
The evolution of squamosal shape in ceratopsid dinosaurs (Dinosauria, Ornithischia)
10
This article was downloaded by: [Society of Vertebrate Paleontology ] On: 12 November 2013, At: 07:17 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Journal of Vertebrate Paleontology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/ujvp20 The evolution of squamosal shape in ceratopsid dinosaurs (Dinosauria, Ornithischia) Leonardo Maiorino a b , Andrew A. Farke c , Paolo Piras a b , Michael J. Ryan d , Kevin M. Terris e & Tassos Kotsakis a b a Department of Sciences , Roma Tre University , Largo S. Leonardo Murialdo 1, 00146 , Rome , Italy b Center for Evolutionary Ecology , Largo S. Leonardo Murialdo 1, 00146 , Rome , Italy c Raymond M. Alf Museum of Paleontology , 1175 West Baseline Road, Claremont , California , 91711-2199 , U.S.A. d Department of Vertebrate Paleontology , Cleveland Museum of Natural History , 1 Wade Oval Drive, University Circle, Cleveland , Ohio , 44106 , U.S.A. e Montana State University , Bozeman , Montana , 59717 , U.S.A. Published online: 12 Nov 2013. To cite this article: Leonardo Maiorino , Andrew A. Farke , Paolo Piras , Michael J. Ryan , Kevin M. Terris & Tassos Kotsakis (2013) The evolution of squamosal shape in ceratopsid dinosaurs (Dinosauria, Ornithischia), Journal of Vertebrate Paleontology, 33:6, 1385-1393 To link to this article: http://dx.doi.org/10.1080/02724634.2013.779922 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
Transcript of The evolution of squamosal shape in ceratopsid dinosaurs (Dinosauria, Ornithischia)

This article was downloaded by: [Society of Vertebrate Paleontology ]On: 12 November 2013, At: 07:17Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Journal of Vertebrate PaleontologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/ujvp20
The evolution of squamosal shape in ceratopsiddinosaurs (Dinosauria, Ornithischia)Leonardo Maiorino a b , Andrew A. Farke c , Paolo Piras a b , Michael J. Ryan d , Kevin M.Terris e & Tassos Kotsakis a ba Department of Sciences , Roma Tre University , Largo S. Leonardo Murialdo 1, 00146 ,Rome , Italyb Center for Evolutionary Ecology , Largo S. Leonardo Murialdo 1, 00146 , Rome , Italyc Raymond M. Alf Museum of Paleontology , 1175 West Baseline Road, Claremont ,California , 91711-2199 , U.S.A.d Department of Vertebrate Paleontology , Cleveland Museum of Natural History , 1 WadeOval Drive, University Circle, Cleveland , Ohio , 44106 , U.S.A.e Montana State University , Bozeman , Montana , 59717 , U.S.A.Published online: 12 Nov 2013.
To cite this article: Leonardo Maiorino , Andrew A. Farke , Paolo Piras , Michael J. Ryan , Kevin M. Terris & TassosKotsakis (2013) The evolution of squamosal shape in ceratopsid dinosaurs (Dinosauria, Ornithischia), Journal of VertebratePaleontology, 33:6, 1385-1393
To link to this article: http://dx.doi.org/10.1080/02724634.2013.779922
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Journal of Vertebrate Paleontology 33(6):1385–1393, November 2013© 2013 by the Society of Vertebrate Paleontology
ARTICLE
THE EVOLUTION OF SQUAMOSAL SHAPE IN CERATOPSID DINOSAURS(DINOSAURIA, ORNITHISCHIA)
LEONARDO MAIORINO,*,1,2 ANDREW A. FARKE,3 PAOLO PIRAS,1,2 MICHAEL J. RYAN,4 KEVIN M. TERRIS,5
and TASSOS KOTSAKIS1,2
1Department of Sciences, Roma Tre University, Largo S. Leonardo Murialdo 1, 00146, Rome, Italy, [email protected];[email protected];
2Center for Evolutionary Ecology, Largo S. Leonardo Murialdo 1, 00146, Rome, Italy, [email protected];3Raymond M. Alf Museum of Paleontology, 1175 West Baseline Road, Claremont, California 91711-2199, U.S.A., [email protected];4Department of Vertebrate Paleontology, Cleveland Museum of Natural History, 1 Wade Oval Drive, University Circle, Cleveland,
Ohio 44106, U.S.A., [email protected];5Montana State University, Bozeman, Montana 59717, U.S.A., [email protected]
ABSTRACT—Ceratopsidae represents one of the last and best-known radiations of non-avian dinosaurs. Interspecific varia-tion is well documented qualitatively with linear measurements, but little has been done to quantify shape differences in thefrill that may indicate functional or evolutionary signals. In order to investigate shape change in the squamosal across Chas-mosaurinae and Centrosaurinae, we applied geometric morphometrics to the outline of the squamosal for 155 specimens rep-resenting 27 ceratopsid species and Protoceratops spp. A Mantel test suggests that the shape of ceratopsid squamosals is highlyconstrained by phylogeny. MANOVA, ANOVA, and their phylogenetic versions suggest that an evolutionarily significant al-lometric signal exists between the two clades, but not within clades. Principal component analysis indicates that centrosaurineshave a uniform squamosal shape, with the exceptions of Spinops and Diabloceratops. Even when accounting for phylogeny,the relationship between squamosal shape and size is significant. Mapping shape onto the phylogeny, we estimated ancestralshapes at nodes. The transition from the non-ceratopsid to ceratopsid condition is characterized by a squamosal with a widerangle between the infratemporal process and the caudoventral margin, and a more dorsoventrally elongated and caudally ex-panded blade. From root to tips, centrosaurine squamosals were found to be conservative, but exhibit a slight dorsoventralexpansion and a narrow angle between the infratemporal process and the caudoventral margin in more derived taxa. Chas-mosaurines, compared with centrosaurines, show a derived morphology, with a trend towards a blade that is strongly expandeddorsoventrally and with a narrower angle between the infratemporal process and the caudoventral margin.
SUPPLEMENTAL DATA—Supplemental materials are available for this article for free at www.tandfonline.com/UJVP
INTRODUCTION
The primary purpose of comparative biology is to understandthe evolution of phenotypic form and infer the phylogenetichistory of shape changes (Klingenberg and Gidaszewski, 2010).The well-documented Late Cretaceous ceratopsids, or horneddinosaurs, had perhaps one of the highest rates of speciation innon-avian dinosaurs, documented by the rapid variation in theirdiagnostic cranial ornamentation, including elaborate horns andfrill morphologies (Dodson et al., 2004; Sampson and Loewen,2010). In a time span of 50–60 million years, ceratopsians diversi-fied from small (<2 m body length) bipedal species to huge (>9 mlong) quadrupedal species. Similarly, skulls of some of the mostderived taxa exceeded 2.5 m in length (Dodson, 1993).
