
The effect of prolonged exposure to morphine on canine cerebral 5-HT2A receptors measured with...
6
The effect of prolonged exposure to morphine on canine cerebral 5-HT2A receptors measured with 123 I-R91150 SPECT $ Antita Adriaens a , Ingeborgh Polis a,n , Simon Vermeire b , Tim Waelbers a , Siska Croubels c , Luc Duchateau d , Sylvia Van Dorpe e , Jos Eersels f , Bart De Spiegeleer e , Kathelijne Peremans b a Medicine and Clinical Biology of Small Animals, Faculty of Veterinary Medicine, Ghent University, Salisburylaan 133, 9820 Merelbeke, Belgium b Medical Imaging of Domestic Animals, Faculty of Veterinary Medicine, Ghent University, Merelbeke, Belgium c Pharmacology, Biochemistry and Toxicology, Faculty of Veterinary Medicine, Ghent University, Merelbeke, Belgium d Physiology and Biometry, Faculty of Veterinary Medicine, Ghent University, Merelbeke, Belgium e Drug Quality & Registration (DruQuaR) Group, Faculty of Pharmaceutical Sciences, Ghent University, Ghent, Belgium f Nuclear Medicine and PET Research, VU University Medical Centre, Amsterdam, The Netherlands Received 21 August 2013; received in revised form 19 December 2013; accepted 19 March 2014 KEYWORDS 5-HT2A; Morphine; 123 I-R91150; SPECT; Dogs Abstract Down-stream neuronal alterations, including changes in the 5-HT-2A receptor system, play an important role in the etiology and treatment of depression. The present study examined the effect of prolonged opioid treatment on cerebral 5-HT2A receptors. Cerebral 5-HT2A receptor availability was estimated in seven healthy five-year-old female neutered Beagle dogs pre and post 10-day morphine treatment (oral sustained release morphine 20 mg twice daily for 10 days) with 123 I-R-91150, a 5-HT2A selective radioligand, and SPECT. 5-HT2A receptor binding indices (BI) for the frontal, parietal, temporal and occipital cortex and the subcortical region were calculated. Statistical analysis was performed using a linear mixed-effect model with treatment as fixed effect and dog as random effect. Morphine treatment significantly (P r0.05) lowered 5-HT2A BIs in the right and left frontal cortex, the right and left temporal cortex, the www.elsevier.com/locate/euroneuro http://dx.doi.org/10.1016/j.euroneuro.2014.03.004 0924-977X/& 2014 Elsevier B.V. and ECNP. All rights reserved. ☆ Financial support: This work was funded by a Special Research Fund Grant from the Ghent University (No. 01J06109). The authors declare no conflict of interest. n Corresponding author. Tel.: + 32 9 264 77 00; fax: + 32 9 264 77 91. E-mail address: [email protected] (I. Polis). European Neuropsychopharmacology (2014) 24, 1133–1138
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The effect of prolonged exposure to morphine on canine cerebral 5-HT2A receptors measured with...

European Neuropsychopharmacology (2014) 24, 1133–1138
http://dx.doi.org/10924-977X/& 2014 E
☆Financial supportno conflict of intere
nCorresponding auE-mail address: I
www.elsevier.com/locate/euroneuro
The effect of prolonged exposureto morphine on canine cerebral 5-HT2Areceptors measured with 123I-R91150 SPECT$
Antita Adriaensa, Ingeborgh Polisa,n, Simon Vermeireb,Tim Waelbersa, Siska Croubelsc, Luc Duchateaud,Sylvia Van Dorpee, Jos Eerselsf, Bart De Spiegeleere,Kathelijne Peremansb
aMedicine and Clinical Biology of Small Animals, Faculty of Veterinary Medicine, Ghent University,Salisburylaan 133, 9820 Merelbeke, BelgiumbMedical Imaging of Domestic Animals, Faculty of Veterinary Medicine, Ghent University,Merelbeke, BelgiumcPharmacology, Biochemistry and Toxicology, Faculty of Veterinary Medicine, Ghent University,Merelbeke, BelgiumdPhysiology and Biometry, Faculty of Veterinary Medicine, Ghent University, Merelbeke, BelgiumeDrug Quality & Registration (DruQuaR) Group, Faculty of Pharmaceutical Sciences,Ghent University, Ghent, BelgiumfNuclear Medicine and PET Research, VU University Medical Centre, Amsterdam, The Netherlands
Received 21 August 2013; received in revised form 19 December 2013; accepted 19 March 2014
KEYWORDS5-HT2A;Morphine;123I-R91150;SPECT;Dogs
0.1016/j.euroneurlsevier B.V. and E
: This work was fust.thor. Tel.: +32 9 2ngeborgh.polis@U
AbstractDown-stream neuronal alterations, including changes in the 5-HT-2A receptor system, play animportant role in the etiology and treatment of depression. The present study examined theeffect of prolonged opioid treatment on cerebral 5-HT2A receptors. Cerebral 5-HT2A receptoravailability was estimated in seven healthy five-year-old female neutered Beagle dogs pre andpost 10-day morphine treatment (oral sustained release morphine 20 mg twice daily for 10days) with 123I-R-91150, a 5-HT2A selective radioligand, and SPECT. 5-HT2A receptor bindingindices (BI) for the frontal, parietal, temporal and occipital cortex and the subcortical regionwere calculated. Statistical analysis was performed using a linear mixed-effect model withtreatment as fixed effect and dog as random effect. Morphine treatment significantly (Pr0.05)lowered 5-HT2A BIs in the right and left frontal cortex, the right and left temporal cortex, the
o.2014.03.004CNP. All rights reserved.
