
The Mycobacterium tuberculosis membrane protein Rv2560 − biochemical and functional studies
13
The Mycobacterium tuberculosis membrane protein Rv2560 ) biochemical and functional studies David F. Plaza 1 , Hernando Curtidor 2,4 , Manuel A. Patarroyo 1,4 , Julie A. Chapeton-Montes 1 , Claudia Reyes 3 , Jose Barreto 3 and Manuel E. Patarroyo 1,5 1 Molecular Biology Department, Fundacion Instituto de Inmunologia de Colombia, Bogota, Colombia 2 Receptor–Ligand Department, Fundacion Instituto de Inmunologia de Colombia, Bogota, Colombia 3 Chemical Synthesis Department, Fundacion Instituto de Inmunologia de Colombia, Bogota, Colombia 4 Universidad del Rosario, Bogota, Colombia 5 Universidad Nacional de Colombia, Bogota, Colombia Tuberculosis (TB) is a major public health problem throughout the world, affecting almost nine million people [1] and causing more than three million deaths per year. An increasing incidence of TB, related to the high risk of developing the disease in immuno- suppressed individuals and the increasing proportion of Mycobacterium tuberculosis drug-resistant strains, has contributed to this problem [2,3]. This makes the Keywords high-activity binding peptide; invasion inhibition; Mycobacterium tuberculosis – host cell interaction; Rv2560 membrane protein Correspondence M. E. Patarroyo, Carrera 50 # 26-00, Bogota, Colombia Fax: +57 (1) 4815269 Tel: +57 (1) 4815219 or +57 (1) 3244672 Ext 125 E-mail: [email protected] (Received 31 July 2007, revised 11 October 2007, accepted 17 October 2007) doi:10.1111/j.1742-4658.2007.06153.x The characterization of membrane proteins having no identified function in Mycobacterium tuberculosis is important for a better understanding of the biology of this pathogen. In this work, the biological activity of the Rv2560 protein was characterized and evaluated. Primers used in PCR and RT-PCR assays revealed that the gene encoding protein Rv2560 is present in M. tuberculosis complex strains, but transcribed in only some of them. Sera obtained from rabbits inoculated with polymer peptides from this pro- tein recognized a 33 kDa band in the M. tuberculosis lysate and a mem- brane fraction corresponding to the predicted molecular mass (33.1 kDa) of this protein. Immunoelectron microscopy analysis found this protein on the mycobacterial membrane. Sixteen peptides covering its entire length were chemically synthesized and tested for their ability to bind to A549 and U937 cells. Peptide 11024 (121VVALSDRATTAYTNTSGVSS140) showed high specific binding to both cell types (dissociation constants of 380 and 800 nm, respectively, and positive receptor–ligand interaction cooperativity), whereas peptide 11033 (284LIGIPVAALIHVYTYRKLS GG304) displayed high binding activity to A549 cells only. Cross-linking assays showed the specific binding of peptide 11024 to a 54 kDa membrane protein on U937. Invasion inhibition assays, in the presence of shared high-activity binding peptide identified for U937 and A549 cells, presented maximum inhibition percentages of 50.53% and 58.27%, respectively. Our work highlights the relevance of the Rv2560 protein in the M. tuberculosis invasion process of monocytes and epithelial cells, and represents a funda- mental step in the rational selection of new antigens to be included as components in a multiepitope, subunit-based, chemically synthesized, anti- tuberculosis vaccine. Abbreviations GRAVY, grand average of hydropathicity; HABP, high-activity binding peptide; SPf66, synthetic Plasmodium falciparum 66; SSP, simple sequence protein; TB, tuberculosis. 6352 FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS
Transcript of The Mycobacterium tuberculosis membrane protein Rv2560 − biochemical and functional studies

The Mycobacterium tuberculosis membrane proteinRv2560 ) biochemical and functional studiesDavid F. Plaza1, Hernando Curtidor2,4, Manuel A. Patarroyo1,4, Julie A. Chapeton-Montes1,Claudia Reyes3, Jose Barreto3 and Manuel E. Patarroyo1,5
1 Molecular Biology Department, Fundacion Instituto de Inmunologia de Colombia, Bogota, Colombia
2 Receptor–Ligand Department, Fundacion Instituto de Inmunologia de Colombia, Bogota, Colombia
3 Chemical Synthesis Department, Fundacion Instituto de Inmunologia de Colombia, Bogota, Colombia
4 Universidad del Rosario, Bogota, Colombia
5 Universidad Nacional de Colombia, Bogota, Colombia
Tuberculosis (TB) is a major public health problem
throughout the world, affecting almost nine million
people [1] and causing more than three million deaths
per year. An increasing incidence of TB, related to the
high risk of developing the disease in immuno-
suppressed individuals and the increasing proportion
of Mycobacterium tuberculosis drug-resistant strains,
has contributed to this problem [2,3]. This makes the
Keywords
high-activity binding peptide; invasion
inhibition; Mycobacterium tuberculosis –
host cell interaction; Rv2560 membrane
protein
Correspondence
M. E. Patarroyo, Carrera 50 # 26-00,
Bogota, Colombia
Fax: +57 (1) 4815269
Tel: +57 (1) 4815219 or +57 (1) 3244672
Ext 125
E-mail: [email protected]
(Received 31 July 2007, revised 11 October
2007, accepted 17 October 2007)
doi:10.1111/j.1742-4658.2007.06153.x
The characterization of membrane proteins having no identified function in
Mycobacterium tuberculosis is important for a better understanding of the
biology of this pathogen. In this work, the biological activity of the
Rv2560 protein was characterized and evaluated. Primers used in PCR and
RT-PCR assays revealed that the gene encoding protein Rv2560 is present
in M. tuberculosis complex strains, but transcribed in only some of them.
Sera obtained from rabbits inoculated with polymer peptides from this pro-
tein recognized a 33 kDa band in the M. tuberculosis lysate and a mem-
brane fraction corresponding to the predicted molecular mass (33.1 kDa)
of this protein. Immunoelectron microscopy analysis found this protein on
the mycobacterial membrane. Sixteen peptides covering its entire length
were chemically synthesized and tested for their ability to bind to A549
and U937 cells. Peptide 11024 (121VVALSDRATTAYTNTSGVSS140)
showed high specific binding to both cell types (dissociation constants of
380 and 800 nm, respectively, and positive receptor–ligand interaction
cooperativity), whereas peptide 11033 (284LIGIPVAALIHVYTYRKLS
GG304) displayed high binding activity to A549 cells only. Cross-linking
assays showed the specific binding of peptide 11024 to a 54 kDa membrane
protein on U937. Invasion inhibition assays, in the presence of shared
high-activity binding peptide identified for U937 and A549 cells, presented
maximum inhibition percentages of 50.53% and 58.27%, respectively. Our
work highlights the relevance of the Rv2560 protein in the M. tuberculosis
invasion process of monocytes and epithelial cells, and represents a funda-
mental step in the rational selection of new antigens to be included as
components in a multiepitope, subunit-based, chemically synthesized, anti-
tuberculosis vaccine.
Abbreviations
GRAVY, grand average of hydropathicity; HABP, high-activity binding peptide; SPf66, synthetic Plasmodium falciparum 66; SSP, simple
sequence protein; TB, tuberculosis.
6352 FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS

development of new therapies and ⁄or vaccines, or the
improvement of existing ones, necessary to control this
disease.
The identification and characterization of the pro-
teins involved in the interaction of M. tuberculosis with
its target cells and the mechanisms for evading an
immune response produced against this pathogen are
thus of crucial interest. Of special interest are those
proteins found on the cell membrane and in the
secreted fraction, as they represent the ‘face’ shown by
the bacterium to its host, thereby leading to its identifi-
cation and elimination.
Although TB is mainly a pulmonary disease, it can
also affect other organs. Few microorganisms can sur-
vive within macrophages because of the abundance of
hydrolytic enzymes contained in acid phagocytic com-
partments [4]. M. tuberculosis has developed evolution-
ary mechanisms that allow it to penetrate, survive and
grow within these compartments [5]. Adhesion and
phagocytosis are the initial steps in the entry of the
bacterium into macrophages, processes triggered by
receptor–ligand interactions. These interactions are
complex and have an important influence on the intra-
cellular fate of M. tuberculosis [6,7], being mediated
mainly by membrane proteins present on the bacillus
and its host [8]. The M. tuberculosis cell membrane,
and that of other prokaryotes, contains proteins func-
tioning as enzymes, ligands, signal transducers or viru-
lence factors, all of great importance for bacterial
survival [9].
