
Quantitation of serum lactate dehydrogenase-5 with monoclonal antibodies
11
Clinica Chimicu Acta, 161 (1986) 315-325 Elsevier 315 CCA 03658 Quantitation of serum lactate dehydrogenase-5 with monoclonal antibodies * Hemant C. Vaidya, David N. Dietzler and Jack H. Ladenson Wushington Universi& School of Medicine, Division of Luhoratoty Medicine, Departments of Puthologv and Medicine, St. Louis, MO (USA) (Received 20 June 1986; revision received 15 August 1986: accepted after revision 29 August 1986) Kerp words: Monoclonal antibodies; Lactate dehydrogenase-5: Isoenzymes Spleen cells from BALB/cJ mice which had been immunized with human lactate dehydrogenase-1 (LDH-1) were fused with SP2/0-Ag14. Two hybridomas were produced which recognized the antigen. Competitive RIA revealed that one anti- body (‘Smit-LDH’) recognized the H subunit of LDH while the other (‘Hem-LDH’) recognized both II and M subunits of LDH. With the use of these antibodies we developed an assay for LDHS activity in which serum is incubated for 30 min at room temperature with the two antibodies (‘Smit-LDH’ and ‘Hem-LDH’ in the ratio 64 : 1.3, pg/ml) immobilized on latex beads to extract LDH-1 through LDH-4. After centrifugation, the LDH activity of the supernatant is measured and repre- sents LDH-5 activity. Latex beads coated with bovine serum albumin were used as control. The LDH-5 activity as determined by our assay correlated well (r = 0.98) with the values obtained by an electrophoresis method. There was no interference due to LDH-1 through LDH3 up to 3000 U/l and LDH-4 up to 350 U/l. Serum samples with total LDH activity above 1000 U/l were appropriately diluted in order to avoid interference by LDH-4. Use of these monoclonal antibodies allows precise, rapid and direct measurement of LDH-5 activity in serum. Introduction Lactate dehydrogenase (LDH, L-lactate: NAD oxidoreductase, EC X1.1.27) exists in serum in five isoenzymic forms, each consisting of a tetramer of two * The antibodies will be made available to qualified investigators on request. Correspondence to: H.C. Vaidya, Washington University School of Medicine, Division of Laboratory Medicine, Departments of Pathology and Medicine, Box 8118, St. Louis, MO 63110, USA. ooO9-8981/86/$03.50 Q 1986 Elsevier Science Publishers B.V. (Biomedical Division)
Transcript of Quantitation of serum lactate dehydrogenase-5 with monoclonal antibodies

Clinica Chimicu Acta, 161 (1986) 315-325
Elsevier 315
CCA 03658
Quantitation of serum lactate dehydrogenase-5 with monoclonal antibodies *
Hemant C. Vaidya, David N. Dietzler and Jack H. Ladenson Wushington Universi& School of Medicine, Division of Luhoratoty Medicine, Departments of Puthologv
and Medicine, St. Louis, MO (USA)
(Received 20 June 1986; revision received 15 August 1986: accepted after revision 29 August 1986)
Kerp words: Monoclonal antibodies; Lactate dehydrogenase-5: Isoenzymes
Spleen cells from BALB/cJ mice which had been immunized with human lactate dehydrogenase-1 (LDH-1) were fused with SP2/0-Ag14. Two hybridomas were produced which recognized the antigen. Competitive RIA revealed that one anti- body (‘Smit-LDH’) recognized the H subunit of LDH while the other (‘Hem-LDH’) recognized both II and M subunits of LDH. With the use of these antibodies we developed an assay for LDHS activity in which serum is incubated for 30 min at room temperature with the two antibodies (‘Smit-LDH’ and ‘Hem-LDH’ in the ratio 64 : 1.3, pg/ml) immobilized on latex beads to extract LDH-1 through LDH-4. After centrifugation, the LDH activity of the supernatant is measured and repre- sents LDH-5 activity. Latex beads coated with bovine serum albumin were used as control. The LDH-5 activity as determined by our assay correlated well (r = 0.98) with the values obtained by an electrophoresis method. There was no interference due to LDH-1 through LDH3 up to 3000 U/l and LDH-4 up to 350 U/l. Serum samples with total LDH activity above 1000 U/l were appropriately diluted in order to avoid interference by LDH-4. Use of these monoclonal antibodies allows precise, rapid and direct measurement of LDH-5 activity in serum.
