
Heterogeneous distribution of pyruvate dehydrogenase in the matrix of mitochondria

The F420H2-dehydrogenase from Methanolobus tindarius : cloningof the ¡d operon and expression of the genes in Escherichia coli
David J. Westenberg a, Annett Braune b, Claudia Ruppert c, Volker Muëller c,Christina Herzberg a, Gerhard Gottschalk a, Michael Blaut b;*
a Institut fuër Mikrobiologie und Genetik, Georg-August-Universitaët Goëttingen, Grisebachstr. 8, D-37077 Goëttingen, Germanyb Gastrointestinale Mikrobiologie, DIfE, Arthur-Scheunert-Allee 114-116, D-14558 Bergholz-Rehbruëcke, Germany
c Institut fuër Genetik und Mikrobiologie, Lehrstuhl fuër Mikrobiologie, Maria-Ward-Str. 1a, D-80638 Muënchen, Germany
Received 17 October 1998; received in revised form 15 November 1998; accepted 18 November 1998
Abstract
The membrane-bound F420H2-dehydrogenase from the methylotrophic methanogen Methanolobus tindarius oxidizes reducedcoenzyme F420 and feeds the electrons into an energy-conserving electron transport chain. Based on the N-terminal amino acidsequence of the 40-kDa subunit of F420H2-dehydrogenase the corresponding gene ffdB was detected in chromosomal DNA ofM. tindarius. Sequence analysis, primer extension, and RT-PCR experiments indicated that ffdB is part of an operon harboringthree additional open reading frames (ffdA, ffdC, ffdD). The corresponding mRNA transcript and transcription start sites weredetermined. All four genes could be heterologously expressed in Escherichia coli. z 1999 Federation of European Micro-biological Societies. Published by Elsevier Science B.V. All rights reserved.
Keywords: Methanolobus tindarius ; F420H2-dehydrogenase; Electron transport; ¡dB gene; Heterologous expression
1. Introduction
The conversion of methanogenic substrates tomethane by methanogenic archaea involves a se-quence of unique reactions [1]. Methyl-coenzyme Mreductase catalyzes the reductive demethylation ofmethyl-coenzyme M (2-methylthioethane sulfonate)with 7-mercaptoheptanoyl threonine phosphate(HS-CoB) as reductant (Eq. 1).
CH3 ÿ Sÿ CoM�HSÿ CoB!CH4
�CoMÿ Sÿ Sÿ CoB �1�
The latter product acts as electron acceptor of themembrane-bound heterodisul¢de reductase whichcouples the reduction of CoM-S-S-CoB (Eq. 2)with the generation of a transmembrane proton po-tential and ATP synthesis [2].
CoMÿ Sÿ Sÿ CoB� 2e3 � 2H�!HSÿ CoB
�HSÿ CoM �2�
One physiological reductant in the latter reactionmay be reduced factor F420 (F420H2) which is formedduring methylotrophic growth and subsequently oxi-dized by the membrane-bound F420H2-dehydrogen-ase [3]. The ¢rst F420H2-dehydrogenase was puri¢edfrom the obligate methylotrophic Methanolobus tin-
0378-1097 / 99 / $19.00 ß 1999 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.PII: S 0 3 7 8 - 1 0 9 7 ( 9 8 ) 0 0 5 6 4 - 3
FEMSLE 8558 13-1-99
* Corresponding author. Tel. : +49 (33200) 88470;Fax: +49 (33200) 88470; E-mail: [email protected]
FEMS Microbiology Letters 170 (1999) 389^398

darius and shown to be composed of ¢ve non-iden-tical subunits of 45, 40, 21, 18 and 17 kDa [4]. Thesecond F420H2-dehydrogenase was puri¢ed fromMethanosarcina mazei Goë1 [5] and shown to transferelectrons to methanophenazine which donates elec-trons for CoM-S-S-CoB reduction as catalyzed byheterodisul¢de reductase [6]. The membrane-boundF420H2 :quinone oxidoreductase complex from thesulfate reducing archaeon Archaeoglobus fulgidus dif-fers from the above enzymes with respect to thenumber of subunits and the physiological electronacceptor [7]. In spite of the important role of thisenzyme in the energy metabolism of methylotrophicmethanogens, there has been no information avail-able on the genes encoding this protein. We thereforeinitiated a study to gain insight into the organizationof the genes coding for F420H2-dehydrogenase fromM. tindarius and to provide a basis for future inves-tigations on structure/function relationship.
