The levels of renin–angiotensin related components are modified in the hippocampus of rats...
9
1 2 The levels of renin–angiotensin related components are modified 3 in the hippocampus of rats submitted to pilocarpine model of epilepsy 4 Telma Luciana Furtado Gouveia a Q1 , Maria Isabel Berzaghi Frangiotti a , Joíse Marques Vieira de Brito a , 5 Eduardo Ferreira de Castro Neto a , Maisa Mayume Sakata c , Andreia Cristina Febba c , Dulce Elena Casarini c , 6 Débora Amado a , Esper Abrão Cavalheiro a , Sandro Soares Almeida e , Martha Trindade Manchini d , 7 Ronaldo Carvalho Araújo e , José Antonio Silva Jr. d , Maria da Graça Naffah-Mazzacoratti a,b,⇑ 8 a Neurology and Neurosurgery Department, Universidade Federal de São Paulo (UNIFESP), São Paulo, Brazil 9 b Biochemistry Department, Universidade Federal de São Paulo (UNIFESP), São Paulo, Brazil 10 c Medicine Department, Nephrology Laboratory, Universidade Federal de São Paulo (UNIFESP), São Paulo, Brazil 11 d Rehabilitation Department, Universidade Nove de Julho, São Paulo, Brazil 12 e Biophysical Department, Universidade Federal de São Paulo (UNIFESP), São Paulo, Brazil 13 14 16 article info 17 Article history: 18 Received 14 September 2011 19 Received in revised form 9 April 2012 20 Accepted 11 April 2012 21 Available online xxxx 22 Keywords: 23 Pilocarpine 24 Angiotensin I 25 Angiotensin II 26 Temporal lobe epilepsy 27 Angiotensin converting enzyme (ACE) 28 29 abstract 30 We previously showed patients with temporal lobe epilepsy (TLE) present an increased expression of 31 angiotensin II (AngII) AT1 and AT2 receptors in the hippocampus, supporting the idea of an upregulation 32 of renin–angiotensin system (RAS) in this disease. 33 This study aimed to verify the relationship between the RAS and TLE during epileptogenesis. Levels of 34 the peptides angiotensin I (AngI), angiotensin II (AngII) and angiotensin 1–7 Ang(1–7), were detected by 35 HPLC assay. Angiotensin AT1 and AT2 receptors, Mas mRNA receptors and angiotensin converting 36 enzyme (ACE), tonin and neutral endopeptidase (NEP) mRNA were also quantified at the hippocampus 37 of Wistar rats by real time PCR, during acute (n = 10), silent (n = 10) and chronic (n = 10) phases of pilo- 38 carpine-induced epilepsy. 39 We observed an increased peptide level of Ang1–7 into acute and silent phases, decreasing importantly 40 (p 6 0.05) in the chronic phase, suggesting that AngI may be converted into Ang(1–7) by NEP, which is 41 present in high levels in these periods. 42 Our results also showed increased peptide level of AngII in the chronic phase of this model. In contra- 43 position, the ACE expression is reduced in all periods. These data suggest that angiotensinogen or AngI 44 may be cleaved to AngII by tonin, which showed increased expression in all phases. We found changes 45 in AT1, AT2 and Mas mRNA receptors levels suggesting that Ang1–7 could act at Mas receptor during 46 the silent period. Herein, we demonstrated for the first time, changes in angiotensin-related peptides, 47 their receptors as well as the releasing enzymes in the hippocampus of rats during pilocarpine-induced 48 epilepsy. 49 Ó 2012 Published by Elsevier Ltd. 50 51 52 1. Introduction 53 Renin–angiotensin system (RAS) is originally known as a blood 54 pressure controller and its role in inflammation has been only re- 55 cently studied. This system is characterized by a precursor mole- 56 cule angiotensinogen, and through the renin enzyme is cleaved 57 to angiotensin I (AngI), which is a decapeptide with little or none 58 biological action. AngI can circulate in blood and tissues being 59 cleaved by a key enzyme called angiotensin converting enzyme 60 (ACE), which removes the dipeptide His-Leu from C-terminal side 61 of the molecule, transforming AngI into AngII, which has an impor- 62 tant biological action (review in de Gasparo et al. (2000)). 63 Although widely distributed in peripheral systems, all RAS com- 64 ponents have also been found in the brain where they actively 65 modulate several functions, including exploratory behavior, stress, 66 anxiety, learning and memory acquisition (Chen et al., 1993; Meff- 67 ert et al., 1996; Saavedra et al., 2005; Wright and Harding, 2004; 68 Yamada et al., 1996). 69 AngII has been considered to be the major RAS active peptide, 70 acting in the periphery as paracrine and endocrine hormone. This 71 polypeptide could also be considered a neurotransmitter/neuro- 72 modulator through its action on AT1 and AT2 receptors present 73 in several brain regions (Wright et al., 2002). 0197-0186/$ - see front matter Ó 2012 Published by Elsevier Ltd. http://dx.doi.org/10.1016/j.neuint.2012.04.012 ⇑ Corresponding author at: Biochemistry Department, Universidade Federal de São Paulo (UNIFESP), São Paulo, Brazil. Tel.: +55 11 55492064; fax: +55 11 55494743. E-mail address: [email protected] (M.d.G. Naffah-Mazzacoratti). Neurochemistry International xxx (2012) xxx–xxx Contents lists available at SciVerse ScienceDirect Neurochemistry International journal homepage: www.elsevier.com/locate/nci NCI 3140 No. of Pages 9, Model 5G 25 April 2012 Please cite this article in press as: Gouveia, T.L.F., et al. The levels of renin–angiotensin related components are modified in the hippocampus of rats sub- mitted to pilocarpine model of epilepsy. Neurochem. Int. (2012), http://dx.doi.org/10.1016/j.neuint.2012.04.012
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The levels of renin–angiotensin related components are modified in the hippocampus of rats...