Introduction
Lactate dehydrogenase (LDH, L-lactate: NAD oxidoreductase, EC X1.1.27) exists in serum in five isoenzymic forms, each consisting of a tetramer of two
* The antibodies will be made available to qualified investigators on request. Correspondence to: H.C. Vaidya, Washington University School of Medicine, Division of Laboratory
Medicine, Departments of Pathology and Medicine, Box 8118, St. Louis, MO 63110, USA.
ooO9-8981/86/$03.50 Q 1986 Elsevier Science Publishers B.V. (Biomedical Division)

316
subunits H or M. The distribution of the isoenzymes varies amongst the tissues. LDH-1 (H4 isoenzyme) is predominant in heart and erythrocytes while LDH-5 (M4
isoenzyme) is predominant in liver and skeletal muscle. Determination of the relative distribution of these isoenzymes in serum is useful in diagnosis of diseases
involving damage to tissues [l-3]. Currently, electrophoresis is the most commonly used procedure for separating and quantitating LDH isoenzymes [4].
In the last two decades, immunochemical properties of LDH isoenzymes have been studied by several investigators [5-g]. A quantitative immunoassay for LDH-1 with use of anti-M subunit specific antiserum was reported by Usategui-Gomez et al
[9] and a diagnostic kit using these polyclonal antibodies is distributed by Roche Diagnostics. However, a quantitative immunoassay for LDH-5 has not been re-
ported. In the present communication, we describe the development of a quantita- tive immunoassay to measure LDH-5 activity in human serum with use of a combination of two different monoclonal antibodies.
Materials and methods
Purification of lactate dehydrogenase (LDH) Human LDH-1 was purified from heart while LDH4 and LDH-5 were purified from human liver. The purification steps included: NH,SO, fractionation of tissue homogenate, separation of isoen- zymes by ion-exchange chromatography [lo], and affinity purification on S’AMP Sepharose4B column [ll]. Human LDH-1, LDH-2, LDH-3, and LDH-5 were also
obtained from Sigma (St. Louis, MO, USA). All the LDH isoenzymes appeared as a single band when checked for purity by sodium dodecyl sulfate polyacrylamide or agarose gel electrophoresis. The specific activity of the purified proteins ranged
from 240-430 (U/mg protein).
Immunization Eight-week-old female BALB/cJ mice, H-2d haplotype (Jackson Laboratories), were injected intraperitoneally with 25 pg of human LDH-1 emulsi- fied in an equal volume of complete Freund’s adjuvant (Sigma). Four and eight weeks later, the same amount of antigen in incomplete Freund’s adjuvant and phosphate buffered saline (PBS, 50 mmol of NaPO,, and 150 mm01 of NaCl/l pH 7.2) was similarly administered. The immunized mice were bled 10 days after the third injection to determine the serum antibody titer. Mice with high antibody titers were intraperitoneally injected four days before the fusion with 25 pg of LDH-1 in PBS. This fourth injection was performed at least 4 weeks after the third injection.
Fusion technique Spleens were aseptically removed from the immunized mice. Spleen cells (log) were fused with SP2/0-Ag14 myeloma cells (107) in the presence of polyethylene glycol (M, 1500), essentially as described by Kijhler and Milstein [12], at the Hybridoma Center of the Washington University School o.f Medicine, St. Louis, MO, USA.

317
Screening of hybridomas Hybridomas secreting antibodies to human LDH-1 were screened by solid phase radioimmunoassay as described earlier [13]. The
antigen was radiolabeled with use of lactoperoxidase-glucose oxidase enzymobeads
(BioRad, NJ, USA) essentially by following the manufacturer’s instructions. The labeled LDH was purified by gel filtration. Hybridomas producing antibodies to LDH-1 were cloned in soft agar and stored under liquid nitrogen in DMEM
(Dulbecco’s modified Eagle’s medium) containing 100 ml of dimethyl sulfoxide and 300 ml of horse serum per liter.
Purification of monoclonal antibodies from ascites Actively growing hybrid cells
(106) were injected i.p. into pristane (2,6,10,14-tetramethyl-pentadecane, Sigma) primed BALB/cJ mice. Ascites fluid produced by each cell line was collected, pooled, and stored at - 20°C after removing the cell debris by centrifugation. Monoclonal antibody from the ascites fluid was purified on a protein ‘A’ affinity
column (MAP@, BioRad). Fractions obtained from the column containing the monoclonal antibody were pooled and concentrated with an ultrafiltration cell (Amicon, USA). The purity of the antibodies was checked by agarose gel electro- phoresis.
Electrophoresis Ascites fluid, purified monoclonal antibody or LDH isoenzymes were electrophoresed on agarose gel (Corning agarose film No. 470100) with diethylbarbital buffer (PHAB, Corning No. 470180) for 30 min at 90 V. For staining
the proteins, the gels were fixed and stained in a 200 ml mixture of methanol, acetic acid and water (40 : 10 : 50 by volume) containing 0.125% Coomassie Brilliant Blue. The background was destained in the same mixture without the dye. LDH isoen- zymes were localized on the agarose gel with use of a LDH staining kit (Sigma).
Determination of isotype Mouse monoclonal antibodies were isotyped by Ouch- terlony double-diffusion technique with use of discs and mouse isotype specific
antisera from Miles Scientific, Naperville, IL, USA.
