
The last but not the least: the origin and significance of trailing adhesions in fibroblastic cells
11
The Last but not the Least: The Origin and Significance of Trailing Adhesions in Fibroblastic Cells Raphaela Rid, 1 Natalia Schiefermeier, 1 Ilya Grigoriev, 1 J. Victor Small, 1 and Irina Kaverina 1,2 * 1 Institute of Molecular Biology, Austrian Academy of Sciences, Salzburg, Austria 2 Vanderbilt University Medical Center, Nashville, Tennessee Mature adhesions in a motile fibroblast can be classified as stationary ‘‘towing’’ adhesions in the front and sliding trailing adhesions that resist the traction force. Adhesions formed at the front of motile fibroblasts rarely reach the trailing zone, due to disassembly promoted by intensive microtubule targeting. Here, we show that the majority of adhesions found at the trailing edge originate within small short-lived protrusions that extend laterally and backwards from the cell edge. These adhesions enlarge by sliding and by fusion with neighboring adhesions. A further subset of trailing adhesions is initiated at a novel site proximal to trailing stress fibre termini. Following tail retraction, trailing adhesions are actively regen- erated and the stress fibre system is remodeled accordingly; the tensile forces ela- borated by the contractile actin system are consequently redirected according to trailing adhesion location. We conclude that persistent and dynamic anchorage of the cell rear is needed for the maintenance of continuous unidirectional movement of fibroblasts. Cell Motil. Cytoskeleton 61:161–171, 2005. ' 2005 Wiley-Liss, Inc. Key words: cell motility; focal contacts; traction force; mechanosensor INTRODUCTION Cell migration is a complex process requiring the continuous formation and disassembly of matrix adhe- sions. Adhesion sites are initiated via signaling through the Rac or Cdc42 pathways, as small focal complexes associated with protrusive activity at the cell margin. Focal complexes are short-lived; they either turn over at the active edge of the cell, or mature to form large focal adhesion sites [for reviews, see Small et al., 1999; Katsumi et al., 2004; Webb et al., 2002] in response to J. Victor Small’s present address is IMBA, Austrian Academy of Sci- ences, Vienna, Austria. *Correspondence to: Irina Kavernia, Dept. of Cell and Developmental Biology, Vanderbilt University Medical Center, U-4213 Learned Lab, MRB III, 465 21st Ave. South, Nashville, TN 37232-8240. E-mail: [email protected] The supplemental materials described in this article can be found at http://www.interscience.wiley.com/jpages/0886-1544/suppmat Contract grant sponsor: Austrian Science Fund; Contract grant num- ber: P16066. Raphaela Rid’s present address is University of Salzburg, Salzburg, Austria. Natalia Schiefermeier’s present address is Medical University of Inns- bruck, Innsbruck, Austria. Ilya Grigoriev’s present address is Erasmus University, Rotterdam, The Netherlands. Received 10 November 2004; Accepted 24 March 2005 Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/cm.20076 ' 2005 Wiley-Liss, Inc. Cell Motility and the Cytoskeleton 61:161–171 (2005)
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The last but not the least: the origin and significance of trailing adhesions in fibroblastic cells

The Last but not the Least: The Originand Significance of Trailing Adhesions
in Fibroblastic Cells
Raphaela Rid,1 Natalia Schiefermeier,1 Ilya Grigoriev,1 J. Victor Small,1
and Irina Kaverina1,2*
1Institute of Molecular Biology, Austrian Academy of Sciences, Salzburg, Austria2Vanderbilt University Medical Center, Nashville, Tennessee
Mature adhesions in a motile fibroblast can be classified as stationary ‘‘towing’’adhesions in the front and sliding trailing adhesions that resist the traction force.Adhesions formed at the front of motile fibroblasts rarely reach the trailing zone,due to disassembly promoted by intensive microtubule targeting. Here, we showthat the majority of adhesions found at the trailing edge originate within smallshort-lived protrusions that extend laterally and backwards from the cell edge.These adhesions enlarge by sliding and by fusion with neighboring adhesions. Afurther subset of trailing adhesions is initiated at a novel site proximal to trailingstress fibre termini. Following tail retraction, trailing adhesions are actively regen-erated and the stress fibre system is remodeled accordingly; the tensile forces ela-borated by the contractile actin system are consequently redirected according totrailing adhesion location. We conclude that persistent and dynamic anchorage ofthe cell rear is needed for the maintenance of continuous unidirectional movementof fibroblasts. Cell Motil. Cytoskeleton 61:161–171, 2005. ' 2005 Wiley-Liss, Inc.
Key words: cell motility; focal contacts; traction force; mechanosensor
INTRODUCTION
Cell migration is a complex process requiring thecontinuous formation and disassembly of matrix adhe-sions. Adhesion sites are initiated via signaling throughthe Rac or Cdc42 pathways, as small focal complexes
associated with protrusive activity at the cell margin.
Focal complexes are short-lived; they either turn over at
the active edge of the cell, or mature to form large focal
adhesion sites [for reviews, see Small et al., 1999;
Katsumi et al., 2004; Webb et al., 2002] in response to
J. Victor Small’s present address is IMBA, Austrian Academy of Sci-
ences, Vienna, Austria.
*Correspondence to: Irina Kavernia, Dept. of Cell and Developmental
Biology, Vanderbilt University Medical Center, U-4213 Learned Lab,
MRB III, 465 21st Ave. South, Nashville, TN 37232-8240.
E-mail: [email protected]
The supplemental materials described in this article can be found at
http://www.interscience.wiley.com/jpages/0886-1544/suppmat
Contract grant sponsor: Austrian Science Fund; Contract grant num-
ber: P16066.
Raphaela Rid’s present address is University of Salzburg, Salzburg,
Austria.
Natalia Schiefermeier’s present address is Medical University of Inns-
bruck, Innsbruck, Austria.
Ilya Grigoriev’s present address is Erasmus University, Rotterdam,
The Netherlands.
Received 10 November 2004; Accepted 24 March 2005
Published online in Wiley InterScience (www.interscience.wiley.com).
DOI: 10.1002/cm.20076
' 2005 Wiley-Liss, Inc.
Cell Motility and the Cytoskeleton 61:161–171 (2005)

actomyosin contraction signaled by the RhoA pathway[for review, see Bershadsky et al., 2003]. Focal adhe-sions remain stationary or they can slide [for review, seeWehrle-Haller and Imhof, 2002]. Stationary adhesionsare found at the front of a motile cell and a subset ofthese serve as the major ‘‘towing’’ sites supporting thetraction forces that drive cell translocation [Beningoet al., 2002]. Trailing ‘‘sliding’’ adhesions at the cell rearresist this towing force.
Despite their analogous protein composition, dif-ferences have been reported in the dynamic propertiesand macro-molecular organization of stationary and slid-ing focal adhesions. Thus, the spacing between integrinmolecules in sliding adhesions is more compact than instationary ones [Ballestrem et al., 2001] and whereas pro-teins are replaced slowly in stationary adhesions, theyundergo fast turnover in sliding adhesions. This fast mol-ecule exchange is thought to be polarized from the distalto the proximal end of sliding adhesions (‘‘adhesiontreadmilling’’); it was proposed to be a mechanism thatunderlies their apparent ‘‘sliding’’ [Wehrle-Haller andImhof, 2002]. Further, as compared to stationary adhe-sions, the connection between integrins and cytoskeletalcomponents in the plaque of sliding adhesions is appa-rently unstable [Laukaitis et al., 2001], such that trailingadhesions can be left behind as integrin-containing ‘‘foot-prints’’ on the substrate [for review, see Webb et al., 2002].
