
The N-terminal part of Als1 protein from Candida albicans specifically binds fucose-containing...
13
The N-terminal part of Als1 protein from Candida albicans specifically binds fucose-containing glycansDagmara S. Donohue, Francesco S. Ielasi, Katty V. Y. Goossens and Ronnie G. Willaert* Laboratory of Structural Biology, Vrije Universiteit Brussel, Pleinlaan 2, 1050 Brussels, Belgium. Summary The opportunistic pathogen Candida albicans expresses on its surface Als (Agglutinin like sequence) proteins, which play an important role in the adhesion to host cells and in the development of candidiasis. The binding specificity of these proteins is broad, as they can bind to various mammalian pro- teins, such as extracellular matrix proteins, and N- and E-cadherins. The N-terminal part of Als proteins con- stitutes the substrate-specific binding domain and is responsible for attachment to epithelial and endothe- lial cells. We have used glycan array screening to identify possible glycan receptors for the binding domain of Als1p-N. Under those conditions, Als1p-N binds specifically to fucose-containing glycans, which adds a lectin function to the functional diversity of the Als1 protein. The binding between Als1p-N and BSA- fucose glycoconjugate was quantitatively charac- terized using surface plasmon resonance, which dem- onstrated a weak millimolar affinity between Als1p-N and fucose. Furthermore, we have also quantified the affinity of Als1p-N to the extracellular matrix proteins proteins fibronectin and laminin, which is situated in the micromolar range. Surface plasmon resonance characterization of Als1p-N–Als1p-N interaction was in the micromolar affinity range. Introduction Candida albicans occurs naturally as a part of the human gastrointestinal flora. In a healthy body, with well- maintained homeostasis, this commensal interaction remains unnoticed. However, when the balance between the immune system and C. albicans is disturbed, com- mensalism may turn into fungal infection (candidiasis) (Mavor et al., 2005; Rupp, 2007). Adherence is an essen- tial determinant of pathogenesis, as it allows C. albicans to attach to host cells and to form biofilms that protect the yeast cells from unfavourable conditions. Candida albicans has developed multiple ways to colo- nize and infect host cells and tissues. One such mecha- nism is the specific ligand–receptor interaction through a whole range of adhesins displayed on the yeast cell wall (Chaffin et al., 1998; Sundstrom, 2002; Filler, 2006). These cell wall proteins are capable of recognizing protein ligands (reviewed in Chaffin et al., 1998), glycolipids (Ghannoum et al., 1986, Jimenez-Lucho et al., 1990; Yu et al., 1994; Cameron and Douglas, 1996) and carbohydrates on mam- malian cells (Sandin et al., 1982; Critchley and Douglas, 1987a; Macura and Tondyra, 1989; Brassart et al., 1991). The ALS gene family of C. albicans encodes eight known large cell surface glycoproteins that are involved in adhesion to mammalian cells (Fu et al., 1998; Sheppard et al., 2004; Hoyer et al., 2008; Zhu and Filler, 2010). Each Als protein possesses an N-terminal region with the secretory signal peptide, followed by a threonine-rich region, then a highly glycosylated central domain rich in tandem repeats and a C-terminus with a glycosylphos- phatidylinositol anchor (Hoyer, 2001; Frank et al., 2010). Homology modelling and experimental studies indicate that the N-terminal part of the Als proteins contain domains that belong to the immunoglobulin superfamily (Sheppard et al., 2004). Each Ig-like domain consists of two sheets of seven stranded anti-parallel b-strands. Microorganisms commonly use this type of structure for cell–cell recognition and signalling functions (Holmgren et al., 1992; Barclay, 2003). The Als1 protein has been shown to be abundantly expressed in various candidiasis models, where it was able to bind to epithelial and endothelial cells (Fu et al., 2002; Kamai et al., 2002; Green et al., 2004; 2006; Sheppard et al., 2004; Cheng et al., 2005). Experimental data on Als1p show that the substrate-specific binding domain is located within the N-terminus (Loza et al., 2004; Sheppard et al., 2004; Phan et al., 2007). It was demonstrated that different Als proteins can bind to extracellular matrix proteins (ECM), epithelial and endot- helial cells (Sheppard et al., 2004). It has been shown using C. albicans and Saccharomyces cerevisiae expressing Als proteins that both Als1p and Als5p rec- ognize peptides containing the structural motif tj+ (a Accepted 19 April, 2011. *For correspondence. E-mail ronnie. [email protected]; Tel. (+32) 2629 1846; Fax (+32) 2629 1963. Molecular Microbiology (2011) doi:10.1111/j.1365-2958.2011.07676.x © 2011 Blackwell Publishing Ltd
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The N-terminal part of Als1 protein from Candida albicans specifically binds fucose-containing...

The N-terminal part of Als1 protein from Candida albicansspecifically binds fucose-containing glycansmmi_7676 1..13
Dagmara S. Donohue, Francesco S. Ielasi,Katty V. Y. Goossens and Ronnie G. Willaert*Laboratory of Structural Biology, Vrije UniversiteitBrussel, Pleinlaan 2, 1050 Brussels, Belgium.
Summary
The opportunistic pathogen Candida albicansexpresses on its surface Als (Agglutinin likesequence) proteins, which play an important role in theadhesion to host cells and in the development ofcandidiasis. The binding specificity of these proteinsis broad, as they can bind to various mammalian pro-teins, such as extracellular matrix proteins, and N- andE-cadherins. The N-terminal part of Als proteins con-stitutes the substrate-specific binding domain and isresponsible for attachment to epithelial and endothe-lial cells. We have used glycan array screening toidentify possible glycan receptors for the bindingdomain of Als1p-N. Under those conditions, Als1p-Nbinds specifically to fucose-containing glycans, whichadds a lectin function to the functional diversity of theAls1 protein. The binding between Als1p-N and BSA-fucose glycoconjugate was quantitatively charac-terized using surface plasmon resonance, which dem-onstrated a weak millimolar affinity between Als1p-Nand fucose. Furthermore, we have also quantified theaffinity of Als1p-N to the extracellular matrix proteinsproteins fibronectin and laminin, which is situated inthe micromolar range. Surface plasmon resonancecharacterization of Als1p-N–Als1p-N interaction wasin the micromolar affinity range.
Introduction
Candida albicans occurs naturally as a part of the humangastrointestinal flora. In a healthy body, with well-maintained homeostasis, this commensal interactionremains unnoticed. However, when the balance betweenthe immune system and C. albicans is disturbed, com-mensalism may turn into fungal infection (candidiasis)(Mavor et al., 2005; Rupp, 2007). Adherence is an essen-
tial determinant of pathogenesis, as it allows C. albicansto attach to host cells and to form biofilms that protect theyeast cells from unfavourable conditions.
Candida albicans has developed multiple ways to colo-nize and infect host cells and tissues. One such mecha-nism is the specific ligand–receptor interaction through awhole range of adhesins displayed on the yeast cell wall(Chaffin et al., 1998; Sundstrom, 2002; Filler, 2006). Thesecell wall proteins are capable of recognizing protein ligands(reviewed in Chaffin et al., 1998), glycolipids (Ghannoumet al., 1986, Jimenez-Lucho et al., 1990; Yu et al., 1994;Cameron and Douglas, 1996) and carbohydrates on mam-malian cells (Sandin et al., 1982; Critchley and Douglas,1987a; Macura and Tondyra, 1989; Brassart et al., 1991).
The ALS gene family of C. albicans encodes eightknown large cell surface glycoproteins that are involved inadhesion to mammalian cells (Fu et al., 1998; Sheppardet al., 2004; Hoyer et al., 2008; Zhu and Filler, 2010).Each Als protein possesses an N-terminal region with thesecretory signal peptide, followed by a threonine-richregion, then a highly glycosylated central domain rich intandem repeats and a C-terminus with a glycosylphos-phatidylinositol anchor (Hoyer, 2001; Frank et al., 2010).Homology modelling and experimental studies indicatethat the N-terminal part of the Als proteins containdomains that belong to the immunoglobulin superfamily(Sheppard et al., 2004). Each Ig-like domain consists oftwo sheets of seven stranded anti-parallel b-strands.Microorganisms commonly use this type of structure forcell–cell recognition and signalling functions (Holmgrenet al., 1992; Barclay, 2003).