Determination of specificity The specificity of the monoclonal antibodies was determined by competitive RIA. This assay was similar to the RIA used’ for
screening hybridoma supematants except that the radiolabeled human LDH-1 was allowed to bind to the immobilized antibody in the presence of various concentra- tions of unlabelled human LDH isoenzymes.
We immobilized 200 ng of goat anti-mouse IgG antibodies (100 ~1 of 2 mg/l solution in 100 mm01 sodium borate buffer containing 150 mmol NaCl/l, pH 8.5) onto 96 well round-bottom microtiter plates by incubation at 37°C for 2 h. The
plates were washed with Tween-saline (0.5 ml of polyoxyethylene [20] sorbitan monolaurate (Tween-20), 8.77 g of NaCl and 0.02 g of NaN,/l). We then added 100 ,ul of affinity purified monoclonal antibody solution (2 mg/l in PBS containing 10 g/l BSA) and incubated and washed the plates as above. For competitive RIA, we added 100 ~1 of a mixture of labelled LDH-1 (100000 counts/mm) and O-1000 ng

318
of competing human LDH isoenzyme per well. The mixture was incubated at 37 o C for 2 h. The plates were then washed, dried, and the radioactivity of the bound
radiolabeled LDH-1 was measured in a Packard gamma-counter.
Clinical samples and standards Serum samples submitted to the Barnes Hospital Clinical Laboratory were assayed within 3-4 h. Total LDH activity and LDH-5 activity were assayed as described below. LDH isoenz mes present in serum
samples were separated and stained using the Parago T electrophoresis system r&
(Beckman, USA). The stained gels were scanned by a Gelman automatic computing densitometer (model No. ACD-18) to determine LDH-5 as a percentage of total
LDH activity.
Immobilization of antibodies on latex beads The two monoclonal antibodies were immobilized by passive adsorption on 0.8 pm diamter latex beads (LB-8 Sigma). The beads were suspended to 10 g/l in coating buffer (100 mmol/l sodium phosphate, pH 6.0) containing 200 mg of ‘Smit-LDH’ and 4 mg of ‘Hem-LDH’ per liter, and were incubated overnight at 4°C. The beads were washed with PBS and suspended in PBS containing bovine serum albumin so that 40 ~1 of the suspension contained 16 pg of ‘Smit-LDH’ and 0.32 pg of ‘Hem-LDH’. More than 98% of the antibodies were immobilized on the beads under these conditions as determined by protein measurement of the supematant after the incubation period.
LDH-5 assay In the LDH-5 assay, LDH-1 through LDH4 isoenzymes are extracted by the monoclonal antibodies immobilized on latex beads and the residual
LDH activity in the supernatant is measured kinetically. The assay was performed as follows. Two hundred microliters of serum sample,
standard solution or control sample was dispensed into two 1.5ml Eppendorf microfuge tubes. The standard solutions (50-900 U/l) were prepared by ap- propriately diluting purified human LDH-5 in PBS containing 10 g/l BSA. LDH- isotrol, used as a control, was obtained from Sigma. Forty microliters of monoclonal antibody-coated latex beads were added to one tube while BSA-coated latex beads were added to the second tube and the volume of both the tubes was brought up to 250 ~1 with PBS. After incubation for 30 min at room temperature the beads were pelleted and LDH-activity of the supematant was measured. The LDH activity measured from the first tube represented the LDH-5 activity while that from the second tube represented total LDH activity. The LDH-5 activity in a serum sample was determined by interpolation from the standard curve and LDH-5 activity was also calculated as a percentage of total LDH activity. Serum samples with total LDH activity greater than 1000 U/l were reassayed after appropria e dilution in PBS. LDH activity at 37OC was measured with the use of a Flexige r& ’ centrifugal analyzer and “Gemini’ LDH reagent (Electronucleonics). The ingredients of the LDH reagent were 75.0 mmol Tris, 5.0 mmol NAD and 50.0 mmol L-lithium lactate per liter of distilled water, pH 9.0. LDH activity measurement was linear up to 1300 U/l.
319
RtSUltS
Serum of immunized BALB/cJ female mice, H-2d haplotype, had a high anti- body titer to human LDH-1. Fusion of the spleen cells from immunized BALB/cJ mice with myeloma cell line SP2/0-Ag14 generated two hyb~domas which pro- duced antibodies to human LDH-1. The cloned hybridomas were injected in- traperitoneally into pristane primed BALB/cJ female mice to produce ascites fluid and the antibodies were purified by protein ‘A’ affinity chromatography.