nded by a Special Research Fund Grant from the Ghent University (No. 01J06109). The authors declare
64 77 00; fax: +32 9 264 77 91.Gent.be (I. Polis).

A. Adriaens et al.1134
right and left parietal cortex, and the subcortical region. The decreased cerebral 5-HT2Areceptor availability following prolonged morphine exposure provides further evidence for aninteraction between the opioid and serotonergic system.& 2014 Elsevier B.V. and ECNP. All rights reserved.
1. Introduction
Interest in the down-stream molecular effects occurringduring depression and antidepressant treatment has grownover the last decade. It has been suggested that abnormal-ities in monoamine receptors and a relative deficiency ofmonoamines may lead to depression (Stahl, 2008). Thesedown-stream neuronal adaptations could also explain thelag-time in onset of response to antidepressant treatment,rather than the immediate synaptic 5-HT elevation itself(Manji et al., 2000).
In depressed patients, adaptive upregulation of postsy-naptic receptors, such as the serotonergic 2A (5-HT2A)receptor, has been observed (Cahir et al., 2007; Sheltonet al., 2009). Furthermore, 5-HT2A receptor expression hasbeen shown to be altered after SSRI treatment in humansand dogs (Gray and Roth, 2001; Peremans et al., 2005) andatypical antipsychotics with 5-HT2A receptor antagonistproperties have been shown to augment the therapeuticeffect of SSRIs (Marek et al., 2003; Nelson and Papakostas,2009; Ostroff and Nelson, 1999; Philip et al., 2008). There-fore, the postsynaptic 5-HT2A receptor is thought to be apossible target for antidepressant therapy.
Mu opioid receptor (MOR) agonists have been reported toincrease serotonin levels in the brain (Tao and Auerbach,1994) and in view of this, the use of opioids as additionalantidepressant therapeutics has been investigated. Indeed,biochemical and behavioral studies suggest the involvementof the opioid system in the mechanism of action of certainantidepressants (Fichna et al., 2007). Since both opioids,like morphine, and 5-HT2A receptors appear to be involvedin similar biological processes, and we have previouslydemonstrated that a single dose of morphine lowered the5-HT2A receptor availability in the canine frontal cortex(Adriaens et al., 2012), the aim of the present study is toassess the influence of prolonged administration of mor-phine on the cerebral 5-HT2A receptors in dogs.
2. Experimental procedures
2.1. Animals
Seven healthy Beagle dogs (neutered females), ranging in weightfrom 6.4 kg to 7.4 kg, and age from 3 to 4 years old were used inthis study. None of the dogs had a history of previous major diseaseor neurological disorder. All procedures and protocols wereapproved by the Ghent University Ethical Committee (EC2010/152).
2.2. Study design
Each dog underwent two SPECT scans. The first scan consistedof a baseline scan (pre-morphine condition) to which data fromthe second scan (post-morphine condition) were compared.
Post-morphine scans were performed after an opioid treatmentthat consisted of an oral sustained release formulation of morphine(20 mg morphine sulfate, MS Contins, Mundipharma, Mechelen,Belgium) administered orally twice daily for 10 days, which iscomparable to a dosage of 2.8670.16 mg/kg twice daily. Dogs weremonitored closely (respiratory rate, heart rate and quality, degreeof sedation, nausea and emesis) for 3 h after morphine administra-tion and side effects were noted if observed.
Food, but not water, was withheld for 12 h before each scan.Morphine plasma concentrations were determined to verify whetherthe given morphine dosage was within clinically relevant ranges.For this, 5 mL of blood was collected from the jugular vein andplaced in a heparinized blood tube the last day of treatment 3 hafter the last morphine administration. Blood was centrifuged at3500 rpm for 5 min after which plasma was collected and kept at�20 1C until analysis was performed.