The Als1 protein has been shown to be abundantlyexpressed in various candidiasis models, where it wasable to bind to epithelial and endothelial cells (Fu et al.,2002; Kamai et al., 2002; Green et al., 2004; 2006;Sheppard et al., 2004; Cheng et al., 2005). Experimentaldata on Als1p show that the substrate-specific bindingdomain is located within the N-terminus (Loza et al.,2004; Sheppard et al., 2004; Phan et al., 2007). It wasdemonstrated that different Als proteins can bind toextracellular matrix proteins (ECM), epithelial and endot-helial cells (Sheppard et al., 2004). It has been shownusing C. albicans and Saccharomyces cerevisiaeexpressing Als proteins that both Als1p and Als5p rec-ognize peptides containing the structural motif tj+ (a
Accepted 19 April, 2011. *For correspondence. E-mail [email protected]; Tel. (+32) 2629 1846; Fax (+32) 2629 1963.
Molecular Microbiology (2011) � doi:10.1111/j.1365-2958.2011.07676.x
© 2011 Blackwell Publishing Ltd
turn-like residue, a bulky hydrophobic residue, and Lysor Arg), which can be found in many proteins (Klotzet al., 2004). However, Als1p does not recognize somepeptides recognized by Als5p. This suggests that differ-ent Als proteins have preference towards differentsequences within the ligands. It has also been shownthat the N-terminal part of Als3 and Als1 proteins bind toepithelial cell E-cadherin and endothelial cell N-cadherinand this induces endocytosis by host cells (Phan et al.,2007).
Host–pathogen interactions depend very often oncarbohydrate–lectin interactions. The presence of alectin-like adhesin with specifity for L-fucose or N-acetylglucosamine (GlcNAc) has been previouslydescribed for C. albicans adhesion, but this lectin activitywas not attributed to any specific protein found on the cellwall of C. albicans (Critchley and Douglas, 1987b; Toshand Douglas, 1992; Vardar-Ünlü et al., 1998).
The goal of this study was to determine if theN-terminal, substrate-specific part of Als1p possesses anaffinity to carbohydrates and thus has lectin activity.Therefore, the N-terminal part of Als1p (Als1p-N) wasoverexpressed in S. cerevisiae, purified, and used in aglycan microarray screening. This experiment revealedthe specificity of Als1p-N towards glycans containing
fucose. Surface plasmon resonance (SPR) experimentsconfirmed the specific interaction between Als1p-N andfucose, which bound to each other with millimolar affinity.Additionally, the Als1p-N interactions with fibronectin andlaminin as well as Als1p-N–Als1p-N interactions werequantitatively investigated using SPR.
Results
Als1p-N binds to fibronectin and laminin withmicromolar affinity
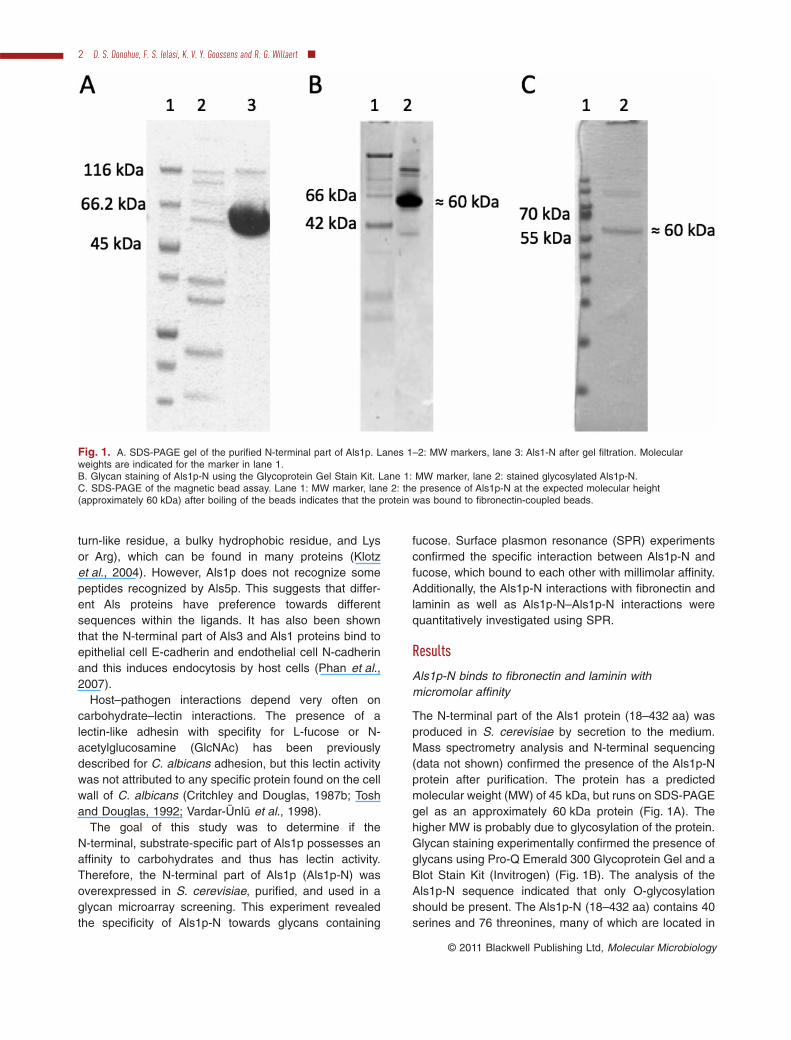
The N-terminal part of the Als1 protein (18–432 aa) wasproduced in S. cerevisiae by secretion to the medium.Mass spectrometry analysis and N-terminal sequencing(data not shown) confirmed the presence of the Als1p-Nprotein after purification. The protein has a predictedmolecular weight (MW) of 45 kDa, but runs on SDS-PAGEgel as an approximately 60 kDa protein (Fig. 1A). Thehigher MW is probably due to glycosylation of the protein.Glycan staining experimentally confirmed the presence ofglycans using Pro-Q Emerald 300 Glycoprotein Gel and aBlot Stain Kit (Invitrogen) (Fig. 1B). The analysis of theAls1p-N sequence indicated that only O-glycosylationshould be present. The Als1p-N (18–432 aa) contains 40serines and 76 threonines, many of which are located in
Fig. 1. A. SDS-PAGE gel of the purified N-terminal part of Als1p. Lanes 1–2: MW markers, lane 3: Als1-N after gel filtration. Molecularweights are indicated for the marker in lane 1.B. Glycan staining of Als1p-N using the Glycoprotein Gel Stain Kit. Lane 1: MW marker, lane 2: stained glycosylated Als1p-N.C. SDS-PAGE of the magnetic bead assay. Lane 1: MW marker, lane 2: the presence of Als1p-N at the expected molecular height(approximately 60 kDa) after boiling of the beads indicates that the protein was bound to fibronectin-coupled beads.
2 D. S. Donohue, F. S. Ielasi, K. V. Y. Goossens and R. G. Willaert �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology
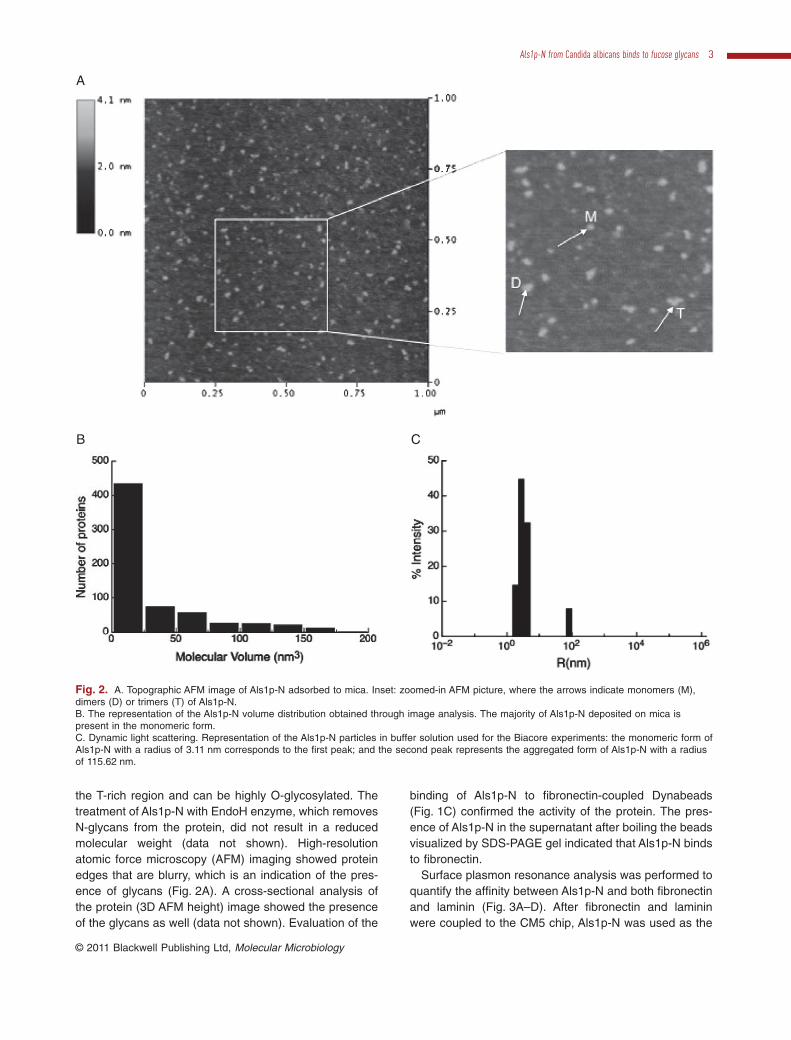
the T-rich region and can be highly O-glycosylated. Thetreatment of Als1p-N with EndoH enzyme, which removesN-glycans from the protein, did not result in a reducedmolecular weight (data not shown). High-resolutionatomic force microscopy (AFM) imaging showed proteinedges that are blurry, which is an indication of the pres-ence of glycans (Fig. 2A). A cross-sectional analysis ofthe protein (3D AFM height) image showed the presenceof the glycans as well (data not shown). Evaluation of the
binding of Als1p-N to fibronectin-coupled Dynabeads(Fig. 1C) confirmed the activity of the protein. The pres-ence of Als1p-N in the supernatant after boiling the beadsvisualized by SDS-PAGE gel indicated that Als1p-N bindsto fibronectin.
