
Bioactive cell-hydrogel microcapsules for cell-based drug delivery
Upload
independentCategory
view
3download
0
The DF-1 Chicken Fibroblast Cell Line: Transformation Induced by Diverse Oncogenes andCell Death Resulting from Infection by Avian Leukosis Viruses
Martin Himly,*,1 Douglas N. Foster,† Ivan Bottoli,* Jason S. Iacovoni,* and Peter K. Vogt*,2
*The Scripps Research Institute, Department of Molecular and Experimental Medicine, 10550 North Torrey Pines Road, BCC239,La Jolla, California 92037; and †Department of Animal Science, University of Minnesota, St. Paul, Minnesota 55108
Received March 31, 1998; returned to author for revision May 4, 1998; accepted June 12, 1998
DF-1 is a continuous cell line of chicken embryo fibroblasts. The cells are free of endogenous sequences related to aviansarcoma and leukosis viruses and have normal fibroblastic morphology. DF-1 cells support the replication of avianretroviruses; diverse oncogenes induce foci of oncogenic transformation on monolayers of DF-1, and avian leukosis virusesof envelope subgroups B, D, and C induce cell death and form plaques. The new cell line will greatly facilitate studies ononcogenic transformation and cell killing by avian viruses. © 1998 Academic Press
INTRODUCTION
Research on avian retroviruses has played an impor-tant role in the development of the retrovirus field andhas often defined the leading edge of retrovirology. Thefocus assay, a first quantitative in vitro measurement ofoncogenic transformation, the discovery of reverse tran-scriptase, and the identification of the first retroviral on-cogene product were all made with an avian virus (Teminand Rubin, 1958; Baltimore, 1970; Temin and Mizutani,1970; Brugge and Erickson, 1977). Other seminal devel-opments originating in avian virology were the demon-stration that acutely oncogenic retroviruses are com-monly defective for replication, determination of retroviralgenome size and maps, retroviral recombination andphenotypic mixing, endogenous viruses, temperature-sensitive mutants affecting oncogenic transformation,and the cellular origin of the retroviral oncogenes (Hana-fusa et al., 1963; Payne and Chubb, 1968; Weiss andPayne, 1971; Vogt, 1971; Weiss et al., 1973; Beemon et al.,1974; Billeter et al., 1974; Duesberg et al., 1975; Coffin andBilleter, 1976; Stehelin et al., 1976; Wang et al., 1976a,b).However, avian retrovirology has long had a peculiarimpediment: the absence of a continuous cell line that isnontransformed and free of endogenous and exogenousviral genomes. Although there have been early and morerecent reports on chicken embryo cell lines (Carrel, 1912;Kaaden et al., 1982), none has resulted in a widely avail-able tool for virus studies. The only continuous avian cell
lines in use are frankly transformed either by Marek’sdisease herpesvirus (Akiyama et al., 1973; Nazerian andWitter, 1975; Nazerian et al., 1977), Rous sarcoma virus(Dinowitz, 1977; Gionti et al., 1989; Barald et al., 1997), ora chemical carcinogen (Moscovici et al., 1977), and theyare therefore of limited utility. This deficiency has been asignificant handicap for molecular biological studies. In-deed, analysis of important molecular aspects of retro-viral infection depended on the availability of continuouscell lines, and this requirement has generally favored theuse of mammalian viruses (Hughes et al., 1978; Steegand Vogt, 1990; Berkowitz and Goff, 1993; Alin and Goff,1996). Recently, a continuous line of chicken embryofibroblasts, DF-1, has been developed (Foster, 1998). It isderived from an EV-0 embryo and is free of endogenoussequences related to the avian sarcoma and leukosisvirus (ALSV) group. It is not transformed but has en-hanced growth potential compared to secondary embryofibroblasts. The present report describes the susceptibil-ity of DF-1 to transformation by various oncogenes and toinfection by ALSV. Related aspects of DF-1/ALSV inter-action are explored in a companion paper (Schaefer-Klein et al., 1998).
RESULTS
Diverse oncogenes induce transformed cell foci incultures of DF-1 cells
Tables 1 and 2 summarize the results of transforma-tion assays on DF-1 cells. Oncogenes that code forcytoplasmic proteins, with the exceptions of crk andv-abl, induce distinct transformed cell foci in DF-1. Theseoncogenes share transforming properties on CEF andDF-1: they induce swelling and rounding of the cell and
1 Present address: Institute of Genetics, University of Salzburg, Hell-brunnerstr. 34, 5020 Salzburg, Austria.
2 To whom reprint requests should be addressed. E-mail: [email protected].
VIROLOGY 248, 295–304 (1998)ARTICLE NO. VY989290
0042-6822/98 $25.00Copyright © 1998 by Academic PressAll rights of reproduction in any form reserved.
295
pronounced multilayering within the focus. These cellu-lar changes in morphology and growth pattern markthese oncogenes as “strong” transformers. DF-1 cellshave higher saturation densities than CEF (2820 6 154versus 736 6 51 cells/mm2 reached under DC3 agaroverlay on day 4 after seeding). Therefore, focus forma-tion on DF-1 must occur against an increased back-ground of multilayering by uninfected cells. Most nuclearoncogenes do not induce foci on DF-1 cells. The twoexceptions are myc and e2f; both also induce cell deathand aberrant growth.