1
2
3
4 Q1
5
6
7
89
101112
1314
1 6
1718192021
22232425262728
2 9
51
52
53
54
55
56
57
58
59
Neurochemistry International xxx (2012) xxx–xxx
NCI 3140 No. of Pages 9, Model 5G
25 April 2012
Contents lists available at SciVerse ScienceDirect
Neurochemistry International
journal homepage: www.elsevier .com/locate /nci
The levels of renin–angiotensin related components are modifiedin the hippocampus of rats submitted to pilocarpine model of epilepsy
Telma Luciana Furtado Gouveia a, Maria Isabel Berzaghi Frangiotti a, Joíse Marques Vieira de Brito a,Eduardo Ferreira de Castro Neto a, Maisa Mayume Sakata c, Andreia Cristina Febba c, Dulce Elena Casarini c,Débora Amado a, Esper Abrão Cavalheiro a, Sandro Soares Almeida e, Martha Trindade Manchini d,Ronaldo Carvalho Araújo e, José Antonio Silva Jr. d, Maria da Graça Naffah-Mazzacoratti a,b,⇑a Neurology and Neurosurgery Department, Universidade Federal de São Paulo (UNIFESP), São Paulo, Brazilb Biochemistry Department, Universidade Federal de São Paulo (UNIFESP), São Paulo, Brazilc Medicine Department, Nephrology Laboratory, Universidade Federal de São Paulo (UNIFESP), São Paulo, Brazild Rehabilitation Department, Universidade Nove de Julho, São Paulo, Brazile Biophysical Department, Universidade Federal de São Paulo (UNIFESP), São Paulo, Brazil
a r t i c l e i n f o
303132333435363738394041
Article history:Received 14 September 2011Received in revised form 9 April 2012Accepted 11 April 2012Available online xxxx
Keywords:PilocarpineAngiotensin IAngiotensin IITemporal lobe epilepsyAngiotensin converting enzyme (ACE)
4243444546474849
0197-0186/$ - see front matter � 2012 Published byhttp://dx.doi.org/10.1016/j.neuint.2012.04.012
⇑ Corresponding author at: Biochemistry DepartmSão Paulo (UNIFESP), São Paulo, Brazil. Tel.: +5555494743.
E-mail address: [email protected] (M.d.G.
Please cite this article in press as: Gouveia, T.L.Fmitted to pilocarpine model of epilepsy. Neuro
a b s t r a c t
We previously showed patients with temporal lobe epilepsy (TLE) present an increased expression ofangiotensin II (AngII) AT1 and AT2 receptors in the hippocampus, supporting the idea of an upregulationof renin–angiotensin system (RAS) in this disease.
This study aimed to verify the relationship between the RAS and TLE during epileptogenesis. Levels ofthe peptides angiotensin I (AngI), angiotensin II (AngII) and angiotensin 1–7 Ang(1–7), were detected byHPLC assay. Angiotensin AT1 and AT2 receptors, Mas mRNA receptors and angiotensin convertingenzyme (ACE), tonin and neutral endopeptidase (NEP) mRNA were also quantified at the hippocampusof Wistar rats by real time PCR, during acute (n = 10), silent (n = 10) and chronic (n = 10) phases of pilo-carpine-induced epilepsy.
We observed an increased peptide level of Ang1–7 into acute and silent phases, decreasing importantly(p 6 0.05) in the chronic phase, suggesting that AngI may be converted into Ang(1–7) by NEP, which ispresent in high levels in these periods.
Our results also showed increased peptide level of AngII in the chronic phase of this model. In contra-position, the ACE expression is reduced in all periods. These data suggest that angiotensinogen or AngImay be cleaved to AngII by tonin, which showed increased expression in all phases. We found changesin AT1, AT2 and Mas mRNA receptors levels suggesting that Ang1–7 could act at Mas receptor duringthe silent period. Herein, we demonstrated for the first time, changes in angiotensin-related peptides,their receptors as well as the releasing enzymes in the hippocampus of rats during pilocarpine-inducedepilepsy.
� 2012 Published by Elsevier Ltd.
50
60
61
62
63
64
65
66
67
68
1. Introduction
Renin–angiotensin system (RAS) is originally known as a bloodpressure controller and its role in inflammation has been only re-cently studied. This system is characterized by a precursor mole-cule angiotensinogen, and through the renin enzyme is cleavedto angiotensin I (AngI), which is a decapeptide with little or nonebiological action. AngI can circulate in blood and tissues beingcleaved by a key enzyme called angiotensin converting enzyme
69
70
71
72
73
Elsevier Ltd.
ent, Universidade Federal de11 55492064; fax: +55 11
Naffah-Mazzacoratti).
., et al. The levels of renin–angchem. Int. (2012), http://dx.doi
(ACE), which removes the dipeptide His-Leu from C-terminal sideof the molecule, transforming AngI into AngII, which has an impor-tant biological action (review in de Gasparo et al. (2000)).
Although widely distributed in peripheral systems, all RAS com-ponents have also been found in the brain where they activelymodulate several functions, including exploratory behavior, stress,anxiety, learning and memory acquisition (Chen et al., 1993; Meff-ert et al., 1996; Saavedra et al., 2005; Wright and Harding, 2004;Yamada et al., 1996).
AngII has been considered to be the major RAS active peptide,acting in the periphery as paracrine and endocrine hormone. Thispolypeptide could also be considered a neurotransmitter/neuro-modulator through its action on AT1 and AT2 receptors presentin several brain regions (Wright et al., 2002).
iotensin related components are modified in the hippocampus of rats sub-.org/10.1016/j.neuint.2012.04.012

74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
2 T.L.F. Gouveia et al. / Neurochemistry International xxx (2012) xxx–xxx
NCI 3140 No. of Pages 9, Model 5G
25 April 2012
It is already known that AngII promotes inflammatory responseinducing oxidative stress, NADPH oxidase and NF-jb activationand consequently production of several cytokines and adhesionmolecules (Benicky et al., 2011). In addition, many experimentalstudies suggest that NF-jb and cytokines participate in the neuro-nal excitability and/or in glial scar formation in epilepsy, showing arelationship between this chronic disease and inflammatoryresponse (Ferder et al., 2003, 2006). In addition, evidences haveemerged indicating RAS participation in some neurodegenerativedisorders such as Alzheimer (Barnes et al., 1991; Tian et al.,2004), Parkinson (Grammatopoulos et al., 2005) and Huntington(Bird, 1980) diseases.
In epilepsy, clinical and experimental findings showed impor-tant role of inflammatory processes linked to this disease (Argana-raz et al., 2004a,b; Vezzani and Granata, 2005), particularly in themechanisms underlying generation of seizures (ictogenesis) andtransformation of a normal neuronal network into a seizure-gener-ating one (Pitkanen and Sutula, 2002). Recently, we observed an in-creased expression of several inflammatory markers such as kininreceptors, cytokines and nitric oxide in limbic areas of subjectswith TLE (Perosa et al., 2007; Silva et al., 2008; Gouveia et al.,2011; Varella et al., 2011). Our group also showed an up-regulationof AT1 and AT2 receptors in the cortex and in the hippocampal for-mation of patients with TLE with mesial temporal sclerosis (MS)(Arganaraz et al., 2008), supporting RAS0 involvement on thispathology.
AngII may be generated in the brain via the classical pathway,through renin and ACE action or can be directly released from angi-otensinogen by cathepsin G or tonin actions (Lippoldt et al., 1995).According to Araujo et al. (2002), tonin may be an alternative path-way to AngII generation in the brain and could participate in thispeptide’s effects.
