
The effect of landscape-scale environmental drivers on the vegetation composition of British...
15
The effect of landscape-scale environmental drivers on the vegetation composition of British woodlands P.M. Corney a, * , M.G. Le Duc a , S.M. Smart b , K.J. Kirby c , R.G.H. Bunce b,1 , R.H. Marrs a a Applied Vegetation Dynamics Laboratory, School of Biological Sciences, University of Liverpool, P.O. Box 147, Liverpool L69 3GS, UK b Merlewood Research Station, Centre for Ecology and Hydrology, Grange-over-sands, Cumbria LA11 6JU, UK c English Nature, Northminster House, Peterborough PE1 1UA, UK Received 4 September 2003; received in revised form 16 March 2004; accepted 26 March 2004 Abstract Assessment of factors influencing woodland vegetation composition across Britain was made using multivariate techniques to analyse data gathered during the 1971 National Woodland Survey. Indirect gradient analysis (unconstrained ordination using detrended correspondence analysis) suggested a gradient strongly associated with nutrient availability and pH. Direct gradient analysis (constrained ordination using canonical correspondence analysis) and variation partitioning were used with over 250 ecophysiologically relevant variables, including climatic, geographical, soil and herbivore data, to model the response of woodland vegetation. Although there was a high degree of multicollinearity between environmental variables, analysis revealed the vegetation composition of surveyed woodlands to be primarily structured by geographical, climatic and soil gradients, in particular rainfall, soil pH and accumulated temperature. The woods have recently been resurveyed. The results of this analysis therefore provide a baseline against which species dynamics can be assessed under a series of conservation threats, such as land use and climate change. Ó 2004 Elsevier Ltd. All rights reserved. Keywords: National Woodland Survey; Environmental factors; Vegetation analysis; Canonical correspondence analysis; Variation partitioning 1. Introduction Successful implementation of local and national woodland conservation management goals requires an understanding of the way in which key environmental factors influence the ability of species to persist. Extant classifications for woodlands in Britain (e.g. Bunce, 1982, 1989; Rodwell et al., 1991; Peterken, 1993; Rackham, 2003) provide useful information regarding species composition, history and management, while Bunce and Shaw (1975) and Bunce (1981) assess the principal gradients behind woodland vegetation in Britain. However, little has been done to quantify the relative contribution of key environmental variables to the determination of woodland species composition. In this paper, we explore the relative impact of climate (e.g. temperature, precipitation, atmospheric pollution), site factors (e.g. altitude, edaphic factors) and land use on woodland composition, using vegetation data collected in 1971. We thus provide a baseline against which to assess changes under a series of conservation threats, such as land use change, global warming and exotic species invasion. In fragmented landscapes, woodlands may act as refugia, harbouring rare woodland plant communities, which can act as a source of propagules for estab- lishment of new communities in the wider landscape. Since both persistence of species and dispersal ability * Corresponding author. Tel.: +44-151-794-4775; fax: +44-151-794- 4940. E-mail address: [email protected] (P.M. Corney). 1 Present address: Alterra, Landscape and Spatial Planning Section, P.O. Box 47, 6700AA Wageningen, The Netherlands. 0006-3207/$ - see front matter Ó 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.biocon.2004.03.022 Biological Conservation 120 (2004) 491–505 www.elsevier.com/locate/biocon BIOLOGICAL CONSERVATION
-
Upload
greenriver -
Category
Documents
-
view
1 -
download
0
Transcript of The effect of landscape-scale environmental drivers on the vegetation composition of British...

BIOLOGICAL
CONSERVATION
Biological Conservation 120 (2004) 491–505
www.elsevier.com/locate/biocon
The effect of landscape-scale environmental drivers on thevegetation composition of British woodlands
P.M. Corney a,*, M.G. Le Duc a, S.M. Smart b, K.J. Kirby c, R.G.H. Bunce b,1,R.H. Marrs a
a Applied Vegetation Dynamics Laboratory, School of Biological Sciences, University of Liverpool, P.O. Box 147, Liverpool L69 3GS, UKb Merlewood Research Station, Centre for Ecology and Hydrology, Grange-over-sands, Cumbria LA11 6JU, UK
c English Nature, Northminster House, Peterborough PE1 1UA, UK
Received 4 September 2003; received in revised form 16 March 2004; accepted 26 March 2004
Abstract
Assessment of factors influencing woodland vegetation composition across Britain was made using multivariate techniques to
analyse data gathered during the 1971 National Woodland Survey. Indirect gradient analysis (unconstrained ordination using
detrended correspondence analysis) suggested a gradient strongly associated with nutrient availability and pH. Direct gradient
analysis (constrained ordination using canonical correspondence analysis) and variation partitioning were used with over 250
ecophysiologically relevant variables, including climatic, geographical, soil and herbivore data, to model the response of woodland
vegetation. Although there was a high degree of multicollinearity between environmental variables, analysis revealed the vegetation
composition of surveyed woodlands to be primarily structured by geographical, climatic and soil gradients, in particular rainfall, soil
pH and accumulated temperature. The woods have recently been resurveyed. The results of this analysis therefore provide a baseline
against which species dynamics can be assessed under a series of conservation threats, such as land use and climate change.
� 2004 Elsevier Ltd. All rights reserved.
Keywords: National Woodland Survey; Environmental factors; Vegetation analysis; Canonical correspondence analysis; Variation partitioning
1. Introduction
Successful implementation of local and nationalwoodland conservation management goals requires an
understanding of the way in which key environmental
factors influence the ability of species to persist. Extant
classifications for woodlands in Britain (e.g. Bunce,
1982, 1989; Rodwell et al., 1991; Peterken, 1993;
Rackham, 2003) provide useful information regarding
species composition, history and management, while
* Corresponding author. Tel.: +44-151-794-4775; fax: +44-151-794-
4940.
E-mail address: [email protected] (P.M. Corney).1 Present address: Alterra, Landscape and Spatial Planning Section,
P.O. Box 47, 6700AA Wageningen, The Netherlands.
0006-3207/$ - see front matter � 2004 Elsevier Ltd. All rights reserved.
doi:10.1016/j.biocon.2004.03.022
Bunce and Shaw (1975) and Bunce (1981) assess the
principal gradients behind woodland vegetation in
Britain. However, little has been done to quantify therelative contribution of key environmental variables to
the determination of woodland species composition. In
this paper, we explore the relative impact of climate (e.g.
temperature, precipitation, atmospheric pollution), site
factors (e.g. altitude, edaphic factors) and land use on
woodland composition, using vegetation data collected
in 1971. We thus provide a baseline against which to
assess changes under a series of conservation threats,such as land use change, global warming and exotic
species invasion.
In fragmented landscapes, woodlands may act as
refugia, harbouring rare woodland plant communities,
which can act as a source of propagules for estab-
lishment of new communities in the wider landscape.
Since both persistence of species and dispersal ability

492 P.M. Corney et al. / Biological Conservation 120 (2004) 491–505
are significantly affected by environmental conditions,
it is important for the purposes of conservation sci-
ence that the effects of biotic and abiotic factors be
quantified.
Current factors affecting woodland composition anddistribution are diverse and their interactions complex.
They range from factors such as climate (Cannell,
2002; Williams et al., 2002) and topography (Mod-
rzynski and Eriksson, 2002), soil parent material and
time of soil development (Jenny, 1980), to management
(Brunet et al., 1996) and grazing by native ungulates,
such as deer (Kirby, 2001). Moreover, it is becoming
apparent that anthropogenic activity leading to climatechange (e.g. Bakkenes et al., 2002; Houghton et al.,
2001; Lasch et al., 2002; Mitchell and Karoly, 2001),
nitrogen deposition and acidification (Bobbink et al.,
1998; Brunet et al., 1998; Hofmeister et al., 2002) may
alter plant distribution, as species climatic envelopes
begin to shift geographically. Modification of local
climatic optima has particular significance for conser-
vation of woodland habitats in Britain, as it has beensuggested that a large proportion (30%) of woodland
plant species may be unable to colonise suitable target
habitats in fragmented areas after a period of 30–40
years (Honnay et al., 2002).
Thus, although management can profoundly effect
woodland vegetation distribution, assessing the true
impact of such can be problematic, as continental, re-
gional, countrywide and local scale environmental fac-tors also exert a considerable influence on the
performance of both higher and lower plants (Furness
and Grime, 1982; Redfern and Hendry, 2002; Yeo and
Blackstock, 2002). Accordingly, this paper reports a
study which aimed to partition the range of variation
within a sample of semi-natural woodlands in Britain in
terms of factors that operate at a range of scales (site to
countrywide) and through a variety of mechanisms(natural to anthropogenic, direct to diffuse), and are
likely to be important in driving the distribution and
composition of native woodlands.
The analysis presented here is based upon data col-
lected during the National Woodland Survey (NWS),
carried out by the Nature Conservancy of Britain in the
summer of 1971 using a standardised methodology
(Bunce and Shaw, 1973; Smart et al., 2001). Whilst thisproject was primarily about classification, the objectives
written in 1969 also specified examining principal un-
derlying environmental factors. Environmental data not
collected during the original survey but likely to be
important in controlling woodland species composition
were obtained from a variety of sources at the site scale.
Multivariate analyses were used to describe variation in
the dataset in relation to selected significant environ-mental variables and relative contribution of selected
groups of environmental variables was assessed using
variation partitioning.
