
The protection of cells from nitric oxide-mediated apoptotic death by mechanochemically synthesized...
10
The protection of cells from nitric oxide-mediated apoptotic death by mechanochemically synthesized fullerene (C 60 ) nanoparticles Maja S. Misirkic a, b,1 , Biljana M. Todorovic-Markovic c,1 , Ljubica M. Vucicevic a, b , Kristina D. Janjetovic a, b , Vukoman R. Jokanovic c , Miroslav D. Dramicanin c , Zoran M. Markovic c, ** , Vladimir S. Trajkovic a, * a Institute of Microbiology and Immunology, School of Medicine, University of Belgrade, Dr. Subotica 1,11000 Belgrade, Serbia b Institute for Biological Research ‘‘Sinisa Stankovic’’, Bulevar Despota Stefana 142,11000 Belgrade, Serbia c Vinca Institute of Nuclear Sciences, POB 522, 11000 Belgrade, Serbia article info Article history: Received 28 November 2008 Accepted 3 January 2009 Available online 5 February 2009 Keywords: Carbon Nanoparticles Nitric oxide Cytotoxicity Apoptosis abstract The influence of fullerene (C 60 ) nanoparticles on the cytotoxicity of a highly reactive free radical nitric oxide (NO) was investigated. Fullerene nanoparticles were prepared by mechanochemically assisted complexation with anionic surfactant sodium dodecyl sulfate, macrocyclic oligosaccharide g-cyclodex- trin or the copolymer ethylene vinyl acetate–ethylene vinyl versatate. C 60 nanoparticles were charac- terized by UV–vis and atomic force microscopy. While readily internalized by mouse L929 fibroblasts, C 60 nanoparticles were not cytotoxic. Moreover, they partially protected L929 cells from the cytotoxic effect of NO-releasing compounds sodium nitroprusside (SNP), S-nitroso-N-acetylpenicillamine (SNAP), S- nitrosoglutathione (GSNO) and 3-morpholino-sydnonimine (SIN-1). C 60 nanoparticles reduced SNP- induced apoptotic cell death by preventing mitochondrial depolarization, caspase activation, cell membrane phosphatidylserine exposure and DNA fragmentation. The protective action of C 60 nano- particles was not exerted via direct interaction with NO, but through neutralization of mitochondria- produced superoxide radical in NO-treated cells, as demonstrated by using different redox-sensitive reporter fluorochromes. These data suggest that C 60 complexes with appropriate host molecules might be plausible candidates for preventing NO-mediated cell injury in inflammatory/autoimmune disorders. Ó 2009 Elsevier Ltd. All rights reserved. 1. Introduction Because of the unique physico-chemical properties, fullerene (C 60 ) is endowed with a puzzling ability to generate reactive oxygen species (ROS) when photoexcited, as well as to quench ROS in the absence of photosensitization [1]. The latter feature makes fullerene a potential candidate for therapeutic cell protection from oxidative stress-mediated tissue destruction during excessive inflammation or following exposure to some toxins [1]. Water- soluble fullerene derivatives with various functional groups (e.g. –OH, –COOH) attached to the fullerene cage are particularly efficient in protecting various cell types from ROS-mediated damage, both in vitro and in vivo [2–6]. However, in contrast to derivatized fuller- enes, water-stable crystalline aggregates of pure C 60 formed by solvent extraction (nano-C 60 ) did not display antioxidant ability [7]. Moreover, nano-C 60 produced high amounts of ROS in the absence of overt photosensitization, killing various cell types in vitro [7–9] and causing damage to aquatic species in vivo [10]. The mechanisms underlying such as distinct behavior of derivatized vs. pure C 60 might involve decrease in ROS generation with increasing derivati- zation of the fullerene cage, as well as contribution of the attached functional groups to ROS quenching [11]. On the other hand, some recent data suggest that ROS-mediated cytotoxicity of nano-C 60 might not be an inherent property of pure C 60 , but rather a result of the residual presence of tetrahydrofuran (THF), the organic solvent intercalated into nano-C 60 crystalline lattice during its preparation [11–13]. Accordingly, pure nano-C 60 suspension prepared by long- term stirring in water, unlike THF-prepared nano-C 60 , did not show acute toxicity to various invertebrates [14]. Furthermore, detergent (Tween)-solubilized pure C 60 was protective in carbon–tetrachlo- ride model of ROS-mediated liver injury in rats [15], while exposure of nano-C 60 to g-irradiation caused decomposition of residual THF, resulting in a shift of nano-C 60 ROS-related properties from proox- idant/cytotoxic to antioxidant/cytoprotective [12]. These data are consistent with the findings that supramolecular C 60 complexes with polyvinylpyrrolidone or g-cyclodextrin displayed potent anti- oxidant activity in different in vitro ROS-generating systems [16,17]. * Corresponding author. Tel.: þ381 11 3643 233; fax: þ381 11 3643 235. ** Corresponding author. Tel.: þ381 11 2455 451; fax: þ381 11 344 0100. E-mail addresses: [email protected] (Z.M. Markovic), [email protected] (V.S. Trajkovic). 1 These authors equally contributed to the work. Contents lists available at ScienceDirect Biomaterials journal homepage: www.elsevier.com/locate/biomaterials 0142-9612/$ – see front matter Ó 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.biomaterials.2009.01.023 Biomaterials 30 (2009) 2319–2328
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The protection of cells from nitric oxide-mediated apoptotic death by mechanochemically synthesized...

lable at ScienceDirect
Biomaterials 30 (2009) 2319–2328
Contents lists avai
Biomaterials
journal homepage: www.elsevier .com/locate/biomateria ls
The protection of cells from nitric oxide-mediated apoptotic death bymechanochemically synthesized fullerene (C60) nanoparticles
Maja S. Misirkic a,b,1, Biljana M. Todorovic-Markovic c,1, Ljubica M. Vucicevic a,b, Kristina D. Janjetovic a,b,Vukoman R. Jokanovic c, Miroslav D. Dramicanin c, Zoran M. Markovic c,**, Vladimir S. Trajkovic a,*
a Institute of Microbiology and Immunology, School of Medicine, University of Belgrade, Dr. Subotica 1, 11000 Belgrade, Serbiab Institute for Biological Research ‘‘Sinisa Stankovic’’, Bulevar Despota Stefana 142, 11000 Belgrade, Serbiac Vinca Institute of Nuclear Sciences, POB 522, 11000 Belgrade, Serbia
a r t i c l e i n f o
Article history:Received 28 November 2008Accepted 3 January 2009Available online 5 February 2009
Keywords:CarbonNanoparticlesNitric oxideCytotoxicityApoptosis
* Corresponding author. Tel.: þ381 11 3643 233; fa** Corresponding author. Tel.: þ381 11 2455 451; fa
E-mail addresses: [email protected] (Z.M. M(V.S. Trajkovic).
1 These authors equally contributed to the work.
0142-9612/$ – see front matter � 2009 Elsevier Ltd.doi:10.1016/j.biomaterials.2009.01.023
a b s t r a c t
The influence of fullerene (C60) nanoparticles on the cytotoxicity of a highly reactive free radical nitricoxide (NO) was investigated. Fullerene nanoparticles were prepared by mechanochemically assistedcomplexation with anionic surfactant sodium dodecyl sulfate, macrocyclic oligosaccharide g-cyclodex-trin or the copolymer ethylene vinyl acetate–ethylene vinyl versatate. C60 nanoparticles were charac-terized by UV–vis and atomic force microscopy. While readily internalized by mouse L929 fibroblasts, C60
nanoparticles were not cytotoxic. Moreover, they partially protected L929 cells from the cytotoxic effectof NO-releasing compounds sodium nitroprusside (SNP), S-nitroso-N-acetylpenicillamine (SNAP), S-nitrosoglutathione (GSNO) and 3-morpholino-sydnonimine (SIN-1). C60 nanoparticles reduced SNP-induced apoptotic cell death by preventing mitochondrial depolarization, caspase activation, cellmembrane phosphatidylserine exposure and DNA fragmentation. The protective action of C60 nano-particles was not exerted via direct interaction with NO, but through neutralization of mitochondria-produced superoxide radical in NO-treated cells, as demonstrated by using different redox-sensitivereporter fluorochromes. These data suggest that C60 complexes with appropriate host molecules mightbe plausible candidates for preventing NO-mediated cell injury in inflammatory/autoimmune disorders.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Because of the unique physico-chemical properties, fullerene(C60) is endowed with a puzzling ability to generate reactive oxygenspecies (ROS) when photoexcited, as well as to quench ROS inthe absence of photosensitization [1]. The latter feature makesfullerene a potential candidate for therapeutic cell protection fromoxidative stress-mediated tissue destruction during excessiveinflammation or following exposure to some toxins [1]. Water-soluble fullerene derivatives with various functional groups (e.g.–OH, –COOH) attached to the fullerene cage are particularly efficientin protecting various cell types from ROS-mediated damage, both invitro and in vivo [2–6]. However, in contrast to derivatized fuller-enes, water-stable crystalline aggregates of pure C60 formed bysolvent extraction (nano-C60) did not display antioxidant ability [7].
x: þ381 11 3643 235.x: þ381 11 344 0100.arkovic), [email protected]
All rights reserved.