Recently it is become evident that another peptide namedAngiotensin (1–7) Ang(1–7) is active in several organs includingthe brain (Alenina et al., 2008), and several endopeptidases suchas neprilysin, thimet and neutral endopeptidase (NEP) may metab-olize the AngII in the brain generating Ang(1–7) (Karamyan andSpeth, 2007).
Mas is the main receptor for Ang(1–7) peptide being a physio-logical antagonist of AT1 receptor (Santos et al., 2003; Kosteniset al., 2005), which is also related to neuronal regulation. Mas-deficient mice exhibit increased anxiety; despite of an enhancedlong-term potentiation in the hippocampus. Furthermore, briefseizure episodes led to a significant and transient increase in MasmRNA in rat hippocampus (Martin and Hockfield, 1993).
Several animal models have been used to study TLE, such as thoseinduced by kainic acid (Ben-Ari et al., 1981), kindling (McNamara,1994) and pilocarpine (Turski et al., 1983) as they show similarityto human phenomena. Pilocarpine administered acutely to rats/ormice produces electrographic and behavioral limbic seizures andstatus epilepticus that last up to 12 h (acute phase). The statusepilepticus is accompanied by widespread brain damage that ismainly observed in limbic areas. This acute period is followed bysilent (seizure-free) period and finally by a chronic period character-ized by spontaneous recurrent seizures (Cavalheiro et al., 1991).
Then, this study was designed to contribute for better under-standing of the relationship between the RAS and TLE during epi-leptogenesis process, without anti-epileptic drugs interference.
188
189
190
191
192
193
194
195
2. Methods
2.1. Animals and SE induction
The experiments were performed under the approval of UNI-FESP Ethical Committee Institutional ethical approval of protocol
Please cite this article in press as: Gouveia, T.L.F., et al. The levels of renin–angmitted to pilocarpine model of epilepsy. Neurochem. Int. (2012), http://dx.doi
and all efforts were made to minimize animal suffering. Wistaradult male rats, weighing 250 g, were housed in groups of threeor four per cage and maintained in controlled room temperature,humidity and light–dark cycle (12:12 h) with chow pellets andtap water available ad libitum.
The rats received a single dose of pilocarpine [350 mg/kg,intraperitoneal (i.p.)]. To prevent peripheral cholinergic effects,scopolamine methylnitrate was injected subcutaneously at adose of 1 mg/kg, 30 min before pilocarpine administration. Onlyrats that displayed status epilepticus (SE) were included in thisstudy.
The SE was monitored and terminated with an injection of diaz-epam (10 mg/kg, ip; Roche, Brazil) 5 h after SE onset. The silent andchronic periods were followed by video-recordings of the animals24 h/day during 60 days. The animals were killed by decapitation;the brain was quickly removed and stored at �80 �C until assay.
Four groups of rats were analyzed: Control group (saline); Acutegroup (rats killed 5 h after SE onset); Silent group (animals killed5 days after pilocarpine injection) and Chronic group (rats killedduring the period of spontaneous seizures, 60 days after pilocar-pine injection). A group of animals, which received pilocarpinebut did not develop SE was initially used as a second control. Asthey did not present statistical difference when compared to con-trol group, they were excluded in this study. There were 10 ani-mals in each group.
2.2. Quantification of angiotensin-related peptides in the hippocampus
Quantification of AngI, AngII and Ang(1–7) were done accordingto assay previously described by Ronchi et al. (2007). Briefly,dissected hippocampi were weighed and homogenized with100 mM phosphate buffer, pH 7.2, containing 340 nM,sucrose, 300 mM NaCl and a mix of proteases inhibitors ethylene-diaminetetraacetic (EFTA) tripotassium salt dehydrate (25 mM),o-phenanthroline (0.44 mM), pepstatin A (0.12 mM) and 4-chloro-mercuribenzoic acid (1 mM). Twenty milliliters of solvent wasadded to 1 g of tissue, followed by centrifugation at 15,000g for20 min. The samples were concentrated in C18 Sep-Pak columnsactivated with methanol (5 mL), tetrahydrofuran (5 mL), hexane(5 mL), methanol (5 mL) and water (10 mL). Peptides were elutedwith ethanol, acetic acid and water (90:4:6).
Samples were then lyophilized and resuspended in 500 lL ofmobile phase A: 5% acetonitrile (50 mL) in 0.1% orthophosphoricacid (1 mL). Peptides were separated in a reverse-phase columnAquapore ODS 300 (250 � 4.6 mm), 7l, using the gradient 5–35%of mobile phase B: 95% acetonitrile in H3PO4 0.1% in the flow1.5 mL/min for 40 min in the Milton Roy System, constituted oftwo constaMetric 3000 pumps, a UV detector spectroMonitor3100, a programmer GM 4000 and a mixer. Synthetic standardswere used and peptide detection was carried out at 214 nm. Theresults were expressed in ng/mL and corrected by the weight ofthe tissue.
2.3. Quantitative real-time PCR for RAS molecules
Dissected hippocampi were frozen in liquid nitrogen and storedat �80 �C. Thawed tissue was homogenized in 1 mL TRIzol reagent(Invitrogen, Carlsbad, CA, USA) and total RNA was isolated accord-ing to the manufacturer’s instructions. RNA was subjected toDNase I digestion, followed by reverse transcription to cDNA andPCR was performed in a 7000 Sequence Detection System (ABIPrism, Applied Biosystems, Foster City, CA) using the SYBRGreencore reaction kit (Applied Biosystems).
Primers used for mRNA quantification were:
iotensin related components are modified in the hippocampus of rats sub-.org/10.1016/j.neuint.2012.04.012

197198
199
200
201202
203
204
205206
207
208
209210
211
212
213214
215
216
217
218219
220
221
222223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
T.L.F. Gouveia et al. / Neurochemistry International xxx (2012) xxx–xxx 3
NCI 3140 No. of Pages 9, Model 5G
25 April 2012
Pm
AT1 (
263
264
lease citeitted to p
GenBank™ Accession No. NM_031009.2)forward primer: 5́-ATGCCAGTGTGTTTCTGCTC-3́reverse primer: 5́-AACTCAACACTCCCCATTGG-3́
265
AT2 (266
267
GenBank™ Accession No. NM012494.3)forward primer: 5́-CAGTGGTCTGCTGGGATTGC-3́reverse primer: 5́-CCATCCAGGTCAGAGCATCC-3́
268
Mas (269
270
GenBank™ Accession No. NM012757.2)forward primer: 5́-ACTGCCGGGCGGTCATCATC-3́reverse primer: 5́-GGTGGAGAAAAGCAAGGAGA-3́
271
NEP (272
273
GenBank Accession No. NM 18272.1)forward primer: 5́-GGATCTTGTAAGCAGCCTCAGC-3́
reverse primer: 5́-AGTTGGCACACCGTCTCCAG-3́
274
Tonin (275
GenBank Accession No. NM 012677.1)forward primer: 5́-CCCTAGTCCTGTCCGTGGGACGAAT-3́reverse primer: 5́-CTTACTACCATCTCAGAGGGGTTG-30
276
ACE (277
GenBank Accession No. NM 001012006.1)forward primer: 5́-GCAGTACAAAGACTTGCCTG-3́reverse primer: 5́-TGGCAGAGGCTGACATGTTA-30
278
GAPDH ( 279280
GenBank Accession No. NM 017008.3)forward primer: 50-GGGCAGCCCAGAACATCAT-30
reverse primer: 50-CCGTTCAGCTCTGGGATGAC-30
281282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
Quantitative values for AT1, AT2, Mas, NEP, Tonin and ACEmRNA transcription were obtained from the threshold cycle num-ber, where the increased signal associated with an exponentialgrowth of PCR products begins to be detected. Melting curves weregenerated at the end of every run to ensure product uniformity.The relative target gene expression level was normalized on thebasis of GAPDH expression as endogenous RNA control.