2. Methods
2.1. National Woodland Survey site selection
In order to cover the range of variation within semi-natural woodlands, selection of sites in 1971 was made
from a survey of 2463 woodland sites (>4.05 ha) car-
ried out for the Nature Conservation Review (Ratcliffe,
1977). This site series represented approximately a 10%
sample of the population of semi-natural woodlands in
Britain (Bunce and Shaw, 1972, 1973). Full historical
background is given by Sheail and Bunce (2003). As-
sociation analysis (Williams and Lambert, 1959) per-formed on this dataset divided the 2463 sites into 103
groups defined by their similarity of plant species
composition (Hill et al., 1975). Numerical analysis of
topographical and climatic data was then used to select
a representative site from each group (Bunce and
Shaw, 1973). These 103 sites were surveyed for NWS
1971. Site locations are shown in Fig. 1, grouped into
Countryside Vegetation System (CVS) classes (Barret al., 1993; Bunce et al., 1999) generated from site
species data using the Modular Analysis of Vegetation
Information System (MAVIS) Plot Analyser, version
1.0 (Smart, 2000).
2.2. NWS methodology
For each of the 103 sites, a sample size of 16 pointswas fixed and marked on 1:25,000 Ordnance Survey
(OS) site maps prior to survey within the delin-
eated woodland area, representing the location of sur-
vey plots (Bunce and Shaw, 1973). Survey of all 1648
14.14� 14.14 m (200 m2) plots was carried out between
June and October 1971 by separate teams of trained
surveyors (Bunce and Shaw, 1973; Hill et al., 1975).
Estimates of cover (5% classes) were made across each200 m2 plot, for individual ground flora species, along
with six additional ground cover categories; bryophyte
cover, litter, dead wood material, rock, bare ground and
standing water (Smart et al., 2001). Woody species
(trees, saplings and shrubs) and bryophyte species
present within each plot were also assessed. Plot slope
and aspect were recorded, along with plot- and site-scale
descriptions, including signs of seven groups of herbiv-orous mammals (sheep, red deer, other deer species,
cattle, horses/ponies, rabbits, squirrels) and boundary
types such as intact walls, ditches and derelict fences. A
single soil sample was taken from each plot for assess-
ment of pH and loss-on-ignition. Diagnostic soil profile
details were recorded in the field (Bunce and Shaw,
1973) and soil subgroup classes were assigned to each
soil sample using Avery’s (1980) classification system.For further description of NWS methods, see Bunce and
Shaw (1973). Table 1 gives a breakdown of those vari-
ables available for, and used in, the present analysis.

Fig. 1. Distribution of sites surveyed in the 1971 National Woodland Survey of Britain. Sites are illustrated according to Countryside Vegetation
System (Barr et al., 1993; Bunce et al., 1999) classes.
P.M. Corney et al. / Biological Conservation 120 (2004) 491–505 493
2.3. Use of NWS botanical data
To facilitate examination of between-site variation,
within-site variation was explicitly omitted by aggre-
gating botanical data for all 16 plots per site. Although
cover data were available for field layer species, data for
the complete vegetation complement (bryophytes, field,
sapling, shrub and tree layer species) were only availableas presence/absence. Analysis therefore proceeded on
the basis of full botanical complement of species present
within each site as present or absent only.
2.4. Potential environmental drivers of woodland compo-
sition
One hundred eighty six additional environmental
variables likely to affect woodland species composi-tion were assembled and amalgamated into groups

Table 1
Description of the nature and transformation of variable sets obtained from National Woodland Survey 1971 data and used in the analysis presented
here
Variable set Description of original data T Description of data used in analysis V No.
Animals Plot level presence or absence
(sight, signs, sounds) of seven
herbivorous mammal species
s Intensity of herbivore grazing pressure by site d 7
Site boundary type Site level presence or absence of different
boundary types
u Site level presence or absence of different
boundary types
b 17
Date Month during which woodland was
surveyed
u Month surveyed. Used as five dummy
variables.
b 1 (5)
Edaphic Plot level soil pH; LOI (%); depth to each soil
horizon (A00, litter layer; A0, organic layer;
A1, mixed mineral/organic layer; A2, leached or
eluviated layer; B, weathered mineral layer)
present (cm)
a Site level soil pH; c 7
g, t Site level LOI;
a Site level cumulative depth to the
bottom of five soil horizons (cm)
Geographic,
regionality
National Grid easting and northing (km)
of site datum
u Easting and northing (km) of site datum c 2
Geographic,
micro-climatic
Plot level slope, the steepest gradient passing
through plot centre (deg) and aspect (deg)
a Average slope of site (deg); c 3
g Transformation (s) of aspect (a, degrees,the site-wise mean), s ¼ sinða� p=180Þ=2,giving a site level southerly aspect;
g Transformation (w) of aspect (a, degrees,the site-wise mean),
w ¼ j sinðða� p=180Þ � p=2Þ=2Þj, giving a site
level westerly aspect
Ground cover
categories
Plot level cover (%) of ground cover estimated
within 14.14� 14.14 m quadrats
g, t Site level estimate of six ground cover types
(bryophyte cover, litter, dead wood material,
rock, bare ground and standing water)
c 6
Soil classification Soil sub groups present in each plot.
Where more than one subgroup occurred
in each plot, the mosaic was assumed
to comprise equal amounts of each constituent
subgroup
a, t Arithmetic average of proportion of
subgroups present in plots, giving the
proportion of 26 soil groups present
within each site;
c 33
a, t As above, giving the proportion of seven
major soil groups present within each site
T – transformation used: a, arithmetic average of plot data by site; g, geometric average of plot data by site; s, sum of plot data by site; t, arcsine
transformation of data (Sokal and Rohlf, 1995); u, untransformed site level data.
V– variable type: b, binary; c, continuous; d, discrete.
No. – number of variables per set used in this analysis.
494 P.M. Corney et al. / Biological Conservation 120 (2004) 491–505
containing landscape, spatial, anthropogenic and cli-
matic variables. Data for 33 variables, the Centre for
Ecology and Hydrology (CEH) satellite Land Cover
Map 2000 (LCM 2000) classes, woodland spatial vari-
ables, nitrogenous deposition data and distance to the
sea, were only available at the site level. However for
other variables (e.g. altitude), plot level data were ob-
tained using plot OS grid references generated using thesoftware package ERDAS IMAGINE version 8.5
(ERDAS, 2001). These results were then aggregated for
each site, to produce an appropriate mean site value for
each variable. Using a full series of 103 digital site map
images, ERDAS IMAGINE was also employed to cal-
culate length of delineated woodland perimeter and
area. These measures of woodland perimeter (Pw) (km)
and area (Aw) (ha) were then used to generate twowoodland shape indices, a perimeter index, after Hinsley
et al. (1995), and an area index. The perimeter index was
calculated as PwPc, where Pc was the perimeter of a hy-
pothetical circular site of the same area. Thus the pe-
rimeter index of a wood is high for woods with scalloped
or jagged edges. The area index was described using an
index calculated as Aw=Pw. This second index contrasts
circular woodlands with those that tend towards a more
ellipsoidal shape. Presence and effect of possible spatial
autocorrelation was assessed using seven terms of qua-dratic and cubic trend surface, derived from site geo-
graphical co-ordinates (e.g. Yeo and Blackstock, 2002).
Distance to the sea, calculated as the minimum dis-
tance from site OS datum point to the nearest coast was
computed with MINDIST2, a FORTRAN77 program
using a set of digitised UK coastal co-ordinates, com-
prising 1550 points (Le Duc et al., 2000).
Plot altitude was extracted from Land-Form PAN-ORAMA Digital Terrain Model contour maps held on
the OS DIGIMAP service (EDINA, 2002), using OS

P.M. Corney et al. / Biological Conservation 120 (2004) 491–505 495
plot co-ordinates generated as outlined above, MAP
MANAGER version 6.2 (ESRI, 2001), ARCVIEW GIS
version 3.2a (ESRI, 2000), and the Avenue Script
GETGRIDVALUE (Elmquist and Davies, 1999).
Climatic data were drawn from two sources: theMeteorological Office and the Forestry Commission.
Monthly and yearly Meteorological Office 5 km� 5 km
grid baseline data were acquired for generated plot co-
ordinates from long term average (LTA) datasets for the
period 1961 to 1990 and 1961 to 2000, respectively,
produced in association with the UK Climate Impacts
Programme, UKCIP (Meteorological Office, 2003). This
Table 2
Description of the 186 additional environmental variables generated or colle
Variable set T Description
Site form, shape u Woodland area (ha); woo
woodland area index (Aw
Spatial u Site (East)2 ; (East)3; (No
(North)2 �East
Geographic u Distance from site to nea
a Altitude (m)
Surrounding landscape u Area (ha) of broad habita
set-aside land, inland roc
Pollution u Site level deposition of N
total N deposition (kgNh
Woodland site conditions
1961–1990 LTA data
a Forest wind climate, calc
resolution; annual accum
moisture deficit (accumul
squares
Climatic monthly 1961–1990 LTA data a Mean monthly daily max
temperature (�C); days of%); total monthly bright
mm (rain days); number o
monthly precipitation (m
Climatic seasonal 1961–1990 LTA
data (accumulated monthly data)
a, s Mean seasonal daily max
temperature (�C); days ofto %); total seasonal brig
P1 mm (rain days); num
total seasonal precipitatio
Climatic annual 1961–1990 LTA
data (Accumulated monthly data)
a, y Mean yearly daily maxim
temperature (�C); days ofto %); total yearly bright
P1 mm (rain days); num
total yearly precipitation
Climatic yearly 1961–2000 LTA data a Annual extreme temperat
(�C); number of growing
negatives, over the summ
temperature being >5 �Cdays; maximum number o
a year; greatest five day p
on rain days (total rain o
of rain days)
All variables are continuous.