In earlier studies [Izzard and Lochner, 1980; Aber-crombie et al., 1977], trailing adhesions in fibroblastswere seen to arise from the spontaneous redirection ofcell migration, such that initially anterior adhesionsbecame localized to the rear. However, these findingshave not dispelled a common assumption that trailingadhesions are survivors of adhesions created at the lead-ing edge, which persist after the cell body translocatesover them. In either case, trailing adhesions have beenattributed with a negative ‘‘resisting’’ role in motility. Inthe present study, we monitored directional movement ofzyxin-GFP-expressing fibroblasts over extended periods,allowing us for the first time to trace assembly and mor-phogenesis of trailing focal adhesions in persistentlymoving cells. We show that trailing adhesions arise bytwo distinct mechanisms. One major subset of trailingadhesions is formed within small short-living protrusionsdirected away from the main vector of cell relocation.The second one is not associated with protrusive activitybut formed underneath pre-existing stress fibres proximalto their trailing ends.
Moreover, our observations suggest that dynamicanchorage of the rear edge is necessary for the develop-ment of the actin network and thereby contributes to cellguidance by defining the net direction of tensile force.These findings support and extend those of Lo et al.[2004], showing that the proper regulation of stress fibre
contractility is critical for the directional force develop-ment during cell movement.
MATERIALS AND METHODS
Cells
Goldfish fin fibroblasts (line CAR, ATCC) weremaintained in basal Eagle medium with Hepes andnon-essential amino acids and with 12% FBS at 278C.They were transfected transiently as described previ-ously [Kaverina et al., 1999].
In brief, the transfection mixture containing 2 to4 lg of DNA and 14ml of Superfect lipofection agent(Qiagen, Chatsworth, CA) in 400 ll of serum-freemedium was incubated for 30 min at room temperature.Then, 1.2 ml of medium containing 5% serum was addedto the mixture, which was then applied to subconfluentmonolayer cultures on 30-mm Petri dishes for 4 h at278C. Then, the medium was replaced by full culturemedium containing 12% serum. After 2 to 12 h, cellswere replated onto fibronectin-coated coverslips andused for microscopy after at least 48 h. Only cells with avery moderate level of expression were used for experi-ments; their morphology and motile behavior did not dif-fer from those of non-transfected cells.
Fibronectin (Sigma, St. Louis, MO) was coatedonto polylysine-treated coverslips by incubation on adrop of 50 mg/ml fibronectin in PBS for 1 h at room tem-perature (RT); after rinsing in PBS, these coverslips wereused without drying. Fibronectin was stored as a stocksolution in 2 M urea at 48C. For polylysine coating, cov-erslips were incubated in aqueous 100 mg/ml polylysinefor 30 min at RT, rinsed with water, dried, and UVsterilized.
Transfection Constructs
Human zyxin in a pEGFP-N1 (Clontech, Palo Alto,CA) vector was kindly provided by Prof. Jurgen Wehlandand coworkers (GBF, Braunschweig, Germany). MurineEB1 in a pEGFP-N1 (Clontech) vector was donated byDr. Anna Akhamanova (Erasmus University, Rotterdam,The Netherlands). Murine H1 calponin in pEGFP-C1vector was kindly provided by Dr. Mario Gimona (Insti-tute of Molecular Biology, Salzburg, Austria).
Microinjection
Injections were performed with sterile Femtotips(Eppendorf) held in a Leitz Micromanipulator with aFemtoJet pressure supply (Eppendorf). Cells wereinjected with a continuous outflow mode from the needleunder a constant pressure of between 20 and 40 hPa.
Tetramethyl Rhodamine (5-TAMRA; MolecularProbes, Eugene, OR) conjugated vinculin from turkey
162 Rid et al.
gizzard was kindly provided by Drs. K. Rottner (GBF,Braunschweig, Germany) and M. Gimona (Institute ofMolecular Biology, Salzburg, Austria). Small aliquots in2 mg sucrose/mg protein were stored at �708C. Beforeuse, the fluorescent vinculin was dialyzed against 2 mMTris-Acetate, pH 7.0, 50 mM KCl, 0.1 mM DTE, andused at a concentration of 1 mg/ml.
Video Microscopy and Image Analysis
CAR cells were observed in an open chamber atroom temperature on an inverted microscope (Axiovert135TV; Zeiss, Austria) equipped for epifluorescence andphase contrast microscopy. Video microscopy was per-formed with a 100�/NA 1.4 Plan-Apochromat with orwithout 1.6� intermediate magnification. Tungstenlamps (100 W) were used for both transmitted and epi-illumination. Data were acquired with a back illumi-nated, cooled CCD camera from Princeton ResearchInstruments driven by IPLabs software (both from Visi-tron Systems, Germany) and stored as 16-bit digitalimages. The microscope was additionally equipped withshutters (Optilas GmbH, Germany) to allow separaterecordings of video sequences in phase contrast and fluo-rescence channels and with a filter wheel for two fluores-cent channels. Times between frames were 5 to 60 sec.