Figures 1 and 2 illustrate transformed cell morpholo-gies and growth patterns induced by some of the onco-genes in DF-1 cultures. In most instances, the foci inDF-1 resembled those in CEF. However, there were somenotable exceptions. On DF-1 clone P3B, the c-mos onco-gene induced the formation of cell rounding as well asmultinucleated cells that originated by cell fusion orendomitosis (Fig. 2C). These cell types are also found inCEF and in chicken yolk sac macrophage cultures ex-pressing c-mos. In contrast, clone 32 of DF-1 respondedto expression of c-mos by becoming elongated and swol-len-fusiform (Fig. 1D); multinucleated cells were not prev-alent. These two types of mos-induced transformationillustrate the extremes of clonal variation within the DF-1cell population. Foci induced by myc consisted of amixed population of rounded, healthy appearing trans-formed cells and dying cells with fragmented nuclei. Thisobservation is in accord with the transforming and ap-optosis-inducing potential of myc (Dolnikov et al., 1996;Fulton et al., 1996; Petropoulos et al., 1996). Cell-killingactivity predominated in MH2, a virus that contains boththe myc and the mil (avian homolog of raf) oncogenes.Similarly, the foci induced by e2f contained a preponder-
ance of dying cells. The E2F protein can have differenteffects on cells depending on the level of expression(Kowalik et al., 1995; DeGregori et al., 1997; Givol et al.,1998). Surprizingly, NK24, a fos-carrying avian retrovirus,also induced cell death rather than oncogenic transfor-mation.
Construct-dependent transformation by Jun
The jun oncogene was originally derived from aviansarcoma virus 17 (ASV17). In this viral genome, it is fusedat its 59 terminus to viral gag sequences generating aGag-Jun fusion protein. Tests with ASV17 in DF-1 culturesfailed to induce foci of distinctly transformed cells. How-ever, when cells of ASV17-infected DF-1 agar colonieswere plated onto plastic, they showed the fusiform andneedle-like morphology characteristic of Jun-transformedCEF. (Maki et al., 1987). Additional experiments with Junconstructs in the RCAS (Hughes et al., 1987; Federspieland Hughes, 1997) and pRV vectors (Kataoka et al., 1993)also revealed a direct focus-forming ability in DF-1 cul-tures. The active constructs were RCAS-vJ1 and pRV-vJ.Both contain the viral Jun of ASV17 but without the Gagsequences. RCAS-vJ0, which, like ASV17, expresses theGag v-Jun fusion protein, did not induce foci. Theseobservations suggest that fusion to Gag may reducefocus-forming ability of Jun in DF-1 cultures. The reasonfor this reduction remains to be determined; it could berelated to protein synthesis or protein stability.
Comparison of efficiencies of focus formation
Table 3 summarizes the efficiencies of focus formationby selected oncogenes on DF-1 cells relative to second-ary CEF cultures. Several oncogenes, namely, src, erbA/erbB, sea, DSH3-abl, mafQ5H, and e2f, show efficiencies
TABLE 2
Focus Formation Induced by Nuclear Oncogenes on DF-1 Cells
OncogenesFocus
formationRepresentativetiter (FFU/ml)
Viruses andconstructs tested
v-gag-jun No ASV17, RCAS-vJ0v-jun Yes 1 3 106 RCAS-vJ1junD No RCAS-JUN Dv-qin No ASV31v-myb/ets No E26v-myb No AMVpax3fkhr No RCASBP-PAX3-FKHRc-myc Yes 9 3 106 RCAS-c-MYCv-myc Yes 5 3 105 RCAS-v-MYCv-fosa No 4 3 106 NK24v-maf No AS42v-maf Q5H Yes 2 3 104 pRV-MAF Q5He2f a No 8 3 106 RCASBP-E2Fv-mil/myca No 3.6 3 106 MH2
a Predominantly cell death.FFU, focus-forming units.
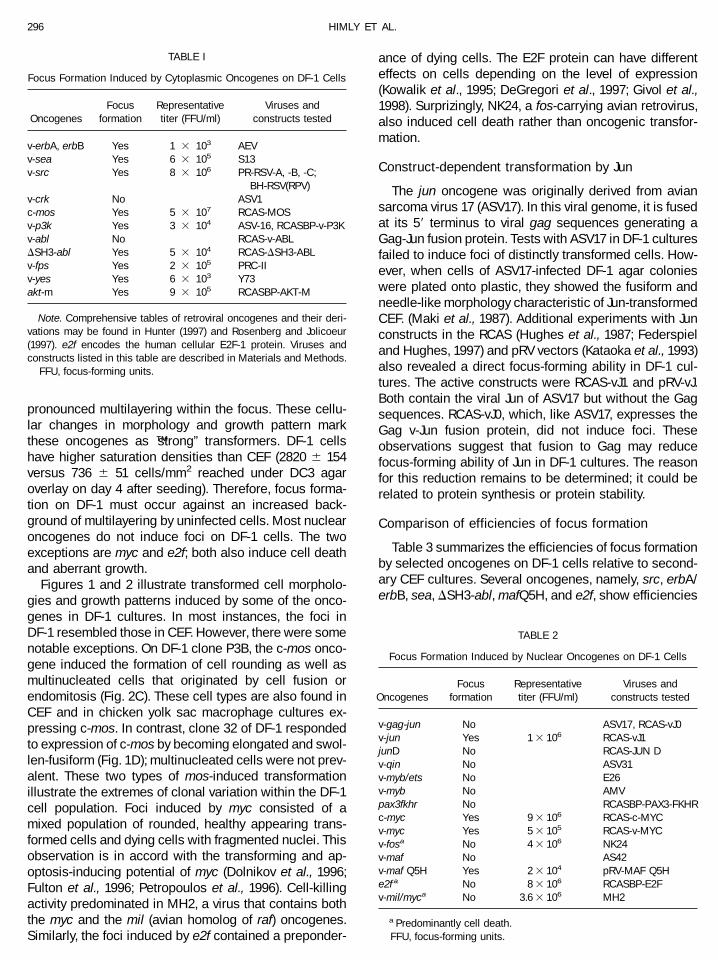
TABLE I
Focus Formation Induced by Cytoplasmic Oncogenes on DF-1 Cells
OncogenesFocus
formationRepresentativetiter (FFU/ml)
Viruses andconstructs tested
v-erbA, erbB Yes 1 3 103 AEVv-sea Yes 6 3 105 S13v-src Yes 8 3 106 PR-RSV-A, -B, -C;
BH-RSV(RPV)v-crk No ASV1c-mos Yes 5 3 107 RCAS-MOSv-p3k Yes 3 3 104 ASV-16, RCASBP-v-P3Kv-abl No RCAS-v-ABLDSH3-abl Yes 5 3 104 RCAS-DSH3-ABLv-fps Yes 2 3 105 PRC-IIv-yes Yes 6 3 103 Y73akt-m Yes 9 3 105 RCASBP-AKT-M
Note. Comprehensive tables of retroviral oncogenes and their deri-vations may be found in Hunter (1997) and Rosenberg and Jolicoeur(1997). e2f encodes the human cellular E2F-1 protein. Viruses andconstructs listed in this table are described in Materials and Methods.