2.3. Tracer
123I-R91150 is synthesized by electrophilic radioiodination on the5-position of the methoxybenzamide group of R91150 (Eerselset al., 2005), followed by purification with high-performance liquidchromatography and was obtained from the Nuclear Medicine andPET research department of Nuclear Medicine and PET research ofthe VU University Medical Centre (Amsterdam, The Netherlands).The product had a radiochemical purity of more than 99% and wassterile and pyrogen free. The administered radioligand had aspecific activity of 370 GBq/mmol. The radioligand is a 5-HT2Aantagonist with high affinity (Kd=0.11 nM) and selectivity for the5-HT2A receptor. The selectivity of the ligand for 5-HT2A receptorswith regard to other neurotransmitter receptors such as other 5-HTreceptors, including 5-HT2C and 5-HT1A, dopamine receptors,adrenergic receptors and histamine receptors is at least a factor50 (Mertens et al., 1994). The tracer is displaceable with ketan-serin, a 5-HT2 antagonist (Catafau et al., 2006) and has beenpreviously shown to be suitable for the use in dogs (Peremans et al.,2002).
2.4. Anesthetic protocol
All dogs were sedated prior to general anesthesia with intramus-cular dexmedetomidine 375 mg/m2 body surface area (Dexdomi-tors, Orion Corporation, Espoo, Finland). After induction with pro-pofol (Propovets, Abbott Laboratories, Queenborough, UnitedKingdom), anesthesia was maintained with isoflurane (Isoflos,Abbott Laboratories) in oxygen. Acquisition started 10 min afterinduction of anesthesia. The dogs were mechanically ventilated byintermittent positive pressure ventilation to maintain eucapnia, i.e.end tidal CO2 within 35–45 mmHg, in order to limit the influence ofthe PaCO2 on cerebral perfusion.
2.5. Image acquisition
The dogs were positioned in ventral recumbence with fixation of thehead in a preformed cushion, to prevent individual positioningartifacts. SPECT was performed with a triple head gamma camera(Triad Trionix, Twinsburg, OH, USA) equipped with ultrahigh resolution
1135The effect of prolonged exposure to morphine on canine cerebral 5-HT2A receptors measured with 123I-R91150 SPECT
parallel hole collimators (FWHM of 8 mm). Images were acquired90 min post-injection of the radioligand, this is the moment at whichpseudoequilibrium is present in dogs (Peremans et al., 2003a). Totalacquisition time was 30 min. For each acquisition, 90 projectionimages were obtained on a 128� 128 matrix using a step-and-shootmode (20 s/step, 41 steps). Camera and table positioning wererecorded to ensure optimal ‘intraindividual’ comparison.
2.6. Image processing protocol
The radioligand data were reconstructed using the HERMES Hybri-dRecon program (HERMES Medical Solutions, Stockholm, Sweden).The reconstruction is based on an ordered subset expectationmaximization-based reconstruction algorithm with attenuation(using a CT transmission scan), collimator response and MonteCarlo-based scatter correction. The details of the reconstructionalgorithm can be found in Sohlberg et al. (2008). Reconstructed imageswere post-filtered using a Butterworth filter (cut-off, 1.6 cycles/cm,order 5).
The 123I-R91150 emission data were fitted to a template, that isbased on previously obtained cerebral perfusion data, using multi-modality software (version 5.0; Nuclear Diagnostic AB) (Peremanset al., 2003a). This software displays images in a dual-windowsetting and allows for manual co-registration by providing tools forscaling, rotating, and translating images in all 3 dimensions. Follow-ing volumes of interest (VOIs) were included: right and left frontal,temporal, parietal and occipital cortices, as well as the subcorticaland cerebellar region. The cerebellum, a region with low densities of5-HT2A receptors, is used as a reference region for non-specificallybound and free radioligand. Consequently, the measured radioactivityin the cortical regions represents the total activity, i.e. specific andnon-specific bound and free radioligand. The binding index (BI) wascalculated as ([counts per pixel in VOI]�[counts per pixel in cere-bellum])/(counts per pixel in cerebellum). This BI is proportional tothe in vivo density of available receptors under pseudoequilibriumconditions (Pazos et al., 1987).