Surface plasmon resonance analysis was performed toquantify the affinity between Als1p-N and both fibronectinand laminin (Fig. 3A–D). After fibronectin and lamininwere coupled to the CM5 chip, Als1p-N was used as the
A
B C
Fig. 2. A. Topographic AFM image of Als1p-N adsorbed to mica. Inset: zoomed-in AFM picture, where the arrows indicate monomers (M),dimers (D) or trimers (T) of Als1p-N.B. The representation of the Als1p-N volume distribution obtained through image analysis. The majority of Als1p-N deposited on mica ispresent in the monomeric form.C. Dynamic light scattering. Representation of the Als1p-N particles in buffer solution used for the Biacore experiments: the monomeric form ofAls1p-N with a radius of 3.11 nm corresponds to the first peak; and the second peak represents the aggregated form of Als1p-N with a radiusof 115.62 nm.
Als1p-N from Candida albicans binds to fucose glycans 3
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

analyte to flow over the chip. The dissociation constant atequilibrium state (KD) was estimated at 1.6 � 0.6 ¥ 10-6 Mfor the Als1p-N–fibronectin interaction and 1.3 � 0.2 ¥10-5 M for the Als1p-N–laminin interaction using the 1:1binding model.
Als1p-N recognizes specificallyfucose-containing glycans
The purified Als1p-N was subjected to a high-throughputscreening allowing the binding to a total of 406 differentsynthetic glycan structures covalently attached to amicroarray glass slide through molecular spacers of differ-ent lengths. The Als1p-N bound selectively to multiplegroups of ligands with terminal fucose as well as to fucosealone (Fig. 4A). Most glycans that gave the highest fluo-rescence signals contained the structure Fuca1-2Galb1-GlcNAc (Fig. 4B). Additionally, Als1p-N selectively bound
to fucose linked to N-acetylglucosamine (GlcNAc) via a1–3and a1–4 linkages. The specificity for a particular stere-ochemistry was high, as no significant binding to othermonosaccharides, such as mannose, galactose, rham-nose, glucose and N-acetylglucosamine was observed.
To find out if the interaction between Als1p-N and theECM proteins laminin and fibronectin is based on a spe-cific interaction with fucose, SPR inhibition experimentsby adding fucose to the Als1p-N solution were performed;glucose and galactose were chosen as negative controls(based on the glycan array results). The results (Table S1)show that fucose exerts a significant – but modest –inhibitory effect on the binding of Als1p-N to both lamininand fibronectin. Glucose and galactose do not have asignificant inhibitory effect on the binding of Als1p-N tolaminin. Unexpectedly glucose and galactose alsoreduced binding of Als1p-N to fibronectin by almost half,relative to binding without added monosaccharide. These
A B
C D
Fig. 3. A. SPR sensorgram of the interaction between fibronectin and Als1p-N (concentration range of 0.02–100 mM).B. Fitting of the data points at equilibrium for the Als1p-N–fibronectin interaction to the steady-state 1:1 model.C. SPR sensorgram of the interaction between laminin and Als1p-N (concentration range of 0.12–62.5 mM).D. Fitting of the data points at equilibrium for the Als1p-N–laminin interaction to the steady-state 1:1 model.
4 D. S. Donohue, F. S. Ielasi, K. V. Y. Goossens and R. G. Willaert �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

results suggest strongly that inhibition of the fucose–lectininteraction is insufficient to disrupt Als1p-N binding tolaminin or fibronectin.
Als1p-N binds to BSA-fucose with millimolar affinity andto Als1p-N with micromolar affinity
The binding of the N-terminal part of Als1p to fucose wasfurther studied by injecting increasing concentrations ofAls1p-N in a BIAcore flow cell over the CM5 chip coatedwith BSA-fucose neoglycoconjugate. A wide concentration
range of Als1p-N (11–355 mM) was used to maximize thesignal-to-noise ratio and to facilitate the evaluation of theresults. Figure 5A shows a typical sensorgram obtained forthe interaction of Als1p-N with BSA-fucose. The equilib-rium dissociation constant was obtained by fitting the datato the 1:1 binding model and the estimated affinity was inmillimolar range with a KD value of 2.1 � 0.3 ¥ 10-4 M(Fig. 5B). Similarly, the N-terminal part of the Als1 proteinwas immobilized on a CM5 chip and increasing concentra-tions of Als1p-N (0.82–210 mM) were injected in the flowcell (Fig. 5C). Fitting of the data using the 1:1 binding
Fig. 4. Interaction of Als1p-N with glycanson the glycan microarray chip.A. Als1p-N glycan array spectrum.Fucose-containing glycans (No 10–11, 55–86,262–279) bind specifically to the N-terminalpart of Als1 protein.B. Consolidated glycan array data. Mostfucose-containing glycans recognized byAls1p-N contained theFuca1-2Galb1-4GlcNAc structure (first groupof glycans), which is a constituent of type-2 Hantigen. The second group of glycans(Fuca1-3GlcNAcb) represents structurespresent in Lewisx and Lewisy and the thirdgroup (Fuca1-4GlcNAcb) contains glycanspresent in Lewisa and Lewisb antigens. Thebinding response of Als1p-N to fucose aloneis shown in the forth group. Glycans with afluorescence response lower than 40 RFUwere not included. Error bars are based onthe standard error of the mean for sixreplicates.
A
B
Als1p-N from Candida albicans binds to fucose glycans 5
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

model, resulted in an estimated affinity KD for Als1p-N –Als1p-N binding of 2 � 0.1 ¥ 10-5 M (Fig. 5D).
Als1p-N in solution exists predominantlyin monomeric form
To find out, if the Als1p-N can form aggregates in bufferused for SPR experiments, the Als1p-N protein (667protein particles/mm2) was adsorbed to mica and visualizedusing tapping mode AFM (Fig. 2A). Figure 2A shows thatmost proteins are monomeric, but also small aggregatescan be observed (some aggregates are pointed witharrows on the inset picture).Arough estimate of the volumedistribution was performed using the method of Ratcliff andErie (2001) (Fig. 2B). The majority of the particles are in amonomeric form (smaller than 25 nm3). The analysis ofAls1p-N by dynamic light scattering confirmed the pres-ence of a very large protein fraction in monomeric form(Fig. 2C). One population corresponded to the monomeric
state of Als1p-N with a radius of 3.11 nm and was respon-sible for 93% of the change in light intensity. The secondpopulation of Als1p-N was in the aggregated form with aradius of 115.62 nm and was responsible for only 7% of thechange in light intensity. Additionally, during gel filtration ofthe Als1p-N into HBS buffer there was no indication of theformation of aggregates, as protein came out of the columnas one peak at 280 nm corresponding to the molecularweight protein of approximately 60 kDa. These resultssuggest that the predominant form of Als1p-N in bufferused for Biacore analysis was in the monomeric, non-aggregated form and justifies the one-to-one bindingmodel used to obtain the affinity constants (KD).