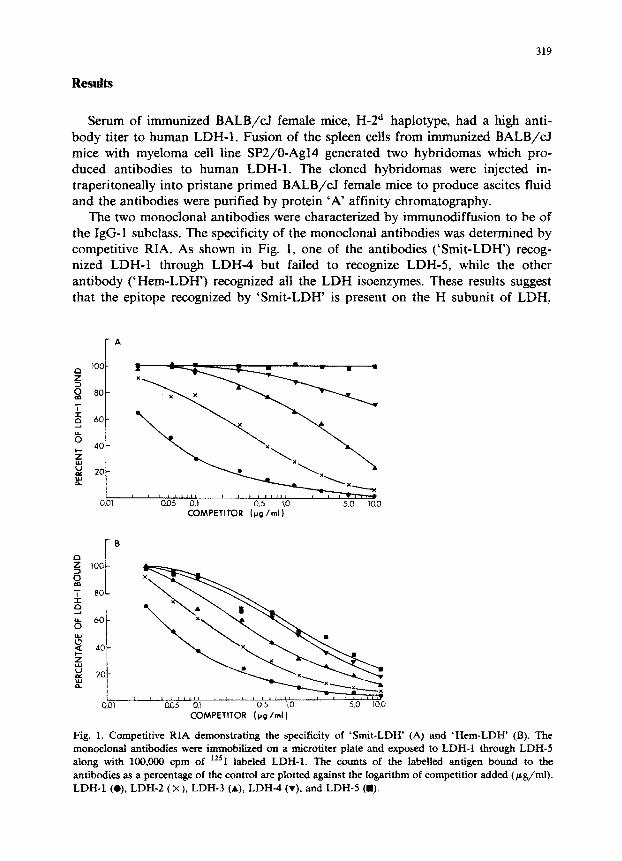
The two monoclonal antibodies were characterized by immun~ffusion to be of the IgG-1 subclass. The specificity of the monoclonal antibodies was determined by competitive RIA. As shown in Fig. 1, one of the antibodies (‘Smit-LDH”) recog- nized LDH-1 through LDH-4 but failed to recognize LDH-5, while the other antibody them-LD~) recognized all the LDH isoenzymes. These results suggest that the epitope recognized by ‘Smit-LDH’ is present on the H subunit of LDH,
COMPETITOR (pg /ml)
rs
5 ii
loo-
T so-
G 8 60-
x s 40-
B 20-
:
COMPETITOR (PQ /ml )
Fig. 1. Competitive RIA demonstrating the specificity of ‘Smit-LDH’ (A) and ‘Hem-LDH’ (B). The monoclonal antibodies were immobilized on a microtiter plate and exposed to LDH-1 through LDH-5 along with 100,000 cpm of %*‘I labeled LDH-1. The counts of the labelled antigen bound to the antibodies as a percentage of the control are plotted against the logarithm of competitior added (pg/ml). LDH-1 (o), LDH-2 (x), LDH-3 (A), LDH-4 (v). and LDH-5 (m).
320
while the epitope for ‘Hem-LDH’ is present on both H and M subunits of LDH. The difference in the recognition of subunits by ‘Hem-LDH’ may be due to partial homology between the epitope on M and H subunits or the two subunits may have the same epitope in different conformation.
Assay development We developed a quantitative assay for LDH-5 in which LDH-5 activity is
measured after extraction of the other isoenzymes with use of the two monoclonal antibodies immobilized on latex beads. We optimized the assay with respect to the concentration and ratio of the two monoclonal antibodies, and with respect to time and temperature of incubation.
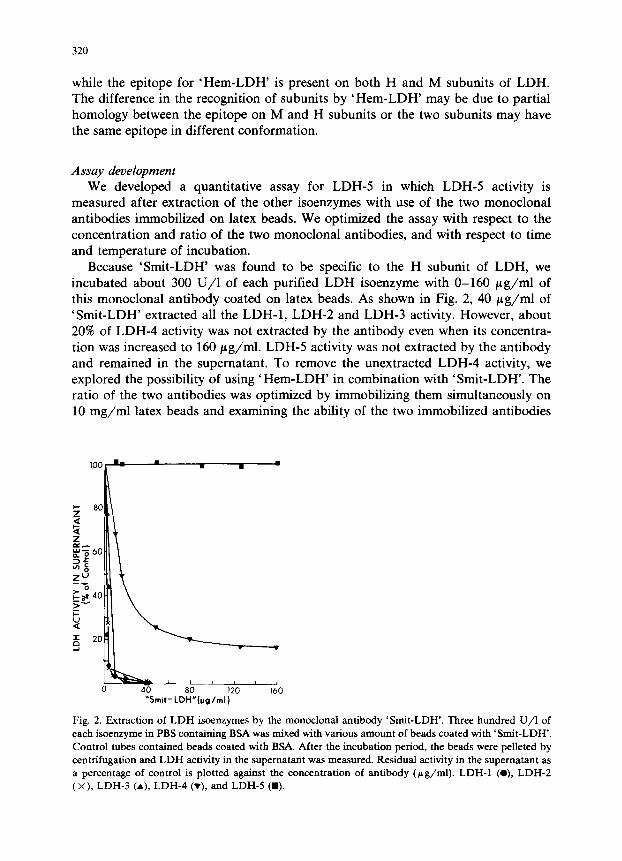
Because ‘Smit-LDH’ was found to be specific to the H subunit of LDH, we incubated about 300 U/l of each purified LDH isoenzyme with O-160 fig/ml of this monoclonal antibody coated on latex beads. As shown in Fig. 2, 40 pg/ml of ‘Smit-LDH’ extracted all the LDH-1, LDH-2 and LDH-3 activity. However, about 20% of LDH4 activity was not extracted by the antibody even when its concentra- tion was increased to 160 pg/ml. LDH-5 activity was not extracted by the antibody and remained in the supematant. To remove the unextracted LDH4 activity, we explored the possibility of using ‘Hem-LDH’ in combination with ‘Smit-LDH’. The ratio of the two antibodies was optimized by immobilizing them simultaneously on 10 mg/ml latex beads and examining the ability of the two immobilized antibodies
“Smit-LDH”(pg/ml)
Fig. 2. Extraction of LDH &enzymes by the monoclonal antibody ‘Smit-LDH’. Three hundred U/l of
each isoenzyme in PBS containing BSA was mixed with various amount of beads coated with ‘Smit-LDH’.