M. tuberculosis invades alveolar epithelial cells,
thereby inducing macropinocytosis, using secreted or
membrane components [10]. The identification of the
bacterial peptides involved in this process has been
poorly studied, and represents an objective of this
work.
Membrane proteins represent a bank of antigens
available for the design of a subunit-based, multiepi-
tope vaccine, as they may be involved in the patho-
genicity of the bacterium and may also be highly
susceptible to immune recognition. The M. tuberculosis
membrane proteome has revealed the presence of mul-
tiple immunogenic components with unknown function
[11–13], which could be used for this purpose.
Bioinformatics represents a preliminary method for
the evaluation of the presence of a determined gene or
protein sequence in different organisms, and for pre-
dicting (with some degree of confidence) the existence
of membrane anchoring or signal export regions within
them, making it a fundamental tool when screening
and ⁄or selecting for a vaccine candidate [14,15]. Fur-
thermore, work carried out on the proteomics of the
bacterium has led to the identification of a large
number of membrane proteins with an as yet undeter-
mined function [11–13].
A previous study has shown the importance of the
melA gene (encoding an Rv2560 homologous protein)
in Mycobacterium marinum invasion of THP-1 cells,
indicating that its homologous gene might be relevant
in M. tuberculosis–host cell interaction [16].
This work has led to the identification of the pres-
ence of the Rv2560 proline- and glycine-rich trans-
membrane protein encoding gene and its transcripts in
the M. tuberculosis complex and clinical isolate strains,
as well as the characterization of the high-activity
binding peptide (HABP) involved in the binding to
and invasion of monocytes (U937) and type II alveolar
epithelial cells (A549), using synthetic peptides. The
protein encoded by the Rv2560 gene was synthesized
in 20-mer-long, nonoverlapping peptides, which were
analysed by a highly specific and robust methodology
(widely used in Plasmodium falciparum research) for
the identification of the HABPs interacting with U937
and A549 cells (in this case), their binding dissociation
constants, their critical binding residues and their role
in cell invasion inhibition, with the specific purpose of
employing them as probable components of a multiepi-
tope, subunit-based, synthetic vaccine. This has impor-
tant biological significance and implications for the
development of strategies to control this disease.
Results
Bioinformatic analysis
Different bioinformatics’ tools were used to predict the
cell location of Rv2560 and its transmembrane topol-
ogy. Although signalp did not predict the existence of
a signal peptide, the high grand average of hydropath-
icity (GRAVY) value (0.488) and the existence of
four possible transmembrane helices in the complete
Rv2560 amino acid sequence suggested its presence on
the bacterial surface. Membrane topology prediction
proposed four transmembrane helices (i103–125o,
o145–167i, i197–228o and o271–293I, where i denotes
inside and o denotes outside), suggesting that the iden-
tified HABPs are located in the protein’s solvent-
exposed region (Fig. 4, topology diagram, see later).
Genomic PCR assay
Two specific primers, flanking an HABP region
(encoding amino acids 80–290), were designed and syn-
thesized to determine the presence or absence of the
gene encoding the Rv2560 protein in M. tuberculosis
complex strains. A single 631 bp amplification band
D. F. Plaza et al. M. tuberculosis Rv2560 protein characterization
FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS 6353

was observed in the following strains: M. tuberculosis
H37Rv, M. tuberculosis H37Ra, Mycobacterium bovis,
M. bovis BCG, Mycobacterium africanum and Myco-
bacterium microti (Fig. 1A, lanes 2–7). tblastn analy-
sis showed similar (although not identical) genes in
M. marinum and Mycobacterium ulcerans.
DNA sequencing
The amplified fragments in the different clinical iso-
lates studied were purified, and their DNA was
sequenced to determine this region’s polymorphism. It
was observed that this region was completely con-
served in all M. tuberculosis clinical isolates analysed,
independent of geographical origin. This result was
confirmed by sequencing each fragment’s complemen-
tary chain (data not shown).
RT-PCR assay
Two specific primers were designed and synthesized to
determine the transcriptional pattern of the Rv2560
gene in M. tuberculosis complex strains. A single
308 bp amplification band was observed in some
M. tuberculosis complex strains (Fig. 1B, lanes 2–7).
This gene’s transcription was exclusive to M. tuberculo-
sis H37Rv, M. tuberculosis H37Ra, M. bovis BCG and
M. africanum strains, but not M. bovis and M. microti,
suggesting that it was not transcribed, in spite of this
gene being present as assessed by PCR. Figure 1C
shows a 360 bp conserved fragment from a mycobacte-
rial genus constitutive gene (the rpoB gene encoding
the RNA polymerase b-subunit) used as transcription
control.
Western blot
Polymerized synthetic peptides corresponding to the
Rv2560 protein peptide 11027 (181ADGKPVTIATFF
RPRNLGLV200) and 11031 (242SPIDSVKASIETV
GSNIGGSVY262) amino acid sequences, located in
the extracellular solvent-exposed region (as predicted
by the TMHMM server), were used as immunogens in
rabbits, employing a 0, 20 and 40 day inoculation
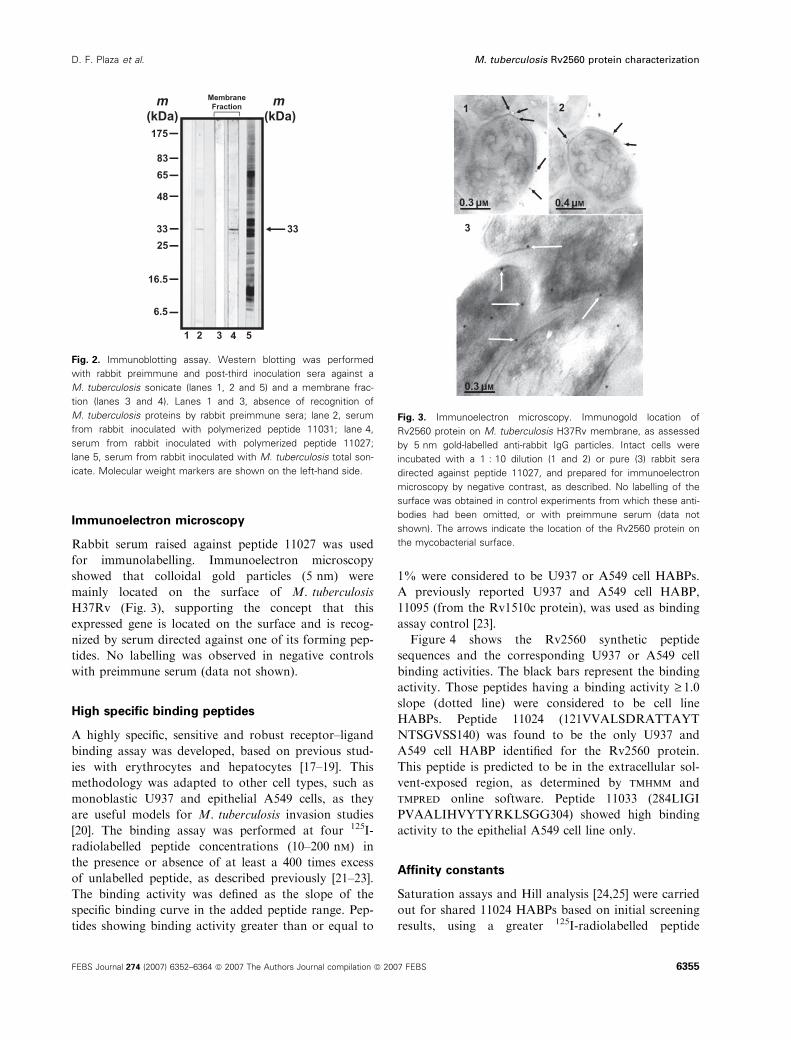
scheme. Western blot (Fig. 2) was performed with
preimmune and post-third inoculation rabbit sera
against M. tuberculosis sonicate and membrane frac-
tion. Sera had previously been adsorbed on columns
with Escherichia coli and Mycobacterium smegmatis
lysates, as well as synthetic Plasmodium falcipa-
rum 66 (SPf66), prior to western blotting to eliminate
the cross-reactivity against bacterial (E. coli), myco-
bacterial (M. smegmatis) and polymerization (SPf66)
neo-antigens.