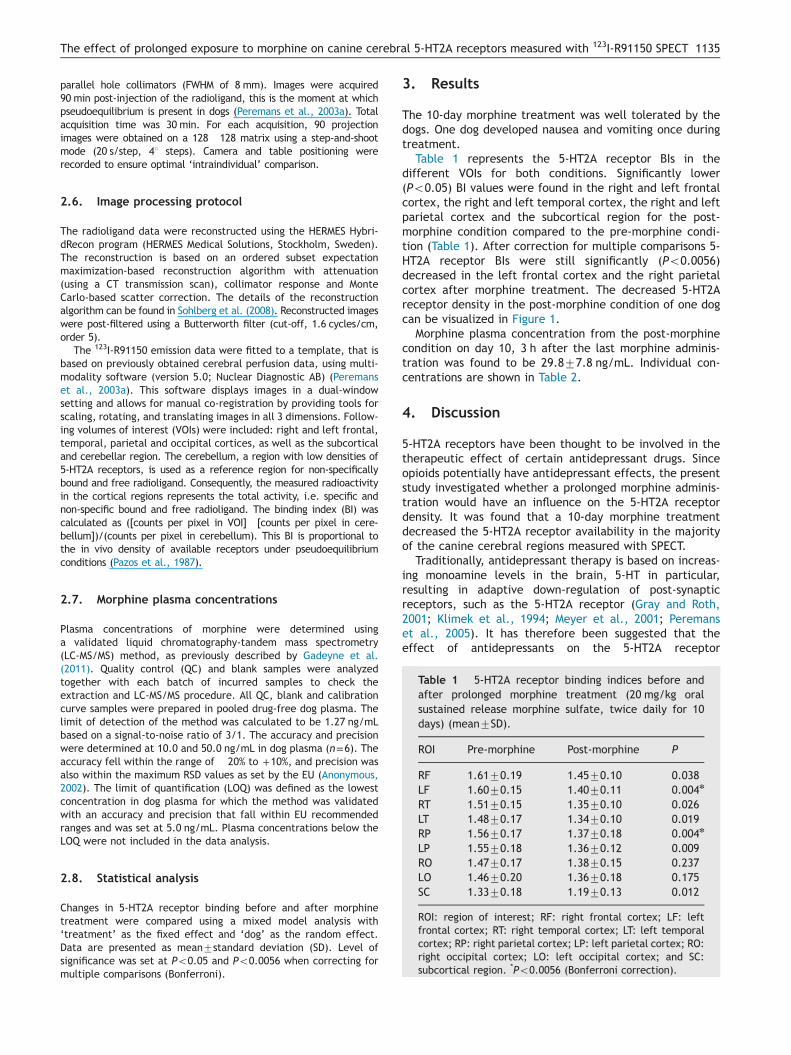
Table 1 5-HT2A receptor binding indices before andafter prolonged morphine treatment (20 mg/kg oralsustained release morphine sulfate, twice daily for 10days) (mean7SD).
ROI Pre-morphine Post-morphine P
RF 1.6170.19 1.4570.10 0.038LF 1.6070.15 1.4070.11 0.004n
RT 1.5170.15 1.3570.10 0.026LT 1.4870.17 1.3470.10 0.019RP 1.5670.17 1.3770.18 0.004n
2.7. Morphine plasma concentrations
Plasma concentrations of morphine were determined usinga validated liquid chromatography-tandem mass spectrometry(LC-MS/MS) method, as previously described by Gadeyne et al.(2011). Quality control (QC) and blank samples were analyzedtogether with each batch of incurred samples to check theextraction and LC-MS/MS procedure. All QC, blank and calibrationcurve samples were prepared in pooled drug-free dog plasma. Thelimit of detection of the method was calculated to be 1.27 ng/mLbased on a signal-to-noise ratio of 3/1. The accuracy and precisionwere determined at 10.0 and 50.0 ng/mL in dog plasma (n=6). Theaccuracy fell within the range of �20% to +10%, and precision wasalso within the maximum RSD values as set by the EU (Anonymous,2002). The limit of quantification (LOQ) was defined as the lowestconcentration in dog plasma for which the method was validatedwith an accuracy and precision that fall within EU recommendedranges and was set at 5.0 ng/mL. Plasma concentrations below theLOQ were not included in the data analysis.
LP 1.5570.18 1.3670.12 0.009RO 1.4770.17 1.3870.15 0.237LO 1.4670.20 1.3670.18 0.175SC 1.3370.18 1.1970.13 0.012
ROI: region of interest; RF: right frontal cortex; LF: leftfrontal cortex; RT: right temporal cortex; LT: left temporalcortex; RP: right parietal cortex; LP: left parietal cortex; RO:right occipital cortex; LO: left occipital cortex; and SC:subcortical region. *Po0.0056 (Bonferroni correction).
2.8. Statistical analysis
Changes in 5-HT2A receptor binding before and after morphinetreatment were compared using a mixed model analysis with‘treatment’ as the fixed effect and ‘dog’ as the random effect.Data are presented as mean7standard deviation (SD). Level ofsignificance was set at Po0.05 and Po0.0056 when correcting formultiple comparisons (Bonferroni).
