G1/S it
appears PLK4 phosphorylation may become critical for inhibiting FBXW5 to stabilize SASS6
for centriole duplication (Puklowski et al., 2011). When co-overexpressed with FBXW5, PLK4
protects SASS6 from degradation, and the kinase activity of PLK4 is required for this effect
(Puklowski et al., 2011). FBXW5 is phosphorylated by PLK4 in vitro, and when the target
serine (151) of FBXW5 is mutated to alanine, the resulting FBXW5 ubiquitinates SASS6
regardless of PLK4 activity (Puklowski et al., 2011). Finally, the loss of centriole duplication
upon PLK4 RNAi is partially rescued by co-depletion of FBXW5, indicating that PLK4

32
regulation of SASS6 stability through FBXW5 de-activation is a critical aspect of centriole
duplication (Puklowski et al., 2011).
Work in Drosophila and later in human cells showed that SAK (Drosophila PLK4) and PLK4
were both negatively regulated by the SCF-complex with the F-box protein Slimb (Drosophila)/
TrCP (Homo Sapiens) (Cunha-Ferreira et al., 2009; Guderian et al., 2010; Rogers et al., 2009).
Accordingly, depletion of Slimb/TrCP drives centriole amplification in both systems, with
elevated SAK/PLK4 protein levels and an increase in centrosome localized SAK/PLK4 (Cunha-
Ferreira et al., 2009; Guderian et al., 2010; Rogers et al., 2009). The targeted degradation of
PLK4 by SCF-Slimb/-TrCP depends on the phosphorylation of each SAK/PLK4 molecule by a
second molecule of SAK/PLK4 in trans (Cunha-Ferreira et al., 2013; Guderian et al., 2010). In
both Drosophila and Homo sapiens, SAK/PLK4 has a conserved phosphodegron with a pair of
residues (S293, T297 in Drosophila,S285 and T289 in Homo sapiens) that along with
phosphorylation sites outside the degron are targets of trans auto-phosphorylation and required
for the interaction with Slimb/-TrCP (Cunha-Ferreira et al., 2013; Guderian et al., 2010; Klebba
et al., 2013). Mutation of the two critical residues in the phosphodegron to alanines prevents the
Slimb/-TrCP interaction, drives centriole overduplication and leads to elevated SAK/PLK4
levels (Cunha-Ferreira et al., 2013; Guderian et al., 2010). In Drosophila and C. elegans, the
phosphorylation of PLK4 is further regulated by protein-phosphatase-2A, which stabilizes PLK4
during mitosis by de-phosphorylating the phosphodegron, allowing PLK4 to be present on
mitotic centrosomes at the same time as SCF-Slimb without being degraded (Brownlee et al.,
2011; Rogers et al., 2009; Song et al., 2011).
The ubiquitin ligase MIB1 is also a regulator of PLK4 stability and function (Cajanek et al.,
2015). MIB1 is localized to the centrosome during S-phase, and enriched there following PLK4
overexpression (Cajanek et al., 2015). When overexpressed, MIB1 drives ubiquitination of
PLK4, and this can occur regardless of -TrCP expression (Cajanek et al., 2015). Depletion of
MIB1 does not cause centriole overduplication in U-2 OS cells the way TrCP depletion does,
but MIB1 overexpression does prevent PLK4 overexpression-induced centriole overduplication
and reduces PLK4 levels in this context (Cajanek et al., 2015). Further, PLK4 ubiquitination by
MIB1 reduced the binding of PLK4 to both CEP192 and CEP152, indicating that in addition to
regulation by degradation, MIB1 ubiquitination of PLK4 can regulate its interactions during

33
centriole duplication (Cajanek et al., 2015). Thus MIB1 along with -TrCP helps regulate PLK4
levels and function in centriole duplication (Cajanek et al., 2015).
PLK4 can positively regulate its own activity in centriole duplication through another
phosphorylation at serine 305 (Sillibourne et al., 2010). A PLK4 peptide where serine 305 is
substituted with glutamic acid to mimic phosphorylation promotes centriole overduplication
more efficiently than wild-type PLK4 when overexpressed (Sillibourne et al., 2010). PLK4
constructs that mimic (S305E) or inhibit (S305A) phosphorylation do not differ in their stability
from wild-type PLK4 when overexpressed, indicating that this phosphorylation is not regulating
PLK4 levels (Sillibourne et al., 2010). When a non-degradable PLK4 construct is overexpressed
it partitions both the soluble and insoluble (centrosome containing) fractions of cell lysates, but
S305-phosphorylated PLK4 is only detectable in the insoluble fractions, suggesting PLK4
phosphorylated at S305 is sequestered to the centrosome (Sillibourne et al., 2010). Therefore
PLK4 can also regulate its own activity through phosphorylation, independent of promoting
degradation.
CPAP is a cell cycle regulated protein, with reduced levels in late mitosis and G1 (Tang et al.,
2009a). The APC co-factor Cdh1 interacts with CPAP, which is ubiquitinated in vivo, and
proteasome inhibition by MG132 stabilizes CPAP throughout the cell cycle (Tang et al., 2009a).
CP110 is likewise regulated during the cell cycle by the SCF-FBXO1/Cyclin-F ubiquitin ligase
complex, so that CP110 levels are decreased beginning in G2 (D’Angiolella et al., 2010). When
a CP110 mutant is expressed that is not targeted by the SCF complex, cells are observed with
extra (more than 4) centrioles (D’Angiolella et al., 2010). The ubiquitination of CP110 by SCF-
FBXO1 is counter-acted by the activity of a deubiquitinating (DUB) enzyme called USP33,
which interacts with CP110, and is present on centrosomes during S and G2 (Li et al., 2013).
When FBXO1 is overexpressed, CP110 becomes ubiquitinated, and this is reversed by
simultaneous overexpression of USP33 (Li et al., 2013). USP33 can also de-ubiquitinate CP110
in vitro (Li et al., 2013). Lastly, when FBXO1 is depleted CP110 accumulates at centrosomes
and centriole overduplication is observed (D’Angiolella et al., 2010; Li et al., 2013). When
USP33 is simultaneously depleted, CP110 levels are reduced at centrosomes and centriole
overduplication is prevented, an affect attributed to residual FBXO1 in the cells following

34
depletion (Li et al., 2013). Therefore, in addition to ubiquitin ligases, DUBS can play a role in
regulating centriole duplication factors.
1.6. CEP120 and SPICE1
Shortly after joining the lab, I was given the opportunity to collaborate on a study of CEP120 as
a potential centriole duplication protein (Hutchins et al., 2010). Prior to this, one function of
CEP120 had been described (Xie et al., 2007). CEP120 was shown to be a centrosomal protein
highly expressed in the brains of mouse embyros (Xie et al., 2007). Further, CEP120 was
important for a process called interkinetic nuclear migration (INM), a microtubule dependent
nuclear migration between opposite ends of a polarized cell during the cell cycle, and loss of
CEP120 led to depletion of the neural progenitor cells (Xie et al., 2007). A yeast-2-hybrid
screen for interactors of CEP120 led to a validated interaction between CEP120 and the
transforming acidic coiled-coil proteins (TACC-2B, TACC3) which are also important for INM
(Xie et al., 2007). CEP120 or TACC3 overexpression increased the size of the microtubule
asters from centrosomes, whereas depletion of CEP120 or TACC3 reduced the size of
microtubule bundles linking the centrosomes to the nuclei in cultured neurons and increased the
distances between the nuclei and the centrosomes in mouse embryonic neural stem cells (Xie et
al., 2007). These results were taken as supportive of a role for CEP120 in centrosome and
microtubule function (Xie et al., 2007). I demonstrated that CEP120 was required for centriole
duplication, and my collaborators demonstrated that CEP120 had an interaction with a protein
called CCDC52, now known as SPICE1(Hutchins et al., 2010). Otherwise, at the beginning of
my Ph.D. nothing was known about how either protein functioned in centriole duplication.
During my time in the lab, a paper came out describing CEP120 function in greater detail
(Mahjoub et al., 2010). This paper supported my conclusion that CEP120 was required for
centriole duplication, and added the insights that CEP120 was preferentially localized to
daughter centrioles in G1, was found on the microtubule walls of centrioles, and was dependent
on its coiled-coil domain for centriole localization (Mahjoub et al., 2010). Subsequent to the
CEP120 paper, another paper from our collaborators in the Luders lab showed that CCDC52,
which they renamed SPICE1, was a centriole and spindle associated protein with separable roles
in centriole duplication and mitotic spindle assembly (Archinti et al., 2010). SPICE1 contains

35
two coiled coil domains, the importance of which was investigated. The N-terminal portion of
SPICE1 (1-450), containing one of two coiled-coil domains, was nuclear in interphase, and
spindle and centriole localized in mitosis (Archinti et al., 2010). The central fragment (350-750)
having both coiled-coil domains of SPICE1, and localized to the centrosome but not the spindle
(Archinti et al., 2010). The C-terminal portion, containing only the second coiled-coil domain
(444-856), did not localize properly to either centrioles or the spindle (Archinti et al., 2010).
Overall, these papers advanced our understanding of how CEP120 and SPICE1 localized, and
validated their roles in centriole duplication.
1.7. Rationale of this Thesis
Answering the question of how CEP120 and SPICE1 participated in centriole duplication was
the first aim of my research. Were they cooperating in the process? Where were they acting
within the centriole assembly pathway? What aspect of centriole structure/assembly were they
important for? What were their interacting partners (if any) in this process? These important
questions formed the rationale for the first data chapter of this thesis. As part of our publication
of a paper addressing these questions (Comartin et al., 2013), we used a screen to identify
proteins that were in the proximity of CEP120 in vivo. This screen yielded a surprising variety
of proteins, some with no or poorly characterized functions. Since then, we have expanded our
search to include proteins in the proximity of SPICE1. The rationale of my second data chapter
was that by investigating proteins identified in the proximity of CEP120 and SPICE1, I would
find proteins required for centriole duplication. Further, within these I might uncover proteins
that functionally cooperate with CEP120/SPICE1 in centriole duplication, centriole elongation,
or mitosis. The overall goal of the thesis presented here is to advance the understanding of
centriole assembly through characterization of proteins critical for this process.

36
1.8. Figures
Figure 1.1

37
Figure 1.1. Structure of the Human Centriole.
(A) A mature mother centriole surrounded by PCM (blue) with distal and subdistal appendages is
shown with an associated procentriole. Note the procentriole does not accumulate PCM proteins
until after passage through mitosis(Wang et al., 2011). The known distal and subdistal
appendage proteins are listed (see text for discussion). (B) (Centre) Schematic of the cross-
sectional view of the proximal end of a procentriole containing a central cartwheel structure and
nine microtubule triplets. (Left) The central hub of the cartwheel and its spokes are made from a
set of nine dimers of SAS-6 (van Breugel et al., 2011; Hirono, 2014; Kitagawa et al., 2011;
Winey and O’Toole, 2014). The cartwheel dimensons, SASS6 dimer dimension, and model are
based on the report of the crystal structure of Chlamydomonas reinhardtii SAS-6 (Kitagawa et
al., 2011). (Right) Two microtubule triplets are shown, with A, B and C microtubules, and the
A-C linker shown (red)., based on cryotomography of Trichonympha centrioles (Diagram based
on Figure 5, Guichard et al., 2013). Each circle of the microtubule represents a protofilament, of
which the A tubule has 13, and the B and C tubules each contain 10 (Guichard et al., 2013;
Winey and O’Toole, 2014). The A-C linker connects protofilament A8 to C9 between each
triplet (Guichard et al., 2013; Winey and O’Toole, 2014). (C) Cross-sectional view of cartwheel
stacks, with physical measurements, as seen in cryotomography based 3D-models of
Trichonympha centrioles (Diagram based on Figures 2, 3 and 6 of Guichard et al., 2013). The
central cartwheel (blue), spoke junction (green), spoke tips (red), pin-heads and pin-feet (blue)
are shown with connection to a microtubule (Guichard et al., 2013; Hirono, 2014; Winey and
O’Toole, 2014).

38
Figure 1.2

39
Figure 1.2 The Centrosome Throughout the Cell Cycle.
The general centrosome duplication cycle is outlined through various stages of the cell cycle.
(From top) In late mitosis, the newly assembled procentrioles from the previous cell cycle lose
their tight connection to the parental centrioles in a process called disengagement, and gain the
ability to recruit PCM proteins. Both of these events are critical for the centriole to participate in
the subsequent round of centriole duplication. After each mitosis, daughter cells enter G1 with a
single centrosome containing two disengaged centrioles connected by a fibrous linker (see text).
In S-phase, a procentriole forms adjacent to each of these centrioles, and these procentrioles
continue to grow and elongate through S and G2. In late G2, the amount of PCM recruited by
the two centrosomes increases (centrosome maturation). Also in late G2, the two parental
centrioles lose their fibrous linker to separate from each other (centrosome separation) in
preparation for bipolar spindle formation in mitosis. During mitosis, the younger of the two
parental centrioles will also complete its maturation with the acquisition of distal and subdistal
appendages.

40
Figure 1.3

41
Figure 1.3. Contemporary Model of Centriole Assembly.
(A) (Top) The process of procentriole assembly on a single parental centriole is shown (see text
for references). (From top-left) In G1 PLK4 is first recruited as a ring to CEP192 on the parental
centriole, then migrates farther away from the centriole barrel by interacting with CEP152 which
is newly recruited in G1. At the start of S-phase, the formation of a SASS6 cartwheel occurs
within the lumen of each disengaged parental centriole. The recruitment of SASS6 into the
parental lumen is mediated through binding to CPAP therein. Subsequently, dependent upon
PLK4 and STIL, SASS6 is released from the parental centrioles and localizes to the proximal
outer wall of the centrioles along with a single focus of PLK4 and STIL. Thereafter, assembly of
a procentriole is initiated requiring the combined activities of -tubulin, /-tubulin, CPAP,
CEP135, Centrobin and CP110. In S-phase, the elongation of the procentrioles is driven by
CPAP, Centrobin and Poc1, whereas in G2 elongation is further promoted by CPAP, Centrobin,
hPOC5 and OFD14. The figure legend is shown below the pathway. (B) A summary of the
proteins known to promote or restrict centriole elongation is shown, with the affected portion of
the centriole indicated. *OFD1 restricts the length of the parental centriole specifically.
(Bottom) Legend is shown.

42
Figure 1.4

43
Figure 1.4. Summary of key Centriole Duplication Proteins and their Localizations.
(Top) A model of a centriole in S-phase, showing an early procentriole (right) adjacent to the
mature centriole (left). Approximate localizations of proteins involved in centriole duplication
are shown in purple, with their localization during duplication shown in green (the procentriole
assembly site). (Bottom) A brief summary of the interactions and functions of select proteins
with critical roles in centriole duplication.

44
2. Chapter II:
CEP120 and SPICE1 Cooperate with CPAP in
Centriole Elongation

45
2.1. Statement of Contributions
Almost all the data in this chapter has been published as a report in the journal Current Biology:
David Comartin*1,2, Gagan D. Gupta1, Eden Fussner3,4, Etienne Coyaud5,6, Monica Hasegan1,
Marco Archinti7, Sally W.T. Cheung1, Deborah Pinchev1, Steffen Lawo1,2, Brian Raught5,6,
David P. Bazett-Jones3,4, Jens Luders8, and Laurence Pelletier1,2. CEP120 and SPICE1 Cooperate
with CPAP in Centriole Elongation, Current Biology.2013 July 22. 23(14): 1360-1366.
DOI: 10.1016/j.cub.2013.06.002.
www.sciencedirect.com.myaccess.library.utoronto.ca/science/article/pii/S0960982213006933
This publication was a collaboration involving researchers from:
(1) The Samuel Lunenfeld Research Institute at Mt. Sinai Hospital, Toronto, Ontario, Canada.
(2) The Department of Molecular Genetics at The University of Toronto, Toronto Ontario
Canada.
(3) The Department of Biochemistry at the University of Toronto, Toronto, Ontario Canada.
(4) The Hospital For Sick Children, Toronto, Ontario, Canada.
(5) The Ontario Cancer Institute, Toronto, Ontario, Canada.
(6) The Department of Medical Biophysics at the University of Toronto, Toronto, Ontario
Canada.
(7) ERA Biotec, Barcelona, Spain.
(8) Institute for Research in Biomedicine (IRB Barcelona), Barcelona, Spain.
Dr. Gagan D. Gupta (Pelletier Laboratory) carried out image analysis and quantification of
various centrosome signals using scripts written in MATLAB. He also generated box and
whisker plots shown in this chapter (Figures 2.8 and 2.13).
Sally Cheung (Pelletier Laboratory) carried out multiple Western blots and co-
immunoprecipitation experiments (Figures 2.5d and 2.16).
Dr. Marco Archinti (Luders Laboratory) made and generously provided SPICE1 DNA in
plasmid form encoding full-length SPICE1 as well as fragments of SPICE1. He further
performed the co-IP experiments shown in Figure 2.3 in their entirety.
Dr. Eden Fussner (Bazett-Jones Laboratory) operated the electron microscope for the acquisition
of all tomography data sets discussed within this thesis. Further, she carried out tomogram
reconstruction and analysis of microtubule length contributing to Figure 2.11.
Doug Holmyard (SickKids Hospital and Mt .Sinai Hospital Toronto Joint Advanced Bioimaging
Centre) fixed and prepared all samples for electron microscopy, and provided training and
assistance on the electron microscope.
Deborah Pinchev (Pelletier Laboratory) performed the rescue experiment for CEP120 siRNA
shown in Figure 2.2D, E. She also generated the cell line used in that experiment.

46
Figure 2.1 of this chapter has been published in part in a paper in the journal Science:
James R. A. Hutchins1*, Yusuke Toyoda2*, Björn Hegemann1*, †Ina Poser2*, Jean-Karim
Hériché 3, 4, Martina M. Sykora1, Martina Augsburg2, Otto Hudecz1, Bettina A. Buschhorn1,
Jutta Bulkescher4, Christian Conrad4, David Comartin5,6, Alexander Schleiffer1, Mihail Sarov2,
Andrei Pozniakovsky2, Mikolaj Michal Slabicki2, Siegfried Schloissnig2,7, Ines Steinmacher1
Marit Leuschner2, Andrea Ssykor2, Steffen Lawo 5,6, Laurence Pelletier5,6, Holger Stark8, Kim
Nasmyth1, ‡Jan Ellenberg4, Richard Durbin3, Frank Buchholz2, Karl Mechtler1, Anthony A.
Hyman2 and Jan-Michael Peters1 Systematic Analysis of Human Protein Complexes Identifies
Chromosome Segregation Proteins. Science, 2010 April 30. 328, 593-599.
Reprinted with permission from AAAJ.
(1) Research Institute of Molecular Pathology (IMP), Dr. Bohr-Gasse 7, A-1030 Vienna, Austria.
(2) Max Planck Institute (MPI) for Molecular Cell Biology and Genetics, Pfotenhauerstrasse
108, D-01307 Dresden, Germany.
(3) Wellcome Trust Sanger Institute, Wellcome Trust Genome Campus, Hinxton, Cambridge
CB10 1HH, UK.
(4) Cell Biology and Biophysics Unit, European Molecular Biology Laboratory, Meyerhofstrasse
1, D-69117 Heidelberg, Germany.
(5) Samuel Lunenfeld Research Institute, Mount Sinai Hospital, 600 University Avenue,
Toronto, Ontario, Canada.
(6) Department of Molecular Genetics, University of Toronto, Toronto, Ontario M5S 1A8,
Canada.
(7) German Cancer Research Center, Im Neuenheimer Feld 280, 69120 Heidelberg,
Germany.
(8) Max Planck Institute for Biophysical Chemistry, Am Fassberg 11, D-37077 Göttingen,
Germany.

47
2.2. Summary
Centrosomes play an important role in maintaining genome stability, and along with the
genome must be duplicated during the cell cycle. Centrioles are the barrel-shaped
microtubule structures that organize the PCM at the centrosome. Procentriole assembly is
achieved by a group of proteins, and centriole lengths are remarkably consistent throughout
human cells. The length of procentrioles is thought to be regulated at least partly through the
balance of the pro-elongation factors such as CPAP and hPOC5 with negative regulators of
centriole length such as CP110 and CEP97 (Azimzadeh et al., 2009; Schmidt et al., 2009;
Spektor et al., 2007; Tang et al., 2009a). How procentriole assembly and growth to a specific
length are achieved is the subject of continuing investigation.
Here, we show that CEP120 and SPICE1 are required for centriole duplication, specifically
impacting the recruitment of CEP135, Centrin, CP110, CEP97 and each other during
assembly. CEP120 and SPICE1 recruitment is dependent upon SASS6, STIL and CPAP
during centriole overduplication. We show that CEP120 complexes with SPICE1 and CPAP
in vivo, and depletion of each of these proteins resulted in procentrioles either lacking
microtubules or with fewer, shorter microtubules. Overexpression of CEP120 or CPAP
drove abnormal centriole elongation in G2 specifically, and CPAP, CEP120 and SPICE1 had
to be expressed for this to occur in either case. Using colchicine to depolymerize
microtubules, I show that CEP135 recruitment during procentriole assembly requires
microtubules whereas SASS6, SPICE1, CEP120, CPAP, and CP110 all localized to the
region of procentriole assembly without microtubules present. Lastly, I demonstrated that
artificially stabilizing microtubules in G2 arrested cells led to elongated centrioles and
procentrioles, but this was not observed when CPAP, CEP120, or SPICE1 were depleted.
Based on these results, I conclude that CEP120 and SPICE1 play a role in procentriole
assembly and that together with CPAP they are critical for delivery or incorporation of
tubulin into centrioles. Overall our findings identify a shared function for CEP120, SPICE1
and CPAP in procentriole assembly and elongation.

48
2.3. Introduction 2.3.1. Combining Super-Resolution Imaging with PLK4 Induced Centriole
Overduplication to Study Procentriole Assembly
The centriole duplication pathway involves several conserved proteins, and the human pathway
was partly illuminated using a system where PLK4, the initiating kinase, was overexpressed
upon addition of tetracycline to cells (Kleylein-Sohn et al., 2007). This system effectively
amplifies centriole duplication from one procentriole per parent to multiple procentrioles per
parent. Much of the data presented in this chapter was obtained using the same PLK4 over-
expression system. There are several advantages to this approach: First, detection of failure of
one procentriole to form is more challenging than detection of 6-8 failing to form, making
defects more obvious experimentally (Kleylein-Sohn et al., 2007). Second, and perhaps more
importantly, the system amplifies the amount of protein localized during duplication, making it
much easier and more reliable to assess the dependencies of recruitment within the pathway
(Kleylein-Sohn et al., 2007). Third, when the parental centriole is oriented such that the observer
is looking down the length of the barrel, PLK4 appears ring-like surrounding the parental
centriole. This provides an important and reliable cue for determining where procentrioles
should form and the orientation of the centrioles and procentrioles being imaged.
In combination with the PLK4 induced centriole overduplication system, I show data within this
chapter that were obtained using a super-resolution microscopy method known as three
dimensional structured illumination microscopy (3D-SIM). The resolution of wide-field
fluorescence microscopy is fundamentally limited by the fact that light diffracts, and thus is often
referred to as ‘diffraction limited’ (Lakadamyali and Cosma, 2015; Oddone et al., 2014;
Yamanaka et al., 2014). The lateral resolution achievable in a quality wide-field microscope is
approximately half the wavelength of the fluorescent light being imaged, on the order of ~200
nm (Lakadamyali and Cosma, 2015; Oddone et al., 2014; Yamanaka et al., 2014). This means
that two coplanar fluorescent point-sources within 200 nm of each other will appear as one spot
in an image (Lakadamyali and Cosma, 2015; Oddone et al., 2014; Yamanaka et al., 2014). With
diffraction limited microscopy, resolving certain structural features of centrosomes, for example
the organization of the PCM proteins, is not possible (Lawo et al., 2012). Several super-
resolution microscopy methods allow resolution beyond the limits of conventional fluorescence

49
(Huang et al., 2009, 2010; Lakadamyali and Cosma, 2015; Oddone et al., 2014; Schermelleh et
al., 2010; Yamanaka et al., 2014). These methods include stimulated-emission-depletion-
microscopy (STED), stochastic optical reconstruction microscopy (STORM) photoactivated
localization microscopy (PALM), and 3D-SIM (Huang et al., 2009; Schermelleh et al., 2010;
Yamanaka et al., 2014). 3D-SIM involves taking multiple images using patterned illumination,
with the pattern shifted or rotated for each image. The combination of the sample’s fluorescence
with the known illumination pattern in multiple positions provides additional information that
can be used to computationally reconstruct an image of higher resolution (Gustafsson, 2000;
Gustafsson et al., 2008; Huang et al., 2009; Schermelleh et al., 2008). 3D-SIM has the advantage
of allowing rapid 3D imaging with multiple fluorophores and achieves double the lateral and
axial resolution of conventional microscopy (Gustafsson, 2000; Huang et al., 2009, 2010;
Schermelleh et al., 2008; Yamanaka et al., 2014). Several studies have used this approach to
visualize features of the centrosomes and centrioles otherwise unresolvable using diffraction
limited approaches (Kim et al., 2013; Lawo et al., 2012; Mennella et al., 2012; Park et al., 2014;
Sonnen et al., 2012). Combining the resolution of 3D-SIM with the benefits of the PLK4 assay
can provide important insights into the process of procentriole assembly.
2.3.2. The Use of Chemical Manipulation of Microtubules to Study Centriole
Elongation
Centrioles are nine-fold symmetrical microtubule based structures, and once assembled these
structures are resilient to chemical destabilization of microtubules, for example by the colchicine
derivative colcemid (Kuriyama, 1982). Normally centrioles reach a consistent length, and
procentrioles elongate through the cell cycle, but when the microtubule stabilizing chemical
taxol is added centrioles undergo abnormal elongation (Kuriyama et al., 1986). Conversely,
microtubule destabilizing chemicals can be used to prevent centriole duplication or limit
procentriole elongation reversibly, depending on concentration used (Kuriyama, 1982). The use
of chemicals to augment or inhibit centriole elongation is in some cases preferable to other
methods that involve protein depletion or overexpression, for example CPAP overexpression or
CP110 depletion (Schmidt et al., 2009). The use of chemicals in this context allows the
relationship between a protein of interest and the process of centriole microtubule nucleation or
elongation to be compared directly, without perturbing that protein’s cofactors. In other cases,

50
such as inferring cooperation or co-dependence for elongation or nucleation, altering the levels
of proteins involved in centriole length regulation would be important. Overall, these tools have
the potential to provide powerful insights into the mechanisms of centriole length regulation.
2.4. Results. 2.4.1. CEP120 and SPICE1 Interact and are Required for Centriole
Duplication in Cycling Cells.
As part of a large scale study of proteins important for mitosis in humans, CEP120 and CCDC52
(now SPICE1) were expressed at near endogenous levels from bacterial artificial chromosomes
(BACs) as fusions to localization-affinity purification tags (LAP-tags) and purified by modified
tandem-affinity-purification for the purpose of identifying interacting proteins (Cheeseman and
Desai, 2005; Hutchins et al., 2010; Poser et al., 2008). Endogenous CEP120 co-purified with
LAP tagged CCDC52, and endogenous CCDC52 co-purified with LAP tagged CEP120
(Hutchins et al., 2010). In addition, both proteins were found by fluorescence microscopy to
localize to the centrosome throughout the cell cycle in HeLa cells (Hutchins et al., 2010). To test
the possibility that CEP120 might play a role in centriole duplication, I used RNAi to deplete
CEP120 for 72 hours in cycling HeLa cells, and quantified the number of centrioles in mitotic
cells (Hutchins et al., 2010) (Figure 2.1A). I observed that CEP120 depletion led to an increase
in the number of cells with less than 4 centrioles in mitosis, indicating that CEP120 is likely
required for centriole duplication (Hutchins et al., 2010). To validate the specificity of the RNAi
silencing reagent, I demonstrated that the number of centrioles in mitosis was restored by
expression of an RNAi-resistant LAP-tagged mouse-CEP120 (Figure 2.1B). To further validate
the role of CEP120 in centriole duplication, I combined RNAi with an established technique for
inducing centriole overduplication by arresting U-2 OS cells in S-phase with hydroxyurea (HU)
(Balczon et al., 1995; Zhu et al., 2008). When CEP120 was depleted in this assay, the frequency
of centriole overduplication was significantly decreased (Hutchins et al., 2010) (Figure 2.1C).
These results indicate that CEP120 is required for centriole duplication in cycling cells, and
centriole overduplication in S-phase arrested cells.

51
Subsequent to our study (Hutchins et al., 2010) it was published that SPICE1 was required for
centriole duplication in cycling HeLa cells and for centriole overduplication in S-phase arrested
U-2 OS cells (Archinti et al., 2010) and elsewhere that CEP120 was required for centriole
duplication (Mahjoub et al., 2010). We established a collaboration with the lab of Dr. Jens
Luders, and I verified that SPICE1 depletion in cycling HeLa cells using their published siRNA,
and a second siRNA, inhibited centriole duplication (Figure 2.2A, B). The same experiment was
performed in HeLa cells to test two different siRNAs against CEP120, and treatment with either
siRNA had a similar effect (Figure 2.2B). To validate the specificity of our unpublished CEP120
siRNA (siRNA-1), Deborah Pinchev generated a HeLa T-REx inducible HeLa cell line that
expressed a FLAG-tagged CEP120 construct resistant to CEP120 siRNA-1. She then
demonstrated that when this siRNA-resistant FLAG-CEP120 was expressed, treatment of cells
with siRNA-1 against CEP120 did not lead to a defect in centriole duplication (Figure 2.2D, E).
Therefore the CEP120 siRNA-1 affects centriole duplication specifically by reducing CEP120
levels. Accordingly, unless otherwise specified the rescued siRNA targeting CEP120 (si-1) and
the published siRNA targeting SPICE1 were used for the remaining work described in this
thesis.
Dr. Marco Archinti from the Luders lab validated the interaction of CEP120 and SPICE1 by
reciprocal co-IP experiments, and mapped the domains of CEP120 and SPICE1 involved in this
interaction (Figure 2.3). A CEP120 fragment was expressed that corresponded to the amino-
terminal portion of the protein (residues 1-700) and a second fragment was expressed that
contained only the carboxy-terminal region including the coiled-coil domain (residues 700 to
986). Unlike the full length CEP120, neither fragment of CEP120 was able to co-IP SPICE1
(Figure 2.3A). SPICE1 fragments containing one or both of its coiled-coil domains were able to
co-IP CEP120, though the fragment containing both coiled-coil domains interacted more
strongly (Figure 2.3B). In summary, CEP120 and SPICE1 interact and are required for centriole
duplication in multiple human cancer cell lines.
2.4.2. CEP120 and SPICE1 are Required for PLK4 Induced Centriole
Overduplication
The creators of the PLK4 induced centriole overduplication assay generously provided us with
their cell line, which when treated with tetracycline will over-express PLK4 fused to a Myc-

52
epitope tag (Kleylein-Sohn et al., 2007). Initially, I sought to confirm that CEP120 and SPICE1
were important for PLK4 induced centriole overduplication, and to place CEP120 and SPICE1
within the centriole duplication pathway beginning with an examination of SASS6 localization to
procentrioles (Figure 2.4). SASS6 is a very early protein in the centriole duplication pathway,
and it forms the cartwheels of centrioles (Pelletier et al. 2006; Kitagawa et al. 2011; Leidel et al.
2005; Kleylein-Sohn et al. 2007). Two phenotypes were observed in each condition: One
population of parental centrioles showed no SASS6 recruitment at all, and a second population
of parental centrioles had SASS6 surrounding them but did not form procentrioles (Figure 2.4A,
B). The two phenotypes (SASS6 recruited, SASS6 not recruited) were also observed in positive
controls where CPAP and CEP135 were depleted in the assay (Figure 2.4A, B). The fact that
centriole duplication was inhibited in cells regardless of SASS6 recruitment to mother centrioles,
and that Western-blots showed CEP120, SPICE1, CPAP and CEP135 depletion, indicated that
the variable phenotypes were not a result of differences in RNAi efficiency (Figure 2.4C). In
trying to understand the variable SASS6 localization, I considered the possibility that differences
in the structure of the parental centrioles might have played a role. The U-2 OS cells were
asynchronous at the time of RNAi, and cycled for 48 hours prior to S-phase arrest. During the
~2 cell cycles of depletion prior to S-phase arrest, a fraction of the parental centrioles that would
later template overduplication were being assembled. If the centrioles assembled during the 48
hours were structurally defective, they might fail in recruiting SASS6, while (older) structurally
sound parental centrioles would retain the ability to recruit SASS6. This could explain the
variable phenotypes of SASS6 localization. To test this possibility I modified the PLK4 induced
centriole overduplication assay to reduce the number of cell cycles following RNAi treatment.
In the modified assay, cells were depleted by RNAi for 24 hours, then arrested in S-phase and
treated with tetracycline for another 24 hours before being released from S-phase by washout and
finally arrested in G2 for 16 hours (hereafter ‘the G2 PLK4 assay’, see Materials and Methods).
This G2 assay was intended to ensure a nearly uniform population of parental centrioles, but also
to allow sufficient time for depletion of target proteins before centriole overduplication.
Western-blots performed by Sally Cheung on whole-cell lysates I collected from the same
experiment showed that each protein was strongly depleted despite the reduced number of cell
cycles in the G2 PLK4 assay (Figure 2.5D). When CEP120, SPICE1, CPAP or CEP135 were
depleted in the G2 PLK4 assay, centriole duplication was still significantly inhibited, based on

53
Centrin recruitment (Figure 2.5A, B). Consistent with my hypothesis, when any of the target
proteins were depleted in the G2 PLK4 assay, SASS6 was localized to the region surrounding the
mother centrioles in the majority of cells where centriole overduplication failed (Figure 2.5A, B).
In summary, my results lead me to the conclusion that CEP120 and SPICE1 act downstream of
SASS6 recruitment during procentriole assembly, as do CPAP and CEP135 as previously
reported (Kleylein-Sohn et al., 2007).
2.4.3. Using 3D SIM to Study Centriole Assembly.
In the G2 PLK4 assay, I observed a subtle difference in SASS6 localization in control cells
compared to cells depleted of centriole assembly proteins. Namely, in controls SASS6
localization was often to clearly resolvable procentrioles, whereas in knock-downs of CEP120,
SPICE1, CPAP or CEP135 SASS6 localization appeared ring-like around the mother centriole
(Figure 2.5A). The frequency of the SASS6 localizations where individual procentrioles could
not be identified (i.e. rings of SASS6) was quantified following CEP120, SPICE1 and CPAP
depletion in the G2 PLK4 assay (Figure 2.5C). I found that depletion of these proteins led to an
increase in the frequency of smooth SASS6 rings surrounding mother centrioles (Figure 2.5C).
Thus, while dispensable for SASS6 recruitment to the mother centrioles, CEP120, SPICE1 and
CPAP depletion each affect the structure of SASS6 surrounding mother centrioles in the G2
PLK4 assay. To gain a better understanding of the ‘rings’ of SASS6, I used 3D SIM to examine
centrosomes following Myc-PLK4 overexpression in asynchronous U-2 OS cells. I imaged
centrosomes labeled with antibodies against SASS6 and the distal centriole markers Centrin and
CP110. Importantly, using PLK4-induced centriole overduplication in combination with 3D-
SIM, it is easy to determine the mother centriole based on the characteristic ‘flower’ shape of the
mother centriole surrounded by procentrioles, when the mother is oriented with its centriole
barrel along the z-axis (Figure 2.6A). Using centrosomes with this orientation, I was able to
compare the lengths of the procentrioles, as evident from both the elongation of SASS6 in some
procentrioles and the distances between the Centrin of the mother centriole and the Centrin in the
distal portion of procentrioles (Figure 2.6B). Centrosomes with procentrioles of varying lengths
were detectable, and representative images are shown in Figure 2.6B. Even using super-
resolution imaging, I found centrosomes where a ring of SASS6 was present, and these
centrosomes had the shortest procentrioles based on CP110 and Centrin localization (Figure

54
2.6B). From this observation, I conclude that SASS6 first localizes in a uniform distribution
around the mother centriole, and then subsequently becomes organized or resolvable into
individual procentrioles (Figure 2.6B). Consistent with this idea, a fraction of control
centrosomes were observed to have SASS6 rings in the G2 PLK4 assay (Figure 2.5C). I next
used 3D-SIM along with PLK4 induced centriole overduplication to examine the localization of
CEP120 and SPICE1 to mother centrioles and procentrioles. Both CEP120 and SPICE1
appeared to label the walls of the procentrioles (Figure 2.6C). This is consistent with the
immuno-gold labelling of CEP120 in EM experiments reported elsewhere during the course of
my research (Mahjoub et al., 2010). I observed a gap between the CEP120/SPICE1 labeling on
the mother centriole and the procentrioles (Figure 2.6C). The gap corresponded to the
localization of Myc-PLK4 around the mother, indicating that 3D-SIM makes it possible to
differentiate between localization of a protein to the mother centriole and localization to a
procentriole when Myc-PLK4 is labeled (Figure 2.6C). In summary, 3D-SIM in combination
with PLK4 induced centriole overduplication provides important insights not possible using
deconvolution fluorescence microscopy. Using this approach I found that SASS6 first localized
as a ring around the mother centriole before becoming resolvable within individual procentrioles
during PLK4 induced centriole overduplication, and that CEP120 and SPICE1 were localized to
the barrels of the procentrioles.
2.4.4. Defining the Roles of CEP120 and SPICE1 in Procentriole Assembly
Having established that CEP120 and SPICE1 are dispensable for SASS6 localization to
procentrioles, I next sought to place CEP120 and SPICE1 within the pathway of centriole
assembly. I repeated the G2 PLK4 assay depleting either CEP120 or SPICE1, and labeled
centrosomes for Myc-PLK4, Centrin, and one of a set of proteins known to be required for
centriole duplication. Using 3D-SIM, I observed that CEP120 or SPICE1 depletion resulted in
SASS6, STIL and CPAP localizing as rings partially overlapping the Myc-PLK4 signal (Figure
2.7). These rings were similar to SASS6 structures seen in the early stages of PLK4 induced
centriole overduplication (compare Figure 2.6B versus Figure 2.7). CEP152 localized as a ring
partly overlapping Myc-PLK4 in control conditions in this assay, but was not observed in the
region outside the Myc-PLK4 ring, consistent with its early role in PLK4 localization (Figure
2.7) (Cizmecioglu et al., 2010; Hatch et al., 2010; Sonnen et al., 2013). CEP120 and SPICE1

55
both appeared to be dispensable for the localization of CEP152 at the parental centriole in this
assay (Figure 2.7). Conversely, CEP120 and SPICE1 depletion resulted in centrosomes lacking
the distal centriole microtubule capping proteins CP110 and CEP97 (Figure 2.7). Lastly, the
localization of hPOC5 was not strongly affected by CEP120 or SPICE1 depletion in the G2
PLK4 assay (Figure 2.7).
When CEP120 or SPICE1 were depleted, I observed CEP135 localization was limited to the
parental centriole (Figure 2.8A). I repeated the G2 PLK4 assay with CEP135, CPAP, CEP120 or
SPICE1 depletion and immunolabeled the cells for Myc-PLK4, CEP135 and Centrin. For each
RNAi condition, I collected images of centrosomes using 3D-SIM under identical imaging and
image processing conditions. From these, I manually selected images where the parental
centriole was oriented along the z-axis such that Myc-PLK4 appeared as a distinct ring. I worked
with Dr. Gagan Gupta to develop an automated image analysis approach to quantify the
fluorescence intensity of CEP135 signal specifically in the region surrounding the parental
centriole outside of or overlapping the Myc-PLK4 signal. We termed this region the PCAR
(ProCentriole Assembly Region) (Figure 2.8B). The Myc-PLK4 signal was used to create a
‘mask’ in which the CEP135 signal that corresponded to the procentrioles was quantified. The
generation of signal masks, quantifications of signals, and generation of dot-plots were
performed by Dr. Gupta using MatLab scripts he authored for this assay. We observed that using
this experimental approach, there was a significant decrease in CEP135 signal intensity in the
PCAR under each of the depletion conditions compared to the control (Figure 2.8B). To validate
the importance of CEP120 and SPICE1 in CEP135 recruitment to procentrioles, I depleted
cycling HeLa cells stably expressing GFP-SASS6 of CEP120 or SPICE1 for 72 hours, then
imaged the S-phase centrosomes using 3D-SIM. Consistent with the results from the G2 PLK4
assay, when CEP120 or SPICE1 were depleted, I observed centrosomes where GFP-SASS6 was
evident adjacent to mother centrioles, but CEP135 was faint or not detectable (Figure 2.8C). To
quantify CEP135 levels at procentrioles in S-phase cells, I synchronized HeLa cells first in G2,
then released them and re-arrested them in S-phase following depletion of CEP120, SPICE1 or
CEP135. Centrosomes were then quantitatively imaged using 3-D SIM, and the frequency of
SASS6 positive procentrioles where CEP135 was not detectable was quantified for each
condition (Figure 2.8D, E). Further, the mean pixel intensity of CEP135 signal overlapping the
procentriole SASS6 signal was quantified for a number of centrosomes from each condition

56
(Figure 2.8D, F). I observed a frequent absence of CEP135 from the SASS6 positive
procentrioles, and a reduced overall average intensity of CEP135 signal concurrent with SASS6
signal, when CEP135, CEP120 or SPICE1 were depleted. Therefore CEP120, SPICE1, and
CPAP all appear to be important for CEP135 recruitment during both procentriole assembly.
Having determined that CEP120 and SPICE1 were important for CEP135 recruitment, I next
sought to establish which centriole assembly proteins were required for CEP120 and SPICE1
localization during centriole overduplication. To do this, I again employed RNAi in combination
with the G2 PLK4 induced centriole overduplication assay and 3D-SIM. I observed that when
CEP152, SASS6, STIL, CPAP or CEP135 were depleted, CEP120 and SPICE1 levels were
reduced in the regions surrounding parental centrioles in the majority of centrosomes (Figure
2.9A). In some cases, CEP120 or SPICE1 were retained on the mother centriole, most strongly
when CEP152/SASS6/STIL were depleted (Figure 2.9A). CEP135 and CPAP depletion reduced
CEP120 and SPICE1 recruitment to procentrioles and reduced their localization at the parental
centrioles (Figure 2.9B). When CEP120 or SPICE1 were depleted, both proteins were strongly
reduced in their localization to the procentriole assembly region (Figure 2.9B). Consistent with
published reports, CEP97 or CP110 depletion in the assay led to procentrioles that were
abnormally elongated (Schmidt et al., 2009; Spektor et al., 2007) (Figure 2.9C). Under CEP97
or CP110 depletion, CEP120 and SPICE1 were localized along the lengths of the abnormally
elongated procentrioles, indicating their localization was independent of CP110 and CEP97
(Figure 2.9C). When hPOC5 was depleted, although Centrin recruitment was affected, CEP120
localized along what appear to be procentrioles, and SPICE1 was also seen in the region of
procentriole assembly (Figure 2.9D). POC1 depletion did not affect CEP120 localization, but
did strongly impact SPICE1 recruitment (Figure 2.9D). Overall, from these results I conclude
that CEP120 and SPICE1 are dependent upon the functions of CEP152, SASS6, STIL, CPAP
and CEP135 for their recruitment to procentrioles, and that CEP120 and SPICE1 are important
for the recruitment of CEP135 and the distal procentriole assembly proteins CP110 and CEP97.
2.4.5. Investigating the Role of CEP120 and SPICE1 in Procentriole
Structure
The depletion of CEP120 or SPICE1 did not prevent accumulation of early procentriole
assembly proteins (SASS6, STIL, CPAP), however, even from super-resolution images it was