Lanes 1 and 3 in Fig. 2 show the lack of preimmune
rabbit sera protein recognition in the M. tuberculosis
sonicate and membrane fraction, respectively. Rabbits
immunized with 11031 polymerized peptide developed
polyclonal antibodies specifically recognizing a 33 kDa
molecular mass band in the M. tuberculosis sonicate,
close to the protein’s molecular mass (33.1 kDa) in
lane 2. Preimmune serum (lane 3) did not recognize
bands in the membrane fraction; however, postimmune
serum from 11027 immunized rabbits strongly recog-
nized a band close to 33 kDa in this fraction (lane 4),
suggesting that this protein becomes enriched in the
cell membrane.
Fig. 1. Genomic PCR and RT-PCR assays. (A) A 631 bp PCR prod-
uct from the gene encoding M. tuberculosis H37Rv Rv2560 protein
was only amplified on DNA from M. tuberculosis complex strains
(lanes 2–7). Lane 1, 1 kb molecular weight marker (Gibco); lane 2,
M. tuberculosis H37Rv; lane 3, M. tuberculosis H37Ra; lane 4,
M. bovis; lane 5, M. bovis BCG; lane 6, M. africanum; lane 7,
M. microti; lane 8, PCR negative control. (B) M. tuberculosis
Rv2560 protein H37Rv 308 bp RT-PCR product obtained from
M. tuberculosis complex strain cDNA. Lane 1, 1 kb molecular
weight marker (Gibco); lane 2, M. tuberculosis H37Rv; lane 3,
M. tuberculosis H37Ra; lane 4, M. bovis; lane 5, M. bovis BCG;
lane 6, M. africanum; lane 7, M. microti; lane 8, negative control
[M. tuberculosis H37Rv DNA treated with DNAse Q (Promega)];
lane 9, PCR positive control (M. tuberculosis H37Rv DNA); lane 10,
PCR negative control. A single 308 bp amplification band was
observed in some M. tuberculosis complex strains. (C) PCR 360 bp
product from the same strains as (B) but with amplification of the
Mycobacterium rpoB gene as positive control for mRNA expression
and cDNA synthesis.
M. tuberculosis Rv2560 protein characterization D. F. Plaza et al.
6354 FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS
Immunoelectron microscopy
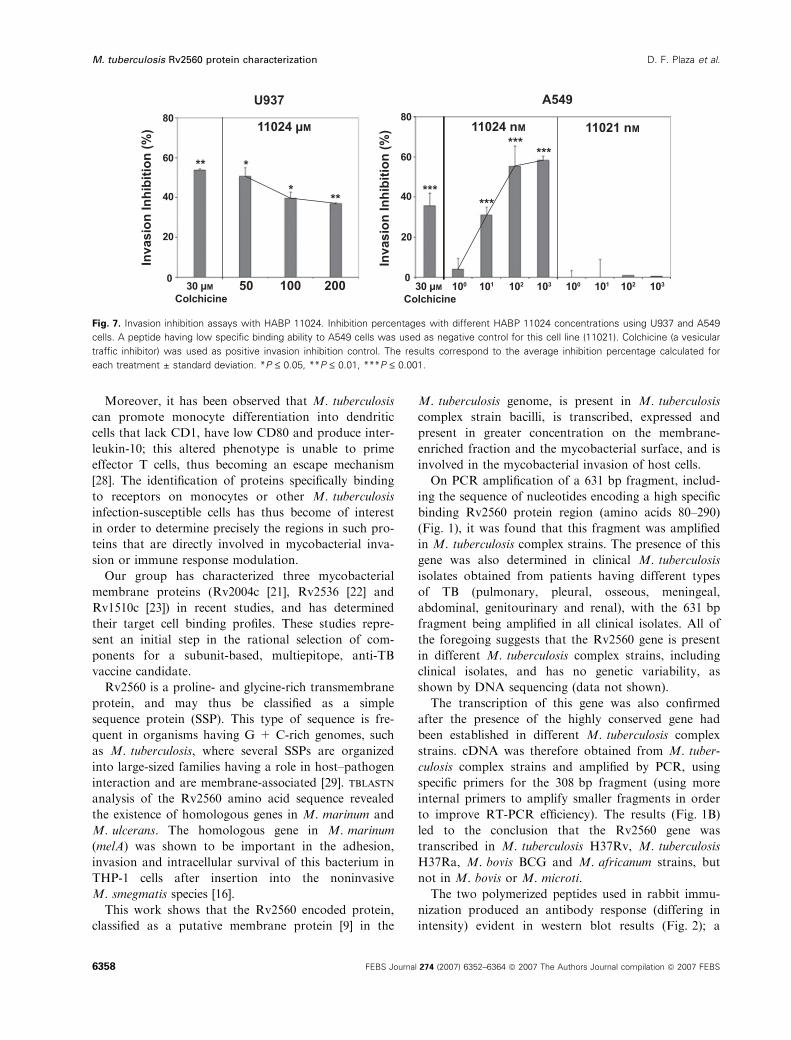
Rabbit serum raised against peptide 11027 was used
for immunolabelling. Immunoelectron microscopy
showed that colloidal gold particles (5 nm) were
mainly located on the surface of M. tuberculosis
H37Rv (Fig. 3), supporting the concept that this
expressed gene is located on the surface and is recog-
nized by serum directed against one of its forming pep-
tides. No labelling was observed in negative controls
with preimmune serum (data not shown).
High specific binding peptides
A highly specific, sensitive and robust receptor–ligand
binding assay was developed, based on previous stud-
ies with erythrocytes and hepatocytes [17–19]. This
methodology was adapted to other cell types, such as
monoblastic U937 and epithelial A549 cells, as they
are useful models for M. tuberculosis invasion studies
[20]. The binding assay was performed at four 125I-
radiolabelled peptide concentrations (10–200 nm) in
the presence or absence of at least a 400 times excess
of unlabelled peptide, as described previously [21–23].
The binding activity was defined as the slope of the
specific binding curve in the added peptide range. Pep-
tides showing binding activity greater than or equal to
1% were considered to be U937 or A549 cell HABPs.
A previously reported U937 and A549 cell HABP,
11095 (from the Rv1510c protein), was used as binding
assay control [23].
Figure 4 shows the Rv2560 synthetic peptide
sequences and the corresponding U937 or A549 cell
binding activities. The black bars represent the binding
activity. Those peptides having a binding activity ‡ 1.0
slope (dotted line) were considered to be cell line
HABPs. Peptide 11024 (121VVALSDRATTAYT
NTSGVSS140) was found to be the only U937 and
A549 cell HABP identified for the Rv2560 protein.
This peptide is predicted to be in the extracellular sol-
vent-exposed region, as determined by tmhmm and
tmpred online software. Peptide 11033 (284LIGI
PVAALIHVYTYRKLSGG304) showed high binding
activity to the epithelial A549 cell line only.
Affinity constants
Saturation assays and Hill analysis [24,25] were carried
out for shared 11024 HABPs based on initial screening
results, using a greater 125I-radiolabelled peptide
Fig. 2. Immunoblotting assay. Western blotting was performed
with rabbit preimmune and post-third inoculation sera against a
M. tuberculosis sonicate (lanes 1, 2 and 5) and a membrane frac-
tion (lanes 3 and 4). Lanes 1 and 3, absence of recognition of
M. tuberculosis proteins by rabbit preimmune sera; lane 2, serum
from rabbit inoculated with polymerized peptide 11031; lane 4,
serum from rabbit inoculated with polymerized peptide 11027;
lane 5, serum from rabbit inoculated with M. tuberculosis total son-
icate. Molecular weight markers are shown on the left-hand side.
Fig. 3. Immunoelectron microscopy. Immunogold location of
Rv2560 protein on M. tuberculosis H37Rv membrane, as assessed
by 5 nm gold-labelled anti-rabbit IgG particles. Intact cells were
incubated with a 1 : 10 dilution (1 and 2) or pure (3) rabbit sera
directed against peptide 11027, and prepared for immunoelectron
microscopy by negative contrast, as described. No labelling of the
surface was obtained in control experiments from which these anti-
bodies had been omitted, or with preimmune serum (data not
shown). The arrows indicate the location of the Rv2560 protein on
the mycobacterial surface.