Two morphologically distinct clades, Chasmosaurinae andCentrosaurinae, constitute Ceratopsidae. Recent discoveries ofnew taxa and revision of historically described taxa have furtherdemonstrated the wide morphological disparity of ceratopsiddinosaur skulls, illuminating the clade’s complex evolutionaryhistory and paleobiogeography (Ryan, 2007; Wu et al., 2007;Sampson et al., 2010; Farke et al., 2011; Longrich, 2011; Ryan
*Corresponding author.
et al., 2012). Although some postcranial differences distinguishthe clades within Ceratopsidae, the most notable and numerousvariations reside within the skull (e.g., Lehman, 1990; Dodsonet al., 2004; Sampson and Loewen, 2010; Maidment and Barrett,2011). Early ceratopsids have elongated postorbital horncores,which are secondarily lost in most centrosaurines and some chas-mosaurines. Chasmosaurines are generally characterized by anelongated parietosquamosal frill in concert with an elaborate setof circumnarial depressions and other features. Centrosaurines,in turn, have a simpler circumnarial region (resembling theplesiomorphic condition of Zuniceratops) and a short but oftenheavily ornamented parietosquamosal frill.
The paired squamosals of the frill have figured prominently indistinguishing the two clades. The squamosal of chasmosaurinesis characterized as elongate and triangular, extending to the pos-terior margin of the frill. Some minor morphological differencesdistinguish taxa, such as the number of marginal ossifications orthe profile of the margin (e.g., Sampson et al., 2010; Sullivan andLucas, 2010). By contrast, the squamosals of centrosaurines havebeen described as remarkably uniform. The free blade of thesquamosal is hemicircular in profile, and the overall shape of thebone is quite broad in comparison with that in chasmosaurines.With the exception of Diabloceratops and Avaceratops, thesquamosal of centrosaurines has not been considered to have
1385
Dow
nloa
ded
by [
Soci
ety
of V
erte
brat
e Pa
leon
tolo
gy ]
at 0
7:17
12
Nov
embe
r 20
13
1386 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 33, NO. 6, 2013
any utility in systematics (e.g., Ryan, 2007). Diabloceratopshas a square squamosal body that resembles the condition ofnon-ceratopsids such as Protoceratops (Kirkland and DeBlieux,2010). Avaceratops is unique among centrosaurines in that ithas an unstepped medial margin of the squamosal (similar tochasmosaurines), rather than a stepped condition in which thesquamosal forms part of the posterior edge of the supratemporalfenestra (Dodson, 1986). Protoceratops is notable in being theonly non-ceratopsid ceratopsian with an overall expanded frilland a squamosal shape resembling that of some ceratopsids (Youand Dodson, 2004); the condition may be similar in taxa such asZuniceratops and Bagaceratops, but the squamosals are poorlyknown for adults of these taxa.
New material, including extensive collections from bonebeds(e.g., Ryan et al., 2001; Ryan, 2007; Wu et al., 2007; Sampson et al.,2010; Farke et al., 2011), have dramatically increased the num-ber of ceratopsian squamosals available for study, with some taxanow represented by dozens of specimens inferred to span ontoge-netic stages ranging from small juveniles to large, mature adults.This large data set now allows us to conduct an in-depth, quanti-tative study of cranial variation in ceratopsids by focusing on thesquamosal. Three specific questions are addressed here: (1) whatis the full range of variation in the ceratopsid squamosal; (2) canthe squamosal be used to distinguish ceratopsid taxa; and (3) whatevolutionary patterns, if any, characterize the squamosal withinceratopsids?
Geometric morphometrics has been used previously to studycranial or postcranial shape changes of ceratopsian dinosaurs.Chapman (1990) analyzed ontogenetic changes in skulls ofBagaceratops rozhdestvenskyi and also explored the sexualdimorphism in skulls and lower jaws of Protoceratops andrewsi bymeans of RFTRA (Resistant-Fit Theta-Rho-Analysis). Dodson(1993) also used RFTRA to investigate the overall evolutionarychanges in skulls across Ceratopsia. Chinnery (2004) used linearmeasurements and shape data to investigate the taxonomic dif-ferences in the ceratopsian postcranial skeleton and to comparegrowth changes with evolutionary changes in postcranial skeletalanatomy.
Despite these contributions, most published studies did notquantitatively analyze morphological variation in Ceratopsidae atthe species level. Here we use geometric morphometrics to inves-tigate phenotypic (shape and size) differences in the squamosalacross ceratopsids, including Protoceratops spp. as the ‘outgroup’to allow us to investigate the transition from the non-ceratopsid tothe ceratopsid condition.
Institutional Abbreviations—AMNH, American Museum ofNatural History, New York, New York, U.S.A.; NHMUK, NaturalHistory Museum, London, U.K.; ROM, Royal Ontario Museum,Toronto, Ontario, Canada; TMP, Royal Tyrrell Museum of Pale-ontology, Drumheller, Alberta, Canada; UMNH, Utah Museumof Natural History, Salt Lake City, Utah, U.S.A.
MATERIALS AND METHODS
Materials
A sample of 155 squamosals, including juvenile, subadult, andadult-sized specimens (juveniles are restricted to the centrosaurinesample), from 11 species of centrosaurines (including one newunnamed taxon, UMNH VP 19469) and 16 species of chas-mosaurines was assembled. The basal neoceratopsians, Protocer-atops andrewsi and P. hellenikorhinus, were included in the sampleto root the analyses and because of the large number of availablesquamosals. Details about the specimens (including repository andidentification) are provided in Supplementary Data 1. A CANON400D camera was used to collect pictures for the sample. Althoughsquamosals can be described as flat bones, we followed the proto-cols of Marcus et al. (2000) and Mullin and Taylor (2002) to mini-mize parallax and measurement error on the photographs.
Methods
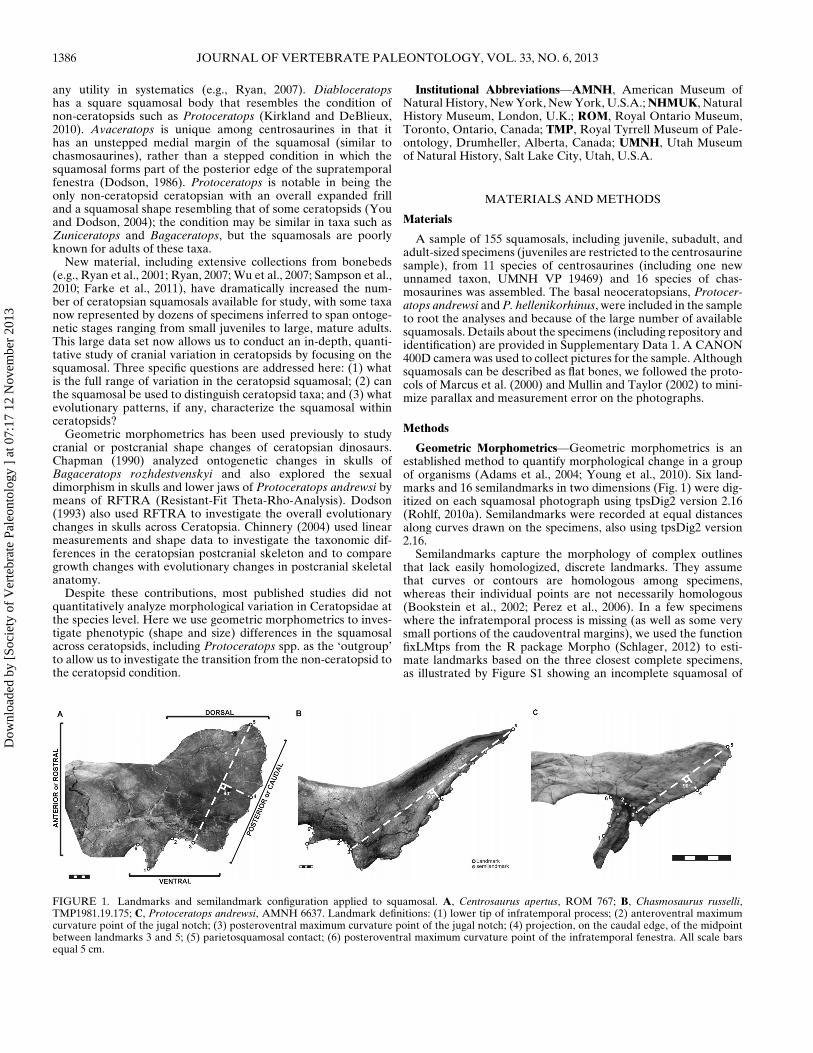
Geometric Morphometrics—Geometric morphometrics is anestablished method to quantify morphological change in a groupof organisms (Adams et al., 2004; Young et al., 2010). Six land-marks and 16 semilandmarks in two dimensions (Fig. 1) were dig-itized on each squamosal photograph using tpsDig2 version 2.16(Rohlf, 2010a). Semilandmarks were recorded at equal distancesalong curves drawn on the specimens, also using tpsDig2 version2.16.