2. Materials and methods
2.1. Bacterial strains, growth conditions, and enzymepuri¢cation
Methanolobus tindarius was grown under strictlyanaerobic conditions and F420H2-dehydrogenasewas puri¢ed as described previously [4]. Amino ter-minal sequence analysis was performed by B.Schmidt (Institute of Biochemistry, University ofGoëttingen) on the puri¢ed enzyme. For cloningand sequencing the following Escherichia coli strainswere used: JM83 [8], DH5K (Promega), NM522 andXL1 Blue (Stratagene), and DK8 [9]. E. coli strainswere cultured in Luria broth (LB) or on LB agar.Ampicillin (100 Wg ml31) was added when required.
2.2. DNA manipulations, sequencing and mRNAanalysis
Extractions of plasmid and chromosomal DNA,restriction enzyme digests, DNA ligations, and trans-formation into E. coli were done by standard meth-ods [10] unless otherwise stated. The following de-generate primers were radiolabeled with [Q-32P]ATPby polynucleotide kinase (Boehringer Mannheim,Mannheim, Germany) and subsequently used for
screening genomic DNA libraries of M. tindarius inpUC18 in order to detect the genes coding for the40- and 45-kDa subunit of the F420H2-dehydrogen-ase: DJW40, 5P-GT(ACGT)AT(ATC)GA(AG)CA-(CT)GA(CT)GT-3P ; DJW45, 5P-CA(AG)CA(CT)C-C(ACGT)ATGATGCC-3P ; DJW45-1, 5P-ATGG-A(CT)GA(AG)AC(ACGT)GT(ACGT)GG(ACGT)-CC-3P ; DJW45-2, 5P-GG(ACGT)CC(ACGT)CA(A-G)CA(CT)CC(ACGT)ATGATG-3P.
Clones hybridizing with one of the probes werefurther analyzed by DNA sequencing [11] using theUSB Sequenase DNA sequencing kit (AmershamBuchler, Braunschweig, Germany). Sequences wereanalyzed with the Genetics Computer Group pack-age Version 9.1, Madison, WI.
To clone the region upstream of ¡dB, PstI-cutchromosomal DNA fragments were circularized byligation, because hybridization of these fragmentswith the 0.14-kb HindIII/PstI fragment of pFFDH1indicated that a 1.24-kb PstI fragment extended froma PstI site upstream of the HindIII site of pFFDH1(uncloned part, 1.1 kb) to the PstI site in ¡dB(cloned part, 0.14 kb). Two oligonucleotide primerswere designed to bind to the known part of the cir-cularized PstI fragment in such a way that the 1.3-kbfragment could be ampli¢ed by PCR. The latter wasrestricted with HindIII and PstI and the resulting1.1-kb PstI/HindIII fragment was used to detect re-striction fragments upstream of the HindIII site.DNA labeling and detection was done with the Ran-dom primed DIG labeling and luminescent detectionkit (Boehringer Mannheim, Mannheim, Germany).
Primer extension was done essentially as described[12] using a 32P-labeled oligonucleotide and reversetranscriptase (Superscript, Gibco BRL). After phe-nol/chloroform extraction and ethanol precipitationthe product was analyzed in a polyacrylamide gel.The ¢rst step in RT-PCR was performed as de-scribed above for primer extension except that otherprimers were used. As a control, RT was omittedfrom the reaction mixture. The cDNA was subse-quently ampli¢ed using T£ DNA polymerase (Bio-zym Diagnostik, Hessisch-Oldendorf, Germany).