Moreover, nano-C60 produced high amounts of ROS in the absence ofovert photosensitization, killing various cell types in vitro [7–9] andcausing damage to aquatic species in vivo [10]. The mechanismsunderlying such as distinct behavior of derivatized vs. pure C60
might involve decrease in ROS generation with increasing derivati-zation of the fullerene cage, as well as contribution of the attachedfunctional groups to ROS quenching [11]. On the other hand, somerecent data suggest that ROS-mediated cytotoxicity of nano-C60
might not be an inherent property of pure C60, but rather a result ofthe residual presence of tetrahydrofuran (THF), the organic solventintercalated into nano-C60 crystalline lattice during its preparation[11–13]. Accordingly, pure nano-C60 suspension prepared by long-term stirring in water, unlike THF-prepared nano-C60, did not showacute toxicity to various invertebrates [14]. Furthermore, detergent(Tween)-solubilized pure C60 was protective in carbon–tetrachlo-ride model of ROS-mediated liver injury in rats [15], while exposureof nano-C60 to g-irradiation caused decomposition of residual THF,resulting in a shift of nano-C60 ROS-related properties from proox-idant/cytotoxic to antioxidant/cytoprotective [12]. These data areconsistent with the findings that supramolecular C60 complexeswith polyvinylpyrrolidone or g-cyclodextrin displayed potent anti-oxidant activity in different in vitro ROS-generating systems [16,17].

M.S. Misirkic et al. / Biomaterials 30 (2009) 2319–23282320
While these findings indicate that pure C60 might also have anti-oxidant properties, their biological significance in terms of cell-protective activity has not been directly assessed.
In addition to ROS such as superoxide anion (O2��) and hydroxyl
radical (�OH), nitric oxide (NO) is a highly reactive, potentially cell-damaging free radical that belongs to both ROS and reactivenitrogen species (RNS). Synthesized intracellularly from L-arginineby NO synthase enzyme family, NO diffuses very rapidly throughwater and membranes, and thus from one cell to another. Whileimplicated in many physiological processes such as smooth musclerelaxation, neurotransmission, immune regulation and hostdefense against microbes and tumors, NO is also involved in hostcell destruction in pathological inflammation during immuneresponses to pathogens or self-antigens in autoimmunity [18]. NOis very reactive and generates a variety of secondary products suchas nitrosonium ion (NOþ), peroxynitrite (ONOO�), nitroxyl anion(NO�) and nitrosothiols. These RNS, including NO, interact withbiomolecules to cause mitochondrial and DNA damage, lipidoxidation, protein modification and alterations in enzyme activity,ultimately leading to apoptotic or necrotic cell death [19]. It hasbeen reported that malonic acid C60 derivatives inhibited thecatalytic activity of NO synthases [20], while C60(OH)24 (fullerol)was able to quench NO and block its biological activity in vivo [21].To the best of our knowledge, similar ability of other C60 nano-particles has not been reported so far.
In the present study, we tested the ability of various C60 nano-particles to influence oxidative stress-mediated cytotoxicity of NO.To overcome C60 insolubility in water, we used anionic surfactantsodium dodecyl sulfate (SDS), macrocyclic oligosaccharide g-cyclodextrin (CDX), or the copolymer ethylene vinyl acetate–ethylene vinyl versatate (EVA–EVV) for mechanochemicalpreparation of C60 nanoparticles.
2. Materials and methods
2.1. Preparation and characterization of C60 nanoparticles
Soft mechanochemical activation was performed in a KHD Humboldt WedagAG vibration mill (type MH954/3), which was reconstructed for the softmechanical synthesis. The mill has a cone-shaped, alumina-coated workingvolume with a maximal-upper cone diameter of 8 cm. Two alumina-coatedceramic balls with the diameter of 3.5 cm were used for fullerene milling. The millvibration rate was 10 000 vibrations per minute, while the energy of the impactwas chosen to be only enough for the disintegration of the fullerene powder intonanoparticles, without destruction of the C60 structure. In the first phase, 20 mg ofC60 (99.9%; Bucky USA, Houston, TX) were milled for 15 min with the half quantityof the total amount (2.5 mg) of the sodium dodecyl sulfate (SDS), g-cyclodextrin(CDX) (both from Sigma, St. Louis, MO) or ethylene vinyl acetate–ethylene vinylversatate (EVA–EVV; kindly provided by Celanese GMBH, Frankfurt, Germany).Thereafter, the small quantity of the ethanol/deionized water mixture (ratio 4:1)was added to sustain homogenization and fullerene coating by the thin film ofSDS/CDX/EVA–EVV. Each mixture was then milled for further 15 min. The rest ofeach solubilizing agent was added in equal four portions, with 7 min-long millingperiods in between and wetting with ethanol/water before adding the last portion.After the last milling, the samples were diluted in deionized water, filteredthrough 0.45 mM filter and concentration of C60 was adjusted to 1 mg/ml. Controlsamples containing only the solubilizing agent without C60 were prepared usingthe same procedure. The UV–vis spectra of C60 suspensions (C60SDS, C60CDX,C60EVA-EVV) were scanned within the wavelength range of 200–550 nm usingAvantes UV–vis spectrophotometer. All UV–vis measurements were carried out at20 �C and automatically corrected for the suspending medium (water). AFMmeasurements were performed with a Quesant (Santa Cruz, CA) microscope in thetapping mode in air, with the cantilever oscillating close to resonance and the tiponly slightly touching the surface. Particles were spread on a fixed mica substrateand imaged after drying. Standard silicon tips (Nano and More, Wetzlar, Germany)with force constant 40 N/m were used.
2.2. Cell cultures
The mouse L929 fibroblast cell line, reportedly sensitive to the cytotoxic actionof NO [22–24], was obtained from the European Collection of Animal Cell Cultures(Salisbury, UK). The cells were maintained at 37 �C in a humidified atmosphere with
5% CO2, in an HEPES (20 mM)-buffered RPMI 1640 cell culture medium (Sigma)supplemented with 5% fetal calf serum, 2 mM L-glutamine, 10 mM sodium pyruvateand penicillin/streptomycin. The cells were prepared for experiments usingconventional trypsinization procedure with trypsin/EDTA and incubated in 96-wellflat-bottom plates (1�104 cells/well) for the cell viability assessment or 24-wellplates (5�104 cells/well) for the flow cytometric analysis. Cells were rested for 24 hand then treated with nitric oxide donors sodium nitroprusside (SNP), S-nitroso-N-acetylpenicillamine (SNAP), S-nitrosoglutathione (GSNO) and 3-morpholino-syd-nonimine (SIN-1) (all from Sigma), in the absence or presence of C60 nanoparticles,as described in Results and figure captions.
2.3. Cell viability and C60 uptake
Cell numbers were determined exactly as previously described, using crystalviolet to stain viable, adherent cells [25]. The crystal violet absorbance, corre-sponding to the number of viable cells, was measured in an automated microplatereader at 570 nm. The results were presented as % of the control value obtained inuntreated cells. The cellular uptake of C60 nanoparticles was determined by flowcytometry as previously described [26], by monitoring the extent of side-scatteringof the laser beam (SSC), which corresponds to the increase in cell granularity.
