
A Single Amino Acid Substitution in the Third Transmembrane ...

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
II
Improving the nutritional quality of Lathyrus sativus L. (grass pea)
for safer consumption
Asnake Fikre Woldemedhin
EIAR

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
III
A Promoters:
Prof. Dr. Godelieve Gheysen Ghent University Department of Molecular Biotechnology Coupure links 653, B-9000, Ghent, Belgium Prof. Dr. Ir. Fernand Lambein Ghent University Institute of Plant Biotechnology for Developing Countries (IPBO), K. L. Ledeganckstraat 35, B-9000, Ghent, Belgium Dean: Prof. Dr. Ir. Herman Van Langenhove
Rector: Prof. Dr. Paul Van Cauwenberge

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
IV
Asnake Fikre Woldemedhin
Improving the nutritional quality of Lathyrus sativus L. (grass pea) for safer consumption
Thesis submitted in partial fulfilment of the requirements for the degree of Doctor (PhD) in Applied Biological Sciences

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
V
Verbetering van de nutritionele kwaliteit van Lathyrus sativus voor een veiliger consumptie Cover illustration : Field grown grass pea in Ethiopia, the green unripe pods that is often consumed by children in fields and diverse seeds from different origin. Citation : Asnake Fikre. 2008. Improving the nutritional quality of Lathyrus sativus L. (grass pea) for safer consumption. PhD thesis. Ghent University, Belgium. ISBN- number: 978-90-5989-255-2 The author and promoter give the authorization to consult and to copy parts of this work for personal use only. No part is to be reproduced without the permission of the author.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
VI
Dedicated to my very first teacher the late Sium Woldemichael who wished if among his students could attain such level, and equally to lathyrism victim children whose life remains in ruins.


Im proving the nutritional quality of grass pea (L athyrus sativus L.)
VII
ACKNOWLEDGMENTS I thank God for making my efforts fruitful. I would like to extend my deepest thanks to professor Dr. Ir. Fernand Lambein whose concern for my work was extremely important. He taught me not only the academics of knowledgeable merit but as well the value of life experience. My deepest thanks is extended to my promoter professor Dr. Godelieve Gheysen whose scientific support was extremely important. I extend my heartfelt thanks to Dr. Yu-Haey Kuo who basically trained me in laboratory work and familiarization with lab equipments. She as well kept me in good caring that made my stay comfortable and in a family feeling. I am grateful to Dr. Nancy Terryn, Dr. Kibebew Asefa, Dr. Alemu Yami, Dr. Reta Leta, Mrs Marijke Van Moorhem and Mr. Hailemariam Teklewold for their comments and/or scientific support of my thesis work. My thanks go to Prof. Dr. Marc Van Montagu, founder of the Institute of Plant Biotechnology for Developing Countries (IPBO), institute established to assist the developing world. Dr. Seid Ahmed, Dr. Million Eshete, Mr. Lijalem Korbu, Mr. Melese Dadi, Mr. Asfaw Taddese, Mrs. Konjit Ayalew, Mr. Nega Alemu, Mr. Tekalign Taddesse, Mr. Wendimagegnehu Weldesemayat, Dr. Sylvia Burssens, Mrs. Veerle van Ongeval, Mrs. Bossena and the late Mr. Kirubel Kora are highly acknowledged for their technical and/or administrative support and facilitation during the course of my work. I thank the Department of Applied Analytical and Physical Chemistry, and Department of Food safety and Food Quality for analysing my samples for micronutrient and allowing me to analyse my samples for proximate composition respectively. I like to extend my greatest thanks to the members of the examination committee: Professor Dr. Els Van Damme, Professor Dr. Ir. Patrick Van Damme, Professor Dr. Patrick Van der Stuyft, Professor Dr. Marcelle Holsters and Professor Dr. Ir. Geert Angenon for their unreserved constructive suggestions and comments. I also extend my thanks to my beloved wife Sr. Meseret Bekele who kept our children, Nahom and Yesufekad, and tolerantly stayed alone in my absence. My thank also goes to my beloved parents Mr. Fikre Woldemedhin Dawitee and Mrs. Mulunesh Habteyohanis Haile, my sisters, my brothers and relatives who were on my side that gave me courage in my work. Last but not least, I appreciably give my thanks to Ethio-Belgian (EIAR-VLIR) Lathyrus Nutritional Improvement Project (2004-2008) which was the financial resource for my study to this end.


Im proving the nutritional quality of grass pea (L athyrus sativus L.)
VIII
Abbreviations and acronyms AMPA α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid ANF Anti Nutritional Factors ANOVA Analysis of Variances AOAC Association of Official Analytical Chemists ASL Above Sea Level BAPN ß-aminopropionitrile BD Bulk Density BIA β-(isoxazolin-5-on-2-yl)-alanine DABA L-2, 4-diaminobutyric acid DAPRO α,ß-diaminopropionic acid DDW Double Distilled Water DF Days to Flowering DM Days to Maturity DG Deep Green DTPA Diethylene Triamine Pentaacetic Acid DVM Doctor of Veterinary Medicine DZARC Debre Zeit Agricultural Research Centre EIAR Ethiopian Institute of Agricultural Research FAO Food and Agricultural Organization FCE Food Conversion Efficiency GFAAS Graphite Furnace Atomic Absorption Spectrophotometry GLM General Linear Model GO Glycolate Oxidase HPLC High-Performance Liquid Chromatography ICARDA International Centre for Agricultural Research in Dry Areas ICP Inductively Coupled Plasma IPGRI International Plant Genetic Resource Institute MCF Moisture Correction Factor MJ Mega joule NSRC National Soil Research Centre OM Organic Matter β-ODAP β-N-oxalyl-L-α,β-diaminopropionic acid pH Potential of Hydrogen PITC Phenyl-iso-thiocyanate SSH Sunshine Hours TN Total Nitrogen VLIR Flemish Interuniversity Council WHO World Health Organization


Im proving the nutritional quality of grass pea (L athyrus sativus L.)
IX
Table of contents
OUTLINE............................................................................................................... XI CHAPTER I .............................................................................................................. 1 Literature review ....................................................................................................... 1
I.1. Introduction..................................................................................................... 3 I.1.1. Characteristic features of grass pea and its cultivation............................... 3 I.1.2. Grass pea consumption habits and neurolathyrism .................................... 7
I.1.2.1. Consumption...................................................................................... 7 I.1.2.2. Neurolathyrism .................................................................................. 7
I.1.3. Factors affecting the level of neuro-excitatory amino acid in grass pea ... 10 I.1.3.1. Climatic factors and ß-ODAP biosynthesis ...................................... 11 I.1.3.2. Growth media and associated changes in ß-ODAP........................... 13 I.1.3.3. Effect of grass pea processing on ß-ODAP reduction and nutritional quality ......................................................................................................... 14 I.1.3.4. Breeding or genetic engineering to reduce β-ODAP in grass pea...... 18
I.1.4. Effect of methionine on toxicity of grass pea .......................................... 21 I.1.5. Rationale of the study and motivation of the investigation ...................... 22 I.1.6. Overall and specific objectives of the study ............................................ 24
General objective......................................................................................... 24 Specific objectives....................................................................................... 24
CHAPTER II........................................................................................................... 25 Level of β-ODAP, methionine and other free and protein amino acids in the seeds of different grass pea (Lathyrus sativus L.) genotypes.................................................. 25
II.1. Introduction ................................................................................................. 27 II.2. Materials and methods ................................................................................. 29 II.3. Results ......................................................................................................... 32 II.4. Discussion.................................................................................................... 36
CHAPTER III ......................................................................................................... 39 Study of environmental factors (climatic, edaphic) affecting yield and toxicity of grass pea grown in Ethiopia in a multilocation experiment ............................................... 39
III.1. Introduction ................................................................................................ 42 III.2. Materials and methods ................................................................................ 44 III.3. Results........................................................................................................ 48 III.4. Discussion .................................................................................................. 55
CHAPTER IV ......................................................................................................... 60 Effect of methionine supplement on physical responses and neurological symptoms of broiler chicks fed grass pea based starter ration ....................................................... 61
IV.1. Introduction................................................................................................ 64 IV.2. Materials and Methods ...............................................................................65 IV.3. Results........................................................................................................ 67 IV.4. Discussion .................................................................................................. 73

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
X
CHAPTER V .......................................................................................................... 79 Effect of methionine supplement on serum amino acids status and trace elements level of broiler chicks fed with grass pea based starter ration ........................................... 79
V.1. Introduction ................................................................................................. 82 V.2. Materials and Methods................................................................................. 83 V.3. Results......................................................................................................... 85 V.4. Discussion ................................................................................................... 90
CHAPTER VI ......................................................................................................... 93 Identification of gamma irradiation derived mutant lines of Lathyrus sativus for higher methionine level ........................................................................................... 93
VI.1. Introduction................................................................................................ 95 VI.2. Materials and methods................................................................................ 97 VI. 3. Results....................................................................................................... 99 VI.4. Discussion .................................................................................................103
CHAPTER VII.......................................................................................................107 General discussion and conclusions........................................................................107
VII.1. Environmental influence on levels of amino acids and crop performances in L. sativus ............................................................................................................109 VII.2. Effect of methionine on toxicity of L. sativus ...........................................114 VII.3. Mutational methionine enrichment in L. sativus........................................117 VII.4. Recommendations and future perspectives ...............................................119
References..............................................................................................................123 Summary - Samenvatting .......................................................................................149
Summary............................................................................................................151 Samenvatting......................................................................................................155
Curriculum Vitae……………………………………………………………………156

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
XI
OUTLINE


Im proving the nutritional quality of grass pea (L athyrus sativus L.)
XII
OUTLINE
Lathyrus sativus L. (grass pea), a drought tolerant legume originating from the Near
East, has emerged with the historical beginning of agricultural practices. However,
this crop, which has been used as a pulse for at least 8000 years, has made little
progress as a food crop during this time. Prolonged and monotonous over-
consumption carries the risk of neurolathyrism, a disease causing irreversible
crippling of the legs. Producers prefer grass pea to many other legumes as production
is insured even under environmental stresses such as drought and flooding. This pulse
crop needs little or no inputs, is cheaply available, has wide adaptability and it is used
as a multipurpose crop for food, feed, forage and green manure.
In CHAPTER I, an overview of the literature is presented. Concerted breeding
research efforts have been exhaustively directed towards reducing or removing the
neuro-excitatory free amino acid β-ODAP (β-N-oxalyl-L-α,β-diaminopropionic acid)
from the plant. This nonprotein amino acid was discovered in 1964 and proposed as
the causal agent of neurolathyrism. The level of this metabolite is affected by
interdependent environmental factors, processing types and genetic factors. This
focused research approach in nutritional improvement of the crop neither resulted in
toxin free lines nor stopped neurolathyrism affliction. Also, other potential uses of the
crop have been under-exploited. In this work, experiments on comparative chemical
characterization of genotypes from different origin, on identification of important
environmental factors affecting grass pea toxicity and yield in Ethiopian context,
effect of methionine on grass pea toxicity, and in vivo methionine enrichment have
been undertaken with the following objectives: (i) Analyse factors affecting the level
of ß-ODAP, methionine and other amino acids in grass pea; (ii) assess the effect of
methionine on toxicity of grass pea in young broiler chicks; and (iii) mutagenize grass
pea seeds and select putative mutant lines with improved methionine content marker.
In CHAPTERS II & III, an extensive study into the influence of altitude, climatic
factors, geographic origin, growth media, genotypes and processing on the level of β-
ODAP, methionine and other amino acids was made. The problem statement was set
against the repeatedly published general belief that genetic variability and drought are
the only factors affecting toxicity, and therefore, hypothesized that there could be
other factors affecting toxicity and crop yield in Ethiopia. Both the free and protein
amino acids of the samples were quantitatively determined using HPLC analysis. Soil

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
XIII
nutrients were determined by flame ionisation analysis. Variability in amino acid
content of genotypes from different origin has been demonstrated. Important
environmental factors affecting the concentration of β-ODAP and yield were
investigated and possible interrelations were discussed. From the results, novel
conclusive remarks were made that several environmental factors interplay in
determining the level of β-ODAP and grass pea yield.
In CHAPTERS IV & V, the role of methionine on grass pea toxicity was studied by
examining chicks in a starter feeding experiment. The hypothesis was that methionine
addition could counteract grass pea toxicity and its aftermath. Performances of 450
chicks subdivided into two grass pea feed levels, each with four different levels of
DL-methionine supplement, were evaluated. Statistical analysis (ANOVA) was
carried out to compare treatment effects on feed intake, weight gain, and feed
conversion efficiency. Development of symptoms such as neck twisting was
investigated and phenotypically described. Serum amino acids and micronutrient
levels in the serum of the young chicks was determined against treatment levels and
time.
In CHAPTER VI, methionine enrichment of grass pea to improve its sulphur amino
acid deficient protein was attempted by gamma-irradiation mutagenesis. According to
Getahun et al. (2005) increased sulphur amino acid content of the diet contributed to
the protection of human consumers from neurolathyrism. The hypothesis is that in
vivo methionine enrichment in grass pea through mutation could be attained and that
it would contribute to food safety. Gamma ray mutagenized grass pea plants
exhibiting different phenotypical appearances were selected by observation in the
growing field. Methionine enhanced putative mutant lines were identified,
characterized and typical features were described. Deep green leaf was demonstrated
to have higher methionine level, therefore showing the typical morphological marker
of methionine enrichment as previously used by others in soybean.
Finally, in CHAPTER VII of the thesis, the main findings of the reported research
were discussed and conclusions drawn. Recommendations were derived and future
perspectives proposed.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
1
CHAPTER I
Literature review

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
2
Lathyrus sativus L. (a) part of a flowering branch, (b) flower in front and side view, (c) dorsal petal (bottom),
wings and keel (from right), and (d) pod with seeds
Adopted from Campbell (1997).

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
3
CHAPTER I
Literature review
I.1. Introduction I.1.1. Characteristic features of grass pea and its cultivation
Lathyrus sativus L., commonly known as grass pea, chickling vetch or chickling pea,
is commercially an important legume in the tribe Viciae of the Fabaceae family. The
genus Lathyrus is characterized by the presence of leaf tendrils in the majority of its
members, an unusual arrangement of vascular tissue in the stem and characteristic
floral parts (Goyder, 1985). It comprises more than 150 species with 15 (Smartt et al.,
1994) or 13 (Kupicha, 1983) intragenic sections where many members, viz L. sativus,
L. hirsutus, L. odoratus, L. latifolius, L. cicera, L. clymenum, etc., have commercial
importance to mankind (Kupicha, 1983; Yamamoto et al., 1986; Yan et al., 2006).
Relationship studies between species in Lathyrus indicated that only two, i.e L.
amphicarpos and L. cicera, gave viable hybrids when crossed with L. sativus while
six others produced pods when crossed with the cultivated species (L. sativus and L.
cicera) which, however, contained either shrivelled seeds or were totally aborted
(Smartt, 1990; Smartt et al., 1994).
The epigrammatic taxonomic features of Lathyrus sativus have been reviewed by
several authors (Duke, 1981; Plitmann et al., 1985 and Campbell, 1997). Owing to the
large variation in several of its characters, only general common features are
discussed.
Grass pea is a herbaceous annual with straggling or climbing growth habit, branched
and with a sub-erect standing of 0.6-0.9 meters long. Stems are slender and
quadrangular with winged margins. Stipules are prominent, narrowly triangular to
ovate with a basal appendage.
The pinnately compounded opposite leaves of 30-150 mm long by 3-11 mm wide are
linear. Leaflets are lanceolate with simple or much-branched tendrils that are modified
from the terminal leaflets.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
4
Flowers having a short (10-15 mm) and slender pedicel are solitary, axilliary, on a
peduncle of 30-60 mm; petals are erect and spreading, ovate 15 x 18 mm, finely
pubescent at upper margin, clawed, flat, and reddish-purple, pink, blue or white in
colour; wings are ovate, 14 x 8 mm, clawed and obtuse at top; keel is slightly twisted,
boat-shaped, 10 x 7 mm, entirely split dorsally, ventrally split near the base; calyx
teeth glabrous and longer than the calyx tube. Stamens are diadelphous (9+1) with
vexillary stamens free, 9 mm long, winged at base, apical part filiform, slightly
winged; anthers are elliptoid, 0.5 mm long and yellow; pollen 49-57 µm long; flowers
are predominantly self-pollinated. Ovary is sessile, thin, 6 mm long, pubescent with 5-
8 ovules; style is abruptly upturned, 6-7 mm long, widening at tip, and bearded below
the stigma; stigma is terminal, glandular-papillate and spatulate.
Pods are oblong, 2.5-4 cm long, flat, slightly curved, dorsally two-winged, with 2-5
seeds per pod. The seed is white, greyish-brown or yellowish, usually spotted or
mottled and sometimes smooth, 3-15 mm long by 7-9 mm diameter, round or angular
wedge-shaped; hilum is elliptic, with 1/15th to 1/16th of the circumference of a seed.
Germination is hypogeal, the epicotyls are purplish-green; the first leaf is small, scale-
like, often fused with two lateral stipulae. The second leaf is sublate, connected at the
base with the stipulae. Chromosomal number is 2x = 2n = 14 (Narayan, 1998). L.
sativus has a well-developed taproot with rootlets often covered by cylindrical
nodules.
The combination of archaeo-botanical and phyto-geographical evidence indicates that
the origin of L. sativus was in the Balkan Peninsula. It was already cultivated in the
early Neolithic period, at the beginning of the 6th millennium BC (Kislev, 1989). This
author also suggested that the practice of cropping annuals, including cereals and
legumes such as pea and lentil, introduced from the Near East around 6000 BC,
enabled domestication of L. sativus in this region. This means that L. sativus is
perhaps among the first crop domesticated in Europe as a consequence of expansion
of agriculture from the Near East. It is probably derived from the genetically nearest
wild species L. cicera (Hopf, 1986).

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
5
Man’s cultivation of grass pea dates back to the prehistoric era, and evidently
originated around the Mediterranean Sea. South East Asia and Ethiopia are traceable
early cultivation sites along with or right after its domestication period (Renfrew,
1979; Hopf, 1986; Kislev, 1989; Kockhar, 1992). Smartt (1984) found it rather
puzzling that a crop, which has been used as a pulse for at least 8000 years and is still
so used, should have made so little evolutionary progress as a grain crop during this
time. He considered that the lack of progress as a pulse crop might have been due to
its possibly more important use as a forage crop. Thus, in many ways, different
selection pressures imposed on the crop may have cancelled each other and must have
resulted in limited improvement of agronomic traits over this long period.
In some countries like Italy, grass pea is receiving renewed attention as a local and
traditional product; it is becoming an exclusive and fashionable food for which
consumers are prepared to pay a higher price than for other pulse products (Polignano
et al., 2005).
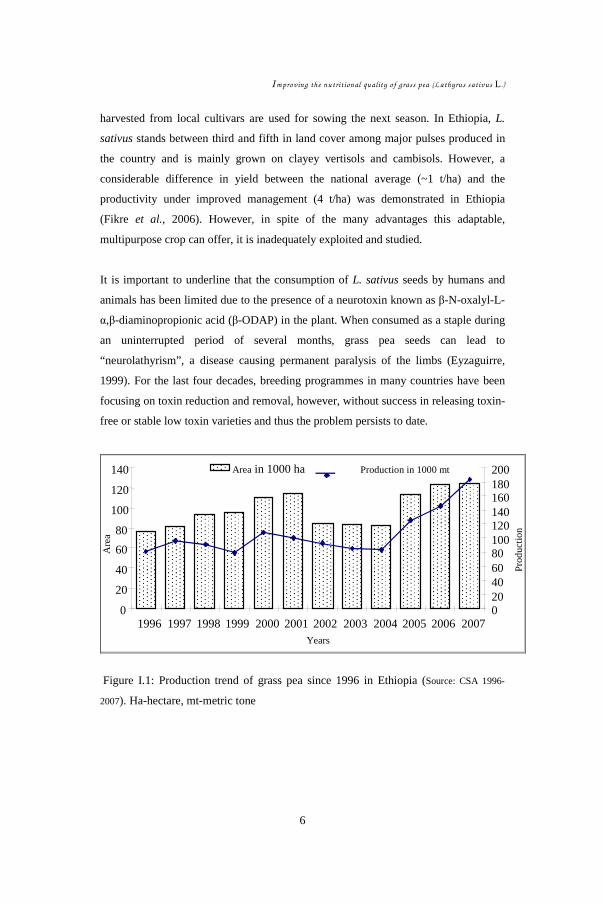
For the specific case of Ethiopia, grass pea has shown increasing trend both in
consumption and production. In the last decade alone its production area has
increased from about 80 thousand hectares to more than 120 thousand hectares
(Figure I.1). It has gained more than 100 % production increase over the last three to
four years, which likely resulted from both improving productivity and an increase in
production area. This could be attributed from the overall increasing trend, for
example, of total agricultural areas by 6 % and grass pea area by 4 % (Dadi et al.,
2003), and also due to increased preference for its insured production under low input
in marginal areas and with such unpredictable climatic situations. Since four years,
grass pea has got momentum of an average 8 % annual increase in production (Figure
I.1). Based on FAO grass pea production statistics (1996-2007), it can be calculated
that Ethiopia accounts for 75.3 % and 9 % in area cover and 85.4 % and 8 % in
production of Africa and the world, respectively.
The most interesting agronomical features of the species are its drought tolerance (as
it can grow with residual soil moisture alone), resistance to pests and diseases,
adaptability to different types of soil as well as to adverse climatic conditions (Noto et
al., 2001). The crop can be produced without any modern inputs whereas seeds
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
6
harvested from local cultivars are used for sowing the next season. In Ethiopia, L.
sativus stands between third and fifth in land cover among major pulses produced in
the country and is mainly grown on clayey vertisols and cambisols. However, a
considerable difference in yield between the national average (~1 t/ha) and the
productivity under improved management (4 t/ha) was demonstrated in Ethiopia
(Fikre et al., 2006). However, in spite of the many advantages this adaptable,
multipurpose crop can offer, it is inadequately exploited and studied.
It is important to underline that the consumption of L. sativus seeds by humans and
animals has been limited due to the presence of a neurotoxin known as β-N-oxalyl-L-
α,β-diaminopropionic acid (β-ODAP) in the plant. When consumed as a staple during
an uninterrupted period of several months, grass pea seeds can lead to
“neurolathyrism”, a disease causing permanent paralysis of the limbs (Eyzaguirre,
1999). For the last four decades, breeding programmes in many countries have been
focusing on toxin reduction and removal, however, without success in releasing toxin-
free or stable low toxin varieties and thus the problem persists to date.
0
20
40
60
80
100
120
140
1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 Years
Are
a
0 20 40 60 80 100 120 140 160 180 200
Pro
du
ctio
n
Area in 1000 ha Production in 1000 mt mmtmtMt
Figure I.1: Production trend of grass pea since 1996 in Ethiopia (Source: CSA 1996-
2007). Ha-hectare, mt-metric tone

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
7
I.1.2. Grass pea consumption habits and neurolathyrism
I.1.2.1. Consumption
Unlike other food legumes, grass pea is used as a staple food. It is an important source
of dietary proteins, calories and minerals for humans (Urga et al., 2005). Grass pea is
a nutritious and stress-surviving crop. Seed of Lathyrus sativus is prepared into
traditional dishes after dehusking and parching or milling. As a drought tolerant food
crop, it is a very promising source of protein, carbohydrate and minerals for drought
prone marginal lands with seeds containing an average of 27 % protein, 0.6 % fat,
58.2 % carbohydrate (about 35 % starch) (Duke, 1981; Williams et al., 1994; Urga et
al., 2005; Vaz Patto et al., 2006). The seeds contain 1.5 % sucrose, 6.8 % pentosans,
3.6 % phytin, 1.5 % lignin, 6.69 % albumin, 1.5 % prolamine, 13.3 % globulin, and
3.8 % glutelin. The concentration of essential amino acids in g per 16 g of N
(equivalent to 100 g of protein) reported were: arginine 7.85, histidine 2.51, leucine
6.57, isoleucine 6.59, lysine 6.94, methionine 0.38, phenylalanine 4.14, threonine
2.34, tryptophane 0.40, and valine 4.68 (Duke, 1981; Williams et al., 1994). Like
other cool season food legumes, grass pea is highly deficient in methionine, cysteine
and tryptophane.
Grass pea has been reported to have toxic metabolites causing irreversible crippling in
humans. Several substances that are toxic to laboratory animals have been identified
from seeds of Lathyrus species including ß-ODAP in L. sativus, the γ-glutamyl
derivative of ß-aminopropionitrile (BAPN) in L. odoratus, L. hirsutus, L. pussilus and
L. roseus, and L-2,4-diaminobutyric acid (DABA) in L. latifolius (Barrow et al.,
1974; Roy and Spencer, 1989; Foster, 1990). In addition to L. sativus, ß-ODAP is
produced in 17 species of Acacia and 13 species of Crotalaria (Quereshi et al., 1977),
and in only one non-legume: Panax ginseng (Long et al., 1996; Kuo et al., 2003).
I.1.2.2. Neurolathyrism
Neurolathyrism is an irreversible disease caused by prolonged dietary dependence on
grass pea (Getahun et al., 2005; Haimanot et al., 2005). A non-protein amino acid,
glutamate analogue, that was identified four decades ago (Rao et al., 1964), ß-N-

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
8
oxalyl-L-α,ß-diaminopropionic acid (ß-ODAP), was proposed as the causative
molecule.
COOH
CO
NH – CH2 – CH – COOH
NH2
Figure I.2. Chemical structure of ß-N-oxalyl-L-α,ß-diaminopropionic acid (ß-ODAP)
Over thirty different physiological and biochemical activities have been ascribed to
this non-protein amino acid (Lambein et al., 2007). The best studied activity of ß-
ODAP is the excitation of a subclass of glutamate receptors on the neuronal cells
(Spencer et al., 1986). This activity triggers the release of the endogenous neuro-
excitant glutamate, whereas ß-ODAP also inhibits the re-uptake of glutamate by the
astrocytes in the central nervous system (CNS) that would ‘detoxify’ glutamate into
glutamine. This not only increases and prolongs the excitation of the receptors, but
also increases the production of nitric oxide (NO), a short-lived molecule that is
responsible for the production of reactive oxygen species (ROS) and oxidative stress.
An interesting activity of ß-ODAP in the plant is reported to be the protection of
glycolate oxidase, an enzyme in plant photosynthesis, against high light intensity. The
level of ß-ODAP in the plant increases with higher light intensity (Zhang et al., 2003).
Faced with drought-triggered crop devastation, people are confronted with a dilemma
of either starvation or survival on the only foods available; i.e. grass pea with its toxic
reputation, knowing that especially the young men, as the breadwinners in the family,
are most apt to succumb to neurolathyrism. The tragic consequences of its unbalanced
consumption may have been recognized shortly after its domestication.
Neurolathyrism mostly affects the poor subsistence farmers families who often are
illiterates and are least informed about the disease. There is a strong perception of the
public about this stigmatising disease that is not normally reported to medical or

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
9
administrative authorities (Getahun et al., 2002c). This underreporting of
neurolathyrism may in part be responsible for its status of ‘neglected disease’.
The typical feature of neurolathyrism is spastic paraparesis of the legs, whereby the
heels can hardly touch the ground, accompanied by the easily recognised ‘scissor
gate’ when the knees touch each other while walking (Ludolph et al., 1988). The term
’’motor neurone disease’’ encompasses independent or combined upper and lower
motor neurone disorders (Donaghy, 1999). There is increasing evidence that the
neurotoxic effects of excitatory amino acids and their analogues are part of the
pathogenesis of this neuronal degeneration primarily affecting corticospinal tracts
regulating the lower limbs (Ludolph et al., 1993). The presence of such a mechanism
in combination with oxidative stress could play a pathogenic role (Getahun, 2004;
Nunn et al., 2005).
Daily overconsumption of grass pea (up to 1 kg during 2-3 months) without
interruption and apparently with no admixture of herbs or condiments is considered to
lead to symmetric crippling of the legs (Getahun et al., 2005). The estimated threshold
intake of ß-ODAP per day per adult person is about 500 mg (Lambein et al., 2001).
The generally suggested cause of neurolathyrism is the agonistic activity of ß-ODAP
at glutamate receptors of neurons, but this cannot explain the extreme inter-species
and intra-species differences in susceptibility to ß-ODAP (Rao, 2001). The later
author indicated that differences in individual physiological activity ascribe for
differences in susceptibility or resistance to the disease in humans.
Neurolathyrism is an important public health problem in some developing countries:
e.g. India, Bangladesh, Nepal, Ethiopia and NW China, which are known for having
high populations and food insufficiency (Haque, 1997). Especially the subsistence
farmers in remote areas that are susceptible to droughts and where grass pea is
considered a survival food are at risk. Hundreds of thousands in the world and tens of
thousands in Ethiopia are living with neurolathyrism. Poverty, illiteracy, malnutrition,
recurrent natural calamities and traditional agriculture are common features of regions
with this problem. The predisposing factors and the sudden onset of neurolathyrism
symptoms are poorly understood, however. Particularily in Ethiopia, sex, age,
illiteracy, poverty and cooking of grass pea foods exclusively in traditional clay pots
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
10
are also reported to be risk factors of neurolathyrism (Getahun et al., 2002a).
Accordingly, they reported, male, young age, illiterates, poor and those using clay
cooking utensils were found to be more susceptible.
Patients of neurolathyrism complain of walking difficulties, leg stiffness and
heaviness of the lower limbs and of frequency of micturition (sense of urination). Gait
abnormalities include spasticity of the calf muscles, scissor gait, toe walking and the
need of walking sticks (Misra et al., 1993; Haque, 1997; Getahun et al., 2005).
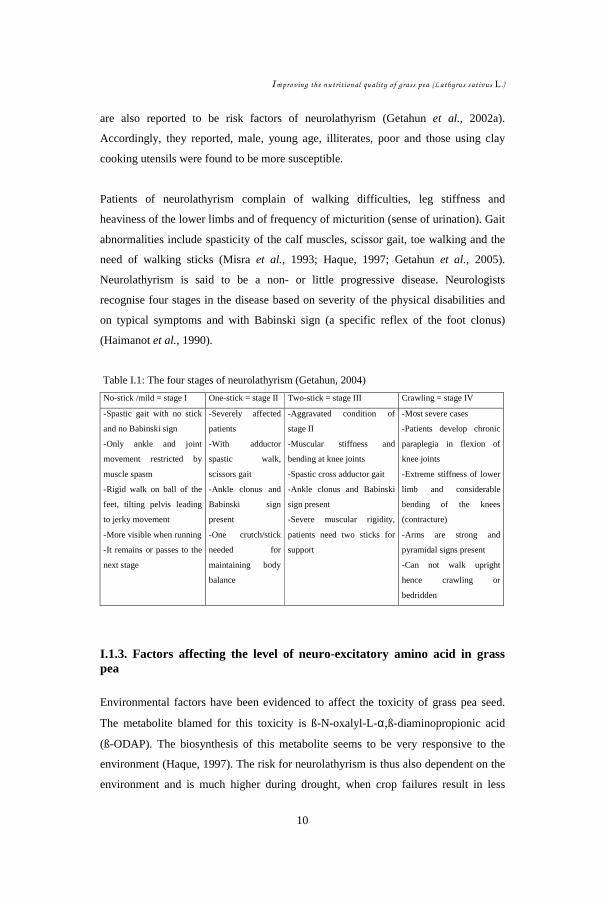
Neurolathyrism is said to be a non- or little progressive disease. Neurologists
recognise four stages in the disease based on severity of the physical disabilities and
on typical symptoms and with Babinski sign (a specific reflex of the foot clonus)
(Haimanot et al., 1990).
Table I.1: The four stages of neurolathyrism (Getahun, 2004)
No-stick /mild = stage I One-stick = stage II Two-stick = stage III Crawling = stage IV
-Spastic gait with no stick
and no Babinski sign
-Only ankle and joint
movement restricted by
muscle spasm
-Rigid walk on ball of the
feet, tilting pelvis leading
to jerky movement
-More visible when running
-It remains or passes to the
next stage
-Severely affected
patients
-With adductor
spastic walk,
scissors gait
-Ankle clonus and
Babinski sign
present
-One crutch/stick
needed for
maintaining body
balance
-Aggravated condition of
stage II
-Muscular stiffness and
bending at knee joints
-Spastic cross adductor gait
-Ankle clonus and Babinski
sign present
-Severe muscular rigidity,
patients need two sticks for
support
-Most severe cases
-Patients develop chronic
paraplegia in flexion of
knee joints
-Extreme stiffness of lower
limb and considerable
bending of the knees
(contracture)
-Arms are strong and
pyramidal signs present
-Can not walk upright
hence crawling or
bedridden
I.1.3. Factors affecting the level of neuro-excitatory amino acid in grass pea
Environmental factors have been evidenced to affect the toxicity of grass pea seed.
The metabolite blamed for this toxicity is ß-N-oxalyl-L-α,ß-diaminopropionic acid
(ß-ODAP). The biosynthesis of this metabolite seems to be very responsive to the
environment (Haque, 1997). The risk for neurolathyrism is thus also dependent on the
environment and is much higher during drought, when crop failures result in less

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
11
balanced diets, food insufficiency and also increasing toxicity of the then possiblly
available grass pea. Based on the current annual production of 100,000 tons grass pea
seeds, the production of ß-ODAP could be in the order of 400 tons annually from
Ethiopian soil alone.
I.1.3.1. Climatic factors and ß-ODAP biosynthesis
Climatic factors play a crucial role in plant adaptation to environments that can be
expressed by growth, production and level of plant biosynthates. There appear clear
differences in concentrations not only for ß-ODAP but also for other antinutritional
factors (ANF) of grass peas such as tannins, phytic acid and trypsin inhibitor when
grown in different agro-ecologies (Urga et al., 2005). Also, some protein and non-
protein amino acids with similar but less potent activities are present in grass pea
(Ikegami et al., 1995). Among these are glutamate, aspartate and BIA or ß-
(isoxazolin-5-on-2-yl)-alanine that are synthesized in the grass pea plant. BIA can
reach high concentrations in the seedling stage (cotyledon removed): up to 2 % of the
dry weight (Lambein et al., 1976). Other metabolites such as total phenolics and
condensed tannins have been shown to be more correlated with cultivars than with
environment (Wang et al., 1998a).
Several authors confirmed the wide variation in ß-ODAP concentration both among
genotypes and environments independent of the method of analysis (Dahiya and
Jeswani, 1975; Leakey, 1979; Ramanujam et al., 1980; Campbell, 1997; DZARC,
2003). Based on an evaluation on three locations with different growing environments
of two common Lathyrus species (407 L. sativus and 96 L. cicera lines) collected
from three geographic origins (Ethiopian, Mediterranean and European) Hanbury and
co-workers (2000a) found that for both species, the genotype was the most important
determinant of ß-ODAP concentration and that environment had less influence.
However, Wuletaw (2003) concluded for L. sativus that genotype and its interaction
with the environment is the most significant determinant for ß-ODAP level. The
repeated multilocation trial of Debre Zeit Agricultural Research Centre (DZARC) on
fields ranging from lower altitude (altitude 1600 m asl, light soil, short rainy period
and a mean annual temperature of 23.3 oC) to higher altitude (altitude 2200 m asl,
vertisol soil, longer rainy period, and a mean annual temperature of 16.7 oC) areas

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
12
(DZARC, 2003) together with specific observations by Campbell (1997) indicated
that ß-ODAP was reduced by more than half for the same cultivars by varying the
environment from high to low drought stress.
A 25 % increment in ß-ODAP in the seeds was observed when planting date was
delayed by two to three weeks (Asfaw et al., 2003). This might have coincided with a
drop in residual moisture in the soil. Cocks and co-workers (2000) found that a
greater influence (up to +100 %) on seed neurotoxin levels resulted from drought
stress applied at early post-anthesis compared to the period of pre-anthesis.
Considering the seed as a physiological sink for ß-ODAP, the amount of grass pea
seed per plant affects the ß-ODAP level. A negative relationship between ß-ODAP
concentration in the seed and total amount of ß-ODAP in the plant was then plausible.
Under optimal growth conditions, the plant produces more pods and seeds than it does
under marginal conditions, in which the reduced ß-ODAP concentrations obtained
may be caused by a dilution of the available toxin over a greater number of seeds.
Even under identical unstressed conditions, plants with only 10 pods produced seeds
with 59 % more ß-ODAP than plants with 100 pods (Cocks et al., 2000).
Haque (1997) studied stress phenomena in grass pea and found a strong correlation
between arginine and homoarginine accumulation in the seed that could be linked to
almost any type of nutritional or environmental stress. He suggested that the non-
protein amino acid homoarginine might act as a central defence for L. sativus to
various stresses. He further proposed that the higher accumulation of ß-ODAP in
drought-stressed L. sativus plants and seeds during the ‘survival phase’ and ‘cell
disorganization phase’ might have resulted from disturbed nitrogen assimilation and
loss of specificity of particular enzymes that might lead to higher synthesis of
asparagine. Asparagine is considered the primary precursor of the isoxazolinone ring
of BIA which is the precursor for ß-ODAP (Kuo et al., 1994 and 1998). During plant
growth, the highest β-ODAP concentration in grass pea shifts from active vegetative
leaf tips to the sinks of the reproductive stage (embryos, cotyledons).
Many plants react to drought stress by increasing their concentration of amino acids
and other osmolytes. Lambein and co-workers (1990) postulated that the free amino

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
13
acids including ß-ODAP might have a role in conveying tolerance to drought that has
made grass pea such a useful species in drought-prone areas over the long period of
its domestication.
I.1.3.2. Growth media and associated changes in ß-ODAP
Grass pea grows on diverse soil types, ranging from nutritionally rich to depleted
marginal lands. In Ethiopia, it grows predominantly on heavy vertisols (that accounts
for about 12 million hectares in the country) under both excessive moisture with poor
rhizosphere aeration and drought stress with typically cracking soils. The
physiological mechanism for survival under such diverse moisture regimes and the
ability of the crop to supply the root with oxygen in poorly-aerated soil are not well-
understood. Eavis (1971) postulated that benefits from additional residual soil water
for plant growth, following a period of water logging or water saturation of the soil,
compensate for the effects of poor aeration. We found no scientific explanation why
grass pea roots that grow under submerged conditions can survive with little oxygen
from the surrounding soil and water or, as in the case of flooded rice and some bog
plants where specialised tissues can facilitate obtaining sufficient supplies of oxygen.
Greenwood (1968) postulated O2 transport through air passages from the aerial parts
to the roots. However, we found no reference to such specialised aerenchyma tissues
for grass pea.
Most soil micro- and macronutrients affect ß-ODAP synthesis and other related amino
acids in grass pea. Haque (1997) found that high Fe2+, Mn2+, B3+, Co2+, and Al3+
increased synthesis and accumulation, while low Zn2+, Mo6+, Cu2+ and Al3+ decreased
the value. High ratios of P2+/Zn2+ and Fe2+/Zn2+ increase the ß-ODAP production. ß-
ODAP has been proposed as carrier molecule for the zinc ion and, therefore, higher
Zn2+ could reduce its synthesis. The same author (Haque, 1997) indicated that
depletion of zinc from the soil results in higher ß-ODAP levels in the plant and in the
seeds.
Once germination is established, grass pea tolerates a certain level of soil salinity,
principally the presence of Na+, K+, Ca2+, and Mg2+. These cations are commonly
linked to the anions Cl- and SO42- (Jackson, 1973). Interestingly, moderate salinity has