D. F. Plaza et al. M. tuberculosis Rv2560 protein characterization
FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS 6355

concentration range (500–2000 nm) (Fig. 5A). Scat-
chard and Hill analyses were then performed and dis-
sociation constants (Kd) were calculated for Rv2560
HABP 11024 for U937 and A549 cell lines (Kd of 380
and 800 nm, respectively). The Hill coefficient (nh) was
1.5 for U937 and 1.2 for A549 cells, suggesting positive
ligand–receptor cooperativity. The numbers of binding
sites per cell calculated for peptide 11024 were 200 000
and 3 000 000 for U937 and A549 cells, respectively.
Critical residues for cell binding in U937 cells
Glycine scanning analogue peptides were synthesized,
and competition cell binding assays were performed
between original radiolabelled peptides and their ana-
logue peptides to identify HABP critical amino acids
for U937 cell binding. Critical residues were those
which, on replacement with glycine, rendered an
invariable decrease of at least 50% in the ability to
compete with original radiolabelled peptide in a bind-
ing assay at two concentrations (16 and 400 nm). This
assumed dramatic changes in peptide binding activity.
Figure 5B shows that critical residues in peptide 11024
binding were VVALSDRATTAYTNTSGVSS (shown
in bold italic in the sequence).
Cross-linking assays
HABP 11024 bound to a protein located on the U937
cell membrane, specifically recognizing a 54 kDa hypo-
thetical receptor. Radiolabelled peptide binding to this
receptor protein was inhibited in the presence of non-
radiolabelled peptide; this was clearly shown by the
band intensities (Fig. 6). Peptide 11024 binding was
target cell specific, as it did not show any binding to
HepG2 cells or erythrocytes (data not shown).
Invasion inhibition assay using HABPs
A549 and U937 cell lines were preincubated in the
presence of HABP 11024 in two independent experi-
ments to evaluate the effect of HABP on bacterial
invasion of target lung cells. It was found that invasion
was inhibited in both cell types. The inhibitory power
of HABP 11024 was greater in assays carried out using
the A549 cell line, where it reached close to 60% inhi-
bition when such cells were preincubated in the pres-
ence of 1 lm peptide. Monocyte invasion assays
revealed around 50% maximum inhibition only when
higher peptide concentrations were used than those
employed for A549 cell assays (Fig. 7). It was also
Fig. 4. Cell binding activity of Rv2560 peptides. Amino acid sequence and specific U937 and A549 cell binding activity for 20-mer long chem-
ically synthesized Rv2560 peptides from the M. tuberculosis H37Rv strain. The peptides are given on the left-hand side, with numbers indi-
cating their position within the native protein; in peptides that did not contain tyrosine, it was added to the carboxy-terminal end. To the
right, the black bar represents each peptide binding activity, determined as the specific binding ⁄ total added peptide ratio. The dotted line
separates peptides having ‡ 1% binding activity. HABP 11095 was used as a binding activity control. Rv2560 transmembrane topology is
presented on the left-hand side. Grey shows the transmembrane helices present in the sequence according to the TMHMM prediction server.
nd, not done because of solubilization problems.
M. tuberculosis Rv2560 protein characterization D. F. Plaza et al.
6356 FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS

observed that the inhibitory activity of HABP 11024
decreased at concentrations equal to or greater than
100 lm. The secondary structure of HABP 11024 in
solution at different concentrations (including those in
which activity was seen to decrease) was verified by
CD, and it was shown that this HABP underwent
conformational changes critical for its function at the
concentrations already established in functional assays
(data not shown).
Discussion
Although the elucidation of the M. tuberculosis gen-
ome sequence has revealed the presence of many gene
families which can be classified according to their
homology to genes having a known function in other
microorganisms, no functional counterparts have been
found for many of these mycobacterial genes. Pro-
teome studies have led to the determination of some of
the proteins present on the membrane for which no
role in M. tuberculosis invasion of the host cell has yet
been determined [12,13].
The infection of epithelial cells in the alveoli requires
the interaction of the bacterium with several surface
receptors, which have been poorly studied to date [26].
Studies of nonpolarized pulmonary epithelial cells
(HEp-2) have revealed the existence of at least five
M. tuberculosis proteins that bind to biotinylated cell
fractions, including the already characterized adhesion
heparin-binding haemagglutinin [27].
A B
Fig. 5. Saturation curves and critical residues of cell binding. (A) Saturation binding curve for HABP 11024 binding to U937 and A549 cells.
The saturation curves resulted from plotting the specifically bound 125I-HABP concentration versus free 125I-HABP. The affinity constants and
maximum number of sites per cell were obtained from these curves. Inset: the abscissa is log F in the Hill plot and the ordinate is
log [B ⁄ Bm ) B], where Bm is the maximum bound peptide, B is the bound peptide and F is free peptide. (B) Competition binding assay with
analogous peptides. Specific original radiolabelled peptide binding inhibited by analogous peptide (at 16 and 400 nM) is shown. Amino acids
underlined in bold represent the critical binding residues, as their binding activity decreased by 50% or more when modified and assayed at
the two concentrations used.
Fig. 6. Cross-linking assay for peptide 11024. Autoradiographs for
U937 proteins specifically cross-linked with radiolabelled peptide
11024. Lanes 1 and 2, U937 cells; lanes 3 and 4, HepG2 cells;
lanes 1 and 3, total radiolabelled peptide binding; lanes 2 and 4,
inhibited binding. The cross-linking assays show that peptide 11024
specifically binds to a 54 kDa membrane protein on U937 cells.
D. F. Plaza et al. M. tuberculosis Rv2560 protein characterization
FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS 6357
Moreover, it has been observed that M. tuberculosis
can promote monocyte differentiation into dendritic
cells that lack CD1, have low CD80 and produce inter-
leukin-10; this altered phenotype is unable to prime
effector T cells, thus becoming an escape mechanism
[28]. The identification of proteins specifically binding
to receptors on monocytes or other M. tuberculosis
infection-susceptible cells has thus become of interest
in order to determine precisely the regions in such pro-
teins that are directly involved in mycobacterial inva-
sion or immune response modulation.
Our group has characterized three mycobacterial
membrane proteins (Rv2004c [21], Rv2536 [22] and
Rv1510c [23]) in recent studies, and has determined
their target cell binding profiles. These studies repre-
sent an initial step in the rational selection of com-
ponents for a subunit-based, multiepitope, anti-TB
vaccine candidate.
Rv2560 is a proline- and glycine-rich transmembrane
protein, and may thus be classified as a simple
sequence protein (SSP). This type of sequence is fre-
quent in organisms having G + C-rich genomes, such
as M. tuberculosis, where several SSPs are organized
into large-sized families having a role in host–pathogen
interaction and are membrane-associated [29]. tblastn
analysis of the Rv2560 amino acid sequence revealed
the existence of homologous genes in M. marinum and
M. ulcerans. The homologous gene in M. marinum
(melA) was shown to be important in the adhesion,
invasion and intracellular survival of this bacterium in
THP-1 cells after insertion into the noninvasive
M. smegmatis species [16].
This work shows that the Rv2560 encoded protein,
classified as a putative membrane protein [9] in the
M. tuberculosis genome, is present in M. tuberculosis
complex strain bacilli, is transcribed, expressed and
present in greater concentration on the membrane-
enriched fraction and the mycobacterial surface, and is
involved in the mycobacterial invasion of host cells.
On PCR amplification of a 631 bp fragment, includ-
ing the sequence of nucleotides encoding a high specific
binding Rv2560 protein region (amino acids 80–290)
(Fig. 1), it was found that this fragment was amplified
in M. tuberculosis complex strains. The presence of this
gene was also determined in clinical M. tuberculosis
isolates obtained from patients having different types
of TB (pulmonary, pleural, osseous, meningeal,
abdominal, genitourinary and renal), with the 631 bp
fragment being amplified in all clinical isolates. All of
the foregoing suggests that the Rv2560 gene is present
in different M. tuberculosis complex strains, including
clinical isolates, and has no genetic variability, as
shown by DNA sequencing (data not shown).
The transcription of this gene was also confirmed
after the presence of the highly conserved gene had
been established in different M. tuberculosis complex
strains. cDNA was therefore obtained from M. tuber-
culosis complex strains and amplified by PCR, using
specific primers for the 308 bp fragment (using more
internal primers to amplify smaller fragments in order
to improve RT-PCR efficiency). The results (Fig. 1B)
led to the conclusion that the Rv2560 gene was
transcribed in M. tuberculosis H37Rv, M. tuberculosis
H37Ra, M. bovis BCG and M. africanum strains, but
not in M. bovis or M. microti.