57
unclear whether there were de facto procentrioles or precursors being assembled under these
conditions. To understand more precisely the structures that exist in the PCAR when CEP120
and SPICE1 are absent, I pursued electron microscopy. Electron microscopy (EM) is the best
method for observing the structure of centrioles and procentrioles in fixed samples. I provided
samples from a G2 PLK4 induced centriole overduplication assay where CEP120, SPICE1, or
CPAP had been depleted to Doug Holmyard at the Mt. Sinai EM facility who prepared them for
transmission electron microscopy (TEM) (see Materials and Methods). CPAP was included as a
positive control because CPAP is known to be required for centriole elongation, and so depletion
in the G2 PLK4 assay would be expected to yield shorter procentrioles (Kohlmaier et al., 2009;
Schmidt et al., 2009; Tang et al., 2009a). Thin sectioning of EM samples (100 nm) results in
centrosomes that are often split into multiple sections, so to ensure proper detection of the full
length of each mother centriole and possible procentrioles, Doug Holmyard prepared serial
sections from each sample. In serial sections, the same sample block is cut and sequential slices
are mounted for viewing in order, allowing for the identification and examination of the same
centrosome in its entirety through a series of slices. Using this approach, I found multiple
centrosomes from each condition that could be examined in their entirety through multiple
sections. Such centrosomes are evident when the cell containing a centrosome of interest can be
identified in a serial section where the centrosome is no longer detectable. Figure 2.10 shows
representative centrosomes imaged through serial sections from each condition. In control
centrosomes, the procentrioles had well defined and resolvable microtubules (Figure 2.10). In
samples where CEP120, SPICE1 and CPAP had been depleted, regions of electron density were
visible adjacent to the parental centrioles (Figure 2.10). In some cases, these regions of electron
density included short but detectable microtubules, though overall no robust procentriole
structure was evident in most cases (Figure 2.10). These observations indicate that CEP120 and
SPICE1, like CPAP, act at an early stage in procentriole assembly, and that despite the presence
of SASS6 and STIL in the PCAR following CEP120 or SPICE1 depletion procentrioles are not
necessarily assembled. Regarding the organization of SASS6 in this region, I was unable to
detect cartwheel structures even in control procentrioles, and thus I cannot determine from these
results whether SASS6 forms cartwheels in the absence of CEP120 or SPICE1.
In the absence of CEP120, CPAP or SPICE1, procentriole precursors sometimes contained
detectable microtubules. I sought to compare the lengths of such microtubules to microtubules

58
in control procentrioles. To better resolve and effectively measure microtubules in three
dimensions, I collaborated with Dr. Eden Fussner from the lab of Dr. David Bazett-Jones to
perform electron tomography on samples from the G2 PLK4 assay. Briefly, electron tomography
involves imaging a thicker sample section at multiple angles by transmission electron
microscopy as it is tilted a number of times within the microscope. The samples are prepared
with gold particles present, and following acquisition of the ‘tilt-series’ of images, software is
used to reconstruct a three-dimensional image of the sample based on the experimental setup and
the positional information provided by the gold-particles. Dr. Fussner performed the image
acquisition for all tomograms, and carried out the processing to produce the final tomograms and
projections. I worked alongside Dr. Fussner selecting the appropriate centrosomes for each
condition. After useable tomograms were obtained for each condition, Dr. Fussner and I
measured detectable microtubules in control procentrioles and the procentriole precursors
observed under experimental conditions (see Materials and Methods). From 3D tomograms we
selected microtubules that we were able to measure along their entire lengths (Figure 2.11A).
We found that in control conditions, procentriole microtubules reached an average length of
>150 nm, whereas when CEP120, SPICE1 or CPAP were absent, the microtubules averaged less
than 100 nm in length (Figure 2.11B). In addition to the length of microtubules, we measured
the width of the procentrioles and precursor procentrioles from the outside of the detectable
microtubules, and found that this metric was similar across all samples (Figure 2.11C). Overall
the results of this experiment validated the observations from transmission electron microscopy
experiments, namely that CEP120, CPAP or SPICE1 depletion resulted in partially assembled
procentrioles that either lacked microtubules, or had short microtubules compared to control
procentrioles. Therefore, CEP120 and SPICE1 act like CPAP at an early stage of procentriole
assembly, and are required for proper formation of procentriole microtubules.
2.4.6. Comparison of the Effects of CEP120 or SPICE1 Depletion versus
Microtubule Depolymerization on PLK4 Induced Centriole Overduplication
Procentriole precursors assembled in the absence of CEP120 or SPICE1 have fewer
microtubules, and when present microtubules are shorter than in control conditions. One
possible explanation for this observation is that centriolar microtubules might not be properly
stabilized when CEP120 or SPICE1 are absent. To test the hypothesis that CEP120 or SPICE1

59
depletion causes defects in centriole duplication by altering microtubule stability, I investigated
the impact of chemical inhibition of microtubule polymerization on procentriole assembly. First,
I repeated the G2 PLK4 assay in the presence or absence of 10 M colchicine, a chemical known
to inhibit microtubule polymerization and thus centriole assembly (Boisvieux-Ulrich et al., 1989;
Kuriyama, 1982). Initially, I verified that treatment with colchicine inhibited centriole
overduplication following PLK4 overexpression. I found that 10 M colchicine effectively
depolymerized cytoplasmic microtubules and prevented the formation of procentriole
microtubules (Figure 2.12A). I noted that CEP135 localization to the PCAR was reduced or
eliminated by the addition of colchicine during PLK4 induced centriole overduplication (Figure
2.12B). I imaged centrosomes using 3D-SIM in this assay, and provided images to Dr. Gupta for
automated quantification of protein signals at the centrosomes using MATLAB. Consistent with
my qualitative observations, there was a significant decrease in total CEP135 signal and total
glutamylated-tubulin signal at the centrosomes when colchicine was present (Figure 2.13A, B).
Conversely, the total levels of SASS6 at the centrosomes were not significantly affected by the
presence of colchicine, suggesting that SASS6 localization was independent of procentriole
microtubule polymerization (Figure 2.13B). Colchicine treatment caused SASS6 to localize in a
ring around the mother centriole, which when imaged using 3D-SIM appeared similar to the
localization of SASS6 following CEP120 or SPICE1 depletion (Figure 2.13A versus Figure 2.7).
I next examined how prevention of procentriole microtubule formation affected the localization
of other centriole assembly proteins following PLK4 overexpression. When colchicine was
present, I observed that CEP120 and SPICE1 could localize to the region of procentriole
assembly, indicating microtubule independent localization (Figure 2.13C). Similarly, CPAP and
CP110 showed robust localization as rings to the PCAR (Figure 2.13C). This was unexpected,
since CEP120 or SPICE1 depletion prevent CP110 localization to the PCAR in the G2 PLK4
assay, and since CP110 localizes to the distal tips of centriole microtubules (Schmidt et al.,
2009). In summary, the initial localization of CP110 is a microtubule and CEP135 independent
process, but requires CEP120 and SPICE1 (Figure 2.13C and Figure 2.7). On the other hand,
recruitment of CEP135 to the PCAR is reduced when microtubules are absent, or when CEP120
or SPICE1 are depleted, in the G2 PLK4 assay.

60
2.4.7. CEP120 and SPICE1 Cooperate with CPAP in Centriole Elongation
The effects of CEP120 and SPICE1 depletion on procentriole assembly in the assays thus far
described are similar to those of CPAP depletion. CPAP is an early procentriole assembly
protein with a known role in procentriole elongation (Kleylein-Sohn et al., 2007; Tang et al.,
2009a). One of the phenotypes that was used to determine the function of CPAP was that when
overexpressed in G2 phase of the cell cycle, CPAP drove abnormal elongation of centrioles and
procentrioles (Tang et al., 2009a). I therefore sought to test whether CEP120 or SPICE1
overexpression gave a similar phenotype. When U-2 OS cells were transiently transfected with
plasmids encoding GFP-fusions to CEP120 or CPAP for 24 hours, a small but reproducible
fraction of cells had abnormally elongated centrioles based on acetylated tubulin and GFP
labeling (Figure 2.14A, B). Overexpression of a GFP-SPICE1 fusion in the same assay did not
result in an elongation of the centrioles, though SPICE1-GFP localized to acetylated cytoplasmic
microtubules (Figure 2.14A). CPAP induced centriole elongation is G2 specific (Kohlmaier et
al., 2009; Tang et al., 2009a). To see if CEP120 induced centriole elongation was cell cycle
specific, I generated a U-2 OS FLP-In TREx cell line that when treated with tetracycline
expressed FLAG-tagged CEP120. Similar to CPAP overexpression, CEP120 overexpression
drove the elongation of centrioles and procentrioles in G2 arrested cells, but not S-phase arrested
cells (Figure 2.14C, D). I next tested the possibility that CEP120 and CPAP were functionally
cooperating by probing whether each was required for centriole elongation following
overexpression of the other. I also tested whether SPICE1 was important for either CEP120 or
CPAP induced centriole elongation. When CPAP or SPICE1 were depleted prior to CEP120-
GFP overexpression, significantly fewer cells had elongated centrioles than in control transfected
cells or cells where SASS6 was pre-depleted as a control procentriole assembly protein not
involved in length regulation (Figure 2.15A, B). Similarly, centriole elongation following CPAP
overexpression required the presence of CEP120 and SPICE1, but was independent of SASS6
(Figure 2.15D, E). Therefore, CEP120 and SPICE1 appear to functionally interact with CPAP,
and to participate in centriole elongation and procentriole assembly along with CPAP.
To test whether CPAP was part of a complex with CEP120 and SPICE1, Sally Cheung
performed co-IP experiments to test whether CEP120 and CPAP interacted in vivo. HeLa cells
were transiently transfected with FLAG-CEP120 or FLAG-USP37 as a control and one of

61
SPICE1-GFP, CPAP-GFP or GFP-USP37 (Figure 2.16). When FLAG-CEP120 was pulled
down, SPICE1-GFP and CPAP-GFP were both detected by Western blot, but neither GFP
construct was detected when FLAG-USP37 was pulled down. Therefore CEP120 functionally
and physically interacts with CPAP and SPICE1 in vivo to drive centriole elongation.
Finally, having shown that colchicine inhibition of microtubule polymerization mimicked some
of the characteristic phenotypes of CEP120 or SPICE1 depletion, I sought to determine whether
CEP120 and SPICE1 were required to elongate centrioles in a context where no protein was
being overexpressed. To achieve this, I used taxol as a chemical agent known to stabilize
microtubules and facilitate microtubule polymerization (Arnal and Wade, 1995; Kumar, 1981;
Schiff et al., 1979). It has been shown previously that taxol treatment leads to elongated
centrioles (Kuriyama et al., 1986). When U-2 OS cells were arrested in G2 for 48 hours in the
presence of taxol, I observed that mother centrioles (as detected by CEP164 staining), daughter
centrioles and procentrioles frequently became elongated (Control, Figure 2.17). Conversely,
depletion of CEP120, CPAP or SPICE1 led to a decrease in the frequency of observed abnormal
centriole elongation affecting both centrioles and procentrioles (Figure 2.17B). In contrast,
SASS6 depletion effectively reduced procentriole formation but had no detectable impact on
centriole elongation (Figure 2.17A, B). Therefore, CEP120, CPAP and SPICE1 are all essential
to the elongation of both mature centrioles and procentrioles, even when elongation is driven by
chemical stabilization of centriolar microtubules themselves.

62
2.5. Discussion
2.5.1. Placing CEP120 and SPICE1 in the Centriole Assembly Pathway
The results described here demonstrate that CEP120 and SPICE1 interact and are required for
centriole duplication in multiple assays and cell types (Comartin et al., 2013; Hutchins et al.,
2010). Independently, our collaborators have shown that SPICE1 is important for centriole
duplication, and during the preparation of this work another group validated our finding that
CEP120 is important for centriole duplication (Archinti et al., 2010; Mahjoub et al., 2010). We
show here that CEP120 and SPICE1 interact, and that this interaction is dependent on the C-
terminal most coiled-coil domain of SPICE1, but stronger when both coiled-coil domains are
present. The fragment of SPICE1 that contains both coiled-coil domains (444-856) localizes to
the centrosome but not the mitotic spindle (Archinti et al., 2010). Taken together with our
findings, this suggests that SPICE1 spindle localization is not mediated by CEP120 interactions
alone. Full-length CEP120 was required to co-IP SPICE1, whereas neither the CTD coiled coil
domain nor NTD domain of CEP120 were sufficient for the interaction. This suggests that either
the full CEP120 protein is required for the interaction, or more likely that the SPICE1 binding
region bridges both of the fragments used here, as is the case for the CPAP binding domain of
CEP120 (416-730) (Lin et al., 2013b). Future work could include co-IPs with different
fragments of CEP120 to more precisely refine the SPICE1 binding domain, for example those
used to define the microtubule binding domain of CEP120 (Lin et al., 2013b). Consistent with
an interaction, depletion of either CEP120 or SPICE1 strongly reduces the recruitment of the
other protein in centriole duplication assays. Thus CEP120 and SPICE1 interact and cooperate
in human centriole duplication.
How CEP120 and SPICE1 act in centriole duplication was not understood at the outset of this
work. Given the previous utility of PLK4 induced centriole overduplication in describing
aspects of the human centriole assembly pathway, I employed PLK4 overexpression to decipher
where CEP120 and SPICE1 acted in the pathway. CEP120 and SPICE1 were recruited
dependent upon CEP152, SASS6, STIL, CEP135 and CPAP. I further observed that CEP120
and SPICE1 were dispensable for SASS6, STIL and CPAP recruitment, consistent with other
reports about CEP120 function in this process (Lin et al., 2013b). I showed that CEP120 and

63
SPICE1 were required for CEP97, CP110 and CEP135 recruitment to the site of procentriole
assembly. CPAP and CEP135 are required for CP110 localization during duplication, so it is
possible that CEP120, SPICE1 and CPAP are impacting CP110 localization indirectly through
localizing CEP135 (Kleylein-Sohn et al., 2007). This is unlikely, however, since in the presence
of colchicine CP110 localizes to the PCAR without CEP135. Alternatively, CEP120/SPICE1
could be directly involved in recruiting CP110 to procentrioles during assembly. Future work
should include investigating possible interactions between CEP120/SPICE1 and CP110/CEP97
by co-IP.
Both CEP120 and SPICE1 localize to procentrioles that are assembled in the absence of hPOC5,
consistent with reports that hPOC5 acts later in procentriole assembly but is important for
Centrin recruitment (Azimzadeh et al., 2009). POC1 depletion strongly affects SPICE1 but not
CEP120 localization to the procentriole regions. This result warrants further investigation, given
that other proteins studied here have similar effects on CEP120 and SPICE1 localization in our
assays. POC1 is important for S-phase elongation of procentrioles, so the requirement for POC1
in SPICE1 localization could explain why procentrioles fail to elongate or are unstable when
POC1 is depleted (Keller et al., 2009; Venoux et al., 2013). We were unable to obtain antibodies
for POC1 that were functional in immunofluorescence, but in future it will be worth
investigating whether CEP120 and SPICE1 are required for POC1A or POC1B localization to
procentrioles, as this may be relevant to the functions of CEP120 and SPICE1 in procentriole
microtubule assembly. In summary, CEP120 and SPICE1 are downstream of SASS6/STIL in
centriole duplication, but are required for CEP135 and CP110 recruitment, similar to CPAP.
2.5.2. CEP120 and SPICE1 Cooperate with CPAP in Centriole Elongation
Overexpression of CEP120, but not SPICE1, gave abnormal centriole elongation in G2 phase of
the cell cycle specifically, a phenotype reminiscent of CPAP induced elongation (Kohlmaier et
al., 2009; Tang et al., 2009a). Depletion of any of these three proteins was sufficient to perturb
this process, whereas SASS6 depletion blocked procentriole elongation presumably by blocking
procentriole formation, but not mature centriole elongation. The fact that we observed both
parental and procentrioles elongating upon CEP120 overexpression, similar to CPAP
overexpression, suggests that the levels of these proteins must be controlled to maintain
appropriate centriole length. Indeed, CPAP is a cell-cycle regulated protein, and an independent

64
study has shown that CEP120 levels are regulated through the cell cycle similar to CPAP levels
(Lin et al., 2013b; Tang et al., 2009a). However, another study of CEP120 and CPAP in
centriole elongation showed that when one was over-expressed, both were localized to the
elongated centrioles (Lin et al., 2013b). This may indicate that both proteins are present at levels
sufficient to drive elongation, but not sufficient to initiate it. Alternatively, the presence of excess
of either protein may stabilize the other, effectively increasing levels of both when either is over-
expressed. It has been shown that Centrobin can stabilize ubiquitinylated CPAP at the centriole
by inhibiting its destruction, so it is possible CEP120 could likewise protect CPAP and vice
versa (Gudi et al., 2015). Further, CEP120 must be able to affect both proximal and distal
portions of the centriole microtubules, since CEP120/CPAP driven elongation occurs distal to
the appendage component CEP164 (see Figure 2.15) and procentrioles are shorter when
CEP120/CPAP/SPICE1 are depleted.
Given the functional interaction, we considered a possible interaction between CEP120 and
CPAP. We found that CEP120 could co-IP CPAP, suggesting they interact in vivo. The same
independent study defined the CPAP binding, microtubule binding, and dimerization domains of
CEP120 (Lin et al., 2013b). Disruption of the microtubule binding domain, but not the
dimerization domain, strongly inhibits CEP120 induced elongation following overexpression
(Lin et al., 2013b). A similar microtubule binding-dependence of CPAP in centriole elongation
was previously reported, suggesting that these domains are both involved in centriole elongation
(Lin et al., 2013b; Tang et al., 2009a). In general, the other published study of CEP120 and
CPAP cooperation is largely in agreement with our study, with the exception that they conclude
that SASS6/STIL are important for centriole elongation following CEP120 overexpression (Lin
et al., 2013b). There are several differences in the experimental protocols used, including that in
our study, we synchronized cells in G2 for a prolonged period, whereas in the other report cells
were asynchronous (Lin et al., 2013b). Since CEP120 and CPAP are primarily active in G2
centriole elongation, our prolonged cell cycle arrest could have sensitized our system to the
effects of overexpression, possibly explaining why we observed mature centrioles elongating
more frequently. It is worth noting that the authors of the other study report 19% of the cells had
elongated mother centrioles despite SASS6 depletion (Lin et al., 2013b). Based on the fact that
SASS6 should not be incorporated in the distal portions of mother centrioles, but CEP120
overexpression drives elongation of that region, I favor a model where SASS6 does not play a

65
direct role in CEP120/SPICE1/CPAP mediated elongation of centrioles. In support of this,
taxol-induce centriole elongation was not inhibited by depletion of SASS6 in our study.
Procentrioles are not assembled when SASS6 is depleted, and SASS6 is required for CPAP,
CEP120 and SPICE1 to localize to these structures. Thus a decrease in overall percentage of
elongated centrioles is expected due to the impact on procentriole elongation alone, but is
probably an indirect effect upstream of CEP120/SPICE1 activity.
2.5.3. CEP120 and SPICE1 are Important for Microtubule Formation during
Procentriole Assembly
We used TEM with serial-sectioning and tomography in combination with the G2 PLK4 assay to
observe the state of microtubules in the procentriole assembly region following CEP120,
SPICE1 or CPAP depletion. Initially, with serial EM we found that in each case, parental
centrioles were surrounded by electron dense regions that either lacked detectable microtubules
or contained very short microtubules. We used tomography to measure the lengths of the
observed procentriole microtubules in three-dimensions, and found that indeed they were
significantly shorter when CEP120, SPICE1 or CPAP were depleted. These results, taken along
with the ring-like localization of centriole assembly proteins, support a model where CEP120
and SPICE1 are required for the assembly of procentriole microtubules or their stability, or both.
In either case, we would expect to see the appearance of some microtubules that are only capable
of reaching short lengths due to the dynamic nature of microtubule assembly. It will be of
interest to test the impact of recombinant CEP120 and SPICE1 on microtubules in vitro, to
determine whether they stabilize microtubules or alter polymerization rates. Combinations of
recombinant CEP120, SPICE1, CEP135 and CPAP could also be tested.
Previous studies have shown that there is a cell-cycle dependent elongation of procentrioles,
continuing through S and G2 until they reach full length in mitosis (Azimzadeh et al., 2009;
Kuriyama and Borisy, 1981). Under depletion of the distal centriole elongation protein hPOC5
for example, the procentriole remains short in G2 and forms only doublet microtubules
(Azimzadeh et al., 2009). While we were not able to detect whether procentrioles had doublets or
triplets, we certainly observed a shorter average length in CEP120/CPAP/SPICE1 depletion
conditions, and fewer microtubules were detectable in procentriole regions. In C. elegans, which
have only single microtubules instead of triplets, there are no identified CEP120, SPICE1 or

66
CEP135 homologs (Carvalho-Santos et al., 2010; Mahjoub et al., 2010). In future, it will be
worth investigating the structure of procentrioles by EM under CEP120/SPICE1 depletion
conditions in more depth, to see if single microtubules, doublet or triplet microtubule are
observed in the absence of CEP120 and SPICE1. Overall, I conclude based on these results that
CEP120 and SPICE1, along with CPAP are important for very early stages of procentriole
microtubule assembly and/or stability. This is consistent with the localization of CEP120 and
SPICE1 along procentriole barrels, as reported here by 3D-SIM and previously by immuno-EM
for CEP120 (Mahjoub et al., 2010).
2.5.4. CEP135 Localization Requires CEP120 and SPICE1, and is a
Microtubule Dependent Event During Centriole Duplication
The application of 3D-SIM to the G2 PLK4 assay gave us the ability to observe features and
structures not normally resolvable by conventional fluorescence microscopy methods. Critically,
we were able to distinguish between localization on parental centrioles versus recruitment to the
region where procentrioles are being assembled, based on localization relative to Myc-PLK4 in
properly oriented centrosomes. This capability made possible the key observation that CEP135
recruitment to procentrioles is lost when CEP120 or SPICE1 are depleted. Using conventional
microscopy, the strong signal from CEP135 stably incorporated into the mother centrioles is not
resolvable from the newly recruited CEP135 participating in procentriole assembly, and this has
been elsewhere reported (Lin et al., 2013a). When we depleted CEP120, SPICE1, CPAP or
CEP135, the levels of CEP135 in the procentriole assembly region decreased significantly.
When colchicine was present, CEP135 levels were reduced in the procentriole assembly region,
but SASS6 levels were not affected. Additionally, CEP120 and SPICE1, along with CP110 and
CPAP were all localized properly despite the lack of procentriole microtubules. One possible
interpretation of this is that CEP135 recruitment/anchoring might be downstream of or dependent
upon procentriole microtubule elongation. CEP135 is thought to be a part of the cartwheel
linkage to the microtubule triplets, so it is possible that trace amounts of CEP135 are present to
establish the first cartwheel(s), but then procentriole microtubule elongation could either precede
or coincide with the incorporation of subsequent cartwheels. Consistent with this, SASS6
structures do not elongate in our CEP120/SPICE1 depletion conditions, but remain as rings or
small puncta, whereas in controls SASS6 becomes resolvable as within distinct procentrioles.

67
Perhaps CEP135 is only stably retained in growing procentrioles once it is incorporated, whereas
other proteins are recruited in excess before assembly of a procentriole. Clearly, CEP135
recruitment to growing procentrioles is distinct from the other proteins examined here, being the
only protein that is strongly affected when microtubules are disrupted. Alternatively, it is
possible that CEP135 delivery to centrosomes is dependent upon a microtubule cytoskeleton.
CEP135 has been shown to bind to p50, a subunit of the dynactin complex which facilitates
microtubule motor-driven cargo transport along microtubules (reviewed in Schroer, 2004), and
importantly both CEP135 and p50 are co-dependent for centrosome localization (Uetake et al.,
2004). Thus CEP135 may be actively transported to centrosomes, but then incorporated or
retained only when procentrioles are being successfully assembled, in a CEP120/SPICE1
dependent process. CPAP also interacts with CEP135, but this is likewise not sufficient for
robust localization in the absence of a microtubule cytoskeleton under colchicine treatment (Lin
et al., 2013a). A model combining microtubule dependent CEP135 localization and
CEP120/SPICE1/CPAP dependent CEP135 incorporation/retention into procentrioles resolves
the observations from the colchicine and RNAi experiments. What is not clear yet is whether
CEP120 or SPICE1 are directly involved in CEP135 incorporation into procentrioles, or are
merely required to build/stabilize procentrioles which then retain CEP135. To date, no
interaction between CEP135 and CEP120 or SPICE1 has been published. Clearly, determining
whether CEP120 and/or SPICE1 bind CEP135 is a critical future experiment.
2.5.5. CEP120 and SPICE1 are Required for Tubulin Incorporation into
Procentrioles
Taxol is a microtubule stabilizing chemical, and upon taxol treatment in G2, U-2 OS cells show
abnormal centriole and procentriole elongation. CEP120 and SPICE1 are necessary for
procentriole microtubule assembly and elongation, and one possible mechanism for their
function could be the stabilization of microtubules. Taxol induced elongation is absent when
CEP120, SPICE1 or CPAP are depleted, but not when SASS6 is depleted. Therefore CEP120,
SPICE1, and CPAP are required for the induced elongation of both mother and procentrioles in
G2 arrested cells. From this, I conclude that CEP120 and SPICE1 have a function beyond
merely stabilizing procentriole microtubules, because taxol cannot facilitate centriole elongation
in their absence.

68
Centriole elongation occurs under a ‘cap’ of CP110 that must be removed for ciliogenesis, but
CP110 remains localized to the plus ends of abnormally elongated centrioles (Kleylein-Sohn et
al., 2007; Schmidt et al., 2009; Spektor et al., 2007). Microtubule incorporation into centrioles
could be driven by diffusion and stabilization, or it could require a chaperoned or active delivery
system. In a simple diffusion model where tubulin is under free exchange and proteins provide
additional stability, taxol stabilization should affect the length of procentrioles regardless of the
presence or absence of other proteins. Thus, the incorporation of free tubulin into centrioles at
the centrosome is dependent upon CEP120, SPICE1 and CPAP. CEP120, CPAP and Centrobin
are all important for centriole elongation, and each of these proteins has a microtubule binding
domain (Gudi et al., 2011; Lin et al., 2013a, 2013b; Tang et al., 2009a). In Drosophila, SAS-4
(CPAP) is a tubulin binding protein that helps deliver PCM components to the centrosome
through a mechanism that depends on binding and release of tubulin by SAS-4 (Gopalakrishnan
et al., 2011, 2012). Perhaps a similar mechanism of tubulin dimer-delivery explains how human
CEP120 and CPAP function in centriole elongation. In CEP120 the microtubule binding domain
is essential for elongation upon overexpression, whereas the tubulin dimer binding domain of
CPAP is essential for elongation upon its overexpression (Lin et al., 2013b; Tang et al., 2009a).
It remains to be shown whether CEP120 binds tubulin dimers, versus microtubules exclusively,
as the assay used to demonstrate the interaction was done in the presence of taxol (Lin et al.,
2013b). SPICE1, although it localizes to the spindle, has not had a microtubule binding activity
demonstrated or characterized in vitro. Addressing how CEP120 and potentially SPICE1 interact
with microtubules and/or tubulin dimers will be a very important step towards understanding
their functions in procentriole elongation. Future experiments should be directed at determining
whether CEP120 and SPICE1 bind free tubulin dimers or polymerized microtubules in vitro, and
whether this is dependent upon the nucleotide bound by the tubulin (GTP vs. GDP), as it is in the
case of Drosophila SAS-4 (Gopalakrishnan et al., 2012). Further, the impact of
CEP120/SPICE1 on polymerized microtubules, specifically whether each protein can
stabilize/destabilize them, or affect polymerization dynamics, should be determined.
Many questions arise from our work. Most importantly, we have very little understanding of the
exact molecular mechanism(s) by which tubulin dimers are incorporated into centrioles. Are
CEP120, CPAP, Centrobin and other proteins each adding tubulin dimers in a redundant yet
robust system of microtubule incorporation? How do the various dependencies and interactions

69
among these proteins relate to their tubulin binding? How does the tubulin incorporation system
link with addition of core centriole or PCM proteins? How do CP110, CEP97 or other negative
regulators of centriole length counterbalance this system? Is it only the A-microtubule of the
triplet that is regulated this way, since it is capped by a -TuRC at one end and CP110 at the
other, whereas B and C microtubules assemble bi-directionally along the A-tubule and are not
nucleated by a -TuRC? (Guichard et al., 2010). Understanding this process is critical to
understanding how centrioles are built to consistent lengths.
While many questions remain, our work here reveals a role for CEP120 and SPICE1, in
cooperating with CPAP in procentriole assembly and elongation. Further, CEP120 and SPICE1
are essential for each other’s recruitment, along with recruitment of CEP135, CP110 and CEP97
during procentriole assembly.

70
2.6. Figures
Figure 2.1.
Figure 2.1. CEP120 is required for centriole duplication in HeLa and U-2 OS cells.
(A) (Left) IF microscopy images of fixed HeLa cells at 72 hours post transfection with esiRNA
against firefly Luciferase (Control) or CEP120. Cells were labeled for DNA (blue), CETN2
(green) and PCNT (red). Boxed insets show 2x magnified centrioles as labeled by CETN2.
Scale bars 5 m. (Right) Histogram showing the percentage of mitotic HeLa cells in which either
0 or 1-4 centrioles were detectable. White histogram bars indicate Luciferase RNAi, black
histogram bars indicate CEP120 RNAi. (B) HeLa cells were treated with esiRNA as in (A), with
or without stable expression of mouse CEP120 fused to a LAP-tag from a BAC. The number of
centrioles were counted in mitotic cells for each condition based on CETN2 labeling, and results
used to generate the histogram shown. No significant differences in centriole number were
observed in the HeLa cells expressing mouse CEP120. (C) Bar graph showing the percentage of
U-2 OS cells in which more than 4 CETN2 foci were observed after 24 hours of RNAi followed
by 68 hours of arrest in S-phase by HU treatment. Error bars represent the standard deviation of
the mean. *p<0.05, **p<0.01 based on unpaired two-tailed student t-test. At least 50 cells were
counted per each replicate, with three replicates per experimental condition.

71
Figure 2.2

72
Figure 2.2. CEP120 and SPICE1 are required for centriole duplication in cycling HeLa
cells.
(A) Immunofluorescence microscopy images of fixed mitotic HeLa cells at 72 hours post
transfection with siRNAs targeting CEP120 or SPICE1, or a control siRNA with no target
sequence. Cells were labeled with DAPI (blue in panels), and antibodies detecting SASS6
(green), CEP135 (red), or CETN2 (blue in insets). (B) Centrioles were counted by examining
CETN2 signal, and the frequency of observed mitotic cells with four or more centrioles is shown
for each condition. (C) Western blot of cell lysates collected from the same experiment showing
(top two rows) CEP120 protein levels under each condition versus tubulin levels under each
condition and (bottom rows) SPICE1 protein levels under each condition versus tubulin levels.
Markers indicate approximate molecular weights based on protein standards. Scale bar in (A) is
10 m, and insets are four-times magnified. (D) HeLa T-Rex cells with or without inducible
expression of siRNA resistant CEP120 were transfected with control or CEP120-si#1 for 24
hours before being induced by addition of tetracycline for 48 hours then fixed in methanol. Cells
were immunolabeled for Centrin (CETN2) and Pericentrin (PCNT) and mitotic cells were
examined. (D) Shows representative images of mitotic cells under the absence of siRNA
resistant CEP120 (left) or in the presence of siRNA-resistant CEP120 (right). Scale bar is 10
m, and insets are approximately 2.5-times magnification. (E) The frequency of mitotic cells
with un-duplicated centrioles was quantified in the experiment described in (D), and bar graph
shows the frequency of cells with failed duplication under each condition. Error bars in (B, E)
indicate SEM from three replicates in which at least 50 cells were counted per condition per
replicate.

73
Figure 2.3.
Figure 2.3. SPICE1 interacts with CEP120 through its coiled-coil domains.
(A) Western blots from IP experiments using CEP120 protein truncations (residues 1-700 and
700-986) or full length CEP120 fused to HA tag as bait. Blot shown using antibodies against
(top) SPICE1 and (bottom) HA following co-IP and for inputs. A model of CEP120 is shown
below indicating the coiled-coil domain and the fragments used for IP. (B) Western blots from
IP experiments using SPICE1 truncations (residues 1-450, 350-750, and 450-856) or full length
SPICE1, each fused to GFP, as baits. (Top) Western blot using antibodies against CEP120 co-IP
in each condition and in respective inputs. (Bottom) Western blot using antibodies against GFP
for each sample. A model of SPICE1 indicating the coiled-coil domains and the fragments used
for IP is shown. Experiments performed by Dr. M. Archinti, Lab of Dr. J. Luders.
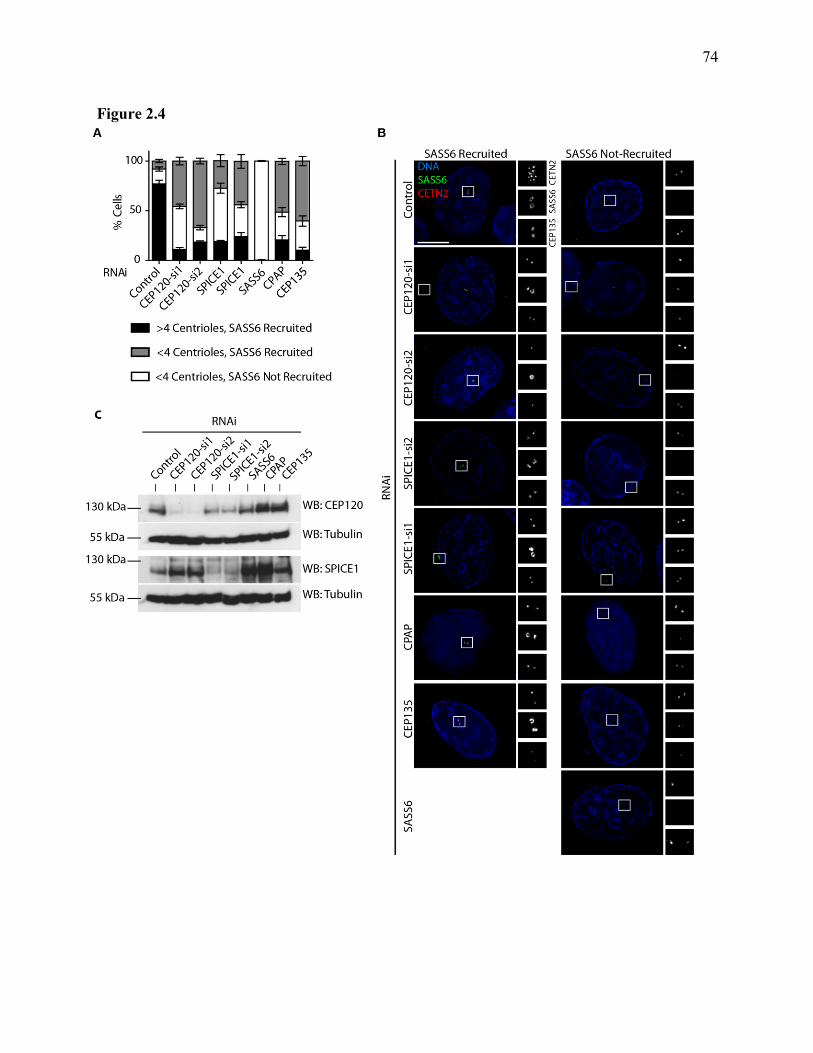
74
Figure 2.4

75
Figure 2.4. CEP120 and SPICE1 Depletion blocks centriole overduplication induced by
PLK4 overexpression in U-2 OS cells.
(A) U-2 OS cells on coverslips were transfected with siRNA or esiRNA against target genes as
indicated, and at 48 hours post-transfection were arrested in S-phase with hydroxyurea and
induced to over-express PLK4 by addition of tetracycline. At 20 hours post induction and arrest,
cells were fixed and lysates were collected for Western blots. Stacked bar graph shows the
frequency of centriole overduplication (black bars) versus non-overduplication (grey and white).
White bars represent the fraction of centrosomes that failed to produce procentrioles and
simultaneously lacked SASS6 localization around the mother centriole. Grey-bars show the
portion of centrosomes where duplication failed but SASS6 was seen localized around the
mother-centrioles. Error bars represent the SEM from three replicates for which at least 100
cells were examined. (B) Representative IF images from each condition where either SASS6
was recruited or SASS6 was not recruited. In large panels, DNA (blue), SASS6 (green) and
CETN2 (red) labels are shown. Insets show CETN2, CEP135 and SASS6 at centrosomes at 2.5x
magnification. No cells were observed with SASS6 recruitment under SASS6 RNAi, so no
image is shown. Scale bar 10 m. (C) Western blots for CEP120 (top) and SPICE1 (third row)
with loading controls (tubulin blots, second and bottom rows respectively) from the same
experiment.

76
Figure 2.5

77
Figure 2.5. CEP120 and SPICE1 depletion block centriole overduplication downstream of
SASS6 recruitment, but lead to abnormal SASS6 localization.
U-2 OS cells expressing Myc-PLK4 under the regulation of the tetracycline-repressor were
transfected with siRNA or esiRNA targeting CEP120, SPICE1, CEP135, CPAP or no-target at all
(control) for 24 hours before being arrested in S-phase by addition of hydroxyurea and induced to
over-express Myc-PLK4 by tetracycline addition. 24 hours after S-phase arrest and induction,
cells were rinsed and arrested in G2 by addition of RO-3066 for 16 additional hours before
fixation and cell lysate collection. (A) Representative IF images of cells from each condition
showing DNA (blue) and SASS6 (green), CETN2 (red). Insets show centrosomes at 4x
magnification (monochromatic) and 8x magnification (colour). Scale bar 10 m. (B) Stacked
bar graph showing the relative frequency of centriole overduplication (black bars) versus non-
overduplication (grey and white). White bars represent the fraction of centrosomes that failed to
produce procentrioles and simultaneously lacked SASS6 localization around the parental
centriole. Grey-bars show the portion of centrosomes where duplication failed but SASS6 was
seen localized around the parental centriole. (C) Quantification of the percentage of cells from
each condition where the SASS6 localization appeared smoothed as a ring-like shape around the
parental centriole, versus punctate allowing distinction of individual procentrioles (see Control
versus CEP120 IF image insets). For (B) and (C), error bars represent the SEM from three
replicates for which at least 100 cells were examined. For (C), significance is indicated as a result
of an unpaired two-tailed student’s t-test against the control. (D) Western blots for proteins
depleted in this experiment were prepared using lysate from the same experiment by Sally
Cheung. Western blots for -tubulin as a loading control are immediately below their respective
blots.

78
Figure 2.6

79
Figure 2.6. Three-dimensional SIM imaging of PLK4 induced centriole overduplication. (A) (Left, Top) Representative immunofluorescence image of S-phase arrested un-induced U-2
OS cells showing cells fixed and immunolabeled for Centrin (CETN2) and Myc-PLK4 versus
(right) representative images from induced U-2 OS cells over-expressing Myc-PLK from the
same experiment. (Bottom) Models showing the presumed orientation of the centrioles from the
un-induced cell (left) versus the easily defined orientation of the procentrioles surrounding
parental centrioles in the induced Myc-PLK4 over-expressing cell (right). (B) 3D-SIM images
of centrosomes after Myc-PLK4 overexpression in U-2 OS. Cells were labeled with antibodies
against CETN2, CP110 and SASS6 for immunofluorescence imaging. Centrosomes were
imaged where the parental centriole was oriented along the z-axis, and thus procentrioles are
seen side-on in the images. Centrosomes were selected at various stages of procentriole
elongation, increasing from left to right, as indicated by the space between CP110/CETN2 and
the mother centriole. (C) Immunofluorescence images of similarly oriented centrosomes from
the same conditions labeled for CEP120 (left) or SPICE1 (right), Myc-PLK4 and CETN2.
Dashed circles indicate the location of the Myc-PLK4 signal throughout the images. White “M”
indicates the location of the mature (parental) centriole for the CEP120 labeled images. Scale
bars are each 1 m.

80
Figure 2.7

81
Figure 2.7. Impact of CEP120 and SPICE1 on the recruitment and localization of centriole
assembly proteins.
3D-SIM images of centrosomes from the G2 PLK4 assay where cells were treated with non-
targeting siRNA (Controls), or siRNA against CEP120 or SPICE1. Cells were labeled with
antibodies against CETN2 and the Myc epitope tag, along with one of the procentriole assembly
proteins CEP152, SASS6, STIL, CPAP, CEP97, CP110 or hPOC5. For each staining, the
specific marker is shown in green at the top of the first column, and the last column where the
inset shows that marker alone. Scale bars represent 1 m. Images are scaled so that intensities
are quantitatively comparable within a given 3x4 panel, with the exception of Myc-PLK4 signal
which has been brightened in each Control to facilitate visibility.
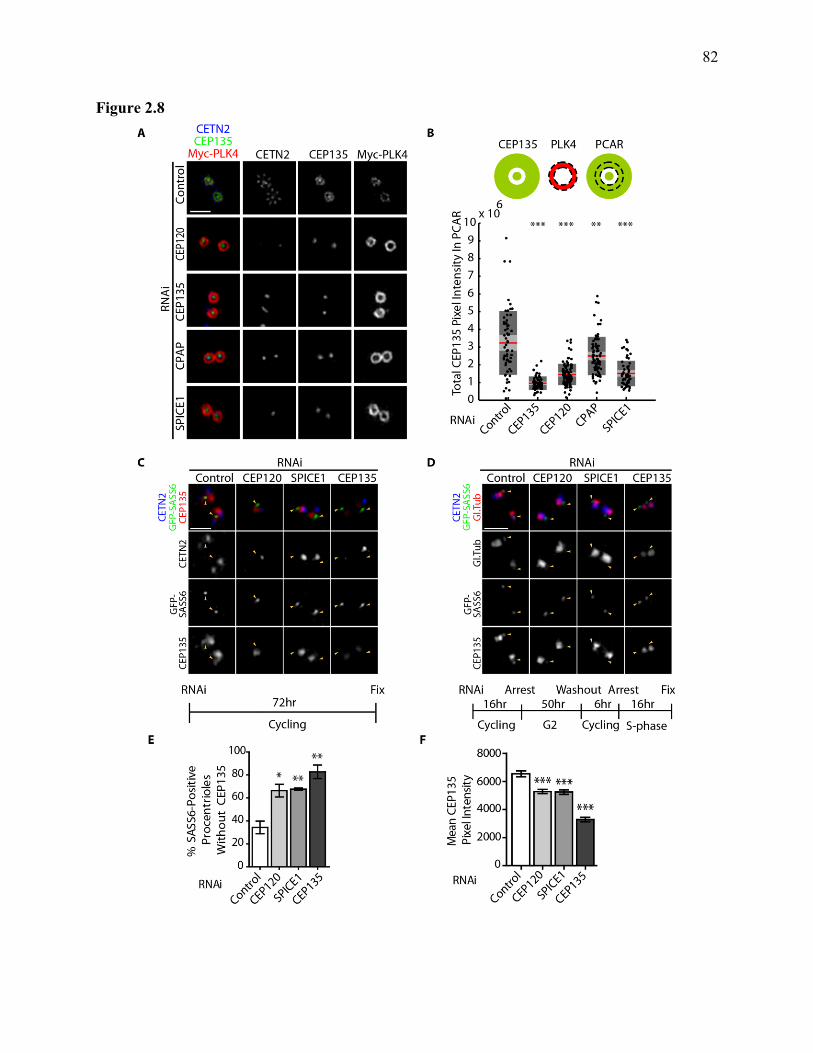
82
Figure 2.8

83
Figure 2.8. Depletion of CEP120 or SPICE1 leads to failure of CEP135 recruitment during
centriole duplication. (A) Representative images of centrosomes observed in the G2 PLK4
assay when CEP120, SPICE1, CPAP or CEP135 were depleted. Centrosomes were labeled with
antibodies against Myc-PLK4, CETN2 and CEP135. (B) Quantification of total pixel intensity
of CEP135 in the procentriole assembly region (PCAR) of 3D-SIM images of correctly oriented
centrosomes in the absence of CEP120, SPICE1, CPAP or CEP135 compared to an un-depleted
control, from a G2 PLK4 assay. Top: Outline of the masking approached used for image
quantification in MatLab by Dr. Gupta. The dashed region defined by Myc-PLK4 signal (red)
was masked and that mask applied to the CEP135 signal (green). The Cep135 intensity within
the Myc-PLK4 defined region was quantified. Bottom: Box and whisker plot showing total pixel
intensities from CEP135 overlapping Myc-PLK4 signal for each centriole from the various
depletion conditions. Multiple centrosome images were chosen for each condition based on
orientation of the centrioles (n=62, 59, 104, 78, 60 from left to right across X axis). Red bar
shows the mean, light-gray represents the 95% confidence interval, and darker gray outlines one
standard deviation. (C) Cycling U-2 OS cells expressing SASS6-GFP were depleted of CEP120,
SPICE1 or CEP135 as indicated for 72 hours. Cells were labeled with antibodies against GFP,
CEP135, and CETN2. Representative centrosomes were selected in S-phase cells for each
condition. Yellow arrowheads indicate position of SASS6 positive procentrioles in each case,
and are positioned consistently in every image within each column. (D) U-2 OS cells were
synchronized as indicated (bottom) to be in S-phase with two mother centrioles at the time of
fixation. Cells were labeled with antibodies as in (C), and arrowheads similarly indicate SASS6
positive procentrioles within each column. (E) Bar graph of the fraction of procentrioles under
each condition from (D) that were observed to have SASS6 but not CEP135. Bar graph shows
mean with error bars indicating the SEM from three experiments, with at least 50 centrosomes
examined for each condition. (F) Bar graph of the mean CEP135 pixel intensity overlapping
SASS6 signal from SASS6 positive procentrioles in cells treated and fixed as in (D). Bar graph
shows the mean from three experiments (total number of centrioles quantified is 158, 164, 112,
166 respectively, from left to right on the x-axis of the graph). Error bars indicate the SEM. All
images are from 3D-SIM experiments acquired quantitatively and are scaled identically for each
channel within a given panel, except for Myc-PLK4 in (A) which is scaled for visibility in the
image from the control condition.