Semilandmarks capture the morphology of complex outlinesthat lack easily homologized, discrete landmarks. They assumethat curves or contours are homologous among specimens,whereas their individual points are not necessarily homologous(Bookstein et al., 2002; Perez et al., 2006). In a few specimenswhere the infratemporal process is missing (as well as some verysmall portions of the caudoventral margins), we used the functionfixLMtps from the R package Morpho (Schlager, 2012) to esti-mate landmarks based on the three closest complete specimens,as illustrated by Figure S1 showing an incomplete squamosal of
FIGURE 1. Landmarks and semilandmark configuration applied to squamosal. A, Centrosaurus apertus, ROM 767; B, Chasmosaurus russelli,TMP1981.19.175; C, Protoceratops andrewsi, AMNH 6637. Landmark definitions: (1) lower tip of infratemporal process; (2) anteroventral maximumcurvature point of the jugal notch; (3) posteroventral maximum curvature point of the jugal notch; (4) projection, on the caudal edge, of the midpointbetween landmarks 3 and 5; (5) parietosquamosal contact; (6) posteroventral maximum curvature point of the infratemporal fenestra. All scale barsequal 5 cm.
Dow
nloa
ded
by [
Soci
ety
of V
erte
brat
e Pa
leon
tolo
gy ]
at 0
7:17
12
Nov
embe
r 20
13

MAIORINO ET AL.—CERATOPSID SQUAMOSAL EVOLUTION 1387
FIGURE 2. Phylogenetic tree with all species considered in this study. The black contour depicts squamosal mean shape of terminal taxa. See Supple-mentary Data 1 for details about topology and branch length calibration.
Spinops sternbergorum (NHMUK R16309) with the reconstructedinfratemporal process. We anticipate here that the infratemporalprocess of the entire sample is not a crucial structure in explainingthe global shape change (see Discussion), because the first princi-pal component explains variation that does not affect particularlythe infratemporal process.
Once the outlines were digitized, shape was analyzed usinggeneralized Procrustes analysis (GPA; Bookstein, 1991), imple-mented in tpsRelw version 1.49 (Rohlf, 2010b). GPA standardizesspecimen landmark configurations for comparison by scaling,aligning, and rotating each configuration with the unit centroidsize (CS = the square root of sum of squared differences betweenlandmarks from their centroid; Bookstein, 1986). We used CSin our analyses to represent individual size. Rotation is initiatedrelative to a reference landmark configuration (usually the firstsampled specimen). After rotation has been accomplished, a meanshape is generated. The rotation is repeated iteratively throughthe entire sample, resulting in a progressively more representativemean shape. This mean-shape rotation ensures that specimenoutlines are maximally comparable by reducing dissimilaritiesbetween specimens due to rotation (Rohlf and Slice, 1990; Piraset al., 2010). A separate sliding semilandmark file was preparedfor tpsRelw to distinguish landmarks from semilandmarks inorder to optimize GPA. In this way, tpsRelw version 1.49 (Rohlf,2010b) performs the relative warp analysis using sliding-landmarkinformation during computation.
After GPA, Procrustes coordinates were averaged by species,and principal component analysis (PCA) was performed on theseProcrustes coordinates to identify orthogonal axes of maximalvariation. This is a standard procedure in geometric morpho-
metric studies (Adams et al., 2004; Claude, 2008; Piras et al.,2010).
Phylogeny
We built a synthetic phylogenetic tree including all species stud-ied here (Fig. 2) using the software Mesquite 2.75 (Maddison andMaddison, 2011). The phylogeny is based on published cladisticanalyses on taxa and on their stratigraphic range (SupplementaryData 1). Branch lengths were calibrated in millions of years (Ma)based on the fossil record (also in Supplementary Data 1).
Linear Models and Comparative Methods
Phenotypic Differences between Clades—Using the pooleddata set, we first performed a Mantel test to assess the relationshipbetween the phylogenetic covariance matrix and Procrustes dis-tance matrix, using the mantel.test function in the ‘ape’ R package(Paradis et al., 2004). We then tested for phenotypic differences(shape and size) between Chasmosaurinae and Centrosaurinae.Analysis of variance (ANOVA; using size as dependent vari-able) and MANOVA (using shape data as dependent variable)were conducted to analysis for linear model and morphospaceoccupation within Ceratopsidae. Protoceratops was excluded inthese tests because it only has two species. Both ANOVA andMANOVA were accompanied by their phylogenetic versionsusing the phy.anova and phy.manova functions available in the‘geiger’ R package (Harmon et al., 2008). A univariate (on size)analysis of differences in variance using Levene’s test and a mul-tivariate (on shape data) dispersion analysis using the betadisper
Dow
nloa
ded
by [
Soci
ety
of V
erte
brat
e Pa
leon
tolo
gy ]
at 0
7:17
12
Nov
embe
r 20
13
1388 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 33, NO. 6, 2013
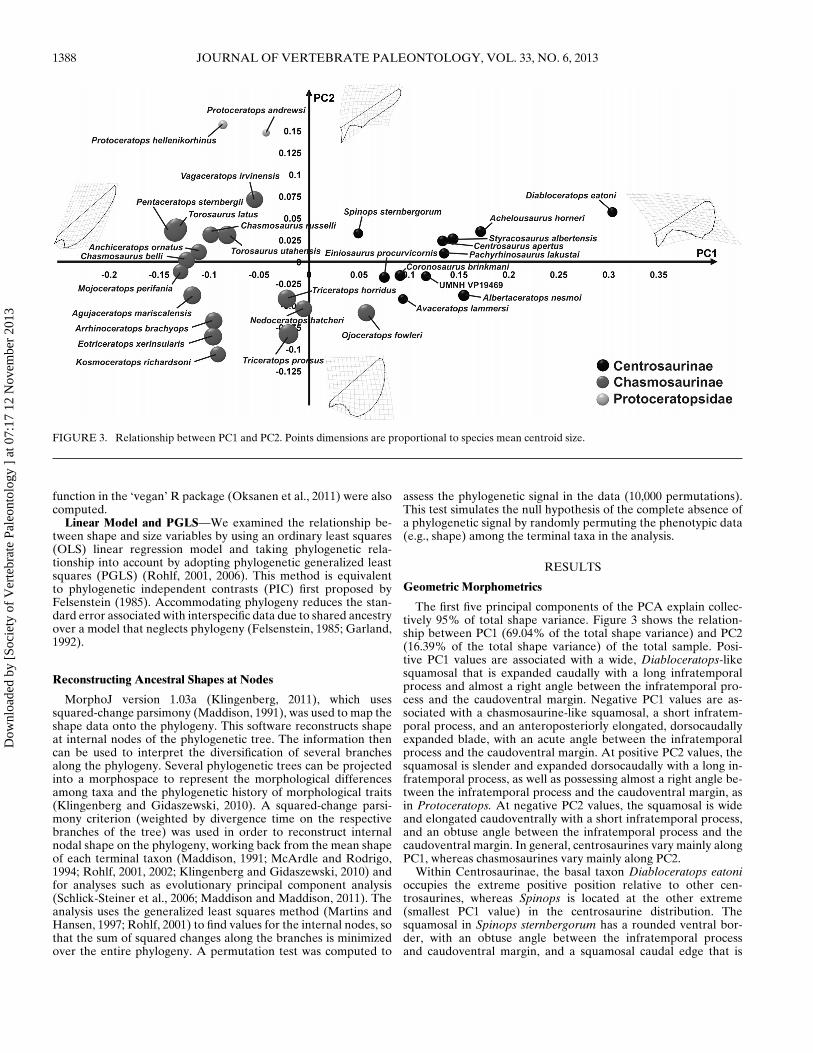
FIGURE 3. Relationship between PC1 and PC2. Points dimensions are proportional to species mean centroid size.