2.3. Heterologous expression in E. coli
For heterologous expression of the ¡d genes, E.coli DK8 transformed with the corresponding plas-
FEMSLE 8558 13-1-99
D.J. Westenberg et al. / FEMS Microbiology Letters 170 (1999) 389^398390

mids, was grown to an OD600 of 1, incubated for 20min with [35S]methionine at 37³C and the proteinswere analyzed by denaturing polyacrylamide gel elec-
trophoresis according to Schaëgger and von Jagow[13] followed by blotting and autoradiographic de-tection of the labeled proteins [9].
FEMSLE 8558 13-1-99
Fig. 2. Southern blot analysis (A) and PCR analysis (B,C) demonstrating contiguity of the cloned regions in pFFDH1 and pFFDH2. (A)Chromosomal DNA from M. tindarius was restricted with AvaI, EcoRV, and HindIII, separated by agarose gel electrophoresis, blottedand subsequently analyzed with DIG-labeled probes to either pFFDH1 or pFFDH2. (B) Agarose gel showing the ampli¢cation productsof PCRs obtained with the four pairs of primers indicated in C. The PCR products (PCR1^PCR4 in C) were analyzed in lanes 1^4;lane 5, DNA-size standards in bp. (C) The map shows the binding position of the primers used in B and gives the size of the expectedPCR products.
Fig. 1. Restriction map of the 7.7-kb EcoRI/HindIII fragment of Methanolobus tindarius containing the ¡dB gene coding for the 40-kDasubunit of the F420H2-dehydrogenase. pFFDH1 to pFFDH6 denote plasmids that contain the fragments indicated. Indicated are also thegenes identi¢ed upstream (¡dA) and downstream (¡dC, ¡dD) of ¡dB.
D.J. Westenberg et al. / FEMS Microbiology Letters 170 (1999) 389^398 391

FEMSLE 8558 13-1-99
Fig. 3.
D.J. Westenberg et al. / FEMS Microbiology Letters 170 (1999) 389^398392

FEMSLE 8558 13-1-99
Fig. 3. (continued)
D.J. Westenberg et al. / FEMS Microbiology Letters 170 (1999) 389^398 393

3. Results and discussion
3.1. Detection of ¡dB
The N-terminal amino acid sequence of the 45-and 40-kDa subunits of the puri¢ed F420H2-dehy-drogenase were determined as MDETVGPSE-MIVHLGPQHPMMPG and AHPKILEVIEHDV-TS(G)G(A)(D)V(S), respectively. Amino acids, givenin parentheses, are not certain. Degenerated oligonu-cleotides (DJW40, DJW45-1, DJW45-2) deducedfrom these sequences were used to probe chromoso-mal DNA from M. tindarius digested with EcoRI,HindIII, BamHI, or PstI. Only the probe for the40-kDa subunit (DJW40) hybridized to a 2.6-kbfragment in HindIII-cut chromosomal DNA. Thisfragment was subsequently cloned into pUC18 andthe resulting plasmid was designated pFFDH1 (Fig.1). Sequencing of the pFFDH1 insert revealed thepresence of three complete and a fourth truncated
open reading frame (ORF). The deduced N-terminalamino acid sequence of the ¢rst ORF was identicalto that of the 40-kDa subunit of F420H2-dehydrogen-ase except for the ¢rst amino acid (methionine)which was missing in the isolated polypeptide. Thisgene was designated ¡dB reminiscent of factorF420H2-dehydrogenase. The two small ORFs down-stream of ¡dB, which were later shown to be cotran-scribed with ¡dB, were tentatively designated ¡dCand ¡dD. The gene coding for the 45-kDa subunitwas absent from the cloned fragment.
3.2. Cloning of the upstream region of ¡dB
A 5-kb DNA fragment harboring the upstreamregion of ¡dB was subsequently cloned into pUC18giving rise to pFFDH2 (Fig. 1). Sequencing revealedthat the insert did not contain the sequence encodingthe N-terminal amino acid sequence of the puri¢ed45-kDa subunit, but suggested the presence of an
FEMSLE 8558 13-1-99
Fig. 3. Primer extension mapping of the 5P-end of the ¡dA mRNA. Sequencing ladders generated with the oligonucleotide used for theprimer extension were loaded next to the reaction mixture. The transcription start sites are indicated by arrows.