The two polymerized peptides used in rabbit immu-
nization produced an antibody response (differing in
intensity) evident in western blot results (Fig. 2); a
Fig. 7. Invasion inhibition assays with HABP 11024. Inhibition percentages with different HABP 11024 concentrations using U937 and A549
cells. A peptide having low specific binding ability to A549 cells was used as negative control for this cell line (11021). Colchicine (a vesicular
traffic inhibitor) was used as positive invasion inhibition control. The results correspond to the average inhibition percentage calculated for
each treatment ± standard deviation. *P £ 0.05, **P £ 0.01, ***P £ 0.001.
M. tuberculosis Rv2560 protein characterization D. F. Plaza et al.
6358 FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS

33 kDa band corresponding to the theoretical molecu-
lar mass of the Rv2560 protein (33.1 kDa) was recog-
nized by these sera, suggesting that this protein is
transcribed and expressed in M. tuberculosis sonicate,
and at much higher concentration in the membrane
fraction, as inferred from the band intensity.
When using serum from rabbits immunized with poly-
merized peptide 11027, immunoelectron microscopy
confirmed the presence of this protein on the mycobac-
terial surface (which has not been reported to date).
Receptor–ligand assays led to the identification of
two high specific binding peptides, one (11024) being
shared by both U937 monoblast and A549 epithelial
cells. Peptide 11024 binding to both cell types was sat-
urable, presenting strong positive cooperativity, as
shown by the dissociation constant and Hill coefficient.
The corresponding analyses for the determination of
the receptor for 11024 HABPs and the identification of
the critical binding residues, and the invasion inhibi-
tion assays, were thus performed exclusively with this
peptide.
This peptide is located in the solvent-exposed central
region of the protein between residues 121 and 140,
displaying the VVALSDRATTAYTNTSGVSS
sequence (where the amino acids shown in bold italic
correspond to those identified as being critical in 11024
binding to U937 cells). These residues may be involved
directly in target cell binding or may be important for
the structural characteristics of the peptide, thereby
allowing specific binding. BLAST analysis of this small
region led to the identification of 78% homology with
Burkholderia cenocepacia-dependent AMP synthase,
which causes respiratory tract infection and is cyto-
toxic for alveolar epithelial cells [30].
A 54 kDa protein was found to be a receptor on
U937 cells; its binding by peptide 11024 was inhibited
by an excess of nonradiolabelled peptide. This receptor
was specific for the U937 cell line, but was not seen in
hepatocytes or erythrocytes. A receptor having a simi-
lar molecular mass (51 kDa) has been described for
the Rv1510c protein HABP 11095; however, further
studies are required to determine whether the same
membrane protein is involved.
Peptide 11024 (for which high monocyte and alveo-
lar epithelial cell binding has already been determined)
induced a 36.96–50.53% decrease in M. tuberculosis
invasion of U937 cells and a 30.98–58.27% decrease in
M. tuberculosis invasion of A549 cells (Fig. 7). This
indicates that this HABP is important in the recogni-
tion and invasion of monocytes and type II alveolar
epithelial cells by the mycobacterium.
It was observed in preliminary assays that HABP
concentrations of less than 100 lm had a greater
inhibitory effect than larger concentrations. Therefore,
additional CD secondary structural studies were per-
formed to verify possible conformational changes at
concentrations greater than 100 lm which were critical
in the inhibitory activity of this peptide. It was found
that the structure of the peptide changed at concentra-
tions equal to or greater than 100 lm (not shown),
thereby explaining the loss of inhibitory potential in
the assay carried out for alveolar epithelial cells.
The above results indicate that the Rv2560 protein
is conserved within the M. tuberculosis complex. It is
also transcribed and expressed in low abundance on
the mycobacterial surface. The protein contains a spe-
cific high binding peptide for U937 and A549 cells,
which plays an important role in mycobacterial inva-
sion. Therefore, it is concluded that the Rv2560 pro-
tein is highly relevant in pathogen–host interaction,
and that the identification of the region directly
involved in such interaction makes it an important
candidate for inclusion in the development of a
subunit-based, multiepitope, chemically synthesized
vaccine.
Experimental procedures
Bioinformatics’ analysis
M. tuberculosis Rv2560 protein sequences were obtained
from the Tuberculist Webserver (http://genolist.pasteur.fr/
TubercuList/). Proteins were aligned by BLAST (http://
www.ncbi.nlm.nih.gov/BLAST/) and clustalw (http://
npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_clu
stalw.html) multiple alignments. Protein features and
GRAVY scores for the Rv2560 amino acid sequence were
calculated using the protparam tool (http://us.expasy.
org ⁄ tools ⁄protparam.html), for which a score of greater
than ) 0.4 (mean score for the cytosolic proteins) suggests
membrane association probability; the higher the score, the
greater the probability [13]. Transmembrane regions were
predicted by tmhmm (http://www.cbs.dtu.dk/services/
TMHMM) and tmpred (http://www.ch.embnet.org/
software/TMPRED_form.html). Cell localization, lipid
attachment sites and signal sequences were predicted using
psort (http://psort.nibb.ac.jp).
Mycobacterial species and strains
The following M. tuberculosis complex species and strains
were used: M. tuberculosis H37Rv (ATCC27294), M. tuber-
culosis H37Ra (ATCC25177), M. bovis (ATCC19210),
M. bovis BCG (ATCC27291, Pasteur sub strain), M. africa-
num (ATCC25420) and M. microti (kindly donated by
F. Portaels, Prince Leopold Institute of Tropical Medicine).
D. F. Plaza et al. M. tuberculosis Rv2560 protein characterization
FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS 6359

Ten M. tuberculosis clinical strains were isolated from speci-
mens obtained from different patients attending the TB pro-
gramme at either the San Juan de Dios Hospital or Santa
Clara Hospital, both in Bogota, Colombia. Patients came
from several geographical areas of Colombia. All mycobacte-
rial strains were grown for 5–15 days in 7H9 Middlebrook
broth (Difco Laboratory, Detroit, MI, USA) with 0.04%
Tween 80 and in Middlebrook agar 7H10 (Difco), both sup-
plemented with oleic acid, albumin, dextrose and catalase
(BBL, Becton Dickinson, Mountain View, CA, USA).
Genomic DNA extraction
Cells were harvested in Tris–EDTA buffer (TE, pH 8.0)
and suspended in 25% sucrose in 10 · TE. Lysozyme
(100 lgÆmL)1) was added and the preparation was incubated
at 37 �C for 2 h with subsequent addition of 100 lgÆmL)1
proteinase K and N-lauroylsarcosine (Sigma, St Louis, MO,
USA) at 1% final concentration. Following 18 h incubation
at 65 �C, the mixture was extracted with chloroform–iso-
amyl alcohol (24 : 1, v ⁄ v), once with phenol–chloroform
(1 : 1, v ⁄ v) and twice more with chloroform–isoamyl alco-
hol. DNA was precipitated with 2-propanol, washed with
70% (v ⁄ v) ethanol and suspended in 1 · TE [31,32].
PCR assay
PCR amplifications were performed in a thermal cycler
GeneAmp PCR System 9600 (Perkin-Elmer Life Sciences
Inc., Boston, MA, USA), using 100 ng mycobacterial geno-
mic DNA for amplifications. The mixture contained 50 mm
KCl, 10 mm Tris ⁄HCl (pH 8.3), 1.5 mm MgCl2, 0.1 mm of
each dNTP, 0.4 mm of both direct 5¢-CAATCGTCG
GGTTTTAGCG-3¢ and reverse 5¢-GTGTAGACGTG
GATGAGCG-3¢ oligonucleotide primers and 1.5 U Taq
DNA polymerase (Invitrogen, Carlsbad, CA, USA).
Twenty-five cycles of the following thermal profile were car-
ried out after DNA had been denatured for 5 min at 94 �C:56 �C for 30 s, 72 �C for 40 s and 95 �C for 40 s. A final
5 min extension cycle was then performed at 72 �C. The
amplification products were loaded onto 1% agarose gel,
electrophoresed at 70 V in 1 · Tris/acetate/EDTA, stained
with ethidium bromide and photographed.
DNA sequencing
The dideoxy chain termination method was used for
sequencing reactions with a Taq FS DyeDeoxy Terminator
cycle sequencing kit (Applied Biosystems, Foster City, CA,
USA) on a GeneAmp PCR System 9600 (Perkin-Elmer Life
Sciences Inc.), and run on a 373 DNA sequencer model
(Applied Biosystems). The sequencing strategy involved for-
ward and reverse specific primers for the 631 bp fragment
of the Rv2560 gene.