Discussion
In this study, we have used the Als1 protein encompass-ing the N-terminal, substrate-specific domain (Ig region ofabout 310 aa) together with the threonine-rich region
A B
C D
Fig. 5. A. SPR sensorgram of the BSA-fucose and Als1p-N (concentration range of 11–355 mM) interaction.B. Fitting of the data points at equilibrium for the Als1p-N–BSA-fucose interaction to the steady-state 1:1 model.C. SPR sensorgram of the Alsp1-N and Als1p-N (concentration range of 0.82–210 mM) interaction.D. Fitting of the data points at equilibrium for the Als1p-N–Als1p-N interaction to the steady-state 1:1 model.
6 D. S. Donohue, F. S. Ielasi, K. V. Y. Goossens and R. G. Willaert �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

(T region of about 105 aa), which has been shown previ-ously to be necessary for proper protein folding andsecretion to the medium (Rauceo et al., 2006). Thebinding activity resides in the N-terminal part of Alsprotein, although tandem repeats have been shown toenhance the binding of the Als1p N-terminal part and alterthe functional characteristics of Als proteins (Loza et al.,2004; Sheppard et al., 2004; Oh et al., 2005; Rauceoet al., 2006). Therefore, we aimed to measure the bindingaffinity that is related to the Ig-T region (18–432 aa).
The interactions between Als1p-N and fibronectin (440–500 kDa) and laminin (900 kDa), which are found in theextracellular matrix surrounding endothelial and epithelialcells (Ruoslahti, 1984; Beck et al., 1990), were quantifiedusing SPR. It was demonstrated that Als1p-N binds tofibronectin with an apparent KD of 1.6 � 0.6 ¥ 10-6 M andto laminin with a KD of 1.3 � 0.2 ¥ 10-5 M (Fig. 3A–D). Thefull-length Als1p was previously shown to be able to bindto fibronectin and laminin (Sheppard et al., 2004; Zhaoet al., 2004). Additionally, the truncated forms of the Als5protein Als5p1-431 and Als5p1-664 were shown, throughsingle antibody ELISA, to bind to fibronectin with anapparent dissociation constant in the nanomolar range(Rauceo et al., 2006; Frank et al., 2010). About 10-foldmore Als5p1-664 bound to fibronectin in comparison withAls5p1-431. This indicates that tandem repeats did not alterthe affinity but affected maximal binding as a result ofself-aggregation.
Using SPR, we have observed binding betweenAls1p-N proteins and estimated the affinity of this interac-tion (KD of 2.0 � 0.1 ¥ 10-5 M) (Fig. 5C and D). AFM visu-alization of Als1p-N showed some self-interaction ofAls1p-N (Fig. 2A and B). Nevertheless, the image analy-sis of the protein volume shows that most proteins arepresent in the non-aggregated form. To obtain KD values,we have used the one-to-one model, which assumesbinding of one molecule of analyte to one molecule ofligand. Taking into account the capability of Als1p-N toform aggregates, this model can only be an estimation ofthe calculated affinity.
In order to gain more information about possible molecu-lar ligands forAls proteins, we have performed glycan arrayscreening, for which Als1p-N was used. The results showthat Als1p-N can recognize fucose-containing glycans(Fig. 4A). The fluorescent signal from the fluorescentlylabelled antibodies obtained in the experiment was ratherlow and suggests a low affinity of Als1p-N for fucose. Thisis typical for lectin–carbohydrate interaction, as in vivothose interactions seem to be highly dependent on prop-erties such as multivalency, which results in an increasedavidity (Monsigny et al., 2000; Loris, 2009).
Fucosyl-specific lectins are found in plants, includingthe Lotus tetragonolobus (Pereira and Kabat, 1974) andUlex europaeus (Matsumoto and Osawa, 1969) lectins,
eel (Anguilla) (Watkins and Morgan, 1952), and in thefungus Rhizopus stalonifer (Oda et al., 2003). Fucose-binding lectins have recently been designated as F-typelectins (or F-lections) and they constitute a newly identi-fied family of fucosyl-binding lectins (Bianchet et al., 2002;Odom and Vasta, 2006), with members also described inthe Japanese horseshoe crab (Saito et al., 1997) and‘fucolectins’ in fish (Honda et al., 2000; Cammarata et al.,2001; Cammarata et al., 2007).
Fucose is present in the L-configuration of many cellsurface-expressed glycan structures found on mammaliancells (Becker and Lowe, 2003; Orczyk-Pawiłowicz, 2007).This carbohydrate is present as a terminal modification ofblood group ABH and Lewis antigens. Such localizationpredisposes fucose to play an important role in host–microbe interactions (Moran, 2008). Blood group antigenshave been pointed before to act as epithelial cell receptorsfor C. albicans (Cameron and Douglas, 1996). It wasreported that the minimum structure required to inhibit C.albicans attachment to buccal cells was the Fuca1-2Galbdeterminant of the H blood group antigen (Brassart et al.,1991). In another study, fucose was also the major inhibitorof C. albicans adhesion to buccal and vaginal epithelialcells (Critchley and Douglas, 1987b). The extracellularpolymeric material from C. albicans was used to isolatea lectin-like mannoprotein that recognized fucose-containing receptors; however, the protein was not identi-fied (Tosh and Douglas, 1992). The analysis of Als1p-Nbinding to glycans coupled to the microarray chip revealedthat Als1p-N bound preferentially to Fuca1-2Galb1-4GlcNAc structures, also when sulphated, followedby fucose linked via a1–3 and a1–4 linkages toN-acetylglucosamine (GlcNAc) (Fig. 4B). Additionally,a-linked fucose coupled via eight- or nine-atoms spacer tothe glycan array chip also gave a significant response.Fuca1-2Galb1-4GlcNAc is a characteristic of the bloodgroup H type-2 determinant, which can be found on redcells, in vascular endothelium, in epithelium of gastrointes-tinal and respiratory tract as well as in the epithelium of theouter layer of the skin (epidermis) (Ravn and Dabelsteen,2000; Varki et al., 2009). By comparison, the response tothe Fuc a1-2Galb1–3 structure, which is a constituent ofthe H antigen type 1, 3 and 4, was much lower. Fuca1–3and a1–4 linked to GlcNAc are typical for Lewis bloodgroup antigens (Varki et al., 2009). The Als1p-N bound tostructures represented by all Lewis antigens, which arepresent in human secretions and fluids, and can beadsorbed to a variety of tissues (Bässler, 1986; Ravn andDabelsteen, 2000). Blood group antigens are commonlyused by pathogens as binding receptors. For example,some Campylobacter jejuni strains and rabbit-specificvirus (RHDV) were shown to recognize and attach to Htype 2 epitopes (Newburg, 2000; Ruvoën-Clouet et al.,2000). In another example, the bacterium Helicobacter
Als1p-N from Candida albicans binds to fucose glycans 7
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

pylori is able to attach to Lewisb antigens displayed onhuman gastric epithelial cells (Moran, 2008). Differentcarbohydrates are also recognized by microorganisms astools to attach to host cells. For example, Epa1 protein fromCandida glabrata has been shown to bind to host-cellasialo-lactosyl-containing carbohydrates, which can befound on epithelial cells (Cormack et al., 1999). Addition-ally, the glycan specifity of Epa proteins from C. glabratausing glycan array and adhesion assays has beendescribed (Zupancic et al., 2008). It was shown that theN-terminal parts of Epa1, Epa6 and Epa7 bound to glycanscontaining a terminal galactose. This binding was inhibitedwhen more complex glycans (e.g. with terminal fucose)were used and blocked the access of Epa proteins togalactose residue. The range of specificity towardsgalactose-containing glycans was different between allthree Epa proteins.
The affinity of the Als1p-N for fucose was determinedquantitatively by SPR (KD of 2.1 � 0.3 ¥ 10-4 M) (Fig. 5Aand B). An affinity in the millimolar range is typicallyobserved for lectin–monosaccharides interactions (Loris,2009). However, it has been shown that some fucosebinding lectins can have an affinity in the micromolarrange (Wimmerova et al., 2003; Kostlánová et al., 2005;Matsumura et al., 2007; Olausson et al., 2008; Okuyamaet al., 2009).