Nitrogen deposition data extracted from deposition maps compiled by C
Woodland site condition data obtained from ESC decision support syste
All other climatic data acquired from the UK Meteorological Office 5 km
association with UK Climate Impact Programme.
T – transformation used: a, arithmetic average of plot data by site; s, a
(winter¼Dec, Jan, Feb; spring¼Mar, Apr, May; summer¼ Jun, Jul, Aug
generate a further eight annual variables; u, untransformed site level data.
No. – number of variables in set.
information is drawn from interpolation models created
from a large network of weather stations across the
country (approximately 3500 stations for rainfall and
500 stations for most other parameters). Data for six
yearly LTA variables (1961–2000) were used, includingnumber of growing degree days and maximum number
of consecutive dry days in a year, along with data for
eight monthly variables (1961 to 1990), such as mean
daily minimum temperature and days of ground frost,
giving 96 variables. Where data for specific LTA vari-
ables were available in such monthly blocks, these were
also accumulated to give a further 32 seasonal and eight
cted for the present study
No.
dland perimeter (m); shape indices,
=Pw); woodland perimeter index (Pw=Pc)4
rth)2; (North)3 ; East�North; (East)2 �North; 7
rest coast (km) 2
t classes (e.g. broadleaved woodland,
k, etc) surrounding site datum
25
Ox (kgNha�1 y�1); deposition of NHx (kgNha�1 y�1);
a�1 y�1)
3
ulated using tatter flags and digital elevation models, 100 m
ulated temperature (degree days/30), 10 km grid squares;
ated excess of evaporation minus rainfall), 10 km grid
3
imum temperature (�C); mean monthly daily minimum
ground frost; mean monthly cloud cover (Oktas converted to
sunshine (h); number of days per month having a rainfall P1
f days per month having a rainfall P10 mm (wet days); total
m)
96
imum temperature (�C); mean seasonal daily minimum
ground frost; mean seasonal cloud cover (Oktas converted
ht sunshine (h); number of days per month having a rainfall
ber of days per month having a rainfall P10 mm (wet days);
n (mm)
32
um temperature (�C); mean yearly daily minimum
ground frost; mean yearly cloud cover (Oktas converted
sunshine (h); number of days per month having a rainfall
ber of days per month having a rainfall P10 mm (wet days);
(mm)
8
ure range (highest daily maximum – lowest daily maximum)
degree days, sum of (daily mean temperature – 4), ignoring
er months; growing season length, bounded by daily mean
for >five consecutive days and <5 �C for >five consecutive
f consecutive dry days (days with less than 1 mm of rain) in
recipitation total in a year (mm); mean rainfall amount (mm)
n rain days/number
6
EH Edinburgh.
m (Ray, 2001).
� 5 km grid baseline datasets for the period 1961–2000 produced in
rithmetic average of monthly data to generate 32 seasonal variables
; autumn¼ Sep, Oct, Nov); y, arithmetic average of monthly data to

496 P.M. Corney et al. / Biological Conservation 120 (2004) 491–505
annual values. Woodland site condition data were ob-
tained from the Ecological Site Classification (ESC)
decision support system published by the Forestry
Commission (Ray, 2001), comprising a windiness score,
accumulated temperature and moisture deficit data forthe 30-year period 1961–1990.
Oxidised (NOx) and reduced (NHx) nitrogen (N)
deposition data (sum of wet, dry and cloud deposition)
for surveyed woodlands were extracted from 5� 5 km
resolution national deposition maps (1995–1997) com-
piled by CEH Edinburgh and derived using methods
described in NEGTAP (2001). NOx and NHx were
summed to generate total N deposition entered as athird pollutant variable in analyses. Although a measure
of total N deposition generated in this way is unlikely to
be independent of its constituent parts, this facilitated
an examination of possible responses to total N load.
Current landscape-scale habitat data constructed
from satellite images were obtained from CEH LCM
2000 (Fuller et al., 2002). LCM subclass level-two hab-
itats found within a circular area of radius of 1500 msurrounding each site OS datum were selected. Table 2
gives a summary of additional environmental variables
used.
2.5. Statistical analyses
All statistical analyses were carried out using CA-
NOCO for Windows version 4.5 (Ter Braak and �Smil-auer, 2002), using default settings and untransformed
species data, unless otherwise specified.
Detrended Correspondence Analysis (DCA) was used
to obtain estimates of gradient lengths in standard de-
viation (SD) units of species turnover (Ter Braak and�Smilauer, 2002), thereby assisting in the decision of
whether to use a linear or unimodal approach to the
data.Principle Components Analysis (PCA) was employed
with both original survey variables (Table 1) and addi-
tional variables (Table 2) for preliminary exploration of
environmental factors only.
The full range of variables was then used, either as
environmental variables or covariables, in different
combinations, along with botanical data, to assess the
influence of environmental factors on vegetation varia-tion, using the CANOCO Canonical Correspondence
Analysis (CCA) forward selection procedure. Forward
selection was used to select significant variables, the
Monte Carlo test being used to assess significance, with
499 permutations for exploratory analyses and 9999 for
final results (Legendre and Legendre, 1998). In all per-
mutation tests, an unrestricted permutation structure
was used. This process was combined with an exami-nation of inflation values, to remove those variables that
were highly multicollinear. If an inflation factor is large,
the variable is likely to be strongly correlated with other
variables (Ter Braak and �Smilauer, 2002). As a result,
the effects of different environmental variables on com-
munity composition cannot be separated out and con-
sequently, canonical coefficients are unstable and do not
merit interpretation (Ter Braak, 1986). Therefore, vari-ables exhibiting high inflation values coupled with low
or non-significance were removed and analysis re-run,
until a set of significant variables was obtained, with
inflation factors of <15. The final model contained 20
variables.
Significant variables selected were divided into three
clearly defined and ecologically meaningful sets; biotic
factors, soil properties and geographical with climatic(geo-climatic) variables. Variation partitioning was
carried out using a standardised procedure (e.g. Borcard
et al., 1992; Marrs and Le Duc, 2000; Yeo and Black-
stock, 2002), variance in the data being partitioned be-
tween these sets and expressed as a percentage of the
total variation explained.
3. Results
3.1. Description of survey sites
Site summary data are presented in Table 3 according
to CVS vegetation classes (Barr et al., 1993; Bunce et al.,
1999; Smart, 2000). The sites covered an extremely wide
geographic range (Fig. 1) and showed considerablephysiographic variability (Table 3), while the wide
range of vegetation types encountered reflects the high
b-diversity within the dataset.
3.2. Range of variation in native semi-natural woodland
communities
Preliminary DCA produced an ordination with a firstaxis gradient length of 3.266 SD units, demonstrating
high b-diversity and suggesting that a unimodal model
such as CCA would adequately describe the relationship
between species and environmental variables (Ter Braak
and Prentice, 1988; Ter Braak and �Smilauer, 2002).