RESULTS
Anterior Adhesions in Motile FibroblastsDo Not Reach the Rear of the Cell
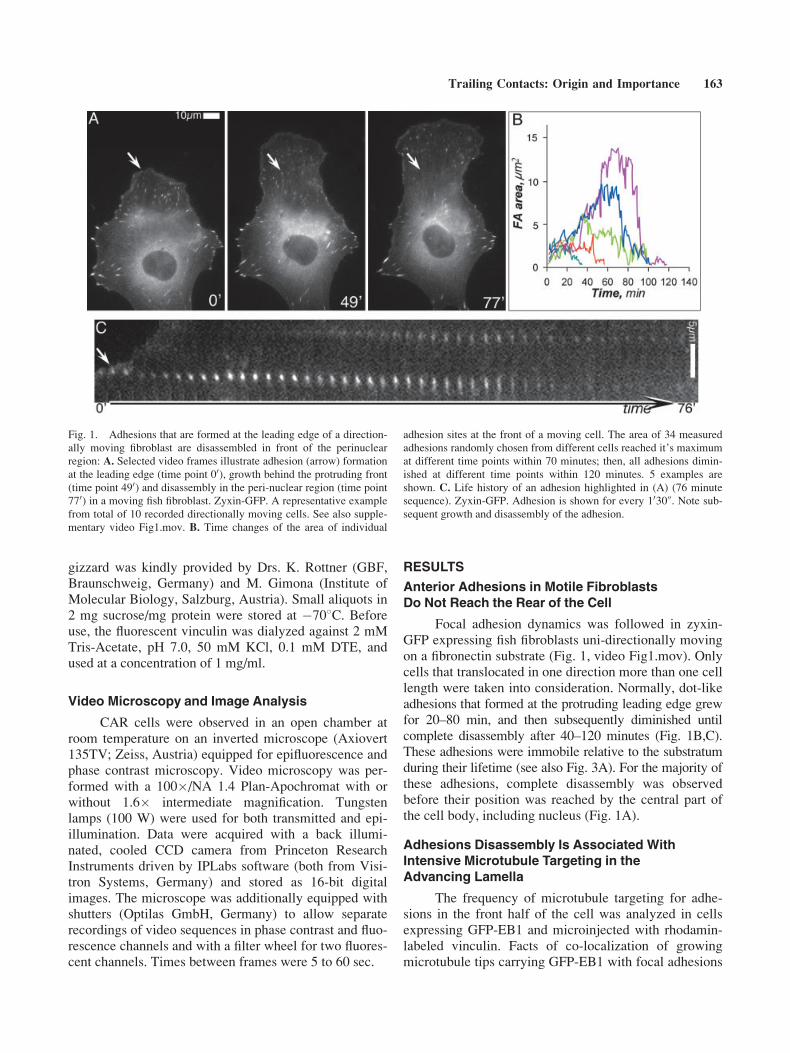
Focal adhesion dynamics was followed in zyxin-GFP expressing fish fibroblasts uni-directionally movingon a fibronectin substrate (Fig. 1, video Fig1.mov). Onlycells that translocated in one direction more than one celllength were taken into consideration. Normally, dot-likeadhesions that formed at the protruding leading edge grewfor 20–80 min, and then subsequently diminished untilcomplete disassembly after 40–120 minutes (Fig. 1B,C).These adhesions were immobile relative to the substratumduring their lifetime (see also Fig. 3A). For the majority ofthese adhesions, complete disassembly was observedbefore their position was reached by the central part ofthe cell body, including nucleus (Fig. 1A).
Adhesions Disassembly Is Associated WithIntensive Microtubule Targeting in theAdvancing Lamella
The frequency of microtubule targeting for adhe-sions in the front half of the cell was analyzed in cellsexpressing GFP-EB1 and microinjected with rhodamin-labeled vinculin. Facts of co-localization of growingmicrotubule tips carrying GFP-EB1 with focal adhesions
Fig. 1. Adhesions that are formed at the leading edge of a direction-
ally moving fibroblast are disassembled in front of the perinuclear
region: A. Selected video frames illustrate adhesion (arrow) formation
at the leading edge (time point 00), growth behind the protruding front
(time point 490) and disassembly in the peri-nuclear region (time point
770) in a moving fish fibroblast. Zyxin-GFP. A representative example
from total of 10 recorded directionally moving cells. See also supple-
mentary video Fig1.mov. B. Time changes of the area of individual
adhesion sites at the front of a moving cell. The area of 34 measured
adhesions randomly chosen from different cells reached it’s maximum
at different time points within 70 minutes; then, all adhesions dimin-
ished at different time points within 120 minutes. 5 examples are
shown. C. Life history of an adhesion highlighted in (A) (76 minute
sequence). Zyxin-GFP. Adhesion is shown for every 1030@. Note sub-
sequent growth and disassembly of the adhesion.
Trailing Contacts: Origin and Importance 163

were considered as targeting events. As shown in Figure2E, the peak of targeting frequency was observed foradhesions that were positioned 20 to 40 lm behind theleading edge. Notably, the most mature (largest) adhe-sions were found in the same zone (Fig. 2D). Hence, areduction of adhesions followed a period of intensive tar-geting by microtubules (Fig. 2A–C; videos Fig2B.movand Fig2C.mov). The extent of adhesion disassemblycorrelated with the number of preceding targetingevents.
Small Protrusions at the Cell Rear Provide theMost Common Source for Trailing Adhesions
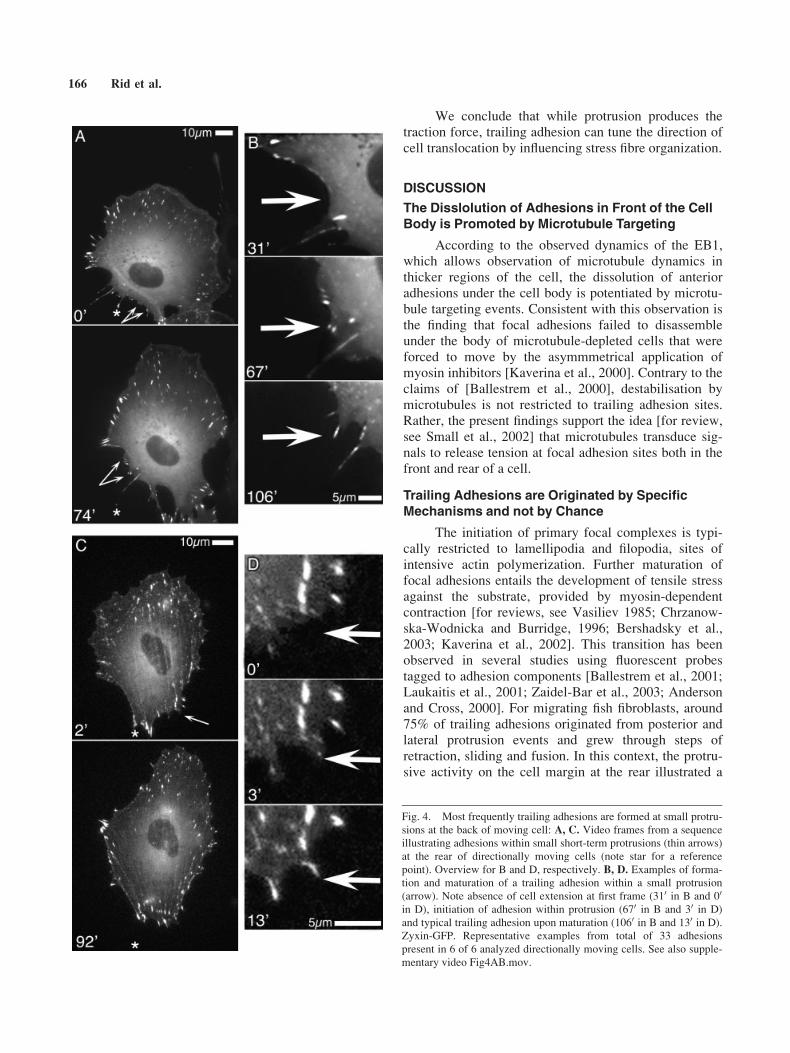
Consistent with the observations above, analysis ofthe life histories of trailing adhesions showed that only8% originated at the leading edge of the cell and then16% originated from lateral regions (Fig. 3B). The mostcommon sites of initiation of trailing adhesions (53%)were protrusions at the rear and flanks of the cell. Com-monly, short-lived lamellipodia extended backwards atan otherwise stable cell edge (Fig. 4, video Fid4AB.-mov). Adhesions formed in these protrusions maturedrapidly in response to tensile force applied from themoving cell body and developed into sliding trailingadhesions. Figure 3A depicts the tracks of trailing adhe-sions during their life cycle and shows that they com-monly arise from the rear and, less frequently, from theflanks of the cell. At later stages, sliding trailing adhe-sions either fused with newly formed ones or detachedfrom the substrate as newer ones took over their function(e.g., complementary video 4).