Some fungal lectins, such as Epa proteins from C.glabrata and flocculins from S. cerevisiae are calcium-dependent for carbohydrate binding (Miki et al., 1982;Cormack et al., 1999). Although, it has been recentlyshown that the N-terminal part of Flo5p contains two
binding sites, where only the high affinity site contained aCa2+ ion (Veelders et al., 2010). Therefore, we have alsotested the influence of calcium on binding (using SPR)between Als1p-N and BSA-fucose, Als1p-N and fibronec-tin and laminin as well as on self-binding of Als1p-N andfound no enhancement of binding (data not shown).Another fungal lectin, the Rhizopus stolonifer lectin, doesalso not require Ca2+ ions for binding (Oda et al., 2003).
Als proteins use different strategies to bind to mamma-lian cells (Fig. 6). In this study, we showed that theN-terminal part of Als1p can bind to fucose-containingglycans, interact with ECM proteins, and self-interact.Because the ECM proteins laminin and fibronectin areglycosylated with fucose containing glycans (Mosessonand Amrani, 1980; Jin et al., 1995), the binding of Als1p-Nto these proteins might in theory depend on interactionwith these glycans. However, SPR inhibition experimentswith fucose, glucose and galactose showed that fucoseinhibited binding of Als1 to laminin only marginally. More-over, a slight inhibition of Als1 binding to fibronectin byfucose was apparently non-specific as glucose or galac-tose inhibited equally well. We interpret this to mean thatbinding of Als1 to laminin or fibronectin is not mediatedentirely by a lectin–glycan interaction, and that additionalcontacts between Als1 and these ECM components mustunderlie the interaction. While our data show that thefucose–lectin interaction is not necessary for bindingthese ECM components, we have not ruled out that itmight contribute under physiological conditions to theAls1p interaction with glycosylated proteins, includinglaminin and fibronectin. Als1p-N has also been indicated
Fig. 6. Hypothetical interactions mediated by the Als1 protein. The N-terminal part of Als1p (indicated in light blue) binds to the extracellularmatrix proteins (laminin and fibronectin), N- and E-cadherins as well as to blood group antigens found on epithelial and endothelial cells.Additionally, Als1p-N binds to Als1p-N expressed on another C. albicans cell. Tandem repeats of Als1p (shown as a zigzag pattern) can formamyloid-like structures by aggregating with tandem repeats of the neighbouring Als1p. Binding of Als1p to gelatin, which is the product of thepartial hydrolysis of collagen has not been indicated (Sheppard et al., 2004).
8 D. S. Donohue, F. S. Ielasi, K. V. Y. Goossens and R. G. Willaert �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

to induce endocytosis of C. albicans by binding toN-cadherin on endothelial cells and E-cadherin on oralepithelial cells (Phan et al., 2007).
The used construct in this study contains the substrate-specific binding domain and the T region. The substrate-specific binding domain contains Ig-like domains thatbelong to the immunoglobulin superfamily (Sheppardet al., 2004). Because the protein was expressed in S.cerevisiae, the T region contains two serine residues(position 303 and 379) instead of two leucine residues asa result of the differently translated CUG codon in C.albicans (Santos and Tuite, 1995; Gomes et al., 2007).However, because the mistranslated codons are notpresent in the N-terminal binding domain, they will haveno significant impact on the investigated substrate-specific binding function. The expression of Als1p-Nin S. cerevisiae results also in a slightly differentO-glycosylation glycan structure (the two end-standingmannose residues are connected via a1–3 in S. cerevi-siae instead of a1–2 in C. albicans) (Varki et al., 2009).Because these expression differences result in minorstructural changes, they will not have a significant effecton the binding characterization results.
Aggregation of Als proteins have also been attributed toamyloid-forming sequences found in the threonine-richand tandem repeats region (Rauceo et al., 2006; Otooet al., 2008; Ramsook et al., 2010). The T region is themost conserved sequence in Als proteins. T regiondomains can bind strongly (unbinding force is larger than250 pN) to other T region domains as T–T interaction wasnecessary to unfold the TR regions of Als5p in an AFMforce spectroscopy experiment (Alsteens et al., 2009).The observed Als1p-N–Als1p-N interaction probablyoccurs through interacting T-domains.
Variation within tandem repeats may alter the functionalcharacteristics of Als proteins (Hoyer et al., 2008; Natherand Munro, 2008). An expanded tandem repeat regionenhances the exposure of the N-terminal functionaldomain in the extracellular space and can explain theenhanced binding. Each tandem repeat sequence foldscompactly to give an independent b-sheet-rich domainwith conserved hydrophobic core and consistent surfacefeatures (Frank et al., 2010). The domain surface pro-motes interactions with a large variety of hydrophobicsurfaces, including other tandem repeat domains.
The expression of the full-length Als1p and Als5p on thesurface of C. albicans and S. cerevisiae demonstratedadherence to immobilized peptides and proteins (Klotzet al., 2004). Adherence was sequence specific becauseall recognized peptides contain the structural motif tj+. Toinvestigate the binding of the full-length Als1p to fucose,we performed a binding assay based on a microfluidicflow cell (Bioflux, Fuxion) with a C. albicans Als1p over-expressing and an Als1p null mutant (Fu et al., 2002)
(data not shown). Binding of the strains to a BSA-fucose-and BSA-coated channel was compared using anincreasing shear stress (up to 10 dyne/cm2) to removeloosely bound cells. Als1p overexpressing cells as well asAls1p null mutant cells could bind to the coated flow cellwall. However, there was no significant differencebetween binding to BSA-fucose and BSA. This result indi-cates that binding to fucose is weak compared with otherbinding events between the cell wall and the coatedchannel wall. The weak binding to fucose-containingglycans and BSA-fucose of the purified Als1p-N was inthis study also experimentally observed using glycanarray screening and SPR respectively.
To sum up, we have shown that the N-terminal bindingdomain of Als1p binds to ECM proteins (fibronectin andlaminin) with micromolar affinity calculated for one-to-one interaction of ligand and analyte. The Als1p-N alsorecognizes fucose-containing glycans, of which struc-tures are typically found in blood group antigens. Thepredominant structure recognized by Als1p-N was theblood group antigen H type 2 and also Lewis antigens.The interaction between Als1p-N and fucose was deter-mined to be in the millimolar range. We have confirmedthat Als1p-N can bind to another Als1p-N and we havecharacterized this interaction quantitatively. Despite thecapability of self-binding of Als1p-N, the AFM and DLSanalysis of Als1p-N sample showed that most proteinparticles are present in non-aggregated form. The mul-tiple interactions mediated by the N-terminal part ofAls1p-N from C. albicans are likely to enhance thechance of efficient colonization of mammalian cells bythis fungus. Although predictions of the Als proteinsstructures were made (Sheppard et al., 2004; Phanet al., 2007), resolving the crystal structure wouldanswer a number of questions regarding the exact posi-tion of binding site(s) for fucose and ECM proteins.
Experimental procedures
Cloning and purification of Als1p-N (17–432 aa)
The N-terminal part of the ALS1 gene was inserted into apYEX-S1 vector using the In-Fusion method (Clontech,Mountain View, CA, USA), which is based on the recombina-tion between homologous sequences of the vector and theamplified gene (Zhu et al., 2007). For this purpose two oligo-nucleotides 5′-TCCTTAGTCAAAAGGAAGACAATCACTGGTGTTTTTGATAGTTTTAATTCAT-3′ and 5′-GGAGATCGGAATTCGTCAGTGATGGTGATGGTGATGTGGAACTTGTACCACCACTGTGT-3′ were designed to amplify the N-terminal partof ALS1 gene with a C-terminal His-tag. The plasmidpYEX-S1 is a yeast-Escherichia coli shuttle vector, regulatedby a strong constitutive PGK promoter with mutated leu-2,URA3 and AmpR as selectable markers (Castelli et al., 1994).The secretion sequence for Als1p (1–17 aa) has not beenincluded during cloning, as the pYEX-S1 vector contains the
Als1p-N from Candida albicans binds to fucose glycans 9
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

full-length leader sequence from Kluyveromyces lactis todirect proteins through the secretory pathway. The S288C-derived BY4147 S. cerevisiae yeast strain has been used totransform plasmids carrying the ALS1 gene. Cells were cul-tivated in rich YPD medium at 30°C with shaking at 150 r.p.m.After 2 days of cultivation, the supernatants were collected bycentrifuging at 4°C, 4000 r.p.m., for 30 min and subsequentlyfiltered to remove residual cells. The pH of the final superna-tant was adjusted to 7.0–7.5 with 1 M Tris base, incubatedwith nickel-nitrilotriacetic acid (Ni-NTA) Sepharose beads for1 h at 4°C, and next applied onto a column containing moreNi-NTA Sepharose beads. To minimize unspecific binding,the beads with bound protein were washed with buffer con-taining 500 mM NaCl, 10 mM imidazol and 20 mM Tris pH7.5. The proteins were eluted with 1 M imidazol in phosphatebuffer saline (PBS), pH 7.4. The eluted protein was collectedand the purity was visualized using sodium dodecyl sulphatepolyacrylamide gel electrophoresis (SDS-PAGE). Fractionshave been pooled together and concentrated using 30 kDamolecular weight cut off (MWCO) membrane concentrators(Amicon Ultra, Millipore). To further purify the proteins andexchange the buffer for future experiments, gel filtration wasapplied. For this purpose, the Superdex 75 HR 10/30 columnwas used with a buffer containing 150 mM NaCl, 20 mM TrispH 7.5 buffer. One millilitre fractions were collected and visu-alized on SDS-PAGE gel. Pure fractions were pooledtogether, concentrated and used immediately for the nextexperiments. The maximum protein yield obtained duringpurification was about 7 mg per litre of culture.