Nevertheless, initial CCA results described using
CANODRAW (Ter Braak and �Smilauer, 2002) revealed
an apparent distortion. Subsequent data explorationrevealed the distortion to be due to a large number (151)
of species, each of which occurred only once within the
dataset (25% of all species recorded) and one outlying
site. As CCA is sensitive to deviant sites when they are
outliers with regard to both species composition and
environment, the 151 species were made supplementary
in further analysis, as recommended by Ter Braak and
Prentice (1988). Species made passive in this way do notinfluence ordination axes, but are added to ordination
diagrams so that their relationship to other species can
be examined. Rare species were also down-weighted,
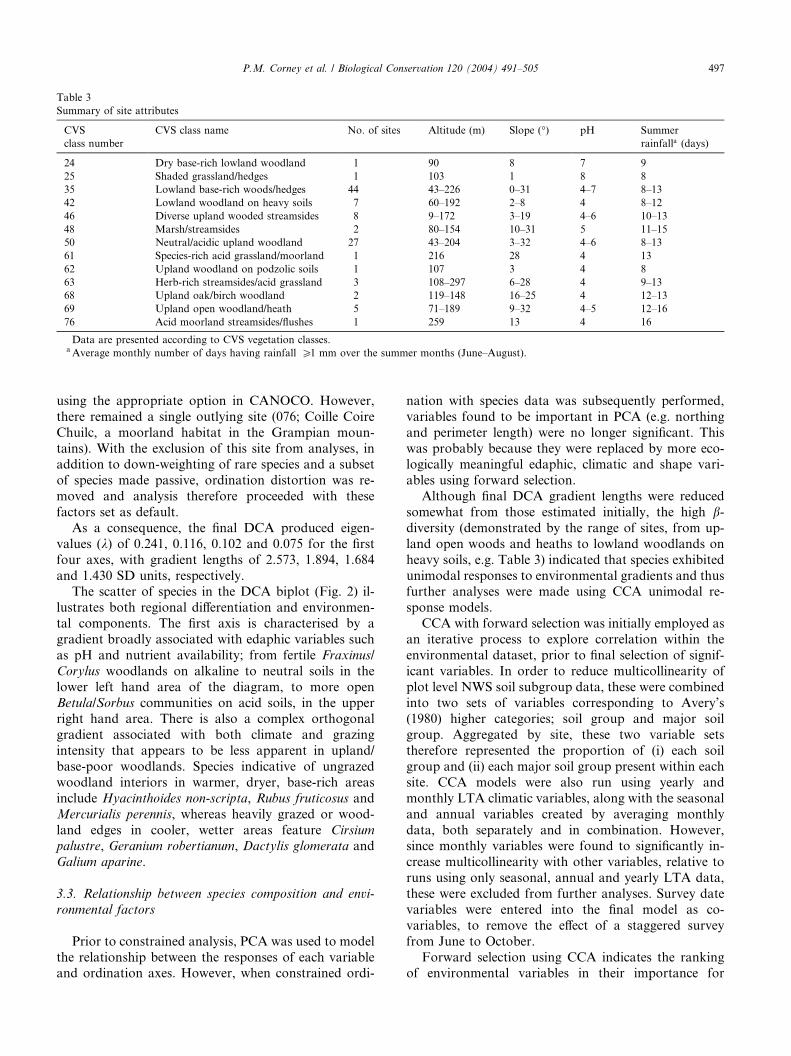
Table 3
Summary of site attributes
CVS
class number
CVS class name No. of sites Altitude (m) Slope (�) pH Summer
rainfalla (days)
24 Dry base-rich lowland woodland 1 90 8 7 9
25 Shaded grassland/hedges 1 103 1 8 8
35 Lowland base-rich woods/hedges 44 43–226 0–31 4–7 8–13
42 Lowland woodland on heavy soils 7 60–192 2–8 4 8–12
46 Diverse upland wooded streamsides 8 9–172 3–19 4–6 10–13
48 Marsh/streamsides 2 80–154 10–31 5 11–15
50 Neutral/acidic upland woodland 27 43–204 3–32 4–6 8–13
61 Species-rich acid grassland/moorland 1 216 28 4 13
62 Upland woodland on podzolic soils 1 107 3 4 8
63 Herb-rich streamsides/acid grassland 3 108–297 6–28 4 9–13
68 Upland oak/birch woodland 2 119–148 16–25 4 12–13
69 Upland open woodland/heath 5 71–189 9–32 4–5 12–16
76 Acid moorland streamsides/flushes 1 259 13 4 16
Data are presented according to CVS vegetation classes.aAverage monthly number of days having rainfall P1 mm over the summer months (June–August).
P.M. Corney et al. / Biological Conservation 120 (2004) 491–505 497
using the appropriate option in CANOCO. However,
there remained a single outlying site (076; Coille Coire
Chuilc, a moorland habitat in the Grampian moun-
tains). With the exclusion of this site from analyses, inaddition to down-weighting of rare species and a subset
of species made passive, ordination distortion was re-
moved and analysis therefore proceeded with these
factors set as default.
As a consequence, the final DCA produced eigen-
values (k) of 0.241, 0.116, 0.102 and 0.075 for the first
four axes, with gradient lengths of 2.573, 1.894, 1.684
and 1.430 SD units, respectively.The scatter of species in the DCA biplot (Fig. 2) il-
lustrates both regional differentiation and environmen-
tal components. The first axis is characterised by a
gradient broadly associated with edaphic variables such
as pH and nutrient availability; from fertile Fraxinus/
Corylus woodlands on alkaline to neutral soils in the
lower left hand area of the diagram, to more open
Betula/Sorbus communities on acid soils, in the upperright hand area. There is also a complex orthogonal
gradient associated with both climate and grazing
intensity that appears to be less apparent in upland/
base-poor woodlands. Species indicative of ungrazed
woodland interiors in warmer, dryer, base-rich areas
include Hyacinthoides non-scripta, Rubus fruticosus and
Mercurialis perennis, whereas heavily grazed or wood-
land edges in cooler, wetter areas feature Cirsium
palustre, Geranium robertianum, Dactylis glomerata and
Galium aparine.
3.3. Relationship between species composition and envi-
ronmental factors
Prior to constrained analysis, PCA was used to model
the relationship between the responses of each variableand ordination axes. However, when constrained ordi-
nation with species data was subsequently performed,
variables found to be important in PCA (e.g. northing
and perimeter length) were no longer significant. This
was probably because they were replaced by more eco-logically meaningful edaphic, climatic and shape vari-
ables using forward selection.
Although final DCA gradient lengths were reduced
somewhat from those estimated initially, the high b-diversity (demonstrated by the range of sites, from up-
land open woods and heaths to lowland woodlands on
heavy soils, e.g. Table 3) indicated that species exhibited
unimodal responses to environmental gradients and thusfurther analyses were made using CCA unimodal re-
sponse models.
CCA with forward selection was initially employed as
an iterative process to explore correlation within the
environmental dataset, prior to final selection of signif-
icant variables. In order to reduce multicollinearity of
plot level NWS soil subgroup data, these were combined
into two sets of variables corresponding to Avery’s(1980) higher categories; soil group and major soil
group. Aggregated by site, these two variable sets
therefore represented the proportion of (i) each soil
group and (ii) each major soil group present within each
site. CCA models were also run using yearly and
monthly LTA climatic variables, along with the seasonal
and annual variables created by averaging monthly
data, both separately and in combination. However,since monthly variables were found to significantly in-
crease multicollinearity with other variables, relative to
runs using only seasonal, annual and yearly LTA data,
these were excluded from further analyses. Survey date
variables were entered into the final model as co-
variables, to remove the effect of a staggered survey
from June to October.
Forward selection using CCA indicates the rankingof environmental variables in their importance for

0.0 3.0
0.0
2.5
Acerpseu
Agrostol
Agrocapi
Ajugrept
Athyfili
Bracsylv
Caresylv
Circlute Cirspalu
Cratmono
Dactglom
Desccesp
Dryodila
Dryofili
Hyacnon-
Epilmont
Fragvesc
Fraxexce
Galiapar
GerarobeGeumurba
Hedeheli
Holclana
HolcmollIlexaqui
Junceffu
Loniperi
Luzupilo
LysinemoMercpere
Oxalacet
Pteraqui
Ranurepe
Rubufrut
Sorbaucu
Stacsylv
Urtidioi
Veromont
Poa 1.2
Quer 1.2
Viol 3.4
Dicrhete
Eurhprae
Eurhstri
Lophbide
Mniuhorn
Plagaspl
Plagdent
Pmniundu
Polysp.
Thuitama
Acerps++
Betusp++
Cratmo++
Fraxex++
Quersp++
Betusp+-
Fraxex+-
Coryav-+
Axis 1
Axi
s 2
Fig. 2. DCA species scatter plot. Ordination of those 59 species with a high level of occurrence in the dataset (species present in 65 or more of the 102
sites). Species found in the ground flora component (including bryophytes) listed by first four letters of genus and species name. Woody species listed
by first four letters of genus, first two letters of species and suffixed appropriately. Suffixes ++ and +) refer to tree and sapling growth stages. The
suffix )+ refers to shrub species. The species are: Acerps++, Acer pseudoplatanus, Acerpseu, A. pseudoplatanus, Agrocapi, Agrostis capillaris,
Agrostol, A. stolonifera, Ajugrept, Ajuga reptans, Athyfili, Athyrium filix-femina, Betusp+), Betula sp., Betusp++, Betula sp., Bracsylv, Brachypo-
dium sylvaticum, Caresylv, Carex sylvatica, Circlute, Circaea lutetiana, Cirspalu, Cirsium palustre, Coryav)+, Corylus avellana, Cratmo++, Cra-
taegus monogyna, Cratmono, C. monogyna, Dactglom, Dactylis glomerata, Desccesp, Deschampsia cespitosa, Dicrhete, Dicranella heteromalla,
Dryodila, Dryopteris dilatata, Dryofili, D. filix-mas, Epilmont, Epilobium montanum, Eurhprae, Eurhynchium praelongum, Eurhstri, E. striatum,
Fragvesc, Fragaria vesca, Fraxex+), Fraxinus excelsior, Fraxex++, F. excelsior, Fraxexce, F. excelsior, Galiapar, Galium aparine, Gerarobe, Ge-
ranium robertianum, Geumurba, Geum urbanum, Hedeheli, Hedera helix, Holclana, Holcus lanatus, Holcmoll, H. mollis, Hyacnon-, Hyacinthoides
non-scripta, Ilexaqui, Ilex aquifolium, Junceffu, Juncus effusus,Loniperi, Lonicera periclymenum, Lophbide, Lophocolea bidentata, Luzupilo, Luzula
pilosa, Lysinemo, Lysimachia nemorum, Mercpere,Mercurialis perennis, Mniuhorn,Mnium hornum, Oxalacet, Oxalis acetosella, Plagaspl, Plagiochila
asplenioides, Plagdent, P. denticulatum, Pmniundu, Plagiomnium undulatum, Polysp., Polytrichum sp., Pteraqui, Pteridium aquilinum, Quersp++,
Quercus sp., Ranurepe, Ranunculus repens, Rubufrut, Rubus fruticosus, Sorbaucu, Sorbus aucuparia, Stacsylv, Stachys sylvatica, Thuitama, Thuidium
tamariscinum, Urtidioi, Urtica dioica, Veromont, Veronica montana. Codes Poa 1.2, Quer 1.2 and Viol 3.4 refer to the species couplets Poa nemoralis
and P.trivialis, Quercus petraea and Q. robur (seedlings) and Viola reichenbachiana and V. riviniana, respectively.