function in the ‘vegan’ R package (Oksanen et al., 2011) were alsocomputed.
Linear Model and PGLS—We examined the relationship be-tween shape and size variables by using an ordinary least squares(OLS) linear regression model and taking phylogenetic rela-tionship into account by adopting phylogenetic generalized leastsquares (PGLS) (Rohlf, 2001, 2006). This method is equivalentto phylogenetic independent contrasts (PIC) first proposed byFelsenstein (1985). Accommodating phylogeny reduces the stan-dard error associated with interspecific data due to shared ancestryover a model that neglects phylogeny (Felsenstein, 1985; Garland,1992).
Reconstructing Ancestral Shapes at Nodes
MorphoJ version 1.03a (Klingenberg, 2011), which usessquared-change parsimony (Maddison, 1991), was used to map theshape data onto the phylogeny. This software reconstructs shapeat internal nodes of the phylogenetic tree. The information thencan be used to interpret the diversification of several branchesalong the phylogeny. Several phylogenetic trees can be projectedinto a morphospace to represent the morphological differencesamong taxa and the phylogenetic history of morphological traits(Klingenberg and Gidaszewski, 2010). A squared-change parsi-mony criterion (weighted by divergence time on the respectivebranches of the tree) was used in order to reconstruct internalnodal shape on the phylogeny, working back from the mean shapeof each terminal taxon (Maddison, 1991; McArdle and Rodrigo,1994; Rohlf, 2001, 2002; Klingenberg and Gidaszewski, 2010) andfor analyses such as evolutionary principal component analysis(Schlick-Steiner et al., 2006; Maddison and Maddison, 2011). Theanalysis uses the generalized least squares method (Martins andHansen, 1997; Rohlf, 2001) to find values for the internal nodes, sothat the sum of squared changes along the branches is minimizedover the entire phylogeny. A permutation test was computed to
assess the phylogenetic signal in the data (10,000 permutations).This test simulates the null hypothesis of the complete absence ofa phylogenetic signal by randomly permuting the phenotypic data(e.g., shape) among the terminal taxa in the analysis.
RESULTS
Geometric Morphometrics
The first five principal components of the PCA explain collec-tively 95% of total shape variance. Figure 3 shows the relation-ship between PC1 (69.04% of the total shape variance) and PC2(16.39% of the total shape variance) of the total sample. Posi-tive PC1 values are associated with a wide, Diabloceratops-likesquamosal that is expanded caudally with a long infratemporalprocess and almost a right angle between the infratemporal pro-cess and the caudoventral margin. Negative PC1 values are as-sociated with a chasmosaurine-like squamosal, a short infratem-poral process, and an anteroposteriorly elongated, dorsocaudallyexpanded blade, with an acute angle between the infratemporalprocess and the caudoventral margin. At positive PC2 values, thesquamosal is slender and expanded dorsocaudally with a long in-fratemporal process, as well as possessing almost a right angle be-tween the infratemporal process and the caudoventral margin, asin Protoceratops. At negative PC2 values, the squamosal is wideand elongated caudoventrally with a short infratemporal process,and an obtuse angle between the infratemporal process and thecaudoventral margin. In general, centrosaurines vary mainly alongPC1, whereas chasmosaurines vary mainly along PC2.
Within Centrosaurinae, the basal taxon Diabloceratops eatonioccupies the extreme positive position relative to other cen-trosaurines, whereas Spinops is located at the other extreme(smallest PC1 value) in the centrosaurine distribution. Thesquamosal in Spinops sternbergorum has a rounded ventral bor-der, with an obtuse angle between the infratemporal processand caudoventral margin, and a squamosal caudal edge that is
Dow
nloa
ded
by [
Soci
ety
of V
erte
brat
e Pa
leon
tolo
gy ]
at 0
7:17
12
Nov
embe
r 20
13
MAIORINO ET AL.—CERATOPSID SQUAMOSAL EVOLUTION 1389
dorsoventrally elongated. Part of the infratemporal process wasreconstructed in Spinops, and we feel that our reconstruction isreasonable, but nonetheless it does not explain the morphology ofthe squamosal margin. The squamosal of Diabloceratops eatoni isroughly rectangular and caudally expanded. The tip of infratem-poral process reaches a much more ventral position than the cau-doventral margin, and the angle between these two structures isobtuse.
Within chasmosaurines, there is not a clear differentiationamong taxa. Their morphology mainly varies along PC2, exceptfor Ojoceratops and Triceratops + Nedoceratops. Ojoceratops fow-leri, placed at negative PC2 values, and low positive PC1 val-ues, has a rounded squamosal that is dorsoventrally and caudallyexpanded, with an obtuse angle between the infratemporal pro-cess and the caudoventral margin. Triceratops + Nedoceratops areplaced at negative PC2 values, but fall in the middle of the PC1distribution.
Protoceratops andrewsi and P. hellenikorhinus are clearly sepa-rated from ceratopsids with positive PC2 values and negative PC1values. Both species are close to each other in the morphospace,indicating their similar morphology. Protoceratops spp. have aslender, blade-like and dorsocaudally expanded squamosal shape,with an acute angle between the infratemporal process and thecaudoventral margin.