84
Figure 2.9

85
Figure 2.9. Recruitment dependencies of CEP120 and SPICE1 during PLK4 induced
centriole overduplication.
For all three panels (A-D), G2 PLK4 assays were performed with RNAi as indicated and cells
were fixed and labeled with antibodies against CEP120 (left) or SPICE1 (right), CETN2, and
Myc-PLK4. Representative quantitatively imaged 3-D SIM micrographs of centrosomes are
shown with equal intensity scaling within each panel, with the exception of Myc signal which is
enhanced for visibility in controls. (A) The early centriole assembly proteins CEP152, STIL or
SASS6 were depleted in the G2 PLK4 assay. Parental centrioles are oriented differently in
images from CEP152 depleted cells to show CEP120 and SPICE1 localization along the parental
centriole barrel. (B) CEP120, SPICE1, CPAP or CEP135 were depleted in the G2 PLK4 assay.
(C) The distal centriole proteins CEP97 and CP110 were depleted in the G2 PLK4 assay. (D)
The centriole elongation proteins POC1 and hPOC5 were depleted in the G2 PLK4 assay. Scale
bars 1 m for each panel.

86
Figure 2.10

87
Figure 2.10. Depletion of CEP120 or SPICE1 results in procentrioles with short or
undetectable microtubules following PLK4 induced centriole overduplication.
A G2 PLK4 assay was performed and the U-2 OS cells were fixed and prepared for serial
electron microscopy (see Materials and Methods). For each condition, two example centrosomes
are shown. From left to right, micrographs of 100 nm serial sections are shown for the same
centrosome. Yellow arrowheads point to approximate locations of procentrioles in controls, or
electron density at the proximal end of the mother centriole in depletion conditions. Scale bar
100 nm.

88
Figure 2.11

89
Figure 2.11. Depletion of CEP120 or SPICE1 results in shorter procentriole microtubules
following PLK4 induced centriole overduplication. (A) Tomogram projections (average projections of 20-30 nm central volume sections) of mother
and procentrioles from cells collected and processed following a G2 PLK4 assay where either no
protein, or CEP120, SPICE1 or CPAP were depleted. (Left) Original tomograms are shown,
with duplicate images (right) where microtubules are highlighted in green. Scale bar represents
100 nm. Bar graph (B) shows mean lengths of individual microtubules of procentrioles and (C)
shows mean cross-sectional widths of detectable procentrioles measured from multiple
tomograms for each condition using IMOD software. For (B) n=16, 12, 16 and 14 microtubules
measured, for (C) n=6, 5, 6, and 3 procentrioles measured for controls, CEP120 depleted,
SPICE1 depleted or CPAP depleted conditions respectively. Widths were determined based on
widest detectable point in tomograms under all conditions. Error bars indicated standard error
of the mean. Tomography acquisition and reconstruction, as well as measurements in IMOD
were performed by Dr. Eden Fussner.

90
Figure 2.12.

91
Figure 2.12. Colchicine treatment effectively depolymerizes cytoplasmic microtubules and
prevents procentriole microtubule assembly. (A) U-2 OS cells over-expressing Myc-PLK4
were arrested in S-phase and induced to over-express Myc-PLK4 for 24 hours before being
rinsed and released into G2 for a final 16 hours before fixation. Fixation was performed with
additional pre-extraction in PBS-triton to remove soluble tubulin, then labeled with antibodies
against CEP135, -tubulin and the Myc epitope tag. Throughout the entire experiment, cells
were maintained with or without 10 M colchicine. (Left) Two examples of cells without
colchicine added, where the top shows a clear microtubule network, and the bottom shows a cell
where the centrosome and centriolar microtubules are visible. (Right) Two examples of cells
maintained with colchicine where centrosomes are visible that include mother centrioles with
detectable microtubules, but lack detectable procentriole microtubules. Images are deconvolved
and projected from wide-field fluorescence microscopy. Scale bar 10 m, insets 4x
magnification. (B) 3D-SIM images of centrosomes from the same experimental setup and
labeling as in (A). Untreated (left) or colchicine treated conditions (right), scale bar 1 m.

92
Figure 2.13

93
Fig 2.13. Localization of centriole assembly proteins to the procentriole assembly region in
the absence of microtubules.
(A) (Left) Representative images from 3D-SIM of U-2 OS induced to over-express PLK4 in the
presence or absence (control) of colchicine. Cells were induced and arrested in S-phase for 24
hours before being released into G2 for another 16 hours prior to fixation. Cells were labeled
with antibodies against CEP135, glutamylated tubulin and SASS6. Scale bar 1 m. (Right):
Box-and-whisker plots of the total integrated intensity of each of the three labels for the whole
centrosome for multiple centrosomes as prepared for (A). At least 50 3D-SIM images were used
for the measurement of each condition. Red bar shows the mean, light-grey the 95% confidence
interval and dark grey bar is 1 standard deviation above and below the mean. (B) Cells were
treated as in (A), but labeled with antibodies against tubulin and a centriole assembly protein
as indicated. Images are representative from 3D-SIM imaging, scale bar 1 m.

94
Figure 2.14

95
Figure 2.14 CEP120 overexpression leads to abnormally elongated centrioles in G2. (A) U-2 OS cells were transiently transfected with plasmids expressing SPICE1, CEP120 or
CPAP as fusions to GFP for 24 hours before being fixed and labeled with antibodies against GFP
and acetylated tubulin. Representative images are shown for each condition. Scale bar 10 m,
insets are 2x magnified. (B) The frequency of abnormally elongated centrioles was quantified
following each overexpression experiment in (A), with three replicates performed per
transfection. Bar graph shows the mean from replicates, with at least 100 cells per condition per
replicate examined, and error bars show the standard error of the mean. (C) A U-2 OS Flp-In
TREx cell line was used for inducible expression of CEP120 fused to a FLAG tag. Cells were
induced to over-express FLAG-CEP120 for 24 hours in the presence of HU (S-phase, left panel)
or RO-3066 (G2, right panel). Cells were fixed and labeled for CEP120 and acetylated tubulin.
Representative images are shown of (top) un-induced cells or (bottom) induced cells in either S-
phase or G2 arrest. (D) Quantification of the frequency of abnormally elongated centrioles in
each condition from (C), with bar graph showing the mean of three replicates for each condition,
with at least 100 cells examined per condition. Error bars show the standard error of the mean,
and asterisks show result of an un-paired two-tailed student’s t-test comparing the induced and
un-induced conditions.

96
Figure 2.15

97
Figure 2.15 CEP120, CPAP and SPICE1 are all required for abnormal centriole elongation
following CEP120 or CPAP overexpression in G2.
U-2 OS cells were depleted of proteins as indicated for 24 hours before being transfected with a
plasmid driving expression of either GFP-CEP120 (A-C) or GFP-CPAP (D-F) for another 24
hours before being arrested in G2 for 48 hours then fixed. (A) Representative images of cells
labeled with antibodies against CEP164, GFP and glutamylated tubulin from each experimental
condition. Scale bars 10 m, insets are 2x magnifications. (B, E) Quantification of the
frequency of abnormal centriole elongation within cells expressing CPAP-GFP (B) or CEP120
GFP (E) from each experimental condition. (C, F) Quantification of the fraction of transfected
cells where the mother centriole was abnormally elongated following the experiment described
in (A) or (D) respectively. Bar graphs show the means from three replicates for which at least
100 cells were counted, for each experiment (B, C, E, and F). Error bars show standard error of
the mean, asterisks are results of unpaired two-tailed student t-test of each condition against the
control.

98
Figure 2.16
Figure 2.16. CPAP interacts with CEP120 in vivo.
Co-immunoprecipitation experiments were performed in HeLa cells transiently transfected
with FLAG-CEP120 or FLAG-USP37 as a control, and one of GFP-USP37, SPICE1-GFP
or CPAP-GFP. Western blots were performed to detect (top) GFP epitope from FLAG-IPs,
(middle) FLAG-epitope from FLAG-IPs, and (bottom) GFP epitope in the lysates from the
experiment before IPs were performed. This experiment was performed by Sally Cheung.

99
Figure 2.17

100
Figure 2.17. CEP120, SPICE1 or CPAP depletion inhibits taxol induced centriole
elongation in G2 arrested U-2 OS cells. U-2 OS cells were depleted of select proteins by RNAi for 24 hours before being arrested in G2
for 48 hours, where taxol was added to 10 M final concentration for the last 24 hours. (A)
Representative images of U-2 OS cells under each depletion condition. Cells were fixed and
labeled with antibodies against CEP164 and glutamylated tubulin. Scale bar represents 10 m,
insets are 4x magnified from the merged pseudo-coloured images. An experimental timeline is
shown underneath the representative images. (B) The fraction of cells under each condition
where at least one centriole or procentriole was elongated abnormally was quantified, bar graph
shows mean from three replicates, where at least 100 cells were examined per replicate per
condition. Error bars show standard error of the mean, asterisks show results of an un-paired
two-tailed student t-tests comparing each condition to the control condition.

101
Figure 2.18

102
Figure 2.18 CEP120 and SPICE1 Functions Centriole Assembly.
(A) (Top) The process of procentriole assembly on a single parental centriole is shown (see text
for references). (From top-left) In G1 PLK4 is first recruited as a ring to CEP192 on the parental
centriole, then migrates farther away from the centriole barrel by interacting with CEP152 which
is newly recruited in G1. At the start of S-phase, the formation of a SASS6 cartwheel occurs
within the lumen of each disengaged parental centriole. The recruitment of SASS6 into the
parental lumen is mediated through binding to CPAP therein. Subsequently, dependent upon
PLK4 and STIL, SASS6 is released from the parental centrioles and localizes to the proximal
outer wall of the centrioles along with a single focus of PLK4 and STIL. From there, assembly
of a procentriole is initiated requiring the combined activities of -tubulin, /-tubulin,
Centrobin, CPAP, CEP120, SPICE1, CEP135 and CP110. In S-phase, the elongation of the
procentrioles is driven by CPAP, CEP120, SPICE1, Centrobin and Poc1, whereas in G2
elongation is further promoted by CPAP, Centrobin,CEP120, SPICE1, hPOC5 and OFD14. The
figure legend is shown below the pathway. (B) A summary of the proteins known to promote or
restrict centriole elongation is shown, with the affected portion of the centriole indicated. *OFD1
restricts the length of the parental centriole specifically. (Bottom) Legend is shown.

103
2.7. Materials and Methods
2.7.1. Cell lines and Tissue Culture
U-2 OS and HeLa cells were grown in Gibco brand McCoy’s 5A media supplemented with fetal
bovine serum (10%) and 2 mM L-glutamine at 37°C. U-2 OS T-REx cells with Tet-inducible
Myc-tagged PLK4 were a kind gift from Dr. E. Nigg, and were maintained as above but with
certified tetracycline-free fetal bovine serum, and G418 at a concentration of 0.5 mg/ml for
selection. FLAG-CEP120 inducible U-2 OS T-REx cell lines were generated as per
manufacturer’s recommendation (Invitrogen). Briefly, Flp-In T-REx U-2 OS cells (Invitrogen)
in 6-well dishes at 80-90% confluence in 1 ml Opti-MEM were transfected with 2 g pOG44
and 200 ng of the pCDNA5-FRT-TO-FLAG-CEP120 plasmid using 2.5 l Lipofectamine-2000
in a total volume of 200 l Opti-MEM media. Reaction mixtures were added to each well for 3
hours before an equal volume (1.2 ml) of McCoy5A with 20% FBS was added. Cells were
incubated overnight, then transferred into 10 cm dishes for another 24 hours before Hygromycin-
B was added to the media (200 g/ml final concentration) for selection. Clones (pools) were
picked using colony-lifts into individual wells, then maintained in tetracycline-free media before
being tested for inducible CEP120 expression. To induce expression of Myc-PLK4 or FLAG-
CEP120 from T-REx cells, tetracycline was added to media to a final concentration of 2 g/ml.
2.7.2. RNA Interference
For a list of primer sequences for esiRNAs and commercially purchased siRNAs used in this
study, see Appendix I: Supplemental Methods. For depletion of CPAP and SASS6 in this study,
esiRNA were synthesized as previously described (Kittler et al., 2005). For all cell assays, cells
were seeded on sterilized glass coverslips in 6-well tissue culture dishes and allowed to settle for
at least 12 hours before treatment with siRNA or esiRNA. RNAi transfection mixes were
prepared using Invitrogen Lipofectamine RNAiMAX. For siRNA, we prepared stocks at
concentrations of 20 M, and for esiRNA we prepared stocks with concentrations of 100 ng/l.
For each well, we prepared one mix with 5 l of siRNA and 245 l Gibco OptiMem media or 15
l esiRNA with 235 l Gibco OptiMem, and mixed these with a separately prepared mixture of
2.5 l of Lipofectamine RNAiMAX in 247.5 l OptiMem. RNA was incubated with

104
Lipofectamine for a minimum of 20 minutes before being added to 2.5 ml of McCoy 5A media
in each well (final volume 3 ml).
For the CEP120 siRNA rescue, either T-REx HeLa or T-REx HeLa expressing siRNA resistant
FLAG-tagged CEP120 were seeded. The next day cells were transfected with siRNA, and at 24
hours post transfection were treated with 1μg/ ml tetracycline for 48 hours before fixation. This
experiment, and generation of the T-REx HeLa cell line expressing FLAG-CEP120, were
performed by Deborah Pinchev.
2.7.3. Cloning of CEP120 siRNA Resistant Construct
CEP120 was mutated at wobble-codon positions using QuikChange II XL site-directed
mutagenesis kit (Agilent Technologies) by Deborah Pinchev. For CEP120 siRNA-1 resistance,
the primers used were 5’- GATGATTATTTGACTCGCCTGATAGAAGAAAGGGATACTTTT
ATGCGGACAGGCGTTTACAATCACGAGGATCGAATAATAAGTGAACTCGACCGACA
G-3’ and 5’- CTGTCGGTCGAGTTCACTTATTATTCGATCCTCGTGATTGTAAACGCCTG
TCCGCATAAAAGTATCCCTTTCTTCTATCAGGCGAGTCAAATAATCATC-3’.
2.7.4. PLK4 Induced Centriole Overduplication Assays
For the S-phase assay initially used (Figure 2.4), U-2 OS T-REx cells with inducible myc-PLK4
were seeded in 6-well dishes and treated with RNAi as described above. At 48 hours post-RNA
addition, cells were arrested in S-phase by addition of hydroxyurea to a final concentration of 8
mM, and induced to over-express PLK4 by addition of tetracycline (2g/mL) for 20 hours
before fixation.
For the ‘G2 PLK4 assay’, RNAi was as above, but at 24 hours post-RNA addition the cells were
arrested in S-phase using HU and induced by tetracycline addition. After 24 hours in S-phase
arrest, cells were rinsed twice with 3 mL sterile PBS (Gibco) and put into fresh McCoy 5A
media containing 10 M RO-3066 (Enzo Life Sciences) to arrest them in G2. At 16 hours post
G2 arrest, cells were fixed on coverslips for microscopy.
For the PLK4 assay in the presence of colchicine, cells were arrested in S-phase and induced to
express PLK4 as described for 24 hours in the presence of colchicine, then rinsed and arrested in

105
G2 as described for 16 hours before fixation. Colchicine (Sigma) was maintained at 10 M from
the time of S-phase arrest until fixation.
2.7.5. Taxol Induced Centriole Elongation
For the taxol induced centriole elongation assay, U-2 OS cells were seeded and transfected with
RNA as described, and at 24 hours post-RNA addition, cells were arrested in G2 using RO-3066
as described. After 24 hours of G2 arrest, taxol (Sigma) was added to 10 M final concentration,
and cells were maintained for another 24 hours in taxol before fixation.
2.7.6. CPAP or CEP120 Induced Centriole Elongation
For inducible FLAG-CEP120 expression in testing of the cell-cycle specificity of elongation,
cells were seeded at equal densities and the next day induced with tetracycline (2g/mL) in the
presence of either 10 M RO-3066 (Enzo Life Sciences) to G2 arrest, or 8 mM HU, and
incubated under these conditions for a subsequent 24 hour period before fixation and preparation
for microscopy. For assays quantifying centriole elongation by CEP120, SPICE1 and CPAP, U-
2 OS cells were seeded on coverslips in 6-well dishes for at least 12 hours before plasmid
addition. 2 g of plasmid was combined with 3 L of Lipofectamine-2000 (Invitrogen) to a final
volume of 500 L in OptiMem, incubated for 20 minutes, then added to 2.5 mL of McCoy-5A
media in each well (final volume 3 mL). At 24 hours post-addition of plasmid, cells were fixed
for microscopy. For overexpression in combination with RNAi, cells were transfected first with
siRNA or esiRNA (described above) for 24 hours, then subsequently with plasmid as described.
At 24 hours post plasmid addition, cells were arrested in G2 using RO-3066 at 10 M (Enzo Life
Sciences) for 48 hours, then fixed as described below.
2.7.7. Immunofluorescence Microscopy (IFM) and 3D-SIM
For IFM, cells on coverslips were fixed by submersion into pure methanol maintained at -20°C,
and incubated at -20°C for a minimum of 15 minutes. Cells were blocked by submersion in PBS
with 0.2% fish-skin gelatin (Sigma) at room temperature for 10 minutes (hereafter PBS-FSG).
Primary antibody solutions were prepared in PBS-FSG, and glass coverslips were inverted onto
droplets of antibody-PBS-FSG solution on clean Parafilm (Parafilm M) in a humidified chamber
for 30-60 minutes at room temperature. Coverslips were then floated off the Parafilm by

106
pipetting 500 l PBS-FSG and transferred back to PBS-FSG for 15 minutes to wash off excess
primary antibody. Secondary antibodies were mixed in PBS-FSG at 1/500 dilution, along with
DAPI or Hoechst 33342 (Sigma) at 1 g/mL, and coverslips were again inverted onto droplets
for 30-60 minutes on clean Parafilm in a humidified chamber at room temperature. Coverslips
were again floated off the Parafilm and submerged in fresh PBS-FSG for a final rinse for 15
minutes. Coverslips were then submerged in ultra-pure distilled water for 1 minute before being
inverted onto drops of ProLong Gold mounting media (Molecular Probes/Invitrogen). A list of
antibodies used is provided in Appendix I.
For any images being compared as quantitative or scaled quantitatively in terms of intensities,
acquisition conditions (exposure time, number of z-sections imaged, light intensity, microscope
used) were identical for each image, and imaging was scaled to avoid pixel saturation. Post-
acquisition, or post reconstruction for 3D-SIM images, images were exported as tiff files of
either 8 or 16 bit depth with identical scaling from SoftWorx, then adjusted identically in Adobe
Photoshop before being used in figures. Images were scaled to avoid saturation at all times.
Image quantifications were performed using MATLAB software by Dr. Gagan Gupta, using
scripts authored by Dr. Gagan Gupta. For 3D-SIM image quantifications, reconstructed and
aligned, maximum-intensity projected non-saturated images were used. For each image set, the
centrosome was identified as being within a region where the maximum overall intensities were
present. Within the region of the centrosome, ‘masks’ were generated using consistent signal-to-
noise and size thresholds for each image channel specific for the protein of interest detected in
that channel. Recruitment to the centrosome for a given protein was quantified as the total pixel
intensity of its masked region. The PCAR mask was defined as the region at the centrosome that
was marked by the Myc-PLK4 label. Recruitment to the PCAR was quantified as the total pixel
intensity of a given label where that label overlapped the PCAR mask. For CEP135
quantification with the SASS6 signal in S-phase synchronized cells (Figure 2.8), SIM images
were exported as quantitatively scaled .tiff files, and opened in IMAGEJ (FIJI build). A defined
size region of interest (ROI) was overlaid on the SASS6 signal from each procentriole centriole,
and the mean CEP135 pixel intensity was obtained for that area from the software. All imaging
was performed on Delta Vision microscopes (GE Healthcare-Applied Precision), with a 40X NA
1.35, 60X NA 1.42, or 100X NA 1.4 plan apochromat oil objective (Olympus). Image scaling

107
and .tiff files were generated by SoftWoRx proprietary software. 3D-SIM was performed using
a Delta Vision OMX microscope, with reconstruction and image alignment performed in
SoftWoRx (GE-Healthcare-Applied Precision) as previously described (Lawo et al., 2012) with
either 60X NA 1.42, or 100X NA 1.4 plan apochromat oil objectives (Olympus).
2.7.8. Cloning
All cloning was performed using KOD DNA polymerase (Toyobio EMD Millipore), restriction
enzymes from New England Biolabs, and T4 DNA ligase from Fermentas (Rapid Ligation Kit).
A complete list of plasmids used in this work is provided in Appendix I. Briefly, the SPICE1
EGFP constructs (full length and fragments) were provided by the lab of Dr. Jens Luders
(Archinti et al., 2010). HA-CEP120 fragments used in the Co-IP experiments were cloned into
pCS2-HA-FA, a pCs2plus based plasmid provided by Guowei Fang (Genetech). Mouse CEP120
plasmid used in rescue experiments was a generous gift from the lab of Dr. Tsai (Xie et al.,
2007).
2.7.9. Statistical Methods
All reported probabilities (p-values) are from two-tailed unpaired student t-tests, performed using
GraphPad Prism 5 software. Experiment sample numbers and replicates are as indicated in
figure legends. The following shorthand is used throughout: *** p<0.001, **p<0.01, *p<0.05.
2.7.10. Western Blots
For RNAi verifications, cell lysates were collected by addition of Laemmli buffer directly to
cells in tissue culture wells following removal of the corresponding coverslip for fixation and
imaging. Proteins were resolved using 8% SDS-Polyacrylamide gels subjected to
electrophoresis (SDS-PAGE), then transferred to PVDF membranes (Immobilon-P, Milipore).
PVDF membranes were dried after transfer as per manufacturer’s recommendations to improve
protein adhesion. Membranes were then incubated in primary antibodies in a Tris-Buffered-
Saline buffer containing 0.1% Tween-20 (TBST), with 5% skim-milk powder (BioShop) and 5%
BSA Fraction V (OmniPur). Primary antibodies were incubated with blots for 12-16 hours
overnight at 4°C. Following primary antibody treatment, membranes were rinsed four times for
ten minutes in TBST alone, then incubated with secondary HRP conjugated antibodies (Mouse,

108
Rabbit, or Goat, Jackson ImmunoResearch) for 1-2 hours at room temperature followed by a
repeated set of four rinses in TBST. Western blots were developed using SuperSignal Pico or
Dura reagents (Pierce/Thermo Scientific) and Amersham-Hyperfilm HCL films were used to
detect signals (GE Healthcare). Blots were digitized using a Canon scanner as tiff images.
2.7.11. Electron Microscopy
Samples were prepared as described in previous publications (Ahmed et al., 2008; Fussner et al.,
2012). Cell samples were provided to Doug Holmyard in Mt. Sinai as pellets fixed in 2%
glutaraldehyde. Doug Holmyard prepared samples using 1% osmium tetroxide followed by
dehydration in an ethanol series, and subsequent infiltration in propylene-Quetol-Spurr resin and
polymerization. 70-100 nm sections were contrast enhanced with 1% uranyl acetate. For
tomography, 10 nm gold particles were added onto samples to act as fiducial markers for
alignment of tilt-series images, an d all samples were coated with 3-5 nm carbon film by direct
evaporation to increase sample stability. Transmission EM images were all collected using a
Tecnai20, FEI operated at 200 kV and collected using an AMT side mount camera. All
tomography data sets were collected by Dr. Eden Fussner based on centrosomes I selected from
each condition. For tomography, zero-loss images were acquired with a post-column Gatan
imaging filter at tilt angles +/- 55 degrees with 2 degree increments using SerialEM
(Mastronarde, 2005). The images were aligned and processed using the weighted back-projection
implementation within the IMOD software program (Kremer et al., 1996; Mastronarde, 2005)
Microtubule length and centriole width measurements were performed in averaged tomographic
slabs from central volumes of 20-30 nm thickness using IMOD. Projections of these central-
slabs are shown in figures.

109
3. Chapter III:
Identification of CEP120 and SPICE1 Associated
Proteins Required for Centriole Duplication and
Elongation

110
3.1. Statement of Contribution, Rights and Permissions
Some of the data in this chapter has been published as part of a report in the journal Current
Biology:
David Comartin*1,2, Gagan D. Gupta1, Eden Fussner3,4, Etienne Coyaud5,6, Monica Hasegan1,
Marco Archinti7, Sally W.T. Cheung1, Deborah Pinchev1, Steffen Lawo1,2, Brian Raught5,6,
David P. Bazett-Jones3,4, Jens Luders8, and Laurence Pelletier1,2. CEP120 and SPICE1 Cooperate
with CPAP in Centriole Elongation, Current Biology.2013 July 22. 23(14): 1360-1366.
This publication was a collaboration involving researchers from:
(1) The Samuel Lunenfeld Research Institute at Mount Sinai Hospital, Toronto Ontario Canada.
(2) The Department of Molecular Genetics at the University of Toronto, Toronto Ontario
Canada.
(3) The Department of Biochemistry at the University of Toronto, Toronto Ontario Canada.
(4) The Hospital for Sick Children, Toronto, Ontario, Canada.
(5) The Ontario Cancer Institute, Toronto, Ontario, Canada.
(6) The Department of Medical Biophysics at the University of Toronto, Toronto, Ontario,
Canada.
(7) ERA Biotec, Barcelona, Spain.
(8) Institute for Research in Biomedicine (IRB Barcelona), Barcelona, Spain.
Sally Cheung (Pelletier Laboratory) prepared and processed all the samples for the BioID
experiment, including cloning BirA*-tagged constructs discussed, generating and validating
inducible BirA* cell lines and preparing cell extracts for mass spectrometry.
Stine Morthorst (Pedersen Laboratory, University of Copenhagen, Denmark) collaborated with
our lab, working on NAP1L1 functional characterization. Stine provided the NAP1L1
expression constructs used here, cloned one of the siRNA resistant NAP1L1 constructs, and
assisted in quantification of some of the PLK4 assays wherein NAP1L1 siRNAs were tested.
Dr. Etienne Coyaud (Raught Laboratory) collaborated to process and analyze all the BioID
experimental samples, including affinity purification and mass spectrometry of cell pellets for
FLAG-BirA* controls, FLAG-BirA*-CEP120 and FLAG-BirA*-SPICE1 samples.
Dr. Mariana Gomez- Ferreria (Pelletier Laboratory) designed PCR primers for amplifying
GAPDH in RT-PCR experiments as a control for total mRNA levels.

111
3.2. Summary
Identification of proteins required for centriole duplication, elongation or mitosis is an important
step towards fully understanding these processes. Here, I describe the use of the BioID method
to identify proteins in the proximity of CEP120 and SPICE1. From these proteins, I identified
several as being required for centriole overduplication following PLK4 overexpression. The
proteins required for overduplication were further tested for possible roles in CEP120/SPICE1
localization. Depletion of several of these proteins led to CEP120 localization to cytoplasmic
microtubules that were abnormally glutamylated. Further, I identified several proteins that were
required for centriole elongation in a taxol induced centriole elongation assay. Overall, I present
here a set of proteins with previously uncharacterized roles in centriole duplication,
CEP120/SPICE1 localization, and centriole elongation. These proteins are strong candidate
interactors for CEP120 and SPICE1, and potentially critical for their function. This work
provides a foundation from which further investigation promises to improve our understanding
of how centriole elongation and duplication are achieved, and how CEP120 and SPICE1
participate in these processes.
In this chapter, we identify a novel role for one of two ubiquitously expressed NAP1L proteins,
NAP1L1, in centriole duplication. Human nucleosome assembly protein-like proteins (NAP1Ls)
chaperone histone proteins into and out of nucleosome core particles, with important roles in the
regulation of DNA organization. NAP1L1 was identified as a prey protein for both CEP120 and
SPICE1 in our BioID experiments. Subsequently, several small-interfering RNAs that targeted
NAP1L1 mRNA were found to prevent centriole duplication. Further, NAP1L4, the other
ubiquitously expressed NAP1L protein, was not required for this process. In support of a novel
role for CEP120/SPICE1 within the nucleus, both CEP120 and SPICE1 BioID identified nuclear
proteins including histones and a component of the INO80 chromatin remodeling complex.
Overall, these findings reveal a novel link between CEP120/SPICE1 and the nucleus, and a
requirement for a nucleosome-assembly protein in centriole duplication.

112
3.3. Introduction
3.3.1. BioID
One of the keys to understanding how a protein functions is to identify its interacting partners.
The interaction of CEP120 with SPICE1 was identified by affinity purification and mass
spectrometry, a method widely used to identify proteins either directly interacting or in complex
with a protein of interest (Hutchins et al., 2010). This approach will robustly detect stable
interactions, but can miss transient or unstable interactions. In this chapter of the thesis, I
describe the characterization of potential CEP120 and SPICE1 interacting partners identified by
another method known as BioID (Roux et al., 2012). BirA is an Escherichia coli biotin ligase
which has strong specificity for its substrates, whereas a mutant (BirA-R118G, hereafter BirA*)
has been identified that abolishes the normal interactions of BirA, and releases a biotin-
derivative that rapidly reacts with amines in its proximity, such as those of lysine residues (Choi-
Rhee et al., 2004; Roux et al., 2012). When fused to a ‘bait’ protein, BirA* will cause
biotinylation of the lysine residues of proteins found around the bait protein (Choi-Rhee et al.,
2004; Roux et al., 2012). Adopted to mammalian cells, the BirA* fusion provides an alternative
method to identify both proteins stably interacting with the bait and proteins that transiently
interact with or are in the proximity of the bait briefly (Roux et al., 2012). The use of the BirA*
fusion to detect proximity interactions has been termed the “BioID” method, wherein a bait of
interest is expressed in fusion to BirA* in the presence of exogenous biotin, and Streptavidin
beads are used to affinity purify biotinylated proteins, which are then identified by mass
spectrometry (Roux et al., 2012). One of the major benefits of this approach is that the covalent
biotin linkage is stable under denaturing conditions, so centrosome proteins can be solubilized
before Streptavidin affinity purification (Firat-Karalar et al., 2014). The potential drawbacks of
BioID include the fact that acceptor proteins will be biased or missed based on the number of
accessible lysines, and the BirA* tag (35 kDa) must not interfere with the function or interactions
of the bait protein (Roux et al., 2012). Another drawback is that BioID detects proteins only
based on their proximity, so some identified proteins will have no functional relationship or
direct interaction with the bait (Roux et al., 2012). Despite the drawbacks, BioID has been
successfully applied since its development, including one study that used known centriole

113
assembly proteins as baits to identify novel proteins required for centriole assembly (Chen et al.,
2015; Dingar et al., 2014; Firat-Karalar et al., 2014; Kim et al., 2014; Lambert et al., 2014).
3.3.2. Human Nucleosome Assembly Proteins
One of the CEP120 and SPICE1 associated proteins we study within this chapter is a nucleosome
assembly protein, NAP1L1. Histones are octamers of histone proteins, and within the nucleus
DNA is wrapped around histones to create structures known as nucleosomes (Akey and Luger,
2003; Burgess and Zhang, 2010; Khorasanizadeh, 2004; Kornberg and Lorch, 1999). Within a
histone octamer, there are two copies of each of the histone proteins (H2A, H2B, H3 and H4)
(Akey and Luger, 2003; Kornberg and Lorch, 1999). The assembly of histones onto newly
synthesized DNA, their removal during replication or transcription, and the exchange of variants
of the histone proteins having different properties are all mediated by histone chaperone proteins
(Burgess and Zhang, 2010; Okuwaki et al., 2005, 2010; Park and Luger, 2006a). One class of
chaperones critical to the function of histones in humans is the nucleosome assembly protein
family. The nucleosome assembly protein NAP1 is found in most eukaryotes, and humans have
five NAP1-like nucleosome assembly proteins (NAP1L1-5) (Attia et al., 2011; Park and Luger,
2006a; Zlatanova et al., 2007). Three of these proteins are restricted in their expression to
specific tissues (NAP1L2,3,5), whereas NAP1L1 and NAP1L4 are expressed in all tissues (Attia
et al., 2011; Okuwaki et al., 2010; Rodriguez et al., 1997; Rougeulle and Avner, 1996; Simon et
al., 1994). Purified NAP1L1 and NAP1L4 have been shown to be capable of chaperoning both
nucleosome assembly and disassembly in vitro, with NAP1L1 showing a stronger ability to
disassemble histones than NAP1L4 (Okuwaki et al., 2005, 2010). There is also evidence that
both proteins interact with H2A-H2B dimers in vivo, and that the NAP1 family proteins can
hetero- or homo-dimerize (Attia et al., 2011; Park and Luger, 2006b). Although the majority of
cellular NAP1L1/NAP1L4 are reported to be cytoplasmic, both proteins must be at least partly
nuclear to function in histone protein exchange, and both contain nuclear-export sequences and
nuclear localization sequences (Attia et al., 2011; Okuwaki et al., 2010; Park and Luger, 2006a,
2006b; Rodriguez et al., 1997). NAP1L1, and no doubt NAP1L4, have critical roles in cellular
function as a consequence of their roles in histone structure and transcriptional regulation (Park
and Luger, 2006a; Zlatanova et al., 2007). NAP1L1 levels increase in proliferating cells,
suggesting that it is a positive regulator of growth (Simon et al., 1994). To date, no published

114
evidence has suggested a direct functional link between NAP1L family proteins and the
centrosome.
3.3.3. The Role of tubulin Glutamylation in Microtubule and Centriole
Stability
The function and stability of microtubules can be heavily regulated by the addition or subtraction
of post-translational modifications to and tubulins (Janke and Bulinski, 2011). Within this
chapter of the thesis, I examine changes to the post-translational modifications of microtubules
and centrioles. Centriolar microtubules are subject to extensive post-translational modification
(PTM), including detyrosination, conversion to 2 tubulin, acetylation and glutamylation (Janke
and Bulinski, 2011). These PTMs are typically observed on long-lived microtubules, and though
they do not directly alter microtubule stability, they can serve to regulate it by affecting the
binding of microtubule-associated-proteins (Janke and Bulinski, 2011). Of particular interest for
the centriole is glutamylation, the addition of short or long polymers of glutamic acid residues to
an existing glutamic acid within the CTD of tubulins (Janke and Bulinski, 2011; Janke and
Kneussel, 2010; Magiera and Janke, 2014; Wloga and Gaertig, 2011). This modification is
limited to cellular contexts where MT stability is critical, including neuronal axons, cilia, the
mitotic spindle and the centrioles (Bobinnec et al., 1998b; Janke and Bulinski, 2011; Janke and
Kneussel, 2010; Magiera and Janke, 2014; Song et al., 2013; Wloga and Gaertig, 2011). It has
been suggested that glutamylated tubulin is absolutely critical for centriole stability, as injection
of an antibody that binds glutamylated tubulin (GT335) completely destabilizes the centrioles,
leading to their destruction in late G2/M (Abal et al., 2005; Bobinnec et al., 1998a; Janke and
Bulinski, 2011). This stabilization may not correlate with elongation in all cases, as OFD1
depletion leads to elongation of centrioles without glutamylation (Singla et al., 2010). The
enzymes that catalyze the addition of glutamic acid residues to tubulins are the TTLL (tubulin-
Tyrosine-Ligase-Like) polyglutamylases, of which there are 13 members in mammals (van Dijk
et al., 2007; Janke and Bulinski, 2011; Janke et al., 2005). Within this family of enzymes, there
are specificities for glutamylation of tubulin versus tubulin, and initiation versus elongation
of glutamic acid chains (van Dijk et al., 2007; Janke and Bulinski, 2011). Of the 13 TTLL
enzymes, TTLL1, TTLL5, TTL6, TTL7, TTLL9 and TTL11 localize to basal bodies (centrioles)
in ciliated mammalian kidney cells (van Dijk et al., 2007). In interphase human cells,

115
glutamylation is restricted to centrioles, and how this is achieved is not well understood. In
addition to being regulated in their expression levels, it was hypothesized that concentration at
the centrosome prevents them from activation at other microtubules in the cell (van Dijk et al.,
2007). The precise mechanisms governing their localization to the centrosome, and which
TTLLs are glutamylating the centrioles, are not well understood.

116
3.4. Results
3.4.1. Identification of Potential Functional Interactors of CEP120 and
SPICE1 by BioID
CEP120 and SPICE1 cooperate with CPAP to drive centriole elongation, and we have shown
that CEP120 interacts with CPAP by co-IP (Comartin et al., 2013; Lin et al., 2013b). In order to
look for other proteins that might functionally associate with CEP120 or SPICE1, Sally Cheung
generated HEK293 cell lines with inducible expression of FLAG and BirA* double-tagged
CEP120 or SPICE1 (Figure 3.1). To validate the localization, expression and activities of the
Flag-BirA*-CEP120/SPICE1 proteins, Sally Cheung examined induced and un-induced cells in
the presence or absence of biotin by IF and Western blot (Figure 3.1). Clear evidence of correct
protein localization and active biotinylation was observed for both constructs, so cell pellets were
collected and sent to Dr. Etienne Coyaud for affinity purification of biotinylated proteins using
Streptavidin, and subsequent mass-spectrometry. I analyzed the BioID data sets using ProHits
(see Materials and Methods) and generated a table of high-confidence prey (hits) for each bait
protein (Table 3.1, Table 3.2, Figure 3.2). The table of CEP120 prey here differs slightly from
our published CEP120 associations (Comartin et al., 2013) due to the use of newer controls and
replicates for the current tables. The CEP120 table included here contains more prey proteins,
and is consistent with the more recent SPICE1 BioID table in terms of controls used for analysis.
From the BioID hits for CEP120 and SPICE1, I chose several proteins for further study.
NAP1L1, MAP7D3, CCDC138, CCDC77, CSPP1, KIAA1731, KIAA0753 and ZC2HC1A were
prey found in both CEP120 and SPICE1 BioID experiments (Table 3.1, Table 3.2). CCDC138,
CCDC77, KIAA0753 and ZC2HC1A had no published functions or characterization at the time
of selection, making them potentially exciting novel proteins involved in centriole biology. Of
the selected proteins, only KIAA1731 had a known role in centriole duplication published at the
time of these experiments, but the characterization was limited to demonstrating a requirement
for KIAA1731 for centriole duplication in RPE-1 cells (Knorz et al., 2010). MAP7D3/Mdp3
was known to play a role in microtubule stability, making it an attractive possible cofactor for
CEP120/SPICE1 function (Sun et al., 2011). CSPP1 (centrosome and spindle-pole associated
protein-1) was known to be localized to the centrosome and spindles, and to be important for
mitosis and for cilia formation, but had not been implicated in centriole duplication (Asiedu et
al., 2009; Patzke et al., 2006, 2010). NAP1L1 was among the strongest peptide hits for both

117
CEP120 and SPICE1, and had no previously reported role in centrosome biology, making it a
protein of particular interest. Two proteins only identified in association with CEP120 were also
chosen; ANK2 because of its high-peptide counts, and KIAA0586 because of a reported role in
cilia formation but not centriole duplication (Bangs et al., 2011; Yin et al., 2009). Several
microtubule associated proteins were identified only in the SPICE1-BioID experiments, and
from these MAP7 and MAP9 were included as potentially important for the microtubule related
functions of SPICE1. No critical cellular function for MAP7/Ensconsin had been described,
whereas MAP9/ASAP (ASter-Associated Protein) is a spindle associated MAP required for
proper mitosis (Bulinski et al., 2001; Saffin et al., 2005; Venoux et al., 2007, 2008). Lastly,
microtubule-associated tumour suppressor protein 1 (MTUS1) was also included as a protein
without a known role in centriole duplication, but was known to be centrosome and microtubule
localized with a role in mitosis, and to be down-regulated in multiple human cancers (Rodrigues-
Ferreira et al., 2009; Yu et al., 2009; Zuern et al., 2010).
For the selected proteins, I first determined whether they were required for centriole duplication
by testing them in the S-phase PLK4 induced centriole duplication assay (see section 2.7.4)
(Kleylein-Sohn et al., 2007). From this primary screen, a subset of proteins were identified that
were required for centriole overduplication (Figure 3.3). The proteins were NAP1L1, ANK2,
KIAA1731, MTUS1, CCDC77, CSPP1, KIAA0586, MAP7D3, MAP7 and MAP9 (Figure 3.3).
These findings demonstrate the potential of the BioID approach to identify proteins with
functional relationships to the FLAG-BirA* fused bait proteins used, in this case CEP120 and
SPICE1.
3.4.2. Characterization of CEP120/SPICE1 Associated Proteins
Having identified CEP120 and SPICE1 proximal proteins required for centriole overduplication,
I sought to investigate how these proteins might be functionally related to CEP120 and SPICE1.
First, I depleted the proteins for 72 hours in U-2 OS cells using RNAi. I then immunolabeled the
cells for CEP120 or SPICE1 and glutamylated tubulin to identify centrioles. I included
Centrobin in these experiments, as I was curious whether Centrobin might impact CEP120 and
SPICE1, given that it is a well-characterized centriole duplication protein that acts upstream of
CPAP in centriole duplication and was found within the CEP120 BioID preys (Gudi et al., 2014)
(Table 3.1). Further, as controls I included SASS6, CPAP, CEP120 and SPICE1 for each

118
condition. Two siRNAs targeting NAP1L1 (NAP1L1-si#1, NAP1L1-si#17) were included,
because as described later in this chapter, multiple siRNAS targeting NAP1L1 were found to
affect centriole duplication. Under normal (control) conditions, CEP120 localization is restricted
to the centrosomes. I observed that depletion of several of these proteins caused CEP120
localization to cytoplasmic microtubules (Figure 3.4). Depletion of MTUS1, MAP7, Centrobin,
CSPP1, ANK2, NAP1L1, CEP135 or SASS6 lead to re-localization of CEP120 onto
microtubules (Figure 3.4). The two NAP1L1 siRNAs used in these experiments gave distinct
phenotypes in terms of CEP120 localization (Figure 3.4). Similar to CEP120, SPICE1 is
localized to the centrosome in interphase when expressed at endogenous levels. Depletion of
KIAA1731, MTUS1, ANK2, CSPP1, KIAA00586, MAP7 or MAP7D3 led to abnormal SPICE1
structures in U-2 OS cells, either resembling aggregates outside the centrosomes, or as long
filaments sometimes associated with the centrosomes (Figure 3.5). From these results, I
conclude that multiple proteins found in proximity to CEP120 and SPICE1 can affect their
localizations. Notably, while some proteins are important for both CEP120 and SPICE1
localization (ANK2, NAP1L1, MTUS1, MAP7, and CSPP1), the re-localizations of CEP120 and
SPICE1 following depletion of some of these proteins appears to be distinct. Depletion of
Centrobin, CEP135, and SASS6 uniquely altered CEP120 without detectably affecting SPICE1,
whereas KIAA0586, KIAA1731, and MAP7D3 depletion appear to have effects on SPICE1
localization without major changes to CEP120 (Figure 3.4 and Figure 3.5).
In the process of examining CEP120 and SPICE1 localization following depletion of BioID
identified proximal proteins, I noted two interesting features of the microtubule landscape under
various depletion conditions. Firstly, while under normal conditions, glutamylation is restricted
to centrioles in U-2 OS cells (see controls in Figure 3.4 and Figure 3.5), depletion of several
proteins led to glutamylation of cytoplasmic non-centriolar microtubules. To quantify this, I re-
analyzed the cells from the CEP120 localization experiment and quantified the frequency of
cytoplasmic microtubule glutamylation (Figure 3.6). Depletion of a number of the proteins led
to abnormal glutamylation, including MAP7, MAP7D3, MTUS1, CSPP1, ANK2, NAP1L1,
CEP120, SPICE1, Centrobin and CEP135 (Figure 3.6). Therefore, along with CEP120 and
SPICE1, CEP135, Centrobin and a number of other proteins tested here are important for
restricting glutamylation of microtubules to the centrioles in U-2 OS cells.