2.4. Determination of apoptosis
Apoptotic cell death was analyzed by double staining with annexin V-FITC andpropidium iodide (PI), in which annexin V binds to early apoptotic cells withexposed phosphatidylserine, while PI labels the late apoptotic/necrotic cells witha membrane damage. Staining was performed according to the instructions by themanufacturer (BD Pharmingen, San Diego, CA). DNA fragmentation, as a hallmarkof apoptotic cell death, was analyzed by measuring the amount of PI-labeled DNAin fixed cells, as previously described [25]. The green/red (FL1/FL2) fluorescence ofannexin/PI- and PI-stained cells was analyzed with FACSCalibur flow cytometer(BD, Heidelberg, Germany). The number of viable (annexin�PI�), early apoptotic(annexinþPI�) and late apoptotic/necrotic (annexinþPIþ) cells, as well as theproportion of cells displaying DNA fragmentation (hypodiploid sub-G compart-ment in the cell cycle analysis), were determined with a Cell Quest Pro software(BD).
2.5. Caspase activation
Activation of caspases, the principal intracellular molecules involved inapoptosis execution, was measured by flow cytometry after labeling the cells witha cell-permeable, FITC-conjugated pan-caspase inhibitor (ApoStat; R&D Systems,Minneapolis, MN) according to the manufacturer’s instructions. The increase ingreen fluorescence (FL1) as a measure of caspase activity within individual cells ofthe treated population was determined using FACSCalibur flow cytometer. Theresults are expressed as % of cells containing active caspases.
2.6. Analysis of mitochondrial membrane potential
Mitochondrial membrane potential was assessed using DePsipher (R&DSystems), a lipophilic cation that has the property of aggregating upon membranepolarization forming an orange–red fluorescent compound. If the potential isdisturbed, the dye cannot access the transmembrane space and remains or revertsto its green monomeric form. The cells were stained with DePsipher as describedby the manufacturer, and the green monomer and the red aggregates weredetected by flow cytometry. The results were presented as a red/green fluores-cence ratio (FL2/FL1, arbitrarily set to 1 in untreated samples), the increase anddecrease of which reflect mitochondrial hyperpolarization and depolarization,respectively.
2.7. Measurement of reactive oxygen species (ROS) and reactive nitrogen species(RNS)
Intracellular production of ROS/RNS was determined by measuring theintensity of green fluorescence emitted by non-selective redox-sensitive dyedihydrorhodamine 123 (DHR; Invitrogen, Carlsbad, CA), able to detect variousROS/RNS, such as hydrogen peroxide, superoxide radical anion (O2
��), NO andONOO� [27]. The production of superoxide and RNS was measured using redox-sensitive fluorochromes dihydroethidium (DHE) and diaminofluorescein (DAF),which are fairly selective for O2
�� [28] and NO/ONOO� [29], respectively. DHR(5 mM) was added to cell cultures at the beginning of treatment, while DHE(20 mM) and DAF (20 mM) were incubated with cells for 30 min at the end of thetreatment. Cells were detached by trypsinization and washed with phosphate-buffered saline. The mean intensity of green (FL1) fluorescence (DHR, DAF) or red(FL2) fluorescence (DHE), corresponding to ROS or RNS levels, was determinedusing a FACSCalibur flow cytometer. Alternatively, DHR and DAF were used todetermine ROS and RNS in the cell-free conditions, using Chameleon (Hidex,Turku, Finland) fluorescence reader (488 nm excitation, 535 nm detection).
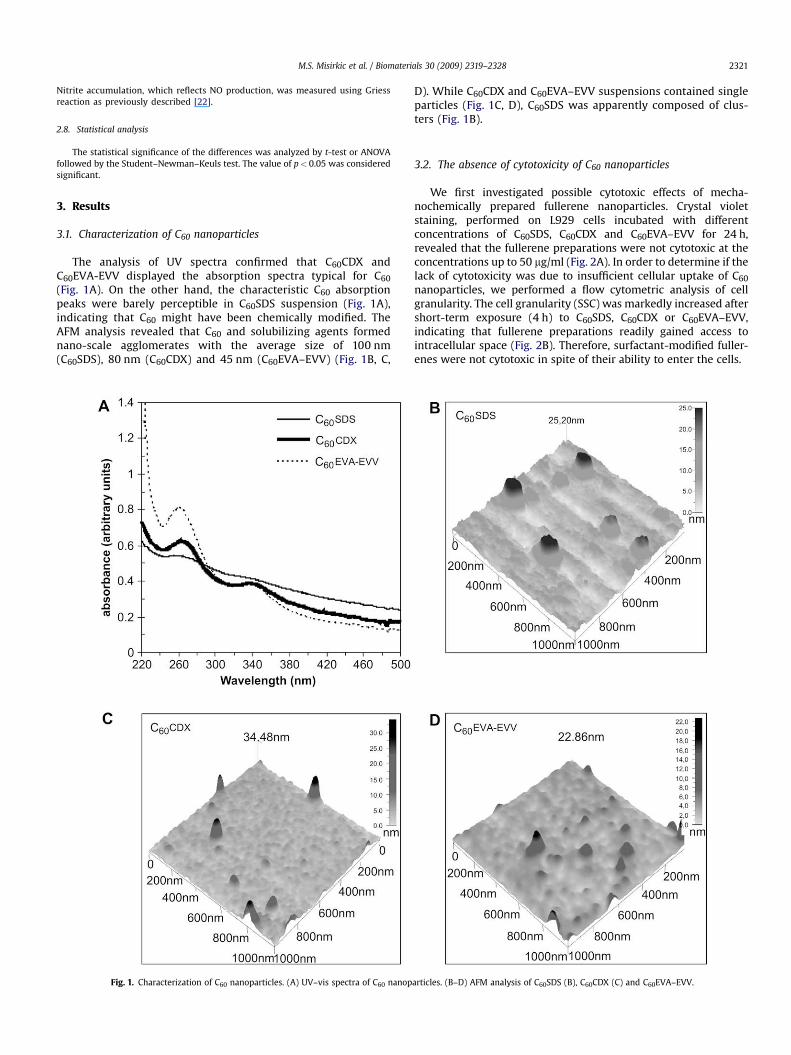
M.S. Misirkic et al. / Biomaterials 30 (2009) 2319–2328 2321
Nitrite accumulation, which reflects NO production, was measured using Griessreaction as previously described [22].
2.8. Statistical analysis
The statistical significance of the differences was analyzed by t-test or ANOVAfollowed by the Student–Newman–Keuls test. The value of p< 0.05 was consideredsignificant.
3. Results
3.1. Characterization of C60 nanoparticles
The analysis of UV spectra confirmed that C60CDX andC60EVA-EVV displayed the absorption spectra typical for C60
(Fig. 1A). On the other hand, the characteristic C60 absorptionpeaks were barely perceptible in C60SDS suspension (Fig. 1A),indicating that C60 might have been chemically modified. TheAFM analysis revealed that C60 and solubilizing agents formednano-scale agglomerates with the average size of 100 nm(C60SDS), 80 nm (C60CDX) and 45 nm (C60EVA–EVV) (Fig. 1B, C,
Fig. 1. Characterization of C60 nanoparticles. (A) UV–vis spectra of C60 nanop
D). While C60CDX and C60EVA–EVV suspensions contained singleparticles (Fig. 1C, D), C60SDS was apparently composed of clus-ters (Fig. 1B).
3.2. The absence of cytotoxicity of C60 nanoparticles
We first investigated possible cytotoxic effects of mecha-nochemically prepared fullerene nanoparticles. Crystal violetstaining, performed on L929 cells incubated with differentconcentrations of C60SDS, C60CDX and C60EVA–EVV for 24 h,revealed that the fullerene preparations were not cytotoxic at theconcentrations up to 50 mg/ml (Fig. 2A). In order to determine if thelack of cytotoxicity was due to insufficient cellular uptake of C60
nanoparticles, we performed a flow cytometric analysis of cellgranularity. The cell granularity (SSC) was markedly increased aftershort-term exposure (4 h) to C60SDS, C60CDX or C60EVA–EVV,indicating that fullerene preparations readily gained access tointracellular space (Fig. 2B). Therefore, surfactant-modified fuller-enes were not cytotoxic in spite of their ability to enter the cells.
articles. (B–D) AFM analysis of C60SDS (B), C60CDX (C) and C60EVA–EVV.