RNA isolation
The bacilli were harvested by spinning at 12 000 g for
15 min at 4 �C. Sodium azide (10 mm) was added to the
culture just before harvesting. The cell pellet was suspended
in 2 mL of cold lysis buffer for each 200 mg wet weight of
cells [33], and sonicated twice for 15 min. Two volumes of
Trizol (Gibco BRL, Gaithersburg, MD, USA) were then
added and extracted according to the manufacturer’s
instructions. The pellet was suspended in 100 lL of distilled
water and stored in aliquots at ) 80 �C.
RT-PCR
Total RNA was quantified by a GeneQuant spectropho-
tometer (Pharmacia Biotech, Piscataway, NJ, USA), treated
with RNAse-free DNAse RQ1 at 37 �C for 3 h, precipi-
tated with isopropanol, washed with 70% ethanol and sus-
pended in distilled water. M. tuberculosis H37Rv DNA was
included as DNAse Q activity control (1 UÆlg)1 DNA).
Target RNA (500 lg) was reverse transcribed in a tube
containing distilled water and 10 lgÆmL)1 random primers
(Gibco BRL). This mixture was incubated for 10 min at
70 �C; 1 · RT buffer, 10 mm dithiothreitol, 0.5 mm dNTPs
and 40 U human placenta ribonuclease inhibitor (Promega,
Madison, WI, USA) were then added on ice; 200 U
M-MLV reverse transcriptase (Gibco-Life Technologies,
Grand Island, NY, USA) was then added at a final volume
of 30 lL. This mixture was kept at 37 �C for 1 h. The
enzyme was finally denatured for 5 min at 95 �C. PCR was
carried out as described above.
The rpoB gene was used as positive transcription control.
This gene, encoding the RNA polymerase b-subunit, is
present in all mycobacterial species [33]. DNAse-Q-treated
M. tuberculosis H37Rv was used as cDNA synthesis nega-
tive control. Distilled water and M. tuberculosis H37Rv
DNA were used as negative and positive PCR controls,
respectively.
Rabbit immunization
Two New Zealand strain rabbits per peptide (previously
determined to be nonreactive to M. tuberculosis sonicate, as
assessed by western blot) were injected with 500 lg of
peptide 11027 (181ADGKPVTIATFFRPRNLGLV200) or
500 lg of peptide 11031 (242SPIDSVKASIETVGSNIG
GSVY262) [polymerized via the addition of cysteine at the
amino and carboxy termini and mixed with Freund’s
Incomplete Adjuvant (Sigma)] on days 0, 20 and 40. Final
bleeding was carried out on day 60 and sera were collected
in accordance with the Fundacion Instituto de Inmunolo-
gia’s Bioethics Committee recommendations. The Rv2560
protein peptide sequences chosen for the immunization of
rabbits were obtained using T-epitope prediction software
M. tuberculosis Rv2560 protein characterization D. F. Plaza et al.
6360 FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS

syfpeithi (15-mer T-epitope for major histocompatibility
complex class II only) downloaded from http://www.
syfpeithi.de/Scripts/MHCServer.dLl/EpitopePrediction.htm.
SDS-PAGE and immunoblotting
M. tuberculosis sonicate or membrane fraction (1 mg) was
loaded in a discontinuous SDS-PAGE system, using a 10–
20% (w ⁄ v) acrylamide gradient, and then transferred to
nitrocellulose paper using the semidry blotting technique
[34]. The filters were incubated with a 1 : 100 (v ⁄ v) dilutionof the sera obtained from rabbits immunized with polymer-
ized 11027 or 11031 peptides. Sera were diluted in NaCl ⁄Tris ⁄Tween 20 (0.02 m Tris-HCl, pH 7.5, 0.05 m NaCl, 1%
Tween 20) and 5% skimmed milk. Incubation for 1 h with
1 : 3000 (v ⁄ v) alkaline phosphatase-conjugated anti-rabbit
IgG (ICN Biomedicals, Costa Mesa, CA, USA) followed
five NaCl ⁄Tris ⁄Tween 20 washes. The reaction was devel-
oped with Nitro Blue tetrazolium ⁄ 5-bromo-4-chloroindol-
2-yl phosphate (KPL, Gaithersburg, MD, USA).
Immunoelectron microscopy
Immunoelectron microscopy studies were carried out on a
Philips CM 10 transmission electron microscope (Philips,
Suresne, Hauts-de-Seine, France). Briefly, a wet pellet
(50 lL) of M. tuberculosis H37Rv was fixed with a 4%
paraformaldehyde)0.5% glutaraldehyde solution for 2 h at
4 �C. After fixation, the pellet was dehydrated in graded
ethanol and then embedded in LR-white hard-grade acrylic
resin (Sigma) for 4 days at 4 �C. Thin slices (400 nm) were
cut and mounted on 300 mesh nickel grids. The slices were
incubated in a saturated sodium metaperiodate solution for
1 h at 20–22 �C for antigen retrieval [35]. The grids were
then floated, section down, in a beaker containing 0.01 m
sodium citrate buffer for 15 min at 80 �C. After 1 h of
blocking in Tris-buffered saline (NaCl ⁄Tris) (0.05 m Tris in
isotonic saline, pH 7.6) containing 0.05% BSA, the grids
were incubated in either 1 : 10 (v ⁄ v) or pure rabbit
polyclonal serum for 1 h at 37 �C. Following an NaCl ⁄Tris ⁄ 0.025% Tween 20 wash, the grids were immersed in a
1 : 50 (v ⁄ v) dilution of 5 nm gold-labelled anti-rabbit IgG
(Sigma) for 1 h at 20–22 �C. The grids were then washed
with NaCl ⁄Tris ⁄Tween 20 and fixed in 2.5% glutaralde-
hyde, followed by 15 min of incubation in 1% uranyl ace-
tate. The grids were washed with distilled water and dried
at 20–22 �C before observation.
Cell culture
A549 human lung epithelial (ATCC CCL-185) and U937
monocyte-like (ATCC CRL-1593.2) cells were grown at
37 �C and 5% CO2 in RPMI 1640 medium (Gibco-BRL)
supplemented with 10% fetal bovine serum (Hyclone,
Logon, UT, USA). A549 cells were dislodged with 0.1%
EDTA–NaCl ⁄Pi. Cell lines were collected in 50 mL tubes,
centrifuged at 1000 g for 5 min and washed with RPMI
1640.
Peptide synthesis
Sixteen sequential 20-mer peptides, corresponding to the
Rv2560 amino acid sequence [9], were synthesized in this
study by a solid-phase multiple peptide system [36,37].
4-Methylbenzhydrylamine resin (0.7 meqÆg)1), t-Boc amino
acids and low–high cleavage techniques were used [38]. Pep-
tide identity and purity were analysed by MALDI-TOF MS
and analytical reverse phase HPLC. An extra tyrosine resi-
due was added to any peptide C-terminus that did not
contain it to enable radiolabelling.
Peptide radiolabelling
Radiolabelling with 125I was performed according to previ-
ously described techniques [24,25,39,40], in which chlor-
amine-T (2.25 mgÆmL)1) and 3.2 lL Na125I (100 mCiÆmL)1)
were added to 5 lL peptide solution (1 lgÆlL)1); 15 lLsodium bisulfite (2.75 mgÆmL)1) and 50 lL NaI (0.16 m)
were added after 5 min of reaction at 18 �C. The radiola-
belled peptide was then separated from reaction by-prod-
ucts on a Sephadex G-10 column (Pharmacia Biotech,
St Albans, UK) (80 · 5.0 mm).
Binding assay
A549 and U937 cells (1 · 106) were incubated with increas-
ing quantities of each Rv2560 m. tuberculosis putative mem-
brane protein 125I-radiolabelled peptide (100–2000 nm) at
100 lL total volume for 90 min at 4 �C, in the presence or
absence of 40 lm unlabelled peptide, to determine the bind-
ing specificity. After incubation, unbound peptide was
removed from the cells by sedimentation through a dioctyl-
phthalate–dibutylphthalate cushion (d ¼ 1.015 gÆmL)1) and
centrifuged at 9000 g for 2 min [18,19]. The assay was car-
ried out in triplicate in identical conditions; bound and free
peptides were determined by measuring the cell-associated
radioactivity on a gamma counter (Gamma Counter
Cobra II, Packard Instrument Co., Meriden, CT, USA);
the average results of the assays performed in triplicate are
reported and shown graphically in Fig. 4.