Control tubes contained beads coated with BSA. After the incubation period, the beads were pelleted by
centrifugation and LDH activity in the supematant was measured. Residual activity in the supematant as
a percentage of control is plotted against the concentration of antibody @g/ml). LDH-1 (O), LDH-2 (x), LDH-3 (A), LDH-4 (v), and LDH-5 (a).
321
TABLE I
Extraction of LDH-4 and LDH-5 isoenzymes by various concentrations of ‘Smit-LDH’ and ‘Hem-LDH
immobilized on latex beads
‘&it-LDH’ ‘Hem-LDH’ @g/ml)
@g/ml) 0 0.16 0.32 0.64 1.3
Residual activity (% of control)
LDH-4 LDH-5 LDH-4 LDH-5 LDH-4 LDH-5 LDH-4 LDH-5 LDH-4 LDH-5
0 100 100 94 98 84 96 73 93 54 89
16 49 102 48 98 43 98 37 94 30 92
32 33 100 30 102 25 100 18 93 15 90
64 22 101 19 102 12 100 9 96 0 89
100 19 98 15 101 10 98 8 97 0 92
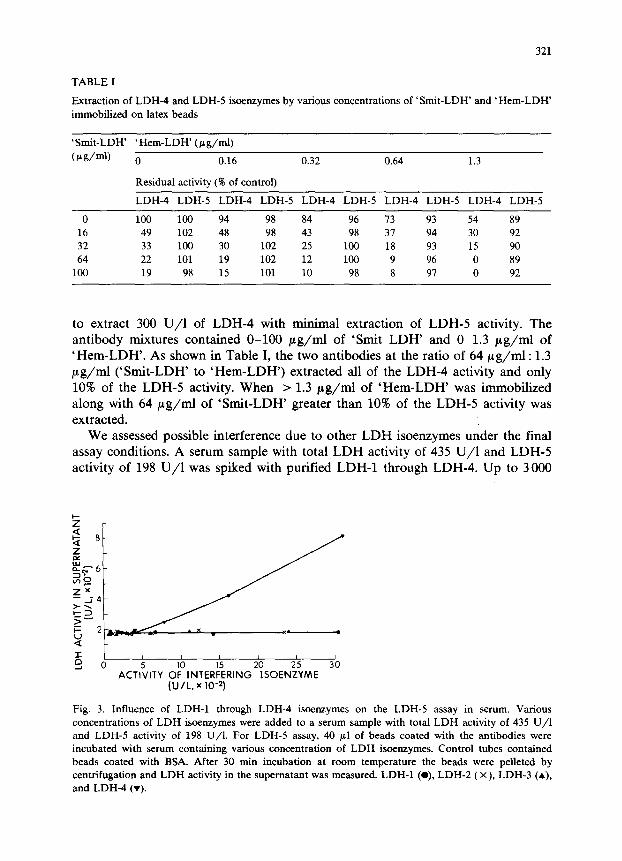
to extract 300 U/l of LDH4 with minimal extraction of LDH-5 activity. The antibody mixtures contained O-100 pg/rnI of ‘Smit LDH’ and O-l.3 pg/mI of ‘Hem-LDH’. As shown in Table I, the two antibodies at the ratio of 64 pg/mI : 1.3 pg/mI (‘Smit-LDH’ to ‘Hem-LDH’) extracted all of the LDH4 activity and only 10% of the LDH-5 activity. When > 1.3 pg/ml of ‘Hem-LDH’ was immobilized along with 64 pg/ml of ‘Smit-LDH’ greater than 10% of the LDH-5 activity was extracted.