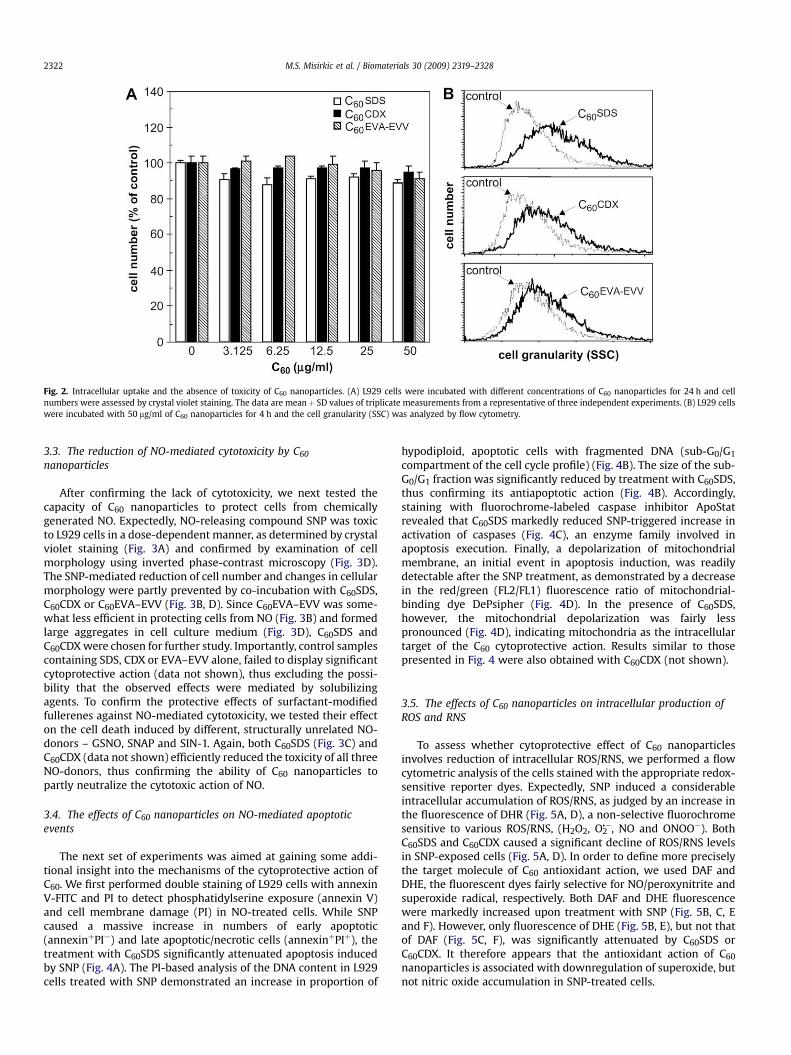
Fig. 2. Intracellular uptake and the absence of toxicity of C60 nanoparticles. (A) L929 cells were incubated with different concentrations of C60 nanoparticles for 24 h and cellnumbers were assessed by crystal violet staining. The data are meanþ SD values of triplicate measurements from a representative of three independent experiments. (B) L929 cellswere incubated with 50 mg/ml of C60 nanoparticles for 4 h and the cell granularity (SSC) was analyzed by flow cytometry.
M.S. Misirkic et al. / Biomaterials 30 (2009) 2319–23282322
3.3. The reduction of NO-mediated cytotoxicity by C60
nanoparticles
After confirming the lack of cytotoxicity, we next tested thecapacity of C60 nanoparticles to protect cells from chemicallygenerated NO. Expectedly, NO-releasing compound SNP was toxicto L929 cells in a dose-dependent manner, as determined by crystalviolet staining (Fig. 3A) and confirmed by examination of cellmorphology using inverted phase-contrast microscopy (Fig. 3D).The SNP-mediated reduction of cell number and changes in cellularmorphology were partly prevented by co-incubation with C60SDS,C60CDX or C60EVA–EVV (Fig. 3B, D). Since C60EVA–EVV was some-what less efficient in protecting cells from NO (Fig. 3B) and formedlarge aggregates in cell culture medium (Fig. 3D), C60SDS andC60CDX were chosen for further study. Importantly, control samplescontaining SDS, CDX or EVA–EVV alone, failed to display significantcytoprotective action (data not shown), thus excluding the possi-bility that the observed effects were mediated by solubilizingagents. To confirm the protective effects of surfactant-modifiedfullerenes against NO-mediated cytotoxicity, we tested their effecton the cell death induced by different, structurally unrelated NO-donors – GSNO, SNAP and SIN-1. Again, both C60SDS (Fig. 3C) andC60CDX (data not shown) efficiently reduced the toxicity of all threeNO-donors, thus confirming the ability of C60 nanoparticles topartly neutralize the cytotoxic action of NO.
3.4. The effects of C60 nanoparticles on NO-mediated apoptoticevents
The next set of experiments was aimed at gaining some addi-tional insight into the mechanisms of the cytoprotective action ofC60. We first performed double staining of L929 cells with annexinV-FITC and PI to detect phosphatidylserine exposure (annexin V)and cell membrane damage (PI) in NO-treated cells. While SNPcaused a massive increase in numbers of early apoptotic(annexinþPI�) and late apoptotic/necrotic cells (annexinþPIþ), thetreatment with C60SDS significantly attenuated apoptosis inducedby SNP (Fig. 4A). The PI-based analysis of the DNA content in L929cells treated with SNP demonstrated an increase in proportion of
hypodiploid, apoptotic cells with fragmented DNA (sub-G0/G1
compartment of the cell cycle profile) (Fig. 4B). The size of the sub-G0/G1 fraction was significantly reduced by treatment with C60SDS,thus confirming its antiapoptotic action (Fig. 4B). Accordingly,staining with fluorochrome-labeled caspase inhibitor ApoStatrevealed that C60SDS markedly reduced SNP-triggered increase inactivation of caspases (Fig. 4C), an enzyme family involved inapoptosis execution. Finally, a depolarization of mitochondrialmembrane, an initial event in apoptosis induction, was readilydetectable after the SNP treatment, as demonstrated by a decreasein the red/green (FL2/FL1) fluorescence ratio of mitochondrial-binding dye DePsipher (Fig. 4D). In the presence of C60SDS,however, the mitochondrial depolarization was fairly lesspronounced (Fig. 4D), indicating mitochondria as the intracellulartarget of the C60 cytoprotective action. Results similar to thosepresented in Fig. 4 were also obtained with C60CDX (not shown).
3.5. The effects of C60 nanoparticles on intracellular production ofROS and RNS
To assess whether cytoprotective effect of C60 nanoparticlesinvolves reduction of intracellular ROS/RNS, we performed a flowcytometric analysis of the cells stained with the appropriate redox-sensitive reporter dyes. Expectedly, SNP induced a considerableintracellular accumulation of ROS/RNS, as judged by an increase inthe fluorescence of DHR (Fig. 5A, D), a non-selective fluorochromesensitive to various ROS/RNS, (H2O2, O2
��, NO and ONOO�). BothC60SDS and C60CDX caused a significant decline of ROS/RNS levelsin SNP-exposed cells (Fig. 5A, D). In order to define more preciselythe target molecule of C60 antioxidant action, we used DAF andDHE, the fluorescent dyes fairly selective for NO/peroxynitrite andsuperoxide radical, respectively. Both DAF and DHE fluorescencewere markedly increased upon treatment with SNP (Fig. 5B, C, Eand F). However, only fluorescence of DHE (Fig. 5B, E), but not thatof DAF (Fig. 5C, F), was significantly attenuated by C60SDS orC60CDX. It therefore appears that the antioxidant action of C60
nanoparticles is associated with downregulation of superoxide, butnot nitric oxide accumulation in SNP-treated cells.