DCt values of the samples were determined by subtracting theaverage Ct value mRNA from the average Ct value of the internalcontrol GAPDH. As it is uncommon to use DCt as a relative datadue to this logarithmic characteristic, the 2�DCt parameter was usedto express the relative expression data.
2.4. Statistical analysis
Concentration of peptides and mRNA expression levels was pre-sented as an n-fold difference relative to the levels expressed andthe different values were compared using the one-way ANOVA fol-lowed by Tukey–Kramer t-test. The data are presented asmean ± SD and p 6 0.05 was considered to be statisticallysignificant.
3. Results
3.1. Animal behavior
Pilocarpine administration induced the following behavioralchanges: akinesia, facial automatisms and limbic seizures consist-ing of forelimb clonus with rearing, salivation, and masticatory jawmovements and falling.
This type of behavior built up progressively into motor limbicseizures that recurred repeatedly evolving to long-lasting SE. Afteracute phase, the rats presented normal behavior, which character-ized the seizure-free period (silent phase), which was recorded andno seizure was detected, at least 24 h before sacrifice. Chronicphase only started with first spontaneous seizure occurrence.
3.2. Quantification of peptides in hippocampus
Decreased levels of hippocampal AngI was observed duringacute, silent and chronic phase (Fig. 1A). The concentration of AngII
this article in press as: Gouveia, T.L.F., et al. The levels of renin–angilocarpine model of epilepsy. Neurochem. Int. (2012), http://dx.doi
was very low in control, acute and silent periods, however, it in-creased dramatically into the chronic phase (p 6 0.001) (Fig. 1B).In addition, the levels of Ang(1–7) increased into acute and silentphases, decreasing importantly into the chronic phase (p 6 0.001)(Fig. 1C).
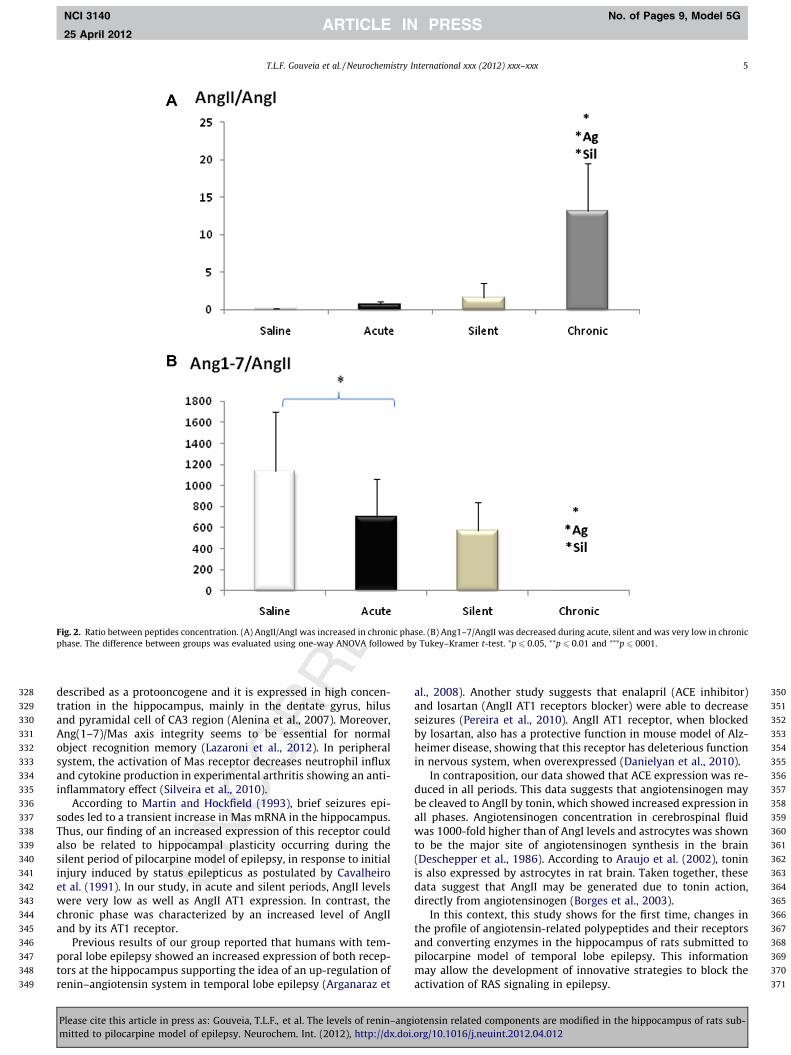
The ratio between AngII/AngI and Ang(1–7)/AngII were alsoanalyzed to verify the conversion rate between these peptides.The data showed increased ratio of AngII/AngI (Fig. 2A) into thechronic phase (p 6 0.05). In contrast, the conversion of AngII toAng(1–7) was decreased during acute (p 6 0.05), silent (p 6 0.05)and even lower at chronic phase (p 6 0.01) (Fig. 2B), in the hippo-campus of rats submitted to pilocarpine-induced model of epi-lepsy. The ratio between peptides amount was used to visualizethe main peptide that remain in the hippocampus during all phasesof this model.
3.3. Expression analysis of RAS -related molecules
The mRNA expression levels of AngII AT1 receptors showed adecreased expression in acute (p 6 0.01) and silent (p 6 0.001)groups (Fig. 3A). In chronic group, an increased expression of AngIIAT1 was observed when compared with control group (p 6 0.001).
AngII AT2 receptors mRNA expression was increased in acute(p 6 0.001) and silent (p 6 0.001) groups, showing no alterationinto chronic group (Fig. 3B). Mas mRNA expression was signifi-cantly increased only into the silent period (p 6 0.001) (Fig. 3C).