Novel Site of Adhesion Initiation inFront of Trailing Adhesions
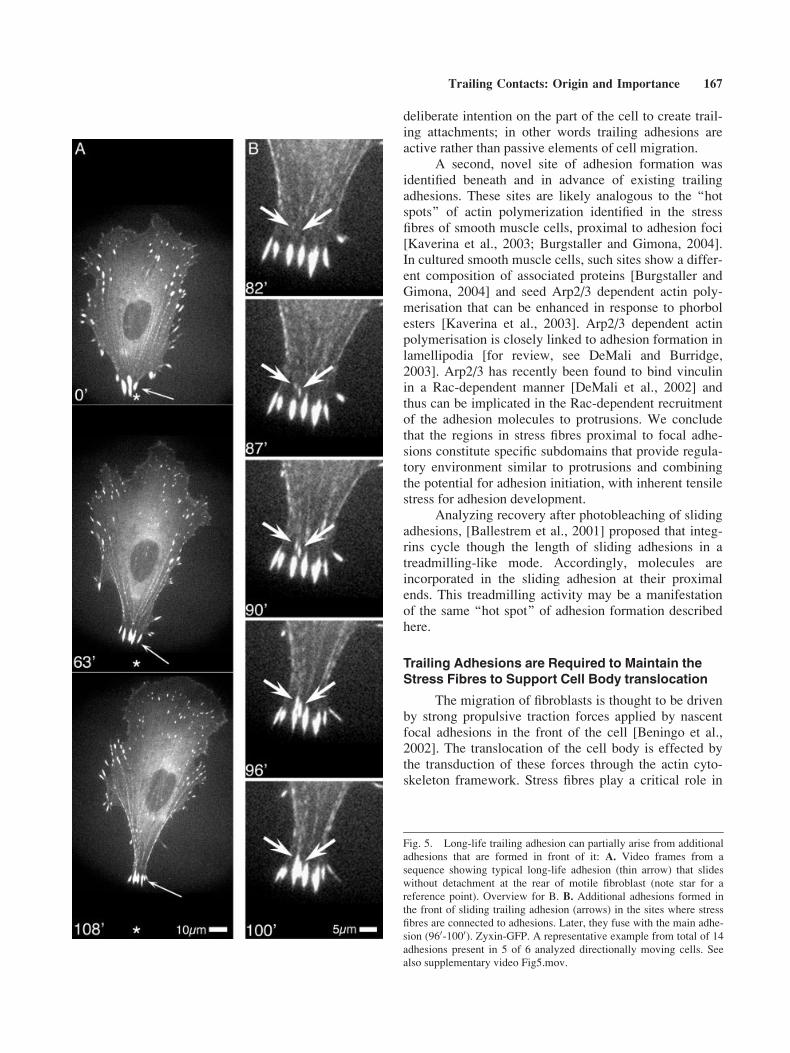
Additional focal adhesions were found to form atthe rear of cell body in front of sliding trailing adhesions(Fig. 5, video Fig5.mov). In contrast to the known adhe-sion types, the origination of these adhesions did notrequire protrusive activity. The sites of their initiationwere located under the proximal parts of stress fibresattached to sliding trailing adhesions. In the process ofmaturation, additional adhesions fused with theapproaching sliding adhesion at the end of the samestress fibre (Fig. 5B). In sum, 23% of nascent adhesionsites that later developed into trailing adhesions wereassociated with stress fibres (Fig. 3B).
Persistent Cell Movement Requires theRegeneration of Trailing Adhesions AfterComplete Detachment of Cell Rear
The locomotory cycle of fibroblastic cells ofteninvolves a stage of complete detachment of cell rear.
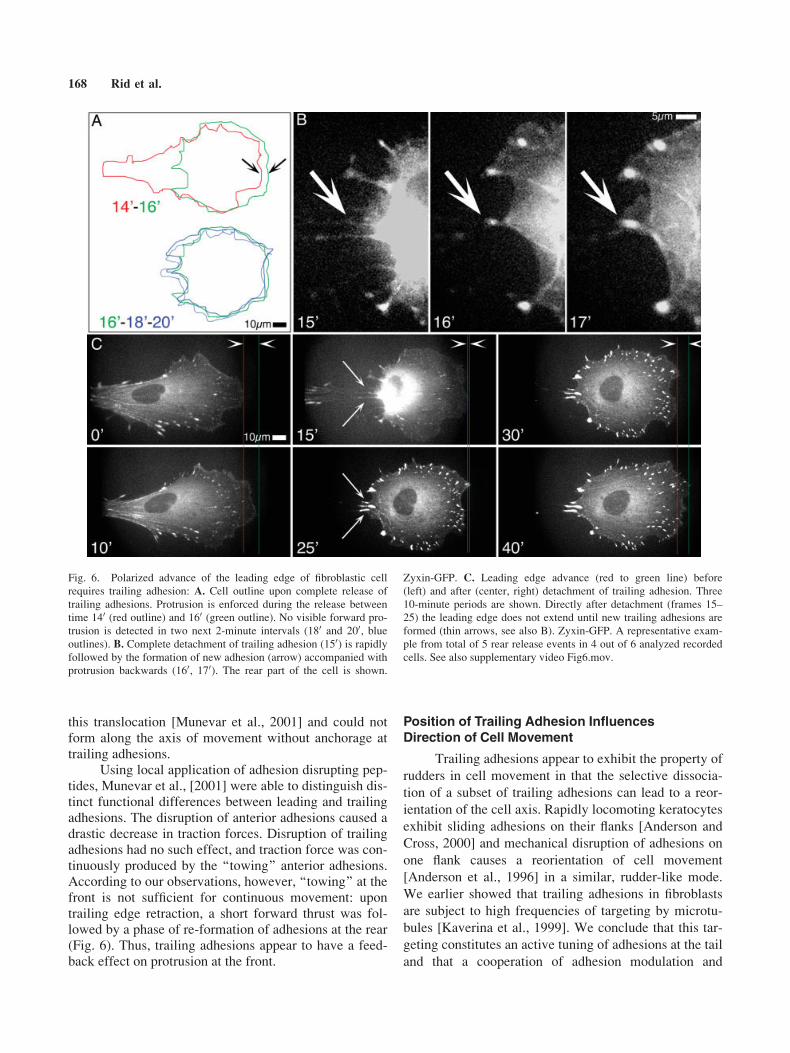
Detachment is followed by enhanced protrusion at theleading edge of the cell [Chen, 1979; Dunn, 1980]. Weobserved that the boost of protrusion was restricted tothe front of the cell only for 1–2 minutes after detach-ment (Fig 6A, time 140–160). This could be explained bya fast redistribution of cell mass to the front due to con-traction of stress fibres upon the release of their rear ends[Dunn, 1980], as well as by increase of towing at thefront adhesions upon loss of resistance [Munevar et al.,2001]. Subsequent to this event, when there was no moredirectional force, we observed formation of protrusionsin a non-polarized manner around the cell perimeter. Inthe example shown, the leading edge of the cell did notadvance significantly for 8–15 minutes (Fig6A, time160–200). Protrusions at the former rear of the cell servedfor initiation of new adhesion sites that provided attach-ment for the trailing edge (Fig. 6B, 160–170). As soon asnew trailing adhesions were formed, leading edgeadvance was restored and polarized movement continued(Fig. 6C, 300–400). See also video Fig6.mov.