119
A second unexpected observation was that depletion of some of the CEP120/SPICE1 associated
proteins led to elongated stretches of CEP120-labeled glutamylated microtubules that appeared
to be elongated centrioles. I quantified the frequency of elongated centrioles within the
experiment and observed that depletion of KIAA0586, CSPP1, ANK2 and MAP7 each resulted
in the formation of elongated structures (Figure 3.7). The elongated structures observed emanate
from paired glutamylated tubulin foci with CEP120 staining, and are therefore most likely to be
elongated centrioles or cilia. To further test the possibility that the proteins depleted in these
studies might be critical for centriole elongation, I tested the impact of depleting these proteins
on taxol-induced centriole elongation in the G2 phase of the cell cycle (see Materials and
Methods, Section 2.7). Two analyses were carried out to examine centriole elongation, the first
was based on glutamylated tubulin structure elongation in the taxol assay (Figure 3.8).
Consistent with my observations in cycling U-2 OS cells (Figure 3.7), depletion of ANK2 and
MAP7 resulted in increased frequency of centriole elongation in the taxol assay above control
conditions (Figure 3.8). Conversely, KIAA0586 and CSPP1 had opposite effects on centriole
length in the taxol assay versus the depletion assay in cycling U-2 OS cells (compare Figure 3.7
and Figure 3.8). All the other proteins depleted in the taxol assay impacted centriole elongation
negatively, with the exceptions being SASS6 and NAP1L1 when si#1 was used. The strongest
effects were observed upon depletion of MAP7D3, CPAP, CEP120, CSPP1, MTUS1 and
NAP1L1 using si#17 (Figure 3.8). For the second analysis, elongation was quantified based on
CEP120 signal, rather than glutamylated tubulin (Figure 3.9). Comparison of the two analyses
yielded three groups of results; glutamylation-specific, CEP120-specific, and results observed
with both markers. MAP7 significantly increased frequency of elongation in both cases, as did
ANK2 though the effect was not quite significant in the case of CEP120-based detection.
SASS6 had no effect on glutamylation, but did have a slight but significant positive effect on
elongation as determined by CEP120 signal. NAP1L1 depletion by siRNA#1 did not
significantly affect elongation in either assay. Several proteins had impacts that were
glutamylation specific: CCDC77, KIAA1731, and CEP135 did not have a significant impact on
CEP120 elongation from centrosomes, but each had a negative impact on centriole elongation
based on examination of glutamylated tubulin structures. Finally, depletion of KIAA0586
promoted centriole elongation in the taxol assay when CEP120 structures were quantified, but
had the opposite effect when glutamylated structures were quantified. This observation was

120
unexpected based on the fact that depletion of KIAA0586 in cycling U-2 OS cells for 72 hours
resulted in apparent elongation of glutamylated tubulin structure resembling centrioles (Figure
3.7). Overall, using the BioID approach we identified CEP120/SPICE1 associated proteins
important for their localizations, centriole elongation, and/or cytoplasmic microtubule regulation
(summarized in Table 3.3).
3.4.3. NAP1L1 is a CEP120 and SPICE1 Associated Protein Required for
Centriole Duplication
One of the proteins identified in the proximity of both CEP120 and SPICE1 using the BioID
approach was the nucleosome assembly protein like protein NAP1L1. In collaboration with
Stine Morthorst, a visiting Ph.D. candidate from the Pedersen lab (University of Copenhagen), I
investigated the potential role of NAP1L1 in centriole duplication. We first investigated whether
NAP1L1 was required for centriole overduplication following PLK4 overexpression, using a set
of four siRNAs from Dharmacon and three siRNAs from LifeTech (Stealth siRNAS). Depletion
of NAP1L1 in the S-phase arrested PLK4 centriole overduplication assay using three of the four
siRNAs from Dharmacon and two of the three siRNAs from LifeTech significantly inhibited
centriole overduplication (Figure 3.10). To validate the role of NAP1L1 in centriole duplication,
I tested the impact of depletion of NAP1L1 using the siRNAS that gave phenotypes in the PLK4
assay on centriole duplication in cycling U-2 OS cells. Depletion of NAP1L1 using Stealth si#1
and Dharmacon siRNA #17 both resulted in cells with less than 4 centrioles in mitosis at 72
hours post-RNAi transfection, whereas the three other siRNAs tested did not significantly impact
centriole duplication under these conditions (Figure 3.11). Initially, NAP1L1 was depleted for
72 hours in HeLa using these two siRNAs, but the impact on centriole duplication was mild.
Therefore, HeLa cells were depleted of NAP1L1 for 96 hours and the frequency of mitotic cells
with fewer than 4 centrioles was quantified (Figure 3.12B). In this approach, both NAP1L1
Stealth si#1 and Dharmacon si#17 effectively inhibited centriole duplication (Figure 3.12). For
this reason, these two siRNAs were used for the assays described earlier in this chapter. Overall,
our observations show that NAP1L1 is important for centriole duplication in cycling cells as well
as in the PLK4 induced centriole overduplication assay.
When siRNA#1 was used to deplete NAP1L1 in the PLK4 induced centriole overduplication
assay, CEP120 was seen on cytoplasmic microtubules specifically encircling the nucleus, and

121
these microtubules were enriched for acetylated tubulin (Figure 3.13). To see if this effect was
specific to the PLK4 overduplication assay, I depleted cycling U-2 OS cells of NAP1L1 for 72
hours by Stealth siRNA#1 and other NAP1L1 siRNAs. NAP1L1 depletion using Stealth-
siRNA#1 caused an increase in CEP120 localization to microtubules in this assay, though
SPICE1 depletion and NAP1L1 depletion with other siRNAs gave minor effects (Figure 3.14B).
To determine whether the re-distribution of CEP120 following NAP1L1 siRNA#1 depletion
resulted from NAP1L1 depletion or an off-target effect, Stine Morthorst generated a NAP1L1-
GFP construct that was resistant to Stealth siRNA#1. I generated a U-2 OS cell line stably
expressing this Stealth siRNA#1 resistant GFP-NAP1L1, and tested whether the presence of
GFP-NAP1L1 could restore the normal localization of CEP120 in U-2 OS cells. I found that
even in cells expressing GFP-NAP1L1 resistant to Stealth siRNA#1, depletion using this siRNA
led to CEP120 mislocalization (Figure 3.15A, B). This indicates that either GFP-NAP1L1 was
not completely functional, was not expressed at levels required for normal cellular functions, or
else that the effect of siRNA#1 on CEP120 localization was due to an off-target effect of that
particular siRNA.
Since NAP1L1 depletion by Dharmacon siRNA#17 also gave strong centriole duplication
defects in all experimental systems tested, and given the failure of GFP-NAP1L1 Stealth-
siRNA# 1 to rescue the mislocalization of CEP120 in U-2 OS, I attempted to rescue the centriole
duplication defects by generating two GFP fusions to NAP1L1 that were resistant to Dharmacon
si#17. In one construct GFP was fused to the amino-terminal end of NAP1L1, whereas the other
construct had GFP fused to the carboxy-terminal end of NAP1L1. I generated stable U-2 OS cell
lines expressing these two constructs, and depleted NAP1L1 for 72 hours in these cell lines using
either Stealth siRNA#1 or Dharmacon siRNA#17 (Figure 3.16). In both cases, the GFP-
NAP1L1 construct was stably expressed and resistant to Dharmacon siRNA#17, but the
expression of these constructs failed to restore centriole duplication to control levels (Figure
3.16).
Given the failure of multiple attempts to rescue the phenotypes associated with NAP1L1
depletion by various siRNAs, I considered the possibilities that the rescue constructs used were
not of the correct isoform of NAP1L1, or that the siRNAs were cross-targeting other NAP1L
protein family members. The NAP1L-family in humans includes NAP1L1, NAP1L2, NAP1L3,

122
NAP1L4 and NAP1L5 (Park and Luger, 2006a). NAP1L2, 3 and 5 are predicted to be expressed
only in neurons, whereas NAP1L1 and NAP1L4 are ubiquitously expressed (Attia et al., 2011;
Park and Luger, 2006a). Since NAP1L4 is ubiquitously expressed I considered whether the
siRNAs used might target NAP1L4 isoforms as well as NAP1L1 isoforms. Using UniProt-
knowledge base (http://www.uniprot.org), I collected the sequences of the two NAP1L1 and two
NAP1L4 isoforms, and used the ClustalW program to align all four sequences
(http://www.ebi.ac.uk/Tools/msa/clustalw2). The two proteins show remarkable similarity
across their sequences, with the only difference between NAP1L1 isoforms 1 and 2 being the C-
terminal region, and the major difference between NAP1L4 and NAP1L1 occurring in the
amino-terminal region where NAP1L4 is lacking two short stretches of conserved amino acids
seen in both NAP1L1 isoforms (Figure 3.17). Like the two NAP1L1 isoforms, the two NAP1L4
isoforms vary from each other only at their CTDs (Figure 3.17). I obtained the targeting
sequences of NAP1L1 siRNAs #1 and #17, and aligned them with the mRNAs for NAP1L1 and
NAP1L4 isoforms, to determine which sequences each siRNA would be predicted to target.
Both siRNAs show 100% alignment with NAP1L1 across both isoforms, whereas each has
limited sequence identity with NAP1L4 (Figure 3.18).
To address the possibility that NAP1L4 and the shorter NAP1L1 isoform were being targeted, I
generated primers for PCR to detect specific sequences unique to each mRNA (see Materials and
Methods). I depleted U-2 OS cells of NAP1L1 using both siRNAs, and in parallel treated
samples with non-targeting siRNAs or a published siRNA specifically targeting NAP1L4
(Okuwaki et al., 2010) for 72 hours. Following depletion, total-RNA was prepared from each
condition using a commercially available kit, RNA concentrations were normalized, and cDNA
was generated from the RNA using random primers. Using equal amounts of the resulting
cDNA as template, parallel PCR reactions were set up to examine relative transcript levels of
NAP1L1 isoforms, NAP1L4, and GAPDH under each depletion condition. I observed that
NAP1L1 siRNA#1 and siRNA#17 resulted in a reduction in the PCR products from both
isoforms of NAP1L1, and the short-isoform of NAP1L1 alone, but did not noticeably decrease
NAP1L4 mRNA abundance (Figure 3.19). NAP1L4 siRNA reduced the abundance of the
NAP1L4 transcripts, consistent with published reports of its specificity (Okuwaki et al., 2010).
NAP1L4 RNAi did, however, slightly reduce the abundance of the PCR product from the short
isoform of NAP1L1 in the assay, and this was unexpected since it targets a conserved sequence

123
in both NAP1L1 isoforms and did not impact the PCR product from both isoforms overall
(Figure 3.19). GAPDH amplification was successful and comparable across all conditions.
Overall, I conclude that the siRNAs used can target the short NAP1L1 isoform resulting in a
decrease in relative abundance of this transcript which may explain the failure of the long-
isoform to rescue the phenotypes of NAP1L1 depletion in both centriole duplication and CEP120
localization. Since both NAP1L1 siRNAs targeted the short and long isoforms to a similar
extent, the unique effect of NAP1L1 siRNA#1 on CEP120 localization is not due to differential
targeting of a NAP1L1 isoform compared to siRNA#17. Importantly, the NAP1L1 siRNAs do
not appear to reduce the abundance of NAP1L4 transcripts.
To directly test the possibility that cross-targeting of NAP1L4 by NAP1L1 siRNAs might
explain the failed rescue of these phenotypes by siRNA resistant GFP-NAP1L1, I examined
whether NAP1L4 depletion affected CEP120 localization or centriole duplication. Using the
same siRNA against NAP1L4 as was used in the PCR experiment, I tested whether NAP1L4
depletion impacted PLK4-induced centriole overduplication or CEP120 localization in the PLK4
assay. I observed that NAP1L4 depletion did not affect CEP120 localization or centriole
overduplication in the S-phase PLK4 induced centriole overduplication assay (Figure 3.20).
Therefore, the phenotypes of NAP1L1 depletion following treatment with siRNA#1 and
siRNA#17 are not due to any cross-targeting of NAP1L4.
Given the failure of rescue experiments to demonstrate conclusively that NAP1L1 RNAi
phenotypes were specific to NAPL1 depletion directly, I obtained an additional, published
siRNA that specifically targeted NAP1L1 but not NAP1L4 (Okuwaki et al., 2010). When
NAP1L1 was depleted using this previously published siRNA reagent (NAP1L1-NEW),
centriole overduplication in the PLK4 assay was significantly inhibited, and although CEP120
was observed on cytoplasmic microtubules, the effect was not as strong as with Stealth siRNA#1
(Figure 3.21). Overall, despite the failure to rescue the effects of NAP1L1 depletion on centriole
overduplication by expression of GFP-NAP1L1, the fact that multiple siRNAs, including a
published siRNA, prevent centriole overduplication in the PLK4 assay strongly supports the
conclusion that one or both NAP1L1 isoforms are critical for centriole duplication (summarized
in Table 3.4).

124
3.5. Discussion
3.5.1. Identification of CEP120 and SPICE1 Associated Proteins Critical for
PLK4 Induced Centriole Duplication
Here, we report the use of the BioID method to identify a panel of candidate interactors for
CEP120 and SPICE1. Several proteins without previously known roles in duplication were
found to be required for PLK4 induced centriole overduplication (Figure 3.3). Those proteins
were NAP1L1, MAP9, CCDC77, CSPP1, MAP7, MAP7D3, KIAA0586, KIAA1731, MTUS1
and ANK2. This work represents a starting point for characterization of these proteins as
centriole duplication proteins. Future work should focus on dissecting how each of these
proteins functions in centriole duplication. For example, where each protein functions in the
centriole assembly pathway, whether these proteins are localized to the centrosomes, and how
they impact CEP120 and SPICE1 recruitment during centriole duplication will be critical
questions to address for each. Additionally, whether or not these proteins are directly interacting
with CEP120 and/or SPICE1 in vivo will be of interest. Overall, the successful identification of
proteins required for centriole duplication by using BioID to identify proteins in the proximity of
CEP120 and SPICE1 demonstrates the potential of this approach to expand our knowledge of
proteins with roles in centriole duplication.
3.5.2. Multiple CEP120 and SPICE1 Associated Proteins Are Required for
CEP120 or SPICE1 localization
The CEP120/SPICE1 associated proteins identified as important for centriole overduplication
following PLK4 overexpression were examined in a variety of assays aimed at identifying
functional relationships with CEP120 and SPICE1. Firstly, each protein was depleted in U-2 OS
cells for 72 hours and CEP120 and SPICE1 localization examined (Figure 3.4 and Figure 3.5).
The observation in this assay that cytoplasmic microtubules were often abnormally glutamylated
led me to quantify the frequency of this phenotype for each condition (Figure 3.6). I also
observed occasional elongation of glutamylated structures from centrioles, which I interpret as
most likely reflecting abnormal elongation of centrioles (Figure 3.7). Strictly speaking, the
possibility that they are cilia cannot be ruled out, so future work should examine these structures
and look for known centriole and cilia proteins, as well as to examine their structure by electron
microscopy as was done for CPAP induced filaments when they were first reported (Kohlmaier

125
et al., 2009; Schmidt et al., 2009; Tang et al., 2009b). For simplicity, I will refer to the structures
as elongated centrioles.
I note that while examining centriole number in mitotic U-2 OS cells was one of the goals of
these experiments, the lack of mitotic cells observed in a many of the conditions made this
impossible. This points to a strong possibility that many of the proteins studied here are critical
for mitosis and/or cell cycle progression, along with centriole duplication. SPICE1 is required
for mitosis and thus these proteins may be functionally linked with SPICE1 in that process
(Archinti et al., 2010). Future work should clearly include examination of the number of
centrioles in synchronized S-phase or G2 cells at 72 hours post RNAi for each condition.
Finally, I utilized taxol induced centriole elongation to test which proteins were required for
elongation, a key function of CEP120 and SPICE1 (Comartin et al., 2013). To determine
whether centrioles were elongated by taxol treatment, I examined both glutamylated tubulin and
CEP120 at the centrioles, and quantified the elongation based on each marker (Figure 3.8 and
Figure 3.9). In some cases, these two approaches yielded conflicting results. In such cases,
glutamylated tubulin is the more stringent/reliable marker for centriole elongation, since CEP120
could form elongated non-centriole structures, though this has not been reported. Alternatively,
it is possible that some proteins affect glutamylation but not centriole elongation, explaining the
difference between the two analyses. CCDC77 is a particularly strong candidate for this, given
the strong impact of its depletion when glutamylated tubulin is examined in the taxol assay,
versus the lack of effect when CEP120 is analyzed (Figure 3.8 and Figure 3.9). It remains
possible that some proteins could promote centriole elongation without glutamylation, in which
case these centrioles would likely be unstable but could persist in the arrested U-2 OS cells (Abal
et al., 2005; Bobinnec et al., 1998a).
The depletion of MTUS1 MAP7, ANK2, NAP1L1, and the well characterized centriole
duplication proteins Centrobin, CEP135 and SASS6 all led to abnormal localization of CEP120
on cytoplasmic microtubules in U-2 OS cells (Figure 3.4). Why does this occur? CEP135 and
SASS6 are required for CEP120 localization in centriole assembly (Comartin et al., 2013). This
suggests the simple possibility that CEP120 localization to microtubules is the result of a loss of
its normal centrosomal binding sites, leading to a lower-affinity interaction with cytoplasmic
microtubules. The fact that CPAP and SPICE1 depletion do not lead to this phenotype argues

126
against this interpretation. Also, depletion of CEP135 or SASS6 has only a minor effect on
CEP120 localization, whereas MTUS1, MAP7, CSPP1, Centrobin, NAP1L1 and especially
ANK2 have more dramatic effects (Figure 3.4). This suggests that these other proteins have an
additional or alternative role in CEP120 localization. Below I will briefly summarize each
protein’s specific effects in our assays, and describe which proteins should be prioritized for
future study.
MTUS1 is an appealing protein for future study as it is a microtubule associated protein that has
not been well characterized functionally, and is clearly down-regulated in cancers (Rodrigues-
Ferreira et al., 2009; Yu et al., 2009; Zuern et al., 2010). In our experiments, MTUS1 depletion
led to significant glutamylation of the microtubule cytoskeleton, mislocalization of CEP120 and
SPICE1, and importantly prevented taxol induced centriole elongation (Figure 3.4, Figure 3.5,
Figure 3.8 and Figure 3.9). From these results, I suggest that MTUS1 is a strong candidate for a
novel protein required for centriole elongation, and possibly a co-factor for CEP120 and SPICE1
in this role. As such, it will be of particular importance to look for a direct interaction between
CEP120 or SPICE1 and MTUS1, as well as to examine in greater depth the role of MTUS1 in
centriole duplication. The impact of MTUS1 depletion on centriole elongation following
CEP120 or CPAP overexpression should also be tested. Whether MTUS1 overexpression alone
can drive centriole elongation in S-phase or G2 arrested cells is worth investigating.
MAP7D3 depletion affected SPICE1 localization, but did not appear to strongly affect CEP120
localization in U-2 OS cells (Figure 3.4 and Figure 3.5). Importantly, in our taxol assay,
MAP7D3 depletion yielded a stronger phenotype than CPAP depletion, strongly suggesting a
role in centriole elongation (Figure 3.8 and Figure 3.9). As mentioned, MAP7D3/Mdp3 has been
shown to stabilize microtubules in vitro, and it is also important for microtubule regrowth in vivo
(Sun et al., 2011). Notably, MAP7D3 contains coiled-coil domains that are a hallmark of
centrosomal proteins, and these domains are indispensable for its interaction with microtubules
in vivo (Sun et al., 2011). As a microtubule stabilizing protein that gave very strong phenotypes
in our centriole elongation assays, MAP7D3 should be prioritized for future study, starting with
determining whether it interacts with CEP120 or SPICE1, and is detectable on centrioles. The
same experiments proposed to test MTUS1 in centriole elongation should also be considered for

127
MAP7D3, as one or both proteins could be important for CEP120 and SPICE1 mediated
centriole elongation.
CSPP1 is critical for proper mitotic spindle formation, cilia formation, and is associated with
mitotic spindle poles (Akizu et al., 2014; Asiedu et al., 2009; Shaheen et al., 2014a). CSPP1
depletion mislocalizes both CEP120 and SPICE1 in U-2 OS cells, and strongly drives
glutamylation of interphase microtubules in our assays (Figure 3.4, Figure 3.5 and Figure 3.6).
CSPP1 depletion leads to abnormal elongation of centriole-like structures in U-2 OS cells,
whereas in the taxol induced centriole elongation assay CSPP1 depletion reduces the frequency
of centriole elongation (Figure 3.7, Figure 3.8 and Figure 3.9). In spite of the seemingly
contradictory phenotypes, we can strongly infer that CSPP1 is important for centriole length
regulation and affects CEP120/SPICE1. The requirement for CSPP1 in centriole duplication and
possibly elongation would be consistent with the loss of cilia formation observed upon depletion
of CSPP1, and may be relevant to its role in Joubert Syndrome (a ciliopathy) (Akizu et al.,
2014). As such, it will be very important to determine how CSPP1 participates in centriole
assembly, and whether it is a legitimate interacting partner of CEP120 and SPICE1.
ANK2 and MAP7 were the only two proteins tested that when depleted gave abnormal centriole
elongation and cytoplasmic MT glutamylation in U-2 OS cycling cells, as well as in the taxol
assay when glutamylated tubulin was used as a marker for centriole length (Figure 3.6, Figure
3.7 and Figure 3.8). Both proteins are also important for correct localization of CEP120 and
SPICE1 (Figure 3.4 and Figure 3.5). In Drosophila, it has been recently discovered that MAP7
is important for centrosome separation and microtubule elongation within the spindle (Gallaud et
al., 2014). It is surprising that our results are consistent with an opposite interpretation; that
MAP7 depletion drives centriole elongation in multiple assays. ANK2 (ankryin-repeat
containing protein-2 or ANK-B) depletion also gave a strong phenotype in the centriole
overduplication assay. ANK2 is important for localization of membrane associated ion channels
in cardiac muscle cells (Cunha et al., 2011; Mohler et al., 2003). Mutations in ANK2
accordingly have been shown to be causal in cardiac arrhythmias (Cunha et al., 2011; Mohler et
al., 2003). Perhaps more relevant, ANK2 interacts with DCTN4, a subunit of the dynactin
complex and plays an important role in DCTN4 mediated transport along axons, as well as being
important for axon length (Ayalon et al., 2008; Lorenzo et al., 2014). It was reported, however,

128
that microtubules were not affected in ANK2-null mouse neurons, so the regulation of axon
length is unlikely to be through microtubule stability (Lorenzo et al., 2014). The dynein-
dynactin complex is a microtubule motor (dynein) coupled with a cargo adaptor (dynactin) that
together play important roles in cellular structure (Kardon and Vale, 2009; Vallee et al., 2004).
Dynein has been implicated in transport of centriolar satellites to the centrosome, and both
CEP120 and SPICE1 had the satellite proteins PCM1 and CEP131 within their BioID preys
(Dammermann, 2002; Kubo et al., 1999; Prosser et al., 2009; Staples et al., 2012). One
possibility is that ANK2 may be linking CEP120/SPICE1 and/or satellites to dynactin for
transport to or from centrosomes. Future work should certainly test this hypothesis. This could
be achieved by live-cell imaging of GFP-PCM1, GFP-SPICE1 or GFP-CEP120 under ANK2
depleted conditions. Overall, these findings identify novel roles for MAP7 and ANK2 in
centriole duplication, and possibly as being important for the restriction of centriole length.
MAP9/ASAP has a critical role in mitotic spindle formation and is localized to the centrosome
and spindles (Bulinski et al., 2001; Eot-Houllier et al., 2010; Saffin et al., 2005; Venoux et al.,
2007, 2008). In one study, the authors report that the spindle assembly defects upon MAP9
depletion were not due to centriole duplication defects (Eot-Houllier et al., 2010). Our
observation of a requirement for MAP9 in centriole overduplication could be a result unique to
the PLK4 assay, or to the siRNA used in our study. Notably, SPICE1 induces spindle defects
prior to the loss of centrioles due to failed duplication, and so the MAP9 phenotypes that are
published could be separable from a role in duplication (Archinti et al., 2010). Further work will
be required to validate the proposed role of MAP9 in centriole assembly. Specifically, it will be
critical to demonstrate a failure of centriole duplication in cycling human cells when MAP9 is
depleted, and to reverse that phenotype with an siRNA resistant MAP9 construct. This may be
challenging as depletion or overexpression of MAP9 can cause mitotic defects (Saffin et al.,
2005).
KIAA0586 and KIAA1731 are both required for centriole overduplication in the PLK4 assay.
Subsequent to my experiments with KIAA0586/Talpid3, it was published to be a CEP120
interacting protein and to affect CEP120 localization to the mother centrioles (Wu et al., 2014).
Additionally, KIAA0586 has been found to interact with CP110, and is involved in multiple
ciliopathies (Alby et al., 2015; Kobayashi et al., 2014; Roosing et al., 2015). We observed a

129
larger effect of KIAA0586 depletion on the localization of SPICE1, and found that depletion of
this protein led to elongated centrioles in U-2 OS cells (Figure 3.4, Figure 3.5 and Figure 3.7).
Within the taxol assay, KIAA0586 caused a slight increase in the formation of CEP120
filaments, but also slightly inhibited the elongation of glutamylated tubulin (Figure 3.8 and
Figure 3.9). KIAA1731/CEP295 was initially described as being required for centriole
duplication, however it has recently been demonstrated that KIAA1731/CEP295 is dispensable
for centriole assembly, but critical for daughter centrioles to become competent for PCM
recruitment and accordingly their stability following loss of their cartwheels (Izquierdo et al.,
2014; Knorz et al., 2010). In the PLK4 assay, the loss of parental centrioles due to loss of
stability following CEP295 depletion could explain why overduplication was inhibited. In our
assays, KIAA1731 also had a slight effect on SPICE1 localization and taxol induced centriole
elongation (Figure 3.5 and Figure 3.8). The possibility of functional cooperation between
KIAA1731/CEP295 or KIAA0586 and CEP120 or SPICE1 warrant further investigation.
CCDC77 depletion did not have a significant impact on CEP120 or SPICE1 localization,
glutamylation of interphase microtubules, or lead to abnormal elongation of centrioles when
depleted in U-2 OS cells (Figure 3.4, Figure 3.5, Figure 3.6 and Figure 3.7). CCDC77 depletion
did moderately inhibit taxol induced centriole elongation based on glutamylated tubulin signal
(Figure 3.8). Additionally, having no published characterization, CCDC77 represents an
attractive protein for study. Understanding the role of this coiled-coil protein in centriole
duplication and potentially elongation, as well as whether it cooperates with CEP120 and
SPICE1, would be of great interest.
Overall, the diversity of proteins identified here and the effects that result from depleting them
point to the possibility of a number of different pathways impacting centriole duplication and
CEP120/SPICE1 localization. These proteins represent a rich data set for future projects that will
yield novel insights into CEP120 and SPICE1 function, and centriole duplication.
3.5.3. Depletion of CEP120 and SPICE1 Associated Proteins Affects the
Microtubule Cytoskeleton
In the experiments described here, there is a correlation between CEP120 re-localization and
abnormal glutamylation of cytoplasmic microtubules (Figure 3.4 and Figure 3.6). ANK2,

130
CSPP1, MTUS1, MAP7, Centrobin and possibly NAP1L1 (*siRNA#1) depletion caused both
phenotypes concurrently in U-2 OS cells. Could recruiting TTLLs to drive glutamylation of
centrioles be a function of CEP120? This possibility should clearly be investigated, given the
paucity of information on how TTLLs are regulated with regard to their localization to the
centrosome. It is known that when expressed above normal levels in HeLa cells, several TTLLs
will bind to and glutamylate cytoplasmic microtubules, so simple release of TTLLs from
centrosomes might explain this effect (van Dijk et al., 2007). Could the loss of centrioles
following CEP120/SPICE1 and other proteins lead to a loss of TTLL restriction to centrioles?
The fact that CPAP and SASS6 depletion in these assays did not have this effect as strongly as
CEP120 or SPICE1 depletion, or the depletion of other proteins, argues against this (Figure 3.6).
CPAP and SASS6 gave the strongest inhibition of duplication in the PLK4 assay, so they would
have caused the strongest glutamylation of cytoplasmic microtubules were loss of centrioles the
driving factor (Figure 3.3). If CEP120 is important for targeting TTLLs to regulate microtubule
glutamylation at centrioles, mislocalized CEP120 on cytoplasmic microtubules could drive
aberrant glutamylation. In this case, the minor but significant glutamylation of cytoplasmic
microtubules following CEP120/SPICE1 depletion would be explained by loss of centrosome-
restriction of TTLL localization. This should be tested by examining whether double-depletion
of CEP120 along with ANK2 or CSPP1 reverses the glutamylation of cytoplasmic microtubules.
An alternative explanation for these results would be that depletion of one or more of the
proteins tested leads directly to glutamylation of microtubules, and CEP120 has a strong affinity
for glutamylated microtubules. The binding of recombinant CEP120 to glutamylated versus non-
glutamylated purified microtubules in vitro should be tested, to see if CEP120 has a higher
affinity for glutamylated tubulins. If TTLL overexpression recruits CEP120 to glutamylated
cytoplasmic microtubules in vivo, this will further support a model where CEP120 preferentially
binds glutamylated microtubules, or TTLL enzymes themselves. Lastly, it is possible that the
mislocalization of CEP120 onto cytoplasmic microtubules stabilizes them, and microtubules that
are long-lived then accumulate post-translational modifications including glutamylation (Janke
and Bulinski, 2011). The stability of microtubules under CEP120 over-expression, and depletion
of ANK2 or CSPP1, should be examined. Testing the resistance of microtubules to cold-shock
induced depolymerization in vivo under these conditions would be a valid approach. It will also

131
be important to look for other microtubule PTMs such as acetylation, to determine whether
glutamylation is being affected specifically.
Depletion of proteins such as ANK2 and CSPP1 lead to accumulation of CEP120 on cytoplasmic
microtubules that were glutamylated, and failed centriole overduplication. It is possible that with
CEP120, other centriole assembly proteins might be localized to cytoplasmic microtubules under
these conditions. Future work should include analysis of the localization of CPAP, Centrobin,
SASS6, hPOC5, Poc1 and CEP135 under such depletion conditions. This could reveal proteins
cooperating with CEP120 in microtubule stabilization or glutamylation. Lastly, CEP120 is
recruited without SPICE1 to cytoplasmic microtubules under ANK2/CSPP1 depletion, indicating
that SPICE1 is dispensable for the glutamylation observed in this context.
In sum, the results presented here reveal a potential link between CEP120, several of the proteins
examined in this study, and the glutamylation of interphase microtubules. It will be of great
interest to understand this link as it may provide insight into the glutamylation of centrioles, as
well as novel functions for the proteins herein described.
3.5.4. NAP1L1 is Required for Centriole Duplication
NAP1L1 has an unreported role in centriole duplication. We have shown here that NAP1L1
depletion by multiple siRNAs, including a published siRNA shown to target NAP1L1, leads to
failed centriole duplication in cycling cells and in the PLK4 assay (Figure 3.21). The difference
in phenotypes between NAP1L1 siRNAs is unexplained, as is the failure of rescue experiments
aimed at validating their effects as being due to NAP1L1 depletion (Figure 3.15 and Figure
3.16). It is possible that the GFP-tag itself might interfere with folding or function of NAP1L1,
and while we mitigated this possibility by using GFP tags on either end of the protein in separate
rescue experiments, we cannot eliminate this possibility. The other possibility is that the
NAP1L1 minor isoform may be critical for centriole duplication, remains open based on the
results of my quantitative PCR experiment showing that the minor isoform is reduced in its
expression by treatment with siRNAs#1 and siRNA#17 against NAP1L1 (Figure 3.19). To
address these possible explanations, future rescue experiments should include use of both
NAP1L1 isoforms, with smaller epitope tags, under more carefully regulated expression.
Insertion of an siRNA resistant NAP1L1 protein at the endogenous locus under its native

132
promoter using CRISPR based genome editing would be an excellent approach (Doudna and
Charpentier, 2014; Gilbert et al., 2014).
NAP1L1 and NAP1L4 are functionally related and may cooperate in their histone-chaperone
activities (Attia et al., 2011; Okuwaki et al., 2010). In our hands, NAP1L4 depletion using a
published siRNA did not affect centriole duplication (Figure 3.20). It will be important to test
whether NAP1L1 interacts with CEP120 or SPICE1 by co-IP, and this should be tested for both
isoforms of the protein. NAP1L1 and NAP1L4 are members of a select group of proteins other
than tubulin that can be glutamylated on their C-termini in vivo (van Dijk et al., 2008; Regnard,
2000). Could this glutamylation mimic a glutamylated microtubule, and facilitate CEP120
binding to regulate CEP120 localization to centrosomes? This is speculative, but since the
original identification of NAP1L1-glutamylation was due to pull-down by an antibody that
detects glutamylated tubulin, such a mechanism is not entirely impossible (Regnard, 2000; Wolff
et al., 1992). The glutamylation of NAP1L1 in Xenopus is essential to its function in mitotic
chromosome organization through histone exchange (Miller and Heald, 2015). NAP1L1
glutamylation sites conserved in the human protein are found within a region of the NTD that is
absent in NAP1L4, and a region of the CTD that is partly truncated in the short-isoform of
NAP1L1 (Miller and Heald, 2015). Therefore it is possible that the NTD glutamylation sites are
relevant to centriole duplication, since NAP1L4 differs only in that region and does not function
in centriole duplication. Testing the role of NAP1L1 glutamylation in centriole duplication
would be worthwhile, especially glutamylation of the NTD region of the protein. Once a rescue
experiment is successful for NAP1L1 depletion, it could be repeated with a NAP1L1 construct
where target glutamic acid residues have been substituted with aspartic acid residues (Miller and
Heald, 2015). This would answer the question of whether NAP1L1 glutamylation in the NTD is
required for centriole duplication. Likewise, if NAP1L1 is found to interact with CEP120 or
SPICE1, whether NAP1L1 glutamylation affected that interaction would be an important
question.
Depletion of NAP1L1 using multiple siRNAs caused CEP120 localization to cytoplasmic
microtubules (Figure 3.14), and preliminary evidence suggests NAP1L1 depletion reduced
CEP120 levels at centrosomes (see Figure 3.10 and Figure 3.11A). It will be important to
quantitatively analyze CEP120 and SPICE1 levels at centrosomes following NAP1L1 depletion,

133
once an siRNA has been successfully validated by rescue experiments. The levels of
CEP120/SPICE1 should also be measured by Western-blot, to determine whether NAP1L1 is
regulating their expression or stability.
The detection of NAP1L1 raises the possibility that a population of CEP120 and SPICE1 might
localize within the nucleus, where they could be associated with NAP1L1 and potentially link
nuclear processes to centriole duplication. The SPICE1 BioID preys included multiple Importin
proteins (IPO4, 5, 7, 8), histone variants (H2AFY, HIST3H2BB), DNA polymerase Eta (POLH)
and Topoisomerases 2A and 2B (Table 3.2 and Figure 3.2). IPO4, 5, 7 and 8 are involved in
transport of histone proteins through the nuclear pore complex, and one of the proposed
functions of NAP1 proteins is the transport of histone variants from the cytoplasm to the nucleus
(Cautain et al., 2015; Rodriguez et al., 1997; Zlatanova et al., 2007). Whether SPICE1 binds
importins, histones or NAP1L1 directly should be investigated. Whether SPICE1 is required for
NAP1L1 or histone import into the nucleus, or vice versa, are also important questions. A
truncated SPICE1 protein (residues 1-450) localizes to the nucleus, in support of this idea
(Archinti et al., 2010). The strongest peptide count among the CEP120 BioID preys was subunit
2 of the INO80 chromatin remodeling complex (INO80B), a complex involved in transcription
regulation, DNA repair and DNA replication (Table 3.1) (Conaway and Conaway, 2009;
Morrison and Shen, 2009). Several nuclear proteins were prey for both CEP120 and SPICE1 in
BioID experiments; NAP1L4, p53 (TP53), p53 binding partner 2 (TP53BP2), and basic
transcription factor 3 (BTF3). NAP1L1 binds multiple transcription factors, including p53, to
help facilitate transcription (Rehtanz et al., 2004). The association of INO80B, p53, NAP1L1
and NAP1L4 suggest that CEP120 and SPICE1 could play as yet unknown roles in
transcriptional regulation within the nucleus. Alternatively, p53 and P53BP2 (ASPP2) are both
associated with centrosomes in mitosis, which could explain their proximity to CEP120 and
SPICE1 (Ciciarello, 2001; Tritarelli et al., 2004; Zhang et al., 2015). Components of the NuRD
chromatin remodeling complex localize to the centrosome, and the CHD3 subunit specifically is
required for Pericentrin recruitment to centrosomes (Sillibourne et al., 2007; Torchy et al., 2015).
Centrin-2 and Pericentrin are also shuttled between the nucleus and the cytoplasm (Keryer et al.,
2003; Prosser et al., 2009). Thus there is precedent for centrosome proteins with nuclear
localizations, and chromatin remodeling complexes affecting centrosome function. Whether
NAP1L1 localizes to the centrosome should be examined carefully, although I failed to detect

134
GFP-NAP1L1 at centrosomes by IF in U-2 OS cells (see Figure 3.15A, right-panel). Further, the
possibility that CEP120 and SPICE1 shuttle into and out of the nucleus should be examined by
IF following inhibition of nuclear export with Leptomycin-B (Keryer et al., 2003; Kudo et al.,
1999; Prosser et al., 2009). A role for CEP120 or SPICE1 in chromatin remodeling, histone
chaperoning or transcriptional regulation in addition to centriole duplication would be a very
interesting discovery.
Overall, we identify NAP1L1 as a protein in the proximity of CEP120 and SPICE1 in vivo.
Depletion of NAP1L1 using multiple siRNAs including previously validated siRNAs inhibits
centriole duplication in multiple assays and cell types. The exact impact of NAP1L1 depletion
on CEP120 localization remains obscured by differences in siRNA effects and failure to rescue
them thus far. Clearly, however, there is an unreported role for NAP1L1 in centriole duplication.
This exciting discovery, and the possible interaction with CEP120 and/or SPICE1, warrant
further investigation.

135
3.6. Figures
Figure 3.1

136
Figure 3.1. Validation of FLAG-BirA*-CEP120 and FLAG-BirA*-SPICE1 biotinylation
activity and localization.
(A) IF images of HEK293 cells that have inducible expression of FLAG-BirA*-CEP120 (left) or
FLAG-BirA*-SPICE1 (right), labeled with antibodies against FLAG (green), PCTN (blue), and
biotin (Streptavidin, red). Cells were incubated for 24 hours in 50 M biotin and tetracycline (1
g/ml) to induce expression (top rows), or tetracycline alone (middle), or neither (bottom). Scale
bars 10 m. (B) Western blots show expression of a FLAG-BirA* peptide and FLAG-BirA*-
CEP120 (left panel), and FLAG-BirA*-SPICE1 (right panel). Bottom panels of each Western
blot show detection of biotinylation of cellular proteins by Streptavidin (SA-HRP) under each
condition. Sally Cheung provided the data shown in this figure, and carried out the experiments
described herein.