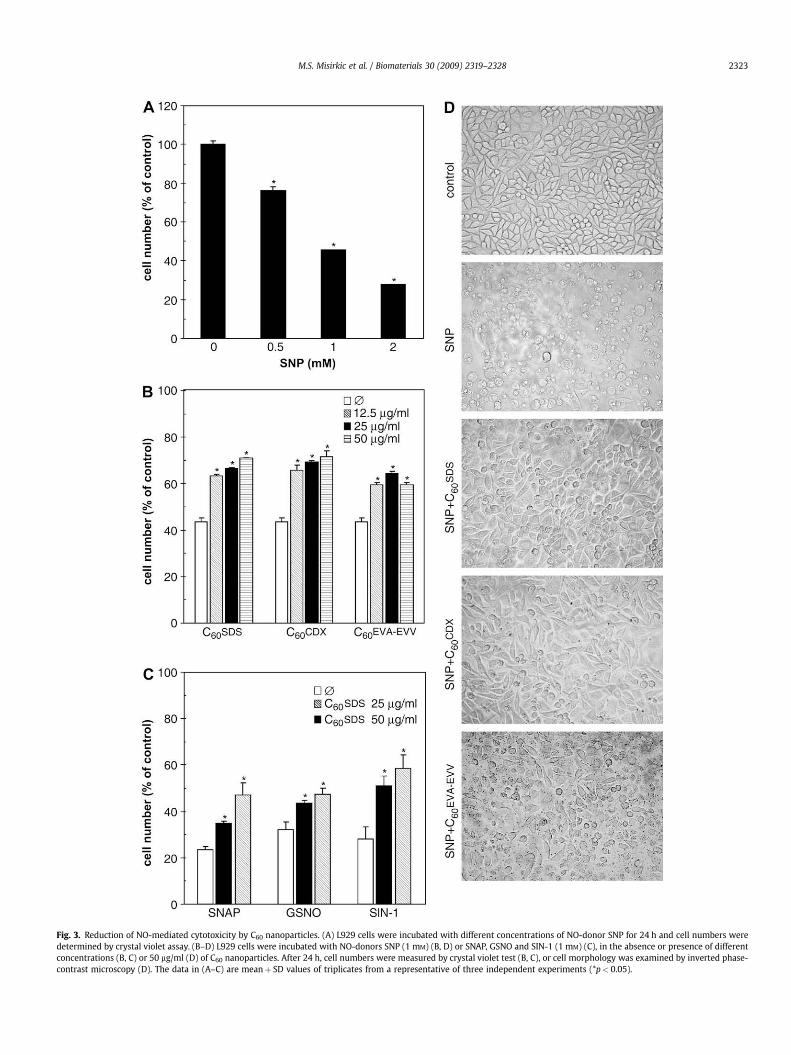
Fig. 3. Reduction of NO-mediated cytotoxicity by C60 nanoparticles. (A) L929 cells were incubated with different concentrations of NO-donor SNP for 24 h and cell numbers weredetermined by crystal violet assay. (B–D) L929 cells were incubated with NO-donors SNP (1 mM) (B, D) or SNAP, GSNO and SIN-1 (1 mM) (C), in the absence or presence of differentconcentrations (B, C) or 50 mg/ml (D) of C60 nanoparticles. After 24 h, cell numbers were measured by crystal violet test (B, C), or cell morphology was examined by inverted phase-contrast microscopy (D). The data in (A–C) are meanþ SD values of triplicates from a representative of three independent experiments (*p< 0.05).
M.S. Misirkic et al. / Biomaterials 30 (2009) 2319–2328 2323
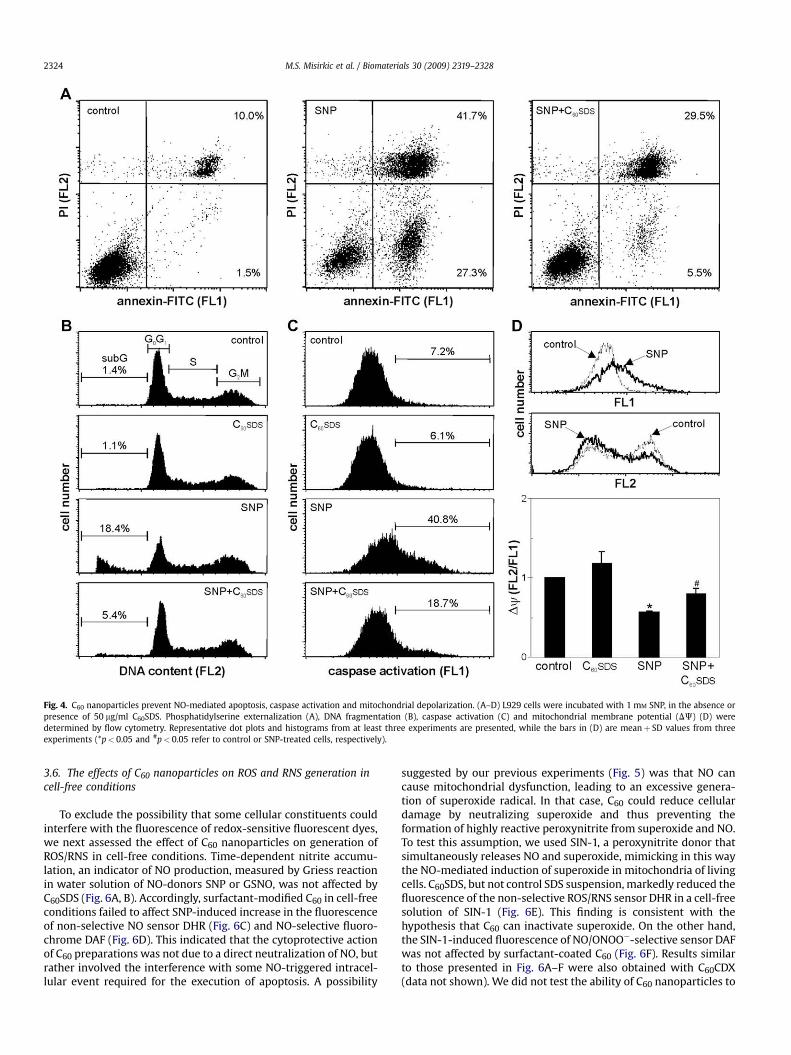
Fig. 4. C60 nanoparticles prevent NO-mediated apoptosis, caspase activation and mitochondrial depolarization. (A–D) L929 cells were incubated with 1 mM SNP, in the absence orpresence of 50 mg/ml C60SDS. Phosphatidylserine externalization (A), DNA fragmentation (B), caspase activation (C) and mitochondrial membrane potential (DJ) (D) weredetermined by flow cytometry. Representative dot plots and histograms from at least three experiments are presented, while the bars in (D) are meanþ SD values from threeexperiments (*p< 0.05 and #p< 0.05 refer to control or SNP-treated cells, respectively).