D.J. Westenberg et al. / FEMS Microbiology Letters 170 (1999) 389^398394
additional reading frame extending about 80 bp be-yond HindIII into pFFDH1 which was designated¡dA. The ¡dA-containing 1.1-kb HindIII/PstI frag-ment of pFFDH2 was subcloned into pUC 9 givingrise to plasmid pFFDH3 (Fig. 1). To prove that bothinserts represented one contiguous piece of DNA onthe M. tindarius chromosome four pairs of oligonu-cleotide primers for PCR ampli¢cation of chromoso-mal DNA were designed. Each pair consisted of oneprimer binding to the insert in pFFDH1 and anotherone binding to the insert in pFFDH2. If the 1.1-kbPstI/HindIII fragment of pFFDH2 and the 2.7-kbHindIII fragment of pFFDH1 were contiguous to
each other, we expected PCR products of about590, 960, 1100, and 800 bp. It is evident from Fig.2B that each reaction (2, 3 and 4) gave rise to onlyone PCR product of the expected size. Only the re-action 1 led to the formation of four bands. How-ever, the dominant band in the latter reaction hadthe expected size. Further support for the contiguityof the pFFDH1 and the pFFDH2 inserts came fromhybridization experiments in which chromosomalDNA digested with AvaI, EcoRV, and HindIII washybridized with the DIG-labeled inserts of pFFDH1and pFFDH2 (Fig. 2A). Both labels detected a 1.65-kb AvaI and a 3-kb EcoRV fragment suggesting that
FEMSLE 8558 13-1-99
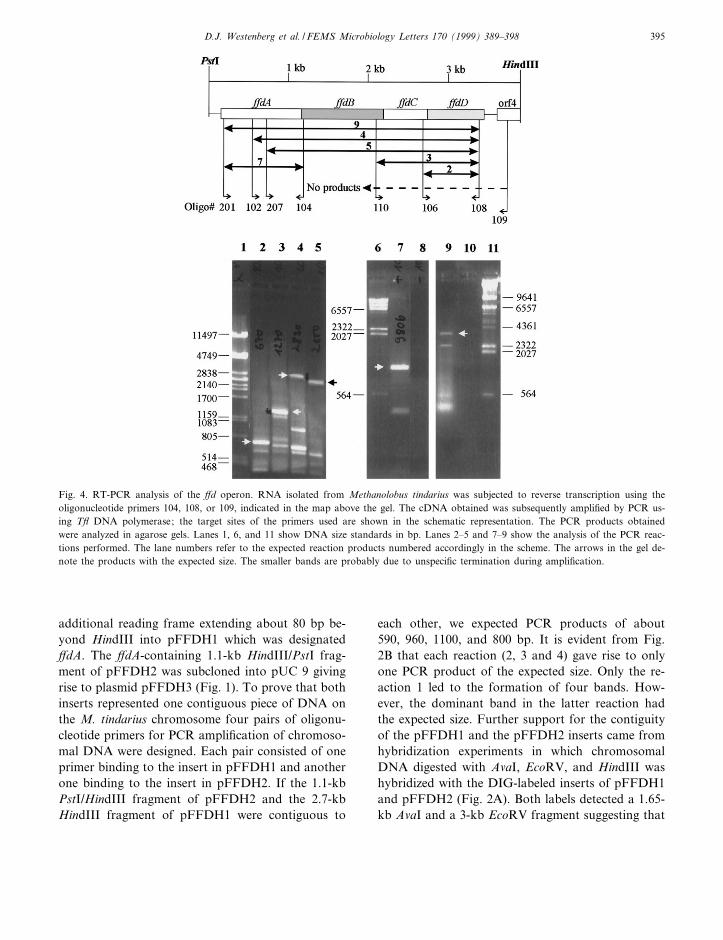
Fig. 4. RT-PCR analysis of the ¡d operon. RNA isolated from Methanolobus tindarius was subjected to reverse transcription using theoligonucleotide primers 104, 108, or 109, indicated in the map above the gel. The cDNA obtained was subsequently ampli¢ed by PCR us-ing T£ DNA polymerase; the target sites of the primers used are shown in the schematic representation. The PCR products obtainedwere analyzed in agarose gels. Lanes 1, 6, and 11 show DNA size standards in bp. Lanes 2^5 and 7^9 show the analysis of the PCR reac-tions performed. The lane numbers refer to the expected reaction products numbered accordingly in the scheme. The arrows in the gel de-note the products with the expected size. The smaller bands are probably due to unspeci¢c termination during ampli¢cation.