3. Results
The 10-day morphine treatment was well tolerated by thedogs. One dog developed nausea and vomiting once duringtreatment.
Table 1 represents the 5-HT2A receptor BIs in thedifferent VOIs for both conditions. Significantly lower(Po0.05) BI values were found in the right and left frontalcortex, the right and left temporal cortex, the right and leftparietal cortex and the subcortical region for the post-morphine condition compared to the pre-morphine condi-tion (Table 1). After correction for multiple comparisons 5-HT2A receptor BIs were still significantly (Po0.0056)decreased in the left frontal cortex and the right parietalcortex after morphine treatment. The decreased 5-HT2Areceptor density in the post-morphine condition of one dogcan be visualized in Figure 1.
Morphine plasma concentration from the post-morphinecondition on day 10, 3 h after the last morphine adminis-tration was found to be 29.877.8 ng/mL. Individual con-centrations are shown in Table 2.
4. Discussion
5-HT2A receptors have been thought to be involved in thetherapeutic effect of certain antidepressant drugs. Sinceopioids potentially have antidepressant effects, the presentstudy investigated whether a prolonged morphine adminis-tration would have an influence on the 5-HT2A receptordensity. It was found that a 10-day morphine treatmentdecreased the 5-HT2A receptor availability in the majorityof the canine cerebral regions measured with SPECT.
Traditionally, antidepressant therapy is based on increas-ing monoamine levels in the brain, 5-HT in particular,resulting in adaptive down-regulation of post-synapticreceptors, such as the 5-HT2A receptor (Gray and Roth,2001; Klimek et al., 1994; Meyer et al., 2001; Peremanset al., 2005). It has therefore been suggested that theeffect of antidepressants on the 5-HT2A receptor

Figure 1 Sagittal 123I-R91150 SPECT images in one dog.Compared to the pre-morphine images (A), 5-HT2A receptoravailability was decreased after 10 days of morphine treatment(B).
Table 2 Morphine plasma concentrations on the lastday of treatment, 3 h after intake (20 mg/kg oral sus-tained release morphine sulfate, twice daily for 10 days).
Dog no. CMorphine (ng/mL)
1 25.32 37.33 19.64 35.35 24.66 40.47 25.8
A. Adriaens et al.1136
contributes to their therapeutic effect (Nelson andPapakostas, 2009; Philip et al., 2008). The results of ourstudy demonstrate a decreased 5-HT2A receptor BI after 10days of morphine treatment. These results are similar to thedecreased 5-HT2A receptor density that was reported indogs after 6-weeks of SSRI treatment, together withreduced impulsive aggression (Peremans et al., 2005). Thisbehavioral improvement associated with the 5-HT2A recep-tor down-regulation underlines the clinical relevance of theobserved findings.
Morphine is a potent mu opioid receptor (MOR) agonistand since MORs and their endogenous ligands, such asendomorphins, are localized in brain regions involved indepression, it has been suggested that the endogenousopioid system could play a role both in the pathogenesisof depression and in patients suffering from treatment-refractory depression (Bodkin et al., 1995; Fichna et al.,2007; Stoll and Reuter, 1999; Tejedor-Real et al., 1995).
The interaction between the serotonergic and opiatesystem has been extensively studied not only in the searchtowards new antidepressants but even more in the field ofanalgesia. Modulation of the serotonergic system has beeninvestigated as a way to enhance the analgesic efficacy of
opioids, and the use of antidepressants is one of the well-established treatment options for chronic pain (Banks et al.,2010; Vadalouca et al., 2006). Additionally, blocking 5-HT2Areceptors restores some of the neurochemical modificationsinduced by the long-term use of drugs of abuse, such asmorphine (Lanteri et al., 2008). Even though our animalswere not physically dependent on morphine, our resultssupport the previously mentioned findings that not onlythere is a serotonergic component in chronic opioid sideeffects, but that, more specifically, the 5-HT2A receptormight be involved as well. Hence, the present findings couldbe of interest for future studies on opioid side effects.
To achieve physical dependence or tolerance to mor-phine, higher dosages are needed (Yoshimura et al., 1993).However, this was not the goal of this study. Moreover, theuse of opioids as potential antidepressant agents would beseverely hindered due to side effects if high doses wererequired. The morphine plasma concentrations determinedin this study correspond to the clinically relevant concen-trations found in literature (Dohoo, 1997).