ACE mRNA levels were decreased in acute (p 6 0.001), silent(p 6 0.001) and chronic (p 6 0.001) phases as shown on Fig. 4A,while NEP mRNA was increased only into acute (p 6 0.05) and si-lent period (p 6 0.05) (Fig. 4B). Tonin mRNA levels was increasedin all periods (Fig. 4C). The Fig. 5 summarizes the main foundresults of comparison between pilocarpine injected group (acute,silent and chronic groups) with control group in several steps ofAngII formation and their related molecules. mRNA expression lev-els of endogenous RNA control (GAPDH) did not change in allgroups during all phases in pilocarpine model of epilepsy.
4. Discussion
Our results showed an increased concentration of AngII in thehippocampus of rats in the chronic phase of pilocarpine epilepsymodel. Usually, angiotensin II (AngII) is the major point of interestof investigations about RAS role on the central nervous system(CNS). This peptide is related to synaptic plasticity since it blockslong-term potentiation (LTP) in the hippocampus (Denny et al.,1991; Armstrong et al., 1996). Moreover, AngII reduces NMDAreceptors signaling through AT2 (angiotensin II receptor type 2)receptors-mediated mechanisms (Schelman et al., 1997, 2004; Jinget al., 2004).
Several other angiotensin-related peptides are recognized asbeing biologically active in brain structures and, among them,Ang(1–7), which is thought to counterbalance most of the AngII ef-fects (Daemen et al., 1991; Strawn et al., 1999; Tallant et al., 2005).In fact, in acute phase, we observed a low quantity of AngII, withlow expression of ACE but an increased amount of Ang(1–7),accompanied by high expression levels of NEP. In chronic phase,the ratio between AngII/AngI was increased, showing a predomi-nant form of AngII. Differently, the ratio between Ang(1–7)/AngIIwas decreased into the chronic phase, demonstrating that the mainpeptide in the hippocampus really is Ang(1–7) at this time.
Previous report suggested an anti-inflammatory action ofAng(1–7) in cardiovascular diseases (Ferrario, 2006). This peptidesignificantly attenuates the inflammatory action of AngII, such asinduced reactive oxygen species generation, c-SRC and ERK1/2activation and other molecules related to the death of endothelial
iotensin related components are modified in the hippocampus of rats sub-.org/10.1016/j.neuint.2012.04.012

322
323
324
325Q2
326
327
Fig. 1. Hippocampal peptides concentration: (A) AngI concentration was decreased during acute, silent and chronic phase of this epilepsy model. (B) AngII concentration wasvery low in control, acute and silent periods, increasing dramatically into the chronic phase. (C) Ang1–7 concentration was increased during acute and silent phases,decreasing importantly in chronic phase.
4 T.L.F. Gouveia et al. / Neurochemistry International xxx (2012) xxx–xxx
NCI 3140 No. of Pages 9, Model 5G
25 April 2012
cells (Sampaio et al., 2005). Previously reports have shown aninvolvement of inflammatory mediators in epilepsy (Naffah-Mazz-acoratti et al., 1995; Arganaraz et al., 2004a,b; Perosa, 2008; Gou-
Please cite this article in press as: Gouveia, T.L.F., et al. The levels of renin–angmitted to pilocarpine model of epilepsy. Neurochem. Int. (2012), http://dx.doi
veia et al., 2011). Thus, the release of Ang1–7 in tissues could belinked to neuroprotection as well as its main receptor (Mas) thatpresented increased expression. Mas receptor was initially
iotensin related components are modified in the hippocampus of rats sub-.org/10.1016/j.neuint.2012.04.012
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
371
Fig. 2. Ratio between peptides concentration. (A) AngII/AngI was increased in chronic phase. (B) Ang1–7/AngII was decreased during acute, silent and was very low in chronicphase. The difference between groups was evaluated using one-way ANOVA followed by Tukey–Kramer t-test. ⁄p 6 0.05, ⁄⁄p 6 0.01 and ⁄⁄⁄p 6 0001.
T.L.F. Gouveia et al. / Neurochemistry International xxx (2012) xxx–xxx 5
NCI 3140 No. of Pages 9, Model 5G
25 April 2012
described as a protooncogene and it is expressed in high concen-tration in the hippocampus, mainly in the dentate gyrus, hilusand pyramidal cell of CA3 region (Alenina et al., 2007). Moreover,Ang(1–7)/Mas axis integrity seems to be essential for normalobject recognition memory (Lazaroni et al., 2012). In peripheralsystem, the activation of Mas receptor decreases neutrophil influxand cytokine production in experimental arthritis showing an anti-inflammatory effect (Silveira et al., 2010).
According to Martin and Hockfield (1993), brief seizures epi-sodes led to a transient increase in Mas mRNA in the hippocampus.Thus, our finding of an increased expression of this receptor couldalso be related to hippocampal plasticity occurring during thesilent period of pilocarpine model of epilepsy, in response to initialinjury induced by status epilepticus as postulated by Cavalheiroet al. (1991). In our study, in acute and silent periods, AngII levelswere very low as well as AngII AT1 expression. In contrast, thechronic phase was characterized by an increased level of AngIIand by its AT1 receptor.
Previous results of our group reported that humans with tem-poral lobe epilepsy showed an increased expression of both recep-tors at the hippocampus supporting the idea of an up-regulation ofrenin–angiotensin system in temporal lobe epilepsy (Arganaraz et
Please cite this article in press as: Gouveia, T.L.F., et al. The levels of renin–angmitted to pilocarpine model of epilepsy. Neurochem. Int. (2012), http://dx.doi
al., 2008). Another study suggests that enalapril (ACE inhibitor)and losartan (AngII AT1 receptors blocker) were able to decreaseseizures (Pereira et al., 2010). AngII AT1 receptor, when blockedby losartan, also has a protective function in mouse model of Alz-heimer disease, showing that this receptor has deleterious functionin nervous system, when overexpressed (Danielyan et al., 2010).
In contraposition, our data showed that ACE expression was re-duced in all periods. This data suggests that angiotensinogen maybe cleaved to AngII by tonin, which showed increased expression inall phases. Angiotensinogen concentration in cerebrospinal fluidwas 1000-fold higher than of AngI levels and astrocytes was shownto be the major site of angiotensinogen synthesis in the brain(Deschepper et al., 1986). According to Araujo et al. (2002), toninis also expressed by astrocytes in rat brain. Taken together, thesedata suggest that AngII may be generated due to tonin action,directly from angiotensinogen (Borges et al., 2003).
In this context, this study shows for the first time, changes inthe profile of angiotensin-related polypeptides and their receptorsand converting enzymes in the hippocampus of rats submitted topilocarpine model of temporal lobe epilepsy. This informationmay allow the development of innovative strategies to block theactivation of RAS signaling in epilepsy.
iotensin related components are modified in the hippocampus of rats sub-.org/10.1016/j.neuint.2012.04.012

Fig. 3. Quantification of mRNA expression of receptors by real-time PCR. (A) AngII AT1 receptor showed a decreased expression in acute and silent groups and an increasedexpression in chronic group. (B) AngII AT2 receptor mRNA expression was increased in acute and silent groups showing no alteration into chronic group. (C) Mas mRNAexpression was increased only in silent period. The difference between groups was evaluated using one-way ANOVA followed by Tukey–Kramer t-test. ⁄p 6 0.05, ⁄⁄p 6 0.01and ⁄⁄⁄p 6 0001.