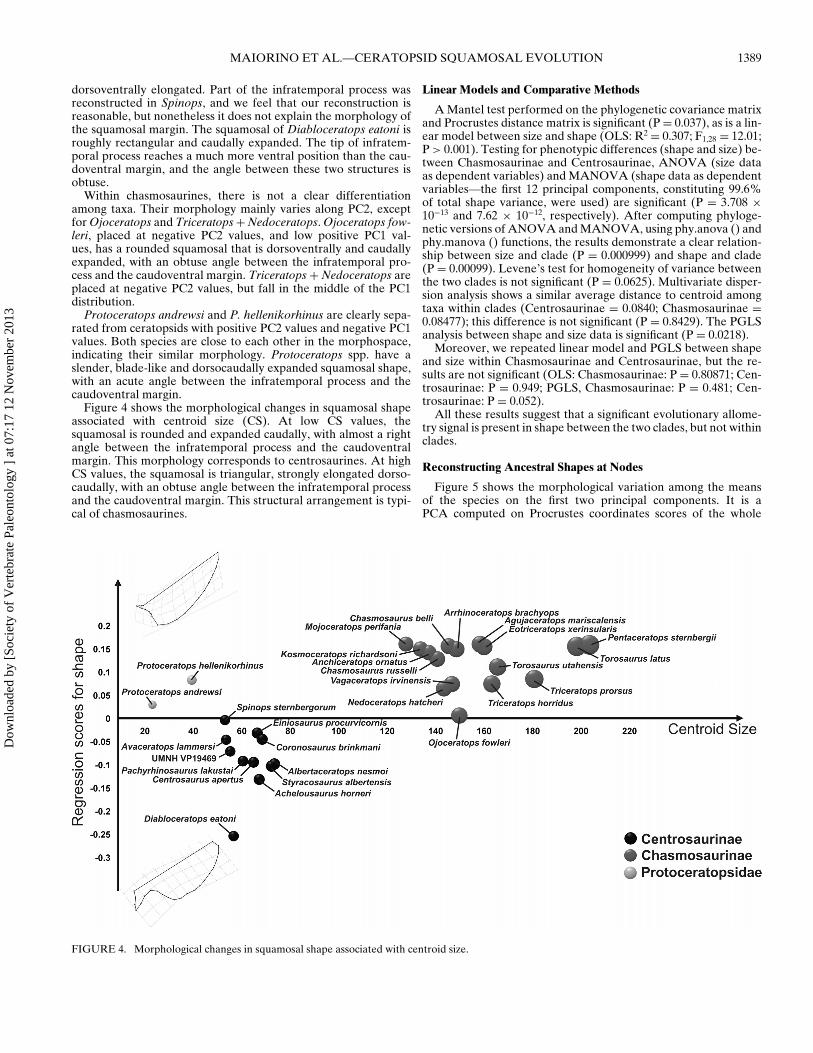
Figure 4 shows the morphological changes in squamosal shapeassociated with centroid size (CS). At low CS values, thesquamosal is rounded and expanded caudally, with almost a rightangle between the infratemporal process and the caudoventralmargin. This morphology corresponds to centrosaurines. At highCS values, the squamosal is triangular, strongly elongated dorso-caudally, with an obtuse angle between the infratemporal processand the caudoventral margin. This structural arrangement is typi-cal of chasmosaurines.
Linear Models and Comparative Methods
A Mantel test performed on the phylogenetic covariance matrixand Procrustes distance matrix is significant (P = 0.037), as is a lin-ear model between size and shape (OLS: R2 = 0.307; F1,28 = 12.01;P > 0.001). Testing for phenotypic differences (shape and size) be-tween Chasmosaurinae and Centrosaurinae, ANOVA (size dataas dependent variables) and MANOVA (shape data as dependentvariables—the first 12 principal components, constituting 99.6%of total shape variance, were used) are significant (P = 3.708 ×10−13 and 7.62 × 10−12, respectively). After computing phyloge-netic versions of ANOVA and MANOVA, using phy.anova () andphy.manova () functions, the results demonstrate a clear relation-ship between size and clade (P = 0.000999) and shape and clade(P = 0.00099). Levene’s test for homogeneity of variance betweenthe two clades is not significant (P = 0.0625). Multivariate disper-sion analysis shows a similar average distance to centroid amongtaxa within clades (Centrosaurinae = 0.0840; Chasmosaurinae =0.08477); this difference is not significant (P = 0.8429). The PGLSanalysis between shape and size data is significant (P = 0.0218).
Moreover, we repeated linear model and PGLS between shapeand size within Chasmosaurinae and Centrosaurinae, but the re-sults are not significant (OLS: Chasmosaurinae: P = 0.80871; Cen-trosaurinae: P = 0.949; PGLS, Chasmosaurinae: P = 0.481; Cen-trosaurinae: P = 0.052).
All these results suggest that a significant evolutionary allome-try signal is present in shape between the two clades, but not withinclades.
Reconstructing Ancestral Shapes at Nodes
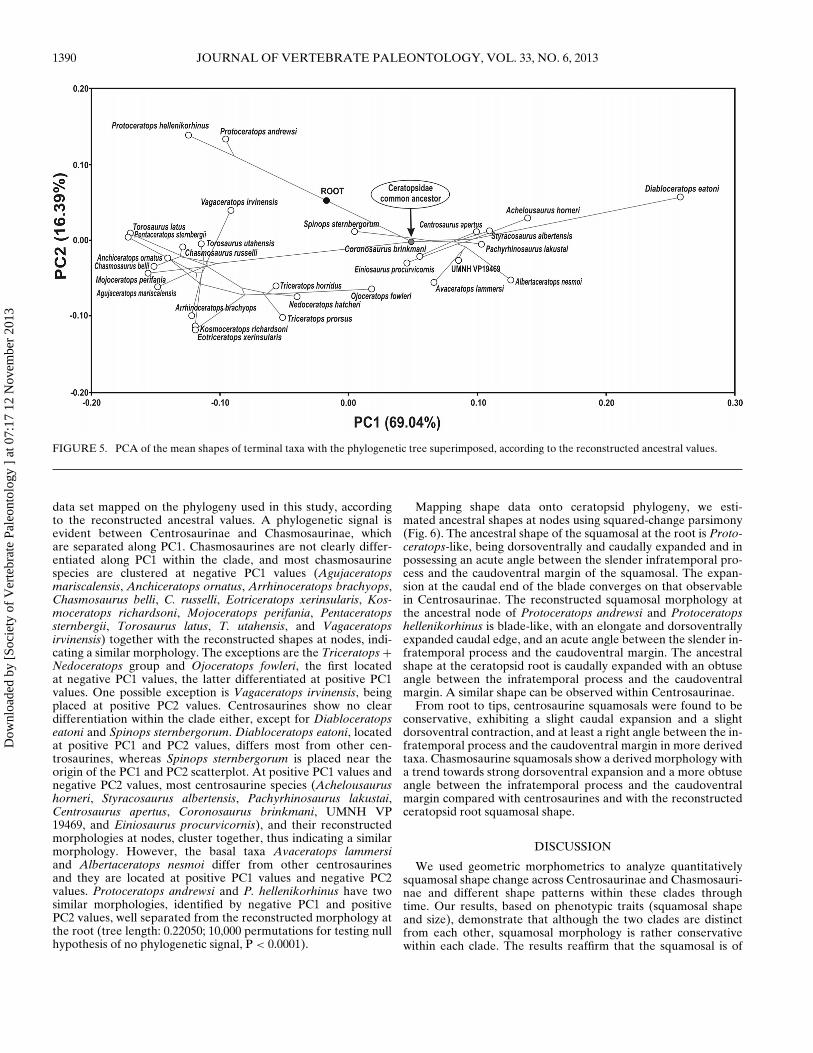
Figure 5 shows the morphological variation among the meansof the species on the first two principal components. It is aPCA computed on Procrustes coordinates scores of the whole
FIGURE 4. Morphological changes in squamosal shape associated with centroid size.