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
14
a decreasing effect (up to 0.6 % NaCl of artificial salinity in hydroponic cultures and
in the field of EC = 13 m mohs/cm) while high salinity has an increasing effect on ß-
ODAP accumulation (Haque, 1997).
I.1.3.3. Effect of grass pea processing on ß-ODAP reduction and nutritional quality
A diversity of recipes that are made from grass peas alone or mixed with other pulses
exists in Ethiopia. Grass pea can be consumed in different ways: roasted, cooked,
boiled, fermented or raw. In various other parts of the world, recipes using grass pea
seeds or green shoot tips are prepared and consumed with condiments (herbs, spices,
salt). Some recipes are used for daily home consumption while some others are only
used on specific festive occasions.
Any grass pea seed processing seems to affect ß-ODAP level present in it (Srivastava
and Khokhar, 1996). The most popular processing of soaking and decanting the water
reduces ß-ODAP content, but this processing also results in the simultaneous
depletion of some other important amino acids and water-soluble vitamins and
mineral nutrients. As a rule, cooking, roasting and fermentation decrease protein
solubility but improve protein digestibility in legumes (Baik et al., 2004).
Fermentation of cereals and their blend with food legumes is a potentially important
processing method that can be expected to improve the balance of essential amino
acids while reducing antinutritional factors (Kuo et al., 1995; Yigzaw et al., 2001),
hence improving their nutritional value. It can decrease certain antinutritional factors
(ANF) like phytates, protease inhibitors or flatulence factors (Kuo et al., 1995).
Significant levels of ANF in soybean, such as trypsin inhibitors and phytic acid, leach
out or are destroyed during soaking and cooking, as well as during fermentation
(Tawali et al., 1998). Yigzaw and co-workers (2001) reported that fungal
fermentation of teff (Eragrostis teff)/grass pea mixtures (teff, a fine seeded crop, is the
traditional staple cereal in Ethiopia) in a ratio of 8/2 was found to be quite comparable
in essential amino acid profile to an ideal reference protein recommended for children
of 2-5 years of age (FAO/WHO, 1985). Solid state fungal fermentation of grass pea
using Rhizopus oligosporus and Aspergillus oryzae strains in succession has shown

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
15
that the neurotoxin β-ODAP was removed by 97 % on average from high-toxin
varieties and by up to 80 % from low-toxin varieties. It was also reported that
fermentation of grass peas the same way as teff is fermented during the preparation of
the Ethiopian traditional injera, is a protective factor in the epidemiology of
neurolathyrism (Getahun et al., 2003). However, the directly baked unleavened bread
or kita is less safe than injera prepared from grass pea as the latter involves
fermentation (Getahun et al., 2003).
Table I.2. Some traditional foods prepared from L. sativus
Cou
ntr
y
Traditional grass pea food
Brief description of the preparation for consumption
Ref
ere
nce
Shiro wot (sauce)
Sauce prepared from roasted L. sativus powder and eaten with pancake (injera) or unleavened bread (kita)
Kik wot Roasted, de-husked and split L. sativus seed made into sauce Kollo (roasted snack)
Roasted seed directly consumed
Nifro (boiled snack)
Boiled seed eaten directly
Kita (unleavened bread)
Unleavened bread usually made from unfermented dough of L. sativus consumed in time of acute food problem
Eshet (green peas)
The seed in the green pod before maturity, usually consumed by children while tending the farm field
Elbet (sauce) Powder of roasted grass pea well spiced and fermented, occationally consumed like in fasting periods of coptic Christians
Seteto Baked kitta after fermentation of grass pea dough usually on clay pans
Eth
iopi
a
Injera (pancake bread)
Fermented and baked, from powder usually adulterated with cereals
Hai
ma
not e
t al.,
19
93;
1995
; G
eta
hun et
al,
2005
; U
rga
et
al.,
200
5; F
ikre
, 200
7
Dahl Aqueous slurry cooked with spices Curry Cooked with vegetables and/or fish Bora Paste ball with onion and spices, deep fried in oil Chapatti /roti Unleavened bread of khesari powder
Ba
ngla
desh
Khichuri Cooked with rice in 3:1 ratio H
ussa
in,
199
4
Chapatti L. sativus flour is routinely used either alone or in combination with wheat, barley or Bengal gram as chapatti
Ghotu L. sativus flour and rice in ratio of 3:1 cooked in water to form a stiff porridge
Dal (soup- like dish)
Cooked whole split or de-husked grains into a soft semi-solid form and eaten as side dish with rice or chapatti
Indi
a
Snacks As substitute of pigeon pea and chick peas Ma
lek,
199
8
W.A
sia
Green vegetable
Young vegetative parts plucked (4-6 cm) and cooked as green vegetable also rolled and dried for off-season use Green pods/seeds eaten directly or salted, cooked and consumed as snacks (India, Bangladesh, Pakistan) C
am
pbe
ll, 1
997

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
16
The neurotoxic amino acid (β-ODAP) is water soluble. As a preventive measure by
consumers, aqueous removal of the soluble neurotoxin from grass pea seed seems to
have co-evolved with its consumption. Grass pea seeds are commonly boiled and
consumed as a pulse. Aqueous detoxification can be done by boiling or by soaking the
seeds overnight and subsequently draining out the supernatant (Bell, 1964; Mohan,
1966). A total replacement of Ethiopian Noug cake (the solids after extracting oil
from niger seed Guizotia abyssinica) by boiled grass pea in a chick diet was found not
to reduce the chicks’ performance significantly (Tadelle et al., 2003). The latter
authors also suggested cooking at 90 °C as a means to improve the nutritional quality
since it showed similar results to the commercial poultry ration (p<0.05) in terms of
feed efficiency. Padmajaprasad and associates (1997) demonstrated a significant
reduction of ß-ODAP by up to 90 % after boiling. Wang (2000) indicated that the
extent of isomerization of the toxin from the naturally occurring toxic ß-form to the
less toxic α-form during cooking might affect the toxicity of the preparation. This
isomerisation is time- and temperature-dependent and results in an equilibrium of 40
% α- and 60 % β-isomer. Toxicity of L. sativus seed due to ß-ODAP is only partially
removed during cooking and a significant proportion of the toxin (about 60%)
remains as the toxic β-form. From comparable experiments, Akalu and co-workers
(1998) reported a statistically significant reduction (57 %) after 60 minute pressure-
cooking at 150oC of grass pea flour and a 39 % reduction after a 30-minute
autoclaving of the dry seeds. Cho and co-workers (2008) reported a remarkable
reduction of the toxin by steaming, which may suggest the need to investigate the
presence or absence of ß-ODAP in the steam from boiling or steamed grass pea.
Roasting also reduces the neurotoxin concentration of grass peas. Akalu and co-
workers (1998) recorded a significant ß-ODAP reduction with up to 30 % after
roasting milled samples and up to 67 % after cooking presoaked seeds compared to
that of whole raw seeds. Girma (1999) indicated that ß-ODAP content in whole seeds
could be reduced by up to 87 % by using various combinations of roasting, cooking,
autoclaving and soaking.
Soaking grass pea seeds prior to cooking can leach out ß-ODAP and decrease the risk
of neurolathyrism by half (Getahun et al., 2005). Soaking (12 h) of the grits (1.5-2
mm) from the cotyledons and roasting at a temperature of 200 °C for a period of 37

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
17
minutes could be recommended for a significant reduction of ß-ODAP, while in vitro
digestibility of starch also improved (Girma, 1999). Likewise, Malek (1998) reported
that soaking seed in water for 7-8 hours and then decanting off the water removes
most ß-ODAP. He further claimed that soaking split seeds overnight and decanting
water made the dal (preparation of pulses that have been stripped of their outer hulls
and split; also referring to the thick, spicy stew prepared therefrom, a mainstay of
Indian, Pakistani, and Bangladeshi cuisine) toxin-free and safe for consumption.
However, when grass pea powder (besan) is used for making pakoras, chapatis or
dalpuri, the danger of neurolathyrism still remains. By soaking mature grass pea seeds
for 72 hours in different soaking media, Kelbessa and Mengistu (1993) found an 11
and 13 % increase of total protein after soaking in plain water and 1 mM HCl,
respectively, and 12 and 16 % decrease in 1 mM NaOH and 0.1 % wood ash
solutions, respectively. They noticed that this processing resulted in the loss of total
solids, non-protein nitrogen, total soluble sugars and reducing sugars. Previous
research indicates that sprouting legumes improved their nutritive value (ability of
food to provide a usable form of nutrients: protein, carbohydrates, vitamins and
minerals) due to an increase of enzyme activity, which breaks down storage protein
and starch in the seed into amino acids, peptides, and simpler carbohydrates needed
for the embryo to grow. Simultaneously, antinutritional factors such as enzyme
inhibitors and other anti-nutrients are brought down to insignificant levels (Vidal-
Valverde et al., 2002).
Extrusion of grass pea seed reduced the ANFs, such as tannins by 77 %, the trypsin
inhibitor to below detection limit, and ß-ODAP by 46.09 % (Masoero et al., 2004). In
peas (Pisum sativum) extrusion causes enzymatic degradability of starch from 11.80
% in meal to 39.70 % in the extruded product while in faba beans (Vicia faba)
extrusion increased starch digestibility from 11.39 % to 85.05 %, whereas in lupines
(Lupinus spp.) it caused a complete starch hydrolysis (Masoero et al., 2004). Some
ANFs like phytates and nutrient components were not affected by extrusion.
Obviously, the amino acid value of a mixed diet containing cereals rich in methionine
and legumes rich in lysine is more complete than either of them consumed
individually. An interesting illustration of this is the detailed report by Kessler (1947)
on a WWII prisoners of war camp in the Ukrainian town of Vapniarca. Inmates were

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
18
given the "horse fodder" left by the Russian cavalry consisting of grass pea. They
received daily rations of 200 g boiled grass pea seed and 200 g barley bread
containing 20 % chaff. The 150 inmates did not develop any sign of lathyrism during
3 to 6 months after which the diet was changed to 400 g boiled grass pea and the same
200 g bread. Within two months after this diet with a higher portion of grass pea was
initiated, cases of crippling started and after 4 months 60 % of the inmates had
developed various degrees of neurolathyrism. Lambein and associates (2001)
estimated a threshold intake level of 500 mg of ß-ODAP per day to develop this high
incidence of neurolathyrism under severe conditions of malnourishment and physical
exhaustion. In an Encyclopaedia of Plants published in 1855 (Loudon, 1855), it was
already reported that bread made from a 50/50 mixture of grass pea and wheat seems
to have no deleterious effect, while bread made only from grass pea caused paralysis
of the legs “when used in continuance”. Thus, if only 0.5 kg of grass pea is consumed
daily in combination with cereals, the tolerance level might be as high as about 2 g of
β-ODAP intake per day (Lambein et al., 2001). Mixing the food with gravy that
contains condiments with antioxidant activity reduced the neurolathyrism cases
manyfold (Getahun et al., 2005). The latter authors confirmed that consumption of
grass pea mixed with cereals rich in sulphur amino acids was also highly
neurolathyrism-protective. Getahun and co-workers (2003) investigated the impact of
cereal aid during drought-triggered famine and reported that consumption of boiled
grass pea and green immature grass pea without cereals was linked to higher
incidence of neurolathyrism.
I.1.3.4. Breeding or genetic engineering to reduce β-ODAP in grass pea
It is rather puzzling that a commonly used crop such as grass pea has made so little
progress as a grain crop in its long cultivation period (Smartt, 1984). The level of β-
ODAP in low toxin varieties is undoubtedly strongly influenced by the environment.
The 40 years of efforts to reduce the crop’s toxicity has produced varieties with a
remarkable reduction in β-ODAP but neither stable nor toxin-free varieties could be
established yet. An Ethiopian-released variety ”Wasie”, claimed to have β-ODAP
concentrations below toxic threshold level (<0.04 %) turned out to be higher in toxin
(>0.2 %) after growing in lowland areas with relatively higher drought stress.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
19
Chromosome numbers of more than 60 species in the genus Lathyrus have been
reported, with only three species having more than 14 chromosomes (Narayan, 1998).
Successful inter-specific hybridisation in the genus has been shown to be extremely
rare. Following a report on the successful crossing of L. hirsutus x L. odoratus
(Barker, 1916), many researchers have attempted inter-specific hybridisation among
species in the genus Lathyrus. Inter-specific hybridisation involving L. sativus has
only been reported as successful with L. cicera (Lwin, 1956) and with L. amphicarpus
(only when the latter was used as female) (Khawaja, 1988). Cytological studies of F1
hybrids between L. amphicarpus x L. sativus, L. amphicarpus x L. cicera and L.
odoratus x L. chloranthus showed 50-70 % chromosome homology and pollen
fertility in conformity with meiotic pairing (Khawaja, 1988). To our knowledge, up
to now there has not been any line developed by crossing that is considered as having
a safe level of toxicity. Successful fertilization of certain cross-combinations
commonly showed embryo abortion during development. Therefore, embryo rescue
techniques need to be employed. This may imply that breeding strategies involving
genetic transfer for the improvement of grass pea should be developed. However,
sofar a high frequency transformation could not be achieved with L. sativus or with
the readily crossable Lathyrus species.
The floral biology of L. sativus is such that it favours self-pollination even if up to
27% outcrossing has been the concern of several breeders (Rahman et al, 1995;
Campbell, 1997). In most breeding programmes, crosses are done under controlled
conditions in the greenhouse or under insect-proof netting. Flowers are emasculated
by removing the anthers in the late bud stage, before flower opening. Male sterility is
uncommon in L. sativus. The morning after, styles are fertilized with pollen from a
pre-selected parental plant as soon as possible following dehiscence of the anthers.
The pollen at this time of the day is orange and rapidly becomes colourless as it loses
its viability. Although this manual pollination is a time-consuming process, it can
rapidly result in a large number of successful pollinations as 3-4 seeds develop from
each successful crossing.
No seed colour or flower colour has been confirmed to associate with the level of β-
ODAP. Also no other phenotypic character can be used as marker for low toxin. From
the variability of toxin levels in F2 populations of many studies, it is confirmed that β-

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
20
ODAP content is inherited quantitatively whereas the presence of cytoplasmic factors
suggested the parental effects on the β-ODAP score (Campbell, 1997). During the last
three decades, a number of low toxin varieties have been released in the world as a
result of conventional breeding. However, the toxin level in seeds of these lines could
vary due to the changes in the growing environment.
Grass pea has a limited secondary gene pool in related species. So far, grass pea only
crosses with L. cicera and L. amphicarpus and then produces a F1 generation to form
the secondary gene pool (Yunus and Jackson, 1991). Tertiary gene pool can be
exploited by crossing species using embryo rescue techniques or protoplast fusion
and/or biotechnological techniques to move genes between species (Smartt et al.,
1994; McCutchan et al., 1999; Durieu and Ochatt, 2000; Ochatt et al., 2001). After
the recent successful regeneration of complete plants from different parts of L.
sativus, somaclones were generated showing tremendous variability with respect to
leaf size, internodal length, flower and seed colour, seed weight, pod morphology and
β-ODAP score (Roy et al., 1991; Mehta and Santha, 1996; Santha et al., 1998; Abd El
Moneim et al., 1999; Zambre et al., 2002). So far, thousands of somaclonal lines
having a remarkable reduction of β-ODAP were developed in many laboratories,
although none were ever found to be toxin-free (Vaz Patto et al., 2006).
The advent of molecular marker techniques may facilitate breeding for improving
important agronomic traits including removal of unwanted metabolites or toxins.
Genetic engineering of the biosynthetic pathway for β–ODAP could be the most
direct method for producing a safe crop. Genetic transformation of L. sativus using
Agrobacterium tumefaciens, or A. rhizogenes, or biolistic techniques have been tried
repeatedly, but with very limited success (Vaz Patto et al., 2006) until the recent
report of an Agrobacterium-based transformation protocol (Barik et al., 2005).
Strategies to engineer the biosynthetic pathway (Kuo et al., 1998) and metabolic
breakdown (Datta, 1995; Metha and Santha, 1997) of β–ODAP have been proposed,
but the tools to achieve this have not yet been developed. Introgression of novel genes
for β-ODAP reduction and increased methionine or the use of antisense RNA
technology to silence or to block the biosynthetic pathway can create opportunities to
develop stable and safe varieties for consumers (Vaz Patto et al., 2006).

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
21
I.1.4. Effect of methionine on toxicity of grass pea
Even before the toxic compound was identified in grass pea, the low level of
methionine in urine of lathyrism patients having consumed grass pea had been
reported (Rudra and Chaudhury, 1952). More recently, Nunn and co-workers (2005)
rediscovered this phenomenon with human volunteers after consumption of grass pea
seed and suggested that a reduced flux of methionine to the central nervous system
(CNS) may sensitise motor neurons to the excito-toxic action of β-ODAP.
Maintaining the proportion of cereals (richer in sulphur amino acids) in the meal to at
least one third of a mixture with grass pea was identified as a protective factor for
neurolathyrism (Getahun et al., 2005).
Oxidative stress is thought to be important in the aetiology of neurolathyrism
development. Excitation of glutamate receptors in the CNS by β-ODAP produces
unstable nitric oxide (NO) that gives rise to free oxygen radicals (ROS) generation
that disrupt cell functions (Schinder et al., 1996). Several studies have clearly
indicated the protective effect of methionine against oxidative stresses caused by the
neurotoxicity of grass pea (Lambein et al., 2001; Nunn et al., 2005; Getahun et al.,
2005). However, the effective concentration level of methionine to sufficiently protect
against neurolathyrism has never been worked out or modelled, and this is one of the
study areas of the present thesis. It could be estimated that a total intake of about 200
g of β-ODAP present in grass pea consumed daily per person over a period of two
months can cripple over 50 % of males under the extreme hardships and malnutrition
of a Second World War camp prisoners in Vapniarca (Lambein et al., 2001), where
only males were present and males are known to be more susceptible to
neurolathyrism. Had it not been for the role of methionine and antioxidants present in
other food components such as teff (the traditional Ethiopian staple cereal) or
condiments used in sauce preparations, the incidence of neurolathyrism would have
been much higher than the dozen or so annual cases occurring during a normal year in
Ethiopia. This annual incidence can, however, get as high as up to hundreds or even
thousands during droughts and famine periods. When considering that the annual
production of β-ODAP in Ethiopia is about 400 tons (in 100,000 tons of grass pea
seed), this could in theory cripple more than a million people annually under extreme
conditions such as those experienced in the Vapniarca camp.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
22
The consumption of grass pea mixed with cereals richer in sulphur amino acids than
legumes was demonstrated to be highly protective from neurolathyrism (Getahun et
al., 2005). The antioxidant activity and enhancing of liver functioning by addition of
Zn and methionine in the molybdenotic rat (receiving sub-lethal doses of
molybdenum) has been documented (Suresh and Archana, 2000). Inhibition of the
production of reactive oxygen species (ROS) was proposed as mode of action in this
experimental system. It was also indicated that antagonists to excitatory amino acid
receptors may offer a therapeutic opportunity for some neurological diseases
(Ludolph et al., 1993).
Nunn and co-workers (2005) noticed that consumption of grass pea seed caused
persistent plasma methionine deficiency in human volunteers. The normal
physiological response to consumption of a methionine-deficient meal would be a
reduced plasma methionine concentration as found after consuming methionine-
limited grass pea and lentils, where around 50 % reduction of plasma methionine was
found after 24 hours. A reduction in the plasma methionine concentration would
cause a reduced methionine flux into the central nervous system as a consequence of
consuming grass pea. This may sensitise these cells to the neurotoxin β-ODAP
present in the grass pea seed. The latter authors concluded that supplementing such a
diet with methionine might protect those at risk from neurolathyrism.
I.1.5. Rationale of the study and motivation of the investigation
Compared to many field legumes, grass pea is a nutritionally rich source of food and
feed, containing high levels of protein, micronutrients and energy. However, its
protein is not well-balanced for it lacks sufficient sulphur-containing amino acids. The
remarkable tolerance to biotic and abiotic stresses, and high nitrogen fixation activity
should make grass pea a promising crop for economic use and for amelioration of
marginal soils. Especially considering global warming, the unique hardiness to grow
under adverse environmental conditions suggests a potential future for this crop in the
already drought-stressed and deteriorating highlands of Ethiopia. At present, in the
developing world over one hundred million poor and often illiterate farmers make

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
23
their living on grass pea cultivation in the face of the hitherto poorly perceived risk
crippling.
Global research on grass pea improvement, unlike with other crops, has been biased
towards taking out the inherent toxic element. Exhaustively undertaken conventional
breeding has been successful in downscaling ß-ODAP manyfolds from the original
0.5 to 1.2 % in seeds to as low as 0.01 %. Due probably to the complexity of the
challenge, and considering the misconceptions prevailing among farmers and the
poorly understood aetiology of the disease, these low-toxin lines had neither the
impact nor the extent of utilization expected. On the other hand, the bias on toxicity
research has masked the other important agronomic and qualitative parameters of the
crop that were not extensively researched or exploited.
Genetic research including contemporary techniques is very little used on grass pea
compared to other field crops. Already in 2004, FAO raised concerns about the need
for gene revolution for so-called ‘orphan crops’ including grass pea if we have to feed
an additional 2 billion people in the next three decades from increasingly fragile
landmasses.
Having potential as an important food source, the holistic approach of improving the
diet needs more attention. Other than toxicity reduction in the plant, the study of the
role of sulphur amino acids and attempts to increase the level of methionine in the
plant could be a novel approach to fundamentally address the problems of the crop. In
addition, assessing environmental influences on toxin concentration could show the
way for improving field management options.
Thus it becomes imperative to look into the importance of methionine as a
counteracting principle against the risk for neurolathyrism from grass pea
consumption. In vivo enrichment of sulphur-containing amino acids and studying
environmental factors affecting biosynthesis and biosynthate levels in grass pea is
anticipated to improve the nutritional quality and the economic value of the crop.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
24
I.1.6. Overall and specific objectives of the study
General objective
Improving the nutritional quality of grass pea for safer consumption and increased
economic benefit for the farmers.
Specific objectives
∎ Identification of gaps in our knowledge of grass pea and neurolathyrism by
reviewing related and relevant literature on the crop.
∎ Identification of climatic and edaphic factors affecting the level of ß-ODAP,
methionine and other amino acids in grass pea relevant to the risk for
neurolathyrism.
∎ To assess the protective effect of methionine on grass pea toxicity.
∎ To attempt increasing methionine in grass pea seed by gamma-irradiation
mutation and screening for methionine-rich mutant lines.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
25
CHAPTER II
Level of ββββ-ODAP, methionine and other free and protein
amino acids in the seeds of different grass pea (Lathyrus
sativus L.) genotypes

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
26
L. sativus seeds from different origin

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
27
CHAPTER II
Level of ββββ-ODAP, methionine and other free and protein amino acids
in the seeds of different grass pea (Lathyrus sativus L.) genotypes
[Fikre A., Korbu L., Kuo Y.-H. and Lambein F. 2008. Food Chemistry 110: 422-427]
Abstract
Free and protein amino acids of nine different genotypes of grass pea (Lathyrus
sativus L.) seeds were analysed by HPLC with precolumn PITC (phenyl
isothiocyanate) derivatisation. Among the free amino acids, homoarginine is
quantitatively the most important (up to 0.8 % of dry seed weight) and stable, while
the neuro-excitatory amino acid β-ODAP (β-N-oxalyl-L-α,β-diaminopropionic acid),
second most abundant, showed highest variation (0.02 % - 0.54 %) in the nine
genotypes examined. For the protein amino acids, glutamate was quantitatively most
significant followed by aspartate, arginine, leucine, lysine and proline. The sulphur
amino acid methionine showed the lowest concentration (~0.1 %) in all L. sativus
genotypes, and also in lentil (Lens culinaris) and in soybean (Glycine max) seeds
analysed at the same time.
II.1. Introduction
Grass pea (Lathyrus sativus L.) is perhaps one of the most environmental stress
resistant legume crop with high tolerance to drought, flooding and insect attacks
(Campbell, 1997). During drought and failure of other crops in Ethiopia, grass pea is
survival food number one for the poor. However, overconsumption of grass pea as a
staple during 2-3 months might cause increased incidence of the crippling disease
neurolathyrism reaching up to 6 % of the rural population. A non-protein amino acid
β-ODAP (β-N-oxalyl-L-α,β-diaminopropionic acid) found in grass pea seeds was
suggested to be the causal agent of this upper motorneurone disease. A recent report
showed that the mechanism of neuro-toxicity of β-ODAP leading to neurolathyrism is

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
28
very complex and oxidative stress might be involved in the pathogenesis (Lambein et
al., 2007).
The ß-ODAP content of grass pea varies widely, both among genotypes and
environments (Dahiya and Jeswani, 1975; Leakey, 1979; Ramanujam et al., 1980;
Campbell, 1997; DZARC, 2003). Based on the evaluation from three Australian
locations growing 407 lines of Lathyrus sativus and 96 lines of Lathyrus cicera
collected from three geographic origins namely Ethiopian, Mediterranean and
European, Hanbury and co-workers (2000a) summarised that for both species,
genotype was the most important determinant of ß-ODAP concentration while
environment had less influence. However, Wuletaw (2003) studied the stability of ß-
ODAP content in Lathyrus sativus and reported the importance of genotype with
environment interaction. The multi-annual multi-location trial by Debre Zeit
Agricultural Research Centre in Ethiopia with growing conditions going from high
altitude 2200 m above sea level (asl), vertisol, low temperature of mean 16 ± 2 oC to
low altitude of 1600 m asl, light soil and higher temperature of mean 24 ± 2 oC
(DZARC, 2003), and also the specific observation from Campbell (1997), indicated
that ß-ODAP level doubles or more for the same grass pea cultivars as the varying
growing environment slides from lower to higher stress conditions. A large variability
of ß-ODAP was seen due to environmental changes (Fikre et al., 2006). Both
environment and genotypes could play an important role in the biosynthesis level of
the toxin.
There appear to be clear differences not only for ß-ODAP content but also for other
antinutritional factors (ANFs) of grass pea. Urga et al. (2005) reported variability in
level of tannins, phytic acid and trypsin inhibitor activities of grass pea grown in
different agro-ecological locations of Ethiopia. Levels of total phenolics and
condensed tannins were reported to be positively correlated and they are determined
by genotypes rather than by environment in grass pea (Wang et al., 1998a). The same
authors also reported that trypsin inhibitor activities in grass pea did not differ among
cultivars or environments (Wang et al., 1998b).
The ß-ODAP content of grass pea seeds analysed by different labs is not always
consistent. The latter variation might be caused by the different analytical methods

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
29
used which include OPT (O-phthalaldehyde) colourimetric assay, automatic amino
acid analyser, HPLC (High Performance Liquid Chromatography), CZE (Capillary
Zone Electrophoresis), NIR (Near Infrared Reflectance Spectroscopy) and GC-MS
(Gas Chromatography-Mass Spectrometry). Some of these methods cannot
differentiate between α- and β-ODAP. However, α-ODAP is present naturally in
minor ratios (< 5%) of the total and was reported to be without neuroexcitatory effect.
For this study, we have collected grass pea seeds from different origins and analysed
their content of β-ODAP and other free amino acids by HPLC with precolumn PITC
(phenyl isothiocyanate) derivatisation as reported previously (Kuo et al., 2003).
The average protein content in legume seeds is 21-25 %. These proteins are rich in
most of the essential amino acids particularly lysine, but usually poor in the sulphur-
containing amino acids methionine and cysteine (Kuo et al., 1995). Monsoor and
Yusuf (2002) reported that the crude protein content in grass pea seeds (29.93 ± 1.26
%) is higher than that of lentil (23.23 ± 1.07 %) and chickpea (20.40 ± 2.27 %). The
objective of this study was to compare the contents of free amino acids, as well as the
protein amino acids among different grass pea genotypes, with special attention for
the neuro-excitatory β-ODAP and for the essential sulphur amino acid methionine.
II.2. Materials and methods
Plant materials: Seeds from nine genotypes of Lathyrus sativus originating from
different parts of the world including Ethiopia (local cultivars purchased from markets
at Debre Zeit; Gonder; Wollo and Bahr Dar), and collections of Institute of Plant
Biotechnology for Developing Countries (IPBO) from India (received from Dr R.L.
Pandey, Indira Gandhi Agricultural University, Raipur), China (black eye cultivar,
received from prof. Yu Jin-Zhong, Soil and Fertility Institute, Yangling, Shaanxii
province), Poland (Derek variety, obtained from prof. Marian Milczak, Lublin) and
Canada (LS 82046 and LS 87124, two low toxin lines bred by Dr. Campbell,
Agriculture Canada, Morden). Two other grain legume seeds, lentil (Lens culinaris)
and soybean (Glycine max) obtained from Turkey and Canada respectively through
commercial sources, were also analysed at the same time for comparison.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
30
Dry seeds were powdered in an electric coffee grinder and sieved. The seed powders
were used for further analysis.
Figure II.1. Variability in size, shape, texture and colour of L. sativus seeds from different origin
Preparation of seed extracts for free amino acids analysis: 200 mg of seed powder
was weighed and mixed with 10 ml of 70% ethanol. In each sample, 50 µl of DL-
allylglycine (100 µmol/ml, Sigma) was added as internal standard. The mixture was
allowed to stand at 4 °C overnight before centrifugation at 34,800 g for 20 min. The
supernatant was collected and the pellet was washed with 2 ml 70 % ethanol and
centrifuged as above. The pooled supernatant was concentrated to 0.5 ml extract with
a rotavapor under vacuum at 45 °C. An aliquot of 50 µl was used for derivatisation
with PITC for HPLC analysis.
Preparation of seed extract for protein amino acid analysis: 100 mg of seed
powder was weighed in a 2 ml glass vacule designed for hydrolysis (Wheaton, USA).
In each vacule, 2 ml of 6 N HCl, 0.01 % mercaptoethanol and 100 µl internal standard
norleucine (100 µmol/ml, Sigma) were added. The mixture was allowed to freeze by
placing the vacule in dry ice (carboglass) for 5-8 min. The vacule was then connected
to a vacuum system with gentle shake for about 10 minutes during the application of
the vacuum. The evacuated vacule was then sealed in a gas flame and placed in an
oven for hydrolysis at 110 °C for 18 hours.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
31
The hydrolysate was transferred to an Eppendorf and centrifuged in an Eppendorf
centrifuge (Hawksley MBC, England). The supernatant was transferred to a conical
evaporatory flask and dried with rotary evaporator under vacuum at 45 °C. This
procedure was repeated twice by adding distilled water after drying to remove the
HCl. Two ml distilled water was added to the flask to dissolve the dried samples and
the mixture was centrifuged. The supernatant was collected and 50 µl aliquot was
used for derivatisation with PITC for HPLC analysis.
PITC (Phenyl isothiocyanate) derivatisation: each 50 µl sample was first dried
under vacuum in an Eppendorf Vacufuge concentrator (5301) at 45 °C. To each dried
sample, 20 µl of coupling buffer (methanol/water/triethylamine: 2/2/1 by volume) was
added, and the whole was mixed and dried in Eppendorf Vacufuge concentrator.
Finally, 30 µl of the PITC reagent
(methanol/water/triethylamine/phenylisothiocyanate: 7/1/1/1 by volume) was added
and left to react at room temperature for 20 min before concentrating to dryness.
To each PITC-derivatised sample, 500 µl of buffer A (0.1 M ammonium acetate, pH
6.5) were added, mixed well and centrifuged. The supernatant was filtered through a
Millipore Millex filter (0.45 µm) and 20 µl aliquot was injected into the HPLC for
analysis. A standard amino acid mixture (AA-S-18, Sigma), L-(+)-homoarginine (99+
%, Janssen Chimica) and synthetic ß-ODAP (obtained from Dr. Rao S.L.N, India)
were also derivatised and prepared as above and injected into the HPLC as standard.
High performance liquid chromatography (HPLC) for amino acid analysis
A Waters 625 LC system with Waters 991 photodiode array detectors was used, as
reported previously (Kuo et al., 2003). A gradient system with buffer A (0.1 M
NH4OAc, pH 6.5) and buffer B (0.1 M NH4OAc, containing acetonitrile and MeOH;
44/46/10 by volume, pH 6.5) with flow rate of 1 ml/min was used for the separation
of amino acids during 50 min. An Alltima C18 column (250 x 4.6 mm I.D., 5 µm
particle size, Alltech, USA) was used with column temperature at 43 °C during
analysis. Absorbance at 254 nm was used for calculations. Results were analysed by
Millennium software (Waters, version 1.10).

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
32
Statistics
All experiments were repeated twice and each sample was injected twice into the
HPLC. Results were expressed as mean ± SD. Data were analysed with ANOVA
using SPSS 12 software. Statistical significance of differences in mean values was
calculated with a confidence level of 95 %.
II.3. Results
Free amino acids profile
The grass pea seeds from different origins showed high variability in morphology,
especially size, shape and colour (Figure II. 1). In this study, we found that the non-
protein amino acid homoarginine (Har) was the most abundant amino acid in all the
genotypes of L. sativus seeds analysed (Table II.1), confirming previous results
obtained from one genotype (Lambein et al., 1992). In addition, this high
concentration (0.68-0.86 %) is rather stable among all genotypes analysed with no
statistical difference. This high level is a unique feature specific for grass pea seeds as
compared to other food legumes (Rozan et al., 2001; Kuo et al., 2004).
Table II.1: Free amino acids (% dry seed weight; n = 4; mean ± SD) in Lathyrus sativus seeds of different origin
Values in a row followed by the same letters are not significantly different (p< 0.05); Har-homoarginine
β-ODAP is quantitatively the second most important free amino acid in L. sativus
seeds but with greatest variation from 0.2 mg/g dry seed in LS-82046 (Canada) to 5.4
mg/g dry seed in Raipur (India). Earlier studies on the influence of nutrient supply on
β-ODAP content in grass pea seeds growing in hydroponic media indicated that
reduction of macro-nutrients such as NO3-, Mg2+, or K+ in Hoagland solution resulted
in a sharp increase of β-ODAP content of the ripe seeds (Kebede et al., 1994).
D.Zeit
(Shoa), Eth
Gonder,
Eth Wollo, Eth B.Dar, Eth India China Poland Ls82046 Ls 87124
Har 0.68±0.11a 0.78±0.16a 0.69±0.09a 0.86±0.18a 0.79±0.05a 0.68±0.20a 0.80±0.32a 0.74±0.10a 0.69±0.15a
ß-ODAP 0.35±0.11ab 0.28±0.09bc 0.35±0.07ab 0.48±0.18ab 0.54±0.06a 0.38±0.13ab 0.18±0.09c 0.02±0.01d 0.02±0.01d
Aspargine 0.04±0.01b 0.08±0.04b 0.06±0.10b 0.05±0.04b 0.03±0.02b 0.07±0.03b 0.15±0.06a 0.06±0.05b 0.03±0.01b
Arginine 0.05±0.07ab 0.03±0.02bc 0.05±0.01ab 0.03±0.03bc 0.06±0.04a 0.04±0.01bc 0.03±0.01bc 0.03±0.01bc 0.01±0.00c
Aspartate 0.02±0.02b 0.03±0.03ab 0.04±0.03a 0.03±0.03ab 0.04±0.04a 0.04±0.02a 0.04±0.01a 0.02±0.01b 0.01±0.00c
Glutamate 0.03±0.03b 0.08±0.06a 0.07±0.04a 0.04±0.03b 0.03±0.03b 0.04±0.02b 0.08±0.02a 0.04±0.01b 0.04±0.02b
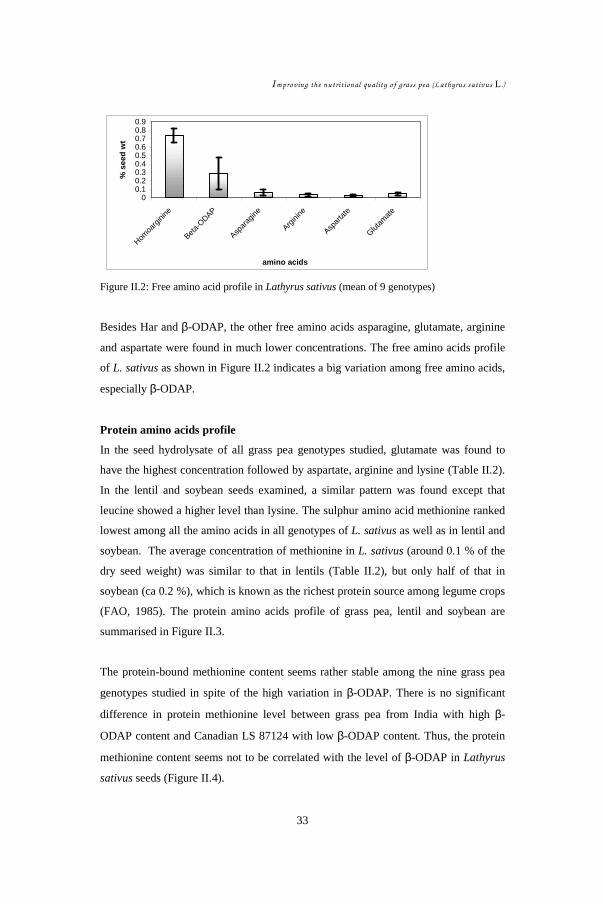
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
33
00.10.20.30.40.50.60.70.80.9
Homoa
rgini
ne
Beta-
ODAP
Aspar
agine
Argini
ne
Aspar
tate
Glutam
ate
amino acids
% s
eed
wt
Figure II.2: Free amino acid profile in Lathyrus sativus (mean of 9 genotypes)
Besides Har and β-ODAP, the other free amino acids asparagine, glutamate, arginine
and aspartate were found in much lower concentrations. The free amino acids profile
of L. sativus as shown in Figure II.2 indicates a big variation among free amino acids,
especially β-ODAP.
Protein amino acids profile
In the seed hydrolysate of all grass pea genotypes studied, glutamate was found to
have the highest concentration followed by aspartate, arginine and lysine (Table II.2).
In the lentil and soybean seeds examined, a similar pattern was found except that
leucine showed a higher level than lysine. The sulphur amino acid methionine ranked
lowest among all the amino acids in all genotypes of L. sativus as well as in lentil and
soybean. The average concentration of methionine in L. sativus (around 0.1 % of the
dry seed weight) was similar to that in lentils (Table II.2), but only half of that in
soybean (ca 0.2 %), which is known as the richest protein source among legume crops
(FAO, 1985). The protein amino acids profile of grass pea, lentil and soybean are
summarised in Figure II.3.
The protein-bound methionine content seems rather stable among the nine grass pea
genotypes studied in spite of the high variation in β-ODAP. There is no significant
difference in protein methionine level between grass pea from India with high β-
ODAP content and Canadian LS 87124 with low β-ODAP content. Thus, the protein
methionine content seems not to be correlated with the level of β-ODAP in Lathyrus
sativus seeds (Figure II.4).
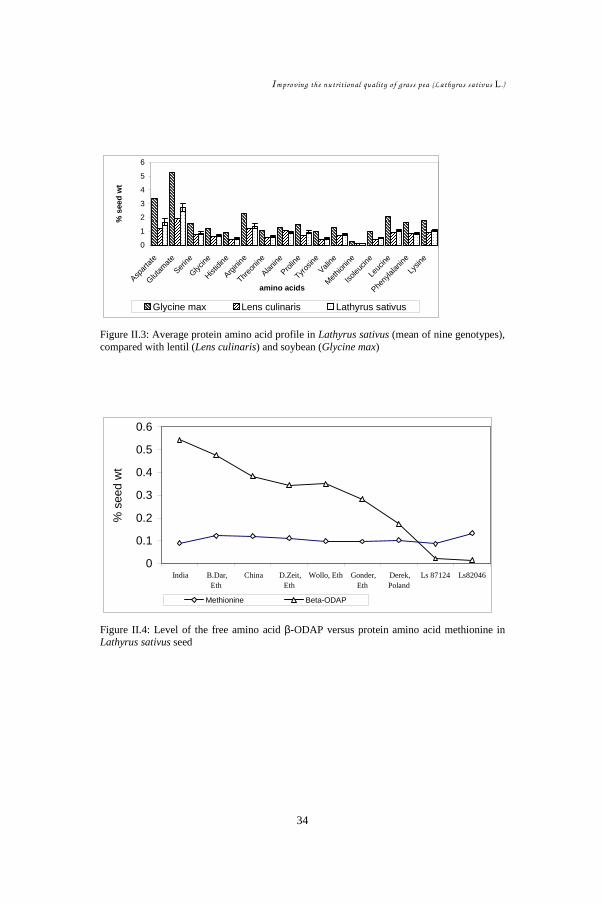
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
34
0
1
2
3
4
5
6
Aspar
tate
Glutam
ate
Serine
Glycine
Histidi
ne
Argini
ne
Threo
nine
Alanine
Prolin
e
Tyros
ine
Valine
Met
hionin
e
Isoleu
cine
Leuc
ine
Pheny
lalan
ine
Lysin
e
amino acids
% s
eed
wt
Glycine max Lens culinaris Lathyrus sativus
Figure II.3: Average protein amino acid profile in Lathyrus sativus (mean of nine genotypes), compared with lentil (Lens culinaris) and soybean (Glycine max)
0
0.1
0.2
0.3
0.4
0.5
0.6
India B.Dar,Eth
China D.Zeit,Eth
Wollo, Eth Gonder,Eth
Derek,Poland
Ls 87124 Ls82046
% s
eed
wt
Methionine Beta-ODAP
Figure II.4: Level of the free amino acid β-ODAP versus protein amino acid methionine in Lathyrus sativus seed