D.J. Westenberg et al. / FEMS Microbiology Letters 170 (1999) 389^398 395

these chromosomal DNA fragments were partlypresent on pFFDH1 and partly on pFFDH2. In or-der to obtain the whole region of interest in a singleclone, the 2.7-kb HindIII insert was excised frompFFDH1, and cloned into the HindIII site ofpFFDH3 resulting in pFFDH4 (Fig. 1). The DNAsequence containing the genes ¡dA, ¡dB, ¡dC, ¡dDhas been deposited in the EMBL database under theaccession number AJ011519. The calculated molecu-lar masses of the assumed gene products are 34.0,37.6, 20.0, and 23.7 kDa. However, only the ¡dBgene could be unambiguously assigned to one ofthe subunits of F420H2-dehydrogenase. Since thegene coding for the 45-kDa subunit was not presentin any of the pFFDH clones it has to be assumedthat it is located in a di¡erent region of the chromo-some.
3.3. De¢nition of the ¡d operon
Primer extension and RT-PCR were performed tode¢ne the start of transcription, the length of thetranscript and the putative genes as part of one op-eron. Possible archaeal promoter consensus sequen-ces were identi¢ed upstream of ¡dA and it wasshown by primer extension that there were two tran-scription start sites located 32 and 57 bp upstream ofthe putative ATG start codon (Fig. 3). A third some-what weaker transcription start site was 27 bp up-stream of this ATG codon. In order to ¢nd outwhich of the open reading frames belong to the pu-tative operon, total RNA from M. tindarius wastranscribed into cDNA with reverse transcriptase.The resulting cDNA served as a template for theampli¢cation by PCR of various parts of the puta-tive ¡d operon. Fig. 4 shows a representation of thecloned region, the sites of primer binding, and thesize of the RT-PCR fragments obtained. RT-PCRproducts of the expected size overlapped the clonedregion. In contrast, RT-PCR products overlappingthe ¡dD/ORF4 intergenic region could not be ob-tained, indicating that ¡dD and ORF4 were tran-scribed into di¡erent mRNAs. These results supportthe notion that ¡dA, ¡dB, ¡dC and ¡dD make upone operon, whereas ORF4 is independently tran-scribed. A possible transcription termination site,an inverted repeat forming a hairpin structure, islocated 94 bp downstream of the ¡dD stop codon.
3.4. Sequence comparison
The predicted amino acid sequences were used tosearch sequence databases. FfdA displays similaritywith the F420-dependent N5,N10-methylene-tetrahy-dromethanopterin reductase from other methano-gens including Methanobacterium thermoautotrophi-cum [14]. The ¢rst 90 amino acids of FfdB weresimilar to numerous ferredoxins, most notably poly-ferredoxins associated with the non-F420 reducinghydrogenase of M. thermoautotrophicum [15], sug-gesting the likely presence of at least two iron^sulfurcenters. FfdB shows also similarities to the L-subu-nits of F420-reducing hydrogenase and F420-reducingformate dehydrogenase from other methanogens andto a subunit of the F420 :quionone oxidoreductasefrom A. fulgidus [16]. FfdC reveals high sequencesimilarity with a putative gene product from Meth-anococcus jannaschii of unknown function. In con-trast to FfdA, FfdB, and FfdC, FfdD appears tobe very hydrophobic and is likely to be an integralmembrane protein. It could serve as membrane an-chor for the hydrophilic polypeptides of the enzyme.