6 T.L.F. Gouveia et al. / Neurochemistry International xxx (2012) xxx–xxx
NCI 3140 No. of Pages 9, Model 5G
25 April 2012
Please cite this article in press as: Gouveia, T.L.F., et al. The levels of renin–angiotensin related components are modified in the hippocampus of rats sub-mitted to pilocarpine model of epilepsy. Neurochem. Int. (2012), http://dx.doi.org/10.1016/j.neuint.2012.04.012

Fig. 4. Quantification of mRNA expression of enzymes by real-time PCR. (A) ACE mRNA level was decreased in acute, silent and chronic phases. (B) NEP mRNA was increasedin acute and silent period. (C) Tonin mRNA levels was increased in chronic phase. The difference between groups was evaluated using one-way ANOVA followed by Tukey–Kramer t-test. ⁄p 6 0.05, ⁄⁄p 6 0.01 and ⁄⁄⁄p 6 0001.
T.L.F. Gouveia et al. / Neurochemistry International xxx (2012) xxx–xxx 7
NCI 3140 No. of Pages 9, Model 5G
25 April 2012
Please cite this article in press as: Gouveia, T.L.F., et al. The levels of renin–angiotensin related components are modified in the hippocampus of rats sub-mitted to pilocarpine model of epilepsy. Neurochem. Int. (2012), http://dx.doi.org/10.1016/j.neuint.2012.04.012

372
373 Q3
374
375
376
377
378
379
380
381
382
383384385386387388389390391392393394395396397398399400401402403404405
406407408409410411412413414415416417418419420421422423424425426427428429430431432433434435436437438439440441442443444445446447448449
Fig. 5. Summary of main found results in all periods of pilocarpine model of epilepsy. " or ; ⁄p 6 0.05, "" or ;;, ⁄⁄p 6 0.01 and """ or ;;; ⁄⁄⁄p 6 0001.
8 T.L.F. Gouveia et al. / Neurochemistry International xxx (2012) xxx–xxx
NCI 3140 No. of Pages 9, Model 5G
25 April 2012
5. Uncited references
De Mello and Frohlich (2011) and Silva et al. (2003).
Acknowledgements
This study was supported by research Grants from Coordenaçãode Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conse-lho Nacional de Desenvolvimento Científico e Tecnológico (CNPq),Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP),Cooperação Interinstitucional de Apoio à Pesquisa sobre o Cérebro(ClnAPCe) and Instituto Nacional de Neurociência Translacional(INNT) (Brazil).
References
Alenina, N., Xu, P., Rentzsch, B., Patkin, E.L., Bader, M., 2008. Genetically alteredanimal models for Mas and angiotensin-(1–7). Exp. Physiol. 93 (5), 528–537.
Araujo, R.C., Lima, M.P., Lomez, E.S., Bader, M., Pesquero, J.B., Sumitani, M., Pesquero,J.L., 2002. Tonin expression in the rat brain and tonin-mediated centralproduction of angiotensin II. Physiol. Behav. 76 (2), 327–333.
Arganaraz, G., Regina Perosa, S., Cristina Lencioni, E., Bader, M., Abrao Cavalheiro, E.,da Graca Naffah-Mazzacoratti, M., Pesquero, J.B., Antonio Silva Jr., J., 2004a. Roleof kinin B1 and B2 receptors in the development of pilocarpine model ofepilepsy. Brain Res. 1013 (1), 30–39.
Arganaraz, G.A., Silva Jr., J.A., Perosa, S.R., Pessoa, L.G., Carvalho, F.F., Bascands, J.L.,Bader, M., da Silva Trindade, E., Amado, D., Cavalheiro, E.A., Pesquero, J.B., daGraca Naffah-Mazzacoratti, M., 2004b. The synthesis and distribution of thekinin B1 and B2 receptors are modified in the hippocampus of rats submitted topilocarpine model of epilepsy. Brain Res. 1006 (1), 114–125.
Arganaraz, G.A., Konno, A.C., Perosa, S.R., Santiago, J.F., Boim, M.A., Vidotti, D.B.,Varella, P.P., Costa, L.G., Canzian, M., Porcionatto, M.A., Yacubian, E.M.,Sakamoto, A.C., Carrete Jr., H., Centeno, R.S., Amado, D., Cavalheiro, E.A.,Junior, J.A., Mazzacoratti Mda, G., 2008. The renin–angiotensin system isupregulated in the cortex and hippocampus of patients with temporal lobeepilepsy related to mesial temporal sclerosis. Epilepsia 49 (8), 1348–1357.
Armstrong, D.L., Garcia, E.A., Ma, T., Quinones, B., Wayner, M.J., 1996. Angiotensin IIblockade of long-term potentiation at the perforant path–granule cell synapsein vitro. Peptides 17 (4), 689–693.
Please cite this article in press as: Gouveia, T.L.F., et al. The levels of renin–angmitted to pilocarpine model of epilepsy. Neurochem. Int. (2012), http://dx.doi
Barnes, N.M., Cheng, C.H., Costall, B., Naylor, R.J., Williams, T.J., Wischik, C.M., 1991.Angiotensin converting enzyme density is increased in temporal cortex frompatients with Alzheimer’s disease. Eur. J. Pharmacol. 200 (2–3), 289–292.
Ben-Ari, Y., Tremblay, E., Riche, D., Ghilini, G., Naquet, R., 1981. Electrographic,clinical and pathological alterations following systemic administration of kainicacid, bicuculline or pentetrazole: metabolic mapping using the deoxyglucosemethod with special reference to the pathology of epilepsy. Neuroscience 6 (7),1361–1391.
Benicky, J., Sánchez-Lemus, E., Honda, M., Pang, T., Orecna, M., Wang, J., Leng, Y.,Chuang, D.M., Saavedra, J.M., 2011. Angiotensin II AT1 receptor blockadeameliorates brain inflammation. Neuropsychopharmacology 36 (4), 857–870.
Bird, E.D., 1980. Chemical pathology of Huntington’s disease. Annu. Rev. Pharmacol.Toxicol. 20, 533–551.
Borges, J.C., Silva Jr., J.A., Gomes, M.A., Lomez, E.S., Leite, K.M., Araujo, R.C., Bader, M.,Pesquero, J.B., Pesquero, J.L., 2003. Tonin in rat heart with experimentalhypertrophy. Am. J. Physiol. Heart Circ. Physiol. 284 (6), H2263–H2268.