Impr
ovin
g th
e nu
trit
iona
l qu
alit
y of
gra
ss p
ea (
Lat
hyru
s sa
tivu
s L
.)
35
Tab
le II
.2:
Pro
tein
am
ino
aci
ds (
% d
ry s
eed
wei
ght;
mea
n ±
SD
; n
= 4
) in
th
e se
eds
of La
thyr
us s
ativ
us o
f diff
eren
t
ge
noty
pes
com
par
ed t
o t
wo
oth
er fo
od
legu
mes
i.e.
lent
il an
d so
ybe
an
Val
ues
in a
row
follo
wed
by
the
sam
e le
tter
are
not
sig
nifi
cant
ly d
iffe
rent
(p<
0.05
)
D
.Ze
it (S
hoa
), E
th
Go
nder
, E
th
Wo
llo,
Eth
B
.Dar
, Eth
In
dia
C
hin
a P
ola
nd
Ls8
20
46
Ls 8
712
4
Len
s cu
lina
ris
Gly
cine
max
Asp
art
ate
1
.40
±0.5
3e 1
.98
±0.5
7b 1
.50
±0.4
5de
1.7
5±0
.63cd
1
.92
±0.5
7bc
1.6
3±0
.55d
1.9
5±0
.65bc
1
.54
±0.3
8de
1.3
9±0
.46e
1.1
8±0
.45ef
3
.36
±1.3
0a
Glu
tam
ate
2
.3±0
.91ef
3.0
7±1
.10bc
2
.43
±0.8
3de
2.9
0±1
.19d
3.0
2±1
.13cd
2
.67
±0.9
3ef
2.9
8±1
.04cd
2
.64
±0.9
2e 2
.48
±0.9
5e 1
.94
±0.7
6f 5
.26
±2.2
9a
Ser
ine
0
.85
±0.3
4cd
0.9
1±0
.04cd
0
.75
±0.0
3e 0
.95
±0.2
4b 0
.94
±0.1
1bc
0.8
8±0
.05cd
0
.93
±0.0
4bc
0.8
6±0
.07cd
0
.63
±0.2
1f 0
.76
±0.0
9de
1.5
6±0
.51a
Gly
cine
0
.69
±0.3
1cd
0.7
1±0
.08bc
0
.62
±0.1
0cd
0.7
6±0
.17b
0.7
2±0
.03bc
0
.74
±0.0
8b 0
.79
±0.0
4b 0
.72
±0.0
3bc
0.5
2±0
.18e
0.6
5±0
.02cd
1
.21
±0.3
8a
His
tidin
e
0.4
5±0
.22cd
0
.50
±0.1
7bc
0.4
1±0
.18e
0.5
1±0
.18bc
0
.51
±0.1
2bc
0.4
9±0
.21cd
0
.52
±0.1
7b 0
.50
±0.1
3c 0
.43
±0.0
8cd
0.4
3±0
.09cd
0
.89
±0.2
7a
Arg
inin
e 1
.32
±0.4
5cd
1.4
7±0
.33bc
1
.21
±0.2
7cd
1.5
1±0
.58bc
1
.56
±0.4
3b 1
.43
±0.2
8c 1
.55
±0.3
6b 1
.41
±0.3
3bc
1.0
4±0
.30d
1.1
9±0
.29d
2.2
6±1
.00a
Thr
eon
ine
0.5
2±0
.23cd
0.6
2±0
.14bc
0
.52
±0.1
3cd
0.6
8±0
.20b
0.6
1±0
.11bc
0
.61
±0.2
1bc
0.6
6±0
.14b
0.5
9±0
.08c
0.4
8±0
.10d
0.5
6±0
.03c
1.0
6±0
.49a
Ala
nin
e
1.0
3±0
.40b
0.8
1±0
.04d
c 0
.83
±0.1
9d 0
.99
±0.4
3cd
0.8
1±0
.14d
e 1
.02
±0.2
8bc
0.9
7±0
.16cd
1
.01
±0.1
4c 0
.76
±0.1
8f 1
.04
±0.2
7b 1
.31
±0.6
2a
Pro
line
0
.90
±0.3
2cde
0.9
3±0
.16cd
0
.80
±0.0
7e 1
.01
±0.0
7bc
1.0
4±0
.27b
0.9
4±0
.21bc
d 1
.06
±0.2
4b 0
.95
±0.2
4c 0
.83
±0.2
6d 0
.67
±0.0
4e 1
.53
±0.7
8a
Tyr
osi
ne
0.4
4±0
.22cd
0.4
9±0
.17c
0.4
3±0
.16d
0.5
3±0
.16b
0.5
1±0
.10bc
0
.49
±0.2
0c 0
.51
±0.1
5bc
0.4
8±0
.12c
0.4
2±0
.08d
0.4
1±0
.08d
0.9
6±0
.55a
Va
line
0.7
3±0
.30cd
0
.79
±0.0
8c 0
.70
±0.1
2e 0
.84
±0.3
7b 0
.78
±0.0c
0.8
1±0
.06bc
0
.81
±0.0
4bc
0.8
1±0
.06bc
0
.66
±0.0
8e 0
.72
±0.0
9d 1
.30
±0.7
7a
Met
hio
nine
0
.11
±0.0
9c 0
.10
±0.0
3d 0
.10
±0.0
3d 0
.12
±0.0
4bc
0.0
9±0
.02e
0.1
2±0
.07bc
0
.10
±0.0
5d 0
.13
±0.0
3b 0
.09
±0.0
2e 0
.10
±0.0
2d 0
.24
±0.0
9a
Iso
leuc
ine
0.4
8±0
.16cd
0.5
8±0
.38b
0.4
9±0
.33cd
0
.57
±0.4
2bc
0.5
5±0
.35bc
0
.53
±0.3
8c 0
.56
±0.3
6bc
0.5
2±0
.34cd
0
.44
±0.2
8d 0
.44
±0.2
9d 0
.99
±0.6
7a
Leuc
ine
0
.96
±0.5
5d 1
.17
±0.3
6b 0
.97
±0.2
9d 1
.15
±0.3
1bc
1.1
5±0
.38bc
1
.07
±0.1
9cd
1.1
3±0
.32c
1.0
5±0
.32cd
0
.92
±0.3
0e 0
.91
±0.3
3e 2
.08
±0.6
1a
Phe
nyl
ala
nine
0
.82
±0.3
2cd
0
.91
±0.2
8b 0
.78
±0.2
6e 0
.91
±0.2
6b 0
.91
±0.3
1b 0
.82
±0.1
9cd
0.8
8±0
.23c
0.8
8±0
.17c
0.7
8±0
.17e
0.8
3±0
.21d
1.6
6±0
.45a
Lysi
ne
0.9
1±0
.59d
1.1
3±0
.22b
0.9
4±0
.23cd
1
.13
±0.2
3b 1
.10
±0.2
5bc
1.1
0±0
.19bc
1
.21
±0.0
2b 1
.01
±0.1
1cd
0.9
6±0
.12cd
0
.88
±0.1
6d 1
.81
±0.1
4ab
To
tal
14.8
8
16.1
7
12.9
8
16.2
9
16.2
2
15.3
5
16.6
1
15.
1
11.8
3
12.7
1
25.9
7

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
36
II.4. Discussion
Our analysis of free amino acids in the different genotypes demonstrated that
homoarginine (Har) was both the most abundant and stable free amino acid in grass pea.
Har has also been found in dry seeds and 4-day-old seedlings of several Lens species but
the amount was very low and negligible (Rozan et al., 2001). The study of the free amino
acids profile of some other common food legumes including pea, bean and lentil (Kuo et
al., 2004) also showed that Har was undetectable in the dry seeds while arginine was
quantitatively the most important free amino acid in pea and bean, and glutamic acid was
the most important amino acid in lentil. Haque (1997) reported that for the low toxin line
of L. sativus LS-82046, when grown in hydroponic solutions with a doubled supply of
trace elements Zn2+, Fe2+, B(OH)4- or Co2+, or with Al3+ at 2 x 10–6 M in the media, or
with increasing salinity up to 0.8 % NaCl (v/w), the level of Har in the seeds increased
dramatically.
Both Har and β-ODAP were detected in the blood plasma and in the urine after human
volunteers ingested a single meal of 200 g cooked grass pea seeds (Nunn et al., 1994).
Har has been reported to antagonise the neurotoxic action of β-ODAP fed to one-day-old
chicks (Yusuf et al., 1995). This might be explained by the modulating effect of L-Har on
nitric oxide synthase (Jyothi and Rao, 1999).
Jiao and co-workers (2006) reported that β-ODAP content in grass pea might be related
to the level of total free nitrogenous compounds whereas nitrogen and phosphate may be
the crucial nutrient factors influencing β-ODAP content under field conditions.
Environmental factors such as drought, zinc deficiency, iron oversupply and the presence
of heavy metals in the soil can considerably increase the level of β-ODAP in the seeds
(Haque, 1997).
The presence of β-ODAP in grass pea seeds was blamed for causing the crippling disease
neurolathyrism and the toxicity is possibly mediated by collective effects of L-β-ODAP
on the AMPA-type receptor, metabotropic glutamate receptors, and NO (nitric oxide)

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
37
production (Kusama-Eguchi et al., 2006). β-ODAP was found to chelate copper and
zinc, which might interfere with the redox homeostasis in the body after heavy
consumption (Davies et al., 1990). However, this multifunctional compound, named as
dencichine in traditional Chinese herbal medicine, was also detected in the longevity
promoting ginseng root (Kuo et al., 2003), and praised for its haemostatic and platelet
increasing properties (Xie et al., 2007).
Efforts in plant breeding research to eliminate β-ODAP from the seeds have produced a
large number of “low toxin” varieties but did not yet result in “toxin-free” varieties.
Post-harvest processing of grass pea seeds by solid state fermentation with Aspergillus
oryzae and Rhizopus microsporus dramatically reduced the level of β-ODAP to a
minimum amount of around 0.1 g kg–1 but could not remove the neurotoxin completely
even with the low toxin lines (Kuo et al., 1995; Kuo et al., 2000).
Grass pea seeds are the major protein source for the poor and become a survival food
during drought or when land productivity becomes marginal in rural areas. Gatta et al.
(2002) reported that the protein content of 161 accessions in the Bari (Bangladesh
Agricultural Research Institute) Lathyrus germplasm collection ranged from 23 to 29.9 %
(mean 26.3 %) with low coefficient of variation (4 %). On the other hand, the protein-
bound methionine content seems to be the least abundant amino acid as reported earlier
(Kuo et al., 1995) but stable among nine grass pea genotypes in spite of high variability
in β-ODAP. Moreover, Nunn and co-workers (2005) reported that consumption of these
seeds limiting in methionine (Kuo et al., 1995) as staple food for a long period leads to
the deprivation of methionine in the bloodstream. The long-term deficiency of
methionine in the bloodstream and thus reduced methionine flux into motor neuron cells
might lead to higher susceptibility of these cells to the excitatory effects of β-ODAP
(Nunn et al., 2005). The concentration of methionine (0.1 %) in the naturally growing
grass pea accounts for only about a fourth of the normal dietary requirement for an
average person and less than a fourth for children (FAO/WHO, 1985). Neurolathyrism is
believed to be caused by the neuro-excitotoxic action of β-ODAP and oxidative stress.
The deficiency of methionine contributes to this oxidative stress. Therefore, for the

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
38
nutritional improvement of Lathyrus sativus and the prevention of neurolathyrism there is
a crucial need for additional sulphur amino acid supplement in grass pea food or for in
vivo enrichment in the plant itself.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
39
CHAPTER III
Study of environmental factors (climatic, edaphic) affecting
yield and toxicity of grass pea grown in Ethiopia in a
multilocation experiment

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
40
L. sativus growing on Ethiopian soil

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
41
CHAPTER III
Study of environmental factors (climatic, edaphic) affecting yield and
toxicity of grass pea grown in Ethiopia in a multilocation experiment
Abstract
Statistical analysis (path coefficient, ANOVA, correlation) was done to identify the
interrelations and the effects of environmental factors (climatic, edaphic, altitudinal) on
yield and β-ODAP level of grass pea grown in Ethiopia. Ten grass pea genotypes grown
for two years (2005- 2006) in five different agro-ecological locations of Ethiopia were
used. Environmental factors including rainfall, sunshine hours, temperature, soil nutrients
and physical soil factors linked to altitudes of the respective locations were studied in
relation to their interrelations or correlations with grass pea yield and β-ODAP level.
Analysis of varience indicated that crop yield and yield related parameters including β-
ODAP showed considerable variability among locations (altitudes), years and genotypes.
Path coefficient analysis indicated that days to maturity was the most influencing factor
having the largest and most positive direct effect on yield. Total rainfall during the
growth period had a large positive direct effect on yield. High levels of soil nutrients
Mn2+ and S2- seem to affect yield negatively at Denbi site. Path analysis for β-ODAP
revealed that Zn2+/P, days to maturity, yield and K+ were variables having a larger direct
effect on the response variable β-ODAP. The dominant variables with high total effect (r
> 0.75) on β-ODAP under field conditions were K+, sunshine hours (ssh) and days to
maturity. We also found that supply of Zn2+ can be optimised to minimise β-ODAP
content, but this is also affected by interactions with other essential soil nutrients. From
linear correlation analysis, we found that K+ and ssh positively, while pH, days to
maturity and yield negatively have the highest negative correlations (>0.70) with β-
ODAP levels among 35 factors considered in this study for Ethiopia. The analysis also
indicated the strongest correlation of sunshine hours with β-ODAP level during the phase
of crop maturity (ssh III). This may suggest that β-ODAP synthesis and translocation to
the final sink (seed) is mainly taking place during crop maturation or late reproductive

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
42
phase. From this we can propose that β-ODAP biosynthesis and its response to
environmental stress could be highest during postanthesis.
III.1. Introduction
Climatic and edaphic factors were found to affect the level of a number of biosynthates
and crop parameters in Lathyrus sativus. Significant differences were reported for crop
performances (plant size, days to maturity, days to flowering, total biomass and grain
production), β-ODAP and other antinutritional constituents of grass pea when grown in
different agro-ecologies. Many studies confirmed that both genotypic and environmental
factors, and their interaction affect the concentration of β-ODAP in grass pea (Campbell,
1997; Hanbury et al., 2000a; Cocks et al., 2000; Wuletaw, 2003; DZARC, 2003; Asfaw
et al., 2003).
Haque (1997) proposed that higher accumulation of β-ODAP in drought-stressed L.
sativus plants and seeds may result from disrupted nitrogen assimilation and loss of
specificity of particular nitrogen assimilating enzymes, leading to higher synthesis of
asparagine. Asparagine is considered the primary precursor of the isoxazolinone ring in
BIA, which was confirmed to be the in vivo and in vitro precursor of β-ODAP in L.
sativus. It was suggested that β-ODAP might be important for the plant by acting as a
defence to various stresses (Lambein et al., 1990; Haque, 1997). This β-ODAP may then
contribute to the hardiness of the crop. Hence, the plant apparently produces less toxin in
the seed under optimal growing conditions than when it grows under marginal conditions.
Under favourable growing conditions, the plant produces more pods and seeds with
reduced β-ODAP concentrations, possibly as a result of a toxin dilution effect. Under
identical unstressed conditions, plants with only 10 pods contained 59 % more β-ODAP
in the seed than plants with 100 pods (Cocks et al., 2000).
From hydroponic pot experiments, Haque (1997) reported that soil nutrients Fe2+, Mn2+,
B3+, Co2+ and high Al3+ were found to enhance accumulation of β-ODAP while Zn2+,

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
43
Mo6+, Cu2+ and low Al3+ decreased the level. The antagonistic effect of Fe2+ and P against
Zn2+availability seem to increase the level of β-ODAP in a manner that higher P/Zn2+ and
Fe2+/Zn2+ ratios would boost toxin production. According to Jiao et al. (2006) β-ODAP
accumulation in grass pea might be related to the level of total free nitrogenous
compounds, whereas nitrogen and phosphate may be the crucial nutrient factors
influencing β-ODAP synthesis and accumulation under field conditions. Grass pea is
reported to be tolerant to a certain level of soil salinity. Occurrence of water soluble salts
refers principally to the precence of four cations Na+, K+, Ca2+, and Mg2+ linked with the
anions Cl- and SO42- (Jackson, 1973). In artificial salinity in sandy soil, grass pea biomass
production increases up to 0.3 % NaCl (EC = 7.8 m.mohs/cm). However, this remains
controversial as salinity has a decreasing effect on β-ODAP accumulation at lower levels
of artificial salinity (up to 0.6% NaCl) and an increasing effect at higher levels of
artificial salinity (Haque, 1997).
Light also influences β-ODAP biosynthesis. Zhang and co-workers (2003) discovered
that β-ODAP can act as a scavenger for hydroxyl radicals in the plant and protects
glycolate oxidase (GO, an enzyme of photorespiration) under high light intensity. In the
Ethiopian highlands high light intensity can obuviously play a role. β-ODAP content
increases while GO activity decreases. The same authors (Zhang et al., 2003) also found
that oxalate, a precursor molecule for β-ODAP, increases β-ODAP content in the seed
when applied in the seedling stage.
Previous studies on grass pea mostly emphasized on genotype improvement with focus
on reducing neurotoxin content, while studies on environmental factors influencing this
metabolite synthesis and subsquent accumulation were hardly undertaken. As a rationale,
understanding the relative importance of environmental factors and their interrelation in
affecting β-ODAP content and crop yield would help developing possible crop
mangement measures aimed at crop improvement and maximizing farmer’s benefit.
Although experiments have previously been carried out in diverse environments in
relation to grass pea toxicity (particularly β-ODAP), those experiments failed to
adequately and inclusively analyse associated factors (climatic, edaphic) influencing seed

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
44
β-ODAP content or crop yield. Hence, the present study was designed to investigate the
effects of various growth factors on grass pea production and its toxicity from different
experimental plots operated by the Ethiopian Institute for Agricultural Research (EIAR).
Environmental and geographic parameters (climatic, edaphic, altitudinal, seasonal) were
measured and monitored. A statistical method called ‘Path Coefficient Analysis’ that was
developed for calculating such cause/effect or predictor/responsive variable relationships
(Gravois and McNew, 1993) was employed. Hence, the study was initiated to identify
correlative effects of environmental (climatic, and edaphic) and genotypic factors and to
determine their level of importance on yield and β-ODAP content of grass pea in
Ethiopia.
III.2. Materials and methods
Field experiments were conducted at five experimental stations (Fig III.1.) in Ethiopia
during the 2005-2006 seasons to measure seed yield and β-ODAP level in 10 genotypes
of grass pea, meteorological and edaphic data. Agronomic parameters and meteorological
data are from the two years (2005 and 2006), however, due to resource limitations, soil
nutrient and amino acid analysis data are presented from the 2005 season only and
analysis was done accordingly. Nine grass pea genotypes that were selected for their wide
environmental adaptation and low toxicity content by International Centre for
Agricultural Research in the Dry Areas (ICARDA), Allepo, Syria were used. These
include ILAT-Ls-590, ILAT-Ls-610, ILAT-Ls-690, ILAT-Ls-Ls-B1, ILAT-Ls-Ls-B2
(newly released variety in Ethiopia named Wasie), ILAT-Ls-k290, ILAT-Ls-k387, ILAT-
Ls-k190, ILAT-Ls-390. Furthermore, cultivar Debre Zeit was included as local control.
ICARDA is an international center that has the mandate for grass pea improvement
research. The experiment was arranged in a randomized complete block design with three
replications. Plot size was 1.6 m x 4.0 m or 6.4 m2. Spacing was 10 cm between plants
and 40 cm between rows. No fertilizer was applied. Plots were weeded as needed. Field
data: sowing date, plant emergence, pest infestation monitoring, days to flowering date,
days to maturity and seed yield were monitored and recorded. Harvesting was done from
the whole plot as plants grow intermingled and separation is very difficult.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
45
Figure. III.1. Growing locations used for analysis in East Shoa Zone in the central high lands of Ethiopia (1*-Alem Tena, 2*-Debre Zeit, 3*-Denbi, 4*-Akaki, 5*-Chefe Donsa. Major roads (yellow) with water bodies (blue patches ) indicated. (See Table III.1 for coordinates). Source: google earth mapping Agro-ecological analysis
Locations altitude were determined using a pocket altimeter (Bariso, Finland) adjusted in
reference points and are given in meters above sea level (m asl). During the growth
period of the crop, the daily measurements of maximum and minimum temperature (oC)
by thermometer (CASSESLA, England), rainfall (mm) by rain gauge (Lamp Cht,
Germany) and daily sunshine hours (the time during which the direct solar radiation
5* 4* 3* 2* 1*

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
46
exceeds the level of 120 W/m2) by radiometer (SIAP, Italy) were taken from the
respective locations’ weather stations.
At different stages of development, the plant may have different sensitivity to stress
(Cocks et al., 2000), with different responses to β-ODAP synthesis levels. Hence, three
growth stages were differentiated and the climatic factor values were analyzed separately
based on these three growth stages of the plant. The growth stages considered were i) pre-
branching or seedling stage: the stage between sowing and branching (first 3-5 weeks); ii)
pre-anthesis (next 3-7 weeks): between branching and anthesis (maximum biomass
accumulation); and iii) post-anthesis (last 5-9 weeks), the period after flowers are fully
open (functional) until crop maturity. The climatic parameters (temperature, rainfall and
sunshine hours) recorded during the respective stages were labeled as tempI, tempII, and
tempIII, and RFI, RFII, RFIII, and sshI, sshII, sshIII.
Soil nutrient and amino acid analysis (2005 season)
Before planting, soil samples were taken using an Auger manual drill in the respective
growth locations (six samples/location) from the surface down to 60 cm depth. Analysis
of soil parameters was done in the national soil laboratory, Ethiopia. Soil micronutrients
(Zn2+, Mn2+, Fe2+, Cu2+) were extracted using the chelating agent Diethylene Triamine
Pentaacetic Acid (DTPA) after which their amount was determined using an atomic
absorption spectrophotometer (Milton Roy Spectronic model No 401, USA) together with
reference standards for each microelement. Published analytical procedures were
followed (Houba et al., 1989) and the content of microelements (Fe2+/Mn2+/Zn2+/Cu2+)
was calculated as:
(mg/kg soil) = (a-b) x 40/S x mcf;
where:
a = conc. of element in sample extract (mg/l)
b = conc. of element in the blank (mg/l)
40 = volume of extractant used per sample (ml)
S = weight of air dry soil (g)
mcf = moisture correction factor.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
47
Sulphur concentration was determined following the procedure described by Cottenie
(1980). Available soil phosphorus content was determined by the 0.5 M sodium
bicarbonate extraction solution (pH 8.5) method of Olsen (1954). C:N ratio was
calculated from the separate determinations of organic carbon (OC) by weight loss on
ignition in a combustion furnace and total nitrogen (TN) using the Kjeldahl method. Soil
pH meter reading was made after suspension in water using calibrations at pH 4.00, 7.00
and 9.00.
Concentration of ß-N-oxalyl-L-α,ß-diaminopropionic acid (ß-ODAP), the neuro-
excitatory free amino acid in the plant known to cause irreversible crippling in humans,
was determined only for the 2005 harvested seed samples using O-phthalaldehyde (OPT)
color reaction as standardized by Rao (1978) at the reference laboratory in ICARDA by
Dr. Abd El Moniem. Protein hydrolysis for amino acids determination using HPLC was
performed as explained in Chapter II. All samples were hydrolysed in duplicate and each
derivatised hydrolysate was analysed twice by HPLC, giving four numbers per data point.
Agronomic parameter analysis
Crop response parameters considered were: days to maturity, 100 seeds weight (g) and
dry seed yield in kg/ha. Days to maturity were determined as the number of days from
seed germination to maturity (>95 pods changed yellow or pale) of the plants in the plot.
Hundred seed weight is the weight in g of 100 seeds taken randomly from the respective
harvested seed lots. Yield in kg/ha was estimated by extrapolation of the yield of all 160
plants of the plot.
Statistics
Analysis of variance for the means was done using statistical analysis software (SAS
Inst., Cary NC) (Kang, 1994). Determination of path coefficients for the path analysis
was done in a multiple step calculation using SPSS. Predictor variables for yield [days to
maturity, copper (Cu2+ in ppm), zinc (Zn2+ in ppm), total rain fall during stage I (RFI)]
and for β-ODAP [soil pH, available K+ (ppm), sshIII, days to maturity, yield (kg/ha), Fe2+

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
48
(ppm), Zn2+/Fe2+, Zn2+/P, soil bulk density (BD/cm3), tempII and RFII] were selected
using stepwise regression after they were found to have significant correlation with
respective dependent variable (yield or β-ODAP). Path coefficient analysis was selected
as the method of choice because it not only shows relationships between predictor and
response variables (direct effect of specific component), but also shows interrelationships
between predictor variables in their effects on the response variable (indirect effect of
specific component on the response variable via other components) (Gravoice and
McNew, 1993). By measuring the extent of effect of one variable on another using
standardized data or a correlation matrix as input, the model, path coefficient, enables
identification of environmental components affecting the response variable (in this case:
grass pea seed yield and β-ODAP level) and to estimate the strength of this effect.
III.3. Results As can be seen in Table III.1 and Figure III. 2, the locations have different altitudinal,
climatic and edaphic features. These differences were correspondingly reflected on the
growth period of grass pea (Table III.2 and Figure III.3) resulting in differences in days
to maturity and ultimate yield among the locations (Tables III.2 and III.4). According to
Bull’s (1996) agro-ecological classification based on length of growth period (L) and
thermal zone (T) as related to altitude, we identified two thermal and two growth length
groups [LI=<90 days, LII = 91-150 days, T3 = 1300-2000 m asl, T4 = 2000-3000 m asl]
in the five locations. This classification was adopted since it is found having important
relevance to the present study. Hence the locations can be grouped into ‘short growth
period LI’ as in Alem Tena and Denbi, and ‘long growth period LII’ as in Debre Zeit,
Akaki and Chefe Donsa (see Table III.1). This effect of the location on length of growth
period (days to maturity) is also found to vary associated with altitude, moisture regime,
temperature range and soil properties (Tables III.3 and III.4 and Figure III.2). In addition
to the light soil texture that may allow fast evapotranspiration, the levels of S2- and Mn2+
were found remarkably high at the poor yielding location of Denbi (Tables III.2 and III.
3).
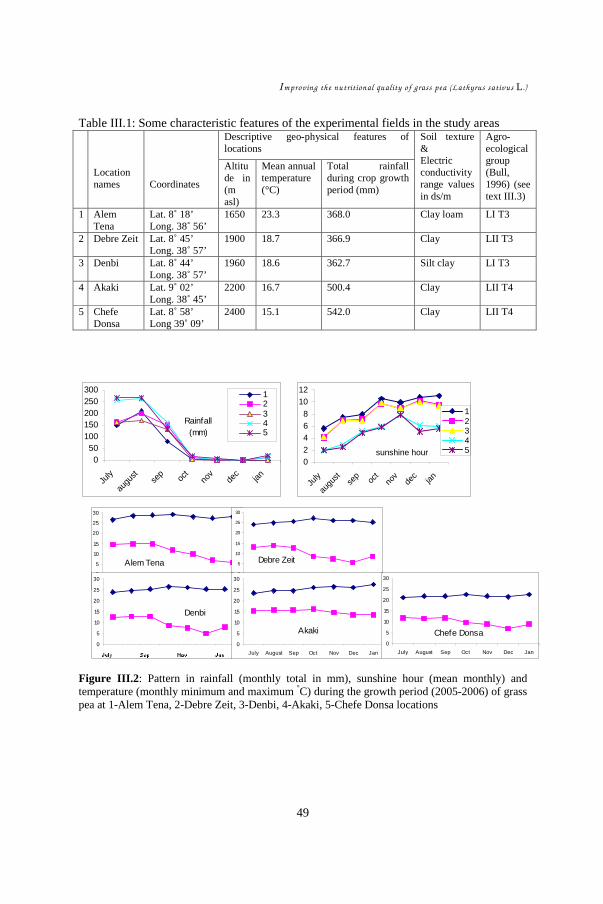
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
49
Table III.1: Some characteristic features of the experimental fields in the study areas Descriptive geo-physical features of locations
Location names
Coordinates
Altitude in (m asl)
Mean annual temperature (°C)
Total rainfall during crop growth period (mm)
Soil texture & Electric conductivity range values in ds/m
Agro- ecological group (Bull, 1996) (see text III.3)
1 Alem Tena
Lat. 8˚ 18’ Long. 38˚ 56’
1650 23.3 368.0 Clay loam
LI T3
2 Debre Zeit Lat. 8˚ 45’ Long. 38˚ 57’
1900 18.7 366.9 Clay
LII T3
3 Denbi
Lat. 8˚ 44’ Long. 38˚ 57’
1960 18.6 362.7 Silt clay
LI T3
4 Akaki
Lat. 9˚ 02’ Long. 38˚ 45’
2200 16.7 500.4 Clay
LII T4
5 Chefe Donsa
Lat. 8˚ 58’ Long 39˚ 09’
2400 15.1 542.0 Clay
LII T4
050
100150200250300
July
augu
st sep oc
tno
vde
cjan
12345
Rainfall (mm)
Figure III.2 : Pattern in rainfall (monthly total in mm), sunshine hour (mean monthly) and temperature (monthly minimum and maximum °C) during the growth period (2005-2006) of grass pea at 1-Alem Tena, 2-Debre Zeit, 3-Denbi, 4-Akaki, 5-Chefe Donsa locations
0
5
10
15
20
25
30
July August Sep Oct Nov Dec Jan
Alem Tena0
5
10
15
20
25
30
July August Sep Oct Nov Dec Jan
Debre Zeit
0
5
10
15
20
25
30
Denbi
0
5
10
15
20
25
30
July August Sep Oct Nov Dec Jan
Akaki0
5
10
15
20
25
30
July August Sep Oct Nov Dec Jan
Chefe Donsa
02468
1012
July
augu
stse
p oct
nov
dec
jan
12345sunshine hour
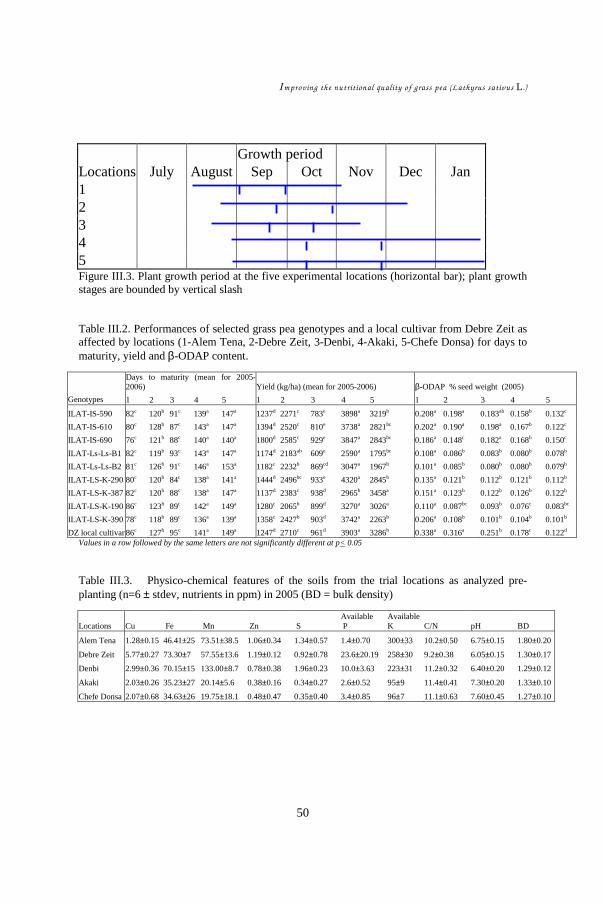
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
50
Growth period Locations July August Sep Oct Nov Dec Jan 1 2 3 4 5 Figure III.3. Plant growth period at the five experimental locations (horizontal bar); plant growth stages are bounded by vertical slash Table III.2. Performances of selected grass pea genotypes and a local cultivar from Debre Zeit as affected by locations (1-Alem Tena, 2-Debre Zeit, 3-Denbi, 4-Akaki, 5-Chefe Donsa) for days to maturity, yield and β-ODAP content.
Days to maturity (mean for 2005-2006) Yield (kg/ha) (mean for 2005-2006) β-ODAP % seed weight (2005)
Genotypes 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5
ILAT-IS-590 82c 120b 91c 139a 147a 1237d 2271c 783e 3898a 3219b 0.208a 0.198a 0.183ab 0.158b 0.132c
ILAT-IS-610 80c 128b 87c 143a 147a 1394d 2520c 810e 3738a 2821bc 0.202a 0.190a 0.198a 0.167b 0.122c
ILAT-IS-690 76c 121b 88c 140a 140a 1800d 2585c 929e 3847a 2843bc 0.186a 0.148c 0.182a 0.168b 0.150c
ILAT-Ls-Ls-B1 82c 119b 93c 143a 147a 1174d 2183ab 609e 2590a 1795bc 0.108a 0.086b 0.083b 0.080b 0.078b
ILAT-Ls-Ls-B2 81c 126b 91c 146a 153a 1182c 2232b 869cd 3047a 1967b 0.101a 0.085b 0.080b 0.080b 0.079b
ILAT-LS-K-290 80c 120b 84c 138a 141a 1444d 2496bc 933e 4320a 2845b 0.135a 0.121b 0.112b 0.121b 0.112b
ILAT-LS-K-387 82c 120b 88c 138a 147a 1137d 2383c 938d 2965b 3458a 0.151a 0.123b 0.122b 0.126b 0.122b
ILAT-LS-K-190 86c 123b 89c 142a 149a 1280c 2065b 899d 3270a 3026a 0.110a 0.087bc 0.093b 0.076c 0.083bc
ILAT-LS-K-390 78c 118b 89c 136a 139a 1358c 2427b 903d 3742a 2263b 0.206a 0.108b 0.101b 0.104b 0.101b
DZ local cultivar 86c 127b 95c 141a 149a 1247d 2710c 961d 3903a 3286b 0.338a 0.316a 0.251b 0.178c 0.122d Values in a row followed by the same letters are not significantly different at p< 0.05
Table III.3. Physico-chemical features of the soils from the trial locations as analyzed pre-planting (n=6 ± stdev, nutrients in ppm) in 2005 (BD = bulk density)
Locations Cu Fe Mn Zn S Available P
AvailableK C/N pH BD
Alem Tena 1.28±0.15 46.41±25 73.51±38.5 1.06±0.34 1.34±0.57 1.4±0.70 300±33 10.2±0.50 6.75±0.15 1.80±0.20
Debre Zeit 5.77±0.27 73.30±7 57.55±13.6 1.19±0.12 0.92±0.78 23.6±20.19 258±30 9.2±0.38 6.05±0.15 1.30±0.17
Denbi 2.99±0.36 70.15±15 133.00±8.7 0.78±0.38 1.96±0.23 10.0±3.63 223±31 11.2±0.32 6.40±0.20 1.29±0.12
Akaki 2.03±0.26 35.23±27 20.14±5.6 0.38±0.16 0.34±0.27 2.6±0.52 95±9 11.4±0.41 7.30±0.20 1.33±0.10
Chefe Donsa 2.07±0.68 34.63±26 19.75±18.1 0.48±0.47 0.35±0.40 3.4±0.85 96±7 11.1±0.63 7.60±0.45 1.27±0.10
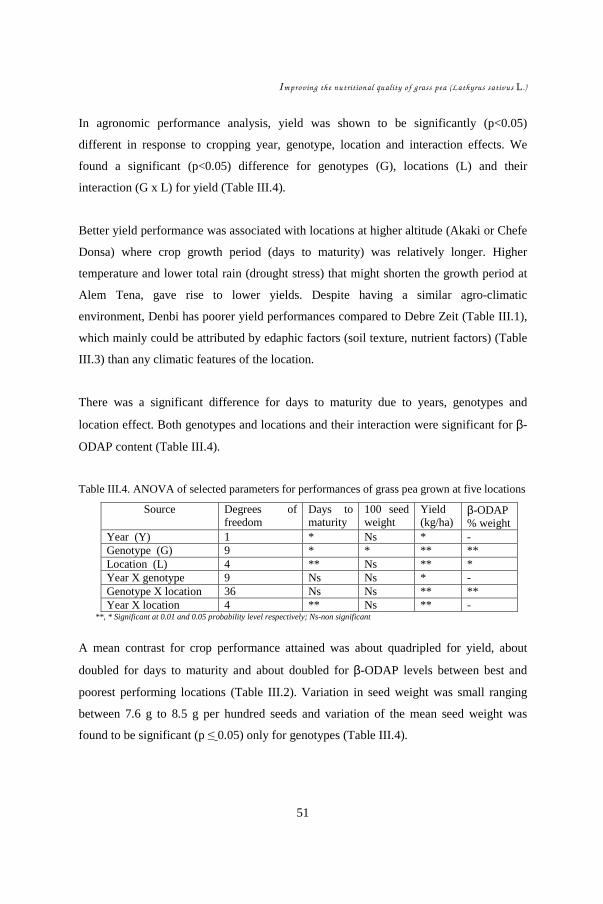
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
51
In agronomic performance analysis, yield was shown to be significantly (p<0.05)
different in response to cropping year, genotype, location and interaction effects. We
found a significant (p<0.05) difference for genotypes (G), locations (L) and their
interaction (G x L) for yield (Table III.4).
Better yield performance was associated with locations at higher altitude (Akaki or Chefe
Donsa) where crop growth period (days to maturity) was relatively longer. Higher
temperature and lower total rain (drought stress) that might shorten the growth period at
Alem Tena, gave rise to lower yields. Despite having a similar agro-climatic
environment, Denbi has poorer yield performances compared to Debre Zeit (Table III.1),
which mainly could be attributed by edaphic factors (soil texture, nutrient factors) (Table
III.3) than any climatic features of the location.
There was a significant difference for days to maturity due to years, genotypes and
location effect. Both genotypes and locations and their interaction were significant for β-
ODAP content (Table III.4).
Table III.4. ANOVA of selected parameters for performances of grass pea grown at five locations
Source Degrees of freedom
Days to maturity
100 seed weight
Yield (kg/ha)
β-ODAP % weight
Year (Y) 1 * Ns * - Genotype (G) 9 * * ** ** Location (L) 4 ** Ns ** * Year X genotype 9 Ns Ns * - Genotype X location 36 Ns Ns ** ** Year X location 4 ** Ns ** -
**, * Significant at 0.01 and 0.05 probability level respectively; Ns-non significant
A mean contrast for crop performance attained was about quadripled for yield, about
doubled for days to maturity and about doubled for β-ODAP levels between best and
poorest performing locations (Table III.2). Variation in seed weight was small ranging
between 7.6 g to 8.5 g per hundred seeds and variation of the mean seed weight was
found to be significant (p ≤ 0.05) only for genotypes (Table III.4).