Fig. 2. Adhesion release in the front of the perinuclear region is pro-
moted by microtubule targeting activity: A. Video frames from a
sequence showing disassembly of adhesions behind the protruding edge
(scattered line). TAMRA-vinculin marked adhesion pattern is shown at
time points 10@ and 8030@. Inset area from frame 10@ is shown at the
right. On this image, EB1 localizations from three subsequent frames
(time points 00 in red, 10@ in green and 20@ in blue) are overlaid; margins
of focal adhesions at time point 10@ are shown in white. Growing micro-
tubule tips target large adhesions (arrows) that disassembly at frame
8030@. A representative example from total of 6 recorded directionally
moving cells is shown. B. Video frames from a sequence showing grow-
ing plus-ends of microtubules (green) approaching adhesions (red).
Localization of EB1 patches according to adhesions suggests that each
of them will target an adhesion in time period from 0@ to 25@ after thecurrent frame (compare with EB1 localization in A). Majority of plus-
ends are concentrated close to the most mature adhesions behind the
leading edge. Note that adhesions in an oval frame undergo more inten-
sive targeting than those in rectangular frame. GFP-EB1 and TAMRA-
vinculin. See also supplementary video Fig2B.mov. C. Long-term obser-
vations of large adhesions in the zone of maximal maturation following
microtubule targeting shown in A. Note effective disassembly of adhe-
sions. Adhesions in oval frame are destroyed faster than those in rectan-
gular frame (compare with A). TAMRA-vinculin. See also supplemen-
tary video Fig2C.mov. In B and C a representative example from total of
8 (other than in A) recorded directionally moving cells is shown. First,
GFP-EB1 and TAMRA-vinculin in each cell were recorded for 3 minutes
at 5 seconds/frame rate and then, TAMRA-vinculin in each cell was
recorded for 20 minutes at 30 seconds/frame rate. D. Correlation
between the distance from the advancing cell edge and the area of indi-
vidual adhesion sites. The area of adhesions located farther than 40 lmfrom the leading edge goes down. E. In the same cells as D, correlation
between the distance from the advancing cell edge and the number of
microtubule tips (targeting events). Most intensive targeting occurs
between 20 and 40 lm from leading edge. Analysis of 4 directional mov-
ing cells (D and E), other than in (A, B and C), recorded at 5 seconds/
frame rate for 8 minutes. Each 5th frame was quantified.
164 Rid et al.
Trailing Adhesions Influence the Direction ofMotility Via Stress Fibre Organization
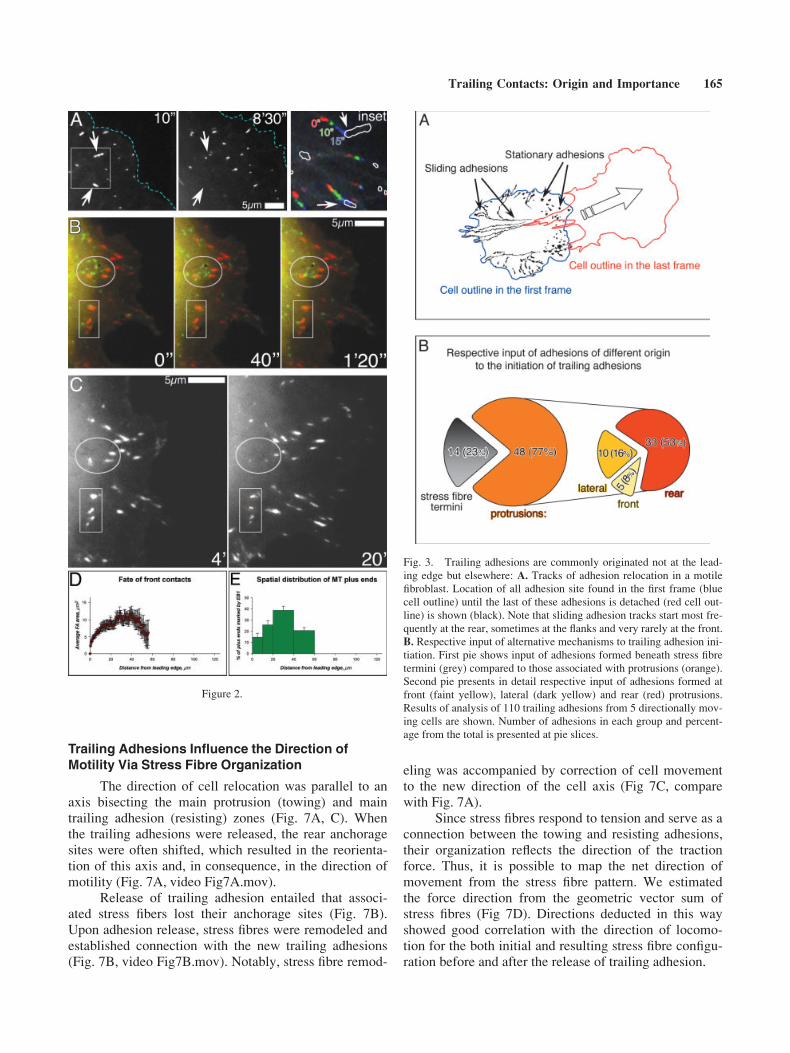
The direction of cell relocation was parallel to anaxis bisecting the main protrusion (towing) and maintrailing adhesion (resisting) zones (Fig. 7A, C). Whenthe trailing adhesions were released, the rear anchoragesites were often shifted, which resulted in the reorienta-tion of this axis and, in consequence, in the direction ofmotility (Fig. 7A, video Fig7A.mov).
Release of trailing adhesion entailed that associ-ated stress fibers lost their anchorage sites (Fig. 7B).Upon adhesion release, stress fibres were remodeled andestablished connection with the new trailing adhesions(Fig. 7B, video Fig7B.mov). Notably, stress fibre remod-
eling was accompanied by correction of cell movementto the new direction of the cell axis (Fig 7C, comparewith Fig. 7A).
Since stress fibres respond to tension and serve as aconnection between the towing and resisting adhesions,their organization reflects the direction of the tractionforce. Thus, it is possible to map the net direction ofmovement from the stress fibre pattern. We estimatedthe force direction from the geometric vector sum ofstress fibres (Fig 7D). Directions deducted in this wayshowed good correlation with the direction of locomo-tion for the both initial and resulting stress fibre configu-ration before and after the release of trailing adhesion.
Fig. 3. Trailing adhesions are commonly originated not at the lead-
ing edge but elsewhere: A. Tracks of adhesion relocation in a motile
fibroblast. Location of all adhesion site found in the first frame (blue
cell outline) until the last of these adhesions is detached (red cell out-
line) is shown (black). Note that sliding adhesion tracks start most fre-
quently at the rear, sometimes at the flanks and very rarely at the front.