Cavalheiro, E.A., Leite, J.P., Bortolotto, Z.A., Turski, W.A., Ikonomidou, C., Turski, L.,1991. Long-term effects of pilocarpine in rats: structural damage of the braintriggers kindling and spontaneous recurrent seizures. Epilepsia 32 (6),778–782.
Chen, L.I., Prakash, O.M., Re, R.N., 1993. The interaction of insulin and angiotensin IIon the regulation of human neuroblastoma cell growth. Mol. Chem.Neuropathol. 18 (1–2), 189–196.
Daemen, M.J., Lombardi, D.M., Bosman, F.T., Schwartz, S.M., 1991. Angiotensin IIinduces smooth muscle cell proliferation in the normal and injured rat arterialwall. Circ. Res. 68 (2), 450–456.
Danielyan, L., Klein, R., Hanson, L.R., Buadze, M., Schwab, M., Gleiter, C.H., Frey, W.H.,2010. Protective effects of intranasal losartan in the APP/PS1 transgenic mousemodel of Alzheimer disease. Rejuvenation Res. 13 (2–3), 195–201.
de Gasparo, M., Catt, K.J., Inagami, T., Wright, J.W., Unger, T., 2000. Internationalunion of pharmacology. XXIII. The angiotensin II receptors. Pharmacol. Rev. 52(3), 415–472.
De Mello, W.C., Frohlich, E.D., 2011. On the local cardiac renin–angiotensin system.Basic and clinical implications. Peptides 32 (8), 1774–1779.
Denny, J.B., Polan-Curtain, J., Wayner, M.J., Armstrong, D.L., 1991. Angiotensin IIblocks hippocampal long-term potentiation. Brain Res. 567 (2), 321–324.
Deschepper, C.F., Bouhnik, J., Ganong, W.F., 1986. Colocalization of angiotensinogenand glial fibrillary acidic protein in astrocytes in rat brain. Brain Res. 374 (1),195–198.
Ferder, L., Inserra, F., Martinez-Maldonado, M., 2006. Inflammation and themetabolic syndrome: role of angiotensin II and oxidative stress. Curr.Hypertens. Rep. 8 (3), 191–198.
Ferder, L.F., Inserra, F., Basso, N., 2003. Effects of renin–angiotensin system blockadein the aging kidney. Exp. Gerontol. 38 (3), 237–244.
iotensin related components are modified in the hippocampus of rats sub-.org/10.1016/j.neuint.2012.04.012

450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484485486487488489490491492493494495496497498499500501502503504505506507
508509510511512513514515516517518519520521522523524525526527528529530531532533534535536537538539540541542543544545546547548549550551552553554555556557558559560561562563564565
T.L.F. Gouveia et al. / Neurochemistry International xxx (2012) xxx–xxx 9
NCI 3140 No. of Pages 9, Model 5G
25 April 2012
Ferrario, C.M., 2006. Role of angiotensin II in cardiovascular disease therapeuticimplications of more than a century of research. J. Renin–AngiotensinAldosterone Syst. 7 (1), 3–14.
Gouveia, T.L., Scorza, F.A., Silva, M.J., Bandeira Tde, A., Perosa, S.R., Arganaraz, G.A.,Silva Mde, P., Araujo, T.R., Frangiotti, M.I., Amado, D., Cavalheiro, E.A., Silva Jr.,J.A., Naffah-Mazzacoratti Mda, G., 2011. Lovastatin decreases the synthesis ofinflammatory mediators in the hippocampus and blocks the hyperthermia ofrats submitted to long-lasting status epilepticus. Epilepsy Behav. 20 (1), 1–5.
Grammatopoulos, T.N., Ahmadi, F., Jones, S.M., Fariss, M.W., Weyhenmeyer, J.A.,Zawada, W.M., 2005. Angiotensin II protects cultured midbrain dopaminergicneurons against rotenone-induced cell death. Brain Res. 1045 (1–2), 64–71.
Jing, G., Grammatopoulos, T., Ferguson, P., Schelman, W., Weyhenmeyer, J., 2004.Inhibitory effects of angiotensin on NMDA-induced cytotoxicity in primaryneuronal cultures. Brain Res Bull. 62 (5), 397–403.
Karamyan, V.T., Speth, R.C., 2007. Identification of a novel non-AT1, non-AT2angiotensin binding site in the rat brain. Brain Res. 1143, 83–91.
Kostenis, E., Milligan, G., Christopoulos, A., Sanchez-Ferrer, C.F., Heringer-Walther,S., Sexton, P.M., Gembardt, F., Kellett, E., Martini, L., Vanderheyden, P.,Schultheiss, H.P., Walther, T., 2005. G-protein-coupled receptor Mas is aphysiological antagonist of the angiotensin II type 1 receptor. Circulation 111(14), 1806–1813.
Lazaroni, T.L., Raslan, A.C., Fontes, W.R., de Oliveira, M.L., Bader, M., Alenina, N.,Moraes, M.F., Dos Santos, R.A., Pereira, G.S., 2012. Angiotensin-(1–7)/Mas axisintegrity is required for the expression of object recognition memory.Neurobiol. Learn Mem. 97 (1), 113–123.
Lippoldt, A., Paul, M., Fuxe, K., Ganten, D., 1995. The brain renin–angiotensinsystem: molecular mechanisms of cell to cell interactions. Clin. Exp. Hypertens.17 (1–2), 251–266.
Martin, K.A., Hockfield, S., 1993. Expression of the mas proto-oncogene in the rathippocampal formation is regulated by neuronal activity. Brain. Res. Mol. BrainRes. 19 (4), 303–309.
McNamara, J.O., 1994. Cellular and molecular basis of epilepsy. J. Neurosci. 14 (6),3413–3425.
Meffert, S., Stoll, M., Steckelings, U.M., Bottari, S.P., Unger, T., 1996. The angiotensinII AT2 receptor inhibits proliferation and promotes differentiation in PC12Wcells. Mol. Cell Endocrinol. 122 (1), 59–67.
Naffah-Mazzacoratti, M.G., Bellissimo, M.I., Cavalheiro, E.A., 1995. Profile ofprostaglandin levels in the rat hippocampus in pilocarpine model of epilepsy.Neurochem. Int. 27 (6), 461–466.
Pereira, M.G., Becari, C., Oliveira, J.A., Salgado, M.C., Garcia-Cairasco, N., Costa-Neto,C.M., 2010. Inhibition of the renin–angiotensin system prevents seizures in a ratmodel of epilepsy. Clin. Sci. (Lond.) 119 (11), 477–482.