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
52
The correlation structure elucidated by the path analysis (Table III.5) with the path
coefficient in the diagonal indicates the level of influence each predictor variable causes
directly on the response variable (seed yield). The direct effects in the correlation reveal
that number of days to maturity has the highest direct effect value and also accounts for
most of the variation (0.94). The second highest value of direct effect was due to rainfall
(RFI) during early growth stage (0.31). Cu2+ and Zn2+ have modest negative direct effects,
whereas the Zn2+/S2- ratio has a very small direct effect. Since no path coefficint values
exceed unity, we conclude that multicolinearity has not produced any inflated path
coefficient values as described in Williams et al. (1990).
Considering the indirect effects on yield in Table III.5, the variable days to maturity have
the greatest indirect effect (-0.34, -0.63, -0.66, 0.43) on yield through other factors.
Rainfall has got modest indirect effects on yield (-0.22, 0.12, -0.18) through all other
factors except Cu2+ (-0.03) which has got the least indirect effect. Smaller indirect effects
(<0.1) were seen for Cu2+, Zn2+ and Zn2+/S2-. Correlation of predictor variables with the
response variable ranged from 0.38 to 0.91 and coefficient of determination was 0.97.
Table III.5: Path analysis of the direct effects (bold) and indirect effects of the most relevant parameters: days to maturity, copper (Cu2+ in ppm), zinc (Zn2+ in ppm), total rain fall during stage I (RFI) on seed yield in grass pea grown at five locations in Ethiopia
Maturity
Cu2+
Zn2+
RFI
Zn2+: S2- r R2
Maturity 0.94 0.07 0.09 -0.22 0.04 0.91** 0.97
Cu2+
-0.34 -0.19 -0.06 -0.03 -0.01 -0.63**
Zn2+
-0.63 -0.08 -0.13 0.12 -0.01 -0.73**
RFI -0.66 0.02 -0.05 0.31 -0.04 -0.43**
Zn2+
/ S2- 0.43 0.03 0.02 -0.18 0.08 0.38* *, **- Significant at respectively 0.05 and 0.01 probability
The higher concentration of micronutrients S2- and Mn2+, at Denbi could be additional
factors to be considered having a negative effect on the yield of the crop, as they appear
to have a negative association (Figure III.4). These nutrients also produced a higher ratio
with Fe2+ and Cu2+ at the same location.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
53
From the graphically illustration of relative distribution of plant and soil parameters
(Figure III.4), it is rather complex to derive a simple linear relation, but some general
trends will be deducted. The concentration of β-ODAP increases from Chefe Donsa
(highland) to Alem Tena (lowland) whereas days to maturity and crop yield (except at
Denbi), decreased in the same direction. Zn2+/ S2- and Zn2+/Mn2+ are higher at higher
altitude whereas the ratios Zn2+/Cu2+ and Zn2+/Fe2+ are higher at locations where we
found higher β-ODAP in the seeds. Association of high Zn2+/Fe2+with high β-ODAP
might be in contradiction of the pot experimentl results by Haque (1997).
0%
20%
40%
60%
80%
100%
alt
B-oda
p %
met
hio m
g/g
pro
mat
urity
days
100s
w
yd kg
/ha
pH: H
2O
Cu ppm
Fe
ppm
Mn p
pm
Zn p
pm
S p
pmZn:C
uZn:F
e
Zn:Mn
Zn:S S:Cu
S:Fe
S:M
n
Mn:
Fe
Mn:
CuFe
:Cu
parameters
rela
tive
valu
es
Alem Tena
Debre Zeit
Denbi
Akaki
Chefe Donsa
Figure III.4. Graphical presentation for relative quantitative distribution of plant parameters and soil nutrients analysed at the five growing locations. Data for each parameter for the five locations were added and considered as 100% and relative values spatially plotted.
Path analysis with 11 predictors for β-ODAP, selected on the basis of stepwise regression
was run the results are shown in Table III.6. Among the selected parmeters, the largest
direct effects were found for Zn2+/P (-0.55), days to maturity (-0.44), yield (-0.39) and K+
(0.39). Smaller direct effects were found for sshIII (0.35), pH (-0.19), Fe2+(0.31), Zn2+/
Fe2+(0.25) soil bulk density (BD) (-0.21), TempII (-0.23) and RFII (0.22). Among larger
indirect effects, yield showed the highest indirect effect through days to maturity (-0.40),
days to maturity showed the highest indirect effect through K+ (-0.34) and K+ showed the
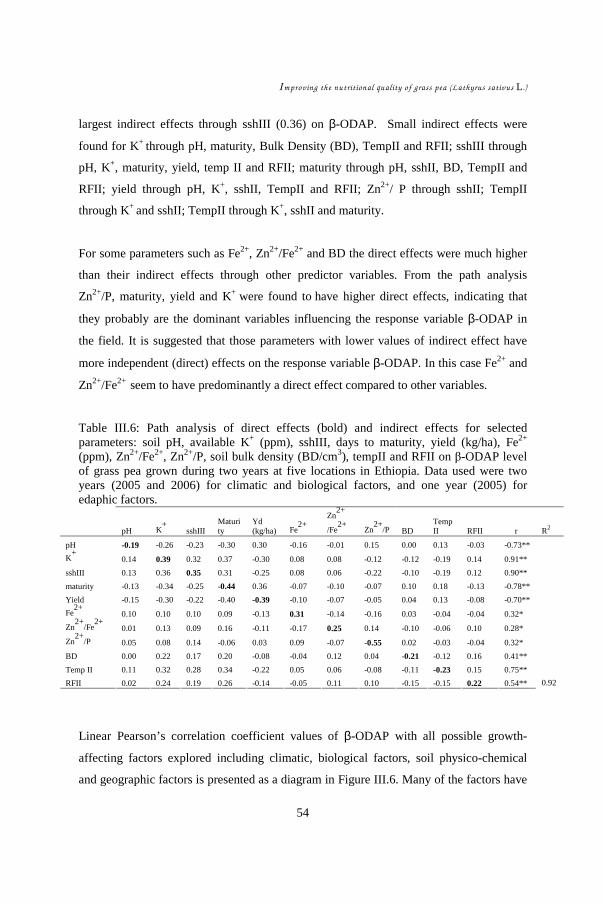
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
54
largest indirect effects through sshIII (0.36) on β-ODAP. Small indirect effects were
found for K+ through pH, maturity, Bulk Density (BD), TempII and RFII; sshIII through
pH, K+, maturity, yield, temp II and RFII; maturity through pH, sshII, BD, TempII and
RFII; yield through pH, K+, sshII, TempII and RFII; Zn2+/ P through sshII; TempII
through K+ and sshII; TempII through K+, sshII and maturity.
For some parameters such as Fe2+, Zn2+/Fe2+ and BD the direct effects were much higher
than their indirect effects through other predictor variables. From the path analysis
Zn2+/P, maturity, yield and K+ were found to have higher direct effects, indicating that
they probably are the dominant variables influencing the response variable β-ODAP in
the field. It is suggested that those parameters with lower values of indirect effect have
more independent (direct) effects on the response variable β-ODAP. In this case Fe2+ and
Zn2+/Fe2+ seem to have predominantly a direct effect compared to other variables.
Table III.6: Path analysis of direct effects (bold) and indirect effects for selected parameters: soil pH, available K+ (ppm), sshIII, days to maturity, yield (kg/ha), Fe2+ (ppm), Zn2+/Fe2+, Zn2+/P, soil bulk density (BD/cm3), tempII and RFII on β-ODAP level of grass pea grown during two years at five locations in Ethiopia. Data used were two years (2005 and 2006) for climatic and biological factors, and one year (2005) for edaphic factors.
pH K+
sshIII
Maturity
Yd (kg/ha) Fe
2+
Zn2+
/Fe2+
Zn2+
/P BD Temp II RFII r R2
pH -0.19 -0.26 -0.23 -0.30 0.30 -0.16 -0.01 0.15 0.00 0.13 -0.03 -0.73**
K+
0.14 0.39 0.32 0.37 -0.30 0.08 0.08 -0.12 -0.12 -0.19 0.14 0.91**
sshIII 0.13 0.36 0.35 0.31 -0.25 0.08 0.06 -0.22 -0.10 -0.19 0.12 0.90**
maturity -0.13 -0.34 -0.25 -0.44 0.36 -0.07 -0.10 -0.07 0.10 0.18 -0.13 -0.78**
Yield -0.15 -0.30 -0.22 -0.40 -0.39 -0.10 -0.07 -0.05 0.04 0.13 -0.08 -0.70**
Fe2+
0.10 0.10 0.10 0.09 -0.13 0.31 -0.14 -0.16 0.03 -0.04 -0.04 0.32*
Zn2+
/Fe2+
0.01 0.13 0.09 0.16 -0.11 -0.17 0.25 0.14 -0.10 -0.06 0.10 0.28*
Zn2+
/P 0.05 0.08 0.14 -0.06 0.03 0.09 -0.07 -0.55 0.02 -0.03 -0.04 0.32*
BD 0.00 0.22 0.17 0.20 -0.08 -0.04 0.12 0.04 -0.21 -0.12 0.16 0.41**
Temp II 0.11 0.32 0.28 0.34 -0.22 0.05 0.06 -0.08 -0.11 -0.23 0.15 0.75**
RFII 0.02 0.24 0.19 0.26 -0.14 -0.05 0.11 0.10 -0.15 -0.15 0.22 0.54** 0.92
Linear Pearson’s correlation coefficient values of β-ODAP with all possible growth-
affecting factors explored including climatic, biological factors, soil physico-chemical
and geographic factors is presented as a diagram in Figure III.6. Many of the factors have

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
55
a significant correlation with the level of β-ODAP in the seed. Altitude (-0.85), days to
maturity (-0.78), soil pH (-0.72) and crop yield (-0.71) have highly significant negative
correlations with β-ODAP content, while K+ (0.86) and ssh III (0.80) have a significant
positive correlation to β-ODAP. The lowest level of β-ODAP was found in neutral to
weakly basic soil pH (6.7-8) conditions. Haque (1997) reported that the lowest value of
β-ODAP was produced at a pH level around neutral. We found the factor sunshine hours
to have a strong positive correlation with β-ODAP, the greatest (r = 0.80** ) being
recorded during post-anthesis phases of the plant (ssh III).
R F II 0 .5 4 **
R F III -0 .5 3 **
S W -0 .5 2 **
te m p II 0 .5 2 **
S :F e -0 .5 2 **
S :C u 0 .5 1 **
Z n ;S -0 .4 9 ** S 0 .4 7 **
te m p I0 .4 4 ** C :N -0 .4 2 **
Z n ;C u 0 .4 1 ** B D 0 .3 9 **
P 0 .3 4 **
F e 0 .3 2 *
Z n :M n -0 .3 2 *
Z n :P 0 .3 1 *
Z n :F e 0 .2 9 *
C u 0 .2 4
Z n :K 0 .0 6 S M n 0 .1 5 F e :C u 0 .1 5
te m p III0 .5 4 ** R F I0 .5 5 *
M n :C u 0 .5 5 * M n :F e 0 .5 5 **
Z n 0 .6 0 ** M n 0 .6 2 ** y ie ld -0 .7 1 **
S S H II0 .7 1 ** P H -0 .7 2 **
S S H I0 .7 7 ** m a tu r ity -0 .7 8 **
S S H III0 .8 0 ** a lt-0 .8 5 ** K 0 .8 6 **
Figure III.6. Pearson’s correlation coefficients of the different environmental and plant variables with β-ODAP in L. sativus (with the strongest correlation for K+ and the weakest for Zn2+/K+)
III.4. Discussion
The experimental plots are located in agro-ecology areas of grass pea production and
hence, the results reflects aspects of grass pea growth conditions in Ethiopia. A number
of important environmental factors were found to affect crop yield and β-ODAP content
in Lathyrus sativus seeds. In general, the soil in the locations was neutral to slightly
acidic or slightly basic, which may favour grass pea growth. Previous work indicated
ββββ----ODAP

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
56
that the soils of the study locations have low levels of organic matter, total nitrogen, P,
Cu2+ and Zn2+ (Abayneh and Demeke, 2003).
The crop growing period gets longer with higher altitude. Higher altitude is also
associated with more cloudiness, an increment in total amount of rain and length of the
rainy period. The better moisture holding capacity of the soil, lower temperature and
smaller sunshine hours (see Table III.1 and Figure III.2) at those higher locations might
have allowed an extention of the reproductive period and thereby better yield. The
number of sunshine hours was less in the highland compared to lowland areas due mainly
to the longer periods of cloud cover in the highlands. Akaki followed by Chefe Donsa
location were found to be the best yielding with longer crop growing period among the
five locations. In addition, this could be associated with the narrower range of max-min
temperatures and the higher C/N that might create better conditions for plant performance
at Akaki.
Days to maturity and amount of rainfall seem to be the dominant effectors for crop yield
as they showed a higher level of direct effect than the other factors considered. The
longer days to maturity period might have allowed effective fertilization of a higher
number of flowers and a longer period of translocation of biosynthates to an increased
number of seed sinks. Thus higher yield would be a resultant effect both from a larger
number of pods and better seed filling. By contrast, a shorter growth period at Alem
Tena and Denbi could be related more to the light soil texture that allow more
evapotranspiration forcing the plant into early maturity and poorer yield. Significant
correlations have been reported between seed yield and length of seed filling period (r =
0.57; Hanson, 1985) whereas a long maturation period is generally associated with
increased seed size (Swank et al., 1987). This was also clear from our own path
coefficient analysis: the greatest effect on yield was found for days to maturity (0.94)
followed by rainfall during crop establishment (0.31). Likewise, in other experiments,
significant (P<0.05) yield variation was reported ranging between 1.2 to 3.9 t/ha due to
locations in Ethiopia (Fikre et al., 2006), between 1.2 to 1.8 in winter and between 1.1 to

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
57
2.2 in spring seasons in Turkey (Düsünceli, 1993) and between 0.1 to 2.5 due to genotype
effect in Bangladesh (Campbell, 1997).
The significance of G x L (genotype by location) interaction for yield suggests that the
different varieties respond differently to the diverse environments as previously reported
by Wuletaw (2003). We found that better yield performance was associated with
locations at higher altitude (Akaki) where the growing season was longer. The longer
growth period must have allowed sufficient translocation of biosynthates to the sink.
There is ample evidence that yield improvement in crops has come from a longer
reproductive phase duration, which leads to a higher seed harvest index (Nelson, 1986;
Smith and Nelson, 1986a and 1986b). In contrast, lower yield recorded in drought stress
environments could have contributed partially to forced earlier maturation, leading to
development of a reduced size of reproductive sink as a physiological phenomenon (Hall,
2004). This condition induces higher concentration of β-ODAP in the relatively smaller
sink size (Cocks et al., 2000).
Nutrient optimization is one crucial element in the soil complex for enhanced
productivity of the crop. If one nutrient is lacking, it may negatively affect the availability
of others. The response of crops to supplementation, in cases where deficiency exists, can
be very pronounced. Micronutrient levels may have negatively affected yield as revealed
by the observation that there is a strong negative correlation between higher
concentration of S2- and Mn2+ and poor yield. The higher abundance of S2- and Mn2+ at
Denbi might explain its toxic levels and possible interferance with the normal
physiological performance of the plants to have lowest yield.
Potassium is absorbed by plants in larger amounts than any other mineral element except
nitrogen and, in some cases, calcium. Yield, Zn2+/P, maturity and K+ have the largest
direct effect on β-ODAP level in the path analysis. The total effect of K+, sshII, days to
maturity, pH and TempII on β-ODAP appear to be considerably high, presupposing their
corresponding influence. However, since these factors have got a substantial level of
indirect effect through other variables, β-ODAP could not be predicted from a single or

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
58
very few factors only. In Pearson’s correlation coefficient analysis, K+ showed the
strongest positive correlation with β-ODAP among all factors considered. This may
suggest that K+, which is reported to be high in Ethiopian soils (Murphy, 1968), could be
an important edaphic factor affecting β-ODAP levels in Ethiopian grass pea. Haque
(1997) reported that excess salinity cations including K+ could enhance the β-ODAP level
while lower salinity levels had a decreasing effect.
Many reports proposed that high concentrations of Zn2+ in the growing medium decrease
the level of β-ODAP in the seed (Lambein et al., 1994; Haque, 1997), possibly by its
chelating action. We found a positive correlation between β-ODAP and Zn2+ per se,
however, this became negative when combined with other nutrients. This conflicting
output might emerge from overall deficiency of Zn2+ in the soil (Abayneh and Demeke,
2003) and/or plants growing in an open field of complex environment could not simply
be reproduced in performance from controlled pot experiments reported earlier (Haque,
1997). Hence, the nutrient balance in the complex field condition could have a more
important role than a single nutrient (Haque, 1997).
In agreement with the results obtained from pot experiments by Haque (1997) the weakly
alkaline soil pH (6.7 – 8.0) appeared to effectively decrease β-ODAP. This could be the
pH where nutrient availability and plant performances are optimized, and whereby
accumulation of the stress metabolite β-ODAP is reduced.
Zhang and co-workers (2003) reported that higher light intensity could promote β-ODAP
levels. In contrast, we found higher β-ODAP values in lowland areas (1600 m asl) with
less light intensity than in highland areas (2200 m asl). In our analysis, the length of shh
period was found to have a strong positive correlation with β-ODAP, the greatest
correlation (r = 0.80** ) being recorded in the maturation phase (ssh III) of the plant. We
propose that the higher light intensity in the highlands combined with shorter light hour
periods (mean = 5 hrs/day with solar irradiance > 120 W/m2) most likely result in lower
light stress than the lower intensity combined with longer light periods (mean = 9.5

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
59
hrs/day) of the lowlands during the crop growth period. In other words, the cloud cover in
the highlands was longer than in the lowlands like Alem Tena.
In general, the effect of stress due to climatic factors (temperature, rain fall, sunshine
hours) on β-ODAP accumulation was apparently stronger during post-anthesis or during
the reproductive phase than at early growth stages. Similarly, Cocks and co-workers
(2000) reported that β-ODAP doubled due to drought stress at post-anthesis compared to
drought stress at pre-anthesis. Likewise, soil factors seem to have substantial influence on
the crop yield and β-ODAP content. Hence, in addition to the effect of genotype, β-
ODAP accumulation in the seed of grass pea is also considerably influenced by
environmental (edaphic and climatic) factors in Ethiopia. These results can be part of the
information needed for better agricultural management to produce a grass pea crop in
more suitable agro-ecologies that may yield seed with better nutritional quality.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
60

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
61
CHAPTER IV
Effect of methionine supplement on physical responses and
neurological symptoms of broiler chicks fed grass pea based
starter ration

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
62
Young chicks feeding on grass pea diet, one of them with torticolis (neck bending) as
neurological symptoms

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
63
CHAPTER IV
Effect of methionine supplement on physical responses and neurological
symptoms of broiler chicks fed grass pea based starter ration
Abstract
Starter feeding experiments of broiler chicks with raw grass pea (Lathyrus sativus L.,
cultivar Debre Zeit local, containing 0.35 % neuro-excitatory amino acid ß-ODAP)
supplemented with different levels of methionine were undertaken during 2005 at EIAR
(Debre Zeit Center, Ethiopia) for 30 days to assess toxicity of grass pea based feed and
to correlate it with animal performances. Four hundred fifty one-day-old broiler chicks
were randomly divided into two groups, each group was given formulations containing
35 % (ration I) or 98.5 % (ration II) grass pea respectively. Each ration included controls
and treatments with four concentrations of methionine (0.025, 0.05, 0.075 and 0.1% of
the feed in ration I; 0.05, 0.1, 0.15 and 0.2 % of the feed in ration II). Each treatment was
conducted with 45 chicks, which were further randomly separated into three replicates of
15 chicks each. Performances of the chicks (feed intake, weight gain, feed conversion
efficiency) were much better in ration I than in ration II. Those parameters were
significantly improved by addition of methionine (0.075 % of the feed in ration I and 0.15
% in ration II respectively). Significant reduction (from 84 % to 2 %) of acute
neurological signs with addition of methionine was observed in grass pea ration II. The
feed intake tolerance for unprocessed grass pea was enhanced with increasing methionine
in the diet and with age. Despite a similarity in the initial intake, a significant (p≤0.05)
difference in the final feed intake level by the chicks with methionine addition compared
to the controls was found in both rations. The total weight gain per head improved by
methionine addition for up to 26.3 % and 65.5 % in ration I and II respectively.
Reversible convulsions developed in some chicks within five days after feeding, the
highest incidence was found in ration II (94 % of the total number of cases) with most

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
64
cases (89 %) in the controls (without addition of methionine) and in chicks on feed with
0.05 % methionine addition. These results suggest that methionine can improve a grass
pea-based diet for the growth of broiler chicks and also can protect young chicks from
neurological symptoms. This drought tolerant, cheap and protein-rich grass pea when
supplemented with optimal methionine can at least partially replace soybean in animal
feed during food crises.
IV.1. Introduction
Dependent consumption of grass pea (L. sativus L.) for more than two months can give
rise to the crippling neurolathyrism in humans, giving the stigma of toxicity to this crop.
Toxicity is blamed on the presence of a non-protein amino acid, β-N-oxalyl-L-α,β-
diaminopropionic acid (ß-ODAP), that acts as a potent agonist of the AMPA subclass of
neuronal glutamate receptors (Spencer et al., 1986). Like all legumes, grass pea is
deficient in the essential sulphur amino acids methionine and cysteine, a deficiency that
might play an important role in its toxicity (Nunn et al., 2005).
Neurological signs can be experimentally induced in animals. In young chicks these signs
are acute and reversible and do not necessarily reflect the aetiology of human
neurolathyrism. Young chicks show specific but reversible signs within one to five days
after grass pea consumption or injection of ß-ODAP; while for primates on a balanced
diet, much higher doses of ß-ODAP are needed for a longer period to develop central
motor neuron deficiency and hind limb spasticity (Spencer et al., 1986). Rao (2001)
demonstrated that there exist extreme inter-species and intra-species differences in
susceptibility to ß-ODAP that may be ascribed to individual metabolic differences.
Currently, both cellular (Kusama-Eguchi et al., 2003) and animal models (Kusama-
Eguchi et al., 2005) for human neurolathyrism are being developed.
Several authors suggested a role for oxidative stress in the aetiology of neurolathyrism
(Lambein et al., 2001; Nunn et al., 2005; Getahun et al., 2005). Depletion of methionine
and cysteine in the bloodstream and in the central nervous system (CNS) can jeopardize

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
65
the defence against oxidative stress and increase the risk of neurolathyrism. A low level
of methionine in urine of lathyrism patients consuming grass pea had been reported
(Rudra and Chaudhury, 1952) even before the toxic compound was identified in grass
pea. Consumption of grass pea by mixing with at least one third of cereals (richer in
sulphur amino acid than legumes) in the diet was identified as a protective factor for
neurolathyrism (Getahun et al., 2005).
Based on a study with human volunteers, Nunn et al. (2005) found that consumption of
grass pea seed resulted in a significant and persistent decrease in plasma methionine. The
normal physiological response to consuming a methionine-deficient meal is suggested to
be reduced plasma methionine concentration. Methionine-deficient grass peas and lentils
were seen to reduce plasma methionine by about 50 % persistently for 24 hours. High
methionine concentrations are found in rat motor neurons, which suggests a high
requirement for methionine by these cells, and perhaps an increased sensitivity to
methionine deficiency. A reduction in plasma methionine concentration could thus cause
a reduced methionine flux into the CNS. As a consequence of prolonged consumption of
grass pea seed as a staple, neuronal cells may be sensitised to the additional oxidative
stress produced by over-excitation of the motor neurons by ß-ODAP and the ensuing
production of nitric oxide (NO) and reactive oxygen species (ROS). Nunn and co-
workers (2005) proposed that supplementing such a diet with methionine might protect
those at risk from neurolathyrism.
The present study was initiated with the hypothesis that methionine plays aginst lathyrism
and lathyrism symptom development in chicks with possible implications in humans. The
objective of the present study was to assess the effect of methionine on grass pea toxicity
by studying neurolathyritic symptoms and general performance of broiler chicks fed with
rations containing grass pea.
IV.2. Materials and Methods The study area The experiments were conducted in the Poultry Research Department at
Debre Zeit Agricultural Research Centre (DZARC), Ethiopia.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
66
Experimental animals and their management Four hundred fifty mixed sex one-day-
old broiler chicks of similar body weight were randomely divided into sets of 225 chicks.
These were subdivided into five groups of 45 receiving the control diet or the four
different levels of methionine addition to the diet. Each group was further randomly sub-
divided into three replicates of 15 chicks and placed in experimental pens at a density of
10 chicks/m2. Chicks on various rations were randomly assigned to pens. Birds were
provided daily with a known amount of feed early in the morning, refusals were weighed
by the end of the day, and the difference calculated as the daily intake value per sub-
group of 15 chicks. Water was provided ad libitum. Routine vaccination and health care
was given as necessary. Chicks’ body weight measurements were taken + 0.1 g for each
replicate group as a whole starting at the beginning of the trial and every week thereafter.
Symptom-developing individual chicks in a replication were marked with different
colours for further monitoring and characterization. No mortality occurred during the
experiment.
Feeding rations Broiler starter rations with two levels of raw grass pea (Debre Zeit local
cultivar containing 0.35 % β-ODAP) as diet were formulated. These served as control
rations. Treatment rations were formulated by adding different levels of DL-methionine
(≥99 %, feed grade) produced by Sumitomo Chemical Company, Tokyo, Japan. Raw
grass pea seeds were coarsely ground with an electric mill and homogeneously mixed
with the other ingredients yielding rations I and II, respectively. The composition of the
two rations is detailed below.
Ration I = 48 % maize, 35 % grass pea, 15.5 % noug cake (protein supplement widely
available in Ethiopia. It is the residue after extraction of oil from niger seed or Guizotia
abyssinica, Asteraceae), 1 % vitamin-mineral premix (commercial premix containing per
kg: 625000 IU vitamin A, 125000 IU vitamin D3, 1250 IU vitamin E, 1500 mg Fe, 1250
mg Cu, 3000 mg Zn, 3750 mg Mn, 15 mg Co and 10 mg Se), 0.5 % NaCl. Four levels of
methionine (as % of feed) were used: 0.025, 0.05, 0.075 and 0.10.
Ration II = 98.5 % grass pea, 1 % vitamin-mineral premix (as in ration I), and 0.5 %

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
67
NaCl. Four levels of methionine (as % of feed) were used: 0.05, 0.1, 0.15, and 0.2.
Chemical analysis Samples of the two feed rations were analysed for proximate
principles based on the standard procedures of AOAC (Williams, 1984) at the
Department of Analytical and Physical Chemistry, Faculty of Bioscience Engineering,
Ghent University. Dry matter (DM) and ash determination were done using the
procedures described in Williams (1984) and De Vleeshauwer et al. (1948). Protein and
fat content were determined using the Kjeldahl method as described in Egan et al. (1981)
and the Weibull method (Pearson’s, 1981) respectively. Carbohydrate values were
derived by differences whereas the energy values (mega joule/ kg) were calculated using
the physiological fuel values of carbohydrate, protein and fat (Table IV.1).
Concentrations (% seed weight) of β-ODAP in the feeding mixtures (ration I and II) were
analysed by HPLC with pre-column PITC derivatisation as described in the previous
chapter.
Table IV.1. Feed component analysis of a starter treatment ration for broiler chicks
% DM % Ash % Protein % Fat Energy (MJ/Kg)
% ß-ODAP
Ration I 89.63 ± 0.35 4.18± 0.02 16.71 ± 0.27 3.16 ± 0.78 16.6 ± 0.016 0.134 + 0.10 Ration II 89.32 ± 0.22 3.91± 0.38 22.81 ± 0.27 1.08 ± 0.46 16.3 ± 0.003 0.293+ 0.19
Statistical analysis
Experiment of the two rations was laid down in a Completely Randomised Design (CRD)
with three replications of 15 chicks each under the four treatment levels and control.
Variance between treatments was analysed using the General Linear Model (GLM)
procedure of the Statistical Analysis System (SAS, 2001). Duncan multiple range test
(Duncan, 1955) was applied to indicate when significant differences between treatments
means exist.
IV.3. Results With increasing methionine in the feed, the chicks’ performance, expressed as weight
gain, feed intake, feed conversion efficiency, showed improvement in both rations. The
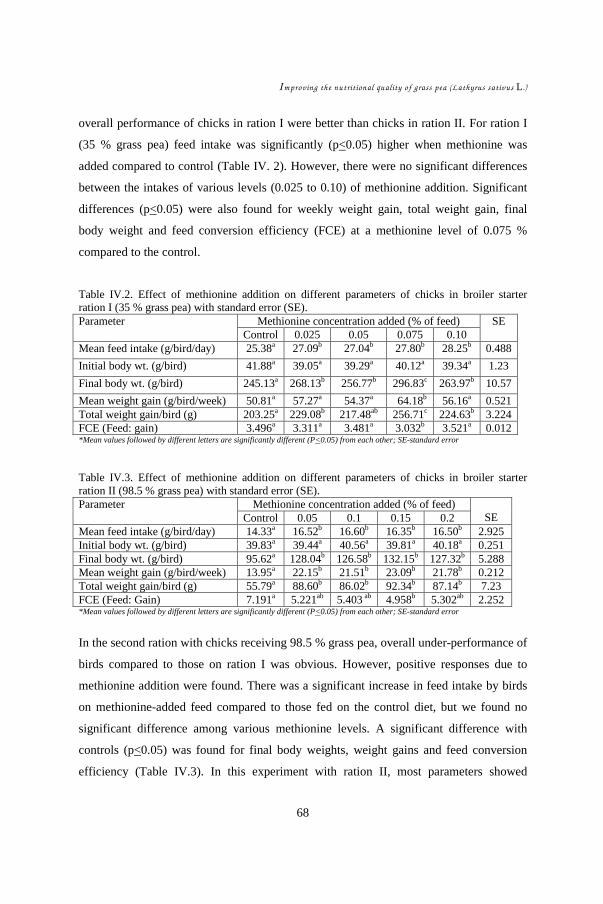
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
68
overall performance of chicks in ration I were better than chicks in ration II. For ration I
(35 % grass pea) feed intake was significantly (p<0.05) higher when methionine was
added compared to control (Table IV. 2). However, there were no significant differences
between the intakes of various levels (0.025 to 0.10) of methionine addition. Significant
differences (p<0.05) were also found for weekly weight gain, total weight gain, final
body weight and feed conversion efficiency (FCE) at a methionine level of 0.075 %
compared to the control.
Table IV.2. Effect of methionine addition on different parameters of chicks in broiler starter ration I (35 % grass pea) with standard error (SE).
Methionine concentration added (% of feed) Parameter Control 0.025 0.05 0.075 0.10
SE
Mean feed intake (g/bird/day) 25.38a 27.09b 27.04b 27.80b 28.25b 0.488
Initial body wt. (g/bird) 41.88a 39.05a 39.29a 40.12a 39.34a 1.23
Final body wt. (g/bird) 245.13a 268.13b 256.77b 296.83c 263.97b 10.57
Mean weight gain (g/bird/week) 50.81a 57.27a 54.37a 64.18b 56.16a 0.521 Total weight gain/bird (g) 203.25a 229.08b 217.48ab 256.71c 224.63b 3.224 FCE (Feed: gain) 3.496a 3.311a 3.481a 3.032b 3.521a 0.012 *Mean values followed by different letters are significantly different (P<0.05) from each other; SE-standard error
Table IV.3. Effect of methionine addition on different parameters of chicks in broiler starter ration II (98.5 % grass pea) with standard error (SE).
Methionine concentration added (% of feed) Parameter Control 0.05 0.1 0.15 0.2
SE
Mean feed intake (g/bird/day) 14.33a 16.52b 16.60b 16.35b 16.50b 2.925 Initial body wt. (g/bird) 39.83a 39.44a 40.56a 39.81a 40.18a 0.251 Final body wt. (g/bird) 95.62a 128.04b 126.58b 132.15b 127.32b 5.288 Mean weight gain (g/bird/week) 13.95a 22.15b 21.51b 23.09b 21.78b 0.212 Total weight gain/bird (g) 55.79a 88.60b 86.02b 92.34b 87.14b 7.23 FCE (Feed: Gain) 7.191a 5.221ab 5.403 ab 4.958b 5.302ab 2.252 *Mean values followed by different letters are significantly different (P<0.05) from each other; SE-standard error
In the second ration with chicks receiving 98.5 % grass pea, overall under-performance of
birds compared to those on ration I was obvious. However, positive responses due to
methionine addition were found. There was a significant increase in feed intake by birds
on methionine-added feed compared to those fed on the control diet, but we found no
significant difference among various methionine levels. A significant difference with
controls (p<0.05) was found for final body weights, weight gains and feed conversion
efficiency (Table IV.3). In this experiment with ration II, most parameters showed