The exact mechanism by which morphine decreasesthe 5-HT2A receptor availability in our study remains tobe elucidated. SPECT imaging is an excellent technique forvisualizing and semi-quantifying neurotransmitter systemsin vivo; however, it cannot provide tangible answers on theexact mechanisms that underlie the alterations in receptorbinding. Since morphine was administered during 10 days,an increased 5-HT tone could account for an adaptive down-regulation of the 5-HT2A receptors. An acute surge (15 minto 48 h) in 5-HT could not provoke reduced 5-HT2A receptornumbers (Van Oekelen et al., 2001); however, chronic SSRItreatment resulted in decreased 5-HT2A receptor density inboth humans and dogs (Meyer et al., 2001; Peremans et al.,2005). Albeit 5-HT release has been found to be reduced inmorphine dependent rodents (Jolas et al., 2000), as statedpreviously, the dogs used in the present study were notconsidered to be dependent on morphine. This possiblyimplies a different alteration of the serotonergic systemduring opioid dependency compared to prolonged low-doseopioid use.
Another possible explanation for the morphine-induceddecrease in 5-HT2A receptor binding could be an interactionbetween the MOR and 5-HT2A receptor. Both receptors arewidely distributed in the brain (Blue et al., 1988; Tempel andZukin, 1987) and it has been shown that 5-HT2A receptoractivation influences MOR internalization, probably through acommon intracellular pathway (López-Giménez et al., 2008).Thus it might be possible as well that MOR activationinfluences 5-HT2A receptor internalization and down-regulation. Competition between the radioligand and mor-phine for the 5-HT2A receptor could also occur; however, theaffinity of morphine for the 5-HT2A receptor is rather low(Leysen et al., 1982). Possible allosteric changes of the5-HT2A receptors due to morphine binding would also be aninteresting subject for future studies.
In conclusion, prolonged exposure to morphine decreasedthe 5-HT2A receptor availability in a large part of the caninecortex. This finding gives further evidence for the existenceof an interaction between the opioid and 5-HT2A receptorsystem and encourages further research on the possiblebeneficial effects of opioids in the treatment of mooddisorders.

1137The effect of prolonged exposure to morphine on canine cerebral 5-HT2A receptors measured with 123I-R91150 SPECT
Role of funding source
This work was funded by a Special Research Fund Grant from theGhent University (No. 01J06109). The Ghent University had nofurther role in the study design; in the collection, analysis andinterpretation of data; in the writing of the report; and in thedecision to submit the paper for publication.
Conflict of interest
All authors declare no conflict of interest.
Contributors
1/ Contributing to conception and design, or acquiring data, oranalyzing and interpreting data.2/ Drafting the manuscript, or critically contributing to or revisingthe manuscript, or enhancing its intellectual content.3/ Approving the final content of the manuscript.
Antita Adriaens: 1-2Ingeborgh Polis: 1-2-3Simon Vermeire: 1-2Tim Waelbers: 1-2Siska Croubels: 1-2Luc Duchateau: 1-2Sylvia Van Dorpe: 2Jos Eersels: 2Bart De Spiegeleer: 2Kathelijne Peremans: 1-2-3
Acknowledgments
The authors gratefully acknowledge the support of An Maes with theanalysis of morphine in plasma and CSF samples.
References
Adriaens, A., Polis, I., Vermeire, S., Waelbers, T., Duchateau, L., Sys, S.,Van Dorpe, S., Eersels, J., De Spiegeleer, B., Peremans, K., 2012.The influence of morphine on cerebral 5-HT2A receptor availabilityin dogs: a SPECT study. J. Nucl. Med. 53 (12), 1969–1973.
Anonymous, 2002. Commission Decision 2002/657/EC implementingCouncil Directive 96/23/EC concerning the performances ofanalytical methods and the interpretation of results.Official Journal of the European Communities. 17.08.2002,Decision of 12 August 2002, European Commission (DirectorateGeneral for Public Health and Consumers Protection (SANCO):L221/8-36).
Banks, M., Rice, K., Negus, S., 2010. Antinociceptive interactionsbetween mu-opioid receptor agonists and the serotonin uptakeinhibitor clomipramine in rhesus monkeys: role of mu agonistefficacy. J. Pharmacol. Exp. Ther. 335, 497–505.
Blue, M.E., Yagaloff, K.A., Mamounas, L.A., Hartig, P.R., Molliver, M.E.,1988. Correspondence between 5-HT2 receptors and serotonergicaxons in rat neocortex. Brain Res. 453, 315–328.
Bodkin, J.A., Zornberg, G.L., Lukas, S.E., Cole, J.O., 1995.Buprenorphine treatment of refractory depression. J. Clin.Psychopharmacol. 15, 49–57.