137
Table 3.1 CEP120 Associated Proteins Detected by BioID.
Prey Gene Counts Control Counts AvgP MaxP SAINT
INO80B 279|173 0|0|0|0|0|0|0|0|0|0 1 1 1
NAP1L1 194|201 25|24|10|12|12|10|34|36|37|40 1 1 1
PCM1 198|180 4|3|0|0|0|0|6|5|2|3 1 1 1
ANK2 247|99 0|0|0|0|0|0|1|0|0|0 1 1 1
XRN1 196|109 13|6|0|7|7|0|8|7|4|0 1 1 1
AZI1 121|59 0|0|0|0|0|0|0|0|0|0 1 1 1
ZC2HC1A 113|54 0|0|0|0|0|0|0|0|0|0 1 1 1
CENPJ 88|65 0|0|0|0|0|0|0|0|0|0 1 1 1
NAP1L4 85|65 9|8|0|12|12|4|14|13|15|14 1 1 1
OFD1 89|58 0|0|0|0|0|0|0|0|0|0 1 1 1
CEP170 87|58 5|8|0|0|0|0|12|18|13|9 1 1 1
SPICE1 62|55 0|0|0|0|0|0|0|0|0|0 1 1 1
CEP95 60|39 0|0|0|0|0|0|0|0|0|0 1 1 1
DVL3 49|48 18|11|8|5|2|8|13|15|0|4 1 1 1
MAP7D3 71|22 1|0|0|3|2|0|4|4|2|3 1 1 1
CCDC138 61|28 0|0|0|0|0|0|0|0|0|0 1 1 1
DVL2 59|29 4|0|0|0|0|8|3|5|2|2 1 1 1
HAUS6 51|36 1|2|0|0|0|0|0|0|1|6 1 1 1
KIAA0586 53|29 0|0|0|0|0|0|0|0|0|0 1 1 1
LRRC49 41|32 0|0|0|0|0|2|9|7|0|0 1 1 1
CSPP1 39|27 0|0|0|0|0|0|0|0|0|0 1 1 1
CEP192 37|28 0|0|0|0|0|0|0|0|0|0 1 1 1
KIAA1671 45|20 0|0|0|0|0|0|0|0|0|0 1 1 1
CEP350 24|41 0|0|0|0|0|0|0|0|0|0 1 1 1
LUZP1 26|33 1|2|0|3|3|0|2|2|0|0 1 1 1
TJP1 27|19 5|2|0|0|0|0|5|3|5|6 1 1 1
GPATCH1 26|20 0|0|4|0|0|3|1|3|3|2 1 1 1
PIBF1 30|14 0|0|0|0|0|0|0|0|0|0 1 1 1
PDE3B 13|27 0|0|0|0|0|0|0|0|0|0 1 1 1
HAUS3 27|12 0|0|0|0|0|0|0|0|0|0 1 1 1
KIAA0753 21|18 0|0|0|0|0|0|0|0|0|0 1 1 1
CCDC66 36|2 0|0|0|0|0|0|0|0|0|0 0.98 1 0.98
CEP97 18|19 0|0|0|0|0|0|1|1|0|0 1 1 1
ALMS1 21|13 0|0|0|0|0|0|0|0|0|0 1 1 1
TP53 17|17 3|4|0|0|0|0|0|0|0|0 1 1 1
HAUS5 18|15 0|0|0|0|0|0|1|1|0|0 1 1 1
SSX2IP 16|16 0|0|0|0|0|0|0|0|0|0 1 1 1
MPHOSPH9 17|11 0|0|0|0|0|0|0|0|0|0 1 1 1
HAUS8 19|8 0|0|0|0|0|0|0|0|0|0 1 1 1
CKAP2 15|12 0|1|0|0|0|0|0|0|0|0 1 1 1
KIAA1009 11|16 0|0|0|0|0|0|0|0|0|0 1 1 1

138
Table 3.1 Continued
Prey Gene Counts Control Counts AvgP MaxP SAINT
CCP110 15|12 0|0|0|0|0|0|0|0|0|0 1 1 1
CEP72 14|12 0|0|0|0|0|0|0|0|0|0 1 1 1
CAMSAP1 13|12 0|0|0|0|2|0|0|0|0|0 1 1 1
WDR67 17|8 0|0|0|0|0|0|0|0|0|0 1 1 1
ARHGAP21 13|12 0|0|0|0|0|0|0|0|0|0 1 1 1
CASC5 9|16 0|0|0|0|3|0|0|0|0|3 0.97 1 0.97
TP53BP2 13|11 0|0|0|0|0|0|1|1|0|0 1 1 1
C2orf44 14|10 0|0|0|0|0|0|0|0|0|0 1 1 1
CKB 4|20 1|0|0|0|0|0|0|0|1|1 0.92 1 0.92
CEP55 10|11 0|0|0|0|0|0|2|2|0|0 1 1 1
WRAP73 11|9 0|0|0|0|0|0|0|0|0|0 1 1 1
TTK 13|6 0|0|0|0|0|0|2|1|0|0 0.99 1 0.99
FGFR1OP 7|10 0|0|0|0|0|0|0|0|0|0 1 1 1
EXOC4 11|6 0|1|0|0|0|0|0|0|0|0 1 1 1
EIF2C2 11|5 0|0|0|0|0|0|0|1|0|0 1 1 1
YEATS2 11|5 0|0|0|0|1|2|0|0|0|0 0.98 1 0.98
CCDC77 9|6 0|0|0|0|0|0|0|0|0|0 1 1 1
UCHL1 10|5 0|0|0|0|0|0|0|0|0|0 1 1 1
UBE2I 9|5 0|0|0|0|0|0|0|0|0|1 1 1 1
CNTROB 6|8 0|0|0|0|0|0|0|0|0|0 1 1 1
EIF4E 8|6 0|0|0|1|0|0|2|0|0|0 0.99 1 0.99
NACAD 5|8 0|0|0|0|0|0|0|0|0|0 1 1 1
HAUS2 7|6 0|0|0|0|0|0|0|0|0|0 1 1 1
TNRC6B 9|4 0|0|0|0|0|0|0|0|0|1 0.99 1 0.99
RPAP2 8|4 0|0|0|0|0|0|0|0|0|0 1 1 1
KIAA1731 6|5 0|0|0|0|0|0|0|0|0|0 1 1 1
IBTK 5|6 1|1|0|0|0|0|0|0|0|0 0.99 1 0.99
AP4B1 2|9 0|0|0|0|0|0|0|0|0|0 0.98 1 0.98
SDCCAG3 4|6 0|0|0|0|0|0|0|0|0|0 1 1 1
SLC9A2 4|6 0|0|0|0|0|0|0|0|0|0 1 1 1
IL20RB 8|2 0|0|0|0|0|0|0|0|0|0 0.98 1 0.98
BTF3 6|4 0|0|0|0|0|0|1|0|0|1 0.97 1 0.97
KIF2A 4|6 1|0|0|0|0|0|0|0|0|1 0.97 1 0.97
IPMK 6|3 0|0|0|0|0|0|0|0|0|0 1 1 1
CEP152 4|4 0|0|0|0|0|0|0|0|0|0 1 1 1
HAUS1 4|4 0|0|0|0|0|0|0|0|0|0 1 1 1
SPATA2 6|2 0|0|0|0|0|0|0|0|0|0 0.98 1 0.98
KIF14 6|2 0|0|0|0|0|0|0|0|0|0 0.98 1 0.98
CDC27 6|2 0|0|0|0|0|0|0|0|0|0 0.98 1 0.98
TTC28 4|3 0|0|0|0|0|0|0|0|0|0 1 1 1
EPPK1 4|3 0|0|0|0|0|0|0|0|0|0 1 1 1

139
Table 3.1: CEP120 Associated Proteins Detected by BioID.
Results from two biological replicates of FLAG-BirA*-CEP120 expression in the presence of
biotin followed by Streptavidin affinity-purification and mass-spectrometry to identify
biotinylated peptides. Control counts are from 10 runs from four biological FLAG-BirA*
expression experiments prepared under the same conditions. (From left to right) Gene Symbol
for identified proteins, the spectral counts for each protein in each of the experimental replicates,
the spectral counts for each protein in the control replicates, the average probability of both
replicates (AvgP), the maximum probability between the two replicates (MaxP), and the overall
SAINT score for both replicates are shown (Choi et al., 2011). Spectra reflect the number of
spectral counts assigned to a prey polypeptide in each MS analysis. Samples were prepared by
Sally Cheung, and BioID-MS and was carried out by Dr. Etienne Coyaud.

140
Table 3.2 SPICE1 Associated Proteins Detected by BioID.
Prey Counts Control Counts AvgP MaxP SaintScore
PCM1 172|208 4|3|0|0|0|0|6|5|2|3 1 1 1
NAP1L1 141|215 25|24|10|12|12|10|34|36|37|40 1 1 1
TOP2A 215|139 0|0|5|54|49|6|0|0|0|0 0.99 1 0.99
MAP7D3 115|142 1|0|0|3|2|0|4|4|2|3 1 1 1
IPO7 89|94 6|7|2|0|0|4|8|9|4|4 1 1 1
CEP120 75|95 0|0|0|0|0|0|0|0|0|0 1 1 1
AZI1 75|93 0|0|0|0|0|0|0|0|0|0 1 1 1
IPO5 72|93 0|0|0|0|0|0|4|4|3|2 1 1 1
KIAA1731 67|97 0|0|0|0|0|0|0|0|0|0 1 1 1
OFD1 65|97 0|0|0|0|0|0|0|0|0|0 1 1 1
CKAP2 72|79 0|1|0|0|0|0|0|0|0|0 1 1 1
TOP2B 95|55 0|0|4|0|0|5|0|0|0|0 1 1 1
LUZP1 63|86 1|2|0|3|3|0|2|2|0|0 1 1 1
LRRC49 69|75 0|0|0|0|0|2|9|7|0|0 1 1 1
NAP1L4 58|71 9|8|0|12|12|4|14|13|15|14 1 1 1
CCDC138 60|61 0|0|0|0|0|0|0|0|0|0 1 1 1
TPGS1 51|53 0|0|6|0|0|5|16|11|0|0 1 1 1
MTUS1 43|54 0|0|0|0|0|0|0|0|0|0 1 1 1
CEP135 41|55 0|0|0|0|0|0|0|0|0|0 1 1 1
KIAA0753 39|54 0|0|0|0|0|0|0|0|0|0 1 1 1
TRIM36 45|45 0|0|0|0|0|0|0|0|0|0 1 1 1
HAUS6 35|52 1|2|0|0|0|0|0|0|1|6 1 1 1
SKA3 35|50 0|1|0|5|0|0|4|2|0|1 1 1 1
SUPT16H 46|36 1|0|1|0|0|0|0|0|0|0 1 1 1
PLEKHA5 30|49 0|0|0|0|0|0|0|0|0|0 1 1 1
WDR67 36|42 0|0|0|0|0|0|0|0|0|0 1 1 1
H2AFY 38|33 15|13|5|3|2|5|0|4|0|11 0.94 1 0.94
KIF7 22|44 0|0|0|0|0|0|0|0|0|0 1 1 1
CAMSAP1 25|37 0|0|0|0|2|0|0|0|0|0 1 1 1
RBMX 34|28 0|0|0|0|0|0|0|0|10|12 0.95 0.97 0.95
SSX2IP 26|35 0|0|0|0|0|0|0|0|0|0 1 1 1
HAUS3 23|37 0|0|0|0|0|0|0|0|0|0 1 1 1
SSRP1 30|28 0|0|0|5|0|1|1|0|0|0 1 1 1
SKA2 24|31 0|0|0|6|4|0|1|1|0|0 1 1 1
NOP58 32|22 3|1|5|2|1|9|6|5|3|4 1 1 1
CSPP1 18|34 0|0|0|0|0|0|0|0|0|0 1 1 1
EML4 21|31 0|0|0|8|11|0|0|0|0|0 0.93 0.97 0.93
HIST3H2BB 26|20 0|0|0|0|0|0|0|0|0|0 1 1 1
WRAP73 22|24 0|0|0|0|0|0|0|0|0|0 1 1 1
PIBF1 20|25 0|0|0|0|0|0|0|0|0|0 1 1 1
HNRNPA3 21|22 2|3|5|1|1|5|3|2|6|7 1 1 1

141
Table 3.2 Continued
Prey Counts Control Counts AvgP MaxP SaintScore
CKB 24|19 1|0|0|0|0|0|0|0|1|1 1 1 1
HAUS5 14|27 0|0|0|0|0|0|1|1|0|0 1 1 1
SKA1 14|20 0|0|0|0|0|0|1|0|1|0 1 1 1
PHIP 27|5 0|0|0|0|0|0|0|0|0|0 1 1 1
CEP55 14|18 0|0|0|0|0|0|2|2|0|0 1 1 1
CEP72 12|19 0|0|0|0|0|0|0|0|0|0 1 1 1
HAUS8 12|18 0|0|0|0|0|0|0|0|0|0 1 1 1
MAP7 10|20 0|0|0|0|0|0|0|0|0|0 1 1 1
POP1 14|15 0|0|7|2|4|6|1|1|1|0 0.96 0.98 0.96
RPS23 15|13 4|4|4|1|2|4|4|4|0|0 0.96 0.99 0.96
MAP7D1 12|15 0|0|0|0|0|0|0|0|0|0 1 1 1
ECH1 16|11 0|0|0|0|0|0|0|0|0|0 1 1 1
IPO8 12|14 0|0|0|0|0|0|0|0|0|0 1 1 1
CEP350 10|16 0|0|0|0|0|0|0|0|0|0 1 1 1
CLASP1 9|16 0|0|0|0|0|0|0|0|0|0 1 1 1
KIF2A 11|14 1|0|0|0|0|0|0|0|0|1 1 1 1
HMMR 11|14 0|0|0|0|0|0|0|0|0|0 1 1 1
TP53 14|11 3|4|0|0|0|0|0|0|0|0 0.97 0.99 0.97
SLC25A13 11|13 0|0|0|0|0|0|5|2|4|3 0.92 0.98 0.92
HAUS7 9|14 0|0|0|0|0|0|1|1|0|0 1 1 1
GTSE1 8|15 0|0|0|0|0|0|0|0|0|0 1 1 1
MAP9 7|15 0|0|0|0|0|0|0|0|0|0 1 1 1
GPI 15|7 0|0|0|0|0|0|0|0|0|0 1 1 1
KIF14 8|13 0|0|0|0|0|0|0|0|0|0 1 1 1
NEDD1 9|12 0|0|0|0|0|0|0|0|0|0 1 1 1
BANF1 10|10 0|0|0|0|1|0|0|0|0|0 1 1 1
SPAG5 6|13 0|0|0|0|0|0|0|0|0|0 1 1 1
MED4 6|13 0|0|0|0|0|0|0|0|0|0 1 1 1
UCHL1 10|9 0|0|0|0|0|0|0|0|0|0 1 1 1
C15orf23 8|10 0|0|0|0|0|0|0|0|0|0 1 1 1
CENPJ 4|14 0|0|0|0|0|0|0|0|0|0 1 1 1
SPC24 4|14 0|0|0|0|0|0|0|0|0|0 1 1 1
HAUS4 4|13 0|0|0|0|0|0|0|0|0|0 1 1 1
PRDX3 9|8 0|0|0|0|0|0|2|0|0|0 0.99 1 0.99
TP53BP2 3|14 0|0|0|0|0|0|1|1|0|0 0.91 1 0.91
RIN3 8|8 0|0|0|0|0|0|0|0|0|0 1 1 1
VPS33B 5|11 0|0|0|0|0|0|0|0|0|0 1 1 1
LOC100508808 8|7 0|0|0|0|0|0|0|0|0|0 1 1 1
ZC2HC1A 4|11 0|0|0|0|0|0|0|0|0|0 1 1 1
RACGAP1 8|7 0|0|2|0|0|3|0|0|0|0 0.94 0.96 0.94
PDZD11 7|7 0|0|0|0|0|0|0|0|0|0 1 1 1

142
Table 3.2 Continued
Prey Counts Control Counts AvgP MaxP SaintScore
NDC80 7|7 0|0|0|0|0|0|0|0|0|0 1 1 1
CCNB1 5|9 0|0|0|0|0|0|0|0|0|0 1 1 1
LOC728307 7|7 0|0|0|0|0|0|0|0|0|0 1 1 1
TXLNG 5|9 0|0|0|0|0|0|0|0|0|0 1 1 1
CKAP2L 5|9 0|0|0|0|0|0|0|0|0|0 1 1 1
HAUS1 2|12 0|0|0|0|0|0|0|0|0|0 0.98 1 0.98
BTF3 7|6 0|0|0|0|0|0|1|0|0|1 1 1 1
MCTS1 6|7 0|0|0|0|0|0|0|0|1|0 1 1 1
KIAA1009 5|8 0|0|0|0|0|0|0|0|0|0 1 1 1
CAMSAP2 2|11 0|0|0|0|0|0|0|0|0|0 0.98 1 0.98
RALY 8|5 0|2|0|0|0|0|0|0|0|0 0.97 0.99 0.97
FAM161A 5|7 0|0|0|0|0|0|0|0|0|0 1 1 1
RPGRIP1L 3|9 0|0|0|0|0|0|0|0|0|0 1 1 1
HAUS2 4|8 0|0|0|0|0|0|0|0|0|0 1 1 1
COPA 8|4 0|0|0|0|0|0|0|0|0|0 1 1 1
CCDC66 7|5 0|0|0|0|0|0|0|0|0|0 1 1 1
TCHP 3|9 0|0|0|0|0|0|1|0|0|0 0.98 1 0.98
ARID1B 6|5 0|0|0|0|0|0|0|0|0|0 1 1 1
CEP44 4|7 0|0|0|0|0|0|0|0|0|0 1 1 1
CEP85 4|7 0|0|0|0|0|0|0|0|0|0 1 1 1
LTV1 5|6 0|0|0|0|0|0|0|0|0|0 1 1 1
HMGN1 6|5 0|1|0|0|0|2|0|0|0|0 0.97 0.99 0.97
DNM1L 7|4 0|1|0|0|0|0|2|0|0|0 0.93 1 0.93
MAP1S 4|6 0|0|0|0|0|0|0|0|0|0 1 1 1
IPO4 6|4 0|0|0|0|0|0|0|0|0|0 1 1 1
ZC3H15 3|7 0|0|0|1|0|0|0|0|0|0 0.98 1 0.98
TRA2B 4|6 0|0|0|0|0|0|0|0|1|2 0.92 0.99 0.92
DIAPH3 5|4 0|0|0|0|0|0|0|0|0|0 1 1 1
TROAP 3|6 0|0|0|0|0|0|0|0|0|0 1 1 1
CC2D1A 3|6 0|0|0|0|0|0|1|0|0|0 0.98 1 0.98
CCDC77 2|7 0|0|0|0|0|0|0|0|0|0 0.98 1 0.98
MPHOSPH9 2|6 0|0|0|0|0|0|0|0|0|0 0.98 1 0.98
ANAPC7 6|2 0|0|0|0|0|0|0|0|0|0 0.98 1 0.98
C1orf35 6|2 0|0|0|1|0|0|0|0|0|0 0.9 1 0.9
SRP72 4|3 0|0|0|0|0|0|0|0|0|0 1 1 1
CDC27 3|4 0|0|0|0|0|0|0|0|0|0 1 1 1
TEX9 2|5 0|0|0|0|0|0|0|0|0|0 0.98 1 0.98
MAP7D2 2|5 0|0|0|0|0|0|0|0|0|0 0.98 1 0.98
UBE2L3 4|3 0|0|0|0|0|0|1|0|0|0 0.97 0.99 0.97
DCLK1 2|5 0|0|0|0|0|0|0|0|1|0 0.9 1 0.9
POLH 2|5 0|0|0|0|0|0|0|1|0|0 0.9 1 0.9

143
Table 3.2: SPICE1 Associated Proteins Detected by BioID.
Results from two biological replicates of FLAG-BirA*-SPICE1 expression in the presence of
biotin followed by affinity-purification and mass-spectrometry to identify biotinylated peptides.
Control counts are from 10 runs from four biological FLAG-BirA expression experiments
prepared under the same conditions. (From left to right) Gene Symbol for identified proteins, the
spectral counts for each protein in each of the experimental replicates, the spectral counts for
each protein in the control replicates, the average probability of both replicates (AvgP), the
maximum probability between the two replicates (MaxP), and the overall SAINT score for both
replicates are shown (Choi et al., 2011). Spectra reflect the number of spectral counts assigned
to a prey polypeptide in each MS analysis. Samples were prepared by Sally Cheung, and BioID-
MS and was carried out by Dr. Etienne Coyaud.

144
Figure 3.2 Cytoscape map of CEP120 and SPICE1 BioID Preys.
The data from BioID experiments (see Tables 3.1 and 3.2) were input into the Cytoscape
Software program and used to generate a map of shared and unique prey for CEP120 and
SPICE1. Each circle within a cluster represents a prey protein, and the thickness of the
connecting line is proportional to the total peptide counts for each protein over the two
experimental replicates. Prey proteins shared by CEP120 and SPICE1 are shown (center), and
prey proteins unique to CEP120 (top) and SPICE1 (bottom) are shown.
Figure 3.2

145
Figure 3.3
Figure 3.3. BioID identified proteins in proximity to CEP120 or SPICE1 are important for
PLK4 induced centriole duplication in U-2 OS cells.
(A, C) In two separate experiments (A,C) U-2 OS cells on coverslips were transfected with
siRNA or esiRNA against target genes as indicated, where gene-symbol is given followed by si
or esi and a number to differentiate between multiple silencing triggers tested for each gene. At
48 hours post-transfection with RNA, cells were arrested in S-phase with 8 mM hydroxyurea and
induced to over-express PLK4 by addition of tetracycline to 2g/ml. At 20 hours post induction
and arrest, cells were fixed in methanol and immunolabeled for imaging and visual analysis. The
frequency of cells with overduplicated centrioles was quantified for each condition, with three
replicates per condition, 100 cells analyzed in each replicate. Bar graph shows frequency of
overduplication, grouped based on p-values from a two-tailed student’s t-test against controls.
Asterisks indicate significance as a result of an unpaired two-tailed student’s t-test against the
control. (B) Sample images of (top) overduplication and (bottom) CEP120 depletion preventing
centriole overduplication from the same assay as in (A).

146
Figure 3.4

147
Figure 3.4 Depletion of CEP120/SPICE1-proximal proteins involved in centriole
duplication results in CEP120 localization outside of the centrosome.
(A) U-2 OS cells were depleted of the indicated proteins for 72 hours by RNAi while cycling.
Cells were fixed and immunolabeled for CEP120 (green) and glutamylated tubulin (red). The
frequency of cells with CEP120 observed on non-centrosomal structures resembling
microtubules was quantified for each condition, and bar graph shows the frequency for each
depletion condition. The experiment was performed in triplicate, with at least 100 cells
examined per condition per replicate. P-values (*s) are results of student’s t-test, two-tailed
compared to control for each condition. (B) Representative micrographs showing CEP120
localization (green, insets) under each condition. Gene name above pictures indicates RNAi
treatment used. Scale bar 10 m.

148
Figure 3.5

149
Figure 3.5 Depletion of CEP120/SPICE1-proximal proteins involved in centriole
duplication results in abnormal SPICE1 structures.
(A) U-2 OS cells were depleted of the indicated proteins for 72 hours by RNAi while cycling.
Cells were fixed and immunolabeled with antibodies against SPICE1 (green) and glutamylated
tubulin (red). The bar graph shows the frequency of cells with SPICE1 forming abnormal
structures for each RNAi condition. The experiment was performed in triplicate, with at least 100
cells examined per condition per replicate. P-values (*s) are results of student’s t-test, two-tailed
compared to control for each condition. (B) Representative micrographs showing SPICE1
localization (green, insets) under each condition. Gene name above pictures indicates RNAi
treatment used. Scale bar 10 m.

150
Figure 3.6

151
Figure 3.6. Depletion of CEP120, SPICE1 and their associated proteins can lead to
cytoskeleton microtubule glutamylation.
(A) U-2 OS cells were depleted of the indicated proteins for 72 hours by RNAi while cycling.
Cells were fixed and immunolabeled for glutamylated tubulin (red) and CEP120 (green). Bar
graph shows the frequency of cells with glutamylated non-centriolar microtubules for each RNAi
condition. The experiment was performed in triplicate, with at least 100 cells examined per
condition per replicate. P-values (*s) are results of a two-tailed student’s t-test compared to
control for each condition. (B) Representative micrographs showing glutamylated tubulin
immunolabeling (red, insets). Gene name above pictures indicates RNAi treatment, scale bar 10
m.

152
Figure 3.7

153
Figure 3.7. Depletion of CEP120/SPICE1 associated proteins can lead to elongated
centriole-like structures.
(A) Bar graph showing the quantification of the frequency of elongated structures that emanated
from centrioles following 72 hours of depletion of the indicated proteins by RNAi in U-2 OS
cells. Quantification was performed in triplicate with at least 100 cells counted per replicate in
each condition. (B) Representative images of centrioles in each condition, with insets showing
glutamylated microtubules in the area of the centrioles (identified by a pair of brightly
glutamylated foci). Scale bars 5 m for both panels.

154
Figure 3.8

155
Figure 3. 8. CEP120/SPICE1 associated proteins are required for taxol induced centriole
elongation.
(A)U-2 OS cells were depleted of the indicated proteins for 24 hours before being arrested in G2
for 48 hours, with taxol added to 10 M for the final 24 of those hours. Cells were fixed and
immunolabeled with antibodies against CEP120, -tubulin and glutamylated tubulin. The
frequency with which elongated glutamylated tubulin structures were observed was quantified
for each condition. Bar graph shows frequency for each condition in decreasing order (from left
to right), with control bar in grey. P-values indicated are results of a two-tailed student’s t-test
compared to controls. For each condition, three replicates were tested and at least 100 cells were
examined per replicate. (B) Representative images from the experiment as described in (A), with
scale bar indicating 10 m and insets showing 4x magnification of centrosomes and
glutamylated tubulin staining.

156
Figure 3.9

157
Figure 3.9. CEP120/SPICE1 associated proteins are required for taxol induced centriole
elongation.
(A) The experiment was carried out as described in Figure 3.7, with the frequency of elongated
CEP120 structures being quantified for each condition. Bar graph shows (from left to right)
increasing inhibition of elongation based on frequency, with control bar in grey. P-values
indicated are results of a two-tailed student’s t-test compared to controls. For each condition,
three replicates were tested and at least 100 cells were examined per replicate. (B) Images used
are from Figure 3.7, with scale bar indicating 10 m and 4x magnified insets, with the exception
that insets in this figure show CEP120 antibody labeling.

158
Table 3.3 Summary of the Phenotypes of CEP120 and SPICE1 Associated Proteins in
Secondary Assays.
Abnormal
Localization Acentriolar MT Elongated
Taxol Induced
Elongation
RNAi CEP120 SPICE1 Glutamylation Structures Glut. Tub. CEP120
ANK2 Y Y Y Y ↑ − MAP7 Y Y Y Y ↑ ↑
CSPP1 Y Y Y Y ↓ ↓
MTUS1 Y Y Y N ↓ ↓
CENTROB Y N Y N ↓ ↓
CEP135 Y N Y N ↓ − NAP1L1-si1 Y N Y N − −
MAP7D3 N Y Y N ↓ ↓
KIAA0586 N Y N Y ↓ ↑
KIAA1731 N Y N N ↓ − SASS6 Y N N N − −
NAP1L1-si17 N N N N ↓ ↓
MAP9 N N N N ↓ ↓
CPAP N N N N ↓ ↓
CCDC77 N N N N ↓ −
Table 3.3. Summary of the Phenotypes of CEP120 and SPICE1 Associated Proteins in
Secondary Assays.
RNAi (Column 1): Names of the proteins including controls and those identified as prey in the
CEP120 and/or SPICE1 BioID datasets that were required for PLK-4 induced centriole
duplication, that were tested in secondary assays. (Columns 2-5): U-2 OS cells were depleted of
the indicated proteins (column 1) by RNAi for 72 hours before fixation.
Abnormal Localization (Columns 2 and 3): Is localization of CEP120 or SPICE1 to
cytoplasmic microtubules observed in this assay significantly increased under each condition
compared to the control (see Figures 3.4, 3.5)? Acentriolar MT Glutamylation (Column 4): Is
cytoplasmic microtubule glutamylation significantly increased under each condition in this assay
compared to the control (see Figure 3.6)? Elongated Structures (Column 5): Are elongated
centriole-like structures observed in more than 5% of cells under each condition (see Figure
3.7)? Taxol Induced Elongation (Columns 6 and 7): Is the taxol driven elongation of
centrioles inhibited (↓), increased (↑) or unchanged (−) by each RNAi compared to controls in
the taxol assay when either glutamylated tubulin (Glut. Tub) or CEP120 (CEP120) are used as
markers for centriole length (see Figures 3.8 and 3.9)?

159
Figure 3.10
Figure 3.10. NAP1L1 Depletion Reduces PLK4 induced centriole overduplication in U-2
OS cells.
In separate experiments, U-2 OS cells with inducible PLK4 expression on coverslips were
transfected with either LifeTech Stealth siRNA against NAP1L1 or Dharmacon siRNAs against
NAP1L1, or controls against CEP135 or CEP120, respectively. At 48 hours post-transfection
cells were arrested in S-phase with hydroxyurea and induced to overexpress PLK4 by addition of
tetracycline. At 20 hours post induction and arrest, cells were fixed and immunolabeled with
antibodies against CEP120 and CETN2. (Top) Representative images from the Stealth siRNA
experiment are shown, with scale bar representing 10 m and insets magnified 4x. (Bottom) Bar
graphs quantifying the frequency of centriole overduplication under each RNAi condition.
Asterisks over bars indicate significance for each sample as compared to control-transfection
based on results of a two-tailed unpaired student’s t-test.

160
Figure 3.11

161
Figure 3.11. NAP1L1 is required for centriole duplication in cycling U-2 OS cells.
U-2 OS cells were depleted of NAP1L1 using one of two LifeTech Stealth siRNAs (NAP1L1-
Stealth-1, NAP1L1 Stealth-2), or one of three Dharmacon siRNAs (NAP1L1-DHRM-2, 17, 18),
or CEP120 or SPICE1 for 72 hours before being fixed and immunolabeled with antibodies
detecting CETN2 and CEP120. Mitotic cells were examined, and the number of centrioles per
mitotic cell was counted based on CETN2 labeling. (A) Representative images are shown from
each RNAi condition, with scale bar representing 10 m and insets 2x magnified below. The
frequency of cells with four or more centrioles was determined under each condition and bar
graph (B) shows the percentage of cells for each RNAi condition, as the average from three
replicates for each of which at least 50 cells were counted. Asterisks indicate significance for
each sample as compared to the control, based on results of two-tailed unpaired student’s t-test,
with RNAi condition indicated on the X-axis.

162
Figure 3.12

163
Figure 3.12. NAP1L1 is required for centriole duplication in cycling HeLa cells.
HeLa cells were depleted of NAP1L1 using multiple siRNAs from LifeTech (NAP1L1-Stealth-1,
NAP1L1 Stealth-2) or Dharmacon (NAP1L1-DHRM-2, 17, 18), or depleted of CEP120 or
SPICE1 for 96 hours before being fixed and immunolabeled with antibodies detecting CETN2
and CEP120. Representative mitotic cells (A) are shown from each condition, with scale-bar
indicating 10 m and insets 2x magnified. The frequency of cells with four or more centrioles in
mitosis was quantified visually based on CETN2 signal, and bar graph (B) shows the average
percentage of cells meeting this criteria for each condition from three independent replicates for
each of which at least 50 mitotic cells were examined. Asterisks indicate the results of a
student’s t-test of significance (two-tailed, unpaired) comparing each condition to the control.

164
Figure 3.13

165
Figure 3.13. NAP1L1 depletion by Stealth siRNA#1 causes CEP120 localization to
perinuclear acetylated microtubules in the PLK4 centriole overduplication assay.
U-2 OS cells that express PLK4 upon treatment with tetracycline were depleted of CEP120 or
NAP1L1 using Stealth siRNA#1 for 48 hours before being arrested in S-phase and induced to
over-express PLK4. At 20 hours post-induction, cells were fixed and immunolabeled for (A)
CEP120 and CETN2, or (B) CEP135 and acetylated tubulin (Ac.Tub). Representative cells are
shown for each immunolabeling, and insets show (A) CEP120 or (B) acetylated tubulin staining
from the experiments. Scale bars 100 m.

166
Figure 3.14

167
Figure 3.14. Depletion of NAP1L1 by Stealth siRNA#1 leads to CEP120 localization on
cytoplasmic structures in U-2 OS cells.
U-2 OS cells were depleted of NAP1L1 using siRNA indicated, or CEP120 or SPICE1 for 72
hours before being fixed and immunolabeled with antibodies against CEP120 and CETN2.
Representative images are shown for each RNAi condition (A) with insets showing the CEP120
label, and scale bar approximating 10 m. The frequency of CEP120 on cytoplasmic
microtubules was quantified for each condition, and the average percentage of cells with CEP120
on cytoplasmic structures from three replicates for each of which at least 100 cells was counted
is shown in the bar graph. (B) Asterisks show significance of differences compared to controls
for each RNAi condition, as determined by unpaired two-tailed student t-test of significance.

168
Figure 3.15

169
Figure 3.15. NAP1L1-Stealth-siRNA#1 resistant GFP-NAP1L1 does not rescue the
phenotype of CEP120 distribution onto cytoplasmic structures.
U-2 OS stable cell lines expressing either GFP alone or a GFP-tagged NAP1L1 that is resistant
to Stealth siRNA#1 (GFP-NAP1L1-siRES) were generated. Both cell lines were treated with
control siRNA or Stealth siRNA#1 targeting NAP1L1 for 72 hours before being fixed and
immunolabeled with antibodies against CEP120 and GFP. Representative images are shown (A)
for each condition, scale bars indicate 100 m, and insets are 3X magnifications showing the
CEP120 signal. GFP-positive cells were analyzed under each condition, and the frequency of
CEP120 being observable on cytoplasmic microtubules was quantified for each condition. Bar
graph (B) shows the average frequency of the CEP120 redistribution phenotype from three
replicates of the experiment, where at least 100 cells were examined per replicate. (C) In a
separate experiment, the U-2 OS cell line described and shown in (A) (right-side) was treated
with Stealth siRNA#1, Dharmacon siRNA#17 (DHRM si#17), or esiRNA against GFP for 72
hours, before whole cell lysates were collected and analyzed by Western blot. Blot shows that
GFP-NAP1L1-siRES is sensitive to RNAi targeting GFP, or DHRM-si#17, but resistant to
Stealth siRNA#1 against NAP1L1.

170
Figure 3.16

171
Figure 3.16. SiRNA resistant NAP1L1-GFP and GFP-NAP1L1 do not restore centriole
duplication in U-2 OS cells treated with siRNA against NAP1L1.
(A, B, D, E) U-2 OS cells stably expressing either GFP or GFP fused to NAP1L1 that is resistant
to Dharmacon siRNA#17 were treated with control or Dharmacon siRNA#17 targeting NAP1L1
for 48 hours before being arrested in S-phase for 24 hours then fixed and immunolabeled with
antibodies against CEP135 and CETN2. (A) Representative images of U-2 OS from the
experiment expressing (left) GFP or (right) siRNA resistant NAP1L1 with GFP fused to its
carboxy-terminal end, under treatment with control or NAP1L1 siRNA#17. CEP135 was
immunolabeled with Alexa-488 and so the strong signal renders GFP signal below background
in the images. Scale bar 10 m, insets show 2X magnifications. (B) The number of centrioles
per cell was counted, and the frequency of cells with four or more centrioles was quantified over
three experiments where at least 100 cells were counted per replicate. Bar graph shows the
average from three experiments for each cell type. Asterisks indicate the result of an unpaired
two-tailed student’s t-test for significance against controls as indicated. (C) Cell lysates were
prepared from the same cell lines as in (A), following the same protocol as in (A). U-2 OS were
treated with Dharmacon siRNA#17, Stealth si#1, esiRNA targeting GFP, or control siRNA and
lysates were collected for Western blot. Western blot shows (top) GFP signal and (bottom)
GAPDH loading controls for each experiment. (D) Representative images from an experiment
as described for (A), but with the NAP1L1 expressed with GFP fused to its amino-terminal end
(right side). (E) The number of centrioles per cell was quantified, and the frequency of cells
with four or more centrioles was averaged from three experiments where at least 100 cells were
counted per replicate. Asterisks indicate the result of an unpaired two-tailed student’s t-test for
significance against controls as indicated. (F) Cell lysates were prepared from U-2 OS cells
treated as in (D), with Dharmacon siRNA#17, Stealth si#1, esiRNA targeting GFP, or control
siRNA. Western blot shows (top) GFP signal and (bottom) GAPDH loading controls for each
experiment.

172
Figure 3.17
N1L1-1 MADIDNKEQSELDQDLDDVEEVEEEETGEETKLKARQLTVQMMQNPQILA 50
N1L1-2 MADIDNKEQSELDQDLDDVEEVEEEETGEETKLKARQLTVQMMQNPQILA 50
N1L4-1 MADHSFSDG----VPSDSVEAAKNASN-------TEKLTDQVMQNPRVLA 39
N1L4-2 MADHSFSDG----VPSDSVEAAKNASN-------TEKLTDQVMQNPRVLA 39
*** . .: *.** .:: .. :.:** *:****::**
ALQERLDGLVETPTGYIESLPRVVKRRVNALKNLQVKCAQIEAKFYEEVH 100
ALQERLDGLVETPTGYIESLPRVVKRRVNALKNLQVKCAQIEAKFYEEVH 100
ALQERLDNVPHTPSSYIETLPKAVKRRINALKQLQVRCAHIEAKFYEEVH 89
ALQERLDNVPHTPSSYIETLPKAVKRRINALKQLQVRCAHIEAKFYEEVH 89
*******.: .**:.***:**:.****:****:***:**:**********
DLERKYAVLYQPLFDKRFEIINAIYEPTEEECEWKPD--EEDEISEELKE 148
DLERKYAVLYQPLFDKRFEIINAIYEPTEEECEWKPD--EEDEISEELKE 148
DLERKYAALYQPLFDKRREFITGDVEPTDAESEWHSENEEEEKLAGDMKS 139
DLERKYAALYQPLFDKRREFITGDVEPTDAESEWHSENEEEEKLAGDMKS 139
*******.********* *:*.. ***: *.**:.: **:::: ::*.
KAKIEDEKKDEEKE-DPKGIPEFWLTVFKNVDLLSDMVQEHDEPILKHLK 197
KAKIEDEKKDEEKE-DPKGIPEFWLTVFKNVDLLSDMVQEHDEPILKHLK 197
KVVVTEKAAATAEEPDPKGIPEFWFTIFRNVDMLSELVQEYDEPILKHLQ 189
KVVVTEKAAATAEEPDPKGIPEFWFTIFRNVDMLSELVQEYDEPILKHLQ 189
*. : :: :* *********:*:*:***:**::***:********:
DIKVKFSDAGQPMSFVLEFHFEPNEYFTNEVLTKTYRMRSEPDDSDPFSF 247
DIKVKFSDAGQPMSFVLEFHFEPNEYFTNEVLTKTYRMRSEPDDSDPFSF 247
DIKVKFSDPGQPMSFVLEFHFEPNDYFTNSVLTKTYKMKSEPDKADPFSF 239
DIKVKFSDPGQPMSFVLEFHFEPNDYFTNSVLTKTYKMKSEPDKADPFSF 239
********.***************:****.******:*:****.:*****
DGPEIMGCTGCQIDWKKGKNVTLKTIKKKQKHKGRGTVRTVTKTVSNDSF 297
DGPEIMGCTGCQIDWKKGKNVTLKTIKKKQKHKGRGTVRTVTKTVSNDSF 297
EGPEIVDCDGCTIDWKKGKNVTVKTIKKKQKHKGRGTVRTITKQVPNESF 289
EGPEIVDCDGCTIDWKKGKNVTVKTIKKKQKHKGRGTVRTITKQVPNESF 289
:****:.* ** **********:*****************:** *.*:**
FNFFAPPEVPESGD-LDDDAEAILAADFEIGHFLRERIIPRSVLYFTGEA 346
FNFFAPPEVPESGD-LDDDAEAILAADFEIGHFLRERIIPRSVLYFTGEA 346
FNFFNPLKASGDGESLDEDSEFTLASDFEIGHFFRERIVPRAVLYFTGEA 339
FNFFNPLKASGDGESLDEDSEFTLASDFEIGHFFRERIVPRAVLYFTGEA 339
**** * :.. .*: **:*:* **:*******:****:**:********
IEDDDD-DYDEEGEEADEEGEEEGDEENDPDYDPKKDQN-PAECKQQ 391
IEDDDD-DYDEEGEEADEVMFTK------------------------ 368
IEDDDNFEEGEEGEEEELEGDEEGEDEDDAEINPKV----------- 375
IEDDDNFEEGEEGEEEELEGDEEGEDEDDAEINPKKEPSQPAECKQQ 386
*****: : .***** : :

173
Figure 3.17. ClustalW Alignment of NAP1L1 and NAP1L4 isoforms. Isoforms for NAP1L1 and NAP1L4 were identified using the UniProt knowledge base online
(http://www.uniprot.org) (Operated by European Bioinformatics Institute (EMBL-EBI), Swiss
Institute of Bioinformatics (SIB), and the Protein Information Resource (PIR)). The FASTA
formatted sequences were then uploaded to ClustalW (http://www.ebi.ac.uk/Tools/msa/
clustalw2) (EMBL-EBI) and aligned using default parameters (slow alignment). Top row is
NAP1L1 isoform 1 (N1L1-1) (canonical), while the second row is NAP1L1 isoform 2 (UniProt
identifiers P55209-1, -2 respectively). Third row and fourth rows are NAP1L4 isoforms 1 and 2,
respectively (Uniprot identifiers Q99733-1,-2 respectively). Fifth row is the result of analysis of
the sequences by ClustalW, where “*” indicates conserved amino acid across all four proteins,
“:” indicates conservation between groups of strongly similar properties (scoring > 0.5 in the
Gonnet PAM 250 matrix), and “.” Indicates conservation between groups of weakly similar
properties (< 0.5 in the Gonnet PAM 250 matrix) (source: http://www.ebi.ac.uk/Tools/msa/
clustalw2/help).

174
Figure 3.18
Figure 3.18. Sequence Alignment of NAP1L1 siRNAs with NAP1L1 and NAP1L4 Long
isoforms.
The coding sequences of NAP1L1 and NAP1L4 long isoforms (NCBI Reference Sequence:
NM_139207.2 and NM_005969.3, respectively) were aligned using ClustalW
(http://www.ebi.ac.uk/Tools/msa/clustalw2) (EMBL-EBI). The target sequences of each
NAP1L1 siRNA were then manually identified within the NAP1L1 sequence. For (A) and (B),
top row of sequence show the NAP1L1 mRNA, bottom row the aligned NAP1L4 mRNA
sequence. Only the relevant regions of the mRNA alignment are shown. (A)The LifeTech
Stealth siRNA#1 (NAP1L1-si#1) target sequence is shown in bold (top), and in bold within the
NAP1L1 sequence. The mismatches with NAP1L4 mRNA are highlighted in red. (B)The
Dharmacon NAP1L1 siRNA #17 (NAP1L1-si#17) sequence is shown in bold. Alignment shown
as described in (A).
NAP1L1 si#1 Target Sequence: GGTAGAAACACCAACAGGATACATT
CAGATGATGCAAAATCCTCAGATTCTTGCAGCCCTTCAAGAAAGACTTGATGGTCTGGTA 180
CAGGTGATGCAGAATCCTCGAGTTCTGGCAGCTTTACAGGAGCGACTTGACAATGTCCCT 147
GAAACACCAACAGGATACATTGAAAGCCTGCCTAGGGTAGTTAAAAGACGAGTGAATGCT 240
CACACCCCTTCCAGCTACATCGAAACTTTACCTAAAGCAGTAAAAAGAAGAATTAATGCA 207
NAP1L1 si#17 Target Sequence: GCGTATAATCCCAAGATCA
TGAAGCTATCCTTGCTGCAGACTTCGAAATTGGTCACTTTTTACGTGAGCGTATAATCCC 1007
TGAATTCACATTAGCCTCTGATTTTGAAATTGGACACTTTTTCCGTGAGCGGATAGTCCC 986
AAGATCAGTGTTATATTTTACTGGAGAAGCTATTGAAGATGATGATGAT---GATTATGA 1064
GCGGGCTGTGCTGTACTTCACTGGGGAGGCCATAGAAGATGATGACAATTTTGAAGAAGG 1046

175
Figure 3.19
Figure 3.19. Relative quantification of NAP1L1 and NAP1L4 transcript levels by PCR.
PCR was performed on cDNA made from normalized amounts of total-RNA from U-2 OS at 72
hours following treatment with siRNA against either NAP1L1 (siRNA#1 or siRNA#17),
NAP1L4, or a control siRNA with no target. Primer sets were designed to target cDNA from
both NAP1L1 isoforms (left lanes), from NAP1L4 specifically (second lane), from the NAP1L1
short isoform (third lane), or GAPDH (fourth lane). (Top) A 2% agarose gel containing
ethidium bromide was run with equal sample loading of each PCR reaction. (Bottom) The lanes
for each sample from the gel (top) were isolated using Photoshop and labelled with their primers
(top) and sample experimental condition (left). Two different exposures of the same gel are
shown (Left versus right lower panels).