FFU, focus-forming units.
296 HIMLY ET AL.

of focus formation on DF-1 that differ from the efficien-cies on CEF by factors of ,10. These differences arewithin 2 SDs of the mean efficiencies determined withCEF from different embryos. Other oncogenes, however,showed efficiencies of focus formation on DF-1 that de-viated from those on CEF by factors of .10. Increasedefficiencies were seen with c-mos, v-myc, c-myc, p3k,and ras. For mos and v-myc, the differences approach100-fold. In contrast, fps and yes transformed CEF muchbetter than DF-1. The very high difference between CEFand DF-1 seen with the combined mil/myc oncogenesexpressed by MH2 virus may represent an enhancementof apoptosis by the cooperation of the oncogenes or anexcess of a cytopathic avian leukosis helper virus. Sucha helper virus may cause no detectable cell damage in
CEF cultures but induce death of DF-1 (see below).Indeed, the virus titer given for MH2 is based on com-bined plaque and focus counts. The reasons for theincreased or decreased susceptibility of DF-1 to theseoncogenes remain to be determined.
Cell death induced in DF-1 cultures by avianleukosis virus
The cytopathic effect of fos and mil/myc in DF-1 cellssuggested an increased susceptibility of the cell line tocell death-inducing viral products. This possibility wasfurther investigated by studying the effects of avian leu-kosis viruses on DF-1. These viruses came from differentenvelope subgroups, some of which are known to have
FIG. 1. Transformation of DF-1 cells clone 32 by retroviral oncogenes. Live cells observed with phase contrast optics and photographed at 1003magnification. Uninfected DF-1 clone 32 cells on day 5 (A) and day 8 (B) after seeding. (C) Transformation by v-ras. (D) Transformation by c-mos. (E)Transformation by c-myc. (F) Transformation by v-src.
297FOCI AND PLAQUES ON DF-1 CELLS

cytopathic potential for CEF (Graf, 1972; Brojatsch et al.,1996). These experiments are summarized in Table 4.Under the present culture conditions, none of the avianleukosis viruses induced detectable changes in cellgrowth or morphology in CEF. In contrast, DF-1 cellswere severely affected by viruses of subgroups B, D, andC, which induced extensive cell death at high virus con-centrations and individual, readily countable plaques atlow concentrations (Figs. 3 and 4). The number of theseplaques was directly proportional to the virus concentra-tion. DF-1 cells can therefore be used for rapid andquantitative assays of subgroup B, D, and C avian leu-kosis viruses. The dying cells in these cultures contained
fragmented nuclei and were positive in the TUNEL assay,suggesting that they were undergoing apoptosis.
Retroviral replication in DF-1 cells
The ability of DF-1 cells to replicate avian retroviruseswas investigated in a one-step growth experiment. Sec-ondary CEF and DF-1 cells were seeded onto separate60-mm dishes at 1 3 106 cells per dish and inoculated ata multiplicity of 1.0 with the Prague strain of Rous sar-coma virus, envelope subgroup A. The cells were kept ina medium consisting of 93% Ham’s F-10, 5% calf serum,and 2% L-glutamine-penicillin-streptomycin. Harvests of
FIG. 2. Transformation of DF-1 cells clone P3B cells by retroviral oncogenes. Live cells observed with phase contrast optics and photographed at1603 magnification. (A) Uninfected DF-1 clone P3B. (B) Transformation by v-ras. (C) Transformation by c-mos. (D) Transformation by v-src. (E)Transformation by c-myc. (F) Transformation by v-myc.
298 HIMLY ET AL.

the culture fluids were taken at daily intervals and as-sayed for focus formation on secondary CEF. Figure 5shows the results. DF-1 cells support the growth of Roussarcoma virus, producing amounts of virus that by days3 and 4 postinfection are roughly equivalent to virusyields from CEF.
DISCUSSION
The properties of DF-1 will encourage widespread useof this cell line for work with diverse avian viruses,especially for molecular biological studies with avianretroviruses. DF-1 cells are susceptible to transformationby numerous oncogenes and efficiently replicate avianretroviruses. Some of the oncogenes that do not inducewell defined oncogenic foci in DF-1 cultures may, similarto gag-jun of ASV17 and of RCAS-vJ0, induce cellulartransformation that can be recognized on subcloning ofthe oncogene-expressing cells. DF-1 cells show an in-creased saturation density, and because of this back-ground of uninfected cell growth, foci of transformedcells become discernible only if distinct changes in cellshape or excessive multilayering are induced by theoncogene. The requirements for focus formation in DF-1cells appear more stringent than the ones in CEF andmay constitute the primary cause for the failure of someoncogenes to induce foci in DF-1 cultures. However,other reasons for inactivity have not been ruled out andshould be investigated; these include low levels of on-cogene expression or instability of the oncogene productor a specific resistance of DF-1 cells to the growthstimulatory signal issued by a particular oncogene. Highsteady state levels of an oncoprotein may be critical; they
provide a likely explanation for the differences in focus-forming ability observed with various Jun constructs.