M.S. Misirkic et al. / Biomaterials 30 (2009) 2319–23282324
3.6. The effects of C60 nanoparticles on ROS and RNS generation incell-free conditions
To exclude the possibility that some cellular constituents couldinterfere with the fluorescence of redox-sensitive fluorescent dyes,we next assessed the effect of C60 nanoparticles on generation ofROS/RNS in cell-free conditions. Time-dependent nitrite accumu-lation, an indicator of NO production, measured by Griess reactionin water solution of NO-donors SNP or GSNO, was not affected byC60SDS (Fig. 6A, B). Accordingly, surfactant-modified C60 in cell-freeconditions failed to affect SNP-induced increase in the fluorescenceof non-selective NO sensor DHR (Fig. 6C) and NO-selective fluoro-chrome DAF (Fig. 6D). This indicated that the cytoprotective actionof C60 preparations was not due to a direct neutralization of NO, butrather involved the interference with some NO-triggered intracel-lular event required for the execution of apoptosis. A possibility
suggested by our previous experiments (Fig. 5) was that NO cancause mitochondrial dysfunction, leading to an excessive genera-tion of superoxide radical. In that case, C60 could reduce cellulardamage by neutralizing superoxide and thus preventing theformation of highly reactive peroxynitrite from superoxide and NO.To test this assumption, we used SIN-1, a peroxynitrite donor thatsimultaneously releases NO and superoxide, mimicking in this waythe NO-mediated induction of superoxide in mitochondria of livingcells. C60SDS, but not control SDS suspension, markedly reduced thefluorescence of the non-selective ROS/RNS sensor DHR in a cell-freesolution of SIN-1 (Fig. 6E). This finding is consistent with thehypothesis that C60 can inactivate superoxide. On the other hand,the SIN-1-induced fluorescence of NO/ONOO�-selective sensor DAFwas not affected by surfactant-coated C60 (Fig. 6F). Results similarto those presented in Fig. 6A–F were also obtained with C60CDX(data not shown). We did not test the ability of C60 nanoparticles to
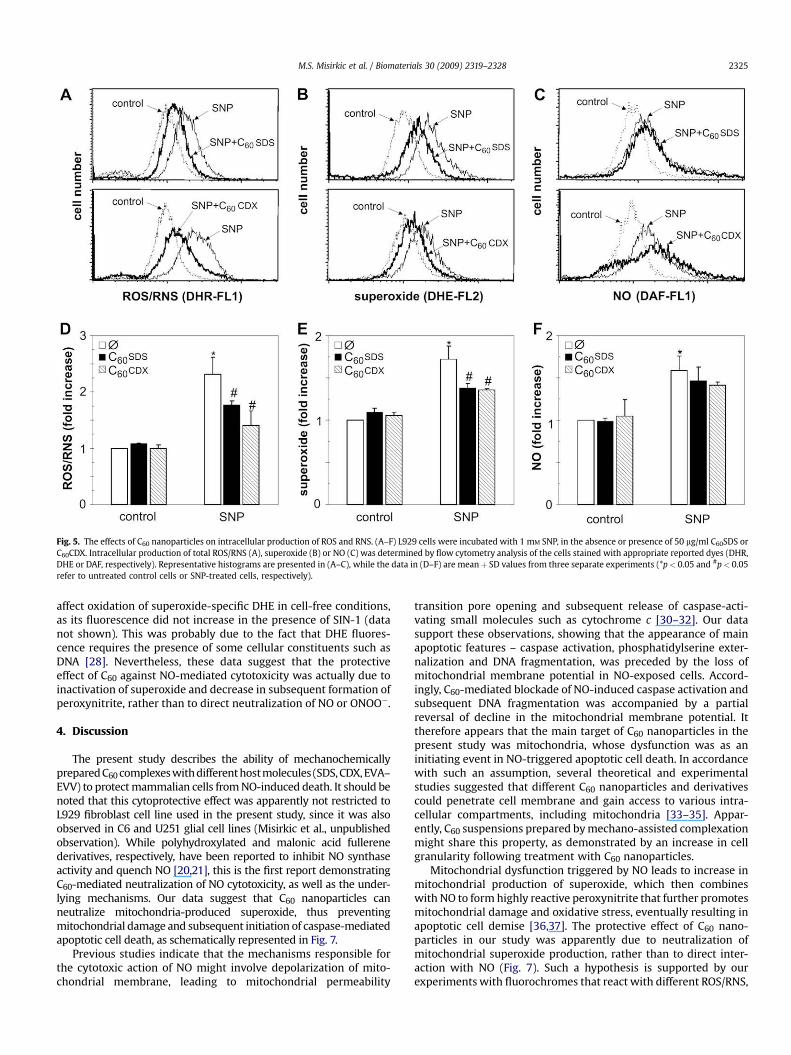
Fig. 5. The effects of C60 nanoparticles on intracellular production of ROS and RNS. (A–F) L929 cells were incubated with 1 mM SNP, in the absence or presence of 50 mg/ml C60SDS orC60CDX. Intracellular production of total ROS/RNS (A), superoxide (B) or NO (C) was determined by flow cytometry analysis of the cells stained with appropriate reported dyes (DHR,DHE or DAF, respectively). Representative histograms are presented in (A–C), while the data in (D–F) are meanþ SD values from three separate experiments (*p< 0.05 and #p< 0.05refer to untreated control cells or SNP-treated cells, respectively).
M.S. Misirkic et al. / Biomaterials 30 (2009) 2319–2328 2325
affect oxidation of superoxide-specific DHE in cell-free conditions,as its fluorescence did not increase in the presence of SIN-1 (datanot shown). This was probably due to the fact that DHE fluores-cence requires the presence of some cellular constituents such asDNA [28]. Nevertheless, these data suggest that the protectiveeffect of C60 against NO-mediated cytotoxicity was actually due toinactivation of superoxide and decrease in subsequent formation ofperoxynitrite, rather than to direct neutralization of NO or ONOO�.
4. Discussion
The present study describes the ability of mechanochemicallyprepared C60 complexes with different hostmolecules (SDS, CDX, EVA–EVV) to protect mammalian cells from NO-induced death. It should benoted that this cytoprotective effect was apparently not restricted toL929 fibroblast cell line used in the present study, since it was alsoobserved in C6 and U251 glial cell lines (Misirkic et al., unpublishedobservation). While polyhydroxylated and malonic acid fullerenederivatives, respectively, have been reported to inhibit NO synthaseactivity and quench NO [20,21], this is the first report demonstratingC60-mediated neutralization of NO cytotoxicity, as well as the under-lying mechanisms. Our data suggest that C60 nanoparticles canneutralize mitochondria-produced superoxide, thus preventingmitochondrial damage and subsequent initiation of caspase-mediatedapoptotic cell death, as schematically represented in Fig. 7.
Previous studies indicate that the mechanisms responsible forthe cytotoxic action of NO might involve depolarization of mito-chondrial membrane, leading to mitochondrial permeability
transition pore opening and subsequent release of caspase-acti-vating small molecules such as cytochrome c [30–32]. Our datasupport these observations, showing that the appearance of mainapoptotic features – caspase activation, phosphatidylserine exter-nalization and DNA fragmentation, was preceded by the loss ofmitochondrial membrane potential in NO-exposed cells. Accord-ingly, C60-mediated blockade of NO-induced caspase activation andsubsequent DNA fragmentation was accompanied by a partialreversal of decline in the mitochondrial membrane potential. Ittherefore appears that the main target of C60 nanoparticles in thepresent study was mitochondria, whose dysfunction was as aninitiating event in NO-triggered apoptotic cell death. In accordancewith such an assumption, several theoretical and experimentalstudies suggested that different C60 nanoparticles and derivativescould penetrate cell membrane and gain access to various intra-cellular compartments, including mitochondria [33–35]. Appar-ently, C60 suspensions prepared by mechano-assisted complexationmight share this property, as demonstrated by an increase in cellgranularity following treatment with C60 nanoparticles.
Mitochondrial dysfunction triggered by NO leads to increase inmitochondrial production of superoxide, which then combineswith NO to form highly reactive peroxynitrite that further promotesmitochondrial damage and oxidative stress, eventually resulting inapoptotic cell demise [36,37]. The protective effect of C60 nano-particles in our study was apparently due to neutralization ofmitochondrial superoxide production, rather than to direct inter-action with NO (Fig. 7). Such a hypothesis is supported by ourexperiments with fluorochromes that react with different ROS/RNS,
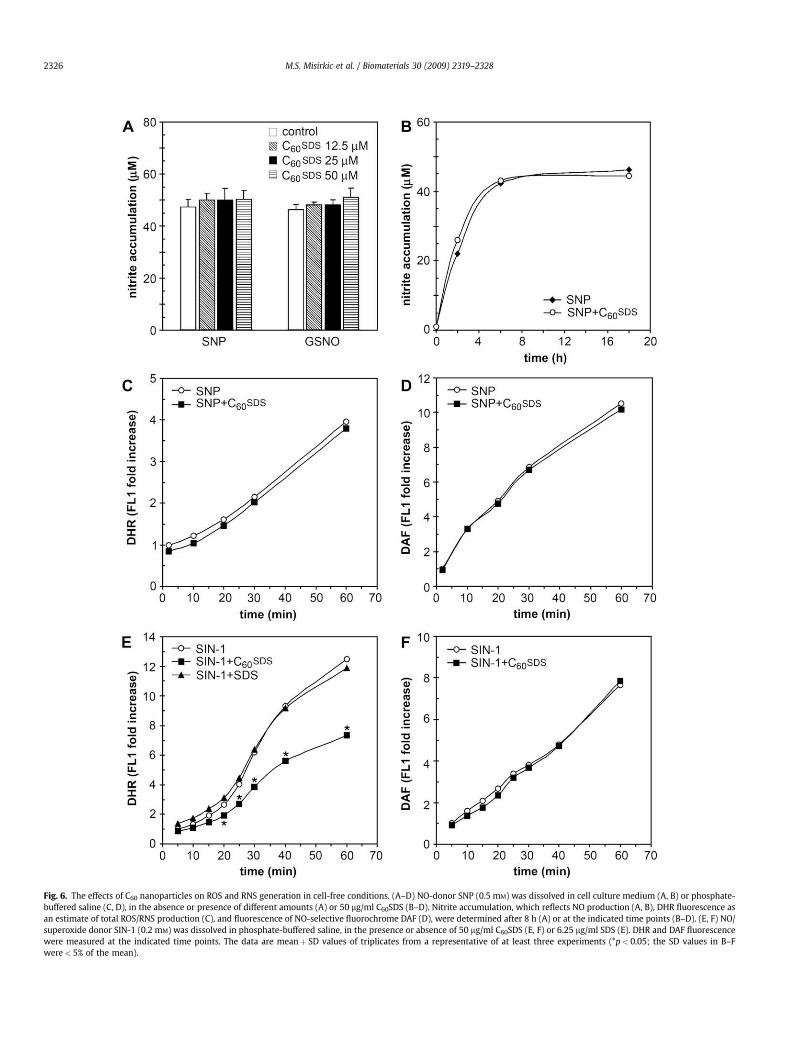
Fig. 6. The effects of C60 nanoparticles on ROS and RNS generation in cell-free conditions. (A–D) NO-donor SNP (0.5 mM) was dissolved in cell culture medium (A, B) or phosphate-buffered saline (C, D), in the absence or presence of different amounts (A) or 50 mg/ml C60SDS (B–D). Nitrite accumulation, which reflects NO production (A, B), DHR fluorescence asan estimate of total ROS/RNS production (C), and fluorescence of NO-selective fluorochrome DAF (D), were determined after 8 h (A) or at the indicated time points (B–D). (E, F) NO/superoxide donor SIN-1 (0.2 mM) was dissolved in phosphate-buffered saline, in the presence or absence of 50 mg/ml C60SDS (E, F) or 6.25 mg/ml SDS (E). DHR and DAF fluorescencewere measured at the indicated time points. The data are meanþ SD values of triplicates from a representative of at least three experiments (*p< 0.05; the SD values in B–Fwere< 5% of the mean).