176
Figure 3.20

177
Figure 3.20. NAP1L4 is not required for PLK4 induced centriole over duplication or
CEP120 localization to centrosomes in U-2 OS cells.
(A) U-2 OS cells with inducible PLK4 expression were transfected with control siRNA, siRNA
targeting CEP120, or siRNA targeting NAP1L4, and at 48 hours post-transfection were arrested
in S-phase and induced to over-express PLK4 by addition of tetracycline. At 20 hours post
induction and arrest, cells were fixed and immunolabeled with antibodies against CEP120 and
CETN2. Representative images (A) are shown from each RNAi condition, with insets shown at
4X magnification and scale bar representing 10 m. (B) The frequency of centriole
overduplication was quantified for each condition, and bar graph shows the average percentage
of cells with centriole overduplication from three replicates for each condition, where at least
100 cells were counted per replicate. (C) U-2 OS cells treated as described for (A), but with
siRNA against NAP1L1 (Stealth si#1) instead of CEP120. (C) Representative images of
populations of cells showing CEP120 localization (insets), scale bar 100 m. (D) The frequency
of CEP120 observed on cytoplasmic structures was quantified for each condition, and the
average percentage of cells with CEP120 on cytoplasmic microtubules was calculated from three
replicates of each experiment with at least 100 cells being examined per replicate. For (B) and
(D), asterisks indicate results of a student’s t-test of significance comparing the condition to the
control condition.

178
Figure 3.21
Figure 3.21. NAP1L1 is required for PLK4 induced centriole overduplication.
(A) U-2 OS cells with inducible PLK4 expression were transfected with control siRNA, or one
of three siRNAs targeting NAP1L1 (Stealth-si#1, Dharmacon si#17, or a third siRNA (see text)).
At 48 hours post-transfection cells were arrested in S-phase and induced to over-express PLK4,
then at 20 hours post induction and arrest, cells were fixed and immunolabeled for CEP120 and
CETN2. Representative images (A) are shown from each RNAi condition, with insets shown at
8X magnification and scale bar representing 10 m. (B) The frequency of centriole
overduplication was quantified for each condition, and bar graph shows the average percentage
of cells with centriole overduplication from three replicates for each condition, where at least
100 cells were counted per replicate. (C) Representative images of populations of cells showing
CEP120 localization, scale bar 10 m. (D) The percentage of cells with CEP120 on cytoplasmic
microtubules was calculated for each of three replicates, with at least 100 cells being examined
per replicate. Bar graph shows average of three replicates for each condition. For (B) and (D),
asterisks indicate results of a student’s t-test of significance comparing the condition to the
control condition.

179
Table 3.4. Summary of the NAP1L1 siRNAs and Their Associated Phenotypes.
Dharmacon siRNAs Stealth siRNAs
Si#2 Si#17 Si#18 Si#1 Si#2 Si#3 Si-New
Isoforms Targeted BOTH BOTH BOTH BOTH BOTH BOTH BOTH
Centriole
Overduplication YES YES YES YES YES NO YES
Centriole Duplication
(72h RNAi, U-2 OS) NO YES NO YES NO NT NT
CEP120 Localization
(72h RNAi, U-2 OS) YES NO YES YES NO NT YES
MT Glutamylation
(72h RNAi, U-2 OS) NT NO NT YES NT NT NT
Table 3.4. Summary of the NAP1L1 siRNAs and Their Associated Phenotypes.
(First Column) Characteristics of different NAP1L1 siRNAs and their impacts in various assays.
Isoforms Targeted: NAP1L1 isoforms predicted to be targeted by the siRNAs based on manual
sequence alignments. Centriole Overduplication: Does siRNA indicated inhibit PLK4 induced
centriole overduplication (see Figures 3.10, 3.21)? Centriole Duplication: Does indicated
siRNA inhibit centriole duplication at 72 hours depletion in U-2 OS cells (see Figure 3.11)?
CEP120 Localization: Does indicated siRNA lead to abnormal CEP120 localization in U-2 OS
at 72 hours (see Figures 3.14 and 3.21)? MT Glutamylation: Does indicated siRNA lead to
abnormal glutamylation of cytoplasmic microtubules in U-2 OS cells at 72 hours RNAi. ‘YES’
indicates statistically significant phenotype in the assay as described. ‘NO’ indicates no
phenotype, or a non-significant phenotype in the assay as described. ‘NT’ indicates the siRNA
was not tested in the assay as described. (Top Row) Indicated siRNAs targeting NAP1L1 that
were used in this study (see Materials and Methods).

180
3.7. Materials and Methods
3.7.1. Cell Culture, RNAi, PLK4 Assay, and Microscopy Sample Preparation
and Imaging
Cell culture conditions, RNA transfections, the PLK4 and taxol assays, and microscopy methods
used in this chapter are as described in detail in Section 2.7 of this thesis (Chapter 2 Materials
and methods).
3.7.2. BioID Analysis
All mass-spectrometry was performed by Etienne Coyaud from the lab of Dr. B. Raught as
previously reported (Comartin et al., 2013). Briefly, analytical columns and pre-columns (75 nm
and 150 nm inner diameters, respectively) were made from silica capillary tubing (InnovaQuartz)
and packed with 100 Angstrom C18-coated silica particles (Magic, Micrhom Bioresources).
Peptides were subjected to liquid chromatography electrospray ionization-MS/MS using 120
minute reversed-phase (100% water 100% acetonitrile, 0.1% formic acid) buffer gradient at 250
nl/min on a Proxeon EASY-nLC pump in line with a hybrid LTQ-Orbitrap Velos mass
spectrometer (Thermo Fisher Scientific). A parent ion scan was performed using a resolving
power of 60,000, then up to the forty most intense peaks were selected for MS/MS (minimum
ion count of 1000 for activation), using standard collision-induced dissociation fragmentation.
Fragment ions were detected in the LTQ. Dynamic exclusion was activated such that MS/MS of
the same m/z (within 15 ppm; exclusion list size =500) detected twice within 15 seconds were
excluded from analysis for 30 seconds. For protein identification, Thermo .RAW files were
converted to the .mzXML format using Proteowizard (Kessner et al., 2008), then searched using
X!Tandem (Craig and Beavis, 2004) against the human RefSeq database. X!Tandem search
parameters were: 15 ppm parent mass error; 0.4 Da fragment mass error; complete
modifications, none; cysteine modification, none; potentials modifications, +16@M and W,
+32@M and W, +42@N-terminus, +1@N and Q. For both CEP120 and SPICE1 datasets, two
technical replicates were used for analysis. Samples were analyzed using the trans-proteomic
pipeline (TPP) in the ProHits software suite (Choi et al., 2011, 2012; Deutsch et al., 2010;
Pedrioli, 2010). Proteins identified were analyzed with the SAINT algorithm, using cut-off
values of AvgP <0.90, MaxP<0.95, and with pre-filtering against heat-shock proteins, ribosomal
proteins, keratins, albumin, and translation elongation factors (Choi et al., 2012). Ten runs of

181
FLAG-BirA* peptide expression pellets were used as controls to remove non-specific
interactions in SAINT, and control counts are reported for each protein.
3.7.3. PLK4 Screen
As described previously in Chapter 2, and similar to the originally published protocol (Kleylein-
Sohn et al., 2007), U-2 OS T-REx cells with inducible Myc-PLK4 expression were seeded on
coverslips in 6-well tissue culture dishes and transfected with siRNA or esiRNA as described in
Materials and Methods Section 2.7.2. At 48 hours post-RNA addition, cells were arrested in S-
phase by addition of hydroxyurea (HU) to 8 mM, and induced to over-express PLK4 by addition
of tetracycline (2g/mL) for 20 hours before fixation.
3.7.4. Analyses of CEP120 and SPICE1 localization, Microtubule
Glutamylation and Centriole Elongation following Depletion of CEP120 and
SPICE1 Associated Proteins in U-2 OS
For the U-2 OS assays wherein the localization of CEP120 or SPICE1were analyzed (Figure 3.4,
Figure 3.5), or wherein the fraction of cells with glutamylated cytoplasmic microtubules or
elongated centrioles were analyzed (Figure 3.6 and Figure 3.7), U-2 OS cells were treated with
siRNA or esiRNA for 72 hours before fixation by ice-cold methanol (-20°C) for at least 10
minutes. Microscopy sample preparation and image processing were as described in Materials
and Methods section 2.7.6. Taxol assays were performed as described in Materials and Methods
section 2.7.4.
3.7.5. Bioinformatic Analysis of NAP1L-family members and Isoforms
UniprotKB (http://www.uniprot.org) (EMBL-EBI, SIB, and the PIR) was used to obtain
information about NAP1L1 and NAP1L4 predicted isoform protein sequences in June 2014. The
FASTA sequences were then uploaded to ClustalW (http://www.ebi.ac.uk/Tools/msa/clustalw2)
(EMBL-EBI) and aligned using default parameters (slow alignment) for protein alignment. For
mRNA alignment of NAP1L1 vs NAP1L4 long isoforms, the NCBI reference sequences
(NM_139207.2 and NM_005969.3) were obtained, the UTRs manually removed and the
sequences double-checked against the protein sequences of the appropriate isoforms by first
translating the mRNA to amino-acid sequence (http://web.expasy.org/translate/, SIB), then
aligning them against the appropriate FASTA sequence from UniProtKB using ClustalW. To

182
align the mRNA sequences of NAP1L1 and NAP1L4, ClustalW was again used (DNA
alignment, default settings, slow alignment). NAP1L1 siRNAs target sequences were manually
identified from within the NAP1L1 sequence, and mismatches were manually identified. For
comparison of the two NAP1L1 isoforms and the NAP1L4 isoforms, all four FASTA protein
sequences were obtained from UniProt KB, and aligned simultaneously in ClustalW using
default parameters for protein alignment (slow method).
3.7.6. Cloning of Rescue Constructs
The NAP1L1 long isoform was mutated at wobble-codon positions using QuikChange II XL
site-directed mutagenesis kit (Agilent Technologies). For NAP1L1 siRNA#1 resistance, the
primers used were 5’- TTTAACTACCCTAGGCAGGCTTTCAATATAACCGGTAGGCGT
CTCGACCAGACCATCAAGTCTTTCTTGAAG G-3’ and 5’-CCTTCAAGAAAGACTTGAT
GGTCTGGTC GAGACGCCTACCGGTTATATTGAAAGCCTGCCTAGGGTAGTTAAA-3’.
For NAP1L1 si#17 resistance, the primers used were 5’- CTTCAATAGCTTCTCCAGTAAAA
TATAACACCGACCTGGGAATGATGCGCTCACGTAAAAAGTGACCAATTTCGAAGT-3’
and 5’-ACTTCGAAATTGGTCACTTTTTACGTGAGCGCATCATTCCCAGGTCGGTGTTA
TATTTTACTGGAGAAGCTATTGAAG-3’.
3.7.7. Quantitative PCR to Detect NAP1L1 and NAP1L4 Transcripts
Following treatment with siRNAs Against NAP1L1
Equal amounts of U-2 OS cells were plated and treated with siRNAs as described in Materials
and Methods section 2.7.2. Cells were harvested and RNA prepared using and OMEGA kit
(OMEGA Bio-Tek) as per manufacturer’s protocols. RNA samples were treated with RNAse-
free DNAse-I (Qiagen) as per manufacturer’s recommendation for 15 minutes at room
temperature before being inactivated by incubation at 65°C in the presence of EDTA for 10
minutes. The concentration of RNA in each sample was quantified using a NanoDrop
spectrophotometer (Thermo-Fischer Scientific), normalized by addition of RNAse-free (DEPC
treated) water using RNAse free pipette tips. The concentrations were then checked again using
the NanoDrop. The SuperScript-3 (LifeTechnologies) reverse transcriptase kit was used
according to manufacturer’s protocols to synthesize cDNA using random oligos provided. For
the semi-quantitative PCR, equal amounts of each cDNA stock were distributed carefully to PCR

183
tubes. A master-mix of PCR reagents (not including primers) was made and distributed evenly
among the wells. Primer mixes were made as pools for each PCR reaction, and equal amounts
distributed to the appropriate wells. PCR was performed using Taq polymerase (LifeTech) as
per manufacturer’s protocols. Equal amounts of each PCR reaction were then carefully loaded
into 2% agarose (BioRad) gel with ethidium bromide, electrophoresis was performed and the
resulting gel was imaged and photographed using UV illumination in an AlphaInnotech
AlphaImager gel imaging system. Printed gel images were then scanned at maximum dpi
greyscale using a Cannon scanner, then contrast adjusted uniformly across all wells in Adobe
Photoshop. Primers were custom designed and checked by alignment against each isoform of
NAP1L1 and NAP1L4 using ClustalW. Control GAPDH primers were designed by Dr. Mariana
Gomez-Ferreria, formerly of the Pelletier Lab. Primers used were:
NAP1L1 (both isoforms): Forward: 5’- GGTGAAGAAACAAAACTCAAAGC-3’
NAP1L1 (both isoforms): Reverse: 5’-CAATGTATCCTGTTGGTGTTTC-3’
NAP1L4 (both isoforms): Forward: 5’- TTCAGATGGGGTTCCTTCAG-3’
Nap1L4 (both isoforms) Reverse: 5’- GAGGGACATTGTCAAGTCGCT-3’
NAP1L1 (short isoform): Forward: 5’- CTTCGAAATTGGTCACTTTTTACG-3’
NAP1L1 (short isoform): Reverse: 5’- TCATTTGGTAAACATCACCTCATC-3’
GAPDH (Control): Forward: 5’- GAAGGTGAAGGTCGGAGTCA-3’
GAPDH (Control) Reverse: 5’-GTTAAAAGCAGCCCTGGTGA-3’

184
Table 3.5 Primary Antibodies Used in this Work.
Primary antibodies Source
Rabbit Anti-CEP120 (Xie et al., 2007)
Rabbit Anti-SPICE1 (Archinti et al., 2010)
Rabbit Anti-CENPJ ProteinTech Group, Inc. 11517-1-AP
Goat Anti-SASS6 Santa-Cruz Biotechnology sc-82360
Rabbit Anti-SASS6 Pelletier Lab
Rabbit Anti-STIL Bethyl Laboratories A302-441A
Rabbit Anti-CP110 Bethyl Laboratories A301-343A-1
Rabbit Anti-CEP152 Bethyl Laboratories A302-480A-1
Mouse Anti-CETN2 Salisbury Laboratory, Mayo Clinic
Mouse Anti-Acetylated tubulin Sigma T 6793
Goat Anti-MYC Abcam Ab19234
Rabbit Anti-GAPDH Sigma G9545
Rabbit Anti-PCNT Abcam Ab4448
Rabbit Anti-FLAG Sigma F7425
Rat Anti--tubulin Serotec MCA77G
Mouse Anti-FLAG Sigma F3165
Mouse Anti-Glutamylated Tub.
(GT335) Gift of Dr. C. Janke,(Bobinnec et al., 1998a)
Mouse -tubulin Sigma T6199
Rabbit anti-CEP135 (Bird and Hyman, 2008)

185
Table 3.6 Secondary Antibodies Used for Microscopy in this Work.
Fluorescent antibodies Source
Donkey Anti-Rabbit Alexa 488 Invitrogen
Donkey Anti-Rabbit Alexa 594 Invitrogen
Donkey Anti-Rabbit Alexa 568 Invitrogen
Donkey Anti-Rabbit Alexa 647 Invitrogen
Donkey Anti-Mouse Alexa 488 Invitrogen
Donkey Anti-Mouse Alexa 594 Invitrogen
Donkey Anti-Mouse Alexa 568 Invitrogen
Donkey Anti-Mouse Alexa 647 Invitrogen
Donkey Anti-Mouse Dylight 405 Jackson ImmunoResearch
Donkey Anti-Goat Alexa 488 Invitrogen
Donkey Anti-Goat Alexa 594 Invitrogen
Donkey Anti-Goat Dylight 647 Jackson ImmunoResearch
Donkey Anti-Rat Dylight 647 Jackson ImmunoResearch

186
Table 3.7 Sequences of Small Interfering RNA Sequences Used in this Work.
Gene Target Sequence (5`-3`) Company, Catalogue Number
Control AUGAACGUGAAUUGCUCAA GE-Dharmacon D-001210-04
CEP135 SMARTpool GE-Dharmacon M-014548-01
SPICE1-si1* GCUGAGAACAAAUGAGUCA Archinti et al, 2010.
SPICE1-si2 GAAAGCGGAGCCACAGGUA GE-Dharmacon siGenome d-017060-03
CEP120-si1* GAUGAGAACGGGUGUGUAU GE-Dharmacon siGenome d-016493-01
CEP120-si2 AAACCGAGCGACAAGAAUU GE-Dharmacon siGenome d-016493-04
STIL SMARTpool GE-Dharmacon M-019717-01
CEP152 SMARTpool GE-Dharmacon M-022241-01
NAP1L1-si18 UAACCAUAGUUCAUCGAAA GE-Dharmacon D-017274-18
NAP1L1-si17* GCGUAUAAUCCCAAGAUCA GE-Dharmacon D-017274-17
NAP1L1-si2 GACCAGAAAUUAUGGGUUG GE-Dharmacon D-017274-02
NAP1L1-si1* GGUAGAAACACCAACAGGAUACAUU LifeTech HSS106946
NAP1L1-si2 AAUGUAUCCUGUUGGUGUUUCUACC LifeTech HSS106946
NAP1L1-si3 CCUGCAAGUUAAAUGUGCACAGAUA LifeTech HSS106946
NAP1L1-New ACCTGCAAGTTAAATGTGCACAGAT (Okuwaki et al., 2010)
CSPP1-si1 GGAAAGGACUAGACAUUGA GE-Dhamacon D-015485-01
CSPP1-si2 GAACGAAUGCGAAGACUGA GE-Dhamacon D-015485-02
CSPP1-si3* AGACAUAUCCUGCCAUUGA GE-Dhamacon D-015485-03
CSPP1-si4 CAUCCCAAGUGCUAAAGUA GE-Dhamacon D-015485-04
MTUS1 SMARTpool GE-Dharmacon M-006848-00
KIAA1731 SMARTpool GE-Dharmacon M-029941-01
MAP9 SMARTpool GE-Dharmacon M-014470-01
MAP7D3-si5* UUACAGGCGUCACCAAUUA GE-Dharmacon J-024948-05
MAP7D3-si6 CUACAUUCGUCUACUGAUA GE-Dharmacon J-024948-06
MAP7D3-si7 UAAGAGUACUGCAGGUAUU GE-Dharmacon J-024948-07
MAP7D3-si8 GAAAGACCAAGCUCCAAUA GE-Dharmacon J-024948-08
NAP1L4 UCAGGUGAUGCAGAAUCCUCGAGUU LifeTech HSS106954
MAP7 SMARTpool GE-Dharmacon M-011563-00
ANK2-si5* GCAUGUAGCAGCCAAGUAU GE-Dharmacon D-0084187-05
ANK2-si2 GAGCAAAUGUUAACGCAAA GE-Dharmacon D-0084187-02
ANK2-si3 GAAAUUAUCUGCGUACCUU GE-Dharmacon D-0084187-03
ANK2-si4 GAAAUUGAACAGACCAUUA GE-Dharmacon D-0084187-04

187
Table 3.7 Sequences of Small Interfering RNA Sequences Used in this Work.
Gene Target Sequence (5`-3`) Company, Catalogue Number
CENTROBIN AGUGCCAGACUGCAGCAACTT (Zou et al., 2005)
POC1 Proprietary Qiagen Flexitube siRNA S103097696
hPOC5 SMARTpool GE-Dharmacon L-016919-02
Table 3.7 Sequences of Small Interfering RNA Sequences Used in this Work. For genes targeted with multiple siRNAs, the * indicates siRNAs used in follow-up experiments.

188
Table 3.8 Primers for esiRNA Used in this Study.
Gene Esi-Forward Primer (5’-3’) Esi-Reverse Primer (5’-3’)
CPAP atgttcctgatgccaaccctc gctccaactgttcggtcttc
SASS6 aagatgttggacagtctcttcg cagggtggctggtatttttg
CEP97 tgccagatatcccaggaaag attgttggctcaggacaagg
CP110 cgagctgccttgtacggtat aagtcttgggcagggaaaat
KIAA1731 esi1 gatgaagcagttaagctgaagg gttctgccgctgataaatatcttg
KIAA1731 esi2 gtttatcacatcttgttttacctcaac tgagggaagagtatcattacttttgg
MTUS1 esi1 tccctgcatgtgtcaacattg cagcagggaacaacacaaga
MTUS1 esi2 tttaaaaggaattgaagaagaaataaaatggcag cctacaagtgcaggacact
MAP7 esi1 aacaaacgctccgggaaatg ttaaatttcagctttggttcttcagaag
MAP7esi2 tttaatccaggcatatggtctttagta aaacagccaaaatacacctactgc
CSPP1 esi1 ttttgcacagttttgtcataaattaggg atacaagtaattacagttcccttcatttc
CSPP1 esi2 ttttgatagacggtatcatagaccag tctgttgctcagccatttgttc
KIAA0753 esi1 acaggtgctgttgagcatga gggctttcgctgtgtttact
KIAA0753 esi2 cagaaccagctgcagtttaatag gacaggctgcttgactctt
CCDC138 esi1 tggactggatttcggatcatc atgcctgagccaggtaatca
CCDC138 esi2 tttatcagtctaagtataagagaagaactc ctgacaggtttcaaaggagag
ZC2HC1A esi1 atactcgaaataccatttccagttaattttg taggcaccaataatacaaatgtttatcc
ZC2HC1A esi2 tttgtttatgattctgatcaactataagacac aaaacagtgctcagtattagataaaacaag
ANK2 esi2 gtgggaagatattacaggaactac ccacttttagttaatggtaccaagttg
CCDC77 esi1 cccgttatgcatgagagtca tatgtgcccgtctattgctg
CCDC77 esi2 gaagatcgtctggccaaac taggtcacttctccagcatct
KIAA0586 esi atcagaagaatcagagaaccaagaaag tacttgggctgaacaaatattgctg
Table 3.8. Primers for esiRNA Used in this study. Note that the forward primers have a non-
target specific lead in sequence of (5’-3’) TCACTATAGGGAGAG, and the reverse primers
have a non-target-specific lead in sequence of (5’-3’) TCACTATAGGGAGAC. These have
been omitted from the above table to show only the gene-specific primer complementary
sequence.

189
Table 3.9 Plasmids Used in this study.
Gene Backbone Source
CENPJ PCDNA5/FRT/TO, PeGFP-N1 Generous gift of Dr. T.K. Tang
SPICE1 PCS2, PCDNA5/FRT/TO,PeGFP-N1 (Archinti et al., 2010)
CEP120 PCS2, PCDNA5,Pegfp-N1,PCDNA5-FLAG/FRT/TO-hBirA* This study.
NAP1L1 pEGFP-C1, pEGFP-N1 This study
USP37 PCDNA5/FRT/TO This study.

190
4.
Chapter IV: Conclusion and Future Directions

191
4.1. The Functions of CEP120 and SPICE1 in
Procentriole Assembly and Elongation
At the outset of my project, CEP120 and SPICE1 were proteins without known roles in centriole
duplication, though CEP120 had been shown to be important in neurons for interkinetic nuclear
migration in mouse neurons, a microtubule dependent process (Xie et al., 2007). My work here
showed for the first time that CEP120 was essential for centriole duplication as part of a
collaboration describing the CEP120/SPICE interaction (Hutchins et al., 2010). Essential roles
for CEP120 and SPICE1 in centriole duplication were subsequently confirmed (Archinti et al.,
2010; Mahjoub et al., 2010). I further characterized CEP120 and SPICE1 function in centriole
duplication, showing that they were required for CEP135 incorporation, as well as CP110/CEP97
and Centrin incorporation into procentrioles. Procentrioles assembled in the absence of SPICE1,
CEP120 or CPAP have shorter than normal microtubules when measured in tomograms. I
showed that CEP120 overexpression induced centriole elongation in G2, further supporting a
role in procentriole microtubule elongation. CPAP and CEP120 cooperated in this process with
SPICE1, as depletion of any of these blocked CEP120 or CPAP induced centriole elongation.
Further, we showed that CEP120 and CPAP interact in vivo. Overall, these results represented
the first characterization of how CEP120 and SPICE1 cooperate with CPAP in procentriole
assembly and elongation. Highlighting the importance of CEP120, mutations have been found in
patients with Jeune asphyxiating thoracic dystrophy (JATD) (Shaheen et al., 2014b). The
mutation in CEP120 identified in these patients leads to shorter cilia and fewer centrioles in
model systems, and recapitulates the JATD patient phenotypes in zebrafish development
(Shaheen et al., 2014b). In addition, mutations in CEP135 and CPAP have been found in
patients with microcephaly (Faheem et al., 2015).
Many questions remain as to how centriole assembly and elongation are achieved, and as to how
exactly CEP120/SPICE1 function in these processes. Specifically of interest is the question of
how exactly tubulin is incorporated into growing procentrioles, since my results are consistent
with the hypothesis that it is a chaperoned process. How do CEP120/SPICE1, along with other
factors like CPAP and Centrobin, cooperate to include tubulin under the cap of CP110 and
CEP97? This will be an important avenue of investigation. Future work should focus on testing
whether CEP120 binds free tubulin dimers or microtubules, whether SPICE1 binds either, and

192
whether nucleotide status of those tubulin dimers is important as it is for SAS-4 (Gopalakrishnan
et al., 2012). How CEP120, SPICE1 and CPAP interactions relate to tubulin binding will be
important to study in vitro. Attempting to drive the elongation of purified centrioles in vitro by
incubation with tubulin and combinations of recombinant CEP120, SPICE1, CPAP, Centrobin
and CEP135 would be a challenging but potentially powerful experimental approach to gain
insights into how centrioles are elongated by these factors. Recently, purified Drosophila
centrioles stripped of PCM have been used with recombinant SAS-4 and tubulin to recapitulate
PCM assembly onto centrioles in vitro, illustrating the potential of such experiments
(Gopalakrishnan et al., 2012). Understanding how CEP120/SPICE1 interact with microtubules
and tubulin in vitro, then determining how they cooperate with other factors to elongate
centrioles in vitro, would provide answers to many of the questions posed in this thesis and
significantly advance our understanding of centriole elongation. To understand the relationship
between CEP120, SPICE1 and CEP135/CP110/CEP97 incorporation during procentriole
assembly, efforts should also include looking for interactions between CEP135, CP110, CEP97
and CEP120/SPICE1. This would be achieved by co-IP experiments immunoprecipitating
epitope-tagged CEP135/CP110/CEP97 proteins, and blotting for CEP120 and SPICE1 within the
immunoprecipitate. Lastly, SPICE1 is important for mitotic spindle formation, and mitosis is
critical for genome stability. Whether CEP120 might cooperate with SPICE1 in regulating
spindle assembly and genome stability, is not well studied and worth examining. The impact (if
any) of CEP120 on spindle-assembly should be examined, for example by looking at mitotic
cells with 4 centrioles following CEP120 depletion as was previously done for SPICE1 (Archinti
et al., 2010). A potential problem with that approach is that younger centrioles could be shorter
in those mitotic cells, so to overcome this the duration of RNAi before fixation could be reduced,
or the cells could be arrested throughout the RNAi to prevent centriole duplication under
CEP120 depleted conditions.
Whether CEP120 and SPICE1 have a microtubule stabilizing activity in addition to aiding
tubulin incorporation is also an important question. In the third chapter of this thesis, I show that
depletion of a variety of proteins found in the proximity of CEP120/SPICE1 leads to CEP120 re-
localization to cytoplasmic microtubules, and the abnormal glutamylation of cytoplasmic
microtubules. How the enzymes responsible for glutamylation localize to the centrosome is not
well studied, nor are the mechanisms that regulate centriole glutamylation (Janke and Bulinski,

193
2011). From these results, I conclude that along with CEP120, tubulin glutamylation enzymes
(TTLLs) are localized to microtubules when the proteins studied here are depleted. Whether this
is direct, or a result of CEP120 increasing microtubule stability leading to long-lived cytoplasmic
microtubules that accumulate PTMs, remains to be tested. Whether microtubules become
stabilized following CEP120/SPICE1 over-expression should be investigated, for example by
exposing cells to cold-shock or colchicine treatment to see if microtubules are resistant to
depolymerization. Whether CEP120 binds glutamylated microtubules following TTLL
overexpression should also be tested in vivo. Can overexpression of CEP120 or SPICE1 drive
abnormal glutamylation of the cytoplasmic microtubules? Are CEP120 and/or SPICE1 required
for the glutamylation of cytoplasmic microtubules observed following depletion of proteins such
as ANK2 (see Figure 3.5)? Are CEP120 and/or SPICE1 required for the glutamylation of
cytoplasmic microtubules observed following over-expression of TTLL enzymes? These are
important experimentally tractable questions that should be addressed using combinations of
protein over-expression and depletion in tissue cultures, with analysis by microscopy.
4.2. Identification of CEP120 and SPICE1 Associated
Proteins with Uncharacterized Roles in Centriole
Assembly
The discovery of novel proteins involved in centriole duplication is a particularly heavy area of
research in centrosome biology, as identifying and characterizing such proteins leads to a more
complete picture of the critical processes of centriole assembly and elongation. Using a
proximity dependent biotinylation assay, we identified proteins that come into close proximity
with CEP120 and SPICE1 in human cells (Roux et al., 2012). From this list of proteins, I
showed that several were important for PLK4 induced centriole overduplication. I identified for
the first time roles for NAP1L1, ANK2, MTUS1, CCDC77, CSPP1, KIAA0586, MAP7D3,
MAP7 and MAP9 in centriole duplication in this assay. These proteins will require extensive
further validation as proteins involved in this process, beginning importantly with testing their
requirement for centriole duplication in non-PLK4 overexpression systems. Each of these
proteins represents a potential CEP120 or SPICE1 interacting protein, and as such could yield

194
insights into the mechanisms by which CEP120/SPICE1 function. Interactions with these
proteins should be tested by expressing them as fusions to epitope tags, and using
immunoprecipitation followed by Western blots to look for co-purified CEP120 and SPICE1 (co-
IPs). Notably, MTUS1 is a tumour suppressor protein that is under-expressed in cancer, and is
required for CEP120 and SPICE1 localization and centriole elongation (Rodrigues-Ferreira et al.,
2009; Yu et al., 2009; Zuern et al., 2010). Thus I have here identified a potential function of an
important tumour suppressor protein in centriole duplication. ANK2 similarly affects both
CEP120 and SPICE1 localization, and may negatively regulate centriole elongation. Future
work should focus on how MTUS1 and ANK2 impact centriole assembly, and whether each
interacts with CEP120 or SPICE1 to facilitate this function. How CEP120 or SPICE1 function
in MTUS1-deficient cancer tissues from patients would also be a particularly interesting and
important question to address.
Several of the proteins identified in my screens as important for centriole duplication have
effects on CEP120 localization, microtubule glutamylation, or cause apparent centriole
elongation. It is important to consider the possibility that these phenotypes could be the result of
a general stress response. Centriolar satellites are molecular complexes containing centriolar and
ciliary proteins, with roles in regulating both centriole duplication and cilia formation (Bärenz et
al., 2011; Kubo, 2003; Kubo et al., 1999; Prosser et al., 2009; Tollenaere et al., 2015). A variety
of cellular stresses such as heat shock and DNA damage impact the function of centriolar
satellites (Löffler et al., 2012; Villumsen et al., 2013). One of the responses to stress is a loss of
AZI1 and PCM1 from satellites, initiated by either p38 activity or inactivation of MIB1, which
can promote ciliogenesis (Villumsen et al., 2013). Several satellite proteins are found in the
BioID prey list for CEP120 and/or SPICE1, including AZI1 and PCM1. It is possible then that
the elongated structures observed following depletion of ANK2, MAP7, CSPP1 or KIAA0586
are actually cilia formed as a result of stress or satellite disruption. To address this possibility,
future work should include staining for cilia-specific proteins such as Arl13b, and ultra-structural
analysis of the structures by electron microscopy (Cevik et al., 2010; Duldulao et al., 2009). The
other phenotypes, such as abnormal glutamylation of cytoplasmic microtubules or CEP120 mis-
localization could also be non-specific. To determine whether stress responses underlie the
phenotypes of depletion of the CEP120/SPICE1 BioID preys, future work should include
simultaneous inhibition of p38 (a stress response kinase) in these assays (Duch et al., 2012;

195
Villumsen et al., 2013). The cell cycle profiles of cells under each RNAi condition should be
analyzed as well, to reveal possible arrests. Most importantly, it should be determined whether
experimentally stressing U-2 OS cells with UV radiation or heat-shock directly causes interphase
mis-localization of CEP120 or SPICE1, or microtubule glutamylation (Villumsen et al., 2013).
Lastly, testing the remaining BioID preys in the PLK4 assay using an siRNA library would be of
particular interest, as yet more novel roles for proteins in centriole duplication are no doubt
waiting there to be discovered. In sum, by identifying CEP120/SPICE1 proximate proteins, we
have generated a set of proteins with potential roles in centriole duplication worthy of further
study.
4.3. A Role for NAP1L1 in Centriole Duplication
The human nucleosome-assembly-like-proteins play important roles in chromatin structure, with
important implications for transcription, and NAP1L1 plays a role in some cancers (Kidd et al.,
2006; Park and Luger, 2006a; Schimmack et al., 2014). We show that NAP1L1, through
depletion with multiple siRNA in multiple assays, is required for centriole duplication. This is a
previously unreported role for NAP1L1, and a link between NAP1L1 and the centrosome is
novel. It will be important to see how the changes in NAP1L1 expression levels reported in
cancer would impact centriole duplication when introduced into non-cancerous cells such as
RPE-1 (Kidd et al., 2006; Park and Luger, 2006a; Schimmack et al., 2014). Understanding how
NAP1L1 functions in centriole duplication is going to be a critical task. Firstly, does NAP1L1
interact with CEP120 or SPICE1 directly? The results of the BioID experiments described
herein suggest that CEP120 and SPICE1 may have nuclear localizations. First, both CEP120 and
SPICE1 are associated with nuclear proteins including NAP1L1, NAP1L4, Histones, Importins,
and INO80B. Second, an amino-terminal fragment of SPICE1 is restricted to the nucleus when
exogenously expressed as GFP-fusion in human cells (Archinti et al., 2010). Third, SPICE1 is
spindle associated in mitosis and plays a role in faithful mitotic chromosome segregation
independent of centriole number (Archinti et al., 2010). The possibility that CEP120 and
SPICE1 shuttle to and from the nucleus should be tested, for example by treatment of cells with
Leptomycin-B to inhibit nuclear export (Kudo et al., 1999), or careful biochemical isolation of
nuclei followed by Western blots for CEP120 and SPICE1. Might the roles of CEP120/SPICE1
in centriole duplication and mitotic spindle assembly be regulated by interactions with NAP1L1?

196
This is an exciting possibility that clearly warrants further investigation. It will be important to
test whether NAP1L1 interacts directly with CEP120 and SPICE1, and to determine where this
interaction occurs within the cell (nuclear, cytoplasmic or both?). This can be achieved by co-IP
experiments using cytoplasmic and nuclear extracts separately. Whether CEP120/SPICE1 are
associated with nucleosomes and DNA should be tested. This can be achieved by chemical
cross-linking of DNA to proteins and subsequent immunoprecipitation from nuclear extracts
using antibodies against histone proteins (i.e. chromatin immunoprecipitation) (Solomon et al.,
1988). These extracts would then be analyzed by Western blot for CEP120 and SPICE1.
Alternatively, chemical cross-linking of proteins to DNA, extraction of nuclei and
immunoprecipitation using antibodies against CEP120/SPICE1 could be attempted. These
extracts could be analyzed for DNA content using ethidium bromide gel electrophoresis, or
tested for the presence of histones or NAP1L1 by Western blot. How does changing the cellular
levels of CEP120/SPICE1 affect NAP1L1 function in transcription, or its localization? One of
the NAP1L1 siRNAs tested here affected CEP120 localization, supportive of a functional link
between the two proteins. This effect, however, was not reversed by the presence of GFP-
NAP1L1. Future work should address these issues, and focus on validating the possible
functional relationship between NAP1L1, CEP120 and SPICE1. Importantly, we report here a
novel link between a cancer-associated transcription regulating histone chaperone and two
proteins involved directly in centriole assembly. In sum, the results described within this thesis
represent significant advances in the field of centrosome biology and centriole duplication, and
open up several new research directions warranting further investigation.

197
5. LITERATURE CITED
Abal, M., Keryer, G., and Bornens, M. (2005). Centrioles resist forces applied on centrosomes
during G2/M transition. Biol. Cell 97, 425–434.
Agircan, F.G., Schiebel, E., and Mardin, B.R. (2014). Separate to operate: control of centrosome
positioning and separation. Philos. Trans. R. Soc. B Biol. Sci. 369, 20130461–20130461.
Ahmed, K., Li, R., and Bazett-Jones, D.P. (2008). Electron spectroscopic imaging of the nuclear
landscape. In The Nucleus, (Springer), pp. 415–423.
Akey, C.W., and Luger, K. (2003). Histone chaperones and nucleosome assembly. Curr. Opin.
Struct. Biol. 13, 6–14.
Akizu, N., Silhavy, J.L., Rosti, R.O., Scott, E., Fenstermaker, A.G., Schroth, J., Zaki, M.S.,
Sanchez, H., Gupta, N., Kabra, M., et al. (2014). Mutations in CSPP1 Lead to Classical Joubert
Syndrome. Am. J. Hum. Genet. 94, 80–86.
Alby, C., Piquand, K., Huber, C., Megarbané, A., Ichkou, A., Legendre, M., Pelluard, F., Encha-
Ravazi, F., Abi-Tayeh, G., Bessières, B., et al. (2015). Mutations in KIAA0586 Cause Lethal
Ciliopathies Ranging from a Hydrolethalus Phenotype to Short-Rib Polydactyly Syndrome. Am.
J. Hum. Genet. 97, 311–318.
Amos, L.A., and Klug, A. (1974). Arrangement of subunits in flagellar microtubules. J. Cell Sci.
14, 523–549.
de Anda, F.C., Pollarolo, G., Da Silva, J.S., Camoletto, P.G., Feiguin, F., and Dotti, C.G. (2005).
Centrosome localization determines neuronal polarity. Nature 436, 704–708.
Andersen, J.S., Wilkinson, C.J., Mayor, T., Mortensen, P., Nigg, E.A., and Mann, M. (2003).
Proteomic characterization of the human centrosome by protein correlation profiling. Nature
426, 570–574.
D’Angiolella, V., Donato, V., Vijayakumar, S., Saraf, A., Florens, L., Washburn, M.P.,
Dynlacht, B., and Pagano, M. (2010). SCFCyclin F controls centrosome homeostasis and mitotic
fidelity through CP110 degradation. Nature 466, 138–142.
Archinti, M., Lacasa, C., Teixido-Travesa, N., and Luders, J. (2010). SPICE - a previously
uncharacterized protein required for centriole duplication and mitotic chromosome congression.
J. Cell Sci. 123, 3039–3046.
Arnal, I., and Wade, R.H. (1995). How does taxol stabilize microtubules? Curr. Biol. 5, 900–
908.
Arquint, C., and Nigg, E.A. (2014). STIL Microcephaly Mutations Interfere with APC/C-
Mediated Degradation and Cause Centriole Amplification. Curr. Biol. 24, 351–360.

198
Arquint, C., Sonnen, K.F., Stierhof, Y.-D., and Nigg, E.A. (2012). Cell-cycle-regulated
expression of STIL controls centriole number in human cells. J. Cell Sci. 125, 1342–1352.
Arquint, C., Gabryjonczyk, A.-M., and Nigg, E.A. (2014). Centrosomes as signalling centres.
Philos. Trans. R. Soc. B Biol. Sci. 369, 20130464–20130464.
Asiedu, M., Wu, D., Matsumura, F., and Wei, Q. (2009). Centrosome/Spindle Pole–associated
Protein Regulates Cytokinesis via Promoting the Recruitment of MyoGEF to the Central
Spindle. Mol. Biol. Cell 20, 1428–1440.
Askham, J.M., Vaughan, K.T., Goodson, H.V., and Morrison, E.E. (2002). Evidence that an
interaction between EB1 and p150Glued is required for the formation and maintenance of a
radial microtubule array anchored at the centrosome. Mol. Biol. Cell 13, 3627–3645.
Attia, M., Förster, A., Rachez, C., Freemont, P., Avner, P., and Rogner, U.C. (2011). Interaction
between Nucleosome Assembly Protein 1-like Family Members. J. Mol. Biol. 407, 647–660.
Ayalon, G., Davis, J.Q., Scotland, P.B., and Bennett, V. (2008). An Ankyrin-Based Mechanism
for Functional Organization of Dystrophin and Dystroglycan. Cell 135, 1189–1200.
Azimzadeh, J. (2014). Exploring the evolutionary history of centrosomes. Philos. Trans. R. Soc.
B Biol. Sci. 369, 20130453–20130453.
Azimzadeh, J., and Marshall, W.F. (2010). Building the Centriole. Curr. Biol. 20, R816–R825.
Azimzadeh, J., Hergert, P., Delouvee, A., Euteneuer, U., Formstecher, E., Khodjakov, A., and
Bornens, M. (2009). hPOC5 is a centrin-binding protein required for assembly of full-length
centrioles. J. Cell Biol. 185, 101–114.
Azimzadeh, J., Wong, M.L., Downhour, D.M., Alvarado, A.S., and Marshall, W.F. (2012).
Centrosome Loss in the Evolution of Planarians. Science 335, 461–463.
Bahe, S. (2005). Rootletin forms centriole-associated filaments and functions in centrosome
cohesion. J. Cell Biol. 171, 27–33.
Bahtz, R., Seidler, J., Arnold, M., Haselmann-Weiss, U., Antony, C., Lehmann, W.D., and
Hoffmann, I. (2012). GCP6 is a substrate of Plk4 and required for centriole duplication. J. Cell
Sci. 125, 486–496.
Balczon, R., Bao, L., Zimmer, W.E., Brown, K., Zinkowski, R.P., and Brinkley, B.R. (1995).
Dissociation of centrosome replication events from cycles of DNA synthesis and mitotic division
in hydroxyurea-arrested Chinese hamster ovary cells. J. Cell Biol. 130, 105–115.
Bangs, F., Antonio, N., Thongnuek, P., Welten, M., Davey, M.G., Briscoe, J., and Tickle, C.
(2011). Generation of mice with functional inactivation of talpid3, a gene first identified in
chicken. Development 138, 3261–3272.