498 P.M. Corney et al. / Biological Conservation 120 (2004) 491–505
predicting the species data (Ter Braak and �Smilauer,
2002). Although analysis performed on NWS 1971 dataincluded variables that were at least partly the result of
woodland processes, such as dead wood material, litter
and depth of AO horizon, these variables were included
because they in turn also affect woodland vegetation
composition.
The final constrained model (CCA) examined spe-
cies response to the 20 significant environmental vari-
ables defined by forward selection. Eigenvalues foraxes 1 and 2 were 0.149 and 0.061, respectively, and
the model was significant according to the Monte
Carlo test (f ¼ 2:115, p6 0:001, 9999 permutations).
Site and species ordinations are given in Fig. 3. The
positions of those environmental variables used in the
final model are illustrated using a biplot (Fig. 3(a)),along with the response to these of all species in the
analysis. The biplot is redrawn (Fig. 3(b)) to show
the positions of sites, classified into CVS classes (Barr
et al., 1993; Bunce et al., 1999) using MAVIS (Smart,
2000). Fig. 3(c) gives response to the 20 environmental
variables of 59 most common species, defined as those
present in 65 or more survey sites. The difference be-
tween the species ordinations (Figs. 3(a) and (c)) is afeature of scaling and dependent on the focus of study.
In Fig. 3(c), for example, the vectors have been ex-
tended so that the biplot shows only the most common
species.
-1.0 1.0
-1.0
1.0
Sheep
Intact walls
Soil pH
Depth,
Depth,
AreaEasting
Sea
Altitude
Slope
Litter
Dead
Bare ground
Rendzinas
Pararendzinas
Brown
Brown
Winter
Summer
Accumulated
Axis 1
Axi
s 2
temp
distance
wood
calcareous
bottom A1H
rain days
bottom A0 index
cloudcover
earths
sands
(a)
Fig. 3. CANODRAW CCA biplots. (a) Full species complement and environmental variables.
P.M. Corney et al. / Biological Conservation 120 (2004) 491–505 499
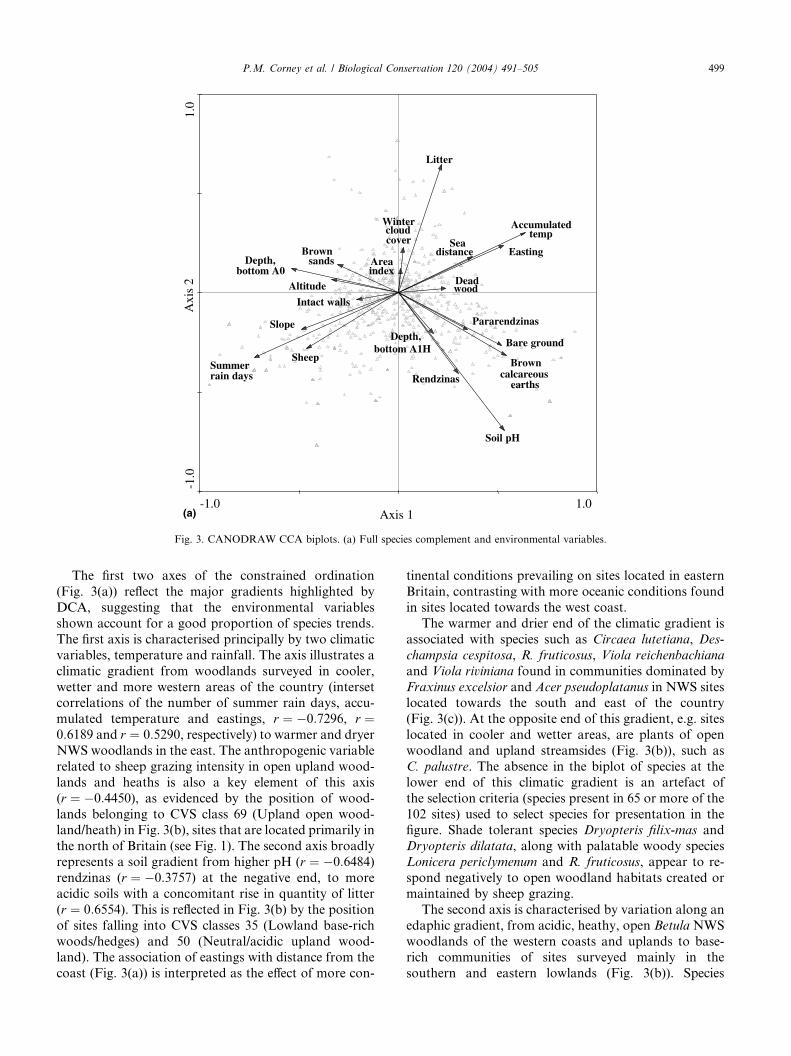
The first two axes of the constrained ordination
(Fig. 3(a)) reflect the major gradients highlighted by
DCA, suggesting that the environmental variables
shown account for a good proportion of species trends.
The first axis is characterised principally by two climatic
variables, temperature and rainfall. The axis illustrates a
climatic gradient from woodlands surveyed in cooler,
wetter and more western areas of the country (intersetcorrelations of the number of summer rain days, accu-
mulated temperature and eastings, r ¼ �0:7296, r ¼0:6189 and r ¼ 0:5290, respectively) to warmer and dryer
NWS woodlands in the east. The anthropogenic variable
related to sheep grazing intensity in open upland wood-
lands and heaths is also a key element of this axis
(r ¼ �0:4450), as evidenced by the position of wood-
lands belonging to CVS class 69 (Upland open wood-land/heath) in Fig. 3(b), sites that are located primarily in
the north of Britain (see Fig. 1). The second axis broadly
represents a soil gradient from higher pH (r ¼ �0:6484)rendzinas (r ¼ �0:3757) at the negative end, to more
acidic soils with a concomitant rise in quantity of litter
(r ¼ 0:6554). This is reflected in Fig. 3(b) by the position
of sites falling into CVS classes 35 (Lowland base-rich
woods/hedges) and 50 (Neutral/acidic upland wood-land). The association of eastings with distance from the
coast (Fig. 3(a)) is interpreted as the effect of more con-
tinental conditions prevailing on sites located in eastern
Britain, contrasting with more oceanic conditions found
in sites located towards the west coast.
The warmer and drier end of the climatic gradient is
associated with species such as Circaea lutetiana, Des-
champsia cespitosa, R. fruticosus, Viola reichenbachiana
and Viola riviniana found in communities dominated by
Fraxinus excelsior and Acer pseudoplatanus in NWS siteslocated towards the south and east of the country
(Fig. 3(c)). At the opposite end of this gradient, e.g. sites
located in cooler and wetter areas, are plants of open
woodland and upland streamsides (Fig. 3(b)), such as
C. palustre. The absence in the biplot of species at the
lower end of this climatic gradient is an artefact of
the selection criteria (species present in 65 or more of the
102 sites) used to select species for presentation in thefigure. Shade tolerant species Dryopteris filix-mas and
Dryopteris dilatata, along with palatable woody species
Lonicera periclymenum and R. fruticosus, appear to re-
spond negatively to open woodland habitats created or
maintained by sheep grazing.
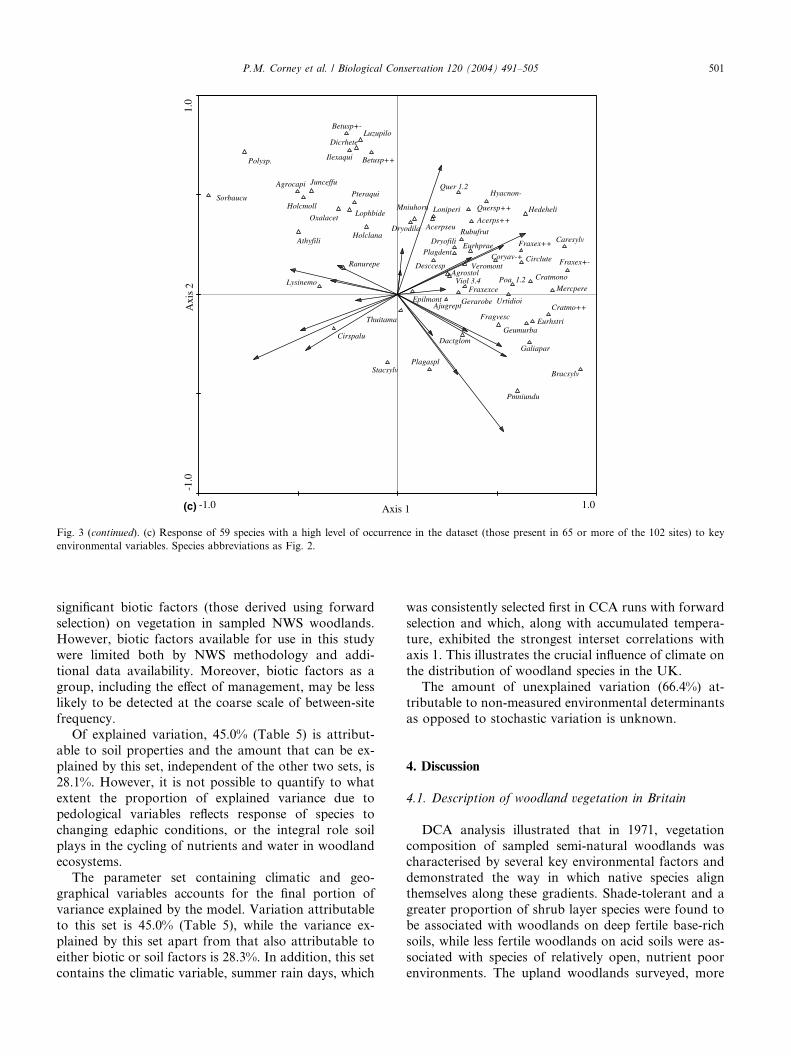
The second axis is characterised by variation along an
edaphic gradient, from acidic, heathy, open Betula NWS
woodlands of the western coasts and uplands to base-rich communities of sites surveyed mainly in the
southern and eastern lowlands (Fig. 3(b)). Species

-1.0 1.0
-1.0
1.0
Axis 1
Axi
s 2
(b)
Fig. 3 (continued). (b) Environmental variables and sites, given in CVS classes. CVS class symbols: 61 Species-rich acid grassland/moorland, ; 25
Shaded grassland/hedges, ; 63 Herb-rich streamsides/acid grassland, ; 46 Diverse upland wooded streamsides, N; 48 Marsh/streamsides, ; 68
Upland oak/birch woodland,j; 62 Upland woodland on podzolic soils, ; 69 Upland open woodland/heath, ; 50 Neutral/acidic upland woodland,
�; 24 Dry base-rich lowland woodland, s; 42 Lowland woodland on heavy soils, d; 35 Lowland base-rich woodland/hedges, .