Saturation assay
A549 and U937 cells (1 · 106) were incubated with increas-
ing concentrations (100–2000 nm) of radiolabelled peptide
at 120 lL total volume for 90 min at 4 �C, in the presence
or absence of 40 lm unlabelled peptide, to determine the
binding specificity. After incubation, unbound peptide was
D. F. Plaza et al. M. tuberculosis Rv2560 protein characterization
FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS 6361

removed from the cells by sedimentation through a dioctyl-
phthalate–dibutylphthalate cushion (d ¼ 1.015 gÆmL)1) and
centrifuged at 9000 g for 2 min. As before, each assay was
performed in triplicate; bound and free peptides were deter-
mined by measuring the cell-associated radioactivity on a
gamma counter. Triplicate assay data were averaged. The
curves obtained were analysed and the dissociation con-
stants were determined by the Hill equation [18,19].
Analogue peptide competition binding assay
HABP 11024 glycine scanning analogues were synthesized
to identify critical residues for binding to U937 cells. For
the competition binding assays, 1 · 106 U937 cells were
incubated with increasing quantities (16 and 400 nm) of
each unlabelled analogue peptide or original unlabelled
peptide in the presence of native 125I-labelled peptide. After
90 min of incubation at 4 �C, the mixture was centrifuged
at 9000 g for 2 min on a dioctylphthalate–dibutylphthalate
cushion (d ¼ 1.015 gÆmL)1). Radioactivity bound to
U937 cells was determined by gamma counter measurement
[17].
Cross-linking assays
U937 cells (2 · 106) were subjected to a conventional bind-
ing assay (90 min of incubation at 4 �C) with HABP 11024.
Following incubation, the cells were washed with NaCl ⁄Pi
and cross-linked with 25 lm bis(sulfosuccinymidyl suberate)
(Pierce, Rockford, IL, USA) for 20 min at 4 �C. The reac-
tion was stopped with 40 nm Tris ⁄HCl (pH 7.4), and washed
again with NaCl ⁄Pi. The cells were then treated with lysis
buffer (5% SDS, 10 nm iodoacetamide, 1% Triton X-100,
100 mm EDTA, 10 mm phenylmethanesulfonyl fluoride).
The membrane proteins so obtained were solubilized in
Laemmli buffer and separated by SDS-PAGE. Proteins
cross-linked with radiolabelled peptides were exposed on a
Bio-Rad Imaging Screen K (Bio-Rad Molecular Imager FX;
Bio-Rad Quantity One Quantitation Software; Bio-Rad,
Hercules, CA, USA) for 7 days.
Mycobacterium culture and staining
for invasion assay
M. tuberculosis H37Rv bacilli were collected during the log-
arithmic phase, washed and suspended in NaCl ⁄Pi. The sus-
pension was briefly sonicated (30 W for 5 s) on a Branson
sonifier (VWR Scientific, Boston, MA, USA); the suspen-
sion turbidity was adjusted to 3 · 108 colony-forming
unitsÆmL)1 according to McFarland’s pattern. M. tuber-
culosis was labelled with SYBR Safe (Invitrogen, Eugene,
OR, USA) at 20 · final concentration at 37 �C for 20 min
in the dark with constant shaking, washed twice and centri-
fuged at 12 000 g for 20 min; excess dye was removed.
Bacteria were then suspended in RPMI 1640, and labelling
was further verified by flow cytometry and fluorescence
microscopy.
Invasion inhibition assay with HABPs
The possible biological relevance of the HABPs identified
in Rv2560 was evaluated by an assay developed by our
group [41]. U937 or A549 cells (1 · 106) suspended in com-
plete RPMI 1640 medium without antibiotics were preincu-
bated for 1 h in the presence of 50, 100 or 200 lm
HABP 11024 for U937 cells and 100)105 nm or 200 lm
HABP 11024 for A549 cells before the addition of 1 · 107
M. tuberculosis H37Rv bacilli (multiplicity of infection,
1 : 10) at a final volume of 200 lL. They were incubated at
37 �C in 5% CO2 for 2 h with constant shaking, and then
placed in 20 lgÆmL)1 amikacin-supplemented medium (ICN
Biomedicals) for 30 min to kill off the extracellular bacte-
ria, whilst the intracellular bacteria remained viable.
Two RPMI 1640 washes preceded centrifugation at 500 g
for 5 min. The cells were fixed in 1% paraformaldehyde
(Merck, Darmstadt, Germany) in RPMI 1640 at 4 �C for
1 h, washed and suspended in 50 lL RPMI 1640. The cells
were counterstained with freshly prepared 3% methylene
blue in 30% ethanol, 5 min before flow cytometry, and
incubated at 20–22 �C. The cells used to determine the
autofluorescence control were treated using the same experi-
mental conditions as those employed for the cells incubated
with bacteria. Invasion control was carried out in the pres-
ence of 30 lm colchicine (Sigma) as inhibitor, added to the
cells for 1 h at 37 �C prior to the addition of bacilli to inhi-
bit cell internalization. The inhibitors were maintained
throughout the 2 h invasion period. Student’s t-test was
used to verify possible differences between peptide treat-
ment and cell control, which had not been preincubated
with peptides or colchicine [20,26].
Flow cytometry quantification
Samples were analysed on a FACScan (Becton Dickinson)
flow cytometer. CellQuest software (Becton Dickinson) was
used for flow cytometry capture and analysis. NaCl ⁄Pi,
which had been filtered with a 0.2 lm membrane, was used
as sheath fluid. Uninfected cells were discriminated from
infected cells on the basis of light characteristics on fluores-
cence channel one. Samples were run at 2000 events per sec-
ond; 40 000 events were collected. The percentage of positive
SYBR safe events in the gated region was determined.
CD spectroscopy
CD was carried out for HABP 11024 and control peptide
11021 to determine whether there were any conformational
changes when the peptide concentration in the solution
M. tuberculosis Rv2560 protein characterization D. F. Plaza et al.
6362 FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS

was increased. CD spectra were recorded at 20 �C on a
Jasco J-810 spectropolarimeter at wavelengths ranging
from 260 to 190 nm in 1 cm cuvettes (Jasco, Tokyo,
Japan). The peptides were dissolved at 5, 25, 50, 100 and
200 lm concentration in 30% trifluoroethanol aqueous
solution. Each spectrum was obtained from the average of
three scans taken at a scan rate of 20 nmÆmin)1 with a
spectral bandwidth of 1 nm, corrected for baseline. The
results were expressed as the mean residue ellipticity [Q].
Acknowledgements
We wish to thank Dr Ramses Lopez, Rosalba Alfonso,
Marisol Ocampo, Martha Calderon and Javier Rosas
who started this work whilst working at Fundacion
Instituto de Immulogia de Colombia (FIDIC). This
work was supported by COLCIENCIAS contract no.
2008. The wholehearted assistance of Gloria P. Barrera
in immunoelectron microscopy, Marı́a H. Duran in
CD experiments and Jason Garry in translating the
manuscript is greatly appreciated.
References
1 World Health Organization (WHO) (2006) World
Health Report 2006. Global Tuberculosis Control: Sur-
veillance, Planning, Financing. WHO, Geneva.
2 Russell DG (2007) Who puts the tubercle in tuberculo-
sis? Nat Rev Microbiol 5, 39–47.
3 Snider DE Jr & Castro KG (1998) The global threat of
drug-resistant tuberculosis. N Engl J Med 338, 1689–
1690.
4 Pillay CS, Elliott E & Dennison C (2002) Endolysoso-
mal proteolysis and its regulation. Biochem J 363, 417–
429.
5 Pieters J (2001) Entry and survival of pathogenic
mycobacteria in macrophages. Microbes Infect 3,
249–255.
6 Bermudez LE & Sangari FJ (2001) Cellular and mole-
cular mechanisms of internalization of mycobacteria by
host cells. Microbes Infect 3, 37–42.
7 El-Etr SH & Cirillo JD (2001) Entry mechanisms of
mycobacteria. Front Biosci 6, D737–D747.
8 Sigler K & Hofer M (1997) Biotechnological aspects of
membrane function. Crit Rev Biotechnol 17, 69–86.