199
Barbelanne, M., and Tsang, W.Y. (2014). Molecular and Cellular Basis of Autosomal Recessive
Primary Microcephaly. BioMed Res. Int. 2014, 1–13.
Bärenz, F., Mayilo, D., and Gruss, O.J. (2011). Centriolar satellites: Busy orbits around the
centrosome. Eur. J. Cell Biol. 90, 983–989.
Barrera, J.A., Kao, L.-R., Hammer, R.E., Seemann, J., Fuchs, J.L., and Megraw, T.L. (2010).
CDK5RAP2 Regulates Centriole Engagement and Cohesion in Mice. Dev. Cell 18, 913–926.
Basto, R., Lau, J., Vinogradova, T., Gardiol, A., Woods, C.G., Khodjakov, A., and Raff, J.W.
(2006). Flies without Centrioles. Cell 125, 1375–1386.
Basto, R., Brunk, K., Vinadogrova, T., Peel, N., Franz, A., Khodjakov, A., and Raff, J.W.
(2008). Centrosome Amplification Can Initiate Tumorigenesis in Flies. Cell 133, 1032–1042.
Bayless, B.A., Giddings, T.H., Winey, M., and Pearson, C.G. (2012). Bld10/Cep135 stabilizes
basal bodies to resist cilia-generated forces. Mol. Biol. Cell 23, 4820–4832.
Bazzi, H., and Anderson, K.V. (2014). Acentriolar mitosis activates a p53-dependent apoptosis
pathway in the mouse embryo. Proc. Natl. Acad. Sci. 111, E1491–E1500.
Bernhard, W., and De Harven, E. (1956). [Presence in certain mammalian cells of an organoid
probably of centriole nature; electron microscopy]. Comptes Rendus Hebd. Séances Académie
Sci. 242, 288–290.
Besson, A., Dowdy, S.F., and Roberts, J.M. (2008). CDK Inhibitors: Cell Cycle Regulators and
Beyond. Dev. Cell 14, 159–169.
Bettencourt-Dias, M., Rodrigues-Martins, A., Carpenter, L., Riparbelli, M., Lehmann, L., Gatt,
M.K., Carmo, N., Balloux, F., Callaini, G., and Glover, D.M. (2005). SAK/PLK4 Is Required for
Centriole Duplication and Flagella Development. Curr. Biol. 15, 2199–2207.
Bettencourt-Dias, M., Hildebrandt, F., Pellman, D., Woods, G., and Godinho, S.A. (2011).
Centrosomes and cilia in human disease. Trends Genet. 27, 307–315.
Bird, A.W., and Hyman, A.A. (2008). Building a spindle of the correct length in human cells
requires the interaction between TPX2 and Aurora A. J. Cell Biol. 182, 289–300.
Blachon, S., Gopalakrishnan, J., Omori, Y., Polyanovsky, A., Church, A., Nicastro, D., Malicki,
J., and Avidor-Reiss, T. (2008). Drosophila asterless and Vertebrate Cep152 Are Orthologs
Essential for Centriole Duplication. Genetics 180, 2081–2094.
Blum, M., and Vick, P. (2015). Left-right asymmetry: cilia and calcium revisited. Curr. Biol. CB
25, R205–R207.
Blum, M., Schweickert, A., Vick, P., Wright, C.V.E., and Danilchik, M.V. (2014). Symmetry
breakage in the vertebrate embryo: When does it happen and how does it work? Dev. Biol. 393,
109–123.

200
Bobinnec, Y., Khodjakov, A., Mir, L.M., Rieder, C.L., Edde, B., and Bornens, M. (1998a).
Centriole disassembly in vivo and its effect on centrosome structure and function in vertebrate
cells. J. Cell Biol. 143, 1575–1589.
Bobinnec, Y., Moudjou, M., Fouquet, J.P., Desbruyeres, E., Edde, B., and Bornens, M. (1998b).
Glutamylation of Centriole and Cytoplasmic Tubulin in Proliferating Non-neuronal Cells. Cell
Motil. Cytoskeleton 39, 223–232.
Bogoyevitch, M.A., Yeap, Y.Y.C., Qu, Z., Ngoei, K.R., Yip, Y.Y., Zhao, T.T., Heng, J.I., and
Ng, D.C.H. (2012). WD40-repeat protein 62 is a JNK-phosphorylated spindle pole protein
required for spindle maintenance and timely mitotic progression. J. Cell Sci. 125, 5096–5109.
Boisvieux-Ulrich, E., Laine, M.C., and Sandoz, D. (1989). In vitro effects of colchicine and
nocodazole on ciliogenesis in qual oviduct. Biol. Cell 67, 67–79.
Bornens, M., Paintrand, M., Berges, J., Marty, M.-C., and Karsenti, E. (1987). Structural and
chemical characterization of isolated centrosomes. Cell Motil. Cytoskeleton 8, 238–249.
Bouckson-Castaing, V., Moudjou, M., Ferguson, D.J., Mucklow, S., Belkaid, Y., Milon, G., and
Crocker, P.R. (1996). Molecular characterisation of ninein, a new coiled-coil protein of the
centrosome. J. Cell Sci. 109, 179–190.
Boveri, T. (2008). Concerning the origin of malignant tumours by Theodor Boveri. Translated
and annotated by Henry Harris. J. Cell Sci. 121 Suppl 1, 1–84.
van Breugel, M., Hirono, M., Andreeva, A., Yanagisawa, H. -a., Yamaguchi, S., Nakazawa, Y.,
Morgner, N., Petrovich, M., Ebong, I.-O., Robinson, C.V., et al. (2011). Structures of SAS-6
Suggest Its Organization in Centrioles. Science 331, 1196–1199.
Brito, D.A., Gouveia, S.M., and Bettencourt-Dias, M. (2012). Deconstructing the centriole:
structure and number control. Curr. Opin. Cell Biol. 24, 4–13.
Brown, N.J., Marjanović, M., Lüders, J., Stracker, T.H., and Costanzo, V. (2013). Cep63 and
Cep152 Cooperate to Ensure Centriole Duplication. PLoS ONE 8, e69986.
Brownlee, C.W., Klebba, J.E., Buster, D.W., and Rogers, G.C. (2011). The Protein Phosphatase
2A regulatory subunit Twins stabilizes Plk4 to induce centriole amplification. J. Cell Biol. 195,
231–243.
Bulinski, J.C., Odde, D.J., Howell, B.J., Salmon, T.D., and Waterman-Storer, C.M. (2001).
Rapid dynamics of the microtubule binding of ensconsin in vivo. J. Cell Sci. 114, 3885–3897.
Burgess, R.J., and Zhang, Z. (2010). Histones, histone chaperones and nucleosome assembly.
Protein Cell 1, 607–612.
Burridge, K., and Wennerberg, K. (2004). Rho and Rac take center stage. Cell 116, 167–179.

201
Cajanek, L., Glatter, T., and Nigg, E.A. (2015). The E3 ubiquitin ligase Mib1 regulates Plk4 and
centriole biogenesis. J. Cell Sci.
Cardozo, T., and Pagano, M. (2004). The SCF ubiquitin ligase: insights into a molecular
machine. Nat. Rev. Mol. Cell Biol. 5, 739–751.
Carvalho-Santos, Z., Machado, P., Branco, P., Tavares-Cadete, F., Rodrigues-Martins, A.,
Pereira-Leal, J.B., and Bettencourt-Dias, M. (2010). Stepwise evolution of the centriole-
assembly pathway. J. Cell Sci. 123, 1414–1426.
Carvalho-Santos, Z., Azimzadeh, J., Pereira-Leal, J.B., and Bettencourt-Dias, M. (2011).
Evolution: Tracing the origins of centrioles, cilia, and flagella. J. Cell Biol. 194, 165–175.
Carvalho-Santos, Z., Machado, P., Alvarez-Martins, I., Gouveia, S.M., Jana, S.C., Duarte, P.,
Amado, T., Branco, P., Freitas, M.C., Silva, S.T.N., et al. (2012). BLD10/CEP135 Is a
Microtubule-Associated Protein that Controls the Formation of the Flagellum Central
Microtubule Pair. Dev. Cell 23, 412–424.
Caussinus, E., and Gonzalez, C. (2005). Induction of tumor growth by altered stem-cell
asymmetric division in Drosophila melanogaster. Nat. Genet. 37, 1125–1129.
Cautain, B., Hill, R., de Pedro, N., and Link, W. (2015). Components and regulation of nuclear
transport processes. FEBS J. 282, 445–462.
Cevik, S., Hori, Y., Kaplan, O.I., Kida, K., Toivenon, T., Foley-Fisher, C., Cottell, D., Katada,
T., Kontani, K., and Blacque, O.E. (2010). Joubert syndrome Arl13b functions at ciliary
membranes and stabilizes protein transport in Caenorhabditis elegans. J. Cell Biol. 188, 953–
969.
Chan, J.Y. (2011). A Clinical Overview of Centrosome Amplification in Human Cancers. Int. J.
Biol. Sci. 7, 1122–1144.
Chang, J., Cizmecioglu, O., Hoffmann, I., and Rhee, K. (2010). PLK2 phosphorylation is critical
for CPAP function in procentriole formation during the centrosome cycle. EMBO J. 29, 2395–
2406.
Chang, J., Seo, S., Lee, K., Nagashima, K., Bang, J., Kim, B., Erikson, R.L., Lee, K.-W., Lee,
H., Park, J.-E., et al. (2013). Essential role of Cenexin1, but not Odf2, in ciliogenesis. Cell Cycle
12, 655–662.
Chang, P., Giddings, T.H., Winey, M., and Stearns, T. (2003). Epsilon-tubulin is required for
centriole duplication and microtubule organization. Nat. Cell Biol. 5, 71–76.
Chavali, P.L., Putz, M., and Gergely, F. (2014). Small organelle, big responsibility: the role of
centrosomes in development and disease. Philos. Trans. R. Soc. B Biol. Sci. 369, 20130468–
20130468.

202
Cheeseman, I.M., and Desai, A. (2005). A combined approach for the localization and tandem
affinity purification of protein complexes from metazoans. Sci STKE 2005, pl1.
Chen, A.L., Kim, E.W., Toh, J.Y., Vashisht, A.A., Rashoff, A.Q., Van, C., Huang, A.S., Moon,
A.S., Bell, H.N., Bentolila, L.A., et al. (2015). Novel Components of the Toxoplasma Inner
Membrane Complex Revealed by BioID. mBio 6, e02357–14.
Chen, J.-F., Zhang, Y., Wilde, J., Hansen, K.C., Lai, F., and Niswander, L. (2014). Microcephaly
disease gene Wdr62 regulates mitotic progression of embryonic neural stem cells and brain size.
Nat. Commun. 5.
Chen, Z., Indjeian, V.B., McManus, M., Wang, L., and Dynlacht, B.D. (2002). CP110, a cell
cycle-dependent CDK substrate, regulates centrosome duplication in human cells. Dev. Cell 3,
339–350.
Choi, H., Larsen, B., Lin, Z.-Y., Breitkreutz, A., Mellacheruvu, D., Fermin, D., Qin, Z.S., Tyers,
M., Gingras, A.-C., and Nesvizhskii, A.I. (2011). SAINT: probabilistic scoring of affinity
purification–mass spectrometry data. Nat. Methods 8, 70–73.
Choi, H., Liu, G., Mellacheruvu, D., Tyers, M., Gingras, A.-C., and Nesvizhskii, A.I. (2012).
Analyzing Protein-Protein Interactions from Affinity Purification-Mass Spectrometry Data with
SAINT. In Current Protocols in Bioinformatics, A.D. Baxevanis, G.A. Petsko, L.D. Stein, and
G.D. Stormo, eds. (Hoboken, NJ, USA: John Wiley & Sons, Inc.),.
Choi, M.C., Raviv, U., Miller, H.P., Gaylord, M.R., Kiris, E., Ventimiglia, D., Needleman, D.J.,
Kim, M.W., Wilson, L., Feinstein, S.C., et al. (2009). Human Microtubule-Associated-Protein
Tau Regulates the Number of Protofilaments in Microtubules: A Synchrotron X-Ray Scattering
Study. Biophys. J. 97, 519–527.
Choi-Rhee, E., Schulman, H., and Cronan, J.E. (2004). Promiscuous protein biotinylation by
Escherichia coli biotin protein ligase. Protein Sci. 13, 3043–3050.
Chrétien, D., Buendia, B., Fuller, S.D., and Karsenti, E. (1997). Reconstruction of the
centrosome cycle from cryoelectron micrographs. J. Struct. Biol. 120, 117–133.
Ciciarello, M. (2001). p53 Displacement from Centrosomes and p53-mediated G1 Arrest
following Transient Inhibition of the Mitotic Spindle. J. Biol. Chem. 276, 19205–19213.
Cizmecioglu, O., Arnold, M., Bahtz, R., Settele, F., Ehret, L., Haselmann-Weiss, U., Antony, C.,
and Hoffmann, I. (2010). Cep152 acts as a scaffold for recruitment of Plk4 and CPAP to the
centrosome. J. Cell Biol. 191, 731–739.
Le Clech, M. (2008). Role of CAP350 in Centriolar Tubule Stability and Centriole Assembly.
PLoS ONE 3, e3855.
Comartin, D., Gupta, G.D., Fussner, E., Coyaud, É., Hasegan, M., Archinti, M., Cheung, S.W.T.,
Pinchev, D., Lawo, S., Raught, B., et al. (2013). CEP120 and SPICE1 Cooperate with CPAP in
Centriole Elongation. Curr. Biol. 23, 1360–1366.

203
Conaway, R.C., and Conaway, J.W. (2009). The INO80 chromatin remodeling complex in
transcription, replication and repair. Trends Biochem. Sci. 34, 71–77.
Conduit, P.T., and Raff, J.W. (2010). Cnn Dynamics Drive Centrosome Size Asymmetry to
Ensure Daughter Centriole Retention in Drosophila Neuroblasts. Curr. Biol. 20, 2187–2192.
Conduit, P.T., Brunk, K., Dobbelaere, J., Dix, C.I., Lucas, E.P., and Raff, J.W. (2010). Centrioles
Regulate Centrosome Size by Controlling the Rate of Cnn Incorporation into the PCM. Curr.
Biol. 20, 2178–2186.
Cottee, M.A., Muschalik, N., Wong, Y.L., Johnson, C.M., Johnson, S., Andreeva, A., Oegema,
K., Lea, S.M., Raff, J.W., and van Breugel, M. (2013). Crystal structures of the CPAP/STIL
complex reveal its role in centriole assembly and human microcephaly. Elife 2.
Craig, R., and Beavis, R.C. (2004). TANDEM: matching proteins with tandem mass spectra.
Bioinformatics 20, 1466–1467.
Cunha, S.R., Hund, T.J., Hashemi, S., Voigt, N., Li, N., Wright, P., Koval, O., Li, J.,
Gudmundsson, H., Gumina, R.J., et al. (2011). Defects in Ankyrin-Based Membrane Protein
Targeting Pathways Underlie Atrial Fibrillation. Circulation 124, 1212–1222.
Cunha-Ferreira, I., Rodrigues-Martins, A., Bento, I., Riparbelli, M., Zhang, W., Laue, E.,
Callaini, G., Glover, D.M., and Bettencourt-Dias, M. (2009). The SCF/Slimb Ubiquitin Ligase
Limits Centrosome Amplification through Degradation of SAK/PLK4. Curr. Biol. 19, 43–49.
Cunha-Ferreira, I., Bento, I., Pimenta-Marques, A., Jana, S.C., Lince-Faria, M., Duarte, P.,
Borrego-Pinto, J., Gilberto, S., Amado, T., Brito, D., et al. (2013). Regulation of
Autophosphorylation Controls PLK4 Self-Destruction and Centriole Number. Curr. Biol. 23,
2245–2254.
Dammermann, A. (2002). Assembly of centrosomal proteins and microtubule organization
depends on PCM-1. J. Cell Biol. 159, 255–266.
David, A., Liu, F., Tibelius, A., Vulprecht, J., Wald, D., Rothermel, U., Ohana, R., Seitel, A.,
Metzger, J., Ashery-Padan, R., et al. (2014). Lack of centrioles and primary cilia in STIL −/−
mouse embryos. Cell Cycle 13, 2859–2868.
Debec, A., Sullivan, W., and Bettencourt-Dias, M. (2010). Centrioles: active players or
passengers during mitosis? Cell. Mol. Life Sci. 67, 2173–2194.
El-Deiry, W. (1993). WAF1, a potential mediator of p53 tumor suppression. Cell 75, 817–825.
Delattre, M., Leidel, S., Wani, K., Baumer, K., Bamat, J., Schnabel, H., Feichtinger, R.,
Schnabel, R., and Gönczy, P. (2004). Centriolar SAS-5 is required for centrosome duplication in
C. elegans. Nat. Cell Biol. 6, 656–664.
Delattre, M., Canard, C., and Gönczy, P. (2006). Sequential Protein Recruitment in C. elegans
Centriole Formation. Curr. Biol. 16, 1844–1849.

204
Delgehyr, N. (2005). Microtubule nucleation and anchoring at the centrosome are independent
processes linked by ninein function. J. Cell Sci. 118, 1565–1575.
Deutsch, E.W., Mendoza, L., Shteynberg, D., Farrah, T., Lam, H., Tasman, N., Sun, Z., Nilsson,
E., Pratt, B., Prazen, B., et al. (2010). A guided tour of the Trans-Proteomic Pipeline.
PROTEOMICS 10, 1150–1159.
van Dijk, J., Rogowski, K., Miro, J., Lacroix, B., Eddé, B., and Janke, C. (2007). A Targeted
Multienzyme Mechanism for Selective Microtubule Polyglutamylation. Mol. Cell 26, 437–448.
van Dijk, J., Miro, J., Strub, J.-M., Lacroix, B., van Dorsselaer, A., Edde, B., and Janke, C.
(2008). Polyglutamylation Is a Post-translational Modification with a Broad Range of Substrates.
J. Biol. Chem. 283, 3915–3922.
Dingar, D., Kalkat, M., Chan, P.-K., Srikumar, T., Bailey, S.D., Tu, W.B., Coyaud, E., Ponzielli,
R., Kolyar, M., Jurisica, I., et al. (2014). BioID identifies novel c-MYC interacting partners in
cultured cells and xenograft tumors. J. Proteomics.
Donehower, L.A., Harvey, M., Slagle, B.L., McArthur, M.J., Montgomery, C.A., Butel, J.S., and
Bradley, A. (1992). Mice deficient for p53 are developmentally normal but susceptible to
spontaneous tumours. Nature 356, 215–221.
Doudna, J.A., and Charpentier, E. (2014). The new frontier of genome engineering with
CRISPR-Cas9. Science 346, 1258096–1258096.
Duch, A., de Nadal, E., and Posas, F. (2012). The p38 and Hog1 SAPKs control cell cycle
progression in response to environmental stresses. FEBS Lett. 586, 2925–2931.
Duldulao, N.A., Lee, S., and Sun, Z. (2009). Cilia localization is essential for in vivo functions of
the Joubert syndrome protein Arl13b/Scorpion. Development 136, 4033–4042.
Dumont, J., and Desai, A. (2012). Acentrosomal spindle assembly and chromosome segregation
during oocyte meiosis. Trends Cell Biol. 22, 241–249.
Dzhindzhev, N.S., Yu, Q.D., Weiskopf, K., Tzolovsky, G., Cunha-Ferreira, I., Riparbelli, M.,
Rodrigues-Martins, A., Bettencourt-Dias, M., Callaini, G., and Glover, D.M. (2010). Asterless is
a scaffold for the onset of centriole assembly. Nature 467, 714–718.
Dzhindzhev, N.S., Tzolovsky, G., Lipinszki, Z., Schneider, S., Lattao, R., Fu, J., Debski, J.,
Dadlez, M., and Glover, D.M. (2014). Plk4 Phosphorylates Ana2 to Trigger Sas6 Recruitment
and Procentriole Formation. Curr. Biol. 24, 2526–2532.
Eot-Houllier, G., Venoux, M., Vidal-Eychenie, S., Hoang, M.-T., Giorgi, D., and Rouquier, S.
(2010). Plk1 Regulates Both ASAP Localization and Its Role in Spindle Pole Integrity. J. Biol.
Chem. 285, 29556–29568.

205
Faheem, M., Naseer, M.I., Rasool, M., Chaudhary, A.G., Kumosani, T.A., Ilyas, A.M.,
Pushparaj, P.N., Ahmed, F., Algahtani, H.A., Al-Qahtani, M.H., et al. (2015). Molecular genetics
of human primary microcephaly: an overview. BMC Med. Genomics 8, S4.
Finley, D. (2009). Recognition and Processing of Ubiquitin-Protein Conjugates by the
Proteasome. Annu. Rev. Biochem. 78, 477–513.
Firat-Karalar, E.N., and Stearns, T. (2014). The centriole duplication cycle. Philos. Trans. R.
Soc. B Biol. Sci. 369, 20130460–20130460.
Firat-Karalar, E.N., Rauniyar, N., Yates, J.R., and Stearns, T. (2014). Proximity Interactions
among Centrosome Components Identify Regulators of Centriole Duplication. Curr. Biol. 24,
664–670.
Fong, C.S., Kim, M., Yang, T.T., Liao, J.-C., and Tsou, M.-F.B. (2014). SAS-6 Assembly
Templated by the Lumen of Cartwheel-less Centrioles Precedes Centriole Duplication. Dev.
Cell.
Fong, K.-W., Choi, Y.-K., Rattner, J.B., and Qi, R.Z. (2008). CDK5RAP2 is a pericentriolar
protein that functions in centrosomal attachment of the γ-tubulin ring complex. Mol. Biol. Cell
19, 115–125.
Fridman, J.S., and Lowe, S.W. (2003). Control of apoptosis by p53. Oncogene 22, 9030–9040.
Friedländer, M., and Wahrman, J. (1970). The spindle as a basal body distributor a study in the
meiosis of the male silkworm moth, Bombyx mori. J. Cell Sci. 7, 65–89.
Fry, A.M., Mayor, T., Meraldi, P., Stierhof, Y.-D., Tanaka, K., and Nigg, E.A. (1998). C-Nap1, a
novel centrosomal coiled-coil protein and candidate substrate of the cell cycle–regulated protein
kinase Nek2. J. Cell Biol. 141, 1563–1574.
Fu, J., and Glover, D.M. (2012). Structured illumination of the interface between centriole and
peri-centriolar material. Open Biol. 2, 120104–120104.
Fu, J., Hagan, I.M., and Glover, D.M. (2015). The Centrosome and Its Duplication Cycle. Cold
Spring Harb. Perspect. Biol. 7, a015800.
Fukasawa, K. (2005). Centrosome amplification, chromosome instability and cancer
development. Cancer Lett. 230, 6–19.
Fukasawa, K. (2008). p53, cyclin-dependent kinase and abnormal amplification of centrosomes.
Biochim. Biophys. Acta BBA - Rev. Cancer 1786, 15–23.
Fukasawa, K., Wiener, F., Vande Woude, G.F., and Mai, S. (1997). Genomic instability and
apoptosis are frequent in p53 deficient young mice. Oncogene 15, 1295–1302.

206
Fussner, E., Strauss, M., Djuric, U., Li, R., Ahmed, K., Hart, M., Ellis, J., and Bazett-Jones, D.P.
(2012). Open and closed domains in the mouse genome are configured as 10-nm chromatin
fibres. EMBO Rep. 13, 992–996.
Gallaud, E., Caous, R., Pascal, A., Bazile, F., Gagne, J.-P., Huet, S., Poirier, G.G., Chretien, D.,
Richard-Parpaillon, L., and Giet, R. (2014). Ensconsin/Map7 promotes microtubule growth and
centrosome separation in Drosophila neural stem cells. J. Cell Biol. 204, 1111–1121.
Ganem, N.J., Godinho, S.A., and Pellman, D. (2009). A mechanism linking extra centrosomes to
chromosomal instability. Nature 460, 278–282.
Gilbert, L.A., Horlbeck, M.A., Adamson, B., Villalta, J.E., Chen, Y., Whitehead, E.H.,
Guimaraes, C., Panning, B., Ploegh, H.L., Bassik, M.C., et al. (2014). Genome-Scale CRISPR-
Mediated Control of Gene Repression and Activation. Cell 159, 647–661.
Godinho, S.A., and Pellman, D. (2014). Causes and consequences of centrosome abnormalities
in cancer. Philos. Trans. R. Soc. B Biol. Sci. 369, 20130467–20130467.
Godinho, S.A., Picone, R., Burute, M., Dagher, R., Su, Y., Leung, C.T., Polyak, K., Brugge, J.S.,
Théry, M., and Pellman, D. (2014). Oncogene-like induction of cellular invasion from
centrosome amplification. Nature 510, 167–171.
Goetz, S.C., and Anderson, K.V. (2010). The primary cilium: a signalling centre during
vertebrate development. Nat. Rev. Genet. 11, 331–344.
Gomez-Ferreria, M.A., Rath, U., Buster, D.W., Chanda, S.K., Caldwell, J.S., Rines, D.R., and
Sharp, D.J. (2007). Human Cep192 Is Required for Mitotic Centrosome and Spindle Assembly.
Curr. Biol. 17, 1960–1966.
Gönczy, P. (2012). Towards a molecular architecture of centriole assembly. Nat. Rev. Mol. Cell
Biol. 13, 425–435.
Gopalakrishnan, J., Mennella, V., Blachon, S., Zhai, B., Smith, A.H., Megraw, T.L., Nicastro,
D., Gygi, S.P., Agard, D.A., and Avidor-Reiss, T. (2011). Sas-4 provides a scaffold for
cytoplasmic complexes and tethers them in a centrosome. Nat. Commun. 2, 359.
Gopalakrishnan, J., Frederick Chim, Y.-C., Ha, A., Basiri, M.L., Lerit, D.A., Rusan, N.M., and
Avidor-Reiss, T. (2012). Tubulin nucleotide status controls Sas-4-dependent pericentriolar
material recruitment. Nat. Cell Biol. 14, 865–873.
Gould, R.R., and Borisy, G.R. (1977). The Pericentriolar Material in Chinese Hamster Ovary
Cells Nucleates Microtubule Formation. J. Cell Biol. 73, 601–615.
Graser, S., Stierhof, Y.-D., Lavoie, S.B., Gassner, O.S., Lamla, S., Le Clech, M., and Nigg, E.A.
(2007). Cep164, a novel centriole appendage protein required for primary cilium formation. J.
Cell Biol. 179, 321–330.

207
Gregan, J., Polakova, S., Zhang, L., Tolić-Nørrelykke, I.M., and Cimini, D. (2011). Merotelic
kinetochore attachment: causes and effects. Trends Cell Biol. 21, 374–381.
Gromley, A. (2003). A novel human protein of the maternal centriole is required for the final
stages of cytokinesis and entry into S phase. J. Cell Biol. 161, 535–545.
Gromley, A., Yeaman, C., Rosa, J., Redick, S., Chen, C.-T., Mirabelle, S., Guha, M., Sillibourne,
J., and Doxsey, S.J. (2005). Centriolin Anchoring of Exocyst and SNARE Complexes at the
Midbody Is Required for Secretory-Vesicle-Mediated Abscission. Cell 123, 75–87.
Guarguaglini, G., Duncan, P.I., Stierhof, Y.D., Holmström, T., Duensing, S., and Nigg, E.A.
(2005). The forkhead-associated domain protein Cep170 interacts with Polo-like kinase 1 and
serves as a marker for mature centrioles. Mol. Biol. Cell 16, 1095–1107.
Guderian, G., Westendorf, J., Uldschmid, A., and Nigg, E.A. (2010). Plk4 trans-
autophosphorylation regulates centriole number by controlling TrCP-mediated degradation. J.
Cell Sci. 123, 2163–2169.
Gudi, R., Zou, C., Li, J., and Gao, Q. (2011). Centrobin-tubulin interaction is required for
centriole elongation and stability. J. Cell Biol. 193, 711–725.
Gudi, R., Zou, C., Dhar, J., Gao, Q., and Vasu, C. (2014). Centrobin-Centrosomal Protein 4.1-
associated Protein (CPAP) Interaction Promotes CPAP Localization to the Centrioles during
Centriole Duplication. J. Biol. Chem. 289, 15166–15178.
Gudi, R., Haycraft, C.J., Bell, P.D., Li, Z., and Vasu, C. (2015). Centrobin-mediated regulation
of CPAP level limits centriole length during elongation stage. J. Biol. Chem. jbc.M114.603423.
Guichard, P., Chrétien, D., Marco, S., and Tassin, A.-M. (2010). Procentriole assembly revealed
by cryo-electron tomography. EMBO J. 29, 1565–1572.
Guichard, P., Desfosses, A., Maheshwari, A., Hachet, V., Dietrich, C., Brune, A., Ishikawa, T.,
Sachse, C., and Gönczy, P. (2012). Cartwheel architecture of Trichonympha basal body. Science
337, 553.
Guichard, P., Hachet, V., Majubu, N., Neves, A., Demurtas, D., Olieric, N., Fluckiger, I.,
Yamada, A., Kihara, K., Nishida, Y., et al. (2013). Native Architecture of the Centriole Proximal
Region Reveals Features Underlying Its 9-Fold Radial Symmetry. Curr. Biol. 23, 1620–1628.
Gul, A., Hassan, M.J., Hussain, S., Raza, S.I., Chishti, M.S., and Ahmad, W. (2006). A novel
deletion mutation in CENPJ gene in a Pakistani family with autosomal recessive primary
microcephaly. J. Hum. Genet. 51, 760–764.
Gustafsson, M.G. (2000). Surpassing the lateral resolution limit by a factor of two using
structured illumination microscopy. J. Microsc. 198, 82–87.

208
Gustafsson, M.G.L., Shao, L., Carlton, P.M., Wang, C.J.R., Golubovskaya, I.N., Cande, W.Z.,
Agard, D.A., and Sedat, J.W. (2008). Three-Dimensional Resolution Doubling in Wide-Field
Fluorescence Microscopy by Structured Illumination. Biophys. J. 94, 4957–4970.
Habedanck, R., Stierhof, Y.-D., Wilkinson, C.J., and Nigg, E.A. (2005). The Polo kinase Plk4
functions in centriole duplication. Nat. Cell Biol. 7, 1140–1146.
Hansford, S., and Huntsman, D.G. (2014). Boveri at 100: Theodor Boveri and Genetic
Predisposition To Cancer. J. Pathol. 234, 142–145.
Haren, L. (2006). NEDD1-dependent recruitment of the -tubulin ring complex to the centrosome
is necessary for centriole duplication and spindle assembly. J. Cell Biol. 172, 505–515.
Haren, L., Stearns, T., and Lüders, J. (2009). Plk1-Dependent Recruitment of γ-Tubulin
Complexes to Mitotic Centrosomes Involves Multiple PCM Components. PLoS ONE 4, e5976.
Hatch, E.M., Kulukian, A., Holland, A.J., Cleveland, D.W., and Stearns, T. (2010). Cep152
interacts with Plk4 and is required for centriole duplication. J. Cell Biol. 191, 721–729.
Hauf, S., Waizenegger, I.C., and Peters, J.-M. (2001). Cohesin cleavage by separase required for
anaphase and cytokinesis in human cells. Science 293, 1320–1323.
He, R., Huang, N., Bao, Y., Zhou, H., Teng, J., and Chen, J. (2013). LRRC45 Is a Centrosome
Linker Component Required for Centrosome Cohesion. Cell Rep. 4, 1100–1107.
Hehnly, H., Chen, C.-T., Powers, C.M., Liu, H.-L., and Doxsey, S. (2012). The Centrosome
Regulates the Rab11- Dependent Recycling Endosome Pathway at Appendages of the Mother
Centriole. Curr. Biol. 22, 1944–1950.
Helps, N., Luo, X., Barker, H., and Cohen, P. (2000). NIMA-related kinase 2 (Nek2), a cell-
cycle-regulated protein kinase localized to centrosomes, is complexed to protein phosphatase 1.
Biochem J 349, 509–518.
Hiraki, M., Nakazawa, Y., Kamiya, R., and Hirono, M. (2007). Bld10p Constitutes the
Cartwheel-Spoke Tip and Stabilizes the 9-Fold Symmetry of the Centriole. Curr. Biol. 17, 1778–
1783.
Hirono, M. (2014). Cartwheel assembly. Philos. Trans. R. Soc. B Biol. Sci. 369, 20130458–
20130458.
Hoover, A.N., Wynkoop, A., Zeng, H., Jia, J., Niswander, L.A., and Liu, A. (2008). C2cd3 is
required for cilia formation and Hedgehog signaling in mouse. Development 135, 4049–4058.
Hsu, W.-B., Hung, L.-Y., Tang, C.-J.C., Su, C.-L., Chang, Y., and Tang, T.K. (2008). Functional
characterization of the microtubule-binding and -destabilizing domains of CPAP and d-SAS-4.
Exp. Cell Res. 314, 2591–2602.

209
Huang, B., Bates, M., and Zhuang, X. (2009). Super-Resolution Fluorescence Microscopy.
Annu. Rev. Biochem. 78, 993–1016.
Huang, B., Babcock, H., and Zhuang, X. (2010). Breaking the Diffraction Barrier: Super-
Resolution Imaging of Cells. Cell 143, 1047–1058.
Huangfu, D., Liu, A., Rakeman, A.S., Murcia, N.S., Niswander, L., and Anderson, K.V. (2003).
Hedgehog signalling in the mouse requires intraflagellar transport proteins. Nature 426, 83–87.
Hung, L.-Y., Chen, H.-L., Chang, C.-W., Li, B.-R., and Tang, T.K. (2004). Identification of a
novel microtubule-destabilizing motif in CPAP that binds to tubulin heterodimers and inhibits
microtubule assembly. Mol. Biol. Cell 15, 2697–2706.
Hurtado, L., Caballero, C., Gavilan, M.P., Cardenas, J., Bornens, M., and Rios, R.M. (2011).
Disconnecting the Golgi ribbon from the centrosome prevents directional cell migration and
ciliogenesis. J. Cell Biol. 193, 917–933.
Hutchins, J.R.A., Toyoda, Y., Hegemann, B., Poser, I., Heriche, J.K., Sykora, M.M., Augsburg,
M., Hudecz, O., Buschhorn, B.A., Bulkescher, J., et al. (2010). Systematic Analysis of Human
Protein Complexes Identifies Chromosome Segregation Proteins. Science 328, 593–599.
Ibrahim, R., Messaoudi, C., Chichon, F.J., Celati, C., and Marco, S. (2009). Electron tomography
study of isolated human centrioles. Microsc. Res. Tech. 72, 42–48.
Insolera, R., Bazzi, H., Shao, W., Anderson, K.V., and Shi, S.-H. (2014). Cortical neurogenesis
in the absence of centrioles. Nat. Neurosci. 17, 1528–1535.
Ishikawa, H., and Marshall, W.F. (2011). Ciliogenesis: building the cell’s antenna. Nat. Rev.
Mol. Cell Biol. 12, 222–234.
Ishikawa, H., Kubo, A., Tsukita, S., and Tsukita, S. (2005). Odf2-deficient mother centrioles
lack distal/subdistal appendages and the ability to generate primary cilia. Nat. Cell Biol. 7, 517–
524.
Izquierdo, D., Wang, W.-J., Uryu, K., and Tsou, M.-F.B. (2014). Stabilization of Cartwheel-less
Centrioles for Duplication Requires CEP295-Mediated Centriole-to-Centrosome Conversion.
Cell Rep. 8, 957–965.
Izraeli, S., Lowe, L.A., Bertness, V.L., Good, D.J., Dorward, D.W., Kirsch, I.R., and Kuehn,
M.R. (1999). The SIL gene is required for mouse embryonic axial development and left-right
specification. Nature 399, 691–694.
Jakobsen, L., Vanselow, K., Skogs, M., Toyoda, Y., Lundberg, E., Poser, I., Falkenby, L.G.,
Bennetzen, M., Westendorf, J., Nigg, E.A., et al. (2011). Novel asymmetrically localizing
components of human centrosomes identified by complementary proteomics methods. EMBO J.
30, 1520–1535.

210
Jana, S.C., Marteil, G., and Bettencourt-Dias, M. (2014). Mapping molecules to structure:
unveiling secrets of centriole and cilia assembly with near-atomic resolution. Curr. Opin. Cell
Biol. 26, 96–106.
Janke, C., and Bulinski, J.C. (2011). Post-translational regulation of the microtubule
cytoskeleton: mechanisms and functions. Nat. Rev. Mol. Cell Biol. 12, 773–786.
Janke, C., and Kneussel, M. (2010). Tubulin post-translational modifications: encoding functions
on the neuronal microtubule cytoskeleton. Trends Neurosci. 33, 362–372.
Janke, C., Rogowski, K., Wloga, D., Regnard, C., Kajava, A.V., Strub, J.M., Temurak, N., van
Dijk, J., Boucher, D., van Dorsselaer, A., et al. (2005). Tubulin Polyglutamylase Enzymes Are
Members of The TTL Domain Protein Family. Science 308, 1758–1762.
Janssen, A., van de Burg, M., Szuhai, K., Kops, G.J.P.L., and Medema, R.H. (2011).
Chromosome Segregation Errors as a Cause of DNA Damage and Structural Chromosome
Aberrations. Science 333, 1895–1898.
Januschke, J., Reina, J., Llamazares, S., Bertran, T., Rossi, F., Roig, J., and Gonzalez, C. (2013).
Centrobin controls mother–daughter centriole asymmetry in Drosophila neuroblasts. Nat. Cell
Biol. 15, 241–248.
Joukov, V., De Nicolo, A., Rodriguez, A., Walter, J.C., and Livingston, D.M. (2010).
Centrosomal protein of 192 kDa (Cep192) promotes centrosome-driven spindle assembly by
engaging in organelle-specific Aurora A activation. Proc. Natl. Acad. Sci. 107, 21022–21027.
Joukov, V., Walter, J.C., and De Nicolo, A. (2014). The Cep192-Organized Aurora A-Plk1
Cascade Is Essential for Centrosome Cycle and Bipolar Spindle Assembly. Mol. Cell 55, 578–
591.
Kardon, J.R., and Vale, R.D. (2009). Regulators of the cytoplasmic dynein motor. Nat. Rev. Mol.
Cell Biol. 10, 854–865.
Keller, L.C., Geimer, S., Romijn, E., Yates, J., Zamora, I., and Marshall, W.F. (2009). Molecular
architecture of the centriole proteome: the conserved WD40 domain protein POC1 is required for
centriole duplication and length control. Mol. Biol. Cell 20, 1150–1166.
Kemp, C.A., Kopish, K.R., Zipperlen, P., Ahringer, J., and O’Connell, K.F. (2004). Centrosome
maturation and duplication in C. elegans require the coiled-coil protein SPD-2. Dev. Cell 6, 511–
523.
Keryer, G., Di Fiore, B., Celati, C., Lechtreck, K.F., Mogensen, M., Delouvée, A., Lavia, P.,
Bornens, M., and Tassin, A.-M. (2003). Part of Ran is associated with AKAP450 at the
centrosome: involvement in microtubule-organizing activity. Mol. Biol. Cell 14, 4260–4271.
Kessner, D., Chambers, M., Burke, R., Agus, D., and Mallick, P. (2008). ProteoWizard: open
source software for rapid proteomics tools development. Bioinformatics 24, 2534–2536.

211
Khodjakov, A., Cole, R.W., Oakley, B.R., and Rieder, C.L. (2000). Centrosome-independent
mitotic spindle formation in vertebrates. Curr. Biol. 10, 59–67.
Khorasanizadeh, S. (2004). The nucleosome: from genomic organization to genomic regulation.
Cell 116, 259–272.
Kidd, M., Modlin, I.M., Mane, S.M., Camp, R.L., Eick, G., and Latich, I. (2006). The Role of
Genetic Markers— NAP1L1, MAGE-D2, and MTA1—in Defining Small-Intestinal Carcinoid
Neoplasia. Ann. Surg. Oncol. 13, 253–262.
Kim, D.I., Kc, B., Zhu, W., Motamedchaboki, K., Doye, V., and Roux, K.J. (2014). Probing
nuclear pore complex architecture with proximity-dependent biotinylation. Proc. Natl. Acad. Sci.
111, E2453–E2461.
Kim, K., Lee, S., Chang, J., and Rhee, K. (2008). A novel function of CEP135 as a platform
protein of C-NAP1 for its centriolar localization. Exp. Cell Res. 314, 3692–3700.
Kim, T.-S., Park, J.-E., Shukla, A., Choi, S., Murugan, R.N., Lee, J.H., Ahn, M., Rhee, K., Bang,
J.K., Kim, B.Y., et al. (2013). Hierarchical recruitment of Plk4 and regulation of centriole
biogenesis by two centrosomal scaffolds, Cep192 and Cep152. Proc. Natl. Acad. Sci. 110,
E4849–E4857.
Kirkham, M., Müller-Reichert, T., Oegema, K., Grill, S., and Hyman, A.A. (2003). SAS-4 Is a<
i> C</i>.< i> elegans</i> Centriolar Protein that Controls Centrosome Size. Cell 112, 575–587.
Kishi, H., Nakagawa, K., Matsumoto, M., Suga, M., Ando, M., Taya, Y., and Yamaizumi, M.
(2001). Osmotic Shock Induces G1 Arrest through p53 Phosphorylation at Ser33 by Activated
p38MAPK without Phosphorylation at Ser15 and Ser20. J. Biol. Chem. 276, 39115–39122.
Kitagawa, D., Vakonakis, I., Olieric, N., Hilbert, M., Keller, D., Olieric, V., Bortfeld, M., Erat,
M.C., Flückiger, I., Gönczy, P., et al. (2011). Structural Basis of the 9-Fold Symmetry of
Centrioles. Cell 144, 364–375.
Kittler, R., Heninger, A.-K., Franke, K., Habermann, B., and Buchholz, F. (2005). Production of
endoribonuclease-prepared short interfering RNAs for gene silencing in mammalian cells. Nat.
Methods 2, 779–784.
Klebba, J.E., Buster, D.W., Nguyen, A.L., Swatkoski, S., Gucek, M., Rusan, N.M., and Rogers,
G.C. (2013). Polo-like Kinase 4 Autodestructs by Generating Its Slimb-Binding Phosphodegron.
Curr. Biol. 23, 2255–2261.
Kleylein-Sohn, J., Westendorf, J., Le Clech, M., Habedanck, R., Stierhof, Y.-D., and Nigg, E.A.
(2007). Plk4-Induced Centriole Biogenesis in Human Cells. Dev. Cell 13, 190–202.
Knorz, V.J., Spalluto, C., Lessard, M., Purvis, T.L., Adigun, F.F., Collin, G.B., Hanley, N.A.,
Wilson, D.I., and Hearn, T. (2010). Centriolar Association of ALMS1 and Likely Centrosomal
Functions of the ALMS Motif–containing Proteins C10orf90 and KIAA1731. Mol. Biol. Cell 21,
3617–3629.

212
Kobayashi, T., Tsang, W.Y., Li, J., Lane, W., and Dynlacht, B.D. (2011). Centriolar Kinesin
Kif24 Interacts with CP110 to Remodel Microtubules and Regulate Ciliogenesis. Cell 145, 914–
925.
Kobayashi, T., Kim, S., Lin, Y.-C., Inoue, T., and Dynlacht, B.D. (2014). The CP110-interacting
proteins Talpid3 and Cep290 play overlapping and distinct roles in cilia assembly. J. Cell Biol.
204, 215–229.
Kohlmaier, G., Loncarek, J., Meng, X., McEwen, B.F., Mogensen, M., Spektor, A., Dynlacht,
B.D., Khodjakov, A., and Gonczy, P. (2009). Overly long centrioles and defective cell division
upon excess of the SAS-4-related protein CPAP. Curr. Biol. 19, 1012–1018.
Kollman, J.M., Polka, J.K., Zelter, A., Davis, T.N., and Agard, D.A. (2010). Microtubule
nucleating γ-TuSC assembles structures with 13-fold microtubule-like symmetry. Nature 466,
879–882.
Kollman, J.M., Merdes, A., Mourey, L., and Agard, D.A. (2011). Microtubule nucleation by γ-
tubulin complexes. Nat. Rev. Mol. Cell Biol. 12, 709–721.
Kornberg, R.D., and Lorch, Y. (1999). Twenty-Five Years of the Nucleosome, Fundamental
Particle of the Eukaryote Chromosome. Cell 98, 285–294.
Korzeniewski, N., Cuevas, R., Duensing, A., and Duensing, S. (2010). Daughter centriole
elongation is controlled by proteolysis. Mol. Biol. Cell 21, 3942–3951.
Kraemer, N., Issa, L., Hauck, S.C.R., Mani, S., Ninnemann, O., and Kaindl, A.M. (2011).
What’s the hype about CDK5RAP2? Cell. Mol. Life Sci. 68, 1719–1736.
Kremer, J.R., Mastronarde, D.N., and McIntosh, J.R. (1996). Computer visualization of three-
dimensional image data using IMOD. J. Struct. Biol. 116, 71–76.
Kubo, A. (2003). Non-membranous granular organelle consisting of PCM-1: subcellular
distribution and cell-cycle-dependent assembly/disassembly. J. Cell Sci. 116, 919–928.
Kubo, A., Sasaki, H., Yuba-Kubo, A., Tsukita, S., and Shiina, N. (1999). Centriolar Satellites
Molecular Characterization, Atp-Dependent Movement toward Centrioles and Possible
Involvement in Ciliogenesis. J. Cell Biol. 147, 969–980.
Kudo, N., Matsumori, N., Taoka, H., Fujiwara, D., Schreiner, E.P., Wolff, B., Yoshida, M., and
Horinouchi, S. (1999). Leptomycin B inactivates CRM1/exportin 1 by covalent modification at a
cysteine residue in the central conserved region. Proc. Natl. Acad. Sci. U. S. A. 96, 9112–9117.
Kumar, N. (1981). Taxol-induced polymerization of purified tubulin. Mechanism of action. J.
Biol. Chem. 256, 10435–10441.
Kuriyama, R. (1982). Effect of colcemid on the centriole cycle in Chinese hamster ovary cells. J.
Cell Sci. 53, 155–171.