Cahir, M., Ardis, T., Reynolds, G.P., Cooper, S.J., 2007. Acute andchronic tryptophan depletion differentially regulate central5-HT1A and 5-HT2A receptor binding in the rat. Psychopharma-cology 190, 497–506.
Catafau, A.M., Danus, M., Bullich, S., Nucci, G., Llop, J., Abanades,S., Cunningham, V.J., Eersels, J., Pavia, J., Farre, M., 2006.Characterization of the SPECT 5-HT2A receptor ligand I-123-R91150 in healthy volunteers: Part 2 – Ketanserin displacement.J. Nucl. Med. 47, 929–937.
Dohoo, S., 1997. Steady-state pharmacokinetics of oral sustained-release morphine sulfate in dogs. J. Vet. Pharmacol. Ther. 20,129–133.
Eersels, J., Travis, M.J., Herscheid, J., 2005. Manufacturing I-123-labelled radiopharmaceuticals. Pitfalls and solutions. J. Label.Compd. Radiopharm. 48, 241–257.
Fichna, J., Janecka, A., Costentin, J., Do Rego, J.C., 2007. Theendomorphin system and its evolving neurophysiological role.Pharmacol. Rev. 59, 88–123.
Gadeyne, C., Van der Heyden, S., Gasthuys, F., Croubels, S.,Schauvliege, S., Polis, I., 2011. The influence of modulation ofP-glycoprotein and/or cytochrome P450 3A on the pharmacoki-netics and pharmacodynamics of orally administered morphinein dogs. J. Vet. Pharmacol. Ther. 34, 417–423.
Gray, J.A., Roth, B.L., 2001. Paradoxical trafficking and regulationof 5-HT(2A) receptors by agonists and antagonists. Brain Res.Bull. 56, 441–451.
Jolas, T., Nestler, E.J., Aghajanian, G.K., 2000. Chronic morphineincreases GABA tone on serotonergic neurons of the dorsal raphenucleus: association with an up-regulation of the cyclic AMPpathway. Neuroscience 95, 433–443.
Klimek, V., Zakknapik, J., Mackowiak, M., 1994. Effects of repeatedtreatment with fluoxetine and citalopram, 5-HT uptake inhibi-tors, on 5-HT1a and 5-HT2 receptors in the rat-brain. J.Psychiatry Neurosci. 19, 63–67.
Lanteri, C., Salomon, L., Torrens, Y., Glowinski, J., Tassin, J.P.,2008. Drugs of abuse specifically sensitize noradrenergic andserotonergic neurons via a non-dopaminergic mechanism. Neu-ropsychopharmacology 33, 1724–1734.
Leysen, J.E., Niemegeers, C.J., Van Nueten, J.M., Laduron, P.M.,1982. [3H]Ketanserin (R 41 468), a selective 3H-ligand forserotonin2 receptor binding sites. Binding properties, braindistribution, and functional role. Mol. Pharmacol. 21, 301–314.
López-Giménez, J.F., Vilaro, M.T., Milligan, G., 2008. Morphinedesensitization, internalization, and down-regulation of the muopioid receptor is facilitated by serotonin 5-hydroxytryptamine2Areceptor coactivation. Mol. Pharmacol. 74, 1278–1291.
Manji, H.K., Moore, G.J., Rajkowska, G., Chen, G., 2000. Neuro-plasticity and cellular resilience in mood disorders. Mol. Psy-chiatry 5, 578–593.
Marek, G.J., Carpenter, L.L., McDougle, C.J., Price, L.H., 2003.Synergistic action of 5-HT2A antagonists and selective serotoninreuptake inhibitors in neuropsychiatric disorders. Neuropsycho-pharmacology 28, 402–412.
Mertens, J., Terriere, D., Sipido, V., Gommeren, W., Janssen, P.M.F.,Leysen, J.E., 1994. Radiosynthesis of a new radioiodinated ligand forserotonin-5HT2-receptors, a promising tracer for gamma-emissiontomography. J. Label. Compd. Radiopharm. 34, 795–806.
Meyer, J.H., Kapur, S., Eisfeld, B., Brown, G.M., Houle, S., Da Silva, J.,Wilson, A.A., Rafi-Tari, S., Mayberg, H.S., Kennedy, S.H., 2001.The effect of paroxetine on 5-HT2A receptors in depression: an[F-18]setoperone PET imaging study. Am. J. Psychiatry 158,78–85.
Nelson, J.C., Papakostas, G.I., 2009. Atypical antipsychoticaugmentation in major depressive disorder: a meta-analysis ofplacebo-controlled randomized trials. Am. J. Psychiatry 166,980–991.
Ostroff, R.B., Nelson, J.C., 1999. Risperidone augmentation ofselective serotonin reuptake inhibitors in major depression. J.Clin. Psychiatry 60, 256–259.