Perosa, S.R., Arganaraz, G.A., Goto, E.M., Costa, L.G., Konno, A.C., Varella, P.P.,Santiago, J.F., Pesquero, J.B., Canzian, M., Amado, D., Yacubian, E.M., Carrete Jr.,H., Centeno, R.S., Cavalheiro, E.A., Silva Jr., J.A., Mazzacoratti Mda, G., 2007. KininB1 and B2 receptors are overexpressed in the hippocampus of humans withtemporal lobe epilepsy. Hippocampus 17 (1), 26–33.
Pitkanen, A., Sutula, T.P., 2002. Is epilepsy a progressive disorder? Prospects for newtherapeutic approaches in temporal-lobe epilepsy. Lancet Neurol. 1 (3), 173–181.
Ronchi, F.A., Irigoyen, M.C., Casarini, D.E., 2007. Association of somatic and N-domain angiotensin-converting enzymes from Wistar rat tissue with renaldysfunction in diabetes mellitus. J. Renin–Angiotensin Aldosterone Syst. 8 (1),34–41.
Saavedra, J.M., Ando, H., Armando, I., Baiardi, G., Bregonzio, C., Juorio, A., Macova, M.,2005. Anti-stress and anti-anxiety effects of centrally acting angiotensin II AT1receptor antagonists. Regul. Pept. 128 (3), 227–238.
Sampaio, R.O., Grinberg, M., Leite, J.J., Tarasoutchi, F., Chalela, W.A., Izaki, M., Spina,G.S., Rossi, E.G., Mady, C., 2005. Effect of enalapril on left ventricular diameters
566
Please cite this article in press as: Gouveia, T.L.F., et al. The levels of renin–angmitted to pilocarpine model of epilepsy. Neurochem. Int. (2012), http://dx.doi
and exercise capacity in asymptomatic or mildly symptomatic patients withregurgitation secondary to mitral valve prolapse or rheumatic heart disease.Am. J. Cardiol. 96 (1), 117–121.
Santos, R.A.S., Simoes e Silva, A.C., Maric, C., Silva, D.M.R., Machado, R.P., De Buhr, I.,Heringer-Walther, S., Pinheiro, S.V.B., Lopes, M.T., Bader, M., Mendes, E.P.,Lemos, V.S., Campagnole-Santos, M.J., Schultheiss, H.-P., Speth, R., Walther, T.,2003. Angiotensin-(1-7) is an endogenous ligand for the G protein-coupledreceptor Mas. Proc. Nat. Acad. Sci. U.S.A. 100 (14), 8258–8263.
Schelman, W.R., Kurth, J.L., Berdeaux, R.L., Norby, S.W., Weyhenmeyer, J.A., 1997.Angiotensin II type-2 (AT2) receptor-mediated inhibition of NMDA receptorsignalling in neuronal cells. Brain Res. Mol. Brain Res. 48 (2), 197–205.
Schelman, W.R., Andres, R., Ferguson, P., Orr, B., Kang, E., Weyhenmeyer, J.A., 2004.Angiotensin II attenuates NMDA receptor-mediated neuronal cell death andprevents the associated reduction in Bcl-2 expression. Brain Res. Mol. Brain Res.128 (1), 20–29.
Silva Jr., J.A., Borges, J.C., Gomes, M.A., Lomez, E.S., Leite, K.M., Araujo, R.C.,Bader, M., Pesquero, J.B., Pesquero, J.L., 2003. Tonin in rat heart withexperimental hypertrophy. Am. J. Physiol. Heart Circ. Physiol. 284 (6),H2263–H2268.
Silva Jr, J.A., Goto, E.M., Perosa, S.R., Argañaraz, G.A., Cavalheiro, E.A., Naffah-Mazzacoratti, M.G., Pesquero, J.B., 2008. Kinin B1 receptors facilitate thedevelopment of temporal lobe epilepsy in mice. Int. Immunopharmacol. 8 (2),197–199.
Silveira, K.D., Coelho, F.M., Vieira, A.T., Sachs, D., Barroso, L.C., Costa, V.V., Bretas, T.L.,Bader, M., de Sousa, L.P., da Silva, T.A., dos Santos, R.A., Simoes e Silva, A.C.,Teixeira, M.M., 2010. Anti-inflammatory effects of the activation of theangiotensin-(1–7) receptor, MAS, in experimental models of arthritis. J.Immunol. 185 (9), 5569–5576.
Strawn, W.B., Ferrario, C.M., Tallant, E.A., 1999. Angiotensin-(1–7) reduces smoothmuscle growth after vascular injury. Hypertension 33 (1 Pt 2), 207–211.
Tallant, E.A., Ferrario, C.M., Gallagher, P.E., 2005. Angiotensin-(1–7) inhibits growthof cardiac myocytes through activation of the mas receptor. Am. J. Physiol.Heart Circ. Physiol. 289 (4), H1560–H1566.
Tian, J., Shi, J., Bailey, K., Harris, J.M., Pritchard, A., Lambert, J.C., Chartier-Harlin,M.C., Pickering-Brown, S.M., Lendon, C.L., Mann, D.M., 2004. A polymorphism inthe angiotensin 1-converting enzyme gene is associated with damage tocerebral cortical white matter in Alzheimer’s disease. Neurosci. Lett. 354 (2),103–106.
Turski, W.A., Cavalheiro, E.A., Schwarz, M., Czuczwar, S.J., Kleinrok, Z., Turski, L.,1983. Limbic seizures produced by pilocarpine in rats: behavioural,electroencephalographic and neuropathological study. Behav. Brain Res. 9 (3),315–335.
Varella, P.P., Santiago, J.F., Carrete Jr., H., Higa, E.M., Yacubian, E.M., Centeno, R.S.,Caboclo, L.O., Castro Neto, E.F., Canzian, M., Amado, D., Cavalheiro, E.A., Naffah-Mazzacoratti Mda, G., 2011. Relationship between fluid-attenuated inversion-recovery (FLAIR) signal intensity and inflammatory mediator’s levels in thehippocampus of patients with temporal lobe epilepsy and mesial temporalsclerosis. Arq. Neuropsiquiatr. 69 (1), 91–99.
Vezzani, A., Granata, T., 2005. Brain inflammation in epilepsy: experimental andclinical evidence. Epilepsia 46 (11), 1724–1743.
Wright, J.W., Harding, J.W., 2004. The brain angiotensin system and extracellularmatrix molecules in neural plasticity, learning, and memory. Prog. Neurobiol.72 (4), 263–293.
Wright, J.W., Reichert, J.R., Davis, C.J., Harding, J.W., 2002. Neural plasticity and thebrain renin–angiotensin system. Neurosci. Biobehav. Rev. 26 (5),529–552.
Yamada, T., Horiuchi, M., Dzau, V.J., 1996. Angiotensin II type 2 receptor mediatesprogrammed cell death. Proc. Natl. Acad. Sci. USA 93 (1), 156–160.
iotensin related components are modified in the hippocampus of rats sub-.org/10.1016/j.neuint.2012.04.012