The high susceptibility of DF-1 cells to the celldeath-inducing activity of certain avian retrovirusesclearly distinguishes DF-1 from CEF and will greatlyfacilitate investigations of virus-induced apoptosis.Some of the observations on cell killing reported heremust be examined further. Although the apoptosis-inducing activities of envelope subgroup B and Dviruses are known to be mediated by a cell surfacereceptor related to the receptor for tumor necrosisfactor (Brojatsch et al., 1996), the cell-killing activitiesof envelope subgroup C viruses are unexpected andunexplained. Barring a trivial explanation (e.g., con-tamination with subgroup B or D viruses), the cell-killing activity of envelope subgroup C could revealinteresting new aspects of apoptosis. The cytopatho-genicity of the fos-expressing avian retrovirus NK24could also be caused by an envelope component or bythe fos oncogene proper, which can function as amediator of stress-induced apoptosis (Pruschy et al.,1997). Similar considerations apply to the extremecell-killing effect of MH2. It could result from the com-bination of the oncogenes mil (raf) and myc expressedby this virus, from a subgroup B or D envelope com-ponent, or from both. Occasionally, we have seen celldeath, presumably apoptosis, in DF-1 cultures evenwith src. This discussion makes it clear that the ex-quisite susceptibility of DF-1 cells to viral cytopathiceffects raises several interesting new questions thatrelate to molecular mechanisms of apoptosis; it alsosuggests that this cell line will prove useful in thestudy of avian and mammalian cytopathic viruses.
TABLE 4
Plaque Formation Induced by Avian Leukosis Viruses on DF-1 Cells
Avian leukosisvirus
Envelopesubgroup
Representativeplaque titer (PFU/ml)
RAV-1 A no CPERAV-3 no CPERAV-2 B 5 3 105
RAV-6 1 3 106
tdPRB 2 3 105
RAV7 C 1 3 105
tdB77 2 3 104
CZAV D 3 3 104
RPV F no CPEGPV G no CPE
Abbreviations: CPE, cytopathic effect; RAV-1, Rous associated vi-rus-1; tdPRB, transformation-defective Prague strain of Rous sarcomavirus envelope subgroup B; tdB77, transformation-defective avian sar-coma virus B77; CZAV, Carr-Zilber-associated virus; RPV, ring neckpheasant virus; GPV, golden pheasant virus (Duesberg and Vogt, 1970;Duesberg and Vogt, 1973; Duff and Vogt, 1969; Fujita et al., 1974; Graf,1972; Weiss, Mason, and Vogt, 1973).
TABLE 3
Efficiencies of Focus Formation on DF-1 Cells
OncogeneFFU or PFU/ml on DF-1FFU or PFU/ml on CEF a
src 1.0sea 3.0p3k 10.0DSH3-abl 2.5fps 0.08yes 0.06ras 12.0mos 75.0c-mycb 11.1v-mycb 84.0v-jun 2.5e2f c 3.0mil/myc (MH2)c 1440.0erbAerbB 0.2
a Data are from a representative experiment. PFU, plaque-formingunits; FFU, focus-forming units.
b Tranformation and cell death.c Predominatly cell death.
299FOCI AND PLAQUES ON DF-1 CELLS

MATERIALS AND METHODS
Viruses and constructs
Avian erythroblastosis virus (AEV) is of the ES4 strain;it carries the erbA and erbB oncogenes (Lai et al., 1979,1980; Debuire et al., 1984). The Prague strains of Roussarcoma virus envelope subgroups A, B, and C havebeen described (Duff and Vogt, 1969). The Bryan hightiter strain of Rous sarcoma virus with ring-neckedpheasant virus helper, BH-RSV(RPV), has also been char-acterized previously (Fujita et al., 1974). ASV-1 is an aviansarcoma virus isolated from a spontaneous tumor. Itcontains the crk oncogene (Tsuchie et al., 1989). PRCIIstands for Poultry Research Centre II; this virus carriesthe fps oncogene, and Y73 contains the yes insert (Neilet al., 1981; Kitamura et al., 1982). ASV-17 and ASV-31with the oncogenes jun and qin, respectively, have beencharacterized recently (Maki et al., 1987; Li and Vogt,1993). E26 and avian myeloblastosis virus (AMV) strainBAI-A are acute avian leukemia viruses carrying themyb/ets and the myb oncogene, respectively (Langlois etal., 1966; Nunn et al., 1983). The fos- and maf-carryingviruses NK24 and AS42 have been isolated and studiedby Kawai and coworkers (Goto and Kawai, 1987; Kawai etal., 1992; Kataoka et al., 1993). S13 is an avian erythro-blastosis virus that carries the sea oncogene (Hayman etal., 1985; Woods et al., 1986; Smith et al., 1989). RCAS is
an avian retroviral expression vector based on the ge-nome of the Prague strain of Rous sarcoma virus(Hughes et al., 1987; Petropoulos and Hughes, 1991;Federspiel and Hughes, 1997). Cells transfected withRCAS constructs release infectious retroviruses that ex-press the insert. Two versions of this vector were used,RCAS and RCASBP. In the latter, the polymerase genehas been replaced by that of the Bryan high titer strain ofRous sarcoma virus. The two versions are noted in theconstructs listed in Tables 1 and 2. The jun-expressingconstructs RCAS-vJ1 and RCAS-vJ0 have been describedpreviously (Bos et al., 1990) The RCAS constructs withchicken c-mos (RCAS-MOS), chicken c-myc (RCAS-c-MYC), and v-myc (RCAS-v-MYC, derived from avian my-elocytoma virus MC29; Mladenov et al., 1967; Duesbergand Vogt, 1979) were kindly provided by Stephen Hughes.They have been characterized previously (Schmidt et al.,1988; Petropoulos et al., 1996). The v-abl and DSH3 ablclones have been described; they were also expressedby RCAS (Kelliher et al., 1990; Maru et al., 1996). RCAS-junD carries a mutationally activated form of the chickenjunD gene (Hartl and Vogt, 1992). RCAS-v-p3k and ASV-16carry a homolog of the catalytic subunit of phosphoino-sitide-3 kinase as an oncogene (Chang et al., 1997).RCAS-AKT-m contains the myristylable form of the mu-rine akt oncogene (Bellacosa et al., 1991; Aoki, 1998).RCAS-PAX3-FKHR expresses a fusion protein consisting
FIG. 3. Cytopathic effects of the envelope subgroup B virus RAV-2. Live cells observed with phase contrast optics and photographed at 1003magnification. (A) Sparse culture with beginning cytopathic changes that include cell vacuolization and cell surface blobbing. (B and C) Later stagesof the same culture show increased cell death but also persistent cell growth. (D) Normal DF-1 cells.