We assessed possible interference due to other LDH isoenzymes under the final assay conditions. A serum sample with total LDH activity of 435 U/l and LDH-5 activity of 198 U/l was spiked with purified LDH-1 through LDH-4. Up to 3000
Fig. 3. Influence of LDH-1 through LDH-4 isoenzymes on the LDH-5 assay in serum. Various
concentrations of LDH isoenzymes were added to a serum sample with total LDH activity of 435 U/l
and LDH-5 activity of 198 U/l. For LDH-5 assay, 40 gl of beads coated with the antibodies were
incubated with serum containing various concentration of LDH isoenzymes. Control tubes contained
beads coated with BSA. After 30 min incubation at room temperature the beads were pelleted by
centrifugation and LDH activity in the supematant was measured. LDH-1 (O), LDH-2 (X), LDH-3 (A),
and LDH-4 (v).
322
U/l of LDH-1, LDH-2, or LDH-3 did not alter the LDH-5 value obtained in the supematant (180 U/l). However, when LDH-4 exceeded 350 U/l the apparent LDH-5 value was spuriously elevated (Fig. 3). It appeared therefore, that LDH-4 activity above 350 U/l in serum samples interfered in the assay. On evaluation of the LDH data of 1570 serum samples submitted to the clinical chemistry laboratory it was found that LDH-4 activity of 16 samples (1.1%) exceeded 350 U/l (350-l 500
U/l) with their total LDH activity ranging from 2 180-8980 U/l. However, the LDH-4 activity of these 16 samples was never > 23% of the total LDH activity.
Thus, if such serum samples are diluted before LDH-5 assay so that their total LDH activity is 1000 U/l or less, LDH-4 activity will be lower than the interference level of 350 U/l. The total LDH activity cut-off value of 1000 U/l was selected for convenience because it is close to the upper limit of measurement of LDH activity by Flexigem centrifugal analyzer.
The influence of incubation period and temperature were assessed. The mono- clonal antibodies extracted the LDH-1 through LDH-4 isoenzymes from serum within 20 min at both room temperature and 37°C. We did not incubate at 4-8°C
because LDH-5 is known to lose activity at lower temperatures [14]. Ten percent of LDH-5 activity was consistently extracted by the antibodies from
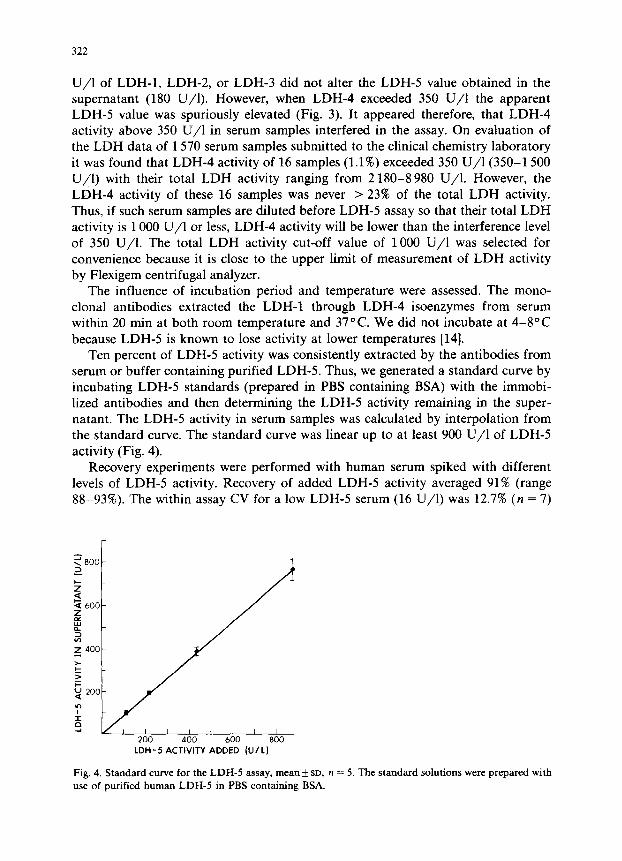
serum or buffer containing purified LDH-5. Thus, we generated a standard curve by incubating LDH-5 standards (prepared in PBS containing BSA) with the immobi- lized antibodies and then determining the LDH-5 activity remaining in the super-
natant. The LDH-5 activity in serum samples was calculated by interpolation from the standard curve. The standard curve was linear up to at least 900 U/l of LDH-5
activity (Fig. 4). Recovery experiments were performed with human serum spiked with different
levels of LDH-5 activity. Recovery of added LDH-5 activity averaged 91% (range 88-93%). The within assay CV for a low LDH-5 serum (16 U/l) was 12.7% (n = 7)
Fig. 4. Standard curve for the LDH-5 assay, mean f SD, n = 5. The standard solutions were prepared with use of purified human LDH-5 in PBS containing BSA.