B. Respective input of alternative mechanisms to trailing adhesion ini-
tiation. First pie shows input of adhesions formed beneath stress fibre
termini (grey) compared to those associated with protrusions (orange).
Second pie presents in detail respective input of adhesions formed at
front (faint yellow), lateral (dark yellow) and rear (red) protrusions.
Results of analysis of 110 trailing adhesions from 5 directionally mov-
ing cells are shown. Number of adhesions in each group and percent-
age from the total is presented at pie slices.
Figure 2.
Trailing Contacts: Origin and Importance 165
We conclude that while protrusion produces thetraction force, trailing adhesion can tune the direction ofcell translocation by influencing stress fibre organization.
DISCUSSION
The Disslolution of Adhesions in Front of the CellBody is Promoted by Microtubule Targeting
According to the observed dynamics of the EB1,which allows observation of microtubule dynamics inthicker regions of the cell, the dissolution of anterioradhesions under the cell body is potentiated by microtu-bule targeting events. Consistent with this observation isthe finding that focal adhesions failed to disassembleunder the body of microtubule-depleted cells that wereforced to move by the asymmmetrical application ofmyosin inhibitors [Kaverina et al., 2000]. Contrary to theclaims of [Ballestrem et al., 2000], destabilisation bymicrotubules is not restricted to trailing adhesion sites.Rather, the present findings support the idea [for review,see Small et al., 2002] that microtubules transduce sig-nals to release tension at focal adhesion sites both in thefront and rear of a cell.
Trailing Adhesions are Originated by SpecificMechanisms and not by Chance
The initiation of primary focal complexes is typi-cally restricted to lamellipodia and filopodia, sites ofintensive actin polymerization. Further maturation offocal adhesions entails the development of tensile stressagainst the substrate, provided by myosin-dependentcontraction [for reviews, see Vasiliev 1985; Chrzanow-ska-Wodnicka and Burridge, 1996; Bershadsky et al.,2003; Kaverina et al., 2002]. This transition has beenobserved in several studies using fluorescent probestagged to adhesion components [Ballestrem et al., 2001;Laukaitis et al., 2001; Zaidel-Bar et al., 2003; Andersonand Cross, 2000]. For migrating fish fibroblasts, around75% of trailing adhesions originated from posterior andlateral protrusion events and grew through steps ofretraction, sliding and fusion. In this context, the protru-sive activity on the cell margin at the rear illustrated a
Fig. 4. Most frequently trailing adhesions are formed at small protru-
sions at the back of moving cell: A, C. Video frames from a sequence
illustrating adhesions within small short-term protrusions (thin arrows)
at the rear of directionally moving cells (note star for a reference
point). Overview for B and D, respectively. B, D. Examples of forma-
tion and maturation of a trailing adhesion within a small protrusion
(arrow). Note absence of cell extension at first frame (310 in B and 00
in D), initiation of adhesion within protrusion (670 in B and 30 in D)
and typical trailing adhesion upon maturation (1060 in B and 130 in D).
Zyxin-GFP. Representative examples from total of 33 adhesions
present in 6 of 6 analyzed directionally moving cells. See also supple-
mentary video Fig4AB.mov.
166 Rid et al.
deliberate intention on the part of the cell to create trail-ing attachments; in other words trailing adhesions areactive rather than passive elements of cell migration.
A second, novel site of adhesion formation wasidentified beneath and in advance of existing trailingadhesions. These sites are likely analogous to the ‘‘hotspots’’ of actin polymerization identified in the stressfibres of smooth muscle cells, proximal to adhesion foci[Kaverina et al., 2003; Burgstaller and Gimona, 2004].In cultured smooth muscle cells, such sites show a differ-ent composition of associated proteins [Burgstaller andGimona, 2004] and seed Arp2/3 dependent actin poly-merisation that can be enhanced in response to phorbolesters [Kaverina et al., 2003]. Arp2/3 dependent actinpolymerisation is closely linked to adhesion formation inlamellipodia [for review, see DeMali and Burridge,2003]. Arp2/3 has recently been found to bind vinculinin a Rac-dependent manner [DeMali et al., 2002] andthus can be implicated in the Rac-dependent recruitmentof the adhesion molecules to protrusions. We concludethat the regions in stress fibres proximal to focal adhe-sions constitute specific subdomains that provide regula-tory environment similar to protrusions and combiningthe potential for adhesion initiation, with inherent tensilestress for adhesion development.
Analyzing recovery after photobleaching of slidingadhesions, [Ballestrem et al., 2001] proposed that integ-rins cycle though the length of sliding adhesions in atreadmilling-like mode. Accordingly, molecules areincorporated in the sliding adhesion at their proximalends. This treadmilling activity may be a manifestationof the same ‘‘hot spot’’ of adhesion formation describedhere.
Trailing Adhesions are Required to Maintain theStress Fibres to Support Cell Body translocation
The migration of fibroblasts is thought to be drivenby strong propulsive traction forces applied by nascentfocal adhesions in the front of the cell [Beningo et al.,2002]. The translocation of the cell body is effected bythe transduction of these forces through the actin cyto-skeleton framework. Stress fibres play a critical role in
Fig. 5. Long-life trailing adhesion can partially arise from additional
adhesions that are formed in front of it: A. Video frames from a
sequence showing typical long-life adhesion (thin arrow) that slides
without detachment at the rear of motile fibroblast (note star for a
reference point). Overview for B. B. Additional adhesions formed in
the front of sliding trailing adhesion (arrows) in the sites where stress
fibres are connected to adhesions. Later, they fuse with the main adhe-
sion (960-1000). Zyxin-GFP. A representative example from total of 14
adhesions present in 5 of 6 analyzed directionally moving cells. See
also supplementary video Fig5.mov.
Trailing Contacts: Origin and Importance 167
this translocation [Munevar et al., 2001] and could notform along the axis of movement without anchorage attrailing adhesions.
Using local application of adhesion disrupting pep-tides, Munevar et al., [2001] were able to distinguish dis-tinct functional differences between leading and trailingadhesions. The disruption of anterior adhesions caused adrastic decrease in traction forces. Disruption of trailingadhesions had no such effect, and traction force was con-tinuously produced by the ‘‘towing’’ anterior adhesions.According to our observations, however, ‘‘towing’’ at thefront is not sufficient for continuous movement: upontrailing edge retraction, a short forward thrust was fol-lowed by a phase of re-formation of adhesions at the rear(Fig. 6). Thus, trailing adhesions appear to have a feed-back effect on protrusion at the front.
Position of Trailing Adhesion InfluencesDirection of Cell Movement
Trailing adhesions appear to exhibit the property of
rudders in cell movement in that the selective dissocia-
tion of a subset of trailing adhesions can lead to a reor-
ientation of the cell axis. Rapidly locomoting keratocytes
exhibit sliding adhesions on their flanks [Anderson and
Cross, 2000] and mechanical disruption of adhesions on
one flank causes a reorientation of cell movement
[Anderson et al., 1996] in a similar, rudder-like mode.