FEMSLE 8558 13-1-99
Fig. 5. Expression of ¡d genes in Escherichia coli. E. coli DK8was transformed with plasmids carrying the genes indicated andminicells grown to an optical density of 1 (600 nm) were isolated.After incubation with [35S]methionine for 20 min, the cellularproteins were separated by SDS-PAGE, blotted and the blotsautoradiographed. The position of molecular size standards areindicated in kDa on the right. Protein extracts of cells trans-formed with pUC18 (lane 1), pFFDH1 (¡dB/C/D preceded bythe lac promoter, lane 2), pFFDH1 (¡dB/C/D in reverse orienta-tion to the lac promoter, lane 3), pFFDH6 (¡dB, in reverse ori-entation to the lac promoter, lane 4), and pFFDH5 (¡dA in re-verse orientation to the lac promoter, lane 5). The bandscorresponding to polypeptides encoded by genes of the ¡d oper-on are indicated by ¢lled arrowheads. Hollow arrowheads pointat pre-L-lactamase (33.5 kDa) and L-lactamase (29 kDa), respec-tively.
D.J. Westenberg et al. / FEMS Microbiology Letters 170 (1999) 389^398396

3.5. Heterologous expression of ¡dA, ¡dB, ¡dC, and¡dD in mini cells
In order to ¢nd experimental support for the no-tion that the putative genes do indeed code for pro-teins, the genes were cloned into pUC18 (Fig. 1) andexpressed in E. coli mini cells. Cells transformed withpFFDH5 (¡dA) formed a polypeptide of 34 kDa,independent of the orientation of the insert (Fig.5). However, the expression was stronger when thegene was preceded by the lac promoter (not shown).When E. coli DK8 was transformed with pFFDH6(¡dB) or pFFDH7 (¡dBCD/ORF4) a polypeptide ofabout 37 kDa, presumably FfdB, was formed. InpFFDH7, two additional weak bands of about 20and 23 kDa appeared. Since these molecular massesare in good agreement with the putative gene prod-ucts of ¡dC and ¡dB it appears likely that they wereindeed heterologously expressed. The orientation ofthe inserts in£uenced the intensity of all three bandsonly slightly, suggesting that there is a sequence infront of ¡dB that allows transcription by E. colipolymerase. The two additional bands of about14 and 14.2 kDa detected in cells transformed withpFFDH7 are assumed to result from the truncatedORF4 which is preceded by a potential methanogenpromoter.
Whereas ¡dB could be clearly assigned to the 40-kDa subunit of F420H2-dehydrogenase and shown tobe part of an operon, it is presently not yet knownwhether FfdA, FfdC, and FfdD are identical to anyother subunit of the enzyme. It is clear, however,that FfdA is not identical to the 45-kDa subunit ofthe puri¢ed enzyme and it can be excluded that thegene encoding this polypeptide is present in thecloned DNA region. It can be speculated thatFfdC and FfdD correspond to two of the three smallsubunits of the puri¢ed F420H2-dehydrogenase.However, further research is required to answer theseopen questions.
Acknowledgments
The ¢nancial support by the `Deutsche For-schungsgemeinschaft' and `Fonds der ChemischenIndustrie' is gratefully acknowledged.
References
[1] Rouvieére, P. and Wolfe, R.S. (1988) Novel biochemistry ofmethanogenesis. J. Biol. Chem. 263, 7913^7916.
[2] Blaut, M., Muëller, V. and Gottschalk, G. (1992) Energetics ofmethanogenesis studied in vesicular systems. J. Bioenerg. Bio-membr. 24, 529^546.
[3] Deppenmeier, U., Blaut, M., Mahlmann, A. and Gottschalk,G. (1990) Reduced coenzyme F420 heterodisul¢de oxidoreduc-tase, a proton-translocating redox system in methanogenicbacteria. Proc. Natl. Acad. Sci. USA 87, 9449^9453.
[4] Haase, P., Deppenmeier, U., Blaut, M. and Gottschalk, G.(1992) Puri¢cation and characterization of F420H2 dehydro-genase from Methanolobus tindarius. Eur. J. Biochem. 203,527^531.
[5] Abken, H.J. and Deppenmeier, U. (1997) Puri¢cation andproperties of an F420H2 dehydrogenase from Methanosarcinamazei Goë1. FEMS Microbiol. Lett. 154, 231^237.