500 P.M. Corney et al. / Biological Conservation 120 (2004) 491–505
typical of more shallow, acid or lighter soils at the
negative end of this gradient (Fig. 3(c)) include Poly-
trichum sp., often found on wet heaths, moorland and
streamsides in woodland, along with Luzula pilosa,
common among heather, leaf-litter, or moss-dominated
sites, and under Sorbus aucuparia. These communities
are also often found at higher altitudes and exhibit a
more open character associated with higher field layerspecies diversity. In contrast, at the positive end of this
gradient are Geum urbanum, M. perennis, Ajuga reptans
and Plagiomnium undulatum, species found in woodland
on deep, fertile, neutral to base-rich soils which are more
common in the lowlands where dense stands and pro-
nounced shrub layers are associated with greater
amounts of bare ground. The Quercus species, Pteridium
aquilinum and D. dilatata, all species that contributesignificantly to woodland litter production, are found to
be strongly associated with this variable (Fig. 3(c)).
3.4. Assessing the relative contribution of environmental
factors to species composition
The 20 significant environmental variables were di-
vided into three sets (biotic factors, geo-climatic vari-
ables and soil properties), each with similar numbers of
variables (Table 4).
Total inertia was 1.748, estimated total variation
explained (Borcard et al., 1992) was 0.587 and total
variation in the data explained by all three variable
sets was 33.6%. At this level, the variables selected can
be considered to be strongly associated with the
structure of the woodland communities sampled, withregard to between-site species coincidence. The results
of variation partitioning indicating the relative im-
portance of sets of variables in determining broad-
scale woodland vegetation patterns are illustrated
(Fig. 4). There was very little overlap in the variation
accounted for by each of the sets in this model, sug-
gesting only modest interaction between the variable
groups used.Of the total variation in the dataset explained,
36.6% (Table 5) can be attributed to biotic factors on
their own, of which anthropogenic influences are a
part. When soil and geo-climatic sets are included,
20.6% is attributable to biotic variables, the remainder
being as likely to be explained by the other variable
sets, because of their inter-correlation. Variation par-
titioning therefore suggests a moderate impact of
-1.0 1.0
-1.0
1.0
Acerpseu
Agrostol
Agrocapi
Ajugrept
Athyfili
Bracsylv
Caresylv
Circlute
Cirspalu
Cratmono
Dactglom
Desccesp
Dryodila
Dryofili
Hyacnon-
Epilmont
Fragvesc
Fraxexce
Galiapar
Gerarobe
Geumurba
Hedeheli
Holclana
Holcmoll
Ilexaqui
Junceffu
Loniperi
Luzupilo
LysinemoMercpere
Oxalacet
Pteraqui
Ranurepe
Rubufrut
Sorbaucu
Stacsylv
Urtidioi
Veromont
Poa 1.2
Quer 1.2
Viol 3.4
Dicrhete
Eurhprae
Eurhstri
LophbideMniuhorn
Plagaspl
Plagdent
Pmniundu
Polysp.
Thuitama
Acerps++
Betusp++
Cratmo++
Fraxex++
Quersp++
Betusp+-
Fraxex+-Coryav-+
Axis 1
Axi
s 2
(c)
Fig. 3 (continued). (c) Response of 59 species with a high level of occurrence in the dataset (those present in 65 or more of the 102 sites) to key
environmental variables. Species abbreviations as Fig. 2.
P.M. Corney et al. / Biological Conservation 120 (2004) 491–505 501
significant biotic factors (those derived using forward
selection) on vegetation in sampled NWS woodlands.
However, biotic factors available for use in this study
were limited both by NWS methodology and addi-
tional data availability. Moreover, biotic factors as a
group, including the effect of management, may be less
likely to be detected at the coarse scale of between-site
frequency.Of explained variation, 45.0% (Table 5) is attribut-
able to soil properties and the amount that can be ex-
plained by this set, independent of the other two sets, is
28.1%. However, it is not possible to quantify to what
extent the proportion of explained variance due to
pedological variables reflects response of species to
changing edaphic conditions, or the integral role soil
plays in the cycling of nutrients and water in woodlandecosystems.
The parameter set containing climatic and geo-
graphical variables accounts for the final portion of
variance explained by the model. Variation attributable
to this set is 45.0% (Table 5), while the variance ex-
plained by this set apart from that also attributable to
either biotic or soil factors is 28.3%. In addition, this set
contains the climatic variable, summer rain days, which
was consistently selected first in CCA runs with forward
selection and which, along with accumulated tempera-
ture, exhibited the strongest interset correlations with
axis 1. This illustrates the crucial influence of climate on
the distribution of woodland species in the UK.
The amount of unexplained variation (66.4%) at-
tributable to non-measured environmental determinants
as opposed to stochastic variation is unknown.
4. Discussion
4.1. Description of woodland vegetation in Britain
DCA analysis illustrated that in 1971, vegetation
composition of sampled semi-natural woodlands wascharacterised by several key environmental factors and
demonstrated the way in which native species align
themselves along these gradients. Shade-tolerant and a
greater proportion of shrub layer species were found to
be associated with woodlands on deep fertile base-rich
soils, while less fertile woodlands on acid soils were as-
sociated with species of relatively open, nutrient poor
environments. The upland woodlands surveyed, more

Table 4
Sets of explanatory variables used in variation partitioning of wood-
land survey data; set allocations are shown along with the f -values ofan unrestricted Monte Carlo test with 499 permutations conducted at
the final automatic forward selection stage
Subset Variable name Order selected f
G Summer rain days 1 6.34**
G Accumulated temperature 3 2.35**
G Easting 6 1.82**
G Distance to sea 8 1.61**
G Winter cloud cover 10 1.53**
G Slope 12 1.43**
G Altitude 18 1.41**
S Soil pH 2 4.42**
S Depth, bottom of A0 horizon 4 1.88**
S Rendzinas 5 1.86**
S Depth, bottom of A1 horizon 7 1.75**
S Brown sands 11 1.49**
S Pararendzinas 14 1.45**
S Brown calcareous earths 20 1.3*
B Dead wood 9 1.58**
B Litter 13 1.49**
B Area index 15 1.41**
B Sheep grazing 16 1.35**
B Bare ground 17 1.43**
B Intact walls 19 1.3*
Significance of the f -test: *p < 0:05; **p < 0:01. Subsets: B, biotic
factors; G, geo-climatic variables; S, soil properties.
Biotic
20.6%
Soil
28.1%
Geo-climatic
28.3%
6.3%6.3%
3.2%
7.2%
Fig. 4. Venn diagram showing the proportion of the total variance
explained by the CCA model in woodland plant community compo-
sition in Britain, attributable to the sets of environmental variables.
502 P.M. Corney et al. / Biological Conservation 120 (2004) 491–505
likely to be grazed by sheep and coincident with geo-
morphological factors such as rocky or shallow soils,were found to support fewer deep-rooted herbs and
shrub species and were not found to express response to
the grazing/woodland edge gradient more apparent in
lowland woodlands.
4.2. Relative contribution of environmental factors to
species composition
At the between-site level, the vegetation composition
of sampled semi-natural woodlands appears to be
structured mainly by climatic, geographical and soil
variables and to a slightly lesser extent, by biotic fac-
tors. This reflects the qualitative interpretation of the
main divisions within the National Vegetation Classi-
fication (Rodwell et al., 1991); for example betweennorth-west and south-east oak woods and ash woods
and within the south-east types, the split between base-
rich, mesotrophic and acidic communities. There was a
relatively small amount of overlap in variation ac-
counted for by each of the sets in this model, indicating
that, in sampled woodlands, interactions between
groups of geo-climatic, soil and biotic variables
are weak in relation to the effects of the variablesthemselves.