We earlier showed that trailing adhesions in fibroblasts
are subject to high frequencies of targeting by microtu-
bules [Kaverina et al., 1999]. We conclude that this tar-
geting constitutes an active tuning of adhesions at the tail
and that a cooperation of adhesion modulation and
Fig. 6. Polarized advance of the leading edge of fibroblastic cell
requires trailing adhesion: A. Cell outline upon complete release of
trailing adhesions. Protrusion is enforced during the release between
time 140 (red outline) and 160 (green outline). No visible forward pro-
trusion is detected in two next 2-minute intervals (180 and 200, blueoutlines). B. Complete detachment of trailing adhesion (150) is rapidlyfollowed by the formation of new adhesion (arrow) accompanied with
protrusion backwards (160, 170). The rear part of the cell is shown.
Zyxin-GFP. C. Leading edge advance (red to green line) before
(left) and after (center, right) detachment of trailing adhesion. Three
10-minute periods are shown. Directly after detachment (frames 15–
25) the leading edge does not extend until new trailing adhesions are
formed (thin arrows, see also B). Zyxin-GFP. A representative exam-
ple from total of 5 rear release events in 4 out of 6 analyzed recorded
cells. See also supplementary video Fig6.mov.
168 Rid et al.
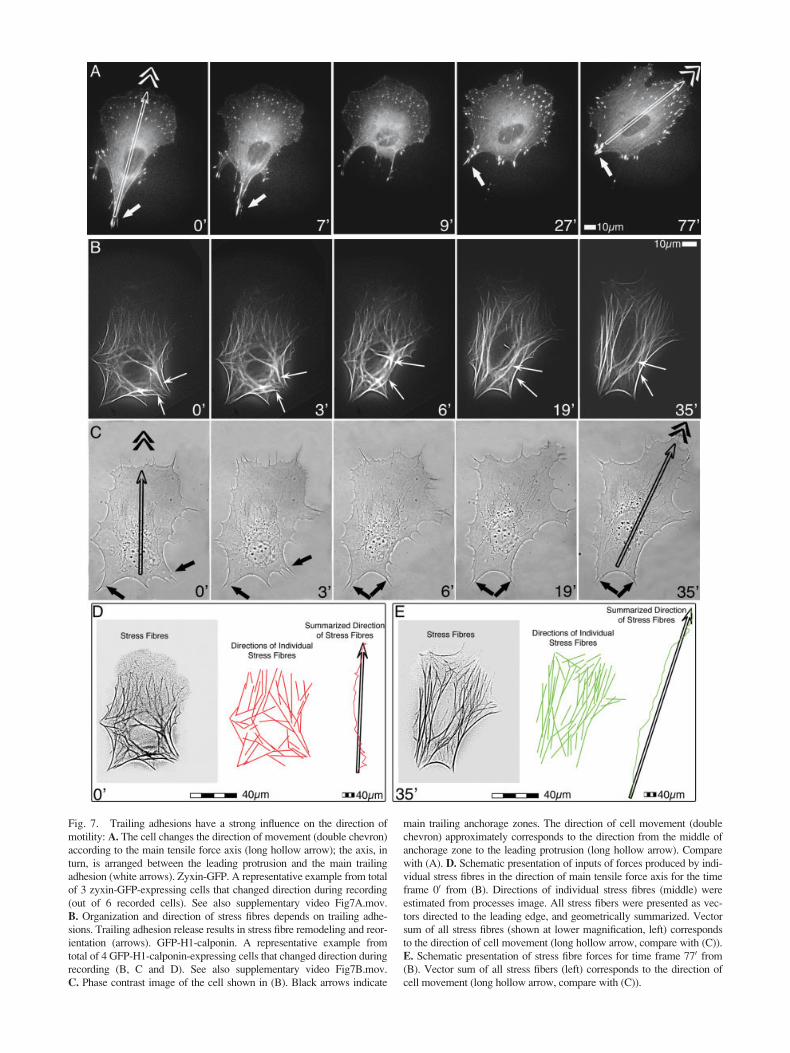
Fig. 7. Trailing adhesions have a strong influence on the direction of
motility: A. The cell changes the direction of movement (double chevron)
according to the main tensile force axis (long hollow arrow); the axis, in
turn, is arranged between the leading protrusion and the main trailing
adhesion (white arrows). Zyxin-GFP. A representative example from total
of 3 zyxin-GFP-expressing cells that changed direction during recording
(out of 6 recorded cells). See also supplementary video Fig7A.mov.
B. Organization and direction of stress fibres depends on trailing adhe-
sions. Trailing adhesion release results in stress fibre remodeling and reor-
ientation (arrows). GFP-H1-calponin. A representative example from
total of 4 GFP-H1-calponin-expressing cells that changed direction during
recording (B, C and D). See also supplementary video Fig7B.mov.
C. Phase contrast image of the cell shown in (B). Black arrows indicate
main trailing anchorage zones. The direction of cell movement (double
chevron) approximately corresponds to the direction from the middle of
anchorage zone to the leading protrusion (long hollow arrow). Compare
with (A). D. Schematic presentation of inputs of forces produced by indi-
vidual stress fibres in the direction of main tensile force axis for the time
frame 00 from (B). Directions of individual stress fibres (middle) were
estimated from processes image. All stress fibers were presented as vec-
tors directed to the leading edge, and geometrically summarized. Vector
sum of all stress fibres (shown at lower magnification, left) corresponds
to the direction of cell movement (long hollow arrow, compare with (C)).
E. Schematic presentation of stress fibre forces for time frame 770 from(B). Vector sum of all stress fibers (left) corresponds to the direction of
cell movement (long hollow arrow, compare with (C)).

protrusive activity determines the net orientation ofmovement.
This cooperation is likely coordinated viamechanical signals transduced through the stress fibrenetwork. The absence of such a coordination mecha-nism may explain the random movement of fibroblastslacking myosin IIb, which is restricted in its localiza-tion to stress fibre bundles [Lo et al., 2004]. These cellsexert disorganized forces on the substrate leading torandom multiple protrusive activity and are thereforeunable to directional movement. In the same context,stable protrusion can be mechanically induced bydirectional application of tensile stress to a cell platedon a flexible substrate [Lo et al., 2000] by substratedeformation.
A number of recent studies reveal possible mech-anisms for mechanosensory facilitating of protrusion.For instance, organization of adhesion pattern in thefront of the cell can be directly regulated by tensileforce [for review, see Geiger and Bershadsky, 2002;Katsumi et al., 2004]. Direction of force decides onlocation, where integrins make strong connection withextracellular matrix. Artificial focal complex formationcan be induced by force application [Galbraith et al.,2002] that stabilizes integrin-cytoskeleton linkage [vonWichert et al., 2003]. New adhesions that come underlocal stress mature earlier than others [Riveline et al.,2001] and provide a stable base for advancing leadingedge.
Also, mechanical force can determine cell motil-ity reactions by means of stretch-activated ion chan-nels. Ca2þ channels are involved in the modulation ofcell movement [Lee et al., 1999] and local concentra-tions of intracellular Ca2þ can enhance traction at thefront of fibroblasts [Munevar et al., 2004]. Accordingly,spatial activation of ion channels has been implicatedto play a role in tension-induced fibroblastic motility.These mechanisms, among others, could contribute tothe fine tuning of protrusion localization by trailingadhesions.