Detection of glycoproteins
To visualize the glycans present on the proteins, the Pro-QEmerald 300 Glycoprotein Gel and Blot Stain Kit (Invitrogen)was used according to manufacturer’s instructions.
Magnetic bead activity assay
To test the binding capacity of the purified Als1p-N, a beadadhesion assay was performed as described previously(Gaur et al., 1999). Tosyl-activated M-280 Dynabeads (Dynal,Invitrogen) were coated with fibronectin (BD Biosciences)following the manufacturer’s instructions. Als1p-N (20 mg permg of Dynabeads) was added to the beads at a concentrationof 3.3 ¥ 108 beads ml-1 and incubated for 2 h at 37°C withvigorous shaking. To remove unbound proteins, the beadswere washed with PBS. To release the bound proteins, thebeads were boiled and the supernatant was subjected toSDS-PAGE to visualize the released proteins.
Glycan array
The N-terminal part of Als1p was screened for binding toglycans printed on a glass slide microarray (version 3.2)developed by the Consortium for Functional Glycomics (Blixtet al., 2004). The immunodetection was performed using fluo-rescently labelled anti-His antibodies. The average relativefluorescent units were obtained for six replicates for eachglycan. Error bars in Fig. 4 are based on the standard error ofthe mean for these replicates.
SPR
The binding measurements of Als1p-N were performed usingSPR, BIAcore 3000 system (GE Healthcare). Fibronectin (BDBiosciences), laminin (Sigma-Aldrich), BSA-fucose (DextraLaboratories) and Als1p-N were covalently immobilized on aCM5 sensor chip via amine coupling using an amine couplingkit (BIAcore, GE Healthcare). Increasing concentrations ofthe analyte (Als1p-N) were allowed to flow across both thereference and ligand-coated flow cell in running buffer con-taining 3 mM EDTA, 150 mM NaCl, 0.05% (v/v) Tween 20,20 mM HEPES pH 7.4 (HBS) or in buffer used for glycanarray screening (2 mM CaCl2, 150 mM NaCl, 20 mM Tris pH7.4). The injection was performed at 25°C using a flow rate of20 ml min-1 for 1.5 min (10 min for Als1p-N over fibronectin).The dissociation was then monitored for 9 min. For the Als1p-N–fibronectin interaction, the surface was regenerated with0.1% (w/v) SDS. No surface regeneration was needed for theinteraction between Als1p-N and laminin, fucose andAls1p-N. Binding was determined by measuring the increasein resonance units after subtraction of the backgroundresponse obtained from the reference flow cell and thesample containing only the buffer. The dissociation constantat equilibrium state (KD) was estimated using followingsteady-state affinity model with a 1:1 ligand-analyte ratio:Req = Rmax (KACA)/(KACA + 1) where Req is the response atequilibrium state, KA is the association constant at equilibriumstate, CA is the analyte concentration and Rmax is themaximum binding capacity (Chaiken et al., 1992). The stan-dard deviation was calculated from the average of thedissociation constants (KD) obtained for two independentexperiments.
For the binding inhibition experiments, Als1p-N–fibronectin and Als1p-N–laminin interactions were analysed– as described before in this section – for 5 mM Als1p-Nsamples containing 100 mM of the monosaccharideL-fucose (Sigma-Aldrich), as potential inhibitor molecule,and 100 mM of the monosaccharides D-glucose orD-galactose (Sigma-Aldrich), as negative controls. For eachexperiment, Req values were retrieved from Als1p-N-onlyand Als1p-N-monosaccharide curves. Averages and stan-dard deviations, based on three independent repetitions,were calculated for these values. Subsequently, in order toevaluate the relative carbohydrate effects on the equilibriumresponse, every mean Req was divided by the one obtainedfor Als1p-N-only condition. The statistical significance wascalculated with the Student’s t-test. All the results wereanalysed with BIAevaluation software version 4.1 (GEHealthcare).
AFM
Topographic images were recorded with a Multimode Nano-scope IIIa AFM instrument (Veeco, Santa Barbara, CA, USA)using tapping mode in air. RTESP cantilevers (Veeco)were used with a nominal spring constant of 20–80 N m-1.The AFM sample was prepared by depositing 10 ml of0.05 mg ml-1 of Als1p-N for 15 min at room temperature ontofreshly cleaved mica. Unbound proteins were removed bywashing with ultrapure water and dried under a gentle streamof nitrogen gas. The image was obtained at a scan rate of
10 D. S. Donohue, F. S. Ielasi, K. V. Y. Goossens and R. G. Willaert �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

1 Hz. Images were processed by first order flattening toremove the background slope. The calculation of the proteinvolume distribution was performed as described previously(Ratcliff and Erie, 2001).
Dynamic light scattering
The Als1p-N sample (0.35 mg ml-1) was prepared in thebuffer that was used for the SPR measurements (HBS). Thelight intensity was measured using the DynaPro DLS PlateReader (Wyatt Technology). The mean radius was calculatedbased on 10 acquisitions for a globular protein shape.
Acknowledgements
We wish to acknowledge the Consortium for Functional Gly-comics (http://www.functionalglycomics.org) for the glycananalysis. We thank Prof J.E. Edwards (Medicine Dept., Divi-sion of Infectious Diseases, Harbor-UCLA, CA, USA) for pro-viding us with the sequence encoding the N-terminal part ofAls1p on pYEX-S1 vector. We acknowledge Prof A.P. Mitchell(Dept. Biological Sciences, Carnegie Mellon University, Pitts-burgh, PA, USA) for the gift of the C. albicans Als1 overex-pression strain and the Als1 null mutant strain. We alsoacknowledge Catherine Stassen and Prof Bart Devreese(L-Probe, Ghent University, Ghent, Belgium) for mass spec-trometry analysis. The Belgian Federal Science Policy Officeand European Space Agency (ESA) PRODEX program, theInstitute for the Promotion of Innovation by Science and Tech-nology in Flanders (IWT) and the Research Council of theVUB, all supported this work.
References
Alsteens, D., Dupres, V., Klotz, S.A., Gaur, N.K., Lipke, P.N.,and Dufrêne, Y.F. (2009) Unfolding individual Als5p adhe-sion proteins on live cells. ACS Nano 3: 1677–1682.
Barclay, A.N. (2003) Membrane proteins withimmunoglobulin-like domains – a master superfamily ofinteraction molecules. Semin Immunol 15: 215–223.
Bässler, G. (1986) Determination of the Lewis blood groupsubstances in stains of forensically relevant body fluids.Forensic Sci Int 30: 29–35.
Beck, K., Hunter, I., and Engel, J. (1990) Structure and func-tion of laminin: anatomy of a multidomain glycoprotein.FASEB J 4: 148–160.
Becker, D.J., and Lowe, J.B. (2003) Fucose: biosynthesisand biological function in mammals. Glycobiology 13:41R–53R.
Bianchet, M.A., Odom, E.W., Vasta, G.R., and Amzel, L.M.(2002) A novel fucose recognition fold involved in innateimmunity. Nat Struct Biol 9: 628–634.
Blixt, O., Head, S., Mondala, T., Scanlan, C., Huflejt, M.E.,Alvarez, R., et al. (2004) Printed covalent glycan array forligand profiling of diverse glycan binding proteins. Proc NatlAcad Sci USA 101: 17033–17038.
Brassart, D., Woltz, A., Golliard, M., and Neeser, J.R. (1991)In vitro inhibition of adhesion of Candida albicans clinicalisolates to human buccal epithelial cells by Fuc alpha1-2Gal beta-bearing complex carbohydrates. Infect Immun59: 1605–1613.