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
69
maximum values at methionine levels of 0.15 % of the feed with a declining trend with
further increasing methionine addition. Total weight gain per bird from 55.79 g to 92.34 g
or an increase of 65 % in this exclusive grass pea ration II proves the importance of
methionine addition. In general, feed conversion efficiency, as expressed by g weight
gain per g consumed feed, of this group was low compared to the chicks on ration I.
However, it was improved with increasing methionine addition. Overall under-
performance of chicks on ration II as compared to that obtained in the birds on ration I
could be the consequence of higher level of other antinutritional factors (ANFs) that are
present in grass pea (Urga et al., 2005) as compared to the higher cereals ratio (48 %
maize) in ration I. Improved feed conversion efficiency resulted in a considerably higher
weight gain per chick yielding a higher total body weight at the end of the four weeks
feeding experiment.
Daily feed intake (difference between offered and refusal) by the chicks showed a drop
during the first three days for the high grass pea feed (ration II) followed by immediate
recovery (Figure IV.1). In both rations, feed intake gradually increased during the
experimental period to reach a maximum by the end of the experiment. An overall low
feed intake in ration II could be the effect of the high amount of antinutritional factors,
which also resulted in poor feed conversion efficiency. Weekly weight gain was
substantially higher in the first ration compared to ration II (that showed poorer FCE)
irrespective of similarity in the daily intake trend of both rations. This weight gain
showed a significant increase with time advance (P<0.05), except for ration II of the last
week that showed a decrease. We can conclude that feed intake increased with age of the
chicks, and was also positively affected by increasing methionine addition in both rations.
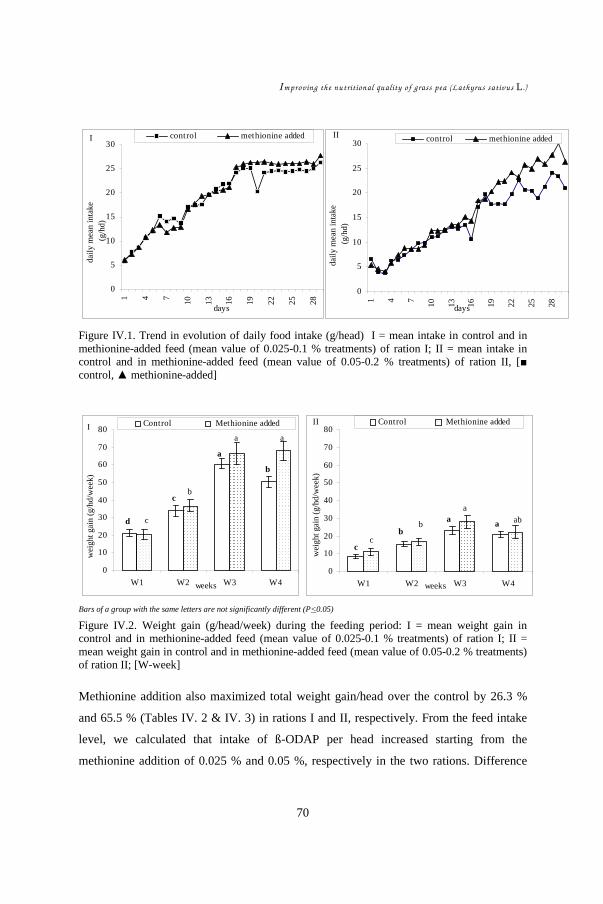
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
70
Figure IV.1. Trend in evolution of daily food intake (g/head) I = mean intake in control and in methionine-added feed (mean value of 0.025-0.1 % treatments) of ration I; II = mean intake in control and in methionine-added feed (mean value of 0.05-0.2 % treatments) of ration II, [■ control, ▲ methionine-added]
Bars of a group with the same letters are not significantly different (P<0.05)
Figure IV.2. Weight gain (g/head/week) during the feeding period: I = mean weight gain in control and in methionine-added feed (mean value of 0.025-0.1 % treatments) of ration I; II = mean weight gain in control and in methionine-added feed (mean value of 0.05-0.2 % treatments) of ration II; [W-week]
Methionine addition also maximized total weight gain/head over the control by 26.3 %
and 65.5 % (Tables IV. 2 & IV. 3) in rations I and II, respectively. From the feed intake
level, we calculated that intake of ß-ODAP per head increased starting from the
methionine addition of 0.025 % and 0.05 %, respectively in the two rations. Difference
I
0
5
10
15
20
25
30
1 4 7 10 13 16 19 22 25 28days
daily
me
an
inta
ke
(g/h
d)
control methionine added
0
5
10
15
20
25
30
1 4 7 10 13 16 19 22 25 28days
daily
me
an
inta
ke
(g/h
d)
control methionine added II
0
10
20
30
40
50
60
70
80
W1 W2 W3 W4weeks
we
ight
ga
in (
g/hd
/we
ek)
Control Methionine addedII
aabb
c
a ab
c
0
10
20
30
40
50
60
70
80
W1 W2 W3 W4weeks
we
ight
ga
in (
g/hd
/we
ek)
Control Methionine addedIa a
b
c
a
b
c
d
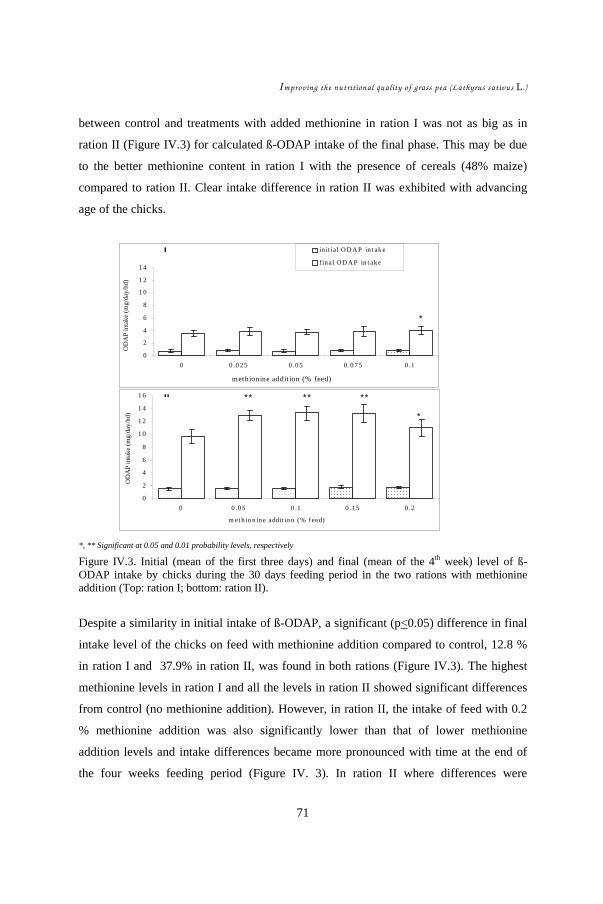
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
71
between control and treatments with added methionine in ration I was not as big as in
ration II (Figure IV.3) for calculated ß-ODAP intake of the final phase. This may be due
to the better methionine content in ration I with the presence of cereals (48% maize)
compared to ration II. Clear intake difference in ration II was exhibited with advancing
age of the chicks.
*, ** Significant at 0.05 and 0.01 probability levels, respectively
Figure IV.3. Initial (mean of the first three days) and final (mean of the 4th week) level of ß-ODAP intake by chicks during the 30 days feeding period in the two rations with methionine addition (Top: ration I; bottom: ration II).
Despite a similarity in initial intake of ß-ODAP, a significant (p<0.05) difference in final
intake level of the chicks on feed with methionine addition compared to control, 12.8 %
in ration I and 37.9% in ration II, was found in both rations (Figure IV.3). The highest
methionine levels in ration I and all the levels in ration II showed significant differences
from control (no methionine addition). However, in ration II, the intake of feed with 0.2
% methionine addition was also significantly lower than that of lower methionine
addition levels and intake differences became more pronounced with time at the end of
the four weeks feeding period (Figure IV. 3). In ration II where differences were
0
2
4
6
8
1 0
1 2
1 4
1 6
0 0 .05 0 .1 0 .1 5 0 .2
m et h io n ine addit io n (% feed)
OD
AP
inta
ke (
mg
/da
y/h
d)
II
*
******
0
2
4
6
8
1 0
1 2
1 4
0 0 .0 25 0 .05 0 .07 5 0 .1
methionine addit ion (% feed)
OD
AP
inta
ke (
mg
/da
y/h
d)
in it ial ODAP in t ak e
f inal ODAP in t ake
I
*
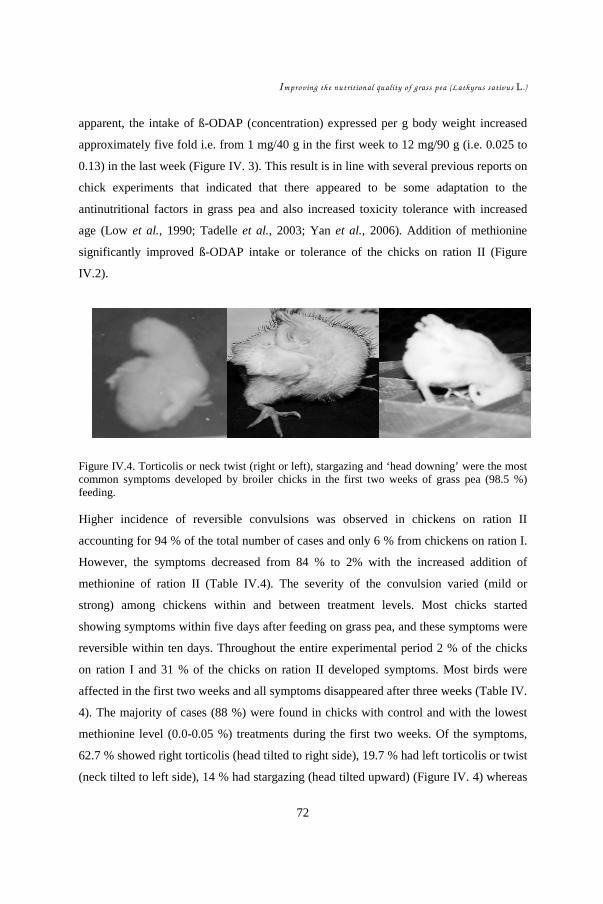
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
72
apparent, the intake of ß-ODAP (concentration) expressed per g body weight increased
approximately five fold i.e. from 1 mg/40 g in the first week to 12 mg/90 g (i.e. 0.025 to
0.13) in the last week (Figure IV. 3). This result is in line with several previous reports on
chick experiments that indicated that there appeared to be some adaptation to the
antinutritional factors in grass pea and also increased toxicity tolerance with increased
age (Low et al., 1990; Tadelle et al., 2003; Yan et al., 2006). Addition of methionine
significantly improved ß-ODAP intake or tolerance of the chicks on ration II (Figure
IV.2).
Figure IV.4. Torticolis or neck twist (right or left), stargazing and ‘head downing’ were the most common symptoms developed by broiler chicks in the first two weeks of grass pea (98.5 %) feeding. Higher incidence of reversible convulsions was observed in chickens on ration II
accounting for 94 % of the total number of cases and only 6 % from chickens on ration I.
However, the symptoms decreased from 84 % to 2% with the increased addition of
methionine of ration II (Table IV.4). The severity of the convulsion varied (mild or
strong) among chickens within and between treatment levels. Most chicks started
showing symptoms within five days after feeding on grass pea, and these symptoms were
reversible within ten days. Throughout the entire experimental period 2 % of the chicks
on ration I and 31 % of the chicks on ration II developed symptoms. Most birds were
affected in the first two weeks and all symptoms disappeared after three weeks (Table IV.
4). The majority of cases (88 %) were found in chicks with control and with the lowest
methionine level (0.0-0.05 %) treatments during the first two weeks. Of the symptoms,
62.7 % showed right torticolis (head tilted to right side), 19.7 % had left torticolis or twist
(neck tilted to left side), 14 % had stargazing (head tilted upward) (Figure IV. 4) whereas
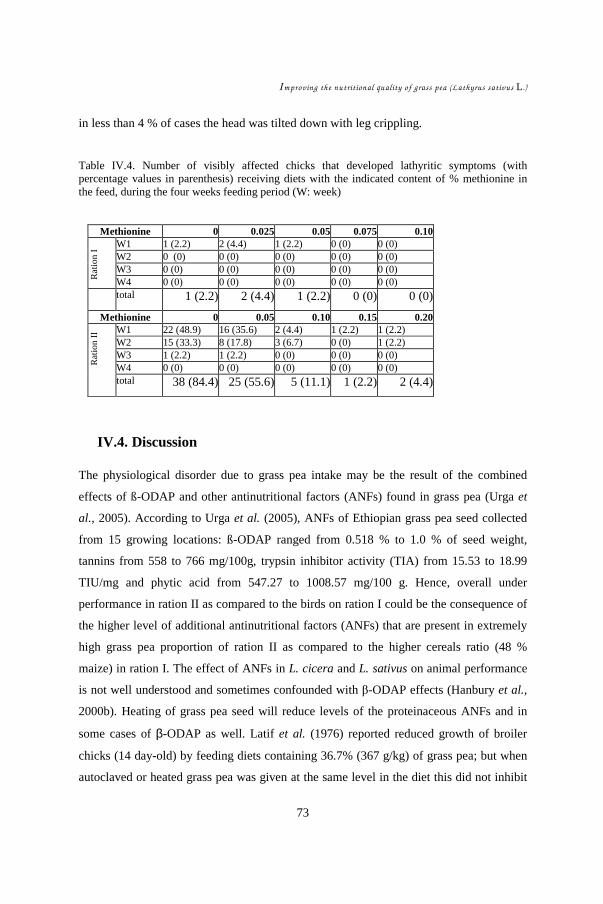
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
73
in less than 4 % of cases the head was tilted down with leg crippling.
Table IV.4. Number of visibly affected chicks that developed lathyritic symptoms (with percentage values in parenthesis) receiving diets with the indicated content of % methionine in the feed, during the four weeks feeding period (W: week)
IV.4. Discussion
The physiological disorder due to grass pea intake may be the result of the combined
effects of ß-ODAP and other antinutritional factors (ANFs) found in grass pea (Urga et
al., 2005). According to Urga et al. (2005), ANFs of Ethiopian grass pea seed collected
from 15 growing locations: ß-ODAP ranged from 0.518 % to 1.0 % of seed weight,
tannins from 558 to 766 mg/100g, trypsin inhibitor activity (TIA) from 15.53 to 18.99
TIU/mg and phytic acid from 547.27 to 1008.57 mg/100 g. Hence, overall under
performance in ration II as compared to the birds on ration I could be the consequence of
the higher level of additional antinutritional factors (ANFs) that are present in extremely
high grass pea proportion of ration II as compared to the higher cereals ratio (48 %
maize) in ration I. The effect of ANFs in L. cicera and L. sativus on animal performance
is not well understood and sometimes confounded with β-ODAP effects (Hanbury et al.,
2000b). Heating of grass pea seed will reduce levels of the proteinaceous ANFs and in
some cases of β-ODAP as well. Latif et al. (1976) reported reduced growth of broiler
chicks (14 day-old) by feeding diets containing 36.7% (367 g/kg) of grass pea; but when
autoclaved or heated grass pea was given at the same level in the diet this did not inhibit
Methionine 0 0.025 0.05 0.075 0.10W1 1 (2.2) 2 (4.4) 1 (2.2) 0 (0) 0 (0) W2 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) W3 0 (0) 0 (0) 0 (0) 0 (0) 0 (0)
Rat
ion
I
W4 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) total 1 (2.2) 2 (4.4) 1 (2.2) 0 (0) 0 (0)
Methionine 0 0.05 0.10 0.15 0.20W1 22 (48.9) 16 (35.6) 2 (4.4) 1 (2.2) 1 (2.2) W2 15 (33.3) 8 (17.8) 3 (6.7) 0 (0) 1 (2.2) W3 1 (2.2) 1 (2.2) 0 (0) 0 (0) 0 (0)
Rat
ion
II
W4 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) total 38 (84.4) 25 (55.6) 5 (11.1) 1 (2.2) 2 (4.4)

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
74
growth whereas showed the same result of a maize-soya diet. Tadelle et al. (2003) fed
broiler starters (0-29 days) with grass pea (at 35 % of the diet) heated in water at 60 oC,
75 oC, 90 oC and boiling temperature respectively, and found better growth performance
of the young chicks after treating the grass pea at increasing temperatures. Boiled grass
pea diet showed no significant difference (p< 0.05) with the diet that composed of
maize/wheat/noug cake. Low et al. (1990) reported that with 800 g/kg grass pea (80 % of
the feed) in the diet for growing chicks over a four week period, chicks’ performance
(feed intake, weight gain) was significantly poorer than that of wheat/soybean meal-based
diet (p<0.05) while it was markedly improved by dietary methionine supplementation at
0.25 % of the feed (p <0.05). Smulikowska et al. (2008) included grass pea var. Krab as
10 % of the feed for broiler chicken (age 8-40 days) and were able to show that growth
performance of the chicks did not differ from the controls that did not receive grass pea in
the diet. In this Polish experiment, birds fed with var. Krab seed had enlarged pancreases
(p< 0.05) and livers in comparison to controls. Rotter et al. (1991) evaluated low (ß-
ODAP 0.13 % of seed weight) and medium (0.22 % ß-ODAP) toxin lines of grass pea in
the diet to feed chicks and concluded that with 40 % grass pea (low or medium ODAP
lines) in the diet, the weight gain showed no significant difference (p<0.05) with the
chicks fed on a wheat-based diet. Hanbury et al. (2000b) observed a 40 % grass pea (0.27
% ß-ODAP) feed tolerance of chicks with insignificant weight reduction. Chowdhury et
al. (2005) studied the nutritional value of grass pea (0.4 % ß-ODAP) for growing and
laying pullets and concluded that inclusion of 10, 15 and 20% of grass pea in the feed,
supported growth performance in pullets at the same level or better than the control feed
(without grass pea). However, for egg-layers, grass pea in the feed was well tolerated
only up to 15 % grass pea without deleterious effects on egg production. A summary of
published feeding trials is presented in Table IV.5. Our study in ration II is the first report
for replacing grass pea as the only energy source (98.5 %) in the chick diet.
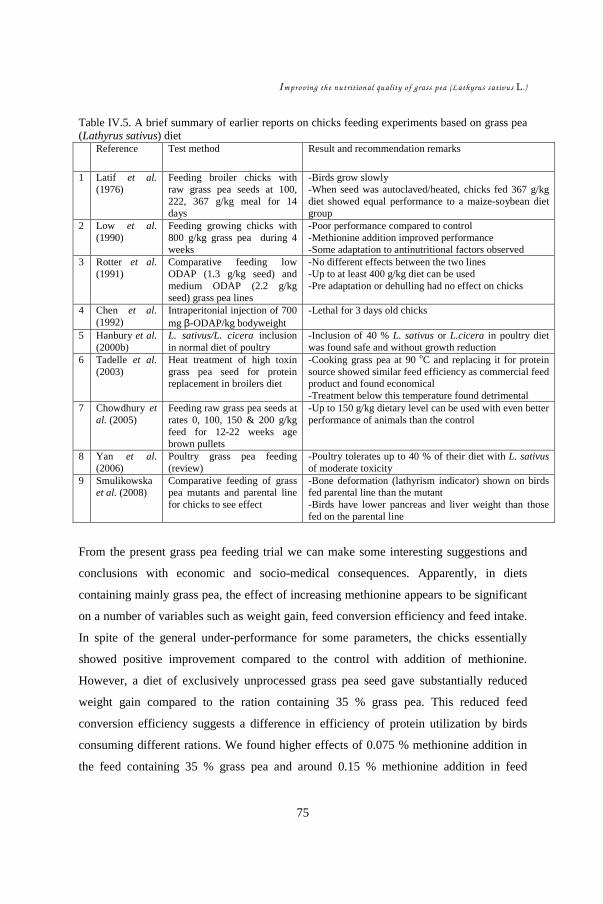
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
75
Table IV.5. A brief summary of earlier reports on chicks feeding experiments based on grass pea (Lathyrus sativus) diet Reference Test method Result and recommendation remarks
1 Latif et al. (1976)
Feeding broiler chicks with raw grass pea seeds at 100, 222, 367 g/kg meal for 14 days
-Birds grow slowly -When seed was autoclaved/heated, chicks fed 367 g/kg diet showed equal performance to a maize-soybean diet group
2 Low et al. (1990)
Feeding growing chicks with 800 g/kg grass pea during 4 weeks
-Poor performance compared to control -Methionine addition improved performance -Some adaptation to antinutritional factors observed
3 Rotter et al. (1991)
Comparative feeding low ODAP (1.3 g/kg seed) and medium ODAP (2.2 g/kg seed) grass pea lines
-No different effects between the two lines -Up to at least 400 g/kg diet can be used -Pre adaptation or dehulling had no effect on chicks
4 Chen et al. (1992)
Intraperitonial injection of 700 mg β-ODAP/kg bodyweight
-Lethal for 3 days old chicks
5 Hanbury et al. (2000b)
L. sativus/L. cicera inclusion in normal diet of poultry
-Inclusion of 40 % L. sativus or L.cicera in poultry diet was found safe and without growth reduction
6 Tadelle et al. (2003)
Heat treatment of high toxin grass pea seed for protein replacement in broilers diet
-Cooking grass pea at 90 oC and replacing it for protein source showed similar feed efficiency as commercial feed product and found economical -Treatment below this temperature found detrimental
7 Chowdhury et al. (2005)
Feeding raw grass pea seeds at rates 0, 100, 150 & 200 g/kg feed for 12-22 weeks age brown pullets
-Up to 150 g/kg dietary level can be used with even better performance of animals than the control
8 Yan et al. (2006)
Poultry grass pea feeding (review)
-Poultry tolerates up to 40 % of their diet with L. sativus of moderate toxicity
9 Smulikowska et al. (2008)
Comparative feeding of grass pea mutants and parental line for chicks to see effect
-Bone deformation (lathyrism indicator) shown on birds fed parental line than the mutant -Birds have lower pancreas and liver weight than those fed on the parental line
From the present grass pea feeding trial we can make some interesting suggestions and
conclusions with economic and socio-medical consequences. Apparently, in diets
containing mainly grass pea, the effect of increasing methionine appears to be significant
on a number of variables such as weight gain, feed conversion efficiency and feed intake.
In spite of the general under-performance for some parameters, the chicks essentially
showed positive improvement compared to the control with addition of methionine.
However, a diet of exclusively unprocessed grass pea seed gave substantially reduced
weight gain compared to the ration containing 35 % grass pea. This reduced feed
conversion efficiency suggests a difference in efficiency of protein utilization by birds
consuming different rations. We found higher effects of 0.075 % methionine addition in
the feed containing 35 % grass pea and around 0.15 % methionine addition in feed

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
76
containing grass pea as an exclusive ( 98.5 % of the feed) protein and energy source. In a
similar way, Molvig and co-workers (1997) mentioned that when a diet with lupine, that
contains ANFs, as major protein source was supplemented with methionine, significant
increases were found in growth of pigs and in efficiency of protein utilization in rats.
Because non-ruminant animals can convert methionine to cysteine, added methionine can
satisfy their total requirement for dietary sulphur containing amino acids. Al-Mayah
(2006) concluded that better immune response could be obtained with adequate
supplementation of methionine at 1 g/L in drinking water or as 0.1 % in the feed for
broiler chicks. This confirms that the compensatory action of methionine supplements for
sulphur amino acid-limited diets such as grass pea can maximize the safe utilization of
this under-utilised but protein rich legume crop (Lambein et al., 2001; Getahun et al.,
2005; Nunn et al., 2005).
The results from the two experiments with different rations containing respectively 35 %
and 98.5 % of grass pea suggest the need for optimising methionine content of feed
formulations for optimal feed conversion efficiency and economical weight gain. Result
also suggests that health feed formulations could be considered using the cheap grass pea
as ingredient.
An important observation was the increase in intake tolerance for unprocessed grass pea
containing ß-ODAP and other ANFs, with increasing methionine in the diet. Our
experiment shows that performance of animals on the exclusive Ethiopian grass pea diet
was improved substantially in food intake and in weight gain by addition of the essential
amino acid methionine. Higher intake of the high toxin grass pea with some
antinutritional factors present in raw seed could be attributed to increase in age (Low et
al., 1990; Chen et al., 1992; Tadelle et al., 2003; Yan et al., 2006) and the addition of
methionine. The added methionine might have suppressed the toxic effects of grass pea
feed. In ration II, methionine at level 0.1 % and above kept the animals safe from chronic
illness and convulsions (Table IV.4). Our results suggests that (1) methionine is an
effective antidote against grass pea toxicity; and (2) its optimisation would improve
chicks performances and thus increase economic gain of farmers.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
77
We can estimate that chicks in ration II, with highest incidence of neurological
symptoms, were using an average of about 600 mg ß-ODAP per kilogram body weight
per day. The ß-ODAP concentration in the blood could be hypothetically considered to be
the amount that would be absorbed by the body to bring about the neurological problem.
Chen and co-workers (1992) reported that intraperitoneal injection of ~700 mg/ kg of
body weights can be lethal for 3-day old chicks.
Our result is in line with earlier suggestions that methionine-rich diet reduces the toxic
action of ß-ODAP ( Getahun et al., 2003, 2005). In agreement with previous work (Mehta
et al., 1979; Chen et al., 1992; Tadelle et al., 2003) also in this study acute neurotoxicity
was more frequent in younger chicks and decreased as age increased. The older the chick,
the less susceptible it is per unit body weight to the toxic amino acids and ANFs in L.
sativus seeds. The common acute convulsive manifestation in chicks was the deformation
or bending of the neck muscle unlike the symptoms seen in human neurolathyrism.
Similar symptoms of head moving downwards or bending, and stiffening of the neck has
been reported in one-day-old chicks by either giving oral doses of β-ODAP (0.1 mmol)
(Shamim et al., 2002) or by intraperitoneal injections of 0.2 to 0.6 ml of L. sativus extract
(5 gm dry seed/ml) per chick (Roy et al., 1963). A direct comparison of toxicity and
protection between the acute reversible signs in chicks and human neurolathyrism
produced by chronic intoxication by grass pea over-consumption (or chronic depletion of
methionine) may not be possible. But the general trend of methionine as protectant
against the toxin is likely to be valid also in humans. Consumption of methionine-rich
foods as protection against the neuro-excitatory action of ß-ODAP and against the
oxidative stress produced by its biochemical and physiological activities has been
proposed before (Lambein et al., 2001; Kusama-Eguchi et al., 2003; Getahun et al., 2005;
Nunn et al., 2005) and is supported by our findings.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
78

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
79
CHAPTER V
Effect of methionine supplement on serum amino acids status
and trace elements level of broiler chicks fed with grass pea
based starter ration

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
80
Healthy chick after feeding on grass pea diet

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
81
CHAPTER V
Effect of methionine supplement on serum amino acids status and trace
elements level of broiler chicks fed with grass pea based starter diet
Abstract
Starter feeding experiments with broiler chicks using grass pea (Lathyrus sativus L.)
supplemented with methionine were undertaken for 4 weeks during 2005 at DZARC,
Ethiopia to (1) assess toxicity by analysing their serum and to (2) assess the evolution of
selected trace elements in serum. The objective was to study the effect of increasing
methionine supplements on grass pea toxicity. Four hundred fifty broiler chicks were
divided into two experimental sets or rations formulations containing (a) 35 % (ration I)
or (b) 98.5 % (ration II) raw grass pea. Each ration include control and four treatments
with methionine (0.025, 0.05, 0.075 and 0.1 % of the feed in ration I; and 0.05, 0.1, 0.15
and 0.2 % in ration II). Blood serum of the chicks of different treatments was collected
each week and analysed for its free amino acids and trace elements content. With addition
of methionine in the feed, some amino acids in the serum showed an increase while
others showed a decrease compared to control (without methionine supplement). ß-
ODAP level in chick serum decreased progressively with increasing methionine
supplement in the feed of ration I. The levels of β-ODAP, homoarginine (Har), arginine
and lysine were much higher in chick serum of ration II than that of ration I due to the
high proportion of grass pea in feed ration II. Trace elements including iron, copper, zinc
and manganese showed different evolution with increasing methionine supplement and
with increasing age. The content of Cu2+ and Fe2+ increased more than double while Zn2+
and Mn2+ showed no difference with control at the end of three weeks. These preliminary
results suggest that methionine supplement in the feed influenced chick’s amino acids
metabolism and trace elements level and this can be important as protective element in a
grass pea based diet.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
82
V.1. Introduction
In Ethiopia, one of the important purposes of grass pea (Lathyrus sativus L) cultivation is
its use for animal feed (Fikre, 2007; Dadi et al., 2003). Grass pea is a cheap source of
protein and smallholder farmers usually use feed rations containing grass pea for
fattening animals.
Neurological symptoms can develop particularly in monogastric animals (Yan et al.,
2006) after prolonged consumption of grass pea. This toxicity is blamed on the presence
of a non-protein amino acid β-N-oxalyl-L-α,β-diaminopropionic acid or ß-ODAP
(Spencer et al., 1986). However, the acute and reversible neurological signs in young
chicks is totally different from the aetiology of human neurolathyrism commonly
manifested by irreversible symptoms of spastic paraparesis in legs. After consumption of
grass pea or injection of ß-ODAP, young chicks show specific but reversible neurological
signs such as torticolis (bending of the neck sidewards) or star gazing (bending of the
neck backward).
For primates on a balanced diet, massive doses of ß-ODAP are needed during an
extended period to develop central motor neuron and hind limb deficiency (Spencer et al.,
1986). Rao (2001) indicated that there exist extreme inter-species and intra-species
differences in susceptibility to ß-ODAP that may be ascribed to metabolic differences,
and he reported that urine samples collected from humans following a Lathyrus meal
showed very little urinary excretion of ß-ODAP (less than 2-5% of what is injested). Kuo
and co-workers (2007) demonstrated that the free amino acids aspartic acid, glycine, β-
aminoisobutyric acid, arginine, α-aminoadipic acid and phenylalanine were significantly
higher (p<0.05) in urine from patients with neurolathyrism than in urine from healthy
controls while β-ODAP was not detected because the consumption of grass pea had been
discontinued at the time of sampling. Nunn and co-workers (2005) found that
consumption of grass pea seed resulted in a significant and persistent decrease in plasma
methionine by about 50 %. A reduction in plasma methionine concentration would cause

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
83
a reduced methionine flux into the central nervous system. As a consequence, cells may
be sensitised to additional oxidative stresses. This oxidative stress and probably the
neuro-toxicity is aggravated by the inherent deficiency of essential sulphur amino acids
methionine and cysteine in grass pea (Nunn et al., 2005). Al-Mayah (2006) reported that
in soybean based feed supplementation of DL-methionine at 1 g/L of drinking water or at
1 g/kg of chick’s feed results in increment in a number of blood parameters: red blood
cell counts, mean cell volume, total serum and liver protein, albumin and globulin. He
suggested that better immune response of chicks could be obtained with adequate
supplementation of methionine in the poultry feed.
The analyses in this study have been conducted with the view that the physical responses
of animals on grass pea rations, low in sulphur amino acids and containing the neuro-
excitatory ß-ODAP, might reflect changes in the serum amino acids and trace elements
comparable to humans on a grass pea diet. Addition of methionine to grass pea rations
may have a modulating effect on these concentration changes and on other physiological
parameters related to redox homeostasis. Hence, the objective of this analytical part of the
feeding study with grass pea was to investigate and to compare the amino acids, β-ODAP
and relevant trace elements levels in the serum of chicks during the first three weeks
period.
V.2. Materials and Methods The study area, the starter broiler chicks, their management and two feeding rations used
(with 35 % and 98.5 % grass pea respectivelly) were as mentioned in Chapter IV.
Serum sampling: a specialized veterinarian collected serum from the wing vein using a 5
cc disposable syringe. This was done for each replication of the treatments on a weekly
basis. However, in the first week, at most four chicks per replication were sacrificed to
get enough volume of blood serum for the analysis. Samples were immediately
centrifuged (5000 rpm) for five minutes and the plasma supernatants was decanted in
standard tubes for freezing until the lab analyses for amino acids and trace elements.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
84
Trace elements analysis: Trace elements were sequentially extracted by nitric acid
(HNO3) and hydrogen peroxide (H2O2) (Quevauviller et al.,1996). Cu2+ and Mn2+ were
determined with Graphite Furnace Atomic Absorption Spectrophotometry (GFAAS)
while Fe2+ and Zn2+ were determined with Inductively Coupled Plasma mass
spectrometry (ICP). These analyses were performed by the Department of Analytical and
Physical Chemistry, Faculty of Bioscience Engineering, Ghent University.
Free amino acid analysis:
Serum samples: 100 µl of serum was mixed with 300 µl of 95 % ethanol. In each
sample, 5 µl of DL-allylglycine (100 µmol/ml, Sigma) was added as internal standard.
The mixture was allowed to stand at 4 °C overnight before centrifugation at 12,000 rpm
for 10 min in an Eppendorf centrifuge (Hawksley MBC). The supernatant was
concentrated in an Eppendorf Vacufuge concentrator 5301 (Germany) at 45 °C under
vacuum. 100 µl ion-free water was added to the concentrate, stirred, centrifuged and the
supernatant collected and dried for PITC derivatisation.
PITC (Phenyl isothiocyanate) derivatisation: in each dried sample, 20 µl of coupling
buffer (methanol/water/triethylamine: 2/2/1 by volume) was added, mixed and dried in
the concentrator. Finally, 30 µl of the PITC reagent (methanol/water/triethylamine/phenyl
isothiocyanate: 7/1/1/1 by volume) was added and reacted at room temperature for 20
min before concentrating to dryness.
To each PITC-derivatised sample, 500 µl of buffer A (0.1 M ammonium acetate, pH 6.5)
was added, mixed well and centrifuged. The supernatant was filtered through a Millipore
Millex filter (0.45 µm) and 20 µl aliquot was injected in the HPLC for analysis. A
standard amino acid mixture (AA-S-18, Sigma), L-homoarginine (99+ %, Janssen
Chimica) and synthetic ß-ODAP (a gift from Dr. Rao S.L.N, India) were also derivatised
and prepared as above and injected in HPLC as standard.
High-performance liquid chromatography (HPLC) for amino acid analysis: a Waters
625 LC system with Waters 991 photodiode array detector was used as previously

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
85
reported (Kuo et al., 2003). A gradient system with buffer A (0.1 M NH4OAc, pH 6.5)
and buffer B (0.1 M NH4OAc, containing acetonitrile and MeOH; 44/46/10 by volume,
pH 6.5) with flow rate of 1 ml/min was used for the separation of amino acids during 50
min. An Alltima C18 column (250x4.6 mm I.D., 5 µm particle size, Alltech, USA) was
used with column temperature at 43 °C during analysis. The absorbance at 254 nm was
used for calculations. Results were analysed by Millennium software (Waters, version
1.10).
Statistical analysis
Amino acids detected in chick serum of the first week for ration I and the three
consecutive weeks for ration II were analysed with ANOVA using SPSS 12 software.
Statistical significance of differences among means was determined using Tukey
Honestly Significant Difference set at 95 % confidence level. Amino acid concentrations
(µmol/100 ml) in serum were presented with their respective standard deviation.
Pearson’s correlation coefficients were calculated for micronutrients, β-ODAP, body
weight and FCE.
V.3. Results Free amino acids in the serum
The free amino acid profiles of chicken serum fed with grass pea ration I (1st week,
samples of second and third weeks could not be analysed) and II (1st, 2nd and 3rd week)
are given in Tables V.1 and V.2 respectively. A number of amino acids showed increase
or decrease along with increasing methionine levels. In ration I, β-ODAP, glutamic acid
(Glu) and homoarginine (Har) showed a decreasing trend with methionine supplements
while arginine (Arg), tyrosine (Tyr) and leucine (Leu) increased compared to the control
without methionine addition (Table V.1). Interestingly, with the increase in methionine
concentration in serum, the corresponding ß-ODAP level in the serum decreased
progressively in the ration I experiment.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
86
Table V.1: Concentrations of free amino acids (µmol/100 ml) in chick serum (mean ± SD, average of 1st week, N = 4) fed with 35 % grass pea (ration I) with different levels of DL-methionine addition (0.025, 0.050, 0.075 and 0.100 % of feed)
0.0 0.025 0.05 0.075 0.1
Asp 10.4±5.8b 16.5±5.0a 10.8±2.8b 13.9±6.6ab 16.8±4.7a
ß-ODAP 16.5±7.3a 8.4±7.7b 7.2±6.2b 7.9±6.8b 4.9±3.0b
Glu 42.8±21.4a 32.3±7.2b 27.5±9.1c 30.0±9.8bc 31.4±5.9bc
α-AAA 8.8±5.4c 7.7±3.1c 14.0±7.1b 13.6±11.2b 22.9±12.8a
Ser 76.2±43b 76.4±29.8b 57.9±18.1c 56.1±25.0c 97.2±44.4a
Gly 75.7±15.9a 62.0±16.9bc 72.0±32.1a 61.6±23.2c 66.5±32.9b
Gln 65.3±46.6b 103.8±29.0a 54.3±25.1c 70.8±35.0b 59.8±29.3c
His 9.5±3.7a 10.1±2.8a 9.5±4.6a 9.2±1.4a 12.2±5.6a
Tau 33.2±17b 25.6±5.6c 27.9±15.6c 33.7±13.6b 70.0±31.8a
Arg 15.4±10.4c 38.7±25.5a 25.6±8.5b 23.7±12.5b 40.7±23.6a
Thr 46.2±35.6a 33.8±18.9b 41.4±11.7a 23.1±19.1c 25.7±16.6c
Ala 55.2±32.6c 55.2±29.8c 54.7±33.5c 73.1±26.0a 66.8±19.3b
Har 139.3±97.8a 66.4±45d 82.3±51.6b 74.7±42.6c 81.2±28.7b
Pro 38.5±24.1a 39.7±17.2a 36.8±21.2a 34.8±18.7b 38.4±20.1a
Tyr 6.9±4.2d 22.1±15.8b 19.7±15.7b 31.1±20.1a 13.5±6.7c
Val 16.7±7.3c 25.7±8.0ab 24.9±8.5b 16.5±9.2c 30.1±14.8a
Met 4.6±2.6b 5.9±1.7ab 3.8±1.8b 7.5±4.4a 6.3±1.3a
Ile 7.1±5.2b 10.5±3.5b 8.8±3.3b 7.7±3.5b 15.1±6.0a
Leu 11.0±5.2b 16.2±2.25a 16.1±3.9a 17.2±4.0a 17.2±10.7a
Phe 14.6±13.5a 14.7±9.1a 10.8±6.8ab 9.0±3.8b 8.9±6.6b
Trp 39.5±25.3a 18.3±10.8bc 16.2±4.8c 21.4±14.1b 37.2±22.1a
Lys 41.3±33.5a 41.5±31.5a 27.3±21.5c 35.5±23.0b 29.3±8.2c
α-AAA: α-amino adipic acid; Har: homoarginine
Values in a row followed by the same letters are not significantly different (P<0.05).