In this study, we show that trailing adhesions,which support stress fibre directionality are necessaryfor persistent forward movement of fibroblasts, and thatthey are actively generated at the rear of the cell. Wesuggest the existence of a feedback, mechanosensingmechanism that maintains trailing adhesion renewal inmoving cells. The nature of this mechanism remains tobe investigated.
ACKNOWLEDGMENTS
We thank Drs. Anna Akhmanova, Mario Gimona,and Juergen Wehland for providing expression constructs.
REFERENCES
Abercrombie M, Dunn GA, Heath JP. 1977. The shape and move-
ment of fibroblasts in culture. Soc Gen Physiol Ser 32:
57–70.
Anderson KI, Cross R. 2000. Contact dynamics during keratocyte
motility. Curr Biol 10:253–260.
Anderson KI, Wang YL, Small JV. 1996. Coordination of protrusion
and translocation of the keratocyte involves rolling of the cell
body. J Cell Biol 134:1209–1218.
Ballestrem C, Hinz B, Imhof BA, Wehrle-Haller B. 2001. Marching
at the front and dragging behind: differential alphaVbeta3-
integrin turnover regulates focal adhesion behavior. J Cell Biol
155:1319–1332.
Ballestrem C, Wehrle-Haller B, Hinz B, Imhof BA. 2000. Actin-
dependent lamellipodia formation and microtubule-dependent
tail retraction control-directed cell migration. Mol Biol Cell
11:2999–3012.
Beningo KA, Lo CM, Wang YL. 2002. Flexible polyacrylamide
substrata for the analysis of mechanical interactions at cell-sub-
stratum adhesions. Methods Cell Biol 69:325–339.
Bershadsky AD, Balaban NQ, Geiger B. 2003. Adhesion-dependent
cell mechanosensitivity. Annu Rev Cell Dev Biol 19:677–695.
Burgstaller G, Gimona M. 2004. Actin cytoskeleton remodelling via
local inhibition of contractility at discrete microdomains. J Cell
Sci 117:223–231.
Chen WT. 1979. Induction of spreading during fibroblast movement.
J Cell Biol 81:684–691.
Chrzanowska-Wodnicka M, Burridge K. 1996. Rho-stimulated con-
tractility drives the formation of stress fibers and focal adhe-
sions. J Cell Biol 133:1403–1415.
DeMali KA, Barlow CA, Burridge K. 2002. Recruitment of the
Arp2/3 complex to vinculin: coupling membrane protrusion to
matrix adhesion. J Cell Biol 159:881–891.
DeMali KA, Burridge K. 2003. Coupling membrane protrusion and
cell adhesion. J Cell Sci 116:2389–2397.
Dunn GA. 1980. The locomotory machinery of fibroblasts. Eur J Can-
cer 16:6–8.
Galbraith CG, Yamada KM, Sheetz MP. 2002. The relationship
between force and focal complex development. J Cell Biol
159:695–705.
Geiger B, Bershadsky A. 2002. Exploring the neighborhood: adhe-
sion-coupled cell mechanosensors. Cell 110:139–142.
Izzard CS, Lochner LR. 1980. Formation of cell-to-substrate contacts
during fibroblast motility: an interference-reflexion study.
J Cell Sci 42:81–116.
Katsumi A, Orr AW, Tzima E, Schwartz MA. 2004. Integrins in
mechanotransduction. J Biol Chem 279:12001–12004.
Kaverina I, Krylyshkina O, Gimona M, Beningo K, Wang YL,
Small JV. 2000. Enforced polarisation and locomotion of
fibroblasts lacking microtubules. Curr Biol 10:739–742.
Kaverina I, Krylyshkina O, Small JV. 1999. Microtubule targeting
of substrate contacts promotes their relaxation and dissociation.
J Cell Biol 146:1033–1044.
Kaverina I, Krylyshkina O, Small JV. 2002. Regulation of substrate
adhesion dynamics during cell motility. Int J Biochem Cell
Biol 34:746–761.
Kaverina I, Stradal TE, Gimona M. 2003. Podosome formation in
cultured A7r5 vascular smooth muscle cells requires Arp2/3-
dependent de-novo actin polymerization at discrete microdo-
mains. J Cell Sci 116:4915–4924.
Laukaitis CM, Webb DJ, Donais K, Horwitz AF. 2001. Differential
dynamics of alpha 5 integrin, paxillin, and alpha-actinin during
formation and disassembly of adhesions in migrating cells.
J Cell Biol 153:1427–1440.
170 Rid et al.

Lee J, Ishihara A, Oxford G, Johnson B, Jacobson K. 1999. Regula-
tion of cell movement is mediated by stretch-activated calcium
channels. Nature 400:382–386.
Lo CM, Buxton DB, Chua GC, Dembo M, Adelstein RS, Wang
YL. 2004. Nonmuscle myosin IIb is involved in the guidance
of fibroblast migration. Mol Biol Cell 15:982–989.
Lo CM, Wang HB, Dembo M, Wang YL. 2000. Cell move-
ment is guided by the rigidity of the substrate. Biophys J
79:144–152.
Munevar S, Wang YL, Dembo M. 2001. Distinct roles of frontal and
rear cell-substrate adhesions in fibroblast migration. Mol Biol
Cell 12:3947–3954.
Munevar S, Wang YL, Dembo M. 2004. Regulation of mechanical
interactions between fibroblasts and the substratum by stretch-
activated Ca2þ entry. J Cell Sci 117:85–92.
Riveline D, Zamir E, Balaban NQ, Schwarz US, Ishizaki T,
Narumiya S, Kam Z, Geiger B, Bershadsky AD. 2001. Focal
contacts as mechanosensors: externally applied local mechani-
cal force induces growth of focal contacts by an mDia1-
dependent and ROCK-independent mechanism. J Cell Biol
153:1175–1186.
Small JV, Geiger B, Kaverina I, Bershadsky A. 2002. How do micro-
tubules guide migrating cells? Nat Rev Mol Cell Biol 3:957–964.
Small JV, Rottner K, Kaverina I. 1999. Functional design in the
actin cytoskeleton. Curr Opin Cell Biol 11:54–60.
Vasiliev JM. 1985. Spreading of non-transformed and transformed
cells. Biochim Biophys Acta 780:21–65.von Wichert G, Haimovich B, Feng GS, Sheetz MP. 2003. Force-
dependent integrin-cytoskeleton linkage formation requiresdownregulation of focal complex dynamics by Shp2. Embo J22:5023–5035.
Webb DJ, Parsons JT, Horwitz AF. 2002. Adhesion assembly, disas-sembly and turnover in migrating cells: over and over and overagain. Nat Cell Biol 4:E97–100.
Wehrle-Haller B, Imhof B. 2002. The inner lives of focal adhesions.
Trends Cell Biol 12:382–389.Zaidel-Bar R, Ballestrem C, Kam Z, Geiger B. 2003. Early molecu-
lar events in the assembly of matrix adhesions at the leadingedge of migrating cells. J Cell Sci 116:4605–4613.
Trailing Contacts: Origin and Importance 171




