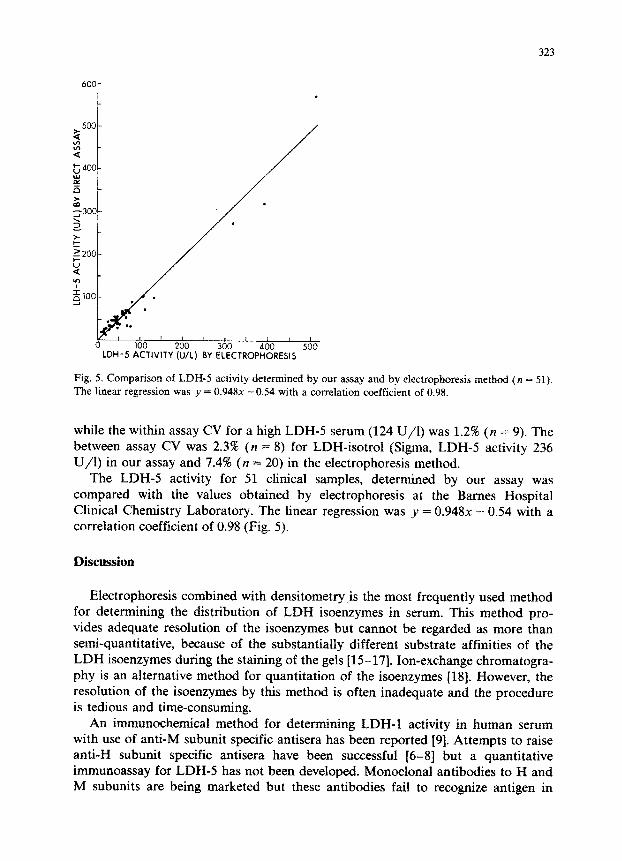
323
Fig. 5. Comparison of LDH-5 activity determined by our assay and by electrophoresis method (n = 51). The linear regression was y = 0.948x -0.54 with a correlation coefficient of 0.98.
while the within assay CV for a high LDH-5 serum (124 U/l) was 1.2% (n = 9). The between assay CV was 2.3% (n = 8) for LDH-isotrol (Sigma, LDH-5 activity 236 U/I) in our assay and 7.4% (n = 20) in the electrophoresis method.
The LDH-5 activity for 51 clinical samples, determined by our assay was compared with the values obtained by electrophoresis at the Barnes Hospital Clinical Chemistry Laboratory. The linear regression was 1’ = 0.948~ - 0.54 with a correlation coefficient of 0.98 (Fig. 5).
Discussion
Electrophoresis combined with densitometry is the most frequently used method for determining the distribution of LDH isoenzymes in serum. This method pro- vides adequate resolution of the isoenzymes but cannot be regarded as more than set-quantitative, because of the subst~tially different substrate affinities of the LDH isoenzymes during the staining of the gels [15-171. Ion-exchange chromatogra- phy is an alternative method for quantitation of the &enzymes [18]. However, the resolution of the isoenzymes by this method is often inadequate and the procedure is tedious and time-consuming.
An immun~he~cai method for dete~ning LDH-1 activity in human serum with use of anti-M subunit specific antisera has been reported [9]. Attempts to raise anti-H subunit specific antisera have been successful [6-81 but a quantitative immunoassay for LDH-5 has not been developed. Monoclonal antibodies to H and M subunits are being marketed but these antibodies fail to recognize antigen in

324
solution [19], a property which precludes their use for immunoassay. The mono- clonal antibodies developed in our laboratory recognize H subunit (‘&it-LDH’) or H and M subunits (‘Hem-LDH’) of LDH in solution as well as on solid phase. In the development of the LDH-5 assay we could not use ‘Smit-LDH’ alone because it
failed to extract about 20% LDH-4 from solution. The most likely reason for this is that the epitope on the H subunit of the unextractable LDH-4 molecules may have been masked by the M subunits. We could extract all of the LDH-4 activity by the use of a combination of our two monoclonal antibodies. With this combination,
only 10% of the LDH-5 activity was extracted under optimal conditions, and this percentage was consistent over a broad range of LDH-5 activities. The complete extraction of LDH-1 through LDH-4 was greatly facilitated by the use of latex particles which could be uniformly suspended in solution and pelleted by centrifu- gation. We believe this is the first assay with use of monoclonal antibodies which allow direct measurement of LDH-5 activity in serum.
Determination of LDH-5 activity by our assay compared well with the values
obtained by electrophoresis and densitometry. Our assay could be further simplified with the use of monoclonal antibodies which would extract all H subunit containing isoenzymes, if such antibodies are possible to obtain.
The procedure described here is simple, precise and less time consuming than
electrophoresis. Our procedure may be of value in rapid clinical diagnosis of liver
and skeletal muscle disorders [1,2] and in the detection of certain human carcinomas [20-241 in which serum LDH-5 activity is elevated.
Acknowledgements
This work was partially supported by NIH training grant 2 T32 ES07066-09 and Monsanto/Washington University Hybridoma Contract. We thank Dr. Curtis Parvin for computer support and Ms. Yvonne Maynard for discussion.
References
1 Vesell ES, Beam AG. Isozymes of lactic dehydrogenase in human tissues. J Clin Invest 1961; 40:
586-591.