300 HIMLY ET AL.

of the DNA binding domain of PAX3 and the transcriptionregulatory domain of the FKHR (forkhead related) gene.This fusion protein is derived from a t(2,13) translocation
that occurs in human rhabdomyosarcoma. The RCAS-PAX3-FKHR construct transforms secondary CEF cul-tures (Scheidler et al., 1996). maf Q5H is a mutated mafoncogene that is more potent in transformation than theviral maf of AS42; it is expressed in the pRV retroviralvector (Kataoka et al., 1993). RCAS-E2F expresses thehuman e2f gene.
Cells
Primary chicken embryo fibroblasts were prepared bystandard techniques (Vogt, 1969) from 10-day embryossupplied by SPAFAS, Inc. (Preston, Connecticut). For vi-rus assays, they were passaged once and are referred toas CEF. These cells are susceptible to envelope sub-groups A, B, C, D, and F of ASLVs and contain genomicsequences related to ASLV genomes (EV sequences).DF-1 cells are a continuous line of fibroblasts derivedfrom an EV-0 chicken embryo lacking endogenous ASLV-like sequences (Foster, 1998). The line developed spon-taneously from a high density seeding of fibroblasts froma 10-day embryo. Cells of this seeding were continuouslypassaged and split when confluent. Eleventh-passagecells appeared highly senescent and were seeded onto30 plates. The cells were refed for several weeks, and
FIG. 4. Plaques produced by avian leukosis viruses. The cells are fixed and stained with crystal violet. (A-C) Photographed with bright fieldillumination at 253 magnification. (D and E) Photographed without magnification. (A) RAV-2, (B) tdB77, (C) MH2, (D) RAV-2, and (E) tdB77.
FIG. 5. One-step growth of PR-RSV-A in CEF compared with DF-1cells.
301FOCI AND PLAQUES ON DF-1 CELLS

eventually a small population of cells began dividing onone of the plates. Initially, the cells grew slowly and hadto be transferred at high seeding density, but by the 40thpassage, they grew more rapidly and formed a homoge-neous population (Foster, 1998). DF-1 cells were origi-nally grown in Dulbecco’s modified Eagle’s medium) sup-plemented with 10% fetal calf serum. During recent pas-sages, they were switched to cloning medium consistingof 83% Ham’s F-10, 10% calf serum, 4.5% chicken serum(heated to 56°C for 60 min), 1% of 1003 vitamin solution,1% of 1.8 mM folic acid, and 0.5% dimethylsulfoxide. Inthis medium, the DF-1 cells showed a more uniform,flatter, and fibroblast-like morphology and more rapidgrowth. Several clones of DF-1 cells were establishedand were selected for flat adherent cell shape andgrowth properties that led to a minimum of multilayering.
Focus assays
Focus assays were carried out on CEF as describedpreviously (Vogt, 1969). Six-well microplates (35-mm di-ameter) were used with 5 3 105 cells seeded per well.The absorption and penetration medium consisted of93% Ham’s F-10, 5% calf serum, and 2% of 1003L-glutamine-penicillin-streptomycin. This medium wasreplaced on the day after inoculation with nutrient agaroverlay (DC3) that contained 43% of 23 Ham’s F-10, 3%fetal calf serum, 1% heat-inactivated chicken serum, 8.5%tryptose phosphate, 1% of 1003 L-glutamine-penicillin-streptomycin, 1% dimethylsulfoxide, and 42.5% of a 1.5%solution of SeaPlaque agar. The procedure was modifiedfor DF-1 cells. Cell seeding was reduced to 3 3 105 cellsper well, and cell growth had to be slowed by reducingthe concentration of fetal calf serum in the nutrient agaroverlay (DC2) to 2.3%. With these modifications, focicould be scored on DF-1 up to 2 weeks after infection.
ACKNOWLEDGMENTS
Stephen H. Hughes first directed our attention to DF-1, freelysharing his advice, insights, and RCAS constructs. We gratefullyacknowledge his generosity. We also thank Irene F. Fu and DouglasW. Geerdes for competent technical assistance and Susan K. Burkefor help with the manuscript. Dr. M. Nishizawa first drew our atten-tion to the DF-1-transforming ability of certain jun constructs. Thiswork was supported by U.S. Public Health Service Research GrantCA42564. Martin Himly received fellowship support from the FederalGovernment of Austria (Bundesministerium fur Wissenschaft undVerkehr). Ivan Bottoli is the recipient of a fellowship from the SwissCancer League. We thank Makoto Nishizawa for alerting us to thedifferences between Gag-Jun and Jun in the transformation of DF-1.
Note added in proof. In recent experiments by Nowakowski and Vogtthe construct RCASBP-PAX3-FKHR was found to induce transformedcell foci in cultures of DF-1 cells (cf. Table 2).