Impr
ovin
g th
e nu
trit
iona
l qu
alit
y of
gra
ss p
ea (
Lat
hyru
s sa
tivu
s L
.)
87
Tab
le V
.2:
Con
cent
ratio
ns o
f fr
ee a
min
o ac
ids
(µm
ol/1
00 m
l) in
chi
ck s
erum
(m
ean
± S
D,
aver
age
of 1
st,
2nd,
3rd w
eeks
, N
=4,
2 a
nd 2
for
w
eeks
1,
2 an
d 3
resp
ectiv
ely)
fed
with
98.
5% g
rass
pea
(ra
tio
n II)
with
diff
eren
t le
vel
of D
L-m
ethi
onin
e ad
ditio
n (0
.05,
0.1
0, 0
.15,
and
0.
20 %
of f
eed)
V
alue
s in
a r
ow fo
llow
ed b
y th
e sa
me
lett
ers
are
not s
igni
fica
ntly
dif
fere
nt (
P<
0.05
).
W
eek
1 W
eek
2 W
eek
3
0.
0 0.
05
0.1
0.15
0.
2 0.
0 0.
05
0.1
0.15
0.
2 0.
0 0.
05
0.1
0.15
0.
2
Asp
17
.7±0
.8b 14
.6±7
.3c 22
.4±1
2.7a
22.3
±8.0a
21.2
±8.3a
11.4
±1.8c
19.8
±2.6b
25.1
±3.4a
23.3
±1.2a
b 15
.4±1
.9bc
14.1
±2.8b
16.1
±11.
2b 18
.6±9
.2b 18
.6±4
.5b 25
.2±1
.7a
β-O
DA
P 22
.2±6
.2c 27
.8±2
1.4b
28.0
±7.3b
35.0
±27.
9a 29
.2±8
.2b 12
.3±1
.3c 12
.4±0
.4c 41
.3±4
.9a 18
.2±0
.3b 12
.4±3
.4c 14
.7±4
.4b 12
.1±0
.5b 15
.8±6
.0b 27
.8±1
5.9a
14.6
±10.
4b
Glu
36
.9±9
.6b 30
.9±6
.0c 44
.1±7
.6ab
23.6
±2.4d
47.6
±7.4a
75.2
±7.3a
42.9
±5.5b
36.8
±4.4c
35.3
±3.9cd
30
.2±2
.3d 35
.0±9
.5ab
32.0
±8.3b
42.3
±0.8a
38.5
±12.
5a 31
.5±6
.6b
α-A
AA
12
.5±8
.8a
8.7±
0.1a
b 8.
0±4.
9b 7.
7±0.
4b 11
.6±2
.1a 11
.1±1
.5b 9.
3±0.
5b 9.
0±1.
3b 10
.4±1
.4b 19
.3±0
.9a 11
.2±1
.3ab
10.4
±2.5a
b 10
.8±0
.8ab
8.3±
1.1b
14.3
±2.1a
Ser
59
.3±4
.5b 56
.3±1
8.1b
71.2
±5.3a
44.2
±4.6c
74.2
±25.
5a 32
.2±3
.6c 62
.2±7
.0b 60
.3±7
.4b 38
.7±2
.9c 75
.2±6
.2a 52
.3±7
.4b 50
.6±1
4.4b
59.4
±2.6b
50.1
±19.
2b 74
.3±1
0.3a
Gly
67
.2±3
.5b 57
.9±8
.1c 67
.5±8
.1b 67
.1±3
.3b 92
.9±2
.9a 73
.1±1
1.1a
b 78
.7±6
.9a 70
.1±6
.7b 55
.9±3
.5c 64
.7±3
.9bc
59.3
±1.1bc
51
.9±4
.5c 64
.6±6
.5b 62
.6±0
.4b 76
.4±8
.6a
Gln
50
.1±1
6.8d
80.3
±18.
8a 58
.0±0
.1c 66
.4±1
0.3b
58.9
±21.
2c 62
.6±2
5.2b
35.7
±4.5d
55.3
±6.8c
88.6
±6.4a
52.6
±4.1c
33.5
±2.0d
73.1
±17.
4a 43
.8±1
2.7c
67.2
±18.
4ab
65.0
±10.
6b
His
21
.6±1
0.3a
11.3
±2.7bc
8.
1±3.
1c 9.
7±2.
3c 13
.9±2
.9b 12
.8±1
.6a 10
.2±2
.8a 11
.2±2
.3a 8.
2±0.
3a 9.
9±0.
1a 5.
9±0.
7b 9.
8±1.
9a 7.
7±3.
1ab
10.3
±3.5a
12.3
±0.2a
Ta
u 49
.5±4
5.9a
45.2
±39.
2b 51
.1±3
9.1a
14.4
±8.6c
43.5
±8.4b
65.7
±6.1a
36.4
±8.8b
18.1
±2.8c
22.4
±1.1c
39.1
±1.2b
31.3
±23.
0ab
28.3
±12.
6b 35
.0±1
7.8a
40.1
±27.
2a 20
.0±1
3.7c
Arg
75
.1±4
5.2a
43.8
±8.2d
57.6
±14.
9c 41
.6±1
4.5d
63.7
±28.
0b 59
.0±6
.6a 41
.1±3
.7b 52
.0±5
.9ab
50.3
±3.7a
b 46
.5±4
.1b 40
.5±3
.5ab
38.4
±3.7b
50.3
±17.
4a 47
.9±3
.6a 47
.3±1
2.4a
Thr
70
.6±5
0.0a
52.1
±26.
2c 50
.6±2
1.2c
61.6
±4.7b
61.3
±26.
8b 10
0.9±
14.4a
88.0
±10.
3b 89
.3±1
5.6b
80.2
±7.2b
82.7
±10.
8bc
31.8
±22.
9e 86
.9±2
8.2b
44.8
±4.7d
67.6
±24.
4c 11
8.0±
23.1a
Ala
70
.6±5
1.5c
82.2
±16.
9b 96
.4±9
.8c 85
.7±2
.2b 93
.2±2
.8c 10
6.0±
15.9a
90.1
±14.
2b 10
5.1±
18.2a
76.8
±7.6c
78.8
±6.7c
56.6
±8.7c
83.7
±32.
3a 67
.9±2
4.6b
85.8
±19.
7a 84
.3±1
0.5a
Har
14
6.4±
5.2a
118.
1±12
.1b 13
8.9±
50.7a
143.
6±28
.6a 14
7.1±
21.4a
157.
5±18
.4b 11
3.3±
22.5c
178.
1±22
.1a 15
6.8±
13.4b
179.
8±16
.7a 59
.1±1
6.1c
81.1
±39.
6c 13
2.4±
43.4b
144.
0±16
.2ab
149.
8±52
.5a
Pro
48
.5±3
.3b 38
.5±7
.9c 50
.4±1
7.7b
43.7
±4.4c
56.3
±7.0a
48.9
±6.1b
52.5
±4.7b
63.1
±7.5a
47.8
±4.0b
52.1
±1.6b
42.4
±5.8b
65.4
±6.1a
46.6
±11.
7b 45
.4±4
.0b 59
.2±5
.4a
Tyr
14
.1±6
.4b 10
.0±7
.2c 21
.0±1
2.9a
12.9
±0.1bc
9.
9±2.
2c 13
.8±2
.2b 19
.8±1
5.3a
b 19
.9±1
2.9a
b 18
.9±1
.3b 24
.5±5
.7a 12
.9±0
.8c 11
.0±8
.5c 12
.8±0
.6c 18
.1±7
.2b 25
.2±5
.7a
Val
55
.8±6
.5a 32
.1±2
4.3b
49.8
±28.
6ab
50.0
±10.
6ab
52.7
±22.
0a 20
.9±3
.3d 44
.4±6
.6b 55
.8±1
0.2a
46.9
±4.8b
34.5
±2.1c
38.2
±17.
6b 44
.7±4
.5ab
44.4
±26.
4ab
48.4
±0.9a
53.0
±0.3a
Met
3.
8±1.
4b 4.
8±1.
2ab
6.4±
4.8a
4.5±
2.3a
b 4.
1±2.
3ab
2.0±
1.5b
3.2±
0.8b
6.7±
1.1a
4.6±
0.2b
8.3±
0.6a
3.2±
0.9b
7.1±
0.9a
5.8±
4.5a
5.5±
1.1a
6.3±
1.3a
Ile
18.7
±3.9b
18.2
±1.2b
23.8
±4.5a
17.9
±5.5bc
14
.3±1
0.1c
8.2±
5.7c
13.7
±1.3b
20.4
±2.5a
19.1
±0.8a
15.5
±4.6b
14.9
±6.8b
17.1
±1.1a
b 17
.5±1
0.4a
b 20
.1±0
.8a 20
.1±1
.3a
Leu
22.8
±2.1a
b 23
.9±0
.2a 25
.2±1
1.4a
22.1
±8.1a
b 20
.3±1
1.9b
13.5
±3.5c
18.9
±1.9
bc 26
.6±2
.5a 22
.6±1
.3ab
19.7
±13.
6b 18
.2±7
.7b 22
.1±1
.3ab
21.7
±12.
7ab
25.5
±0.7a
27.7
±1.5a
Phe
39
.1±2
7.6b
16.0
±12.
0c 44
.0±3
1.3a
37.0
±26.
9b 48
.0±2
4.1a
25.2
±22.
7c 49
.0±3
5.2a
24.0
±19.
9c 37
.7±2
.7b 18
.0±8
.9d 28
.1±1
7.3bc
20
.6±8
.1c 21
.6±1
2.2c
29.8
±4.5b
40.0
±28.
5a
Trp
50
.3±2
5.8b
39.0
±18.
5c 40
.0±3
5.4c
46.2
±1.7b
57.8
±67.
0a 17
.5±1
2.4c
41.7
±16.
6a 27
.9±1
9.4b
38.2
±16.
0a 42
.1±2
3.0a
42.3
±33.
5b 58
.5±1
8.0a
18.0
±13.
2c 36
.0±1
2.4bc
50
.0±1
3.0a
b
Lys
94.3
±6.8b
73.1
±6.6c
107.
7±37
.5a 93
.4±2
.4b 10
1.1±
34.2a
90.3
±63.
9b 68
.7±7
.2c 84
.9±1
1.5b
87.9
±10.
0b 10
8.1±
14.1a
67.2
±32.
8c 85
.0±3
5.7b
80.8
±52.
0b 87
.5±1
3.5b
112.
3±21
.7a

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
88
In ration II which contains 98.5 % grass pea, ß-ODAP level in the chick’s serum
increased with methionine supplements at the end of the first week while it decreased
with time or with age of the chicks later on (Table V.2). After the first week, a general
change trend from lower to higher levels along the methionine addition was found for β-
ODAP, aspartic acid (Asp) and glutamine (Gln) while histidine (His), arginine (Arg) and
threonine (Thr) showed a decreasing trend compared to control. The β-ODAP level was
high in the first week with methionine addition and gradually decreased after the second
week, irrespective of methionine addition. Obviously, chickens with 98.5 % grass pea in
the feed required a period of several weeks to adapt to such diet. Homoarginine (Har), the
main amino acid in grass pea seeds (see chapter II), was the most abundant amino acid
particularly in ration II serums.
Trace elements in the serum
Among the trace elements we analysed in the chick serum, Fe2+ showed the highest
concentration (2.70-19.0 mg/kg) followed by Zn2+ (1.57-4.04 mg/kg), Cu2+ (0.16-0.62
mg/kg) and Mn2+ (0.027-0.186 mg/kg). The concentrations of trace elements in chick
serum from ration II experiment seem to be affected by time (age) and methionine
concentration in the feed (Figure V.1 and Table V.3) although there is no clear overall
trend. Most changes were found at higher methionine supplement from 0.1 % to 0.2 %
between week 1 and week 3. At the end of the 3rd weeks, Cu2+ and Fe2+ increased more
than double in the chick serum while Zn2+ and Mn2+ had less change compared to the first
week.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
89
Figure V.1. Contents of trace elements Cu2+, Zn2+, Mn2+ and Fe2+ (mg/kg) detected in chick serum with supplement of methionine in the feed (0, 0.05, 0.10, 0.15 and 0.20 %) after week 1 (W1), week 2 (W2) and week 3 (W3) in grass pea ration II fed broilers
Table V.3. Level of trace elements (mg/kg) detected (one sample) in chick serum in ration II with increasing methionine supplements at the end of 1st, 2nd and 3rd weeks
Met supplement 0 0.05 % 0.10 % 0.15 % 0.20 % Fe2+ Week 1 3.000 6.000 5.400 9.500 3.890 Week 2 7.620 7.790 8.200 19.000 14.300 Week 3 10.700 7.970 10.400 9.500 15.700 Mn2+ Week 1 0.027 0.052 0.042 0.170 0.047 Week 2 0.035 0.029 0.025 0.047 0.072 Week 3 0.033 0.031 0.043 0.060 0.046 Zn2+ Week 1 1.740 2.300 1.990 1.900 2.220 Week 2 2.120 3.070 4.040 2.850 2.830 Week 3 2.070 1.860 2.380 2.620 2.360 Cu2+ Week 1 0.300 0.220 0.170 0.200 0.160 Week 2 0.230 0.220 0.130 0.210 0.200 Week 3 0.370 0.290 0.380 0.490 0.400
00.020.040.060.08
0.10.120.140.160.18
0 0.05 0.1 0.15 0.2
Mn W 1 Mn W 2 Mn W 3
00.05
0.10.15
0.20.25
0.30.35
0.40.45
0.50.55
0 0.05 0.1 0.15 0.2
Cu W 1 Cu W 2 Cu W 3
02468
1 01 21 41 61 82 0
0 0 .0 5 0 .1 0 .1 5 0 .2
Fe W 1 Fe W 2 Fe W 3
00.5
11.5
22.5
33.5
44.5
0 0.05 0.1 0.15 0.2
Z n W 1 Z n W 2 Z n W 3
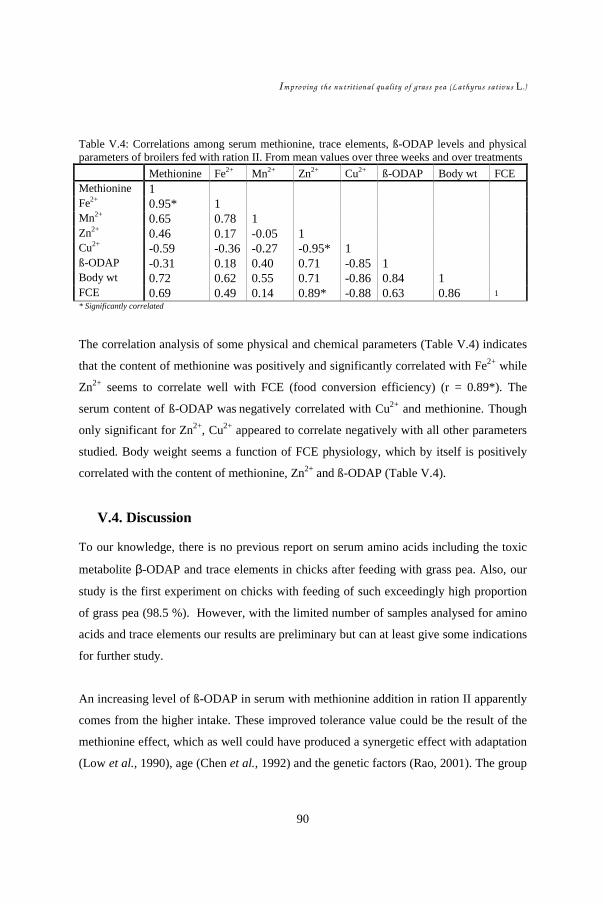
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
90
Table V.4: Correlations among serum methionine, trace elements, ß-ODAP levels and physical parameters of broilers fed with ration II. From mean values over three weeks and over treatments Methionine Fe2+ Mn2+ Zn2+ Cu2+ ß-ODAP Body wt FCE Methionine 1
Fe2+ 0.95* 1
Mn2+ 0.65 0.78 1
Zn2+ 0.46 0.17 -0.05 1
Cu2+ -0.59 -0.36 -0.27 -0.95* 1
ß-ODAP -0.31 0.18 0.40 0.71 -0.85 1
Body wt 0.72 0.62 0.55 0.71 -0.86 0.84 1
FCE 0.69 0.49 0.14 0.89* -0.88 0.63 0.86 1
* Significantly correlated
The correlation analysis of some physical and chemical parameters (Table V.4) indicates
that the content of methionine was positively and significantly correlated with Fe2+ while
Zn2+ seems to correlate well with FCE (food conversion efficiency) (r = 0.89*). The
serum content of ß-ODAP was negatively correlated with Cu2+ and methionine. Though
only significant for Zn2+, Cu2+ appeared to correlate negatively with all other parameters
studied. Body weight seems a function of FCE physiology, which by itself is positively
correlated with the content of methionine, Zn2+ and ß-ODAP (Table V.4).
V.4. Discussion To our knowledge, there is no previous report on serum amino acids including the toxic
metabolite β-ODAP and trace elements in chicks after feeding with grass pea. Also, our
study is the first experiment on chicks with feeding of such exceedingly high proportion
of grass pea (98.5 %). However, with the limited number of samples analysed for amino
acids and trace elements our results are preliminary but can at least give some indications
for further study.
An increasing level of ß-ODAP in serum with methionine addition in ration II apparently
comes from the higher intake. These improved tolerance value could be the result of the
methionine effect, which as well could have produced a synergetic effect with adaptation
(Low et al., 1990), age (Chen et al., 1992) and the genetic factors (Rao, 2001). The group

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
91
of chicks on ration II, having the highest incidence of neurological symptoms, were
taking on average about 600 mg ß-ODAP per kilogram body weight per day of the
feeding period. The ß-ODAP concentration level in the blood could reflect the amount
absorbed by the body. In comparison, Chen and co-workers (1992) reported that
intraperitoneal injection of ~700 mg/kg body weight can be lethal for 3-day old chicks.
A number of protein and non-protein amino acids assayed in serum of chicks consuming
grass pea feed showed differences between the control and methionine-added feeds.
However, the changes were neither unidirectional nor consistent for all of them. As
compared to controls, several amino acids showed an increase that may be indicating the
improvement in the animal’s physiological performances in response to methionine
addition. The much higher concentration of lysine and arginine found in serum of chicks
on ration II than that in chicks on ration I might be related to the higher intake of
homoarginine in ration II. Homoarginine can be metabolised into lysine and can also
replace arginine in some metabolic reactions. Homoarginine has been reported to
significantly depress feed intake and also to influence basic tissue amino acids
concentration and arginase activity in chickens (Angkanaporn et al., 1997). Har was also
reported to compete with lysine for transport across the blood brain barrier in rats (Tews
and Harper, 1986). The later authors found that feeding Har to the rats lowered the
concentration of lysine, ornithine and arginine in the brains, while glycine concentration
increased.
Consumption of methionine-rich foods as protection against the neuro-excitatory action
of ß-ODAP and the oxidative stress produced by its biochemical and physiological
activities has been proposed before (Lambein et al., 2001; Kusama-Eguchi et al., 2003;
Getahun et al., 2005; Nunn et al., 2005) and is supported by our findings. However,
additional experiments are needed to obtain statistically significant results.
Of all micronutrients determined in the serum samples, Fe2+ was found at the highest and
Mn2+ at the lowest concentrations. High Fe2+ intake might have an aggravating effect in
neurodegeneration as reported by Jellinger (1999). Cu2+ was reported to be significantly

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
92
higher (p < 0.05) in the urine of neurolathyrism patients (0.266 ± 0.117 mg/kg) compared
to healthy controls (0.042 ± 0.009 mg/kg) (Kuo et al., 2007). High copper content in
grass pea seeds compared to that in cereal grains like wheat and barley (Kuo et al., 2007)
might also lead to the higher concentrations of Cu2+ in chick serum after being fed
exclusively with grass pea in ration II. In general, however, the micronutrients level
detected in the serum did not show a clear trend with time and with the concentration of
methionine in the feed. Since only one serum sample of each treatment was analysed this
result should be considered preliminary. More data are needed for more clearer
conclusions.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
93
CHAPTER VI
Identification of gamma irradiation derived mutant lines of
Lathyrus sativus for higher methionine level

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
94
Putative mutant of L. sativus with higher methionine in the seed

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
95
CHAPTER VI
Identification of gamma irradiation derived mutant lines of Lathyrus
sativus selected for higher methionine level
Abstract
Methionine is a limiting essential amino acid in legumes. Methionine supplement was
suggested to protect against neurolathyrism caused by Lathyrus sativus (grass pea) based
diets. Seeds of Lathyrus sativus from different origins were irradiated using 40 KRs
gamma cell irradiation at DZARC, Ethiopia during 2005. Procedures to identify
methionine-overproducing mutants by phenotypic performance analysis were adapted
from similar work on soybean (Glycine max). Common phenotypic features of mutants in
the field include plant size reduction, reduction of branching, colour intensity, leaf
deformation (curling), infertility and absence of flower setting. Mutagenic events were
estimated at a rate of 1-3 per thousand irradiated seeds. Three distinct phenotype mutants
meeting the typical changes of phenotypic features described for soybean mutants with
increased methionine were identified, propagated and further characterized. They all have
glittering deep green leaves, stem and pods, a relatively smaller plant size when
compared to the parent. It gave samaller but a sensible yield and seed size comparable to
the parental line (cv. Debre Zeit). Analysis of seeds revealed that the putative mutants
have about 60 % higher methionine levels compared to parental controls.
VI.1. Introduction
Lathyrus sativus L. (grass pea) is an environmentally hardy multipurpose legume
growing in many parts of the world since almost as long as the existence of agriculture.
Grass pea is endowed with environmental hardiness that ensures production even when
other crops fail. It is responsible for the survival of millions of subsistence farmers during
drought and famine. There is a strong positive association between drought stress and the

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
96
biosynthesis of a neurotoxic metabolite, β-N-oxalyl-L-α,β-diaminopropionic acid (β-
ODAP). Grass pea is the only legume used as a staple food when other crops fail.
Ethiopia is the largest producer (85 %) and consumer of grass pea in Africa and is also
the most affected by the neurotoxicity it induces (Fikre, 2007). Grass pea is a good source
of protein (27 - 32 %) (Urga et al., 2005) but this protein is limited in the essential amino
acids methionine and cysteine (Kuo et al., 1995), which counts only for a quarter of the
amount in standard egg proteins (Coates et al., 1985). It was mentioned that developing
high sulphur-containing protein soybean lines could save huge financial resources
commonly used to purchase synthetic methionine added to feed in the livestock industry
(Imsande, 2001).
Neurolathyrism is an irreversible crippling disease resulting from prolonged grass pea
overconsumption. The suggested causative molecule β-ODAP is a free amino acid and its
biosynthesis is affected both by genetic and environmental factors. The neurotoxicity
mechanism of β-ODAP is counteracted by adding methionine-rich cereals or condiments
(Getahun et al., 2005) that make a better balanced diet (Molvig et al., 1997). Methionine
(like cysteine) functions as an antioxidant and is a key component for the regulation of
cellular redox homeostasis. Methionine is readily oxidised to methionine sulfoxide by
reactive oxygen species (ROS). The oxidation of methionine residues exposed at the
surface of proteins serves to protect other functionally essential residues from oxidative
damage. Methionine sulfoxide reductase enzymes have the potential to reduce those
residues back to methionine, increasing the scavenging efficiency of the system.
Interconversion of methionine and methionine sulfoxide can regulate the biological
activity of proteins through alteration of the catalytic efficiency and modulation of the
hydrophobicity of the protein (Levine et al., 2000).
Cysteine is involved in methionine biosynthesis in plants and its internal concentration
may contribute to the regulation of methionine biosynthesis (Kim et al., 1999). It was
reported that in soybean the internal concentration of O-acetyl serine (OAS), a precursor
of cysteine, regulates the relative abundance of the β-subunit of β-conglycinin, a soybean
storage protein that is low in sulphur amino acid content (Kim et al., 1999). Rosa and co-

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
97
workers (2000) investigated the storage proteins from L. sativus. These represent 20 %
of the seed dry weight and are composed of globulins and albumins. A single, 24 kDa
polypeptide comprises more than half of the protein present in the albumin fraction. The
globulins may be fractionated into -lathyrin, β-lathyrin, and -lathyrin components. Two
major polypeptides are glycosylated and exhibit structural similarity with β-conglutin
from Lupinus albus. One of these is a lectin. The amino acid composition of these storage
proteins is not known. A strategically located mutation that would favour storage proteins
with higher methionine or in the genes encoding either cystathionine γ-synthase or serine
acetyl-transferase may promote higher methionine production and thus improve the
sulphur amino acid deficient grass pea. The aim of this study was to isolate methionine-
enriched grass pea mutant phenotypes by γ-irradiation of seeds and screening the plants
grown from the irradiated seeds for higher methionine content.
VI.2. Materials and methods
Gamma irradiation
A total of 17,800 seeds of Lathyrus sativus from different origins were gamma irradiated
at 40 kilorads at DZARC, Ethiopia in 2005. The effectiveness of the irradiation dose was
established from previous similar work experience in the Debre Zeit Agricultural
Research Center. The seeds used were from the collections of IPBO and EIAR: 3,360
seeds from India (Raipur), 4,370 seeds from Bangladesh (Mymensingh), 1870 seeds from
Poland, 660 seeds from China (black eye) and 7,540 seeds from Ethiopia (3000 Debre
Zeit local, 2,700 Bahir Dar, 970 Wello, 870 Gondar).
Irradiated seeds (M1) were planted in rows in the field with 10 cm between seeds and 40
cm between rows on a plot of size 4 m by 10 m. Parental line was planted in a separate
plot comparatively to monitor general performances. The climatic and soil conditions of
DZARC were as described in Chapter III. Field managements (weeding, cultivation)
were performed as needed. Progeny seeds (M2) from plots were bulk-harvested for the
next generation planting, from which selection would be made.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
98
Selection for morpho-phenotypic appearance in the field
About half of the 140,000 obtained M2 seeds were planted along parental check in field
plots at DZARC. Field plot preparation and planting procedures were as described in the
above section. In comparison with parental check, plants grown from M2 seeds were
screened on the basis of morpho-phenotypic markers inspired from similar work on
soybean (Imsande, 2001). This established procedure in identifying soybean methionine-
rich lines was conceived to have a similar applicability in grass pea. Selections from the
field screening were planted in separate plots of smaller size in an insect proof screen
house for a further two subsequent generations (M3 and M4) to confirm whether the
altered phenotypes were stable mutants compared to the parents. However, the Chinese
and Polish cultivars did not give viable seeds and were excluded from further screening.
Plant parameters on mean basis were determined from randomly selected plants in the
plot. After germination, days to flowering was determined at 50 % of plants flowering in
the plot, whereas days to maturity is detrmined when 95 % of the pods changed to yellow
(pale). Plant height (cm) is measured from five ramdomely selected plants in the
respective plot at maturity. Seed number per plant is mean seed count from five plants.
Hundred seed weight (g) was measured twice from random 100 seeds of the respective
line and mean value taken. Yield per plant (g) was determined from total plant yield in
the plot divided by the number of plants. Leaflet number is the count of five plant leaflets
and expressed in mean. Mean leaf area, leaf width, leaf shape, leaf length were
determined using CI-202 Portable Leaf Area Meter (model 4901, CID,USA) by measuring
100 leaves. Leaf area index and leaf length to leaf width ratios were derived from
measured values. Descriptive statistics for the parameters were performed on the basis of
means for characterization of mutants against the parent.
Selection by ethionine resistance
Ethionine is the higher homologue of methionine and is toxic for plants and all living
systems. Hence, plants producing more methionine are assumed to be more tolerant to
ethionine (Imsande, 2001). The second half of the harvested progeny seeds (M2) were
taken for an in vitro screening, based on root growth in ethionine media following

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
99
procedure published in Imsande (2001) for soybean. DL-ethionine (99 %) was purchased
from Acros (Belgium) and dissolved in monodistilled water at a concentration range of
12.5, 50, and 100 ppm. However, this method could not be applied successfully in grass
pea mainly due to lack of fertility of the screened materials. So a workable screening
procedure for grass pea was failed although its application with soybean was reported
(Imsande, 2001). Therefore, there would not be further reports in the result section on
this.
Evaluation of the selections
After selections were identified and isolated, free and protein-bound amino acid contents
of randomly selected 10-14 M4 seeds and parental seeds were analysed using High
Performance Liquid Chromatography as described in Chapter II.
VI. 3. Results
Morpho-phenotypic variations were observed in plants grown from irradiated grass peas
in the field as indicated in Figure VI.1 and Table VI.1. Irradiation increased the range of
variation of traits in mutants obtained as compared to control. Mutagenic events observed
included decreased plant height, reduced branching, leaf colour intensity, leaf
deformation (leaf curling), infertility and lack of flower setting. The frequency of events
observed at M2 ranged between 1 and 3 per thousand irradiated seeds. More variations
were observed among the Ethiopian cultivars, which produced more biomass compared
to the Indian and Bangladesh cultivars. The later two cultivars behaved as dwarf types
under Ethiopian growth conditions.
The phenotypic feature of selections were the glittering deep green colour of leaf, stem
and pods (Figure VI.1) that was clearly observed in the field and was similar to what was
reported in soybean for methionine-enriched mutants (Imsande, 2001). From the progeny
of irradiated Debre Zeit parental seeds, three single plants were visually identified and
selected in the field exhibiting clearly distinctive phenotypes. They are sub-erect, have a
smaller biomass, smaller leaf size, comparable seed size, and slightly smaller grain

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
100
productivity compared to the parent line (Debre Zeit). They were selected for generation
advancement, characterization and were designated as deep greens (DGA, DGB and
DGC). These putative mutants are discussed in this chapter.
Figure VI.1: Phenotypic appearance of control (right) and one of the mutants (left) - DGA (M4). Differences in colour, glitterness and plant morphology were among phenotypic markers used as selection criteria in the field.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
101
Figure VI.2: Chromatograms of protein amino acids; the methionine [↓] peak at elution time 37.3 min in the parent Debre Zeit seed, deep green mutant line and in soybean seeds, respectively Several agronomic and qualitative parameters analysed in the field showed variations
between parental and putative mutant lines. The parental line has got greater plant height
(97 + 5.6 cm) surpassing the mutants by about 61 %, 38 % and 15 % for DGA, DGB and
DGC, respectively (Table VI.1). Yield parameters showed a wide variation in the
selections with DGC scoring best for early flowering, albeit later than the parent, and
having the longest reproductive period (from flowering to maturity) that possibly allowed
more translocation and accumulation of photosynthates into the sink. With efficient
fertilization this line may have higher harvest index as can be seen from the higher
number of seeds per plant. Leaf area index, a quantitative indicator of photosynthesis, is
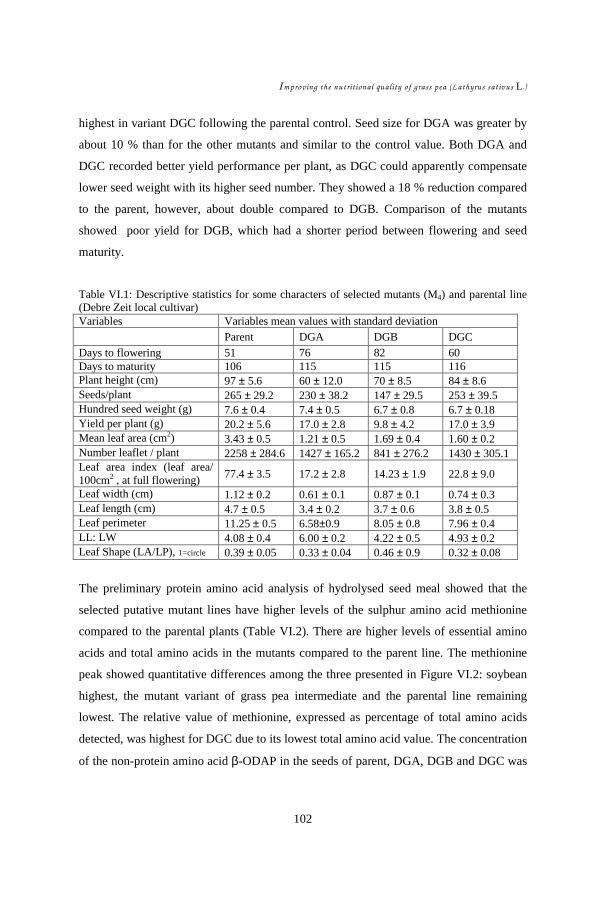
Im proving the nutritional quality of grass pea (L athyrus sativus L.)
102
highest in variant DGC following the parental control. Seed size for DGA was greater by
about 10 % than for the other mutants and similar to the control value. Both DGA and
DGC recorded better yield performance per plant, as DGC could apparently compensate
lower seed weight with its higher seed number. They showed a 18 % reduction compared
to the parent, however, about double compared to DGB. Comparison of the mutants
showed poor yield for DGB, which had a shorter period between flowering and seed
maturity.
Table VI.1: Descriptive statistics for some characters of selected mutants (M4) and parental line (Debre Zeit local cultivar) Variables Variables mean values with standard deviation Parent DGA DGB DGC Days to flowering 51 76 82 60 Days to maturity 106 115 115 116 Plant height (cm) 97 ± 5.6 60 ± 12.0 70 ± 8.5 84 ± 8.6 Seeds/plant 265 ± 29.2 230 ± 38.2 147 ± 29.5 253 ± 39.5 Hundred seed weight (g) 7.6 ± 0.4 7.4 ± 0.5 6.7 ± 0.8 6.7 ± 0.18 Yield per plant (g) 20.2 ± 5.6 17.0 ± 2.8 9.8 ± 4.2 17.0 ± 3.9 Mean leaf area (cm2) 3.43 ± 0.5 1.21 ± 0.5 1.69 ± 0.4 1.60 ± 0.2 Number leaflet / plant 2258 ± 284.6 1427 ± 165.2 841 ± 276.2 1430 ± 305.1 Leaf area index (leaf area/ 100cm2 , at full flowering) 77.4 ± 3.5 17.2 ± 2.8 14.23 ± 1.9 22.8 ± 9.0
Leaf width (cm) 1.12 ± 0.2 0.61 ± 0.1 0.87 ± 0.1 0.74 ± 0.3 Leaf length (cm) 4.7 ± 0.5 3.4 ± 0.2 3.7 ± 0.6 3.8 ± 0.5 Leaf perimeter 11.25 ± 0.5 6.58±0.9 8.05 ± 0.8 7.96 ± 0.4 LL: LW 4.08 ± 0.4 6.00 ± 0.2 4.22 ± 0.5 4.93 ± 0.2 Leaf Shape (LA/LP), 1=circle 0.39 ± 0.05 0.33 ± 0.04 0.46 ± 0.9 0.32 ± 0.08
The preliminary protein amino acid analysis of hydrolysed seed meal showed that the
selected putative mutant lines have higher levels of the sulphur amino acid methionine
compared to the parental plants (Table VI.2). There are higher levels of essential amino
acids and total amino acids in the mutants compared to the parent line. The methionine
peak showed quantitative differences among the three presented in Figure VI.2: soybean
highest, the mutant variant of grass pea intermediate and the parental line remaining
lowest. The relative value of methionine, expressed as percentage of total amino acids
detected, was highest for DGC due to its lowest total amino acid value. The concentration
of the non-protein amino acid β-ODAP in the seeds of parent, DGA, DGB and DGC was

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
103
also determined by HPLC as 0.35 %, 0.38 %, 0.22 % and 0.36 % of seed weight
respectively.
Table VI.2: A preliminary result (single analyses) of protein amino acids (% seed dry weight) levels of mutants (DGA, DGB, DGC) and parental line (Debre Zeit local cultivar). Values in parenthesis are percentage of total amino acids detected
Parent DGA DGB DGC Aspartate 2.02 (11.50) 3.37 (13.00) 3.36 (11.92) 2.37 (11.54) Glutamate 3.15 (18.00) 4.24 (16.85) 5.06 (17.94) 3.29 (16.02) Serine 1.03 (5.88) 1.42 (5.64) 1.60 (5.67) 1.05 (5.11) Glycine 0.77 (4.40) 1.09 (4.33) 1.21 (4.29) 0.80 (3.90) Histidine 0.52 (2.97) 0.73 (2.90) 0.79 (2.80) 0.52 (2.53) Arginine 1.57 (8.97) 2.46 (9.78) 2.60 (9.21) 1.56 (7.60) Threonine 0.64 (3.66) 1.02 (4.05) 1.11 (3.93) 0.74 (3.60) Alanine 0.87 (4.97) 1.57 (6.40) 1.56 (5.53) 1.05 (5.11) Proline 1.23 (7.03) 1.59 (6.31) 1.87 (6.63) 1.15 (5.60) Tyrosine 0.54 (3.09) 0.75 (2.98) 0.78 (2.77) 0.61 (2.97) Valine 0.83 (4.74) 1.15 (4.57) 1.28 (4.53) 0.84 (4.09) Methionine 0.11 (0.61) 0.18 (0.71) 0.19 (0.67) 0.16 (0.77) Isoleucine 0.75 (4.29) 1.05 (4.17) 1.16 (4.11) 0.79 (3.85) Leucine 1.32 (7.54) 1.60 (6.36) 2.20 (7.80) 2.43 (11.83) Phenylalanine 0.86 (4.91) 1.17 (4.65) 1.32 (4.68) 0.88 (4.28) Lysine 1.31 (7.49) 1.77 (7.04) 2.05 (7.27) 1.30 (6.38) Total 17.50 (100) 25.16 (100) 28.21 (100) 20.54 (100)
VI.4. Discussion
In general, the traditional crop plant populations arose from natural variation and sexual
recombination. Contemporary plant breeding is based on creating variation, selection,
evaluation and multiplication of selected genotypes. One of the elements of variation
creation is spontaneous mutation, which naturally occurs at very low frequency and is
difficult to recognize. To create additional variation, the use of chemical and physical
mutagens is an interesting method, which has become an established technology
(Waghmare and Mehra, 2000).
Our results confirm the effectiveness of mutagens in inducing a wide variability of grass
pea traits, which was also observed by other investigators (Waghmare and Mehra, 2000;
Rybinski, 2001). The selected mutants exhibited variability in important yield
parameters. From an agricultural viewpoint, reduction in plant size with reasonable yield

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
104
as seen in mutants is highly needed as grass pea has important yield losses from the
inherent high lodging problem. The per plant basis yield of the mutant lines could be
acceptable, provided the methionine level is improved. Waghmare and Mehra (2000)
using gamma rays and EMS (ethyl methanesulfonate), induced polygenic variability in
grass pea and found plant heights of 26-120 cm in M2 and 37-118 cm in M3.
A visual phenotypic screening established for methionine overproduction in soybean
experiments (Imsande, 2001) appears to be applicable in grass pea. This method has
become instrumental in the identification and characterization of methionine-enriched
mutants. Selected mutants maintained the phenotypical markers over the subsequent
generations. The isolation of methionine overproducing lines both by phenotypic
screening procedures (Imsande, 2001) and chemical analyses resulted in three lines with
a promising increase of methionine. Considering that the sulfur amino acid limitation of
grass pea is believed to be an important factor in the aetiology of a neurodegenerative
disease (Getahun et al., 2005), this result may well have important medical consequences.
A promising increase in the level of methionine by mutagenesis is now reported for the
first time in grass pea, although more analyses should be done to confirm this result. This
would improve the methionine content of grass pea seed from one-fourth of the dietary
requirement of the standard diet (FAO/WHO/UNU-1985) to a more healthy level. This
could probably imply a considerable reduction of the potential toxicity effect in seeds of
grass pea. As methionine biosynthesis depends on sulphur fixation into the cysteine
pathway (Molvig et al., 1997), the mutagenic event might have altered gene expression
and/or enzyme activity in favor of cysteine/methionine biosynthesis.
Enhancing methionine levels via genetic engineering has shown to increase the nutritive
value of legume seeds (Molvig et al., 1997). The increased level of essential amino acids
including methionine in the mutants may considerably improve the nutritional quality of
grass pea towards a better-balanced food and feed. Mutants with lower total amino acid
values appear to have higher relative methionine levels. This relative value of essential
amino acids determines the quality of protein. Supplementary feeding experiments need

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
105
to verify the biological value of the protein from mutants with the improved methionine
level.
The higher methionine selections (DGA, DGB, DGC) still have elevated β-ODAP levels
that are similar to the parent line (around 0.35 % of seed weight). Simultaneous reduction
in the level of the neurotoxin needs to be worked out by crossing of our mutant selections
with the presently available low toxin lines.
Sufficient dietary methionine and even addition of synthetic methionine improves the
nutritional quality of a diet with pulses in general (Molvig et al., 1997). Even more
importantly, higher methionine can be protective against neurolathyrism for consumers of
grass pea (Getahun et al., 2005). Compared to the parental line, the putative mutants
obtained in this study have around 60 % higher methionine level, which would improve
their nutritional quality. This also implies that the antioxidant activity of food or feed
made from these seeds would be higher, with potentially lower risk for neurolathyrism.
This considerable improvement is, however, not yet sufficient to reach the optimal
balance of essential amino acids. Hence, selection of adaptive, methionine
overproducing, low β-ODAP lines is becoming a more promising field of research to
develop nutritionally improved grass pea lines that would be safe for consumers.
Evaluation of these mutants in different environmental conditions together with
nutritional or feeding tests need to be undertaken to determine comparative benefits. The
new lines could be further used in breeding programmes and for genetic analysis. Such
approaches need to be further strengthened to achieve the goal of preventing
neurolathyrism in Ethiopia and in other countries.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
106

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
107
CHAPTER VII
General discussion and conclusions