[6] Baumer, S., Murakami, E., Brodersen, J., Gottschalk, G.,Ragsdale, S.W. and Deppenmeier, U. (1998) TheF420H2 :heterodisul¢de oxidoreductase system from Methano-sarcina species. 2-Hydroxyphenazine mediates electron trans-fer from F420H2 dehydrogenase to heterodisul¢de reductase.FEBS Lett. 29, 295^298.
[7] Kunow, J., Linder, D., Stetter, K.O. and Thauer, R.K. (1994)F420H2 :quinone oxidoreductase from Archaeoglobus fulgidus.Characterization of a membrane-bound multisubunit complexcontaining FAD and iron^sulfur clusters. Eur. J. Biochem.223, 503^511.
[8] Vieira, J. and Messing, J. (1982) The pUC plasmid, anM13mp7-derived system for insertion mutagenesis and se-quencing with synthetic universal primers. Gene 19, 259^268.
[9] Ruppert, C., Wimmers, S., Lemker, T. and Muëller, V. (1998)The A1A0 ATPase from Methanosarcina mazei : cloning of the5P end of the aha operon encoding the membrane domain andexpression of the proteolipid in a membrane-bound form inEscherichia coli. J. Bacteriol. 180, 3448^3452.
[10] Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) MolecularCloning: A Laboratory Manual. Cold Spring Harbor Labo-ratory Press, Cold Spring Harbor, NY.
[11] Sanger, F., Nicklen, S. and Coulson, A.R. (1977) DNA se-quencing with chain-terminating inhibitors. Proc. Natl. Acad.Sci. USA 74, 5463^5467.
[12] Kellmann, J.-W., Piechersky, E. and Piechulla, B. (1990)Analysis of the diurnal expression patterns of the tomatochlorophyll a/b binding protein genes. In£uences of lightand characterization of the gene family. Photochem. Photo-biol. 52, 35^41.
[13] Schaëgger, H. and von Jagow, G. (1987) Tricine-sodium do-decyl sulfate-polyacrylamide gel electrophoresis for the sepa-ration of proteins in the range from 1 to 100 kDa. Anal.Biochem. 166, 368^379.
[14] Vaupel, M. and Thauer, R.K. (1995) Coenzyme F420-depend-ent N5,N10-methylenetetrahydromethanopterin reductase(Mer) from Methanobacterium thermoautotrophicum strainMarburg. Cloning, sequencing, transcriptional analysis, and
FEMSLE 8558 13-1-99
D.J. Westenberg et al. / FEMS Microbiology Letters 170 (1999) 389^398 397

functional expression in Escherichia coli of the mer gene. Eur.J. Biochem. 231, 773^778.
[15] Reeve, J.N., Beckler, G.S., Cram, D.S., Hamilton, P.T.,Brown, J.W. and Krzycki, J.A. (1989) A hydrogenase-linkedgene in Methanobacterium thermoautotrophicum strain vHencodes a polyferredoxin. Proc. Natl. Acad. Sci. USA 86,3031^3035.
[16] Klenk, H.P., Clayton, R.A., Tomb, J.F., White, O., Nelson,K.E., Ketchum, K.A., Dodson, R.J., Gwinn, M., Hickey,
E.K., Peterson, J.D., Richardson, D.L., Kerlavage, A.R.,Graham, D.E., Kyrpides, N.C., Fleischmann, R.D., Quacken-bush, J., Lee, N.H., Sutton, G.G., Gill, S., Kirkness, E.F.,Dougherty, B.A., McKenney, K., Adams, M.D., Loftus, B.and Venter, J.C. (1997) The complete genome sequence ofthe hyperthermophilic, sulphate-reducing archaeon Archaeo-globus fulgidus (published erratum appears in Nature 1998Jul 2;394(6688):101). Nature 390, 364^370.
FEMSLE 8558 13-1-99
D.J. Westenberg et al. / FEMS Microbiology Letters 170 (1999) 389^398398
Copyright © 2022 FDOKUMEN




