4.3. Effects of biotic factors
Of biotic variables found to be significant using
forward selection, only two were primarily causal fac-
tors (sheep grazing intensity and area index) and these
were of principally anthropogenic influence. Highgrazing pressure was found to be associated with spe-
cies typical of western uplands; grassland, moorland
mosaics and open woodland, whilst low pressures in
the south and east allowed palatable species such as
L. periclymenum and R. fruticosus to thrive. Similar
results have been found in the Atlantic deciduous
woodlands of the western Pyrenees where sheep actively
select woodlands containing an abundant herbaceousunderstorey and preferentially browse Rubus, Ilex,
Vaccinium, forbs and Festuca-like graminoids, while
preferentially rejecting Brachypodium and Luzula sp.
(Garin et al., 2000). Conversely, no significant botanical
response to presence of deer or other herbivorous
mammal species recorded during NWS 1971 was found
in this study.
Management of the wider countryside, defined asLCM 2000 broad habitats, did not correlate with species
distribution, although this may be expected with the
non-contemporaneous nature of these data.
4.4. Effects of soil properties
This analysis also indicates the extent to which soil
type determines vegetation composition of sampledwoodlands. As expected, a strong environmental gradi-
ent related to soil pH was found, from upland
communities on acid soils with low biotic activity, to

Table 5
Variation partitioned between sets of environmental parameters, showing the proportion of explained variance in woodland community composition
at the between-site level explained by each set and grouping of parameters
Variable set NotationP
canonical kn Variation accounted for by variable
set from total variation explained (%)
All variable sets {B[S[G} 0.587 100.0
Biotic set {B} 0.215 36.6
Soil set {S} 0.264 45.0
Geo-climatic set {G} 0.264 45.0
Biotic without effect of soil and geo-climatic {B\S0\G0} 0.121 20.6
Soil without effect of biotic and geo-climatic {S\B0\G0} 0.165 28.1
Geo-climatic without effect of biotic and soil {G\B0\S0} 0.166 28.3
Interaction of biotic and geo-climatic {B\S0} 0.037 6.3
Interaction of soil and biotic {S\G0} 0.037 6.3
Interaction of geo-climatic and soil {G\B0} 0.042 7.2
Interaction of all variable sets {B\S\G} 0.019 3.2
P.M. Corney et al. / Biological Conservation 120 (2004) 491–505 503
species-rich lowland forests on more base-rich soils
featuring a more rapid decomposition of plant material.
Increasing N availability, and consequent soil acidifi-
cation has been shown to affect woodland ground veg-
etation composition in continental oak forests even at
relatively low N loads (e.g. Brunet et al., 1998; Tybirk
and Strandberg, 1999). Nitrogen enrichment can lead to
competitive exclusion of woodland species by more ni-trophilous species, while acidification favours calcifuge
species (Bobbink et al., 1998; Brunet et al., 1998; Hof-
meister et al., 2002; Ling, 2003). However, although no
significant species response to N deposition was found in
this analysis, the finding is limited by the time lapse
between NWS 1971 and the compilation of national
deposition maps.
4.5. Effects of climatic and geographical factors
Summer rainfall and growing degree-days (accumu-
lated temperature) were found to be the most important
factors driving vegetation composition in woodlands
sampled. This effect is well documented; temperature is
one of the primary factors controlling distribution of
Tilia cordata (Pigott and Huntley, 1978), whilst the east–west oceanic-continental rainfall gradient determines
native fern and bryophyte distribution, which is highest
in western oak woods but declines towards the south-
east. However, countrywide influence of both tempera-
ture and rainfall gradients on woodlands in Britain may
be changing as a result of global warming. Over the last
century, average global surface temperatures have risen
by ca. 0.6� (Houghton et al., 2001), while precipitationand cloud cover have both increased by between 0.5%
and 2% over land in the Northern Hemisphere (Nicholls
et al., 1996; Folland and Karl, 2001; Houghton et al.,
2001). Moreover, it has been predicted that these trends
will continue over the next 50–100 years (Cubasch and
Meehl, 2001). If climate change does alter plant distri-
bution considerably and species do not achieve the speed
of migration required to colonise new areas, wide
ranging review of conservation policy may become
necessary.
4.6. Limitations of this analysis
Analysis of landscape-scale environmental factors
affecting vegetation of woodlands surveyed across Brit-ain must necessarily be an analysis at the site level,
where much plot level variation driven by micro-climatic
factors is lost. Nevertheless, analysing vegetation at the
between-site scale has enabled that part of the variation
pertaining to the geographical range of species sampled
in woodlands to be specifically targeted and will both
balance and inform subsequent analysis of plot level
variation.
4.7. Relevance for future woodland management
This study has provided a baseline assessment of a
stratified sample of native semi-natural woodlands in
Britain with particular reference to environmental fac-
tors that control species distribution at the between-site
level. Original NWS sites have recently been resurveyed(2002/3) and the analysis presented in this paper will
provide an opportunity for change to be gauged in a
range of ways. Resurvey will facilitate an assessment of
the way in which plant communities have developed
over time, along with an examination of whether envi-
ronmental variables that were selected as important in
1971 have remained so in 2003. It will also allow in-
vestigation of whether variables such as land use change,N deposition and deer grazing are becoming more im-
portant in the context of semi-natural woodland con-
servation. Irrespective of future benefits, this initial
analysis provides a useful conservation tool, and a
methodology for describing and assessing the important
factors that are likely to affect native semi-natural
woodlands in Britain.

504 P.M. Corney et al. / Biological Conservation 120 (2004) 491–505
Acknowledgements
The authors thank Jane Hall (CEH Monks Wood),
Dr. David Howard (CEH Merlewood), Duncan Ray
(Forest Research), Mary Thorp and Dr. Hugh McAll-ister (University of Liverpool) for technical support.
Financial support for Philip Corney was provided by
EN and CEH in the form of a CASE research stu-
dentship. We are grateful to all who read previous drafts
and supplied useful comments, in particular the valuable
reviews of two anonymous referees which were greatly
appreciated in the final preparation of this manuscript.
References
Avery, B.W., 1980. Soil classification for England and Wales [Higher
categories]. Soil Survey Technical Monograph No.14, Rothamsted
Experimental Station, Harpenden.
Bakkenes, M., Alkemade, J.R.M., Ihle, F., Leemans, R., Latour, J.B.,
2002. Assessing effects of forecasted climate change on the diversity
and distribution of European higher plants for 2050. Global
Change Biology 8, 390–407.
Barr, C.J., Bunce, R.G.H., Clarke, R.T., Fuller, R.M., Furse, M.T.,
Gillespie, M.K., Groom, G.B., Hallam, C.J., Hornung, M.,
Howard, D.C., Ness, M., 1993. Countryside Survey 1990: main
report. Department of the Environment, London.
Bobbink, R., Hornung, M., Roelofs, J.G.M., 1998. The effects of air-
borne nitrogen pollutants on species diversity in natural and semi-
natural European vegetation. Journal of Ecology 86, 717–738.
Borcard, D., Legendre, P., Drapeau, P., 1992. Partialling out the
spatial component of ecological variation. Ecology 73, 1045–1055.
Brunet, J., Diekmann, M., Falkengren-Grerup, U., 1998. Effects of
nitrogen deposition on field layer vegetation in south Swedish oak
forests. Environmental Pollution 102, 35–40.
Brunet, J., FalkengrenGrerup, U., Tyler, G., 1996. Herb layer
vegetation of south Swedish beech and oak forests – effects of
management and soil acidity during one decade. Forest Ecology
and Management 88, 259–272.
Bunce, R.G.H., 1981. British woodlands in an European context. In:
Last, F.T., Gardiner, A.S. (Eds.), Institute of Terrestrial Ecology
Symposium No. 8. Nature of Tree and Woodland Resources.
Natural Environment Research Council.
Bunce, R.G.H., 1982. A Field Key for Classifying British Woodland
Vegetation. Part 1. Institute of Terrestrial Ecology, Cambridge.
Bunce, R.G.H., 1989. A Field Key for Classifying British Woodland
Vegetation. Part 2. Institute of Terrestrial Ecology, Cambridge.
Bunce, R.G.H., Barr, C.J., Gillespie, M.K., Howard, D.C., Scott,
W.A., Smart, S.M., van de Poll, H.M., Watkins, J.W., 1999.
Vegetation of the British countryside – the Countryside Vegetation
System. ECOFACT Volume 1. Department of Environment,
Transport and the Regions, London.
Bunce, R.G.H., Shaw, M.W., 1972. Classifying woodland for conser-
vation. Forestry and Home Grown Timber 1, 23–25.
Bunce, R.G.H., Shaw, M.W., 1973. A standardized procedure for
ecological survey. Journal of Environmental Management 1, 239–
258.
Bunce, R.G.H., Shaw, M.W., 1975. Classification of semi-natural
woodlands in Britain. Institute of Terrestrial Ecology Annual
Report 1974. Natural Environmental Research Council, p. 43.
Cannell, M., 2002. Impacts of climate change on forest growth. In:
Broadmeadow, M. (Ed.), Climate Change: Impacts on UK forests.
Forestry Commission, Edinburgh, pp. 141–149.
Cubasch, U., Meehl, G.A., 2001. Projections of future climate change.
In: Houghton, J.T., Ding, Y., Griggs, D.J., Noguer, M., van der
Linden, P.J., Dai, X., Maskell, K., Johnson, C.A. (Eds.), Climate
Change 2001; The Scientific Basis. Cambridge University Press,
Cambridge, pp. 525–582.