Pazos, A., Probst, A., Palacios, J.M., 1987. Serotonin receptors inthe human brain–IV. Autoradiographic mapping of serotonin-2receptors. Neuroscience 21, 123–139.

A. Adriaens et al.1138
Peremans, K., Audenaert, K., Jacobs, F., Dumont, F., De Vos, F.,Van De Wiele, C., Vandecapelle, M., van Bree, H., Verschooten, F.,Slegers, G., Mertens, J., Dierckx, R., 2002. Biodistribution anddisplacement studies of the selective 5-HT2A receptor antagonist123I-5-I-R91150 in the normal dog. Nucl. Med. Commun. 23,1019–1027.
Peremans, K., Audenaert, K., Coopman, F., Jacobs, F., Dumont, F.,Slegers, G., Verschooten, F., van Bree, H., Mertens, J., Dierckx, R.,2003a. Regional binding index of the radiolabeled selective 5-HT2Aantagonist 123I-5-I-R91150 in the normal canine brain imaged withsingle photon emission computed tomography. Vet. Radiol. Ultra-sound 44, 344–351.
Peremans, K., Audenaert, K., Hoybergs, Y., Otte, A., Goethals, I.,Gielen, I., Blanckaert, P., Vervaet, M., van Heeringen, C.,Dierckx, R., 2005. The effect of citalopram hydrobromide on5-HT2A receptors in the impulsive–aggressive dog, as measuredwith 123I-5-I-R91150 SPECT. Eur. J. Nucl. Med. Mol. Imaging 32,708–716.
Philip, N.S., Carpenter, L.L., Tyrka, A.R., Price, L.H., 2008.Augmentation of antidepressants with atypical antipsychotics:a review of the current literature. J. Psychiatr. Pract. 14,34–44.
Shelton, R.C., Sanders-Bush, E., Manier, D.H., Lewis, D.A., 2009.Elevated 5-HT 2A receptors in postmortem prefrontal cortex inmajor depression is associated with reduced activity of proteinkinase A. Neuroscience 158, 1406–1415.
Sohlberg, A., Watabe, H., Iida, H., 2008. Three-dimensional SPECTreconstruction with transmission-dependent scatter correction.Ann. Nucl. Med. 22, 549–556.
Stahl, S.M., 2008. Mood disorders. In: Stahl, S.M. (Ed.), Stahl'sEssential Psychopharmacology: Neuroscientific Basis and PracticalApplications 3rd ed. Cambridge University Press, New York,pp. 453–510.
Stoll, A.L., Rueter, S., 1999. Treatment augmentation with opiatesin severe and refractory major depression. Am. J. Psychiatry156, 2017.
Tao, R., Auerbach, S.B., 1994. Increased extracellular serotonin inrat brain after systemic or intraraphe administration of mor-phine. J. Neurochem. 63, 517–524.
Tejedor-Real, P., Mico, J.A., Maldonado, R., Roques, B.P., Giber-trahola, J., 1995. Implication of endogenous opioid system inthe learned helplessness model of depression. Pharmacol.Biochem. Behav. 52, 145–152.
Tempel, A., Zukin, R.S., 1987. Neuroanatomical patterns of the mu,delta, and kappa opioid receptors of rat brain as determined byquantitative in vitro autoradiography. Proc. Natl. Acad. Sci. USA84, 4308–4312.
Vadalouca, A., Siafaka, I., Argyra, E., Vrachnou, E., Moka, E., 2006.Therapeutic management of chronic neuropathic pain – anexamination of pharmacologic treatment. Ann. N. Y. Acad. Sci.1088, 164–186.
Van Oekelen, D., Jurzak, M., Van de Wiel, D., Van Hecke, G.,Luyten, W.H., Leysen, J.E., 2001. Different regulation of rat5-HT(2A) and rat 5-HT(2C) receptors in NIH 3T3 cells uponexposure to 5-HT and pipamperone. Eur. J. Pharmacol. 425, 21–32.
Yoshimura, K., Horiuchi, M., Konishi, M., Yamamoto, K., 1993.Physical-dependence on morphine induced in dogs via the use ofminiosmotic pumps. J. Pharmacol. Toxicol. Methods 30, 85–95.

![Striatal amphetamine-induced dopamine release in patients with schizotypal personality disorder studied with single photon emission computed tomography and [123I]iodobenzamide](https://static.fdokumen.com/doc/165x107/631cb9a45a0be56b6e0e579d/striatal-amphetamine-induced-dopamine-release-in-patients-with-schizotypal-personality.jpg)


