9 Cole ST, Brosch R, Parkhill J, Garnier T, Churcher C,
Harris D, Gordon SV, Eiglmeier K, Gas S, Barry CE
3rd, et al. (1998) Deciphering the biology of Mycobacte-
rium tuberculosis from the complete genome sequence.
Nature 393, 537–544.
10 Garcia-Perez BE, Mondragon-Flores R & Luna-Herrera
J (2003) Internalization of Mycobacterium tuberculosis
by macropinocytosis in non-phagocytic cells. Microb
Pathog 35, 49–55.
11 Chakravarti DN, Fiske MJ, Fletcher LD & Zagursky
RJ (2000) Application of genomics and proteomics for
identification of bacterial gene products as potential
vaccine candidates. Vaccine 19, 601–612.
12 Gu S, Chen J, Dobos KM, Bradbury EM, Belisle JT &
Chen X (2003) Comprehensive proteomic profiling of
the membrane constituents of a Mycobacterium tubercu-
losis strain. Mol Cell Proteomics 2, 1284–1296.
13 Sinha S, Kosalai K, Arora S, Namane A, Sharma P,
Gaikwad AN, Brodin P & Cole ST (2005) Immunogenic
membrane-associated proteins of Mycobacterium tuber-
culosis revealed by proteomics. Microbiology 151, 2411–
2419.
14 Cardoso FC, Pinho JM, Azevedo V & Oliveira SC
(2006) Identification of a new Schistosoma mansoni
membrane-bound protein through bioinformatic analy-
sis. Genet Mol Res 5, 609–618.
15 Kyte J & Doolittle RF (1982) A simple method for dis-
playing the hydropathic character of a protein. J Mol
Biol 157, 105–132.
16 El-Etr SH, Subbian S, Cirillo SL & Cirillo JD (2004)
Identification of two Mycobacterium marinum loci that
affect interactions with macrophages. Infect Immun 72,
6902–6913.
17 Garcia JE, Puentes A, Suarez J, Lopez R, Vera R,
Rodriguez LE, Ocampo M, Curtidor H, Guzman F,
Urquiza M, et al. (2002) Hepatitis C virus (HCV) E1
and E2 protein regions that specifically bind to HepG2
cells. J Hepatol 36, 254–262.
18 Ocampo M, Urquiza M, Guzman F, Rodriguez LE,
Suarez J, Curtidor H, Rosas J, Diaz M & Patarroyo
ME (2000) Two MSA 2 peptides that bind to human
red blood cells are relevant to Plasmodium falciparum
merozoite invasion. J Pept Res 55, 216–223.
19 Ocampo M, Vera R, Eduardo Rodriguez L, Curtidor
H, Urquiza M, Suarez J, Garcia J, Puentes A, Lopez R,
Trujillo M, et al. (2002) Plasmodium vivax Duffy bind-
ing protein peptides specifically bind to reticulocytes.
Peptides 23, 13–22.
20 Danelishvili L, McGarvey J, Li YJ & Bermudez LE
(2003)Mycobacterium tuberculosis infection causes differ-
ent levels of apoptosis and necrosis in human macrophag-
es and alveolar epithelial cells. Cell Microbiol 5, 649–660.
21 Forero M, Puentes A, Cortes J, Castillo F, Vera R,
Rodriguez LE, Valbuena J, Ocampo M, Curtidor H,
Rosas J, et al. (2005) Identifying putative Mycobacte-
rium tuberculosis Rv2004c protein sequences that bind
specifically to U937 macrophages and A549 epithelial
cells. Protein Sci 14, 2767–2780.
22 Garcia J, Puentes A, Rodriguez L, Ocampo M, Curti-
dor H, Vera R, Lopez R, Valbuena J, Cortes J, Vanegas
M, et al. (2005) Mycobacterium tuberculosis Rv2536
protein implicated in specific binding to human cell
lines. Protein Sci 14, 2236–2245.
D. F. Plaza et al. M. tuberculosis Rv2560 protein characterization
FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS 6363

23 Vera-Bravo R, Torres E, Valbuena JJ, Ocampo M,
Rodriguez LE, Puentes A, Garcia JE, Curtidor H, Cor-
tes J, Vanegas M, et al. (2005) Characterising Mycobac-
terium tuberculosis Rv1510c protein and determining its
sequences that specifically bind to two target cell lines.
Biochem Biophys Res Commun 332, 771–781.
24 Hulme E (1993) Receptor–Ligand Interactions. A Practi-
cal Approach. IRL Press, Oxford.
25 Weiland GA & Molinoff PB (1981) Quantitative analy-
sis of drug–receptor interactions. I. Determination of
kinetic and equilibrium properties. Life Sci 29, 313–330.
26 Bermudez LE & Goodman J (1996) Mycobacterium
tuberculosis invades and replicates within type II alveo-
lar cells. Infect Immun 64, 1400–1406.
27 Reddy VM & Hayworth DA (2002) Interaction of
Mycobacterium tuberculosis with human respiratory epi-
thelial cells (HEp-2). Tuberculosis (Edinb) 82, 31–36.
28 Gagliardi MC, Lemassu A, Teloni R, Mariotti S, Sar-
gentini V, Pardini M, Daffe M & Nisini R (2007) Cell
wall-associated alpha-glucan is instrumental for Myco-
bacterium tuberculosis to block CD1 molecule expression
and disable the function of dendritic cell derived from
infected monocyte. Cell Microbiol 9, 2081–2092.
29 Subramanyam MB, Gnanamani M & Ramachandran S
(2006) Simple sequence proteins in prokaryotic proteo-
mes. BMC Genomics 7, 141.
30 Cheung KJ Jr, Li G, Urban TA, Goldberg JB, Griffith
A, Lu F & Burns JL (2007) Pilus-mediated epithelial cell
death in response to infection with Burkholderia ceno-
cepacia. Microbes Infect 9, 829–837.
31 Del Portillo P, Murillo LA & Patarroyo ME (1991)
Amplification of a species-specific DNA fragment of
Mycobacterium tuberculosis and its possible use in diag-
nosis. J Clin Microbiol 29, 2163–2168.
32 Katoch VM & Cox RA (1986) Step-wise isolation of
RNA and DNA from mycobacteria. Int J Lepr Other
Mycobact Dis 54, 409–415.
33 Lee H, Park HJ, Cho SN, Bai GH & Kim SJ (2000)
Species identification of mycobacteria by PCR-
restriction fragment length polymorphism of the rpoB
gene. J Clin Microbiol 38, 2966–2971.
34 Kyhse-Andersen J (1984) Electroblotting of multiple
gels: a simple apparatus without buffer tank for rapid
transfer of proteins from polyacrylamide to nitrocellu-
lose. J Biochem Biophys Methods 10, 203–209.
35 Stirling JW & Graff PS (1995) Antigen unmasking for
immunoelectron microscopy: labeling is improved by
treating with sodium ethoxide or sodium metaperiodate,
then heating on retrieval medium. J Histochem Cyto-
chem 43, 115–123.
36 Houghten RA (1985) General method for the rapid
solid-phase synthesis of large numbers of peptides: spec-
ificity of antigen–antibody interaction at the level of
individual amino acids. Proc Natl Acad Sci USA 82,
5131–5135.
37 Merrifield RB (1969) Solid-phase peptide synthesis. Adv
Enzymol Relat Areas Mol Biol 32, 221–296.
38 Tam JP, Heath WF & Merrifield RB (1983) SN 1 and SN
2 mechanisms for the deprotection of synthetic peptides
by hydrogen fluoride. Studies to minimize the tyrosine
alkylation side reaction. Int J Pept Protein Res 21, 57–65.
39 Vera Bravo R, Marin V, Garcia J, Urquiza M, Torres
E, Trujillo M, Rosas J & Patarroyo ME (2000) Amino
terminal peptides of the ring infected erythrocyte surface
antigen of Plasmodium falciparum bind specifically to
erythrocytes. Vaccine 18, 1289–1293.
40 Yamamura H (1978) Neurotransmitter Receptor Binding.
Raven Press, New York, NY.
41 Chapeton-Montes JA, Plaza DF, Barrero CA & Patar-
royo MA (2007) Quantitative flow cytometric monitor-
ing of invasion of epithelial cells by Mycobacterium
tuberculosis. Front Biosci 30, 650–656.
M. tuberculosis Rv2560 protein characterization D. F. Plaza et al.
6364 FEBS Journal 274 (2007) 6352–6364 ª 2007 The Authors Journal compilation ª 2007 FEBS




