M.S. Misirkic et al. / Biomaterials 30 (2009) 2319–23282326

Fig. 7. A hypothetical mechanism of C60 nanoparticle-mediated cell protection fromNO-induced apoptosis.
M.S. Misirkic et al. / Biomaterials 30 (2009) 2319–2328 2327
showing that both C60SDS and C60CDX were able to reduce thelevels of intracellular and chemically generated superoxide. Thesedata are consistent with the findings that both O2
�� and �OH wereefficiently scavenged by polyvinylpirrolidone- or CDX-solubilizedC60 in the cell-free systems [16,17]. The putative C60-mediatedneutralization of superoxide in NO-exposed cells presumably pre-vented O2
�� reaction with NO and subsequent generation of highlyreactive, cell-damaging peroxynitrite (ONOO�) (Fig. 7). On theother hand, C60 nanoparticles did not directly interact with eitherintracellular NO or that produced by NO-releasing agents in cell-free conditions. While this apparently contradicts the datadescribing the ability of polyhydroxylated C60 to readily scavengeSNP-generated NO in cell-free conditions [21], the reasons for thedifferent behavior of mechanochemically solubilized C60 in ourstudy remain to be resolved.
While previous and our data confirm the antioxidant capacity ofdifferent C60 preparations, the precise explanation of the mecha-nisms underlying C60-mediated ROS neutralization is currentlylacking. Although it has been described that up to 34 benzyl radicalscan readily add to the fullerene cage [38], such a direct interactionhas not been reported for ROS [1]. Interestingly, the O2
�� removal bymalonic acid-derivatized C60 was not via stoichiometric ‘‘scav-enging,’’ as expected, but through catalytic dismutation of super-oxide, in which electron-deficient regions on the C60 sphere workedin concert with the attached malonyl groups to electrostaticallyguide and stabilize superoxide, thus promoting dismutation [39].Accordingly, some sort of cooperation between the solubilizingagent and the embedded C60 might be envisaged to explain ROSneutralization in our study. This is consistent with the relativelyhigh ROS-quenching capacity of both SDS and CDX [40], and it haseven been proposed that a substantial part of �OH quenching byCDX-C60 host-guest complex can be attributed to the macromo-lecular host [41]. In our hands, however, neither CDX nor SDS atrelevant concentrations were able to mimic C60 nanoparticle-mediated ROS/RNS neutralization and subsequent cell protection,thus confirming that the host alone was not responsible for theseactions in our experimental system. While the UV–vis analysisindicates that C60 was not chemically modified by complexationwith CDX or EVA–EVV, the disappearance of typical UV absorptionpeaks in C60SDS suggests a chemical reaction between C60 and the
surfactant. If one assumes that alkyl chain part of surfactantmolecules lies on the surface of fullerene cage, then it is energeti-cally favourable for the chains to bend around the cage whenadsorbing onto small-diameter fullerene surface. In this way, vander Waals interactions are established among alkyl chains andfullerene surface, while ionic sulfate part of surfactant moleculeinteracts with water. Similar model has been proposed for SDS-coated nanotubes dispersed in water [42]. We are currentlyinvestigating the possible contribution of C60 interactions with thehost molecules to ROS-scavenging activity of C60 nanoparticles, aswell as the mechanisms underlying these interactions.
5. Conclusions
The present report describes the ability of the C60 mecha-nochemically embedded in different solubilizing agents, to protectmammalian cells against in vitro cytotoxicity of the biologicallyrelevant free radical NO. The cytoprotective action of C60 nano-particles was not mediated by direct interaction with NO, but ratherthrough neutralization of mitochondria-derived superoxide radicalanion. As a consequence, generation of peroxynitrite, as well assubsequent mitochondrial damage, caspase activation andapoptotic cell death were all reduced in C60-treated cells. In light ofthe proposed role of NO in causing tissue destruction duringexcessive inflammation, our findings suggest that C60 complexeswith appropriate host molecules might have potential for furtherdevelopment as therapeutic agents against RNS/ROS-mediatedcellular injury in various inflammatory/autoimmune disorders.
Acknowledgements
This work was supported by the Ministry of Science of theRepublic of Serbia (grant no. 145073).
References
[1] Markovic Z, Trajkovic V. Biomedical potential of the reactive oxygen speciesgeneration and quenching by fullerenes (C60). Biomaterials 2008;29:3561–73.
[2] Dugan LL, Turetsky DM, Du C, Lobner D, Wheeler M, Almli CR, et al. Carboxy-fullerenes as neuroprotective agents. Proc Natl Acad Sci USA 1997;94:9434–9.
[3] Dugan LL, Gabrielsen JK, Yu SP, Lin TS, Choi DW. Buckminsterfullerenol freeradical scavengers reduce excitotoxic and apoptotic death of cultured corticalneurons. Neurobiol Dis 1996;3:129–35.
[4] Jin H, Chen WQ, Tang XW, Chiang LY, Yang CY, Schloss JV, et al. Poly-hydroxylated C60, fullerenols, as glutamate receptor antagonists and neuro-protective agents. J Neurosci Res 2000;62:600–7.
[5] Lin AM, Chyi BY, Wang SD, Yu HH, Kanakamma PP, Luh TY, et al. Carboxy-fullerene prevents iron-induced oxidative stress in rat brain. J Neurochem1999;72:1634–40.
[6] Monti D, Moretti L, Salvioli S, Straface E, Malorni W, Pellicciari R, et al. C60carboxyfullerene exerts a protective activity against oxidative stress-inducedapoptosis in human peripheral blood mononuclear cells. Biochem Biophys ResCommun 2000;277:711–7.
[7] Isakovic A, Markovic Z, Todorovic-Markovic B, Nikolic N, Vranjes-Djuric S,Mirkovic M, et al. Distinct cytotoxic mechanisms of pristine versus hydroxy-lated fullerene. Toxicol Sci 2006;91:173–83.
[8] Sayes C, Fortner J, Lyon D, Boyd A, Ausman K, Tao Y, et al. The differentialcytotoxicity of water-soluble fullerenes. Nano Lett 2004;4:1881–7.
[9] Sayes CM, Gobin AM, Ausman KD, Mendez J, West JL, Colvin VL. Nano-C60cytotoxicity is due to lipid peroxidation. Biomaterials 2005;26:7587–95.
[10] Oberdorster E. Manufactured nanomaterials (fullerenes, C60) induce oxidativestress in the brain of juvenile largemouth bass. Environ Health Perspect2004;112:1058–62.