2 Wilkinson JH. The principles and practice of diagnostic enzymology. London: Edward Arnold/UK,
1976.
3 Wilkinson JH. Isoenzymes. Philadelphia: J.P. Lippincott Co./PA USA, 1970: 134-203.
4 Rosalki SB. Standardisation of isoenzyme assays with special reference to lactate dehydrogenase
isoenzyme electrophoresis. Clin Biochem 1974, 7: 29-40.
5 Nisselbaum JS, Bodansky 0. Immunochemical studies of human lactic dehydrogenase. Ann NY Acad Sci 1963; 103: 930-937.
6 Markert CL, Appella E. Immunochemical properties of lactate dehydrogenase isozymes. Ann NY
Acad Sci 1963; 103: 915-929.
7 Burd JF, Usategui-Gomez M. Immunochemical studies on lactate dehydrogenase. Biochim Biophys
Acta 1973; 310: 238-247.
8 Burd JF, Usategui-Gomez M, Fernandez de Castro A, Mhatre NS, Yeager FM. Immunochemical studies on lactate dehydrogenase. Preparation and characterization of anti-M lactate dehydrogenase
antiserum. Development of an immunochemical assay for lactate dehydrogenase isoenzymes in human serum. Clin Chim Acta 1973; 46: 205-216.

9
10
11
12
13
14
15
16
17
18
19
325
Usategui-Gomez M, Wicks RW, Warshaw M. Immunochemical determination of the heart isoenzyme
of lactate dehydrogenase (LDH-1) in human serum. Clin Chem 1979; 25: 729-734.
Fritz PJ, Morrison WJ, White EL, Vesell ES. Comparative study of methods for quantitation of
lactate dehydrogenase isozymes. Anal Biochem 1970; 36: 443-453.
Lindberg M, Larsson PO, Misbach K. A new immobilized NAD+ analogue, its application in affinity
chromatography and as a functioning coenzyme. Eur J Biochem 1973; 40: 187-193.
Kohler G, Milstein C. Continuous culture of fused cells secreting antibody of predefined specificity.
Nature (London) 1975; 256: 495-497.
Vaidya HC, Maynard Y, Dietzler DN, Ladenson JH. Direct measurement of creatine kinase-MB
activity in serum after extraction with a monoclonal antibody specific to the MB isoenzyme. Clin
Chem 1986; 32: 657-663.
Kreutzer HH, Fennis WHS. Lactic dehydrogenase isoenzymes in blood serum after storage at
different temperatures. Clin Chim Acta 1964; 9: 64-71.
Rosa& SB, Wilkinson JH. Reduction of a-ketobutyrate by human serum. Nature (London) 1960;
188: 1110-1111.
Plummer DT, Elliott BA, Cooke KB, Wilkinson JH. Organ specificity and lactate dehydrogenase
activity 1. The relative activities with pyruvate and a-oxobutyrate of electrophoretically separated
fractions. Biochem J 1963; 87: 416-422.
Buhl SN, Jackson KY, Lubinski R, Vanderlinde RE. Optimal conditions for assaying human lactate
dehydrogenase by the lactate-to-pyruvate reaction: Arhenius relationships for lactate dehydrogenase
isoenzymes 1 and 5. Clin Chem 1977; 23: 1289-1295.
Hsu MY, Kohler MM, Barolia L, Bonder RJL. Separation of five isoenzymes of serum lactate
dehydrogenase by discontinuous gradient elution from a miniature ion exchange column. Clin Chem
1979; 25: 1453-1458.
Vaidya HC, Dietzler DN, Ladenson JH. Inadequacy of traditional ELISA for screening hybridoma
supematants for murine monoclonal antibodies. Hybridoma 1985; 4: 271-276.
20 Wood DC, Varela V, Palmquist M, Weber F. Serum lactate dehydrogenase and isoenzyme changes in
clinical cancer. J Surg Oncol 1973; 5: 251-257.
21 Kolaric K, Roguljic A, Petrinovic R. Comparative investigations of some enzymatic parameters and
liver scanning in the early detection of the malignant liver process. Clin Chim Acta 1975; 60:
109-111.
22 Carda-Abella P, Perez-Cuadrado S, Lara-baruque S, Gil-Grande L, Nunez Puertas A. LDH isoen-
zyme patterns in tumors, polyps, and uninvolved mucosa of human cancerous colon. Cancer 1982; 49: 80-83.
23 Vergnon JM, Guidollet J, Gateau 0, Rip011 JP, Collet P, Louisot P, Brune J. Lactic dehydrogenase
isoenzyme electrophoretic patterns in the diagnosis of pleural effusion. Cancer 1984; 54: 507-511.
24 Stefanini M. Enzymes, isozymes, and enzyme variants in the diagnosis of cancer. Cancer 1985; 55: 1931-1936.




