REFERENCES
Akiyama, Y., Kato, S., and Iwa, N. (1973). Continuous cell culture fromlymphoma of Marek’s disease. Biken J. 16, 177–179.
Alin, K., and Goff, S. P. (1996). Mutational analysis of interactionsbetween the Gag precursor proteins of murine leukemia viruses.Virology 216, 418–424.
Baltimore, D. (1970). RNA-dependent DNA polymerase in virions of RNAtumor viruses. Nature 226, 1209–1211.
Barald, K. F., Lindberg, K. H., Hardiman, K., Kavka, A. I., Lewis, J. E.,Victor, J. C., Gardner, C. A., and Poniatowski, A. (1997). Immortalizedcell lines from embryonic avian and murine otocysts: Tools for mo-lecular studies of the developing inner ear. Int. J. Dev. Neurosci. 15,523–540.
Beemon, K., Duesberg, P., and Vogt, P. (1974). Evidence for crossing-over between avian tumor viruses based on analysis of viral RNAs.Proc. Natl. Acad. Sci. USA 71, 4254–4258.
Bellacosa, A., Testa, J. R., Staal, S. P., and Tsichlis, P. N. (1991). Aretroviral oncogene, akt, encoding a serine-threonine kinase contain-ing an SH2-like region. Science 254, 274–277.
Berkowitz, R. D., and Goff, S. P. (1993). Point mutations in Moloneymurine leukemia virus envelope protein: Effects on infectivity, virionassociation, and superinfection resistance. Virology 196, 748–757.
Billeter, M. A., Parsons, J. T., and Coffin, J. M. (1974). The nucleotidesequence complexity of avian tumor virus RNA. Proc. Natl. Acad. Sci.USA 71, 3560–3564.
Bos, T. J., Monteclaro, F. S., Mitsunobu, F., Ball, A. R., Jr., Chang, C. H.,Nishimura, T., and Vogt, P. K. (1990). Efficient transformation ofchicken embryo fibroblasts by c-Jun requires structural modificationin coding and noncoding sequences. Genes Dev. 4, 1677–1687.
Brojatsch, J., Naughton, J., Rolls, M. M., Zingler, K., and Young, J. A.(1996). CAR1, a TNFR-related protein, is a cellular receptor for cyto-pathic avian leukosis-sarcoma viruses and mediates apoptosis. Cell87, 845–855.
Brugge, J. S., and Erickson, R. L. (1977). Identification of a transforma-tion-specific antigen induced by an avian sarcoma virus. Nature 269,346–348.
Carrel, A. (1912). The permanent life of tissue outside of the organism.J. Exp. Med. 15, 516.
Chang, H. W., Aoki, M., Fruman, D., Auger, K. R., Bellacosa, A., Tsichlis,P. N., Cantley, L. C., Roberts, T. M., and Vogt, P. K. (1997). Transfor-mation of chicken cells by the gene encoding the catalytic subunit ofPI 3-kinase. Science 276, 1848–1850.
Coffin, J. M., and Billeter, M. A. (1976). A physical map of the Roussarcoma virus genome. J. Mol. Biol. 100, 293–318.
Debuire, B., Henry, C., Bernissa, M.,Biserte, G., Claverie, J. M., Saule, S.,Martin, P., and Stehelin, D. (1984). Sequencing the erbA gene of avianerythroblastosis virus reveals a new type of oncogene. Science 224,1456–1459.
DeGregori, J., Leone, G., Miron, A., Jakoi, L., and Nevins, J. R. (1997).Distinct roles for E2F proteins in cell growth control and apoptosis.Proc. Natl. Acad. Sci. USA 94, 7245–7250.
Dinowitz, M. (1977). A continuous line of Rous sarcoma virus-trans-formed chick embryo cells. J. Natl. Cancer Inst. 58, 307–312.
Dolnikov, A., Ward, R. L., Hawkins, N. J., and Symonds, G. (1996).Apoptosis in v-myc transformation of myelomonocytic cells and itsmodulation by CSF-1. Oncogene 12, 1189–1198.
Duesberg, P., Vogt, P. K., Beemon, K., and Lai, M. (1975). Avian RNAtumor viruses: mechanism of recombination and complexity of thegenome. Cold Spring Harb. Symp. Quant. Biol. 39, 847–857.
Duesberg, P. H., and Vogt, P. K. (1979). Avian acute leukemia virusesMC29 and MH2 share specific RNA sequences: Evidence for asecond class of transforming genes. Proc. Natl. Acad. Sci. USA 76,1633–1637.
Duff, R. G., and Vogt, P. K. (1969). Characteristics of two new aviantumor virus subgroups. Virology 39, 18–30.
Federspiel, M. J., and Hughes, S. H. (1997). Retroviral gene delivery.Methods Cell Biol. 52, 179–214.
Foster, D. N. (1998). Development of a spontaneously immortalizedchicken embryo fibroblastic cell line.
Fujita, D. J., Chen, Y. C., Friis, R. R., and Vogt, P. K. (1974). RNA tumor
302 HIMLY ET AL.

viruses of pheasants: Characterization of avian leukosis subgroups Fand G. Virology 60, 558–571.
Fulton, R., Gallagher, R., Crouch, D., and Neil, J. C. (1996). Apparentuncoupling of oncogenicity from fibroblast transformation and apo-ptosis in a mutant myc gene transduced by feline leukemia virus.J. Virol. 70, 1154–1162.
Gionti, E., Jullien, P., Pontarelli, G., and Sanchez, M. (1989). A continuousline of chicken embryo cells derived from a chondrocyte cultureinfected with RSV. Cell Differ. Dev. 27, 215–223.
Givol, I., Givol, D., and Hughes, S. H. (1998). Overexpression of p21waf1/cip1
arrests the growth of chicken embryo fibroblasts that overexpress E2F1.Oncogene 16, 3115–3122.