Dow
nloa
ded
by [
Soci
ety
of V
erte
brat
e Pa
leon
tolo
gy ]
at 0
7:17
12
Nov
embe
r 20
13
1390 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 33, NO. 6, 2013
FIGURE 5. PCA of the mean shapes of terminal taxa with the phylogenetic tree superimposed, according to the reconstructed ancestral values.
data set mapped on the phylogeny used in this study, accordingto the reconstructed ancestral values. A phylogenetic signal isevident between Centrosaurinae and Chasmosaurinae, whichare separated along PC1. Chasmosaurines are not clearly differ-entiated along PC1 within the clade, and most chasmosaurinespecies are clustered at negative PC1 values (Agujaceratopsmariscalensis, Anchiceratops ornatus, Arrhinoceratops brachyops,Chasmosaurus belli, C. russelli, Eotriceratops xerinsularis, Kos-moceratops richardsoni, Mojoceratops perifania, Pentaceratopssternbergii, Torosaurus latus, T. utahensis, and Vagaceratopsirvinensis) together with the reconstructed shapes at nodes, indi-cating a similar morphology. The exceptions are the Triceratops +Nedoceratops group and Ojoceratops fowleri, the first locatedat negative PC1 values, the latter differentiated at positive PC1values. One possible exception is Vagaceratops irvinensis, beingplaced at positive PC2 values. Centrosaurines show no cleardifferentiation within the clade either, except for Diabloceratopseatoni and Spinops sternbergorum. Diabloceratops eatoni, locatedat positive PC1 and PC2 values, differs most from other cen-trosaurines, whereas Spinops sternbergorum is placed near theorigin of the PC1 and PC2 scatterplot. At positive PC1 values andnegative PC2 values, most centrosaurine species (Achelousaurushorneri, Styracosaurus albertensis, Pachyrhinosaurus lakustai,Centrosaurus apertus, Coronosaurus brinkmani, UMNH VP19469, and Einiosaurus procurvicornis), and their reconstructedmorphologies at nodes, cluster together, thus indicating a similarmorphology. However, the basal taxa Avaceratops lammersiand Albertaceratops nesmoi differ from other centrosaurinesand they are located at positive PC1 values and negative PC2values. Protoceratops andrewsi and P. hellenikorhinus have twosimilar morphologies, identified by negative PC1 and positivePC2 values, well separated from the reconstructed morphology atthe root (tree length: 0.22050; 10,000 permutations for testing nullhypothesis of no phylogenetic signal, P < 0.0001).
Mapping shape data onto ceratopsid phylogeny, we esti-mated ancestral shapes at nodes using squared-change parsimony(Fig. 6). The ancestral shape of the squamosal at the root is Proto-ceratops-like, being dorsoventrally and caudally expanded and inpossessing an acute angle between the slender infratemporal pro-cess and the caudoventral margin of the squamosal. The expan-sion at the caudal end of the blade converges on that observablein Centrosaurinae. The reconstructed squamosal morphology atthe ancestral node of Protoceratops andrewsi and Protoceratopshellenikorhinus is blade-like, with an elongate and dorsoventrallyexpanded caudal edge, and an acute angle between the slender in-fratemporal process and the caudoventral margin. The ancestralshape at the ceratopsid root is caudally expanded with an obtuseangle between the infratemporal process and the caudoventralmargin. A similar shape can be observed within Centrosaurinae.
From root to tips, centrosaurine squamosals were found to beconservative, exhibiting a slight caudal expansion and a slightdorsoventral contraction, and at least a right angle between the in-fratemporal process and the caudoventral margin in more derivedtaxa. Chasmosaurine squamosals show a derived morphology witha trend towards strong dorsoventral expansion and a more obtuseangle between the infratemporal process and the caudoventralmargin compared with centrosaurines and with the reconstructedceratopsid root squamosal shape.
DISCUSSION
We used geometric morphometrics to analyze quantitativelysquamosal shape change across Centrosaurinae and Chasmosauri-nae and different shape patterns within these clades throughtime. Our results, based on phenotypic traits (squamosal shapeand size), demonstrate that although the two clades are distinctfrom each other, squamosal morphology is rather conservativewithin each clade. The results reaffirm that the squamosal is of
Dow
nloa
ded
by [
Soci
ety
of V
erte
brat
e Pa
leon
tolo
gy ]
at 0
7:17
12
Nov
embe
r 20
13

MAIORINO ET AL.—CERATOPSID SQUAMOSAL EVOLUTION 1391
FIGURE 6. Reconstruction of ancestral squamosal shapes at nodes using the squared-change parsimony criterion. The gray contour is the squamosalshape at root, and the black contours are squamosal shapes at the node.
limited taxonomic utility, confirming previous anecdotal and non-quantitative observations (Lehman, 1990; Dodson et al., 2004;Maidment and Barrett, 2011).
The Chasmosaurinae squamosal is morphologically conserva-tive except for the derived taxa Ojoceratops and Triceratops +Nedoceratops. The element is also conservative in centrosaurinesexcept for the basal and derived centrosaurines Diabloceratopseatoni and Spinops sternbergorum, respectively.
The position in morphospace of Triceratops + Nedoceratopsindicates a similar squamosal morphology among these taxa, assuggested by Scannella and Horner (2011); however, the Nedocer-atops squamosal does seem to reflect a shape variation with Tricer-atops spp. Ojoceratops is located between the Triceratops + Nedo-ceratops group and the basal centrosaur Avaceratops. Ojoceratopshas a squamosal shape, with a large and broad lateral outline, aspreviously noted (Sullivan and Lucas, 2010). Its width resembles,partly, a centrosaurine-like shape.
The inclusion of Protoceratops spp. in the analysis allows ten-tative reconstruction of the transition from the non-ceratopsidplesiomorphic condition (immediately outside of Ceratopsidae)to the derived ceratopsid condition of the squamosal. At theroot, the ancestral morphology (non-ceratopsid condition) is rep-resented by a squamosal that is Protoceratops-like in being elon-gated dorsoventrally, but also partly caudally expanded, converg-ing on a centrosaurine-like shape, with an acute angle betweenthe short infratemporal process and the caudoventral margin. Un-fortunately, the squamosal is not well preserved in specimens of
the immediate outgroup to Ceratopsidae (Zuniceratops and Tu-ranoceratops), so confirmation of this reconstructed morphologyawaits better fossil material.