213
Kuriyama, R., and Borisy, G.G. (1981). Centriole cycle in Chinese hamster ovary cells as
determined by whole-mount electron microscopy. J. Cell Biol. 91, 814–821.
Kuriyama, R., Dasgupta, S., and Borisy, G.G. (1986). Independence of centriole formation and
initiation of DNA synthesis in Chinese hamster ovary cells. Cell Motil. Cytoskeleton 6, 355–362.
Lakadamyali, M., and Cosma, M.P. (2015). Advanced microscopy methods for visualizing
chromatin structure. FEBS Lett.
Lambert, J.-P., Tucholska, M., Go, C., Knight, J.D.R., and Gingras, A.-C. (2014). Proximity
biotinylation and affinity purification are complementary approaches for the interactome
mapping of chromatin-associated protein complexes. J. Proteomics.
Lane, H.A., and Nigg, E.A. (1996). Antibody microinjection reveals an essential role for human
polo-like kinase 1 (Plk1) in the functional maturation of mitotic centrosomes. J. Cell Biol. 135,
1701–1713.
Lawo, S., Hasegan, M., Gupta, G.D., and Pelletier, L. (2012). Subdiffraction imaging of
centrosomes reveals higher-order organizational features of pericentriolar material. Nat. Cell
Biol. 14, 1148–1158.
Lawson, C.D., and Burridge, K. (2014). The on-off relationship of Rho and Rac during integrin-
mediated adhesion and cell migration. Small GTPases 5, e27958.
Lechler, T., and Fuchs, E. (2005). Asymmetric cell divisions promote stratification and
differentiation of mammalian skin. Nature 437, 275–280.
Lee, J.D., and Anderson, K.V. (2008). Morphogenesis of the node and notochord: The cellular
basis for the establishment and maintenance of left-right asymmetry in the mouse. Dev. Dyn.
237, 3464–3476.
Lee, K., and Rhee, K. (2011). PLK1 phosphorylation of pericentrin initiates centrosome
maturation at the onset of mitosis. J. Cell Biol. 195, 1093–1101.
Lee, K., and Rhee, K. (2012). Separase-dependent cleavage of pericentrin B is necessary and
sufficient for centriole disengagement during mitosis. Cell Cycle 11, 2476–2485.
Leidel, S., and Gönczy, P. (2003). SAS-4 is essential for centrosome duplication in C. elegans
and is recruited to daughter centrioles once per cell cycle. Dev. Cell 4, 431–439.
Leidel, S., Delattre, M., Cerutti, L., Baumer, K., and Gönczy, P. (2005). SAS-6 defines a protein
family required for centrosome duplication in C. elegans and in human cells. Nat. Cell Biol. 7,
115–125.
Lettman, M.M., Wong, Y.L., Viscardi, V., Niessen, S., Chen, S., Shiau, A.K., Zhou, H., Desai,
A., and Oegema, K. (2013). Direct Binding of SAS-6 to ZYG-1 Recruits SAS-6 to the Mother
Centriole for Cartwheel Assembly. Dev. Cell 25, 284–298.

214
Li, J., D’Angiolella, V., Seeley, E.S., Kim, S., Kobayashi, T., Fu, W., Campos, E.I., Pagano, M.,
and Dynlacht, B.D. (2013). USP33 regulates centrosome biogenesis via deubiquitination of the
centriolar protein CP110. Nature 495, 255–259.
Li, S., Fernandez, J.-J., Marshall, W.F., and Agard, D.A. (2012). Three-dimensional structure of
basal body triplet revealed by electron cryo-tomography. EMBO J. 31, 552–562.
Lin, Y.-C., Chang, C.-W., Hsu, W.-B., Tang, C.-J.C., Lin, Y.-N., Chou, E.-J., Wu, C.-T., and
Tang, T.K. (2013a). Human microcephaly protein CEP135 binds to hSAS-6 and CPAP, and is
required for centriole assembly. EMBO J.
Lin, Y.-N., Wu, C.-T., Lin, Y.-C., Hsu, W.-B., Tang, C.-J.C., Chang, C.-W., and Tang, T.K.
(2013b). CEP120 interacts with CPAP and positively regulates centriole elongation. J. Cell Biol.
202, 211–219.
Lindemann, C.B., and Lesich, K.A. (2010). Flagellar and ciliary beating: the proven and the
possible. J. Cell Sci. 123, 519–528.
Löffler, H., Fechter, A., Liu, F.Y., Poppelreuther, S., and Krämer, A. (2012). DNA damage-
induced centrosome amplification occurs via excessive formation of centriolar satellites.
Oncogene 32, 2963–2972.
Lorenzo, D.N., Badea, A., Davis, J., Hostettler, J., He, J., Zhong, G., Zhuang, X., and Bennett, V.
(2014). A PIK3C3-Ankyrin-B-Dynactin pathway promotes axonal growth and multiorganelle
transport. J. Cell Biol. 207, 735–752.
Louie, R.K. (2004). Adenomatous polyposis coli and EB1 localize in close proximity of the
mother centriole and EB1 is a functional component of centrosomes. J. Cell Sci. 117, 1117–
1128.
Lüders, J., Patel, U.K., and Stearns, T. (2006). GCP-WD is a γ-tubulin targeting factor required
for centrosomal and chromatin-mediated microtubule nucleation. Nat. Cell Biol. 8, 137–147.
Magiera, M.M., and Janke, C. (2014). Post-translational modifications of tubulin. Curr. Biol. 24,
R351–R354.
Mahjoub, M.R., and Stearns, T. (2012). Supernumerary Centrosomes Nucleate Extra Cilia and
Compromise Primary Cilium Signaling. Curr. Biol. 22, 1628–1634.
Mahjoub, M.R., Xie, Z., and Stearns, T. (2010). Cep120 is asymmetrically localized to the
daughter centriole and is essential for centriole assembly. J. Cell Biol. 191, 331–346.
Manandhar, G. (2005). Centrosome Reduction During Gametogenesis and Its Significance. Biol.
Reprod. 72, 2–13.
Mardin, B.R., Lange, C., Baxter, J.E., Hardy, T., Scholz, S.R., Fry, A.M., and Schiebel, E.
(2010). Components of the Hippo pathway cooperate with Nek2 kinase to regulate centrosome
disjunction. Nat. Cell Biol. 12, 1166–1176.

215
Mardin, B.R., Agircan, F.G., Lange, C., and Schiebel, E. (2011). Plk1 Controls the Nek2A-PP1γ
Antagonism in Centrosome Disjunction. Curr. Biol. 21, 1145–1151.
Marthiens, V., Rujano, M.A., Pennetier, C., Tessier, S., Paul-Gilloteaux, P., and Basto, R.
(2013). Centrosome amplification causes microcephaly. Nat. Cell Biol. 15, 731–740.
Mastronarde, D.N. (2005). Automated electron microscope tomography using robust prediction
of specimen movements. J. Struct. Biol. 152, 36–51.
Matsuo, K., Ohsumi, K., Iwabuchi, M., Kawamata, T., Ono, Y., and Takahashi, M. (2012).
Kendrin Is a Novel Substrate for Separase Involved in the Licensing of Centriole Duplication.
Curr. Biol. 22, 915–921.
Matsuura, K. (2004). Bld10p, a novel protein essential for basal body assembly in
Chlamydomonas: localization to the cartwheel, the first ninefold symmetrical structure appearing
during assembly. J. Cell Biol. 165, 663–671.
Mayor, T., Stierhof, Y.-D., Tanaka, K., Fry, A.M., and Nigg, E.A. (2000). The centrosomal
protein C-Nap1 is required for cell cycle–regulated centrosome cohesion. J. Cell Biol. 151, 837–
846.
Mennella, V., Keszthelyi, B., McDonald, K.L., Chhun, B., Kan, F., Rogers, G.C., Huang, B., and
Agard, D.A. (2012). Subdiffraction-resolution fluorescence microscopy reveals a domain of the
centrosome critical for pericentriolar material organization. Nat. Cell Biol. 14, 1159–1168.
Mennella, V., Agard, D.A., Huang, B., and Pelletier, L. (2014). Amorphous no more:
subdiffraction view of the pericentriolar material architecture. Trends Cell Biol. 24, 188–197.
Meraldi, P., and Nigg, E.A. (2001). Centrosome cohesion is regulated by a balance of kinase and
phosphatase activities. J. Cell Sci. 114, 3749–3757.
Meurer-Grob, P., Kasparian, J., and Wade, R.H. (2001). Microtubule Structure at Improved
Resolution †. Biochemistry (Mosc.) 40, 8000–8008.
Mikule, K., Delaval, B., Kaldis, P., Jurcyzk, A., Hergert, P., and Doxsey, S. (2007). Loss of
centrosome integrity induces p38—p53—p21-dependent G1—S arrest. Nat. Cell Biol. 9, 160–
170.
Miller, K.E., and Heald, R. (2015). Glutamylation of Nap1 modulates histone H1 dynamics and
chromosome condensation in Xenopus. J. Cell Biol.
Mogensen, M.M., Malik, A., Piel, M., Bouckson-Castaing, V., and Bornens, M. (2000).
Microtubule minus-end anchorage at centrosomal and non-centrosomal sites: the role of ninein.
J. Cell Sci. 113, 3013–3023.
Mohler, P.J., Schott, J.J., Gramolini, A.O., Dilly, K.W., Guatimosim, S., duBell, W.H., Song,
L.S., Haurogne, K., Kyndt, F., Ali, M.E., et al. (2003). Ankyrin-B Mutation Causes Type 4 Long
QT Cardiac Arrhythmia and Sudden Cardiac Death. Nature 421, 634–638.

216
Morrison, A.J., and Shen, X. (2009). Chromatin remodelling beyond transcription: the INO80
and SWR1 complexes. Nat. Rev. Mol. Cell Biol. 10, 373–384.
Morrison, S.J., and Kimble, J. (2006). Asymmetric and symmetric stem-cell divisions in
development and cancer. Nature 441, 1068–1074.
Mottier-Pavie, V., and Megraw, T.L. (2009). Drosophila bld10 is a centriolar protein that
regulates centriole, basal body, and motile cilium assembly. Mol. Biol. Cell 20, 2605–2614.
Nakazawa, Y., Hiraki, M., Kamiya, R., and Hirono, M. (2007). SAS-6 is a Cartwheel Protein
that Establishes the 9-Fold Symmetry of the Centriole. Curr. Biol. 17, 2169–2174.
Nigg, E.A. (2002). Centrosome Aberrations: Cause or Consequence of Cancer Progression? Nat.
Rev. Cancer 2, 815–825.
Nigg, E.A., and Raff, J.W. (2009). Centrioles, Centrosomes, and Cilia in Health and Disease.
Cell 139, 663–678.
Nigg, E.A., Čajánek, L., and Arquint, C. (2014). The centrosome duplication cycle in health and
disease. FEBS Lett. 588, 2366–2372.
Nogales, E., Whittaker, M., Milligan, R.A., and Downing, K.H. (1999). High-resolution model
of the microtubule. Cell 96, 79–88.
Nonaka, S., Tanaka, Y., Okada, Y., Takeda, S., Harada, A., Kanai, Y., Kido, M., and Hirokawa,
N. (1998). Randomization of left–right asymmetry due to loss of nodal cilia generating leftward
flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell 95, 829–837.
Nonaka, S., Shiratori, H., Saijoh, Y., and Hamada, H. (2002). Determination of left-right
patterning of the mouse embryo by artificial nodal flow. Nature 418, 96–99.
O’Connell, K.F., Caron, C., Kopish, K.R., Hurd, D.D., Kemphues, K.J., Li, Y., and White, J.G.
(2001). The C. elegans zyg-1 gene encodes a regulator of centrosome duplication with distinct
maternal and paternal roles in the embryo. Cell 105, 547–558.
Oddone, A., Vilanova, I.V., Tam, J., and Lakadamyali, M. (2014). Super-resolution imaging
with stochastic single-molecule localization: Concepts, technical developments, and biological
applications: Super-Resolution Imaging with Single-Molecule Localization. Microsc. Res. Tech.
77, 502–509.
Ohkura, H. (2015). Meiosis: An Overview of Key Differences from Mitosis. Cold Spring Harb.
Perspect. Biol. a015859.
Ohta, T. (2002). Characterization of Cep135, a novel coiled-coil centrosomal protein involved in
microtubule organization in mammalian cells. J. Cell Biol. 156, 87–100.

217
Ohta, M., Ashikawa, T., Nozaki, Y., Kozuka-Hata, H., Goto, H., Inagaki, M., Oyama, M., and
Kitagawa, D. (2014). Direct interaction of Plk4 with STIL ensures formation of a single
procentriole per parental centriole. Nat. Commun. 5, 5267.
Okuwaki, M., Kato, K., Shimahara, H., Tate, S. -i., and Nagata, K. (2005). Assembly and
Disassembly of Nucleosome Core Particles Containing Histone Variants by Human Nucleosome
Assembly Protein I. Mol. Cell. Biol. 25, 10639–10651.
Okuwaki, M., Kato, K., and Nagata, K. (2010). Functional characterization of human
nucleosome assembly protein 1-like proteins as histone chaperones. Genes Cells 15, 13–27.
Pagan, J.K., Marzio, A., Jones, M.J.K., Saraf, A., Jallepalli, P.V., Florens, L., Washburn, M.P.,
and Pagano, M. (2014). Degradation of Cep68 and PCNT cleavage mediate Cep215 removal
from the PCM to allow centriole separation, disengagement and licensing. Nat. Cell Biol. 17,
31–43.
Paintrand, M., Moudjou, M., Delacroix, H., and Bornens, M. (1992). Centrosome organization
and centriole architecture: their sensitivity to divalent cations. J. Struct. Biol. 108, 107–128.
Paoletti, A., Moudjou, M., Paintrand, M., Salisbury, J.L., and Bornens, M. (1996). Most of
centrin in animal cells is not centrosome-associated and centrosomal centrin is confined to the
distal lumen of centrioles. J. Cell Sci. 109, 3089–3102.
Park, Y.-J., and Luger, K. (2006a). Structure and function of nucleosome assembly proteins.
Biochem. Cell Biol. 84, 549–549.
Park, Y.-J., and Luger, K. (2006b). The structure of nucleosome assembly protein 1. Proc. Natl.
Acad. Sci. U. S. A. 103, 1248–1253.
Park, S.-Y., Park, J.-E., Kim, T.-S., Kim, J.H., Kwak, M.-J., Ku, B., Tian, L., Murugan, R.N.,
Ahn, M., Komiya, S., et al. (2014). Molecular basis for unidirectional scaffold switching of
human Plk4 in centriole biogenesis. Nat. Struct. Mol. Biol.
Patzke, S., Stokke, T., and Aasheim, H.-C. (2006). CSPP and CSPP-L associate with
centrosomes and microtubules and differently affect microtubule organization. J. Cell. Physiol.
209, 199–210.
Patzke, S., Redick, S., Warsame, A., Murga-Zamalloa, C.A., Khanna, H., Doxsey, S., and
Stokke, T. (2010). CSPP is a ciliary protein interacting with Nephrocystin 8 and required for cilia
formation. Mol. Biol. Cell 21, 2555–2567.
Pearson, C.G., and Winey, M. (2010). Plk4/SAK/ZYG-1 in the regulation of centriole
duplication. F1000 Biol. Rep. 2.
Pearson, C.G., Osborn, D.P.S., Giddings, T.H., Beales, P.L., and Winey, M. (2009). Basal body
stability and ciliogenesis requires the conserved component Poc1. J. Cell Biol. 187, 905–920.

218
Pedrioli, P.G.A. (2010). Trans-Proteomic Pipeline: A Pipeline for Proteomic Analysis. In
Proteome Bioinformatics, S.J. Hubbard, and A.R. Jones, eds. (Totowa, NJ: Humana Press), pp.
213–238.
Pelletier, L., Özlü, N., Hannak, E., Cowan, C., Habermann, B., Ruer, M., Müller-Reichert, T.,
and Hyman, A.A. (2004). The Caenorhabditis elegans centrosomal protein SPD-2 is required for
both pericentriolar material recruitment and centriole duplication. Curr. Biol. 14, 863–873.
Pelletier, L., O’Toole, E., Schwager, A., Hyman, A.A., and Müller-Reichert, T. (2006). Centriole
assembly in Caenorhabditis elegans. Nature 444, 619–623.
Pennekamp, P., Menchen, T., Dworniczak, B., and Hamada, H. (2015). Situs inversus and ciliary
abnormalities: 20 years later, what is the connection? Cilia 4.
Peters, J.-M. (2006). The anaphase promoting complex/cyclosome: a machine designed to
destroy. Nat. Rev. Mol. Cell Biol. 7, 644–656.
Phillips, D.M. (1967). Giant centriole formation in Sciara. J. Cell Biol. 33, 73–92.
Piehl, M., Tulu, U.S., Wadsworth, P., and Cassimeris, L. (2004). Centrosome maturation:
measurement of microtubule nucleation throughout the cell cycle by using GFP-tagged EB1.
Proc. Natl. Acad. Sci. U. S. A. 101, 1584–1588.
Pihan, G.A. (2013). Centrosome Dysfunction Contributes to Chromosome Instability,
Chromoanagenesis, and Genome Reprograming in Cancer. Front. Oncol. 3.
Poser, I., Sarov, M., Hutchins, J.R.A., Hériché, J.-K., Toyoda, Y., Pozniakovsky, A., Weigl, D.,
Nitzsche, A., Hegemann, B., Bird, A.W., et al. (2008). BAC TransgeneOmics: a high-throughput
method for exploration of protein function in mammals. Nat. Methods 5, 409–415.
Prosser, S.L., Straatman, K.R., and Fry, A.M. (2009). Molecular Dissection of the Centrosome
Overduplication Pathway in S-Phase-Arrested Cells. Mol. Cell. Biol. 29, 1760–1773.
Puklowski, A., Homsi, Y., Keller, D., May, M., Chauhan, S., Kossatz, U., Grünwald, V.,
Kubicka, S., Pich, A., Manns, M.P., et al. (2011). The SCF–FBXW5 E3-ubiquitin ligase is
regulated by PLK4 and targets HsSAS-6 to control centrosome duplication. Nat. Cell Biol. 13,
1004–1009.
Qiao, R., Cabral, G., Lettman, M.M., Dammermann, A., and Dong, G. (2012). SAS-6 coiled-coil
structure and interaction with SAS-5 suggest a regulatory mechanism in C. elegans centriole
assembly. EMBO J. 31, 4334–4347.
Quintyne, N.J., Reing, J.E., Hoffelder, D.R., Gollin, S.M., and Saunders, W.S. (2005). Spindle
multipolarity is prevented by centrosomal clustering. Science 307, 127–129.
Regnard, C. (2000). Polyglutamylation of Nucleosome Assembly Proteins. J. Biol. Chem. 275,
15969–15976.

219
Rehtanz, M., Schmidt, H.-M., Warthorst, U., and Steger, G. (2004). Direct Interaction between
Nucleosome Assembly Protein 1 and the Papillomavirus E2 Proteins Involved in Activation of
Transcription. Mol. Cell. Biol. 24, 2153–2168.
Ring, D., Hubble, R., and Kirschner, M. (1982). Mitosis in a cell with multiple centrioles. J. Cell
Biol. 94, 549–556.
Rios, R.M. (2014). The centrosome-Golgi apparatus nexus. Philos. Trans. R. Soc. B Biol. Sci.
369, 20130462–20130462.
Roberts, A.J., Kon, T., Knight, P.J., Sutoh, K., and Burgess, S.A. (2013). Functions and
mechanics of dynein motor proteins. Nat. Rev. Mol. Cell Biol. 14, 713–726.
Rodrigues-Ferreira, S., Di Tommaso, A., Dimitrov, A., Cazaubon, S., Gruel, N., Colasson, H.,
Nicolas, A., Chaverot, N., Molinié, V., Reyal, F., et al. (2009). 8p22 MTUS1 Gene Product
ATIP3 Is a Novel Anti-Mitotic Protein Underexpressed in Invasive Breast Carcinoma of Poor
Prognosis. PLoS ONE 4, e7239.
Rodrigues-Martins, A., Bettencourt-Dias, M., Riparbelli, M., Ferreira, C., Ferreira, I., Callaini,
G., and Glover, D.M. (2007). DSAS-6 Organizes a Tube-like Centriole Precursor, and Its
Absence Suggests Modularity in Centriole Assembly. Curr. Biol. 17, 1465–1472.
Rodriguez, P., Munroe, D., Prawitt, D., Chu, L.L., Bric, E., Kim, J., Reid, L.H., Davies, C.,
Nakagama, H., Loebbert, R., et al. (1997). Functional Characterization of Human Nucleosome
Assembly Protein-2 (NAP1L4) Suggests a Role as a Histone Chaperone. Genomics 44, 253–265.
Rogers, G.C., Rusan, N.M., Roberts, D.M., Peifer, M., and Rogers, S.L. (2009). The SCFSlimb
ubiquitin ligase regulates Plk4/Sak levels to block centriole reduplication. J. Cell Biol. 184, 225–
239.
Roosing, S., Hofree, M., Kim, S., Scott, E., Copeland, B., Romani, M., Silhavy, J.L., Rosti, R.O.,
Schroth, J., Mazza, T., et al. (2015). Functional genome-wide siRNA screen identifies KIAA0586
as mutated in Joubert syndrome. eLife 4.
Roque, H., Wainman, A., Richens, J., Kozyrska, K., Franz, A., and Raff, J.W. (2012).
Drosophila Cep135/Bld10 maintains proper centriole structure but is dispensable for cartwheel
formation. J. Cell Sci. 125, 5881–5886.
Rougeulle, C., and Avner, P. (1996). Cloning and characterization of a murine brain specific
gene Bpx and its human homologue lying within the Xic candidate region. Hum. Mol. Genet. 5,
41–49.
Roux, K.J., Kim, D.I., Raida, M., and Burke, B. (2012). A promiscuous biotin ligase fusion
protein identifies proximal and interacting proteins in mammalian cells. J. Cell Biol. 196, 801–
810.
Sadok, A., and Marshall, C.J. (2014). Rho GTPases: Masters of cell migration. Small GTPases 5,
1–7.

220
Saffin, J.-M., Venoux, M., Prigent, C., Espeut, J., Poulat, F., Giorgi, D., Abrieu, A., and
Rouquier, S. (2005). ASAP, a human microtubule-associated protein required for bipolar spindle
assembly and cytokinesis. Proc. Natl. Acad. Sci. U. S. A. 102, 11302–11307.
Schatten, H., and Sun, Q.-Y. (2010). The role of centrosomes in fertilization, cell division and
establishment of asymmetry during embryo development. Semin. Cell Dev. Biol. 21, 174–184.
Scheer, U. (2014). Historical roots of centrosome research: discovery of Boveri’s microscope
slides in Wurzburg. Philos. Trans. R. Soc. B Biol. Sci. 369, 20130469–20130469.
Schermelleh, L., Carlton, P.M., Haase, S., Shao, L., Winoto, L., Kner, P., Burke, B., Cardoso,
M.C., Agard, D.A., Gustafsson, M.G., et al. (2008). Subdiffraction multicolor imaging of the
nuclear periphery with 3D structured illumination microscopy. Science 320, 1332–1336.
Schermelleh, L., Heintzmann, R., and Leonhardt, H. (2010). A guide to super-resolution
fluorescence microscopy. J. Cell Biol. 190, 165–175.
Schiff, P.B., Fant, J., and Horwitz, S.B. (1979). Promotion of microtubule assembly in vitro by
taxol. Nature 277, 665–667.
Schimmack, S., Taylor, A., Lawrence, B., Alaimo, D., Schmitz-Winnenthal, H., Büchler, M.W.,
Modlin, I.M., and Kidd, M. (2014). A mechanistic role for the chromatin modulator, NAP1L1, in
pancreatic neuroendocrine neoplasm proliferation and metastases. Epigenetics Chromatin 7, 15.
Schmidt, T.I., Kleylein-Sohn, J., Westendorf, J., Le Clech, M., Lavoie, S.B., Stierhof, Y.-D., and
Nigg, E.A. (2009). Control of Centriole Length by CPAP and CP110. Curr. Biol. 19, 1005–1011.
Schöckel, L., Möckel, M., Mayer, B., Boos, D., and Stemmann, O. (2011). Cleavage of cohesin
rings coordinates the separation of centrioles and chromatids. Nat. Cell Biol. 13, 966–972.
Schroer, T.A. (2004). DYNACTIN. Annu. Rev. Cell Dev. Biol. 20, 759–779.
Shaheen, R., Shamseldin, H.E., Loucks, C.M., Seidahmed, M.Z., Ansari, S., Ibrahim Khalil, M.,
Al-Yacoub, N., Davis, E.E., Mola, N.A., Szymanska, K., et al. (2014a). Mutations in CSPP1,
Encoding a Core Centrosomal Protein, Cause a Range of Ciliopathy Phenotypes in Humans. Am.
J. Hum. Genet. 94, 73–79.
Shaheen, R., Schmidts, M., Faqeih, E., Hashem, A., Lausch, E., Holder, I., Superti-Furga, A.,
UK10K Consortium, Mitchison, H.M., Almoisheer, A., et al. (2014b). A founder CEP120
mutation in Jeune asphyxiating thoracic dystrophy expands the role of centriolar proteins in
skeletal ciliopathies. Hum. Mol. Genet.
Sharma, N., Berbari, N.F., and Yoder, B.K. (2008). Chapter 13 Ciliary Dysfunction in
Developmental Abnormalities and Diseases. In Current Topics in Developmental Biology,
(Elsevier), pp. 371–427.
Sharp, D.J., McDonald, K.L., Brown, H.M., Matthies, H.J., Walczak, C., Vale, R.D., Mitchison,
T.J., and Scholey, J.M. (1999). The bipolar kinesin, KLP61F, cross-links microtubules within

221
interpolar microtubule bundles of Drosophila embryonic mitotic spindles. J. Cell Biol. 144, 125–
138.
Sherr, C.J., and Roberts, J.M. (1999). CDK inhibitors: positive and negative regulators of G1-
phase progression. Genes Dev. 13, 1501–1512.
Shiloh, Y. (2001). ATM and ATR: networking cellular responses to DNA damage. Curr. Opin.
Genet. Dev. 11, 71–77.
Shinohara, H., Sakayori, N., Takahashi, M., and Osumi, N. (2013). Ninein is essential for the
maintenance of the cortical progenitor character by anchoring the centrosome to microtubules.
Biol. Open 2, 739–749.
Sillibourne, J.E., Delaval, B., Redick, S., Sinha, M., and Doxsey, S.J. (2007). Chromatin
remodeling proteins interact with pericentrin to regulate centrosome integrity. Mol. Biol. Cell 18,
3667–3680.
Sillibourne, J.E., Tack, F., Vloemans, N., Boeckx, A., Thambirajah, S., Bonnet, P., Ramaekers,
F.C., Bornens, M., and Grand-Perret, T. (2010). Autophosphorylation of polo-like kinase 4 and
its role in centriole duplication. Mol. Biol. Cell 21, 547–561.
Silverman, J.S., Skaar, J.R., and Pagano, M. (2012). SCF ubiquitin ligases in the maintenance of
genome stability. Trends Biochem. Sci. 37, 66–73.
Simon, H.U., Mills, G.B., Kozlowski, M., Hogg, D., Branch, D., Ishimi, Y., and Siminovitch,
K.A. (1994). Molecular characterization of hNRP, a cDNA encoding a human nucleosome-
assembly-protein-I-related gene product involved in the induction of cell proliferation. Biochem
J 297, 389–397.
Singla, V., Romaguera-Ros, M., Garcia-Verdugo, J.M., and Reiter, J.F. (2010). Ofd1, a Human
Disease Gene, Regulates the Length and Distal Structure of Centrioles. Dev. Cell 18, 410–424.
Sivakumar, S., and Gorbsky, G.J. (2015). Spatiotemporal regulation of the anaphase-promoting
complex in mitosis. Nat. Rev. Mol. Cell Biol. 16, 82–94.
Skaar, J.R., Pagan, J.K., and Pagano, M. (2013). Mechanisms and function of substrate
recruitment by F-box proteins. Nat. Rev. Mol. Cell Biol. 14, 369–381.
Solomon, M.J., Larsen, P.L., and Varshavsky, A. (1988). Mapping protein-DNA interactions in
vivo with formaldehyde: evidence that histone H4 is retained on a highly transcribed gene. Cell
53, 937–947.
Song, M.H., Liu, Y., Anderson, D.E., Jahng, W.J., and O’Connell, K.F. (2011). Protein
Phosphatase 2A-SUR-6/B55 Regulates Centriole Duplication in C. elegans by Controlling the
Levels of Centriole Assembly Factors. Dev. Cell 20, 563–571.

222
Song, Y., Kirkpatrick, L.L., Schilling, A.B., Helseth, D.L., Chabot, N., Keillor, J.W., Johnson,
G.V.W., and Brady, S.T. (2013). Transglutaminase and Polyamination of Tubulin:
Posttranslational Modification for Stabilizing Axonal Microtubules. Neuron 78, 109–123.
Sonnen, K.F., Schermelleh, L., Leonhardt, H., and Nigg, E.A. (2012). 3D-structured illumination
microscopy provides novel insight into architecture of human centrosomes. Biol. Open 1, 965–
976.
Sonnen, K.F., Gabryjonczyk, A.-M., Anselm, E., Stierhof, Y.-D., and Nigg, E.A. (2013). Human
Cep192 and Cep152 cooperate in Plk4 recruitment and centriole duplication. J. Cell Sci. 126,
3223–3233.
Spektor, A., Tsang, W.Y., Khoo, D., and Dynlacht, B.D. (2007). Cep97 and CP110 Suppress a
Cilia Assembly Program. Cell 130, 678–690.
Staples, C.J., Myers, K.N., Beveridge, R.D.D., Patil, A.A., Lee, A.J.X., Swanton, C., Howell,
M., Boulton, S.J., and Collis, S.J. (2012). The centriolar satellite protein Cep131 is important for
genome stability. J. Cell Sci. 125, 4770–4779.
Stevens, N.R., Dobbelaere, J., Brunk, K., Franz, A., and Raff, J.W. (2010). Drosophila Ana2 is a
conserved centriole duplication factor. J. Cell Biol. 188, 313–323.
Stinchcombe, J.C., and Griffiths, G.M. (2014). Communication, the centrosome and the
immunological synapse. Philos. Trans. R. Soc. B Biol. Sci. 369, 20130463–20130463.
Stinchcombe, J.C., Majorovits, E., Bossi, G., Fuller, S., and Griffiths, G.M. (2006). Centrosome
polarization delivers secretory granules to the immunological synapse. Nature 443, 462–465.
Strnad, P., Leidel, S., Vinogradova, T., Euteneuer, U., Khodjakov, A., and Gönczy, P. (2007).
Regulated HsSAS-6 Levels Ensure Formation of a Single Procentriole per Centriole during the
Centrosome Duplication Cycle. Dev. Cell 13, 203–213.
Sun, X., Shi, X., Liu, M., Li, D., Zhang, L., Liu, X., and Zhou, J. (2011). Mdp3 is a novel
microtubule-binding protein that regulates microtubule assembly and stability. Cell Cycle 10,
3929–3937.
Tang, C.-J.C., Fu, R.-H., Wu, K.-S., Hsu, W.-B., and Tang, T.K. (2009a). CPAP is a cell-cycle
regulated protein that controls centriole length. Nat. Cell Biol. 11, 825–831.
Tang, C.-J.C., Fu, R.-H., Wu, K.-S., Hsu, W.-B., and Tang, T.K. (2009b). CPAP is a cell-cycle
regulated protein that controls centriole length. Nat. Cell Biol. 11, 825–831.
Tang, C.-J.C., Lin, S.-Y., Hsu, W.-B., Lin, Y.-N., Wu, C.-T., Lin, Y.-C., Chang, C.-W., Wu, K.-
S., and Tang, T.K. (2011). The human microcephaly protein STIL interacts with CPAP and is
required for procentriole formation. EMBO J. 30, 4790–4804.

223
Tanos, B.E., Yang, H.-J., Soni, R., Wang, W.-J., Macaluso, F.P., Asara, J.M., and Tsou, M.-F.B.
(2013). Centriole distal appendages promote membrane docking, leading to cilia initiation.
Genes Dev. 27, 163–168.
Thauvin-Robinet, C., Lee, J.S., Lopez, E., Herranz-Pérez, V., Shida, T., Franco, B., Jego, L., Ye,
F., Pasquier, L., Loget, P., et al. (2014). The oral-facial-digital syndrome gene C2CD3 encodes a
positive regulator of centriole elongation. Nat. Genet.
Tobin, J.L., and Beales, P.L. (2009). The nonmotile ciliopathies. Genet. Med. 11, 386–402.
Tollenaere, M.A.X., Mailand, N., and Bekker-Jensen, S. (2015). Centriolar satellites: key
mediators of centrosome functions. Cell. Mol. Life Sci. 72, 11–23.
Torchy, M.P., Hamiche, A., and Klaholz, B.P. (2015). Structure and function insights into the
NuRD chromatin remodeling complex. Cell. Mol. Life Sci.
Tritarelli, A., Oricchio, E., Ciciarello, M., Mangiacasale, R., Palena, A., Lavia, P., Soddu, S., and
Cundari, E. (2004). p53 localization at centrosomes during mitosis and postmitotic checkpoint
are ATM-dependent and require serine 15 phosphorylation. Mol. Biol. Cell 15, 3751–3757.
Tsou, M.-F.B., and Stearns, T. (2006). Mechanism limiting centrosome duplication to once per
cell cycle. Nature 442, 947–951.
Tsou, M.-F.B., Wang, W.-J., George, K.A., Uryu, K., Stearns, T., and Jallepalli, P.V. (2009).
Polo Kinase and Separase Regulate the Mitotic Licensing of Centriole Duplication in Human
Cells. Dev. Cell 17, 344–354.
Uetake, Y., and Sluder, G. (2010). Prolonged Prometaphase Blocks Daughter Cell Proliferation
Despite Normal Completion of Mitosis. Curr. Biol. 20, 1666–1671.
Uetake, Y., Terada, Y., Matuliene, J., and Kuriyama, R. (2004). Interaction of Cep135 with a
p50 dynactin subunit in mammalian centrosomes. Cell Motil. Cytoskeleton 58, 53–66.
Vallee, R.B., Williams, J.C., Varma, D., and Barnhart, L.E. (2004). Dynein: An ancient motor
protein involved in multiple modes of transport. J. Neurobiol. 58, 189–200.
Venoux, M., Basbous, J., Berthenet, C., Prigent, C., Fernandez, A., Lamb, N.J., and Rouquier, S.
(2007). ASAP is a novel substrate of the oncogenic mitotic kinase Aurora-A: phosphorylation on
Ser625 is essential to spindle formation and mitosis. Hum. Mol. Genet. 17, 215–224.
Venoux, M., Delmouly, K., Milhavet, O., Vidal-Eychenié, S., Giorgi, D., and Rouquier, S.
(2008). Gene organization, evolution and expression of the microtubule-associated protein ASAP
(MAP9). BMC Genomics 9, 406.
Venoux, M., Tait, X., Hames, R.S., Straatman, K.R., Woodland, H.R., and Fry, A.M. (2013).
Poc1A and Poc1B act together in human cells to ensure centriole integrity. J. Cell Sci. 126, 163–
175.

224
Villumsen, B.H., Danielsen, J.R., Povlsen, L., Sylvestersen, K.B., Merdes, A., Beli, P., Yang, Y.-
G., Choudhary, C., Nielsen, M.L., Mailand, N., et al. (2013). A new cellular stress response that
triggers centriolar satellite reorganization and ciliogenesis. EMBO J. 32, 3029–3040.
Voges, P., Zwickl, P., and Baumeister (1999). The 26S Proteasome: A Molecular Machine
Designed For Controlled Proteolysis. Annu. Rev. Biochem. 68, 1015–1068.
Vorobjev, I.A., and YuS, C. (1982). Centrioles in the cell cycle. I. Epithelial cells. J. Cell Biol.
93, 938–949.
Vulprecht, J., David, A., Tibelius, A., Castiel, A., Konotop, G., Liu, F., Bestvater, F., Raab,
M.S., Zentgraf, H., Izraeli, S., et al. (2012). STIL is required for centriole duplication in human
cells. J. Cell Sci. 125, 1353–1362.
Wang, W.-J., Soni, R.K., Uryu, K., and Bryan Tsou, M.-F. (2011). The conversion of centrioles
to centrosomes: essential coupling of duplication with segregation. J. Cell Biol. 193, 727–739.
Wang, X., Tsai, J.-W., Imai, J.H., Lian, W.-N., Vallee, R.B., and Shi, S.-H. (2009). Asymmetric
centrosome inheritance maintains neural progenitors in the neocortex. Nature 461, 947–955.
Wang, Z., Wu, T., Shi, L., Zhang, L., Zheng, W., Qu, J.Y., Niu, R., and Qi, R.Z. (2010).
Conserved Motif of CDK5RAP2 Mediates Its Localization to Centrosomes and the Golgi
Complex. J. Biol. Chem. 285, 22658–22665.
Waters, A.M., and Beales, P.L. (2011). Ciliopathies: an expanding disease spectrum. Pediatr.
Nephrol. 26, 1039–1056.
Winey, M., and O’Toole, E. (2014). Centriole structure. Philos. Trans. R. Soc. B Biol. Sci. 369,
20130457–20130457.
Wloga, D., and Gaertig, J. (2011). Post-translational modifications of microtubules. J. Cell Sci.
124, 154–154.
Wolff, A., de Néchaud, B., Chillet, D., Mazarguil, H., Desbruyères, E., Audebert, S., Eddé, B.,
Gros, F., and Denoulet, P. (1992). Distribution of glutamylated alpha and beta-tubulin in mouse
tissues using a specific monoclonal antibody, GT335. Eur. J. Cell Biol. 59, 425–432.
Wong, Y.L., Anzola, J.V., Davis, R.L., Yoon, M., Motamedi, A., Kroll, A., Seo, C.P., Hsia, J.E.,
Kim, S.K., Mitchell, J.W., et al. (2015). Reversible centriole depletion with an inhibitor of Polo-
like kinase 4. Science.
Woodruff, J.B., Wueseke, O., and Hyman, A.A. (2014). Pericentriolar material structure and
dynamics. Philos. Trans. R. Soc. B Biol. Sci. 369, 20130459–20130459.
Wu, C., Yang, M., Li, J., Wang, C., Cao, T., Tao, K., and Wang, B. (2014). Talpid3-Binding
Centrosomal Protein Cep120 Is Required for Centriole Duplication and Proliferation of
Cerebellar Granule Neuron Progenitors. PLoS ONE 9, e107943.

225
Xie, Z., Moy, L.Y., Sanada, K., Zhou, Y., Buchman, J.J., and Tsai, L.-H. (2007). Cep120 and
TACCs Control Interkinetic Nuclear Migration and the Neural Progenitor Pool. Neuron 56, 79–
93.
Yadav, S., and Linstedt, A.D. (2011). Golgi Positioning. Cold Spring Harb. Perspect. Biol. 3,
a005322–a005322.
Yadav, S., Puri, S., and Linstedt, A.D. (2009). A primary role for Golgi positioning in directed
secretion, cell polarity, and wound healing. Mol. Biol. Cell 20, 1728–1736.
Yamanaka, M., Smith, N.I., and Fujita, K. (2014). Introduction to super-resolution microscopy.
Microscopy 63, 177–192.
Yamashita, Y.M., and Fuller, M.T. (2008). Asymmetric centrosome behavior and the
mechanisms of stem cell division. J. Cell Biol. 180, 261–266.
Yamashita, Y.M., Jones, D.L., and Fuller, M.T. (2003). Orientation of asymmetric stem cell
division by the APC tumor suppressor and centrosome. Science 301, 1547–1550.
Yamashita, Y.M., Mahowald, A.P., Perlin, J.R., and Fuller, M.T. (2007). Asymmetric
Inheritance of Mother Versus Daughter Centrosome in Stem Cell Division. Science 315, 518–
521.
Yan, X., Habedanck, R., and Nigg, E.A. (2006). A complex of two centrosomal proteins,
CAP350 and FOP, cooperates with EB1 in microtubule anchoring. Mol. Biol. Cell 17, 634–644.
Ye, X., Zeng, H., Ning, G., Reiter, J.F., and Liu, A. (2014). C2cd3 is critical for centriolar distal
appendage assembly and ciliary vesicle docking in mammals. Proc. Natl. Acad. Sci. 111, 2164–
2169.
Yin, Y., Bangs, F., Paton, I.R., Prescott, A., James, J., Davey, M.G., Whitley, P., Genikhovich,
G., Technau, U., Burt, D.W., et al. (2009). The Talpid3 gene (KIAA0586) encodes a centrosomal
protein that is essential for primary cilia formation. Development 136, 655–664.
Yu, J., Liu, X., Ye, H., and Zhou, X. (2009). Genomic characterization of the human
mitochondrial tumor suppressor gene 1 (MTUS1): 5’ cloning and preliminary analysis of the
multiple gene promoters. BMC Res. Notes 2, 109.
Zhang, Y., Wang, Y., Wei, Y., Ma, J., Peng, J., Wumaier, R., Shen, S., Zhang, P., and Yu, L.
(2015). The tumor suppressor proteins ASPP1 and ASPP2 interact with C-Nap1 and regulate
centrosome linker reassembly. Biochem. Biophys. Res. Commun. 458, 494–500.
Zhu, F., Lawo, S., Bird, A., Pinchev, D., Ralph, A., Richter, C., Müller-Reichert, T., Kittler, R.,
Hyman, A.A., and Pelletier, L. (2008). The Mammalian SPD-2 Ortholog Cep192 Regulates
Centrosome Biogenesis. Curr. Biol. 18, 136–141.
Zlatanova, J., Seebart, C., and Tomschik, M. (2007). Nap1: taking a closer look at a juggler
protein of extraordinary skills. FASEB J. 21, 1294–1310.

226
Zou, C., Li, J., Gunning, W.T., Wazer, D.E., Band, V., and Gao, Q. (2005). Centrobin: a novel
daughter centriole-associated protein that is required for centriole duplication. J. Cell Biol. 171,
437–445.
Zuern, C., Heimrich, J., Kaufmann, R., Richter, K.K., Settmacher, U., Wanner, C., Galle, J., and
Seibold, S. (2010). Down-regulation of MTUS1 in human colon tumors. Oncol. Rep. 23, 183–
189.
Copyright © 2022 FDOKUMEN




