Graf, T. (1972). A plaque assay for avian RNA tumor viruses. Virology 50,567–578.
Hanafusa, H., Hanafusa, T., and Rubin, H. (1963). The defectiveness ofRous sarcoma virus. Proc. Natl. Acad. Sci. USA 49, 572–580.
Hartl, M., and Vogt, P. K. (1992). A rearranged junD transforms chickenembryo fibroblasts. Cell Growth Differ. 3, 909–918.
Hayman, M. J., Kitchener, G., Vogt, P. K., and Beug, H. (1985). Theputative transforming protein of S13 avian erythroblastosis virus is atransmembrane glycoprotein with an associated protein kinase ac-tivity. Proc. Natl. Acad. Sci. USA 82, 8237–8241.
Hughes, S. E., Shank, P. R., Spector, D. H., Kung, H.-J., Bishop, J. M.,Varmus, H. E., Vogt, P. K., and Breitman, M. L. (1978). Proviruses ofavian sarcoma virus are terminally redundant: Co-extensive withunintegrated linear DNA and integrated at many sites. Cell 15,1397–1410.
Hughes, S. H., Greenhouse, J. J., Petropoulos, C. J., and Sutrave, P.(1987). Adaptor plasmids simplify the insertion of foreign DNA intohelper-independent retroviral vectors. J. Virol. 61, 3004–3012.
Hunter, T. (1997). Oncogenes, growth suppressor genes, and cancer. In“Exploring Genetic Mechanisms” (M. Singer, and P. Berg, Eds.), pp.203–269. University Science Books, Sausalito.
Kaaden, O. R., Lange, S., and Stiburek, B. (1982). Establishment andcharacterization of chicken embryo fibroblast clone LSCC-H32. InVitro 18, 827–834.
Kataoka, K., Nishizawa, M., and Kawai, S. (1993). Structure-functionanalysis of the maf oncogene product, a member of the b-Zip proteinfamily. J. Virol. 67, 2133–2141.
Kawai, S., Goto, N., Kataoka, K., Saegusa, T., Shinno-Kohno, H., andNishizawa, M. (1992). Isolation of the avian transforming retrovirus,AS42, carrying the v-maf oncogene and initial characterization of itsgene product. Virology 188, 778–784.
Kelliher, M. A., McLaughlin, J., Witte, O. N., and Rosenberg, N. (1990).Induction of a chronic myelogenous leukemia-like syndrome in micewith v-abl and BCR/ABL. Proc. Natl. Acad. Sci. USA 87, 6649–6653.
Kitamura, N., Kitamura, A., Toyoshima, K., Hirayama, Y., and Yoshida, M.(1982). Avian sarcoma virus Y73 genome sequence and structuralsimilarity of its transforming gene product to that of Rous sarcomavirus. Nature 297, 205–208.
Kowalik, T. F., DeGregori, J., Schwarz, J. K., and Nevins, J. R. (1995). E2F1overexpression in quiescent fibroblasts leads to induction of cellularDNA synthesis and apoptosis. J. Virol. 69, 2491–2500.
Lai, M. M., Hu, S. S., and Vogt, P. K. (1979). Avian erythroblastosis virus:Transformation-specific sequences form a contiguous segment of3.25 kb located in the middle of the 6-kb genome. Virology 97,366–377.
Lai, M. M., Neil, J. C., and Vogt, P. K. (1980). Cell-free translation of avianerythroblastosis virus RNA yields two specific and distinct proteinswith molecular weights of 75,000 and 40,000. Virology 100, 475–483.
Langlois, A. J., Bonar, R. A., Bolognesi, D. P., Beard, D., Beard, J. W., andRao, P. R. (1966). BAI strain A avian (myeloblastosis) leukosis virusfrom myeloblast tissue culture. Proc. Soc. Exp. Biol. Med. 123, 286–290.
Li, J., and Vogt, P. K. (1993). The retroviral oncogene qin belongs to thetranscription factor family that includes the homeotic gene fork head.Proc. Natl. Acad. Sci. USA 90, 4490–4494.
Maki, Y., Bos, T. J., Davis, C., Starbuck, M., and Vogt, P. K. (1987). Aviansarcoma virus 17 carries the jun oncogene. Proc. Natl. Acad. Sci.USA 84, 2848–2852.
Maru, Y., Witte, O. N., and Shibuya, M. (1996). Deletion of the ABL SH3domain reactivates de-oligomerized BCR-ABL for growth factor inde-pendence. FEBS Lett. 379, 244–246.
Mladenov, Z., Heine, U., Beard, D., and Beard, J. W. (1967). Strain MC29avian leukosis virus. Myelocytoma, endothelioma, and renal growths:Pathomorphological and ultrastructural aspects. J. Natl. Cancer Inst.38, 251–285.
Moscovici, C., Moscovici, M. G., Jimenez, H., Lai, M. M., Hayman, M. J.,and Vogt, P. K. (1977). Continuous tissue culture cell lines derivedfrom chemically induced tumors of Japanese quail. Cell 11, 95–103.
Nazerian, K., Stephens, E. A., Sharma, J. M., Lee, L. F., Gailitis, M., andWitter, R. L. (1977). A nonproducer T lymphoblastoid cell line fromMarek’s disease transplantable tumor (JMV). Avian Dis. 21, 69–76.
Nazerian, K., and Witter, R. L. (1975). Properties of a chicken lympho-blastoid cell line from Marek’s disease tumor. J. Natl. Cancer Inst. 54,453–458.
Neil, J. C., Delamarter, J. F., and Vogt, P. K. (1981). Evidence for threeclasses of avian sarcoma viruses: Comparison of the transformation-specific proteins of PRCII, Y73, and Fujinami viruses. Proc. Natl.Acad. Sci. USA 78, 1906–1910.