The ancestral ceratopsid shape is centrosaurine-like, with asquamosal expanded caudally and a more obtuse angle betweenthe infratemporal process and the caudoventral margin than seenin chasmosaurines. A Mantel test between the phylogenetic co-variance matrix and Procrustes distance matrix demonstrates thatdisparity in ceratopsid squamosals is highly constrained by phy-logeny. The regression between size and shape of the pooled sam-ple, ANOVA and MANOVA, and their phylogenetic versions,between shape and size against the discrete variable clade, sug-gests a strong evolutionary allometric signal in squamosal mor-phology between Chasmosaurinae and Centrosaurinae, but notwithin these clades. Even when accounting for phylogeny (phy-logenetic generalized least squares), the relationship betweensquamosal shape and size is significant. Mapping shape data ontophylogeny confirms the conservative squamosal shape patternin Centrosaurinae and Chasmosaurinae, even if this latter cladeshows a derived morphology, as compared with Centrosaurinae.The overall shape of the squamosal in Diabloceratops is at leastsuperficially similar to the condition in Protoceratops (Kirklandand DeBlieux, 2010) in some aspects (such as a squared poste-rior margin), but the analysis here indicates that many aspectsof the squamosal shape in Diabloceratops are unique. Diablocer-atops has a rectangular squamosal expanded caudally, with a rightor slightly obtuse angle between the infratemporal process and
Dow
nloa
ded
by [
Soci
ety
of V
erte
brat
e Pa
leon
tolo
gy ]
at 0
7:17
12
Nov
embe
r 20
13

1392 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 33, NO. 6, 2013
FIGURE 7. Comparison between Diabloceratops and Protoceratopssquamosal shape. Gray line indicates the average shape for Protoceratopsand the black line indicates Diabloceratops (UMNH VP 16099).
the caudoventral margin, as described in Kirkland and DeBlieux(2010). Protoceratops, by contrast, has a slender, blade-like, anddorsocaudally elongated squamosal, with an acute angle betweenthe infratemporal process and the caudoventral margin (Fig. 7).Relative to Diabloceratops, the lateral profile is greatly expandedin centrosaurines such as Centrosaurus and Albertaceratops.
The underlying causes for the differences in squamosal shapebetween the two ceratopsid clades are uncertain, but may be re-lated to modification for visual display or thermoregulatory func-tions (Farlow and Dodson, 1975; Sampson et al., 1997; Hone et al.,2011), or the size of the area needed for the insertion of the m.longissimus capitis in the depressions on the ventral surface of thesquamosal blade (Tsuihiji, 2010). Additional work is needed onthis issue.
This paper is the first to use geometric morphometrics to quanti-tatively investigate shape changes of squamosal bones in ceratop-sids. It will be interesting in future work to expand the sample,including juveniles, subadults, and adults of all ceratopsid speciesand of all basal ceratopsian species as well, when possible, to in-vestigate shape changes of the squamosals, disparity rates, and theontogenetic variation of ceratopsians through time.
ACKNOWLEDGMENTS
We are grateful to many institutions and their staff for allowingus access to the collection in their care. We are also grateful toS. D. Sampson and E. K. Lund for allowing access to UMNH VP19469. Portions of this work were supported under National Sci-ence Foundation grant EAR 0819953. We also thank M. Loewenand an anonymous reviewer for their comments that improved themanuscript.
LITERATURE CITED
Adams, D. C., F. J. Rohlf, and D. E. Slice. 2004. Geometric morphomet-rics: ten years of progress following the ‘revolution’. Italian Journalof Zoology 71:5–16.
Bookstein, F. L. 1986. Size and shape spaces for landmark data in two di-mensions. Statistical Science 1:181–242.
Bookstein, F. L. 1991. Morphometric Tools for Landmark Data: Geome-try and Biology. Cambridge University Press, Cambridge (U.K.) andNew York, 456 pp.
Bookstein, F. L., A. P. Streissguth, P. D. Sampson, P. D. Connor, andH. H. Barr. 2002. Corpus callosum shape and neuropsychologicaldeficits in adult males with heavy fetal alcohol exposure. NeuroIm-age 15:233–251.
Chapman, R. E. 1990. Shape analysis in the study of dinosaur morphol-ogy; pp. 21–42 in K. Carpenter and P. J. Currie (eds.), Dinosaur Sys-tematics: Approaches and Perspectives. Cambridge University Press,Cambridge (U.K.) and New York.
Chinnery, B. 2004. Morphometric analysis of evolutionary trends in theceratopsian postcranial skeleton. Journal of Vertebrate Paleontology24:591–609.
Claude, J. 2008. Morphometrics with R. Springer, New York, 318 pp.Dodson, P. 1986. Avaceratops lammersi: a new ceratopsid from the Judith
River Formation of Montana. Proceedings of the Academy of Natu-ral Sciences of Philadelphia 138:305–317.
Dodson, P. 1993. Comparative craniology of the Ceratopsia. AmericanJournal of Science 293A:200–234.
Dodson, P., C. A. Forster, and S. D. Sampson. 2004. Ceratopsidae;pp. 494–513 in D. B. Weishampel, P. Dodson, and H. Osmolska(eds.), The Dinosauria, second edition. University of California Press,Berkeley, California.
Farke, A. A., M. J. Ryan, P. M. Barrett, D. H. Tanke, D. R. Braman,M. A. Loewen, and M. R. Graham. 2011. A new centrosaurine fromthe Late Cretaceous of Alberta, Canada, and the evolution of pari-etal ornamentation in horned dinosaurs. Acta Paleontologia Polonica56:691–702.
Farlow, J. O., and P. Dodson. 1975. The behavioral significance of frill andhorn morphology in ceratopsian dinosaurs. Evolution 29:353–361.
Felsenstein, J. 1985. Phylogenies and the comparative method. AmericanNaturalist 125:1–15.
Garland, T., Jr. 1992. Rate tests for phenotypic evolution using phyloge-netically independent contrasts. American Naturalist 140:509–519.
Harmon, L. J., J. Weir, C. Brock, R. E. Glor, and W. Challenger.2008. Geiger: investigating evolutionary radiations. Bioinformatics24:129–131.
Hone, D. W. E., D. Naish, and I. C. Cuthill. 2011. Does mutual sexual selec-tion explain the evolution of head crests in pterosaurs and dinosaurs?Lethaia 45:139–156.
Kirkland, J. I., and D. D. DeBlieux. 2010. New basal centrosaurinae cer-atopsian skull from the Wahweap Formation (Middle Campanian),Grand Staircase-Escalante Monument, Southern Utah; pp. 117–140in M. J. Ryan, B. J. Chinnery-Allgeier, and D. A. Eberth (eds.),New Perspectives on Horned Dinosaurs. Indiana University Press,Bloomington, Indiana.
Klingenberg, C. P. 2011. MorphoJ: an integrated software package forgeometric morphometrics. Molecular Ecology Research 11:353–357.
Klingenberg, C. P., and N. A. Gidaszewski. 2010. Testing and quantifyingphylogenetic signals and homoplasy in morphometric data. System-atic Biology 59:245–261.
Lehman, T. M. 1990. The ceratopsian subfamily Chasmosaurinae: sexualdimorphism and systematics; pp. 211–229 in K. Carpenter and P. J.Currie (eds.), Dinosaur Systematics: Approaches and Perspectives.Cambridge University Press, Cambridge (U.K.) and New York.
Longrich, N. R. 2011. Titanoceratops ouranos, a giant horned dinosaurfrom the late Campanian of New Mexico. Cretaceous Research32:264–276.
Maddison, W. P. 1991. Squared-change parsimony reconstructions of an-cestral states for continuous-valued characters on a phylogenetic tree.Systematic Zoology 40:304–314.
Maddison, W. P., and D. R. Maddison. 2011. Mesquite: A Modu-lar System for Evolutionary Analysis, version 2.75. Available athttp://mesquiteproject.org/mesquite/mesquite.html. Accessed Octo-ber 3, 2012.
Maidment, S. C. R., and P. M. Barrett. 2011. A new specimen of Chas-mosaurus belli (Ornithischia: Ceratopsidae), a revision of the genus,and the utility of postcrania in the taxonomy and systematics of cer-atopsid dinosaurs. Zootaxa 2963:1–47.