[11] Markovic Z, Todorovic-Markovic B, Kleut D, Nikolic N, Vranjes-Djuric S,Misirkic M, et al. The mechanism of cell-damaging reactive oxygen generationby colloidal fullerenes. Biomaterials 2007;28:5437–48.
[12] Isakovic A, Markovic Z, Nikolic N, Todorovic-Markovic B, Vranjes-Djuric S,Harhaji L, et al. Inactivation of nanocrystalline C60 cytotoxicity by gamma-irradiation. Biomaterials 2006;27:5049–58.
[13] Henry TB, Menn FM, Fleming JT, Wilgus J, Compton RN, Sayler GS. Attributingeffects of aqueous C60 nano-aggregates to tetrahydrofuran decompositionproducts in larval zebrafish by assessment of gene expression. Environ HealthPerspect 2007;115:1059–65.

M.S. Misirkic et al. / Biomaterials 30 (2009) 2319–23282328
[14] Oberdorster E, Zhu S, Blickley TM, McClellan-Green P, Haasch ML. Ecotox-icology of carbon-based engineered nanoparticles: effects of fullerene (C60) onaquatic organisms. Carbon 2006;44:1112–20.
[15] Gharbi N, Pressac M, Hadchouel M, Szwarc H, Wilson SR, Moussa F.[60]Fullerene is a powerful antioxidant in vivo with no acute or subacutetoxicity. Nano Lett 2005;5:2578–85.
[16] Xiao L, Takada H, Maeda K, Haramoto M, Miwa N. Antioxidant effects of water-soluble fullerene derivatives against ultraviolet ray or peroxylipid throughtheir action of scavenging the reactive oxygen species in human skin kerati-nocytes. Biomed Pharmacother 2005;59:351–8.
[17] Takada H, Kokubo K, Matsubayashi K, Oshima T. Antioxidant activity ofsupramolecular water-soluble fullerenes evaluated by beta-carotene bleachingassay. Biosci Biotechnol Biochem 2006;70:3088–93.
[18] Thippeswamy T, McKay JS, Quinn JP, Morris R. Nitric oxide, a biological double-faced janus–is this good or bad? Histol Histopathol 2006;21:445–58.
[19] Brown GC, Borutaite V. Nitric oxide inhibition of mitochondrial respirationand its role in cell death. Free Radic Biol Med 2002;33:1440–50.
[20] Wolff DJ, Papoiu AD, Mialkowski K, Richardson CF, Schuster DI, Wilson SR.Inhibition of nitric oxide synthase isoforms by tris-malonyl-C60-fullereneadducts. Arch Biochem Biophys 2000;378:216–23.
[21] Mirkov SM, Djordjevic AN, Andric NL, Andric SA, Kostic TS, Bogdanovic GM,et al. Nitric oxide-scavenging activity of polyhydroxylated fullerenol,C60(OH)24. Nitric Oxide 2004;11:201–7.
[22] Harhaji L, Vuckovic O, Miljkovic D, Stosic-Grujicic S, Trajkovic V. Iron down-regulates macrophage anti-tumour activity by blocking nitric oxide produc-tion. Clin Exp Immunol 2004;137:109–16.
[23] Mijatovic S, Maksimovic-Ivanic D, Radovic J, Popadic D, Momcilovic M,Harhaji L, et al. Aloe-emodin prevents cytokine-induced tumor cell death: theinhibition of auto-toxic nitric oxide release as a potential mechanism. Cell MolLife Sci 2004;61:1805–15.
[24] Harhaji L, Popadic D, Miljkovic D, Cvetkovic I, Isakovic A, Trajkovic V. Acidosisaffects tumor cell survival through modulation of nitric oxide release. FreeRadic Biol Med 2006;40:226–35.
[25] Mijatovic S, Maksimovic-Ivanic D, Radovic J, Miljkovic D, Kaludjerovic GN,Sabo TJ, et al. Aloe emodin decreases the ERK-dependent anticancer activity ofcisplatin. Cell Mol Life Sci 2005;62:1275–82.
[26] Bhattarai SR, Kc RB, Kim SY, Sharma M, Khil MS, Hwang PH, et al. N-hexanoylchitosan stabilized magnetic nanoparticles: implication for cellular labelingand magnetic resonance imaging. J Nanobiotechnol 2008;6:1.
[27] Hempel SL, Buettner GR, O’Malley YQ, Wessels DA, Flaherty DM. Dihydro-fluorescein diacetate is superior for detecting intracellular oxidants:comparison with 20 ,70-dichlorodihydrofluorescein diacetate, 5(and 6)-car-boxy-20 ,70-dichlorodihydrofluorescein diacetate, and dihydrorhodamine 123.Free Radic Biol Med 1999;27:146–59.
[28] Zhao H, Kalivendi S, Zhang H, Joseph J, Nithipatikom K, Vasquez-Vivar J, et al.Superoxide reacts with hydroethidine but forms a fluorescent product that isdistinctly different from ethidium: potential implications in intracellularfluorescence detection of superoxide. Free Radic Biol Med 2003;34:1359–68.
[29] Roychowdhury S, Luthe A, Keilhoff G, Wolf G, Horn TF. Oxidative stress in glialcultures: detection by DAF-2 fluorescence used as a tool to measure peroxy-nitrite rather than nitric oxide. Glia 2002;38:103–14.
[30] Hortelano S, Dallaporta B, Zamzami N, Hirsch T, Susin SA, Marzo I, et al. Nitricoxide induces apoptosis via triggering mitochondrial permeability transition.FEBS Lett 1997;410:373–7.
[31] Ushmorov A, Ratter F, Lehmann V, Droge W, Schirrmacher V, Umansky V.Nitric-oxide-induced apoptosis in human leukemic lines requires mitochon-drial lipid degradation and cytochrome c release. Blood 1999;93:2342–52.
[32] Bal-Price A, Brown GC. Nitric-oxide-induced necrosis and apoptosis in PC12cells mediated by mitochondria. J Neurochem 2000;75:1455–64.
[33] Qiao R, Roberts AP, Mount AS, Klaine SJ, Ke PC. Translocation of C60 and itsderivatives across a lipid bilayer. Nano Lett 2007;7:614–9.
[34] Porter AE, Muller K, Skepper J, Midgley P, Welland M. Uptake of C60 by humanmonocyte macrophages, its localization and implications for toxicity: studiedby high resolution electron microscopy and electron tomography. Acta Bio-mater 2006;2:409–19.
[35] Foley S, Crowley C, Smaihi M, Bonfils C, Erlanger BF, Seta P, et al. Cellularlocalisation of a water-soluble fullerene derivative. Biochem Biophys ResCommun 2002;294:116–9.
[36] Keller JN, Kindy MS, Holtsberg FW, St Clair DK, Yen HC, Germeyer A, et al.Mitochondrial manganese superoxide dismutase prevents neural apoptosisand reduces ischemic brain injury: suppression of peroxynitrite production,lipid peroxidation, and mitochondrial dysfunction. J Neurosci 1998;18:687–97.
[37] Ghafourifar P, Schenk U, Klein SD, Richter C. Mitochondrial nitric-oxide syn-thase stimulation causes cytochrome c release from isolated mitochondria.Evidence for intramitochondrial peroxynitrite formation. J Biol Chem1999;274:31185–8.
[38] Krusic PJ, Wasserman E, Keizer PN, Morton JR, Preston KF. Radical reactions ofC60. Science 1991;254:1183–5.
[39] Ali SS, Hardt JI, Quick KL, Kim-Han JS, Erlanger BF, Huang TT, et al. A biolog-ically effective fullerene (C60) derivative with superoxide dismutase mimeticproperties. Free Radic Biol Med 2004;37:1191–202.
[40] The Radiation Chemistry Data Center of the Notre Dame Radiation Laboratory,http://www.rcdc.nd.edu.
[41] Guldi DM, Asmus KD. Activity of water-soluble fullerenes towards �OH-radicalsand molecular oxygen. Radiat Phys Chem 1999;56:449–56.
[42] Islam MF, Rojas E, Bergey DM, Johnson AT, Yodh AG. High weight fractionsurfactant solubilization of single-wall carbon nanotubes in water. Nano Lett2003;3:269–73.




