Nishizawa, M., Goto, N., and Kawai, S. (1987). An avian transformingretrovirus isolated from a nephroblastoma that carries the fos geneas the oncogene. J. Virol. 61, 3733–3740.
Nunn, M. F., Seeburg, P. H., Moscovici, C., and Duesberg, P. H. (1983).Tripartite structure of the avian erythroblastosis virus E26 transform-ing gene. Nature 306, 391–395.
Payne, L. N., and Chubb, R. C. (1968). Studies on the nature and geneticcontrol of antigen in normal chick embryos which reacts in theCOFAL test. J. Gen. Virol. 3, 379–391.
Petropoulos, C. J., Givol, I., and Hughes, S. H. (1996). Comparativeanalysis of the structure and function of the chicken c-myc and v-mycgenes: v-myc is a more potent inducer of cell proliferation andapoptosis than c-myc. Oncogene 12, 2611–2621.
Petropoulos, C. J., and Hughes, S. H. (1991). Replication-competentretrovirus vectors for the transfer and expression of gene cassettesin avian cells. J. Virol. 65, 3728–3737.
Pruschy, M., Shi, Y. Q., Crompton, N. E., Steinbach, J., Aguzzi, A.,Glanzmann, C., and Bodis, S. (1997). The proto-oncogene c-fos me-diates apoptosis in murine T-lymphocytes induced by ionizing radi-ation and dexamethasone. Biochem. Biophys. Res Commun. 241,519–524.
Rosenberg, N., and Jolicoeur, P. (1997). Retroviral pathogenesis. In“Retroviruses” (J. M. Coffin, S. H. Hughes, and H. E. Varmus, Eds.), pp.475–585. Cold Spring Harbor Laboratory Press, Cold Spring Harbor,New York.
Schaefer-Klein, J., Givol, I., Barsov, E. V., Whitcomb, J. M., VanBrocklin,M., Foster, D. N., Federspiel, M. J., and Hughes, S. H. (1988). The EV-0derived cell line DF-1 supports the efficient replication of ASLVviruses and vectors. Virology 248, 305–311.
Scheidler, S., Fredericks, W. J., Rauscher, F., Jr., Barr, F. G., and Vogt, P. K.(1996). The hybrid PAX3-FKHR fusion protein of alveolar rhabdomyo-sarcoma transforms fibroblasts in culture. Proc. Natl. Acad. Sci. USA93, 9805–9809.
Schmidt, M., Oskarsson, M. K., Dunn, J. K., Blair, D. G., Hughes, S.,Propst, F., and Vande Woude, G. F. (1988). Chicken homolog of themos proto-oncogene. Mol. Cell. Biol. 8, 923–929.
Smith, D. R., Vogt, P. K., and Hayman, M. J. (1989). The v-sea oncogeneof avian erythroblastosis retrovirus S13: Another member of theprotein-tyrosine kinase gene family. Proc. Natl. Acad. Sci. USA 86,5291–5295.
Steeg, C. M., and Vogt, V. M. (1990). RNA-binding properties of thematrix protein (p19gag) of avian sarcoma and leukemia viruses.J. Virol. 64, 847–855.
Stehelin, D., Varmus, H. E., Bishop, J. M., and Vogt, P. K. (1976). DNA
303FOCI AND PLAQUES ON DF-1 CELLS

related to the transforming gene(s) of avian sarcoma viruses ispresent in normal avian DNA. Nature 260, 170–173.
Temin, H. M., and Mizutani, S. (1970). RNA-dependent DNA polymerasein virions of Rous sarcoma virus. Nature 226, 1211–1213.
Temin, H. M., and Rubin, H. (1958). Characteristics of an assay for Roussarcoma virus and Rous sarcoma cells in tissue culture. Virology 6,669–688.
Tsuchie, H., Chang, C. H., Yoshida, M., and Vogt, P. K. (1989). A newlyisolated avian sarcoma virus, ASV-1, carries the crk oncogene. On-cogene 4, 1281–1284.
Vogt, P. K. (1969). Focus assay of Rous sarcoma virus. In “FundamentalTechniques in Virology” (K. Habel and N. P. Salzman, Eds.), pp.198–211. Academic Press, New York.
Vogt, P. K. (1971). Genetically stable reassortment of markers duringmixed infection with avian tumor viruses. Virology 46, 947–952.
Wang, L.-H., Duesberg, P. H., Kawai, S., and Hanafusa, H. (1976a). Thelocation of envelope-specific and sarcoma-specific oligonucleotides
on the RNA of Schmidt-Ruppin Rous sarcoma virus. Proc. Natl. Acad.Sci. USA 73, 447–451.
Wang, L.-H., Duesberg, P. H., Mellon, P., and Vogt, P. K. (1976b). Distri-bution of envelope-specific and sarcoma-specific nucleotide se-quences from different parents in the RNAs of avian tumor virusrecombinants. Proc. Natl. Acad. Sci. USA 73, 1073–1077.
Weiss, R. A., Mason, W. S., and Vogt, P. K. (1973). Genetic recombinantsand heterozygotes derived from endogenous and exogenous avianRNA tumor viruses. Virology 52, 535–552.
Weiss, R. A., and Payne, L. N. (1971). The heritable nature of the factorin chicken cells which acts as a helper virus for Rous sarcoma virus.Virology 45, 508–515.
Woods, C. M., Boyer, B., Vogt, P. K., and Lazarides, E. (1986). Controlof erythroid differentiation: Asynchronous expression of the aniontransporter and the peripheral components of the membraneskeleton in AEV- and S13-transformed cells. J. Cell Biol. 103,1789–1798.
304 HIMLY ET AL.
Copyright © 2022 FDOKUMEN




