Marcus, L. F., E. Hingst-Zaher, and H. Zaher. 2000. Application of land-marks morphometrics to skull representing the orders of living mam-mals. Hystrix 11:27–47.
Martins, E. P., and T. F. Hansen. 1997. Phylogenies and the compar-ative method: a general approach to incorporating phylogenetic
Dow
nloa
ded
by [
Soci
ety
of V
erte
brat
e Pa
leon
tolo
gy ]
at 0
7:17
12
Nov
embe
r 20
13

MAIORINO ET AL.—CERATOPSID SQUAMOSAL EVOLUTION 1393
information into the analysis of interspecific data. American Natu-ralist 149:646–667.
McArdle, B. H., and A. G. Rodrigo. 1994. Estimating the ancestral statesof a continuous-valued character using squared-change parsimony: ananalytical solution. Systematic Biology 43:573–578.
Mullin, S. K., and P. J. Taylor. 2002. The effects of parallax on geometricmorphometric data. Computers in Biology and Medicine 32:455–464.
Oksanen, J., F. G. Blanchet, R. Kindt, P. Legendre, P. R. Minchin, R.B. O’Hara, G. L. Simpson, P. Solymos, M. H. H. Stevens, and H.Wagner. 2011. Vegan: Community Ecology Package. R package, ver-sion 2.0-2. Available at http://CRAN.R-project.org/package=vegan.Accessed June 20, 2012.
Paradis, E., J. Claude, and K. Strimmer. 2004. APE: analyses of phyloge-netics and evolution in R language. Bioinformatics 20:289–290.
Perez, S. I., V. Bernal, and P. N. Gonzalez. 2006. Differences betweensliding semi-landmark methods in geometric morphometrics, with anapplication to human craniofacial and dental variation. Journal ofAnatomy 208:769–784.
Piras, P., L. Maiorino, P. Raia, F. Marcolini, D. Salvi, L. Vignoli,and T. Kotsakis. 2010. Functional and phylogenetic constraints inRhinocerotinae craniodental morphology. Evolutionary Ecology Re-search 12:897–928.
Rohlf, F. J. 2001. Comparative methods for the analysis of continuous vari-ables: geometric interpretations. Evolution 55:2143–2160.
Rohlf, F. J. 2002. Geometric morphometrics and phylogeny; pp. 175–193in N. MacLeod and P. L. Forey (eds.), Morphology, Shape, and Phy-logeny. Taylor and Francis, London.
Rohlf, F. J. 2006. A comment on phylogenetic corrections. Evolution60:1509–1515.
Rohlf, F. J. 2010a. tpsDig2, version 2.16. Available at http://life.bio.sunysb.edu/morph/. Accessed December 11, 2011.
Rohlf, F. J. 2010b. tpsRelw, version 1.49. Available at http://life.bio.sunysb.edu/morph/. Accessed December 11, 2011.
Rohlf, F. J., and D. E. Slice. 1990. Extensions of the Procrustes methodfor the optimal superimposition of landmarks. Systematic Biology39:40–59.
Ryan, M. J. 2007. A new basal centrosaurinae ceratopsid from theOldman Formation, southeastern Alberta. Journal of Paleontology81:376–396.
Ryan, M. J., D. C. Evans, and K. M. Shepherd. 2012. A new ceratopsidfrom the Foremost Formation (middle Campanian) of Alberta. Cana-dian Journal of Earth Sciences 49:1251–1262.
Ryan, M. J., A. P. Russell, D. A. Eberth, and P. J. Currie. 2001. Thetaphonomy of a Centrosaurus (Ornithischia: Certopsidae) bone bedfrom the Dinosaur Park Formation (Upper Campanian), Alberta,Canada, with comments on cranial ontogeny. Palaios 16:482–506.
Sampson, S. D., and M. A. Loewen. 2010. Unraveling a radiation: a reviewof the diversity, stratigraphic, distribution, biogeography, and evolu-
tion of horned dinosaurs (Ornithischia: Ceratopsidae); pp. 405–427in M. J. Ryan, B. J. Chinnery-Allgeier, and D. A. Eberth (eds.), NewPerspectives on Horned Dinosaurs. Indiana University Press, Bloom-ington, Indiana.
Sampson, S. D., M. J. Ryan, and D. H. Tanke. 1997. Craniofacial ontogenyin centrosaurine dinosaurs (Omithischia: Ceratopsidae): taxonomicand behavioral implications. Zoological Journal of the Linnean Soci-ety 121:293–337.
Sampson, S. D., M. A. Loewen, A. A. Farke, E. M. Roberts, C. A.Forster, J. A. Smith, and A. L. Titus. 2010. New horned dinosaursfrom Utah provide evidence for intracontinental dinosaur endemism.PLoS ONE 5:e12292.
Scannella, J. B., and J. R. Horner. 2011. “Nedoceratops”: an example of atransitional morphology. PLoS ONE 6:e28705.
Schlager, S. 2012. Morpho. R package. Available at http://www.uniklinik-freiburg.de/anthropologie/live/forschung/Shape en.html. AccessedJuly 19, 2012.
Schlick-Steiner, B. C., F. M. Steiner, K. Moder, B. Seifert, M. Sanetra, E.Dyreson, C. Stauffer, and E. Christian. 2006. A multidisciplinary ap-proach reveals cryptic diversity in Western Palearctic Tetramoriumants (Hymenoptera: Formicidae). Molecular Phylogenetics and Evo-lution 40:259–273.
Sullivan, R. M., and S. G. Lucas. 2010. A new chasmosaurine (Ceratopsi-dae, Dinosauria) from the Upper Cretaceous Ojo Alamo Formation(Naashoibito Member), San Juan Basin, New Mexico; pp. 169–180in M. J. Ryan, B. J. Chinnery-Allgeier, and D. A. Eberth (eds.), NewPerspectives on Horned Dinosaurs. Indiana University Press, Bloom-ington, Indiana.
Tsuihiji, T. 2010. Reconstructions of the axial muscle insertions in theoccipital region of dinosaurs: evaluations of past hypotheses onMarginocephalia and Tyrannosauridae using the Extant PhylogeneticBracket approach. The Anatomical Record 293:1360–1386.
Wu, X.-C., D. B. Brinkman, D. A. Eberth, and D. R. Braman. 2007. A newceratopsid dinosaur (Ornithischia) from the uppermost HorseshoeCanyon Formation (upper Maastrichtian), Alberta, Canada. Cana-dian Journal of Earth Sciences 44:1243–1265.
You, H.-L., and P. Dodson. 2004. Basal Ceratopsia; pp. 478–593 in D. B.Weishampel, P. Dodson, and H. Osmolska (eds.), The Dinosauria,second edition. University of California Press, Berkeley, California.
Young, M. T., S. L. Brusatte, M. Ruta, and M. B. De Andrade. 2010.The evolution of Metriorhynchoidea (Mesoeucrocodylia, Thalatto-suchia): an integrated approach using geometric morphometrics,analysis of disparity, and biomechanics. Zoological Journal of Lin-nean Society 158:801–859.
Submitted November 8, 2012; revisions received February 8, 2013;accepted February 18, 2013.Handling editor: Randall Irmis.
Dow
nloa
ded
by [
Soci
ety
of V
erte
brat
e Pa
leon
tolo
gy ]
at 0
7:17
12
Nov
embe
r 20
13




