EDINA, 2002. Digimap online mapping service (http://edina.ac.uk/
digimap). � Crown Copyright. An EDINA Digimap/JISC supplied
service.
Elmquist, M., Davies, J., 1999. Getgridvalue ArcView Avenue Script.
ERDAS, 2001. ERDAS IMAGINE, version 8.5. ERDAS, Inc.,
Atlanta, GA.
ESRI, 2000. ArcView GIS, version 3.2a. Environmental Systems
Research Institute, Redlands, CA.
ESRI, 2001. Map Manager version 6.2. ESRI, Redlands, CA.
Folland, C.K., Karl, T.R., 2001. Observed climate variability and
change. In: Houghton, J.T., Ding, Y., Griggs, D.J., Noguer, M.,
van der Linden, P.J., Dai, X., Maskell, K., Johnson, C.A. (Eds.),
Climate Change 2001; The Scientific Basis. Cambridge University
Press, Cambridge, pp. 99–181.
Fuller, R.M., Smith, G.M., Sanderson, R.A., Hill, R.A., Thomson,
A.G., 2002. The UK Land Cover Map 2000: Construction of a
parcel-based vector map from satellite images. Cartographic
Journal 39, 15–25.
Furness, S.B., Grime, J.P., 1982. Growth rate and temperature
responses in bryophytes. II. A comparative study of species of
contrasted ecology. Journal of Ecology 70, 525–536.
Garin, I., Aldezabal, A., Herrero, J., Garcia-Serrano, A., 2000.
Understorey foraging and habitat selection by sheep in mixed
Atlantic woodland. Journal of Vegetation Science 11, 863–870.
Hill, M.O., Bunce, R.G.H., Shaw, M.W., 1975. Indicator species
analysis, a divisive polythetic method of classification and its
application to a survey of native pinewoods in Scotland. Journal of
Ecology 63, 597–613.
Hinsley, S.A., Bellamy, P.E., Newton, I., 1995. Bird species turnover
and stochastic extinction in woodland fragments. Ecography 18,
41–50.
Hofmeister, J., Mihaljevic, M., Hosek, J., Sadlo, J., 2002. Eutrophi-
cation of deciduous forests in the Bohemian Karst (Czech
Republic): the role of nitrogen and phosphorus. Forest Ecology
and Management 169, 213–230.
Honnay, O., Verheyen, K., Butaye, J., Jacquemyn, H., Bossuyt, B.,
Hermy, M., 2002. Possible effects of habitat fragmentation and
climate change on the range of forest plant species. Ecology Letters
5, 525–530.
Houghton, J.T., Ding, Y., Griggs, D.J., Noguer, M., van der Linden,
P.J., Dai, X., Maskell, K., Johnson, C.A., 2001. Climate Change
2001; The Scientific Basis. Contribution of Working Group I to the
Third assessment Report of the Intergovernmental Panel on
Climate Change. Cambridge University Press, Cambridge.
Jenny, H., 1980. The Soil Resource. Springer, Berlin.
Kirby, K.J., 2001. The impact of deer on the ground flora of British
broadleaved woodland. Forestry 74, 219–229.
Lasch, P., Lindner, M., Erhard, M., Suckow, F., Wenzel, A., 2002.
Regional impact assessment on forest structure and functions
under climate change – the Brandenburg case study. Forest
Ecology and Management 162, 73–86.
Le Duc, M.G., Pakeman, R.J., Marrs, R.H., 2000. Vegetation
development on upland and marginal land treated with herbicide,
for bracken (Pteridium aquilinum) control, in Great Britain. Journal
of Environmental Management 58, 147–160.
Legendre, P., Legendre, L., 1998. Numerical Ecology. Elsevier,
Amsterdam.
Ling, K.A., 2003. Using environmental and growth characteristics of
plants to detect long-term changes in response to atmospheric
pollution: some examples from British beech woods. Science of the
Total Environment 310, 203–210.

P.M. Corney et al. / Biological Conservation 120 (2004) 491–505 505
Marrs, R.H., Le Duc, M.G., 2000. Factors controlling vegetation
change in long-term experiments designed to restore heathland in
Breckland, UK. Applied Vegetation Science 3, 135–146.
Meteorological Office, 2003. UK 5 km� 5 km grid baseline datasets
for the period 1961 to 2000. Available from: (http://www.metof-
fice.com/index.html).
Mitchell, J.F.B., Karoly, D.J., 2001. Detection of climate change and
attribution of causes. In: Houghton, J.T., Ding, Y., Griggs, D.J.,
Noguer, M., van der Linden, P.J., Dai, X., Maskell, K., Johnson,
C.A. (Eds.), Climate Change 2001; The Scientific Basis. Cambridge
University Press, Cambridge, pp. 695–738.
Modrzynski, J., Eriksson, G., 2002. Response of Picea abies popula-
tions from elevational transects in the Polish Sudety and Carpa-
thian mountains to simulated drought stress. Forest Ecology and
Management 165, 105–116.
NEGTAP (National Expert Group on Transboundary Air Pollution),
2001. Transboundary Air Pollution: Acidification, Eutrophication
and Ground-Level Ozone in the UK. Report of the National
Expert Group on Transboundary Air Pollution (NEGTAP) for the
UK Department for Environment, Food and Rural Affairs,
Scottish Executive, The National Assembly for Wales/Cynulliad
Cenedlaethol Cymru and the Department of the Environment for
Northern Ireland. CEH, Edinburgh.
Nicholls, N., Gruza, G.V., Jouzel, J., Karl, T.R., Ogallo, L.A.,
Parker, D.E., 1996. Observed climate variability and change. In:
Houghton, J.T., Meira Filho, L.G., Callander, B.A., Harris, N.,
Kattenberg, A., Maskell, K. (Eds.), Climate Change 1995; The
Science of Climate Change. Contribution of Working Group I to
the Second Assessment Report of the Intergovernmental Panel on
Climate Change. Cambridge University Press, Cambridge, pp.
133–192.
Peterken, G.F., 1993. Woodland Conservation and Management.
Chapman & Hall, London.
Pigott, C.D., Huntley, J.P., 1978. Factors controlling the distribution
of Tilia cordata at the northern limits of its geographical range.
New Phytologist 81, 429–441.
Rackham, O., 2003. Ancient Woodland, New ed. Castlepoint Press,
Dalbeattie.
Ratcliffe, D.A., 1977. A nature conservation review: the selection of
biological sites of national importance to nature conservation in
Britain. Cambridge University Press, Cambridge.
Ray, D., 2001. Ecological Site Classification, version 1.7 A PC-based
Decision Support System for British Forests. Forestry Commis-
sion, Edinburgh.
Redfern, D., Hendry, S., 2002. Climate change and damage to trees
caused by extremes of temperature. In: Broadmeadow, M. (Ed.),
Climate Change: Impacts on UK forests. Forestry Commission,
Edinburgh, pp. 29–39.
Rodwell, J., Pigott, C.D., Ratcliffe, D.A., Malloch, A.J.C., Birks,
H.J.B., Proctor, M.C.F., Shimwell, D.W., Huntley, J.P., Radford,
E., Wigginton, M.J., Wilkins, P., 1991. British Plant Communities,
vol. 1. Woodlands and Scrub. Cambridge University Press,
Cambridge.
Sheail, J., Bunce, R.G.H., 2003. The development and scientific
principles of an environmental classification for strategic ecological
survey in the united Kingdom. Environmental Conservation 30,
147–159.
Smart, S.M., 2000. Modular Analysis of Vegetation Information
System (MAVIS) Plot Analyser, version 1.00. CEH, Merlewood,
Cumbria.
Smart, S.M., Bunce, R.G.H., Black, H.I.J., Ray, N., Bunce, F., Kirby,
K.J., Watson, R., Singleton, D., 2001. Measuring long term change
in Biodiversity in British woodlands (1971–2000) – a pilot re-survey
of 14 sites from the 1971 Nature Conservancy ‘Bunce’ woodland
survey and two from the 1971 Native Pinewood Survey. Centre for
Ecology and Hydrology, Merlewood, Cumbria.
Sokal, R.R., Rohlf, F.J., 1995. Biometry, third ed. Freeman, New
York.
Ter Braak, C.J.F., 1986. Canonical correspondence analysis: a new
eigenvector technique for multivariate direct gradient analysis.
Ecology 67, 1167–1179.
Ter Braak, C.J.F., Prentice, C.I., 1988. A theory of gradient analysis.
Advances in Ecological Research 18, 271–313.
Ter Braak, C.J.F., �Smilauer, P., 2002. CANOCO Reference Manual
and CANODRAW for Windows User’s Guide version 4.5.
Microcomputer Power, Ithaca, NY.
Tybirk, K., Strandberg, B., 1999. Oak forest development as a result of
historical land-use patterns and present nitrogen deposition. Forest
Ecology and Management 114, 97–106.
Williams, J.W., Post, D.M., Cwynar, L.C., Lotter, A.F., Levesque,
A.J., 2002. Rapid and widespread vegetation responses to past
climate change in the North Atlantic region. Geology 30, 971–974.
Williams, W.T., Lambert, J.M., 1959. Multivariate methods in plant
ecology. Association analysis in plant communities. Journal of
Ecology 47, 83.
Yeo, M.J.M., Blackstock, T.H., 2002. A vegetation analysis of the
pastoral landscapes of upland Wales, UK. Journal of Vegetation
Science 13, 803–816.




