Cameron, B.J., and Douglas, L.J. (1996) Blood group gly-colipids as epithelial cell receptors for Candida albicans.Infect Immun 64: 891–896.
Cammarata, M., Vazzana, M., Chinnici, C., and Parrinello, N.(2001) A serum fucolectin isolated and characterized fromsea bass Dicentrarchus labrax. Biochim Biophys Acta1528: 196–202.
Cammarata, M., Benenati, G., Odom, E.W., Salerno, G.,Vizzini, A., Vasta, G.R., and Parrinello, N. (2007) Isolationand characterization of a fish F-type lectin from gilt headbream (Sparus aurata) serum. Biochim Biophys Acta 1770:150–155.
Castelli, L.A., Mardon, C.J., Strike, P.M., Azad, A.A., andMacreadie, I.G. (1994) High-level secretion of correctlyprocessed beta-lactamase from Saccharomyces cerevi-siae using a high-copy-number secretion vector. Gene142: 113–117.
Chaffin, W.L., López-Ribot, J.L., Casanova, M., Gozalbo, D.,and Martínez, J.P. (1998) Cell wall and secreted proteins ofCandida albicans: identification, function, and expression.Microbiol Mol Biol Rev 62: 130–180.
Chaiken, I., Rosé, S., and Karlsson, R. (1992) Analysis ofmacromolecular interactions using immobilized ligands.Anal Biochem 201: 197–210.
Cheng, G., Wozniak, K., Wallig, M.A., Fidel, P.L., Jr, Trupin,S.R., and Hoyer, L.L. (2005) Comparison betweenCandida albicans agglutinin-like sequence gene expres-sion patterns in human clinical specimens and models ofvaginal candidiasis. Infect Immun 73: 1656–1663.
Cormack, B.P., Ghori, N., and Falkow, S. (1999) An adhesinof the yeast pathogen Candida glabrata mediating adher-ence to human epithelial cells. Science 285: 578–582.
Critchley, I.A., and Douglas, L.J. (1987a) Role of glycosidesas epithelial cell receptors for Candida albicans. J GenMicrobiol 133: 637–643.
Critchley, I.A., and Douglas, L.J. (1987b) Isolation and partialcharacterization of an adhesin from Candida albicans.J Gen Microbiol 133: 629–636.
Filler, S.G. (2006) Candida–host cell receptor–ligandinteractions. Curr Opin Microbiol 9: 333–339.
Frank, A.T., Ramsook, C.B., Otoo, H.N., Tan, C., Soybelman,G., Rauceo, J.M., et al. (2010) Structure and Function ofGlycosylated Tandem Repeats from Candida albicans ALSAdhesins. Eukaryot Cell 9: 405–414.
Fu, Y., Rieg, G., Fonzi, W.A., Belanger, P.H., Edwards, J.E.,Jr, and Filler, S.G. (1998) Expression of the Candida albi-cans gene ALS1 in Saccharomyces cerevisiae inducesadherence to endothelial and epithelial cells. Infect Immun66: 1783–1786.
Fu, Y., Ibrahim, A.S., Sheppard, D.C., Chen, Y.C., French,S.W., Cutler, J.E., et al. (2002) Candida albicans Als1p: anadhesin that is a downstream effector of the EFG1 filamen-tation pathway. Mol Microbiol 44: 61–72.
Gaur, N.K., Klotz, S.A., and Henderson, R.L. (1999) Overex-pression of the Candida albicans ALA1 gene in Saccharo-myces cerevisiae results in aggregation followingattachment of yeast cells to extracellular matrix proteins,adherence properties similar to those of Candida albicans.Infect Immun 67: 6040–6047.
Ghannoum, M.A., Burns, G.R., Elteen, K.A., and Radwan,S.S. (1986) Experimental evidence for the role of lipids in
Als1p-N from Candida albicans binds to fucose glycans 11
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

adherence of Candida spp. to human buccal epithelialcells. Infect Immun 54: 189–193.
Gomes, A.C., Miranda, I., Silva, R.M., Moura, G.R., Thomas,B., Akoulitchev, A., and Santos, M.A. (2007) A geneticcode alteration generates a proteome of high diversity inthe human pathogen Candida albicans. Genome Biol 8:R206.
Green, C.B., Cheng, G., Chandra, J., Mukherjee, P., Ghan-noum, M.A., and Hoyer, L.L. (2004) RT-PCR detection ofCandida albicans ALS gene expression in the reconstitutedhuman epithelium (RHE) model of oral candidiasis and inmodel biofilms. Microbiology 150: 267–275.
Green, C.B., Marretta, S.M., Cheng, G., Faddoul, F.F.,Ehrhart, E.J., and Hoyer, L.L. (2006) RT-PCR analysis ofCandida albicans ALS gene expression in a hyposalivatoryrat model of oral candidiasis and in HIV-positive humanpatients. Med Mycol 44: 103–111.
Holmgren, A., Kuehn, M.J., Brändén, C.I., and Hultgren, S.J.(1992) Conserved immunoglobulin-like features in a familyof periplasmic pilus chaperones in bacteria. EMBO J 11:1617–1622.
Honda, S., Kashiwagi, M., Miyamoto, K., Takei, Y., andHirose, S. (2000) Multiplicity, structures, and endocrine andexocrine natures of eel fucose-binding lectins. J Biol Chem275: 33151–33157.
Hoyer, L.L. (2001) The ALS gene family of Candida albicans.Trends Microbiol 9: 176–180.
Hoyer, L.L., Green, C.B., Oh, S.H., and Zhao, X. (2008)Discovering the secrets of the Candida albicans agglutinin-like sequence (ALS) gene family-a sticky pursuit. MedMycol 46: 1–15.
Jimenez-Lucho, V., Ginsburg, V., and Krivan, H.C. (1990)Cryptococcus neoformans, Candida albicans, and otherfungi bind specifically to the glycosphingolipid lactosylce-ramide (Gal beta 1-4Glc beta 1-1Cer), a possible adhesionreceptor for yeasts. Infect Immun 58: 2085–2090.
Jin, F., Chammas, R., Engel, J., and Reinhold, V. (1995)Structure and function of laminin 1 glycans; glycanprofiling. Glycobiology 5: 157–158.
Kamai, Y., Kubota, M., Kamai, Y., Hosokawa, T., Fukoka, T.,and Filler, S.G. (2002) Contribution of Candida albicansALS1 to the pathogenesis of experimental oropharyngealcandidiasis. Infect Immun 70: 5256–5258.
Klotz, S.A., Gaur, N.K., Lake, D.F., Chan, V., Rauceo, J., andLipke, P.N. (2004) Degenerate peptide recognition byCandida albicans adhesins Als5p and Als1p. Infect Immun72: 2029–2034.
Kostlánová, N., Mitchell, E.P., Lortat-Jacob, H., Oscarson, S.,Lahmann, M., Gilboa-Garber, N., et al. (2005) The fucose-binding lectin from Ralstonia solanacearum. A new type ofbeta-propeller architecture formed by oligomerizationand interacting with fucoside, fucosyllactose, and plantxyloglucan. J Biol Chem 29: 27839–27849.
Loris, R. (2009) Structural features of lectins and their bindingsites. In Bioinformatics for Glycobiology and Glycomics: AnIntroduction. Von der Lieth, C.W., Lütteke, T., and Frank, M.(eds). Chichester: Wiley-Blackwell, pp. 415–431.
Loza, L., Fu, Y., Ibrahim, A.S., Sheppard, D.C., Filler, S.G.,and Edwards, J.E., Jr (2004) Functional analysis of theCandida albicans ALS1 gene product. Yeast 21: 473–482.
Macura, A.B., and Tondyra, E. (1989) Influence of some
carbohydrates and concanavalin A on the adherence ofCandida albicans in vitro to buccal epithelial cells. ZentralblBakteriol 272: 196–201.
Matsumoto, I., and Osawa, T. (1969) Purification and char-acterization of an anti-H(O) phytohemagglutinin of Ulexeuropeus. Biochim Biophys Acta 194: 180–189.
Matsumura, K., Higashida, K., Ishida, H., Hata, Y., Yama-moto, K., Shigeta, M., et al. (2007) Carbohydrate bindingspecificity of a fucose-specific lectin from Aspergillusoryzae: a novel probe for core fucose. J Biol Chem 282:15700–15708.
Mavor, A.L., Thewes, S., and Hube, B. (2005) Systemicfungal infections caused by Candida species: epidemiol-ogy, infection process and virulence attributes. Curr DrugTargets 6: 863–874.