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
108

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
109
CHAPTER VII
General discussion and conclusions
From the viewpoint of improving the nutritional quality of grass pea, it is felt that a
holistic approach integrating the social, environmental and biological components could
be chosen for solving the nutritional and toxicity problems of the crop. Aside from topics
presented in individual chapters of this thesis, the author has made important
observations while touring lathyrism-prone areas during field work in Ethiopia. It is
worth mentioning that grass pea consumption habit (with or without cereals or fish),
processing, level of education, immature green pea consumption frequency are all
variables existing in the society that seem to affect the incidence of neurolathyrism. The
present work describes observations and experiments that contribute to our understanding
of this crop, the aetiology of neurolathyrism and to its prevention. It also highlights some
properties of grass pea that are less frequently in focus of research on grass pea and
neurolathyrism.
VII.1. Environmental influence on levels of amino acids and crop performances in L. sativus
Efforts to improve this important legume crop have been underway since the discovery of
the neuro-excitatory amino acid β-ODAP in the seeds (Rao et al., 1964). Since this
discovery the focus was on the reduction of this metabolite. No holistic approach on the
more general improvement of the nutritional quality was performed and other merits of
the crop were neglected. As a consequence of the stigma of neurolathyrism and the lack
of incentive to improve the agronomic properties, not much progress has been achieved
to make grass pea a more productive food crop. Probably because the crop was normally
used for forage and only becoming important for human consumption during droughts
and famines, the crop has undergone very little evolutionary progress in its adaptive
environment.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
110
Analysis of the free amino acids of grass pea seeds (chapter II) revealed that the non-
protein amino acid homoarginine (Har), which is absent from most other legumes, was
the most abundant amino acid among all grass pea genotypes from diverse origin,
supporting previous results (Lambein et al., 1992). The concentration of this metabolite
was rather similar among all the genotypes, which marks a unique feature specific to
grass pea seeds. The study of the profile of free amino acids in grass pea and in some
other common food legumes including pea, bean and lentil (Kuo et al., 2004) showed that
Har was undetectable in the dry seeds of these species where quantitatively the most
important free amino acid was arginine (in pea and bean) or glutamic acid (in lentil).
Haque (1997) reported that the level of Har in the seeds increased dramatically in the low
toxin line of L. sativus LS-82046, when growing in Hoagland solution under stress
conditions with supply of the double amount of the trace elements Zn2+, Fe2+, B(OH)-4
and Co2+ in hydroponic media or with Al3+ at 2 x 10 –6 M in the media, or with increasing
salinity up to 0.8 % NaCl (w/v).
Har has been reported to antagonise the neurotoxic action of β-ODAP fed to one-day-old
chicks (Yusuf et al., 1995; Shamim et al., 2002). This might be explained by the
modulating effect of L-Har on nitric oxide synthase (Jyothi & Rao, 1999).
β-ODAP is quantitatively the second most abundant free amino acid in L. sativus seeds
but with greatest variation from 0.2 mg/g dry seed in LS-82046 (Canada) to 5.4 mg/g dry
seed in Raipur (India) (chapter II and Fikre et al., 2008). The presence of β-ODAP in
grass pea seeds was blamed for causing the crippling disease neurolathyrism and the
toxicity is possibly mediated by collective effects of L-β-ODAP on the AMPA-type
receptor, metabotropic glutamate receptors, and nitric oxide production (Kusama-Eguchi
et al., 2006). This free amino acid is reported to be highly variable among genotypes and
environments (Campbell, 1997; Wuletaw, 2003). We have been able to demonstrate the
high variability of this molecule as its level is affected by changes in temperature,
moisture regime, light duration, plant phenology, soil nutrients and altitudinal variations
(chapter III). Increases up to 100 % in levels were exhibited by the same cultivars grown

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
111
in environments with a marked alteration of altitudinal and associated climatic and
edaphic factors in Ethiopia (chapter III).
Earlier studies on the influence of nutrient supply on the β-ODAP content in grass pea
seeds growing in hydroponic media indicated that reduction of the macro-nutrients such
as Mg2+, or K+ in the Hoagland solution resulted in sharp changes of β-ODAP content in
the ripe seeds (Kebede et al., 1994). Conversely, actual environmental factor analysis
(chapter III) showed that K+, known to be high in Ethiopian soil (Murphy, 1968), has a
strong positive correlation with β-ODAP. This strong dependency on environmental
factors indicates the potential for improved crop quality under better-controlled
environmental conditions. We have demonstrated that K+, Zn2+, and P are crucial nutrient
factors influencing β-ODAP content under field conditions while Jiao et al. (2006)
reported the same importance for N and P in Gansu province of China.
Drought stress was reported to have maximal influence on β-ODAP concentration in
grass pea (Lambein et al., 1990; Campbell, 1997; Haque, 1997; Cocks et al., 2000;
Asfaw et al., 2003; DZARC, 2003). From studies on interrelations between
environmental factors we propose that drought stress influences the β-ODAP level in a
multifaceted interplay of links to which the plant responds by over-synthesizing β-ODAP
as a protective molecule or a stress metabolite. The mechanism could be summarized as:
Directly Indirectly Plant responses/processes
Drought
stress
Rain fall distribution
Temperature and wind
Soil moisture availability
Altitude
Evapotranspiration
Nutrient uptake
pH
Root exploration restricted
Physiological retardation
Forced maturity
Yield and metabolites synthesis
Low productivity
High conc. of β-ODAP → seed
sink
Efforts in plant breeding research to eliminate β-ODAP from the seeds have produced
large numbers of “low toxin” varieties but did not yet result in “toxin-free” ones. A
number of post-harvest processing methods of grass pea seeds dramatically reduced the

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
112
level of β-ODAP but could not remove the neurotoxin completely (Srivastava and
Khokhar, 1996; Kuo et al., 2000).
In seed hydrolysates of all grass pea genotypes glutamate was found to have the highest
concentration followed by aspartate, arginine and lysine (chapter II). The sulphur amino
acid methionine ranked the lowest among all the amino acids in all genotypes of L.
sativus as well as in lentil and soybean. The average level of protein methionine in L.
sativus was 0.1 % of the seed weight, and seems rather stable among genotypes of
diverse origin in spite of the high variation of β-ODAP. There is no significant difference
in protein methionine level between grass pea genotypes from India with high β-ODAP
content and Canadian line LS 87124 with low β-ODAP content. Thus protein methionine
content seems not to be correlated with the level of β-ODAP in Lathyrus sativus seeds,
although a biochemical link seems to exist because O-acetylserine (OAS) is involved in
the biosynthesis of both cysteine and BIA (β-(isoxazolin-5-on-2-yl)-alanine) through
cysteine synthase (see Figure VII.1). BIA, present in seedlings of species of the genera
Pisum, Lens, most Lathyrus species and some Vicia species, was confirmed to be the
precursor of β-ODAP in grass pea (Ikegami et al., 1993; Kuo et al., 1998).
Figure VII.1: Biosynthetic pathways of the sulphur containing amino acids, Cystine and Methionine. The enzymes involved in the pathways are: CS (Cys synthase); Cgs (Cysthathionine g-synthase); Cbl (Cystathionine beta-lyase); MS (Met synthase); SAMS (S-adenosylmethionine synthase). Scheme adapted from Noji and Saito (2003).
Acetyl-CoA Ser
OAS CS BIA ↓ ODAP
SO42- ↓ ↓ S2- ↓ CS Cys ↓ Protein Protein
↓ Cgs Cysthathionine ↓ Cbl Homocystein ↓ MS ← Met ↓ SAMS SAM

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
113
Grass pea seeds are the major protein source for the poor and occasionally become the
survival food during crop failure. Gatta et al. (2002) reported that the protein content of
161 Bangladesh accessions ranged from 23 to 29.9 % with a mean value of 26.3 %. We
found 24.1 % protein content in Debre Zeit local cultivar and Urga et al. (2005) reported
a protein range of 27.3-32 % in fifteen cultivars collected in Ethiopia.
Agronomically, the dominant factors affecting the yield of grass pea in studied locations
of Ethiopia were the days to maturity and moisture (chapter III). Yield and various yield
components have shown variability in response to cropping seasons (years), genotype,
locations and their interactions. Better yield performance was associated to locations at
higher altitude where the crop growth period was relatively longer. The exceptional low
yield harvest at Denbi might be attributed to the shorter maturity period, and high
concentration of sulphur and manganese in the soil that showed a significant negative
correlation to yield. Despite their individual negative correlation the interaction Zn2+/S
produced a positive correlation to yield, suggesting that an optimal ratio can be found for
better yield realization.
In summarizing this part, it is very well established that grass pea is an important legume
grain food and a life saving crop (Fikre et al., 2006) but it is also controversial as the
cause of crippling lathyrism. It is well known that genotypic variability exists for the
level of the neuro-excitatory amino acid β-ODAP in the seed (Wuletaw, 2003). Those
earlier studies were done using a colourimetric method that cannot distinguish between
the non-toxic α-ODAP and the toxic β-ODAP. In our comparative study with two other
food legume species (lentil and soybean), the levels of β-ODAP, methionine and other
amino acids were determined using the more accurate HPLC-method (Fikre et al., 2008).
This is also the first comparative study on free and protein amino acids in a set of
genotypes collected from different parts of the world. However, the concentration of β-
ODAP (with a wide range) and methionine (narrow range) did not show any specific
correlation between these two amino acids, although one enzyme, cysteine synthase, is
involved in the biosynthesis of both.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
114
In Ethiopia, recurrent climatic disasters (drought) are a common phenomenon, and this is
known to be associated with increasing β-ODAP in L. sativus (Asfaw et al., 2003) with a
potential increase of neurolathyrism incidence to epidemic proportions (Haimanot et al.,
2005). However, the role of environmental factors on the toxicity (β-ODAP level) of the
crop has never been comprehensively studied. From the present path analysis using two
growing seasons at five locations, a number of additional factors including both climatic
and edaphic ones were shown to have a strong relation with β-ODAP level and crop yield
in Ethiopia. Previously, other authors did not consider these factors. The results suggest
novel crop management options to better control the particular agro-ecology for safer
utilization and better exploitation of this underutilized crop. However, such short period
of data collection in the unstable and changing climatic condition of Ethiopia may not
allow long-term predictions. Also, the results may not be applicable to countries such as
Bangladesh with different edaphic (lowland sedimental soil as opposed to volcanic soil in
Ethiopia) and climatic (higher precipitation and even flooding) conditions and also with
different geographic locations (different light quality).
VII.2. Effect of methionine on toxicity of L. sativus
This study was initiated with the hypothesis that methionine could neutralize the
antinutritional nature of grass pea and would contribute better to its overall food quality
improvement than focusing only on toxicity reduction/elimination alone. When
comparing neurolathyrism with konzo (an irreversible crippling disease undistiguishable
from neurolathyrism and caused by prolonged overconsumption of cassava root
preparations) it was suggested that the common feature of the diets leading to those
similar diseases, namely methionine deficiency could play a role in the aetiology of
neurolathyrism (Defoort, 2004; Lambein et al., 2004). The impetus was then to confirm
the hypothesis that methionine would affect grass pea toxicity and also to estimate the
level of its effect. A feeding experiment with methionine supplements in the diet was
done with broiler chicks and the results are presented in section IV.3.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
115
Nunn and co-workers (2005) reported that consumption of grass pea seeds deficient in
methionine as staple food for a long period leads to the deprivation of methionine in the
plasma. They suggested that long-term deficiency of methionine in plasma and thus
reduced methionine flux into motor neurone cells might lead to the higher susceptibility
of these cells to the excitatory effects of β-ODAP.
Among the essential amino acids in grass pea the level of methionine and cysteine in the
seed reaches only about 1/4th of the requirement for a normal balanced diet. Diets with
grass pea as the staple are therefore a poor source of methionine and cysteine amino acids
(Kuo et al., 1995). The antioxidant effect of methionine and its protection against the
oxidative stress induced by the neuro-excitatory activity of ß-ODAP leading to
neurolathyrism have been reported (Kumar and Rao, 1990; Getahun et al., 2005; Nunn et
al., 2005).
From the grass pea feeding trial (chapter IV) the effect of increasing methionine addition
was evident in bringing significant change to the chick performances and phenotypic
responses. In spite of the general under-performances for some parameters when the
chicks received exclusively grass pea feed (98.5 %), the broiler chicks showed
improvement along with the increasing addition of methionine. Although this study did
not address the economics of grass pea in animal husbandry, it has shown that a positive
outcome can be expected from a focussed economic study, which can be undertaken
using different formulations as treatment inputs.
The highest effects were found at methionine addition of 0.075 % when grass pea
accounts for 35 % of the feed and 0.15 % when grass pea (98.5 %) became almost the
exclusive feed source. If this would be valid also for humans, then this may suggest that
methionine optimisation in the diet could play a central role for the safe utilization of this
under-utilised but protein rich grass pea (Lambein et al., 2001; Getahun et al., 2005;
Nunn et al., 2005).

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
116
It was also demonstrated that the β-ODAP intake tolerance of chicks increased with the
addition of methionine in the diet (chapter IV). On average, the performance (feed intake,
weight gain) of the chicks fed on the high β-ODAP Ethiopian grass peas was
significantly improved. The improved performance in feed intake and body weight could
likely be the indirect consequence of enhanced FCE and/or from the fact that methionine
protects the animals from oxidative stress induced by ß-ODAP. In an earlier study
Hanbury and co-workers (2000b) had observed a tolerance in chicks of up to 40 % grass
pea (containing 0.27 % ß-ODAP) with minor weight reduction.
With the increase of methionine in the feed, ß-ODAP was found to decrease in serum of
chicks on ration I (35% grass pea), which may result from improved physiological
condition and metabolism of the chicks (Chapter V). Individual variability in tolerance to
acute neurological symptoms was seen in chicks having the same level of exposure. Rao
(2001) and Rudra and co-workers (2004) suggested that there exist extreme inter and
intra species differences in susceptibility to ß-ODAP that may be ascribed to individual
metabolic differences. To our knowledge a genetic basis for such differences has only
been proposed by Getahun and co-workers (2002b) who indicated that blood type could
be a factor of susceptibility to neurolathyrism in humans.
From the analysis of trace elements in chick’s serum (chapter V) we found a gradual
increase of Fe2+, Zn2+, and Cu2+ with time but not for Mn2+. In agreement with previous
studies, in this study acute neurological symptoms were observed more in younger chicks
and becoming less as age increased. From the data of feed intake over the feeding period
we calculated that the convulsive dose for young chicks was about 600 mg of ß-ODAP
per kilo of body weight in ration II (98.5% grass pea) while Chen and co-workers (1992)
reported that intraperitoneal injection of ~700 mg ß-ODAP/kg body weight is lethal for
3-day old chicks. Unlike the manifestations of neurolathyrism in humans, the common
convulsive symptoms in chicks were mainly spasticity of the neck muscles. Differences
also include the reversibility of symptoms in chicks and the irreversible symptoms of leg
spasticity in man with sudden onset after prolonged consumption of mainly grass pea
seed during two months or more.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
117
Our study attempted to estimate the particular effect of methionine on grass pea toxicity
as feed. Deficiency of methionine in L. sativus (Kuo et al., 1995) and its potential
involvement in neurolathyrism were reported (Kumar and Rao, 1990; Nunn et al., 2005;
Getahun et al., 2005). The absence of a suitable animal model is claimed to be a barrier
for better understanding of neurolathyrism (Getahun et al., 2004). A limited study was
undertaken on the effect of methionine on chicks fed a L. sativus based ration (Table
IV.5). Unlike previous experiments, in this thesis the effect of methionine per se on grass
pea toxicity was studied not only from the viewpoint of animal performance, but also on
neurological symptoms. Chicks were symptomatically characterized, their performance
recorded and amino acids in the serum analyzed. The study contributes to the
understanding of the effect of methionine on L. sativus toxicity and confirms
unequivocally its protective effect against grass pea toxicity in young chicks. It also
indicates the possibility that methionine could be implicated as protective factor in other
animals and/or human neurolathyrism as well. However, the experimental setup did not
allow distinguishing between the toxic effect of β-ODAP and of other ANFs, as only the
raw unprocessed seeds were used.
VII.3. Mutational methionine enrichment in L. sativus
The sulphur containing amino acids methionine and cysteine are in general deficient in
legumes and consumption of legumes as the sole protein source in the human diet is not
advised. The deficiency of these amino acids (Kuo et al., 1995) is of particular
importance in neurolathyrism disease development that is caused by monotonous
consumption of L. sativus.
By adopting phenotypic markers for methionine enriched lines as has been established for
soybean (Imsande, 2001) we have identified and selected L. sativus variants in the field
from γ-irradiated seeds (chapter VI). The phenotypic variants exhibited phenological and
morphological appearances different from the parental line. Unlike the parent line they
have a smaller size, later maturity and glittering deep green phenotypic appearance.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
118
Amino acid analysis of seeds from the mutants has shown a 60 % higher methionine
content compared to the parent. Unlike other legumes, the sulfur amino acid limitation of
grass pea apparently has medical implications by increasing the susceptibility of subjects
to a neurodegenerative disease caused by dependent consumption of the crop (Getahun et
al., 2005). Hence, it is assumed that in vivo methionine improvement potentially
enhances the nutritional quality as well as the safety of grass pea food preparations or
feed. However, the ß-ODAP level in the putative mutants we obtained is not lower than
that in the parent. Further work should aim at the simultaneous reduction of the β-ODAP
level and increase of methionine as a promissing goal. While genetic transformation
could be utilized to reach this goal, the present selections can at least be used in breeding
programs to achieve lines with increased nutritional quality and adapted to marginal or
drought prone lands.
In conclusion, in vivo methionine enrichment in L. sativus could be a plausible approach
in the process of making it a more healthy food. This has never been tried effectively thus
far. Among methods that could produce genetic alteration, mutagenesis was attempted by
irradiating several thousand seeds where mutants with a distinct phenotypic appearance
were advanced over four subsequent generations and were chemically and phenotypically
characterized. From the preliminary analysis, some selected putative mutants have
revealed higher methionine content and desirable agronomic performances. Though the
work is at its early phase, this novel approach might give promising prospects for
improvement of the crop with a better balance of essential amino acids. Moreover, the
potential introduction of such mutants would not be subjected to the stringent regulations
that virtually stop the use of even the most promising genetically modified food plants.
Limited experience in this area has put a challenge in the course of the work. The work is
not yet complete/conclusive as analysis and characterization has to go over several
generations to confirm this preliminary result. Supportive genetic and molecular studies
(mode of inheritance, molecular characterization) are needed to further characterize and
utilize the selections.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
119
VII.4. Recommendations and future perspectives This thesis attempts to identify and study gaps in our knowledge of the traditional legume
crop plant Lathyrus sativus and the human crippling disease neurolathyrism. The ultimate
goal is to prevent neurolathyrism by establishing varieties with improved nutritional
quality without the risk for the irreversible crippling neurolathyrism. A secondary goal is
to suggest ways to optimise the use of this underutilized crop. The environmental and
biological aspects have been studied after a thorough literature review of already
established knowledge. Recommendations and suggestions of particular importance or
usefulness are forwarded.
In the interaction of various factors, the process of neurolathyrism development could be
summarized in the following simplified scheme (Figure VII.2).
Figure VII.2. Hypothetical scheme of neurolathyrism (NL) development by interaction of various factors (human, environment, socio-economics and grass pea plant). Some factors are directly linked to NL; others are via affecting β–ODAP.
The level of factor’s effect = * lowest, ***** highest A-age, B- sex, C- physical labor, D- health status, E- individual metabolism/genetics, F- moisture stress, G- soil nutrient balance, H- salinity, I-light hours, J- genetic interventions, K- crop management/ productivity, L- stressed growth, M- processing, N-green pea O- traditional perception and precaution measures Vs scientific information, P- poverty (± option), Q- consumption style and intensity, R- diet status (± methionine and antioxidant rich condiments). Our study demonstrated that crop parameters such as yield, ß-ODAP and other amino
acids of grass pea are affected by various environmental factors. Particularly, the unstable
ß-ODAP was reported to be affected by moisture stresses, nutrient levels, genetic factors
and processing. However, these parameter effects should also be studied in other areas
prone to neurolathyrism, as there could be certain specificity from country to country. In

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
120
this part of the study, interrelations between amino acid levels and environmental
conditions of grass pea have been demonstrated. Especially the great variability of ß-
ODAP level in response to varying environment, media and processing was verified
under Ethiopian conditions. As the sulphur containing amino acid methionine has been
proposed to protect cells against β-ODAP toxicity, we also focused on the level of
methonine in seeds (± 0.1 % of dry weight). It was shown that methionine levels are
much less variable compared to ß-ODAP in grass pea and it is present in concentrations
that may lead to dietary deficiency if grass pea is consumed as staple. Especially, this
study revealed that, among several factors considered, the soil factors (K+, pH) and the
photosynthetic factor (sunshine hours) have a strong positive relation with β-ODAP level
while days to maturity, yield and altitude have negative correlations. Further studies on
the fertilization and other factors affecting crop yield, Rhizobium nodulation and
adaptation across environments and resistance to biotic and abiotic stresses are
anticipated to provide better information to optimize the crop use. This drought tolerant
traditional crop is well suited for adaptation to the increasing water deficits Ethiopia may
face due to the global warming and to become an important asset to overcome food
insufficiency if the stigma of neurolathyrism can be removed.
The antioxidant activity of the essential sulphur containing amino acids cysteine and
methionine is confirmed in a variety of research reports. Especially the motor neurones
seem to require high methionine for their integrity. Some experimental studies indicated
that sufficient cereals (1/3rd of the diet) rich in methionine could protect against
neurolathyrism. There are also some results indicating that methionine antagonizes the
antinutritional effects of β-ODAP. In a broiler chick feeding experiment we have
demonstrated that methionine addition in grass pea feed improved feed intake, weight
gains and general animal performances. This is in support of the hypothesis that the
antioxidant activity of methionine counteracts the oxidative stresses induced by various
physiological and biochemical activities of β-ODAP. Developing a reproducible and
reliable animal model would allow more detailed studies to advance our knowledge on
the aetiology of human neurolathyrism with potential prevention implication. Such an
animal model would be important also for the study of other nutrition related

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
121
neurological diseases such as konzo (a neurological disease with symptoms
undistinguishable from neurolathyrism but caused by unbalanced overconsumption of
methionine-deficient cassava roots).
Apparently, as a potential protective factor against β-ODAP toxicity, methionine enriched
food such as cereals or addition of synthetic methionine or genetically enriched grass pea
varieties are believed to confer such protection. Methionine deficiency in legumes
including grass pea is long established. Grass pea can only supply about a fifth to a fourth
of the methionine required for an optimal balanced diet. There is hardly any ongoing
research on genetic enrichment of methionine in grass pea. We have demonstrated for the
first time a methionine-enriched selection after γ-irradiation of the seeds and we
described the mutant isolates. Phenotypically distinct putative mutants showed a medium
biomass, have comparably smaller and deeper-glittering green leaf, medium seed size and
reasonable grain productivity. This study is now in the M5 stage and mutant lines are
designated as deep greens (DGA, DGB, DGC) referring to their typical phenotype. They
have more than 50 % methionine increase against the parent line. A subsequent study into
its environmental adaptation, the mode of methionine inheritance, crossing ability and
molecular characterization of the genetic alteration would enhance the incentive to
change the crop into a promising healthy food and feed. Further studies on mutation and
molecular techniques are essential in grass pea research. Evaporates of grass pea have to
be studied for potential toxic constituent, as there is no clear evidence of this in spite of
the strongly entrenched traditional conception that the vapour from flowering plants or
from cooking the seed are toxic. Safe varieties of the drought tolerant grass pea with a
better-balanced complement of essential sulfur amino acids can become a wonder crop
for subsistence farmers in marginal areas of Ethiopia and South-East Asia. This would lift
the socio-economic status of many poor farmers and it would also be of enormous benefit
to the ecology of areas with marginal agricultural productivity or with adverse climates.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
122