Miki, B.L., Poon, N.H., James, A.P., and Seligy, V.L. (1982)Possible mechanism for flocculation interactions governedby gene FLO1 in Saccharomyces cerevisiae. J Bacteriol150: 878–889.
Monsigny, M., Mayer, R., and Roche, A.C. (2000) Sugar-lectin interactions: sugar clusters, lectin multivalency andavidity. Carbohydr Lett 4: 35–52.
Moran, A.P. (2008) Relevance of fucosylation and Lewisantigen expression in the bacterial gastroduodenal patho-gen Helicobacter pylori. Carbohydr Res 343: 1952–1965.
Mosesson, M.W., and Amrani, D.L. (1980) The structure andbiologic activities of plasma fibronectin. Blood 56: 145–158.
Nather, K., and Munro, C.A. (2008) Generating cell surfacediversity in Candida albicans and other fungal pathogens.FEMS Microbiol Lett 285: 137–145.
Newburg, D.S. (2000) Oligosaccharides in human milk andbacterial colonization. J Pediatr Gastroenterol Nutr 30:S8–S17.
Oda, Y., Senaha, T., Matsuno, Y., Nakajima, K., Naka, R.,Kinoshita, M., et al. (2003) A new fungal lectin recognizingalpha(1-6)-linked fucose in the N-glycan. J Biol Chem 278:32439–32447.
Odom, E.W., and Vasta, G.R. (2006) Characterization of abinary tandem domain F-type lectin from striped bass(Morone saxatilis). J Biol Chem 281: 1698–1713.
Oh, S.H., Cheng, G., Nuessen, J.A., Jajko, R., Yeater, K.M.,Zhao, X., et al. (2005) Functional specificity of Candidaalbicans Als3p proteins and clade specificity of ALS3alleles discriminated by the number of copies of thetandem repeat sequence in the central domain. Microbiol-ogy 151: 673–681.
Okuyama, S., Nakamura-Tsuruta, S., Tateno, H., Hiraba-yashi, J., Matsubara, K., and Hori, K. (2009) Strict bindingspecificity of small-sized lectins from the Red Alga Hypneajaponica for core (a1-6) fucosylated N-glycans. Biosci Bio-technol Biochem 73: 912–920.
Olausson, J., Tibell, L., Jonsson, B.H., and Påhlsson, P.(2008) Detection of a high affinity binding site in recombi-nant Aleuria aurantia lectin. Glycoconj J 25: 753–762.
Orczyk-Pawiłowicz, M. (2007) The role of fucosylation ofglycoconjugates in health and disease. Postepy Hig MedDosw 61: 240–252.
Otoo, H.N., Lee, K.G., Qiu, W., and Lipke, P.N. (2008)Candida albicans Als adhesins have conserved amyloid-forming sequences. Eukaryot Cell 7: 776–782.
12 D. S. Donohue, F. S. Ielasi, K. V. Y. Goossens and R. G. Willaert �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

Pereira, M.E., and Kabat, E.A. (1974) Specificity ofpurified hemagglutinin (lectin) from Lotus tetragonolobus.Biochemistry 13: 3184–3192.
Phan, Q.T., Myers, C.L., Fu, Y., Sheppard, D.C., Yeaman,M.R., Welch, W.H., et al. (2007) Als3 is a Candida albicansinvasin that binds to cadherins and induces endocytosis byhost cells. PLoS Biol 5: e64.
Ramsook, C.B., Tan, C., Garcia, M.C., Fung, R., Soybelman,G., Henry, R., et al. (2010) Yeast cell adhesion moleculeshave functional amyloid-forming sequences. Eukaryot Cell9: 393–404.
Ratcliff, G.C., and Erie, D.A. (2001) A novel single-moleculestudy to determine protein–protein association constants.J Am Chem Soc 123: 5632–5635.
Rauceo, J.M., De Armond, R., Otoo, H., Kahn, P.C., Klotz,S.A., Gaur, N.K., et al. (2006) Threonine-rich repeatsincrease fibronectin binding in the Candida albicansadhesin Als5p. Eukaryot Cell 5: 1664–1673.
Ravn, V., and Dabelsteen, E. (2000) Tissue distribution ofhisto-blood group antigens. APMIS 108: 1–28.
Ruoslahti, E. (1984) Fibronectin in cell adhesion andinvasion. Cancer Metastasis Rev 3: 43–51.
Rupp, S. (2007) Interactions of the fungal pathogen Candidaalbicans with the host. Future Microbiol 2: 141–151.
Ruvoën-Clouet, N., Ganiére, J.P., André-Fontaine, G., Blan-chard, D., and Le Pendu, J. (2000) Binding of rabbit hem-orrhagic disease virus to antigens of the ABH histo-bloodgroup family. J Virol 74: 11950–11954.
Saito, T., Hatada, M., Iwanaga, S., and Kawabata, S. (1997)A newly identified horseshoe crab lectin with binding speci-ficity to O-antigen of bacterial lipopolysaccharides. J BiolChem 272: 30703–30708.
Sandin, R.L., Rogers, A.L., Patterson, R.J., and Beneke, E.S.(1982) Evidence for mannose-mediated adherence ofCandida albicans to human buccal cells in vitro. InfectImmun 35: 79–85.
Santos, M.A., and Tuite, M.F. (1995) The CUG codon isdecoded in vivo as serine and not leucine in Candidaalbicans. Nucleic Acids Res 23: 1481–1486.
Sheppard, D.C., Yeaman, M.R., Welch, W.H., Phan, Q.T., Fu,Y., Ibrahim, A.S., et al. (2004) Functional and structuraldiversity in the Als protein family of Candida albicans. J BiolChem 279: 30480–30489.
Sundstrom, P. (2002) Adhesion in Candida spp. Cell Micro-biol 4: 461–469.
Tosh, F.D., and Douglas, L.J. (1992) Characterization of afucoside-binding adhesin of Candida albicans. InfectImmun 60: 4734–4739.
Vardar-Ünlü, G., MsSharry, C., and Douglas, J.L. (1998)Fucose-specific adhesins on germ tubes of Candidaalbicans. FEMS Immunol Med Microbiol 20: 55–67.
Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley,P., Bertozzi, C.R., et al. (2009) Essentials of Glycobiology,2nd edn. New York: Cold Spring Harbor Laboratory Press.
Veelders, M., Brückner, S., Ott, D., Unverzagt, C., Mösch,H.U., and Essen, L.O. (2010) Structural basis of flocculin-mediated social behavior in yeast. Proc Natl Acad Sci USA107: 22511–22516.
Watkins, W.M., and Morgan, W.T. (1952) Neutralization of theanti-H agglutinin in eel serum by simple sugars. Nature169: 825–826.
Wimmerova, M., Mitchell, E., Sanchez, J.F., Gautier, C., andImberty, A. (2003) Crystal structure of fungal lectin: six-bladed beta-propeller fold and novel fucose recognitionmode for Aleuria aurantia lectin. J Biol Chem 278: 27059–27067.
Yu, L., Lee, K.K., Sheth, H.B., Lane-Bell, P., Srivastava, G.,Hindsgaul, O., et al. (1994) Fimbria-mediated adherence ofCandida albicans to glycosphingolipid receptors on humanbuccal epithelial cells. Infect Immun 62: 2843–2848.
Zhao, X., Oh, S.H., Cheng, G., Green, C.B., Nuessen, J.A.,Yeater, K., et al. (2004) ALS3 and ALS8 represent a singlelocus that encodes a Candida albicans adhesin; functionalcomparisons between Als3p and Als1p. Microbiology 150:2415–2428.
Zhu, B., Cai, G., Hall, E.O., and Freeman, G.J. (2007)In-fusion assembly: seamless engineering of multidomainfusion proteins, modular vectors, and mutations. Biotech-niques 43: 354–359.
Zhu, W., and Filler, S.G. (2010) Interactions of Candida albi-cans with epithelial cells. Cell Microbiol 12: 273–282.
Zupancic, M.L., Frieman, M., Smith, D., Alvarez, R.A., Cum-mings, R.D., and Cormack, B.P. (2008) Glycan microarrayanalysis of Candida glabrata adhesin ligand specificity. MolMicrobiol 68: 547–549.
Supporting information
Additional supporting information may be found in the onlineversion of this article.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials suppliedby the authors. Any queries (other than missing material)should be directed to the corresponding author for thearticle.
Als1p-N from Candida albicans binds to fucose glycans 13
© 2011 Blackwell Publishing Ltd, Molecular Microbiology




