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
123
References

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
124

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
125
References
Abayneh, E. and Demeke, T. 2003. Soil of Debre Zeit agricultural research center and its
sub-centers. Technical paper No 79. National Soil Research Center. Addis Ababa.
Abd El Moneim, A. Van Dorrestine, B. Baum, M. Ryan, J. and Bejiga. G. 2001. Role of
ICARDA in improving the nutritional quality and yield potential of grass pea for
subsistence farmers in dry areas. Lathyrus lathyrism newsletter 2(2): 55-58.
Abd El Moneim, A., Van Dorrestein, B., Baum, M. and Mulugeta, W. 1999. Role of
ICARDA in improving the nutritional quality and yield potential of grass pea (Lathyrus
sativus L.) for subsistence farmers in developing countries. Los Banos, The Philippines,
5-6 October. pp 1-11.
Akalu, G., Tufvesson, F., Jonsson, C. and B.M. Nair. 1998. Physico-chemical
characteristics and functional properties of starch and dietary fiber in prass pea seeds.
Starch/Stärke 50: 374–382.
Al-Mayah, A.A.S. 2006. Immune response of broiler chicks to DL-methionine
supplementation at different ages. International Journal of Poultry Science 5: 169-172.
Angkanaporn, K, Ravindran, V., Mollah Y. and Bryden W.L. 1997. Homoarginine
influences voluntary feed intake, tissue basic amino acid concentrations and arginase
activity in chickens. Journal of Nutrition 127: 1128-1136.
Asfaw, T., Mulugeta, T. and Lijalem, K. 2003. Breeding grass pea, fenugreek and lupine
for wide adaptation in Ethiopia. Proceedings of the second workshop on pulses research
in Ethiopia. Addis Ababa. pp 102-103.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
126
Baik, B.K., Han, I.H., McPhee, K.E. and Paszczynska, B. 2004. Cooking, roasting and
fermentation of chickpeas, lentils and peas for fortification of pan bread. IFT annual
meeting, July 12-16. Las Vegas, USA. Abstract 67F-6.
http://ift.confex.com/ift/techprogram/paper_24394.htm
Barik, D.P., Mohapatra, U. and Chand, P.K. 2005. Transgenic grass pea (Lathyrus sativus
L): Factors influencing Agrobacterium mediated transformation and regeneration. Plant
Cell Reports 24: 523-531.
Barker, B.T.P. 1916. Sweet pea hybrids. Gardeners’ Chronicle Series 3, 60: 156-157.
Barrow, M.V., Simpson, C.F. and Miller, E.J. 1974. Lathyrism: A review. Quarterly
Review of Biology 49: 101-128.
Bell, EA. 1964. Relevance of biochemical taxonomy to the problem of lathyrism. Nature
203: 378-380.
Bull, T.A. 1996. Agro-ecological assesment of Ethiopian vertisol. FAO Proceedings,
ILCA, Addis Ababa.
http://www.fao.org./wairdocs/IIRI/x5493E/x5493e07.htm#TopOfPage
Campbell, C.G. 1997. Grass pea (Lathyrus sativus L). Promoting the conservation and
use of underutilized and neglected crops. Vol 18. Institute of Plant Genetics and Crop
Plant Research, Gatersleben & International Plant Genetic Resources Institute, Rome,
Italy. pp 1-90.
Central Statistics Authority (CSA) of Ethiopia. (1996-2007). Crop production and area
statistics. Addis Ababa, Ethiopia.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
127
Chen, Y.Z., Li, Z.X., Lu, F.H., Bao, X.G., Liu, S.Z., Liu, X.E., Zhang, G.W. and Li, Y.R.
1992. Studies on the screening of Lathyrus, analysis of toxins and toxicology. Journal of
Lanzhou University (Natural Science) 28: 93-98.
Cho, E. J, Piao, X.L., Jang, M.H., Baek, S.H., Kim, H.Y., Kang, K.S., Kwon, S.W. and
Park, J.H. 2008. The effect of steaming on the free amino acid contents and antioxidant
activity of Panax ginseng. Food Chemistry 107(2): 876-882.
Chowdhury, S.D, Sultana, Z., Ahammed, M., Chowdhury, B.L., Das, S.C. and Roy, B.C.
2005. The Nutritional Value of Khesari (Lathyrus sativus) for Growing and Laying
Pullets. The Journal of Poultry Science 42(4): 308-320.
Coates, J.B., Medeiros, J.S., Thanh, V.H. and Nielsen, N.C. 1985. Characterization of the
subunits of β-conglycinin. Archives of Biochemistry and Biophysics 243: 184-194.
Cocks, P., Siddique, K., and Hanbury, C. 2000. Lathyrus a new grain legume. A report
for the Rural Industries Research and Development Corporation. Faculty of Agriculture
The University of Western Australia, Nedlands, WA, 6907. pp 5-15.
Cottenie, A. 1980. Soil and plant testing as a basis of fertilizer recommendation. FAO
soils bulletin 38(2). Rome, Italy.
Dadi, L., Teklewold, H., Aw- Hassan, A., Moneim, A.A. and Bejiga, G. 2003. The socio-
economic factors affecting grass pea consumption and influence of lathyrism in Ethiopia.
Integrated National Resources Management Research Report Series, No.4. ICARDA,
Aleppo, Syria. pp 7-11.
Dahiya, B.S. and Jeswani, L.M. 1975. Genotype & environment interactions for
neurotoxic principle (BOAA) in grass-pea. Indian Journal of Agricultural Sciences 45(9):
437-439.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
128
Datta, A. 1995. Strategy to develop transgenic Lathyrus with low-level neurotoxin by
recombinant DNA technology. In: Yusuf HKM, and Lambein F (eds). Lathyrus sativus
and human lathyrism. Progress and prospects. University of Dhaka, Bangladesh. pp 173-
176.
Davies, A.J., Nunn, P.B., O’Brien, P., Pettit, L.D. and Wang, G.F. 1990. Facile isolation
from Lathyrus sativus seed of the neurotoxin β-N-oxalyl-L-α,β-diaminopropionic acid as
the copper complex and studies of the coordination chemistry of copper and zinc with the
amino acid in aqueous solution. Journal of Inorganic Biochemistry 39: 209-216.
Defoort, B. Biochemische en fysiologische vergelijking tussen neurolathyrisme en konzo.
Research thesis, Faculty of Medicine and Health Sciences, Ghent University, Belgium.
De Vleeshauwer, A., Hendrickx, H. and Heyndricks, G. 1948. Bepalen van de droge stof.
Onderzoeksmethoden voor zuivelprodukten, Standaard Boekhandel, Antwerpen.
Dixit, S., Khanna, S.K., Das, M. 2008. Non-uniform implementation of ban on Lathyrus
cultivation in Indian states leading to unwarranted exposure to consumers. Current
Science (India) 94: 570-572.
DZARC (Debre Zeit Agri. Research Centre) annual research report. (2001-2004). Debre
Zeit, Ethiopia.
Donaghy, M. 1999. Classification and clinical features of motor neurone diseases and
motor neuropathies in adults. Journal of Neurology 246(5): 331–333.
Duke, J.A. 1981. Handbook of legumes of world economic importance. Plenum Press,
New York. pp 199-265.
Duncan, D. B. 1955. Multiple range test and multiple F tests. Biometrics 11: 1-42.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
129
Durieu, P. and Ochatt, S.J. 2000. Efficient intergeneric fusion of pea (Pisum sativum) and
grass pea (Lathyrus sativus) protoplasts. Journal of Experimental Botany 51: 1237-1242.
Düsünceli, F. 1993. Prospects for production of Lathyrus sativus and L. cicera in
southwest Anatolia in Turkey. In: Lathyrus sativus and Human Lathyrism: Progress and
Prospects (Yusuf H.K.M., and Lambein F., eds.) Proc. 2nd Int. Colloq. Lathyrus/
Lathyrism, Dhaka, 10-12 December 1993. University of Dhaka. pp 131-137.
Eavis B.W. 1971. Effects of flooding on sugarcane growth. Benefits during subsequent
drought. Proc. Internat. Soc. Sugar Cane Technologists 14th Congress. pp 715-721.
Egan, H., Kirk, R. and Swayer, R. 1981. Pearson’s chemical analysis of foods. 8th edition.
Churchill Livingstone. pp 20-23.
Eyzaguirre, P.B., Padulosi, S. and Hodgkin, T. 1999. IPGRI’s strategy for neglected and
underutilized species and the human dimension of agrobiodiversity. In: Priority setting
for underutilized and neglected plant species of the Mediterranean region (Padulosi S,
ed.). Report of the IPGRI Conference, 9-11 February 1998, ICARDA, Aleppo, Syria.
FAO. 2004. Calls for Biotechnology. From AgBioView e-mail news May 17, 2004.
www.agbioworld.org
FAO. 1996-2005. Crop production statistics. Rome, Italy.
FAO/WHO/UNU expert consultation. 1985. Energy and protein requirements. World
Health Organization Technical Report Series 724:1-204.
(http://www.nutrition.org./cgi/content/full/130/7/18415).
Fikre, A. 2007. Grass pea (Guaya). A research review handbook. Ethiopian Institute of
Agricultural Research, Addis, Ababa. pp 1-31.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
130
Fikre, A., Lambein, F. and Gheysen, G. 2006. A life saving food plant producing more
neurotoxin under environmental stresses. Communications in agriculture and applied
biological sciences. Ghent University, Belgium 71(1): 79-82.
Fikre A., Korbu L., Kuo Y.-H. and Lambein F. (2008) The contents of the neuro-
excitatory amino acid ß-ODAP (β-N-oxalyl-L-α,β-diaminopropionic acid), and other free
and protein amino acids in the seeds of different genotypes of grass pea (Lathyrus sativus
L.). Food Chemistry 110: 422-427.
Foster, J.G. 1990. Flatpea (L. sylvestris L.): a new forage species; a comprehensive
review. Advances in Agronomy 43: 317-348.
Gatta, C. D., Polignano, G.B. and Bisignano, V. 2002. Variation for protein content and
seed weight in grass pea (Lathyrus spp.) germplasm. PGR Newsletter. FAO-Biodiversity
issue 132: 30-34.
Getahun, H. 2004. Public health importance of neurolathyrism and epidemiological risk
factors: evidence from Ethiopia. Ph D thesis, Faculty of Medicine and Health Sciences,
Ghent University, Belgium.
Getahun, H., Lambein, F., Vanhoorne, M. and Van der Stuyft, P. 2002a. Pattern and
associated factors of the neurolathyrism epidemic in Ethiopia. Tropical Medicine and
International Health 7: 118-124.
Getahun, H., Lambein, F. and Van der Stuyft, P. 2002b. ABO blood groups, types of
grass pea preparation and neurolathyrism in Ethiopia. Transactions of the Royal Society
of Tropical Medicine and Hygiene 96: 700-703.
Getahun, H., Lambein, F. and Vanhoorne, M. 2002c. Neurolathyrism in Ethiopia:
assessment and comparison of knowledge and attitude of health workers and rural
inhabitants. Social Science & Medicine 54(10): 1513-1524.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
131
Getahun, H., Lambein, F., Vanhoorne, M. and Van der Stuyft, P. 2003. Food-aid cereals
to reduce neurolathyrism related to grass pea preparations during famine. Lancet 362:
1808–1810.
Getahun, H., Lambein, F., Vanhoorne, M. and Van der Stuyft, P. 2005. Neurolathyrism
risk depends on type of grass pea preparation and on mixing with cereals and
antioxidants. Tropical Medicine and International Health 10(2): 169-178.
Girma, A. 1999. Development of a process for reducing the content of β-N-oxalyl-α,β-
diaminopropionic acid (ß-ODAP) in grass pea (Lathyrus sativus) seeds: nutritional
consequences and technological aspects. Ph D dissertation. Lund University, Sweden.
Goyder, D.J. 1985. The Genus Lathyrus. In Lathyrus and lathyrism. Kaul, A.K., and
Combes, D. (eds). The Third World Medical Research Foundation, New York. pp 3-7.
Gravois, K.A. and McNew, R.W. 1993. Genetic relationships among and selection for
rice yield and yield components. Crop Sciences 33(2): 249-252.
Greenwood, D.J. 1968. Effect of oxygen distribution in the soil on plant growth. In: Root
Growth, ed. W.J. Whittington. Proc. 15th Easter School, University of Nottingham. pp
202-223.
Haimanot, R., Abegaz, B.M., Wuhib, E., Kassina, A., Kidane, Y., Kebede, N., Alemu, T.
and Spencer, P.S., 1993. Pattern of Lathyrus sativus (grass pea) consumption and β-N-
Oxalyl-α,β-diaminopropionic acid (β-ODAP) content of food samples in the lathyrism
endemic region of northwest Ethiopia. Nutrition Research 13: 1113–1126.
Haimanot, R.T., Abegaz, B.M., Wuhib, E., Kassina, A., Kidane, Y., Kebede, N., Alemu
T. and Spencer, P.S. 1995. Nutritional neuro- toxicological surveys of Lathyrus sativus
consumption in Northern Ethiopia. in Lathyrus sativus and Human Lathyrism : Progress
and Prospects (H.K.M. Yusuf and F. Lambein, eds.). Proc. 2nd Int. Colloq.
Lathyrus/Lathyrism. Dhaka Pp 41-45.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
132
Haimanot, R.T., Feleke, A. and Lambein, F. 2005. Is lathyrism still endemic in northern
Ethiopia? – The case of Legambo Woreda (district) in the South Wollo Zone, Amhara
National Regional State. Ethiopian Journal of Health Development 19: 230-235.
Haimanot, R.T., Kidane, Y., Wuhib, E., Kalissa, A., Alemu, T., Zein, Z.A. and Spencer,
P.S. 1990. Lathyrism in rural northern Ethiopia: A highly prevalent neurotoxic disorder.
International Journal of Epidemiology 19: 664–672.
Hall, A.E. 2004. Comparative ecophysiology of cowpea, common bean and peanut.
In:”Physiology and Biotechnology Integration for Plant Breeding” p 271-326. University
of California, Riverside, USA.
Hanbury, C. D., Siddique, K.H.M., Galwey, N.W. and Cocks, P.S. 2000a. Genotype-
environment interaction for seed yield and ODAP concentration of Lathyrus sativus L.
and L. cicera L. in Mediterranean type environments. Euphytica 11: 45-60.
Hanbury, C.D., White, C.L., Mullan, B.P. Siddique, K.H.M. 2000b. A review of the
potential of Lathyrus sativus L. and L. cicera L. grain as animal feed. Animal Feed
Science and Technology 87: 1-27.
Hanson, W.D. 1985. Association of seed yield with partitioned lengths of the
reproductive period in soybean genotypes. Crop Science 25: 525-529.
Haque, R. 1997. A study on the effect of nutritional and agro-ecological factors on the
accumulation of neurotoxin β-N-oxalyl-L-α, β-diaminopropionic acid (ODAP) and other
amino acids in Lathyrus sativus (Khesari) aimed at repressing its toxin synthesis. Ph D
dissertation, Bangladesh Agricultural University, Mymensingh.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
133
Hopf, M. 1986. Archaeological evidence of the spread and use of some members of the
Leguminosae family. In The Origin and Domestication of Cultivated Plants (C.
Barigozzi, ed.). Elsevier, Oxford, New York and Tokyo. pp 35-60.
Houba, V.J.G., Van Der Lee, J.J., Novozamasky, I. and I.Wallinga. 1989. Soil and plant
analysis, a series of Syllabi. Part 5. Soil analysis procedure. Dep. Soil Science and Plant
Nutrition, Wageningen. Agricultural University, the Netherlands.
Hussain, M. 1994. Agro-ecological factors affecting the concentration of ODAP in
Lathyrus sativus seeds. Annual Report. Bangladesh-Belgium collaborative project on
Lathyrus and lathyrism. pp 1-17.
Ikegami, F., Ongena, G., Sakai, R., Itagaki, S., Kobori, M., Ishikawa, T., Kuo, Y.H.,
Lambein, F. and Murakoshi, I. 1993. Biosynthesis of beta-(isoxazolin-5-on-2-yl)-alanine,
the precursor of the neurotoxin beta-N-oxalyl-L-alpha,beta-diaminopropionic acid, by
cysteine synthase in Lathyrus sativus. Phytochemistry 33: 93-98.
Ikegami, F., Kusama-Eguchi, K., Kusama, T., Watanabe, K. and Lambein, F. 1995.
Neurological action of β-ODAP and related Lathyrus toxins: A possible relationship to
neurolathyrism. Proceeding of International Conference on Lathyrus and lathyrism, a
decade of progress. Addis Ababa, Ethiopia, 27-29 November 1995. pp 3-7.
Imsande, J. 2001. Selection of soybean mutants with increased concentration of seed
methionine and cysteine. Crop Science 41: 510-515.
Jackson, M.L. 1973. Plant tissue analysis. In: Soil Analysis Chemistry. Prentiss Hall of
India Pvt. Ltd. New Delhi. pp 326-338.
Jellinger, K. 1999. The role of iron in neurodegeneration. Prospects for pharmacotherapy
of Parkinson's disease. Drugs & Aging 14: 115–140.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
134
Jiao, C.-J., Xu, Q.-L., Wang, C.-Y., Li, F.-M., Li, Z.-X. and Wang, Y.-F. 2006.
Accumulation pattern of toxin β-ODAP during lifespan and effect of nutrient elements on
β-ODAP content in Lathyrus sativus seedlings. Agricultural Science 144: 369-375.
Jyothi, P. and Rao, S.L.N. 1999. Sustained nitric oxide generation with L-homoarginine.
Research Communications in Biochemistry & Cell & Molecular Biology 3: 223-232.
Kang, M.S. 1994. Applied quantitative genetics. M.S. Kang, Publ., Baton Rouge pp 1-
157.
Kebede, N., Haque, R., Kuo, Y.-H. and Lambein, F. 1994. Influence of nutrient supply on
the toxicity of Lathyrus sativus. Acta Botanica Neerlandica 43: 295.
Kelbessa, U., and Mengistu, G. 1993. The effect of soaking time and soaking solution on
the nutritional quality of grass pea seeds. Ethiopian Journal of Health Development 7(2):
79-83.
Kessler, A. 1947. Lathyrismus. Monatschrifte Psychiatrie und Neurologie 113: 345-376.
Khawaja, H.I.T. 1988. A new interspecific Lathyrus hybrid to introduce the yellow
flower character into sweet pea. Euphytica 37(1): 69-75.
Kim, H., Hirai, M.Y., Hayashi, H., Chino, M., Naito, S. and Fugiwara, T. 1999. Role of
O-acetyl-L-serine in the coordinated regulation of the expression of soybean seed storage
protein gene by sulphur and nitrogen nutrition. Planta 209: 282–289.
Kislev, M.E. 1989. Origins of the cultivation of Lathyrus sativus and L. cicera
(Fabaceae). Economic Botany 43: 262-270.
Kockhar, S.L. 1992. Economic botany in the tropics. Macmillan India Ltd., Madras. pp
141-142.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
135
Kumar, P.S. and Rao, S.L.N. 1990. Modulation of different forms of glutamine
synthetases in Rhizobium phaseoli and their possible role in nitrogen fixation. Current
Microbiology 21(3): 199-204.
Kuo, Y. -H., Bau, H.M., Quemener, B., Khan, J.K. and Lambein, F. 1995. Solid state
fermentation of Lathyrus seeds using Aspergillus oryzae and Rhizopus oligosporus sp T3 to
eliminate the neurotoxin ß-ODAP without loss of nutritional value. Journal of the Science of
Food and Agriculture 69: 81-89.
Kuo, Y.-H., Bau, H.-M., Rozan, P., Chowdhury B. and Lambein, F. 2000. Reduction
efficiency of the neurotoxin β-ODAP in low-toxin varieties of Lathyrus sativus seeds by
solid state fermentation with Aspergillus oryzae and Rhizopus microsporus var Chinensis.
Journal of the Science of Food and Agriculture 80: 2209-2215.
Kuo, Y.H., Defoort, B., Getahun, H., Teklehaimanot, R. and Lambein, F .2007.
Comparison of urinary amino acids and trace elements (copper, zinc and manganese) of
recent neurolathyrism patients and healthy controls from Ethiopia. Clinical Biochemistry
40: 397-402.
Kuo, Y.-H., Ikegami, F. and Lambein, F. 2003. Neuroactive and other free amin acids in
seed and young plants of Panax ginseng. Phytochemistry 62: 1087-1091.
Kuo,Y.-H., Ikegami, F. and Lambein, F. 1998. Metabolic route of β-(isoxazolin-5-on-2-
yl)-L-alanine (BIA), the precursor of neurotoxin ODAP (β-N-oxalyl-L-α,β-
diaminopropionic acid), in different legume seedlings. Phytochemistry 49 (1): 43-48.
Kuo, Y.-H., Khan J.K. and Lambein, F. 1994. Biosynthesis of the neurotoxin β-ODAP in
developing pods of Lathyrus sativus. Phytochemistry 35: 911-913.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
136
Kuo, Y.-H., Rozan, P., Lambein, F., Frias, J. and Vidal-Valverde, C. 2004. Effects of
different germination conditions on the content of free protein and non-protein amino acids
of commercial legumes. Food Chemistry 86 (4): 537-545.
Kupicha, F. 1983. The intragenic structure of Lathyrus. Notes Royal Botanical Gardens
41: 209-244.
Kusama-Eguchi, K., Ikegami F., Kusama, T., Suda, A., Ogawa,Y., Igarashi, K. and
Watanabe, K. 2005. A rat model of neurolathyrism: repeated injection of L-β-ODAP
induces the paraparesis of the hind legs. Amino Acids 28: 139-143.
Kusama-Eguchi, K., Suda, A., Ikegami, F., Kusama, T., Ogawa, Y. and Watanabe, K. 2006.
Neurotoxicity and pharmacology of Lathyrus sativus extracts of high- and low-toxicity
strains. Journal of Natural Medicine 60: 107-112.
Kusama-Eguchi, K., Yamamoto, M., Kusama,T., Ito, Y., Ikegami F., Igarashi K., Lambein
F. and Watanabe, K. 2003. Toxicity of 3-N-oxalyl-2,3-diaminopropanoic acid (L-Beta-
ODAP) in rat spinal cord in relation to human neurolathyrism. Korean Journal of
Physiological Pharmacology 7: 64-72.
Lambein, F., Defroot, B., Kuo, Y-.H. 2004. Konzo or neurolathyrism: is there a
difference?. Cassava cyanide diseases network news 3: 2-3.
Lambein, F., Haque, R., Khan, J.K., Kebede, N. and Kuo, Y.H. 1994. From soil to brain:
zinc defficiency increases the neurotoxicity of Lathyrus sativus and may affect the
susceptibility for the motor-neuron disease neurolathyrism. Toxicon 32: 461-466.
Lambein, F., Khan, J.K. and Kuo, Y.-H. 1992. Free amino acids and toxins in Lathyrus
sativus seedlings. Planta Medica 58: 380-381.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
137
Lambein, F., Kuo, Y.H., Ikegami, F. and Murakoshi, I. 1990. Toxic and non-toxic non-
protein amino acids in the Vicieae. In ‘Amino Acids, Chemistry, Biology and medicine
(Eds. G. Lubec and Rosenthal). Pp 21-28.
Lambein, F., Kuo, Y.-H., Kusama-Eguchi, K. and Ikegami, F. 2007. 3-N-oxalyl-L-2,3-
diaminopropanoic acid, a multifunctional plant metabolite of toxic reputation. Arkivoc, ix:
45-52.
Lambein, F., Kuo, Y.H. and Van Parijs, R. 1976. Isoxazolin-5-ones, chemistry and
biology of a new class of plant compounds. Heterocycles 4: 567-593.
Lambein, F., Ngudi, D.D., and Kuo, Y.H. 2001. Vapniarca revisited: Lessons from an
inhuman human experience. Lathyrus Lathyrism Newsletter. 2(1): 5-7.
(http://www.clima.uwa.edu.au/news/lathyrus)
Latif, M.A., Morris, T.R. and Jayne-Williams, D.J. 1976. Use of khesari (Lathyrus
sativus) in chick diets. British Poultry Science 17 (5): 539-546.
Leakey, C. 1979. Khesari Dhal - The poisonous pea. Appropriate Technology 6: 15-16.
Levine, R.L., Moskovitz, J., and Stadtman, E.R. (2000). Oxidation of methionine in
proteins: roles in antioxidant defense and cellular regulation. IUBMB Lif e 50: 301-307.
Long, Y.C, Ye,Y.H., and Xing, Q.Y. 1996. Studies on the neurotoxins beta-N-oxalyl-L-
α,β-diamonopropionic acid its isomer α,β-diaminopropionic acid from the root of Panax
species. International Journal of Peptide Protein Research 47: 42-46.
Loudon. 1855. Encyclopedia of plants. Longman, Brown, Green and Longman, London.
Pp 618-621.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
138
Low, R.K-C., Rotter, R.G., Marquardt, R.R., and Campbell, G.C. 1990. Use of Lathyrus
Sativus L. (var. seminis albi) as a foodstuff for poultry. British Poultry Science 31(3):
615-625.
Ludolph, A. C., Hugon, J., Román, G. C., Spencer, P.S. and Schoenberg, B.S. 1988. A
clinical neurophysiologic study of tropical spastic paraparesis. Muscle & Nerve 11 (4):
392–397.
Ludolph, A. C., Riepe, M. and Ullrich K. 1993. Excitatoxicity, energy metabolism and
neurodegeneration. Journal of Inherited Metabolic Diseases 16 (4): 716-723.
Lwin, S. 1956. Studies in genus Lathyrus. M Sc Thesis, Manchester University.
Masoero, F., Pulimeno, A.M. and Rossi, F. 2005. Effect of extrusion, espansion and
toasting on the nutritional value of peas, faba beans and lupins. Italian Journal of Animal
Sciences 4: 177-189.
Malek, M.A. 1998. Genetic resource of grass pea (Lathyrus sativus) in Bangladesh. In
Mathur, P.N., Ramanatha Rao, V. and Arora, R.K., (eds). 1998. Lathyrus Genetic
Resources Network: Proceedings of IPGRI-ICARDA-ICAR Regional Working Group.
New Delhi, India. pp 7-11.
McCutchan, J.S., Larkin, P.J., Stoutjesdijk, P.A., Morgan, E.R. and Taylor, P.W.J. 1999.
Establishment of shoots and suspension cultures for protoplast isolation in Lathyrus
sativus. SABRAO Journal of Breeding and Genetics 31: 43-50.
Mehta, S.L. and Santha, I.M. 1997. Plant biotechnology for development of non-toxic
strains on Lathyrus sativus. In: Lathyrus Genetic Resources in Asia: Proceedings of a
Regional Workshop, 27-29 December 1995, Indira Gandhi Agricultural University,
Raipur, India. pp 129-138.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
139
Mehta, T., Zarghami, N.S, Parker, A.J. and Cusick, B.E. 1979. Neurotoxicity of orally or
intraperitoneally administered L-3-oxalylamino-2-aminopropionic acid in the mouse.
Toxicology and Applied Pharmacology 48: 1-9.
Misra, U.K., Sharma, V.P. and Singh, V.P. 1993. Clinical aspects of neurolathyrism in
Unnao, India. Paraplegia 31(4): 249-254.
Mohan, V.S., Nagarajan, V. and Gopalan, C. 1966. A simple and specific method for
detection of adulteration with Lathyrus sativus. Indian Journal of Medicinal Research 54:
410-414.
Molvig, L., Tabe, L.M., Eggum, B.O., Moore, A.E., Graig, S., Spencer, D. and Higgins,
T.J.V. 1997. Enhanced methionine levels and increased nutritive value of seeds of
transgenic lupines (Lupinus angustifolus L.) expressing a sunflower seed albumin gene.
Proceedings of the National Academy of Sciences. USA 94: 8393-8398.
Monsoor, M.A. and Yusuf, H.K.M. 2002. In vitro protein digestibility of Lathyrus pea
(Lathyrus sativus), lentil (Lens culinaris), and chickpea (Cicer arietinum). International
Journal of Food Science and Technology 37: 97-99.
Murphy, H.F.1968. A report on the fertility status and other data on some soils of
Ethiopia. Dire Dawa, Ethiopia. pp 135.
Narayan, R.1998. The role of genomic constraints upon evolutionary changes in genome
size and chromosome organization. Annals of Botany 82: 57-66.
Nelson, R.L. 1986. Defining the seed-filling period in soybeans to predict yield. Crop
Science 26: 132–135.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
140
Noji, M. and Saito, K. 2003. Sulphur amino acids: biosynthesis of cysteine and
methionine. In Yash, P. Abol and Altaf Ahmad (eds). Kluwer academic publisher, The
Netherlands. pp 135-144.
Noto, F., Poma, I., Gristina, L., Venezia, G. and Ferrotti, F. 2001. Bioagronomic and
qualitative characteristics in Lathyrus sativus lines. In: Proceedings 4th European
Conference on Grain Legumes (eds. AEP), 8-12 July 2001, Cracow, Poland. pp 183.
Nunn, P.B., James, R.A., Lyddiard, C.P.W., Perera, C. and Bell E.A. 2005. Grass pea-
induced plasma methionine deficiency effects on motor neurons in neurolathyrism.
Journal of Food, Agriculture & Environment 3(2): 347.
Nunn, P.B., Perera, C.K.P.W., Bell, E.A. and Lambein, F. 1994. Appearance of the
neurotoxin β-N-oxalyl-α,β-diaminopropionic acid (β-ODAP) and of homoarginine in
systemic blood and urine following the consumption of cooked Lathyrus sativus by
human volunteers. Archives Internationales de Physiologie et de Biochimie 102, B89.
Ochatt, S.J., Mousset-Declas, C. and Rancillac, M. 2001. Fertile pea plants regenerate
from protoplasts when calluses have not undergone endoreplication. Plant Science 156:
177-183.
Olsen, S.R., Cole, C.V., Watanabe F.S., Dean L. 1954. Estimation of available P in soils
by extraction with sodium bicarbonate. USDA circular, Washington DC.
Padamajaprasad, V., Kaladhar, M. and Bhat, R.V. 1997. Thermal isomerization of β-N-
oxalyl-α,β-diaminopropionic acid, the neurotoxin in Lathyrus sativus, during cooking.
Phytochemistry 59(1): 77-80.
Plitmann, U., Heyn, C.C. and Weinberger, H. 1985. Comparative taxonomy of some wild
species allied to Lathyrus sativus. The Third World Medical Research Foundation, New
York, pp.69-79.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
141
Polignano, G. B, Uggenti, P., Olita, G., Bisignano, V., Alba, V. and Perrino, P. 2005.
Characterization of grass pea (Lathyrus sativus L.) entries by means of agronomically
useful traits. Lathyrus Lathyrism Newsletter 4: 10-14.
Quereshi, M.Y., Pilbeam, D.J.,Evans, C.S. and Bell, E.A.. 1977. Neurolathyrism - α,β-
diaminopropionic acid in legume seeds. Phytochemistry 16: 477-479.
Quevauviller, P., M.Lachica, B.Barahona, G.Raure, A.M.Ure, A.Gomez, A.and H.Mun
au. 1996. Inter laboratory comparison of EDTA and DTPA procedures prior to
certification of extractable trace elements in calcareous soils.Science Total Environment
78:127-132.
Rahman, M.M., Kumar, J., Rahaman, M.A. and Ali Afzal, M. 1995. Natural out-crossing.
Euphytica 83: 33–42.
Ramanujam, K.L., Sethi, K.L. and Rao, S.L.N. 1980. Stability of neurotoxin content in
Khesari. Indian Journal of Genetics and Plant Breeding 40(1): 300-304.
Rao, S.L.N. 1978 . A sensitive and specific colourimetric method for the determination of
α,β-diaminopropionic acid and the Lathyrus sativus neurotoxin. Analytical Biochemistry
86: 386–395.
Rao, S.L.N. 2001. Do we need more research on neurolathyrism? Lathyrus Lathyrism
Newsletter 2(1): 2-4. (http://www.clima.uwa.edu.au/news/lathyrus)
Rao, S.L.N., Adiga, P.R. and Sarma, P.S. 1964. The isolation and characterization of
beta-N-oxalyl-L-alpha,beta-diaminopropionic acid: a neurotoxin from the seeds of
Lathyrus sativus. Biochemistry 3: 432-436.
Renfrew, J. 1979. The first farmer in South-east Europe. Archaeo-physika 8: 243-265.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
142
Rosa, M.J.S., Ferreira , R.B. and Teixeira, A.R. 2000. Storage proteins from Lathyrus
sativus seeds. Journal of Agricultural and Food Chemistry 48: 5432-5439
Rotter, R.G., Marquardt, R.R., Campbell, C.G. 1991. The nutritional value of low
lathyrogenic Lathyrus (Lathyrus sativus) for growing chicks. British Poultry Science 32:
1055-1067.
Roy, D.N., Nagarajam, V. and Gopalan, C. 1963. Production of neurolathyrim in chicks
by the injection of Lathyrus sativus concentrates. Current Science (India) 32: 116-118..
Roy, D.N. and Spencer, P.S. 1989. Lathyrogens: Toxicants of plant origin. In: Cheeks,
P.R. (ed), Proteins and Amino Acids. CRC press Bocca Raton, FL. pp. 3: 170-201.
Roy, P.K., Singh, B., Metha, S.L, Barat, G.K., Gupta, N., Kirti, P.B. and Chopra, V.L.
1991. Plant regeneration from leaf discs of Lathyrus sativus. Indian Journal of
Experimental Biology 29: 327-330.
Rozan, P., Kuo, Y.-H. and Lambein, F. 2001. Amino acids in seeds and seedlings of the
genus Lens. Phytochemistry 58: 281-289.
Rudra, M.N. and Chaudhury, L.M. 1952. Preliminary note on methionine excretion in
lathyrism. Current Science (India) 21(4): 99-100.
Rudra, M.P.P., Singh, M.R., Junaid, M.A., Jyothy, P. and Rao, S.L.N. 2004. Metabolism
of dietary ODAP in humans may be responsible for the low incidence of neurolathyrism.
Clinical Biochemistry 37(4): 318-322.
Rybiñski W. 2001. Mutants of grass-pea (Lathyrus sativus L.) obtained after use of
chemomutagens. Lathyrus Neurolathyrism Newsletter (abstract). 2: 41.
Saito, K. 2004. Sulphur assimilatory metabolism. The long and smelling road. Plant
Physiology 136(1): 2443–2450.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
143
Santha, I.M., Ali, K. and Metha, S.L. 1998. Performance of low ODAP somaclones of
Lathyrus sativus. Lathyrus Genetic Resources Network: Proceedings of an IPGRI-
ICARDA-ICAR Regional Working Group Meeting. New Delhi. pp 74-79.
Schinder, A., Olson, E., Spizer, N., and Montal, M. 1996. Mitochondrial dysfunction is a
primary event in the glutamate neurotoxicity. Journal of Neuroscience 19: 6125-6133.
Shamim, M.Z., Hossain, M.S., Islam, K., Yusuf, H.K.M., Lambein, F. 2002. Mechanism
of ODAP toxicity in one-day-old chicks. Dhaka University. Journal of Biological
Sciences 11: 1-7
Smartt, J. 1984. Evolution of Grain Legumes. I. Mediterranean Pulses. Experimental
Agriculture 20: 275-296.
Smartt, J. 1990. Grain Legumes: Evolution and genetic resources. Cambridge University
Press, Cambridge, UK. pp 200.
Smartt, J., Kaul, A., Araya, W.A., Rahamanand, M.M. and Kearney, J. 1994. Grass pea
(Lathyrus sativus) as a potentially safe food legume crop. In: F.J.Muehlbaurer and
W.J.Kaiser(eds). Expanding the production and use of cool season food legumes. Kluwer
Academic Press, Dordrecht, The Netherlands. pp 144-155.
Smith, J.R. and Nelson, R.L. 1986a. Selection for seed-filling period in soybean. Crop
Science 26: 466–469.
Smith, J.R. and Nelson, R.L. 1986b. Relationship between seed-filling period and yield
among soybean breeding lines. Crop Science 26: 469–472.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
144
Smulikowska A., Rybinski W., Czerwinski J., Taciak M., Mieczkowska, A. 2008.
Evaluation of selected mutants of grass pea (Lathyrus sativus L.) var. Krab as an
ingredient in broiler chicken diet. Journal of Animal and Feed Sciences 17(1): 75-87.
Spencer, P.S, Roy, D.N, Ludolph, A., Hugon, J., Dwivedi, M.P. & Schaumburg,
H.H.1986. Lathyrism: evidence for role of the neuroexcitatory amino acid BOAA. Lancet
2:1066-1067.
Srivastava, S. and Khokhar, S. 1996. Effects of processing on the reduction of β-ODAP
(β-N-oxalyl-L-2, 3-diaminopropanoic acid) and anti-nutrients of khesari dhal (Lathyrus
sativus). Journal of the Science of Food and Agriculture 71(1): 50-58.
Suresh, V.S.R. and Archana, C. 2000. Influence of methionine and zinc on liver collagen
in molybdenotic rats. Biological Trace Element Research 73 (1): 85-91.
Swank J.C., Egli, D.B. and Pfeiffer, T.W. 1987. Seed growth characteristics of soybean
genotypes differing in duration of seed fill. Crop Science 27: 85-89.
Tadelle, D., Alemu, Y., Nigusie, D. and Peters, K. J. 2003. Evaluation of processing
methods on the feeding value of grass pea to broilers. International Journal of Poultry
Science 2(2): 120-127.
Tawali, A.B., Hain, J.U. and Schwedt, G. 1998. Determination of phytic acid content of
soybeans during tempeh production using capillary electrophoresis. Deutsche
Lebensmittel-Rundschau 94: 28–30.
Tews, J.K., Harper, A.E. 1986. Induction in rats of lysine imbalance by dietary
homoarginine. Journal of Nutrition 116: 1910-1921.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
145
Urga, K., Fufa, H., Biratu, E. and Husain, A. 2005. Evaluation of Lathyrus sativus
cultivated in Ethiopia for proximate composition, mineral and β-ODAP and
antinutritional components. African Journal of Food, Agriculture, Nutrition and
Development 5(1): 1-15.
Vaz Patto, M.C, Skiba, B., Pang, E.C.K., Ochatt, S.J., Lambein, F. and Rubiales, D.
2006. Lathyrus improvement for resistance against biotic and abiotic stresses: from
classical breeding to molecular assisted selection. Euphytica 147 (1): 133-147.
Vidal-Valverde, C., Frias, J., Sierra, I., Blazquez, I., Lambein, F. and Kuo, Y.H. 2002.
New functional foods by germination: Effect on the nutritive value of beans, lentils and
peas. European Food Research and Technology 215: 472-477.
Waghmare V.N. and Mehra R.B. 2000. Induced mutations in grass-pea (Lathyrus sativus
L.). Lathyrus Lathyrism Newsletter 1: 21-24.
Wang, F. 2000. HPLC determination of neurotoxin β-N-oxalyl-L-α,β–diaminopropionic
acid and its α-isomer in Lathyrus sativus by precolumn derivatization with 1-fluoro-2,4-
dinitrobenzene. Chinese Chemical Letters 11(5): 435-438.
Wang, X., Warkentin, T. D., Briggs, C. J., Oomah B. D., Campbell, C. G. and Woods, S.
1998a. Total phenolics and condensed tannins in field pea (Pisum sativum L.) and grass
pea (Lathyrus sativus L.). Euphytica 101(1): 97-102.
Wang, X., Warkentin, T.D., Briggs, C.J., Oomah, B.D., Campbell, C.G. and Woods, S.
1998b. Trypsin inhibitor activity in field pea (Pisum sativum L.) and grass pea (Lathyrus
sativus L.). Journal of Agricultural and Food Chemistry 46(7): 2620-2623.
Williams, P.C., Bhatty, R.S., Deshpande, S.S., Hussein, L.A. and Savage, G.P. 1994.
Improving nutritional quality of cool season food legumes. In: Muehlbauer, F.J. and

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
146
Kaiser, W.J (eds.).1994. Expanding the Production and Use of Cool Season Food
Legumes. Kluwer Academic Publishers, Dordrecht Netherlands. pp 113-129.
Williams, S. 1984. Moisture. Official Methods of Analysis, AOAC.
Williams, W. A, Jones, M.B. and Demment, M.W. 1990. A concise table for path
analysis statistics. Agronomy Journal 82: 1022-1024.
Wuletaw, T. 2003. Stability of grass pea (Lathyrus sativus L.) varieties for β-ODAP
content and grain yield in Ethiopia. Lathyrus Lathyrism Newsletter 3: 32-34.
Xie, G.-X., Qiu, Y.-P., Qiu, M.-F., Gao, X.-F., Liu, Y.-M. and Jia, W. 2007. Analysis of
dencichine in Panax notoginseng by gas chromatography-mass spectrometry with ethyl
chloroformate derivatization. Journal of Pharmaceutical and Biomedical Analysis 43:
920-925.
Yamamoto, K., Fujiware, T. and Blumenreich, I.D. 1986. Isozyme variation and
interspecific crossability in annual species of the genus Lathyrus L. In: Lathyrus and
lathyrism. Kaul, A.K.and Combes, D. (eds). The third medical research foundation, New
York. pp 118-129.
Yan, Z.Y, Spencer, P.S., Li, Z.X., Liang Y. M., Wang, Y.F., Wang, C.Y., and Li, F.M.
2006. Lathyrus sativus (grass pea) and its neurotoxin ODAP. Phytochemistry 67: 107-
121.
Yigzaw, Y., Gorton, L., Akalu, G. and Solomon, T. 2001. Fermentation of teff
(Eragrostis tef), grass-pea (Lathyrus sativus), and their mixtures: Aspects of nutrition and
food safety. Lathyrus Lathyrism Newsletter 2(1): 8-10.
Yunus, A.G. and Jackson, M.T. 1991. The gene pool of the grass pea (Lathyrus sativus
L). Plant breeding 106: 319-328.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
147
Yusuf, H.K.M., Hoque, K., Uddin, A., Roy, B.C. and Lambein, F. 1995. Homoarginine
antagonizes the toxicity of Lathyrus toxin in 1-day-chicks. Bangladesh Journal of
Physiology and Pharmacology 10(2): 74-75.
Zambre, M., Chowdhury, B., Kuo Y.H, Van Montagu, M., Angenon, G. and Lambein, F.
2002. Prolific regeneration of fertile plants from green nodular callus induced from
meristematic tissues in Lathyrus sativus L (grass pea). Plant Science 163: 1107-1112.
Zhang, J., Xing, G. M., Yan, Z. Y. and Li, Z. X. 2003. β-N-Oxalyl-L-α,β-
Diaminopropionic Acid Protects the Activity of Glycolate Oxidase in Lathyrus sativus
Seedlings under High Light. Russian Journal of Plant Physiology 50(5): 618-622.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
148

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
149
Summary - Samenvatting

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
150

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
151
Summary
Grass pea (Lathyrus sativus L.) is economically the most important species in the genus
Lathyrus. Hundreds of millions of people in Asia, Africa and the Near East use grass pea
in their daily diet. Typically and unlike other legumes, it is used as a staple food. Its
adaptation to environmental stresses, productivity under low inputs, and capacity to
restore soil fertility capacity underline its agricultural potential. Despite its hardiness it is
a dependable and promising source of protein, carbohydrate and minerals. Grass pea is
used as a pulse since the early history of agriculture and is still so used. However, it has
made little progress as a grain crop during this long history.
Unbalanced overconsumption of grass pea during months can lead to neurolathyrism, an
irreversible crippling of the legs in humans. The disease can become epidemic,
particularly if accompanied by malnutrition, drought, insufficient processing or raw
consumption. The causative metabolite is a neuro-excitatory amino acid β-N-oxalyl-L-α-
β-diaminopropionic acid (β-ODAP). In grass pea this metabolite is highly affected by
environmental factors, processing and genetic factors. Hence, the level of β-ODAP in the
seed seems to be related to a physiological reaction of the plant to environmental stresses
and may be considered as a stress metabolite. Methionine has a protective effect against
the toxicity of grass pea, but is very limited in this plant.
The study tries to integrate the environmental and biological aspects towards the same
goal of improving the nutritional quality of grass pea. In the first part of the thesis
interrelations between amino acids in grass pea and environmental conditions were
studied. Unique among legume species, grass pea is an important source of the non-
protein amino acids homoarginine, followed in concentration by the presumed cause of
neurolathyrism ß-ODAP. These metabolites are suggested to confer the plant with some
degree of tolerance against different biotic and abiotic stresses. There was big variation of
ß-ODAP level in response to genotypes and environments. Grass pea has a sufficient
level of most essential amino acids but methionine is extremely low, covering less than
25% of the recommended level for a healthy diet. Methionine levels are less variable than

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
152
ß-ODAP in grass pea. Path-analysis with selected predictors for ß-ODAP showed a larger
direct effect due to K+, high light intensity during maturity phase (sshIII), maturity, yield
and Zn2+:P. Ethiopian soil is often very high in potassium, and this has been
demonstrated to have a strong enhancing activity for β-ODAP in Ethiopia. Pearson’s
correlation coefficient showed that altitude, maturity period, soil pH and crop yield have
a strong negative correlation while K+ and shh have strong positive correlation with β-
ODAP. High light intensity, supposedly found at higher elevation, is found to increase β-
ODAP biosynthesis. However, in this study the effect of high light intensity in high land
was modulated by the cloudiness and thus shorter period of high light intensity resulting
in higher ssh in lowlands producing higher β-ODAP than high lands with reduced total
ssh in the plant growth periods. Apart from these we found the greatest direct effect of
maturity period (0.94) followed by rainfall during crop establishment (0.31) for crop
yield. There was also a significance of Genotype x Environment combination for grass
pea yield, indicating the different responsiveness of varieties to the varying environment.
The effect of methionine on grass pea toxicity was investigated using broiler chicks. The
responses of chicks to dietary DL-methionine supplements to grass pea feed were
evaluated. This supported the hypothesis that the antioxidant activity of methionine
counteracted the oxidative stresses generated by various physiological activities of β-
ODAP. In agreement with other reports, some chicks developed reversible convulsions of
the neck region at early age and showed resistance afterwards. Symptoms occurred only
in maximum 1/3rd of chicks receiving the same exposure to β-ODAP, consistent with
earlier suggestions that individual differences exist. There was a much-reduced level of
β-ODAP in the chick’s serum compared to the corresponding intake level. This might
suggest the reduced uptake or increased metabolic breakdown in the presence of higher
methionine, which might also have individual variation. The improved performances of
the chicks in response to methionine supplementation was partially due to the indirect
effect through FCE improvement. It was learned that the tolerance level of the animals
remarkably improved with age, suggesting that adaptation and stronger detoxification
mechanism might have developed. There was a trend of ß-ODAP reduction with

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
153
increasing methionine supplement, suggesting the counteracting nature of the two amino
acids.
Methionine enrichment in grass pea is perceived as a key element to reduce
neurolathyrism development. Mutation and selection for increased methionine has been
adapted from similar work with other legumes. Three phenotypically distinct putative
mutant lines have been selected from gamma-irradiated grass pea seeds. The M4
regenerants of these putative mutant lines have 63% higher methionine in the seed
compared to the control parental line. Characterization of the three selections showed that
they are sub-erect with medium biomass, have comparably smaller and deeper-glittering
green leaves, medium seed size, and reasonable grain productivity. They were designated
as deep greens (DGA, DGB, DGC) referring to their typical phenotype. However, the ß-
ODAP level in the new mutants is not different from the wild type. The methionine level
in the mutant lines may improve the nutritional quality and safety of the seed in food and
feed. Hence, the market price of grass pea and the financial situation of the farmers may
improve. These lines would serve as attractive genetic resource for future breeding.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
154

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
155
Samenvatting
Zaailatyrus (Lathyrus sativus L.) is economisch een van de meest interessante
peulvruchten binnen het genus Lathyrus. Het onderscheidt zich van andere peulvruchten
door het feit dat het door honderden miljoenen mensen in Azië, Afrika en het
Middenoosten dagelijks als voedsel gebruikt wordt. Het aanpassingsvermogen van deze
peulvrucht aan stress uit de omgeving, de hoge opbrengst die relatief weinig moeite kost,
en de capaciteit om de bodem te verbeteren, geeft zaailatyrus een toekomstperspectief in
het milieu van de toekomst met meer droogte en verzilting. Naast de mogelijkheid om in
extreme omstandigheden te overleven is het ook een rijke bron van proteïnen,
koolhydraten en mineralen. Toch heeft zaailatyrus, dat sinds millennia als peulvrucht
wordt gebruikt en tot nu toe weinig uitgebuit werd, tot op heden als landbouwgewas
weinig vooruitgang geboekt.
Wanneer mensen maandenlang afhankelijk zijn van de consumptie van zaailatyrus kan
dit leiden tot neurolathyrisme, een irreversibele verlamming van de onderste ledematen.
Deze aandoening kan voorkomen onder de vorm van een epidemie, voornamelijk in
combinatie met ondervoeding, droogte, inefficiënte verwerking of consumptie van de
rauwe zaden. Het vrij aminozuur β-N-oxalyl-L-α-β-diaminopropionzuur (β-ODAP) in
zaailatyrus wordt verantwoordelijk geacht voor die aandoening. De concentratie van dit
aminozuur in de zaden wordt sterk beïnvloed door omgevingsfactoren, de
bereidingswijze van de zaden of genetische factoren. Zo blijkt er een verband te bestaan
tussen de concentratie van die ongewone metaboliet en diverse omgevingsfactoren.
Methionine, dat deficiënt is in peulvruchten en vooral in latyrus kan bescherming bieden
tegen de neurotoxische werking van langdurige latyrus overconsumptie; waarschijnlijk
als beschermer tegen oxidatieve stress.
Deze studie probeert de socio-economische aspecten te integreren met de
omgevingsgerelateerde en biologische aspecten met het oog op de preventie van
neurolathyrisme en de verbetering van de voedingswaarde van zaailatyrus. In het eerste

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
156
deel beschrijven we een grondig literatuuroverzicht waarbij de aandacht gaat naar
ontbrekende schakels in onze kennis van het lathyrisme en de consumptie van latyrus.
In het tweede deel van de studie werden onderlinge verbanden aangetoond tussen de
niveau’s van aminozuren in zaailatyrus en de omgeving. In vergelijking met andere
peulvruchten is zaailatyrus een unieke bron van het vrije aminozuren homoarginine,
kwantitatief gevolgd door het vermoedelijk toxische ß-ODAP. Deze moleculen worden
verondersteld het gewas een brede tolerantie te verlenen tegen verschillende vormen van
omgevingsstress. Het ß-ODAP gehalte varieerde sterk naargelang het genotype, ecotype
en de soort voedselbereiding. Zaailatyrus bevat voldoende essentiële aminozuren,
behalve methionine en cysteine. Het methioninegehalte is heel laag, namelijk minder dan
25% van de normale behoefte, en het gehalte is minder variabel dan dat van ß-ODAP. In
optimale groeimedia vertonen beide aminozuren een duidelijk tegengestelde gunstige
wijziging (methionine stijgt en ß-ODAP daalt), wijzend op de mogelijkheid tot simultane
verbetering van deze twee negatieve factoren in de voedingskwaliteit van de plant. De
stress-vrije conditie zou de plant minder stimuleren om ß-ODAP te synthetiseren en meer
aanzetten tot de biosynthese van essentiële aminozuren. Er werd ook aangetoond dat de
manier van bereiding van voedsel de twee aminozuren gunstig beïnvloedt, mogelijk door
het uitwassen van het toxine. Pad-analyse met geselecteerde predictoren toonde tevens
een groter direct effect van K+, uren zonlicht, maturiteit, opbrengst en de verhouding
Zn2+:P op de synthese van ß-ODAP. Er werd aangetoond dat K+, dat vaak in overmaat
aanwezig is in de Ethiopische bodem, de hoeveelheid ß-ODAP sterk kan verhogen. De
Pearson’s correlatiecoefficiënt toonde een sterk negatieve correlatie voor de hoogte, de
maturiteitsperiode, de pH van de bodem en de opbrengst van het gewas, terwijl K+ en shh
een sterk positieve correlatie met ß-ODAP vertonen. De hogere lichtintensiteit op grotere
hoogte, zou de biosynthese van ß-ODAP verhogen. Toch bleek in deze studie de invloed
van het licht redelijk compenserend te zijn, aangezien de lager gelegen percelen in totaal
meer uren zon hadden en de planten er meer ß-ODAP produceerden dan in het gebergte
met een verminderde totale duur zonneschijn in de groeiperiode van de plant. Naast deze
effecten waren de maturiteitsperiode (0.94), gevolgd door regenval tijdens de
ontwikkeling van het jonge het gewas (0.31), de voornaamste directe effecten op de

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
157
opbrengst van het gewas. Ook de combinatie Genotype x Omgeving vertoonde een
significant effect op de opbrengst van zaailatyrus, wijzend op de verschillende reactie van
de variëteiten op een veranderende omgeving.
Het effect van methionine op β-ODAP toxiciteit werd onderzocht aan de hand van
kuikens die gevoed werden met zaailatyrus. De fysiologische en gedrags-responsen van
de kuikens op toevoeging van DL-methionine aan hun dieet van zaailatyrus werden
onderzocht. Dit bevestigde dat de antioxiderende werking van methionine de oxidatieve
stress van β-ODAP tegenwerkt. Naar analogie met andere rapporten verkregen de
kuikens het reversibel convulsief torticolis van de nek bij jonge leeftijd en vertoonden
resistentie bij oudere leeftijd. Symptomen traden enkel op bij een derde van dezelfde
groep kuikens die werd blootgesteld, consistent met eerdere suggesties dat er individuele
verschillen bestaan. Er was een sterk verminderde hoeveelheid β-ODAP in serum ten
opzichte van de ingenomen hoeveelheid, wat fysiologische mechanismen kan suggereren
die kunnen bijdragen tot individuele efficiëntie om zich te wapenen tegen de ziekte. De
verbeterde resistentie van de kuikens bij toevoeging van methionine was deels te wijten
aan de indirecte verbetering van de voedselconversie. Er werd aangetoond dat de graad
van tolerantie van de dieren opmerkelijk verbeterde met de leeftijd, wat kan wijzen op
aanpassing en ontwikkeling van sterkere detoxificatiemechanismen. Bij kippen met
neurologische symptomen werd veel meer β-ODAP aangetroffen in het serum naast een
veel lagere concentratie methionine, wat de antagonistische aard van deze twee
aminozuren suggereert.
Het verrijken van zaailatyrus met methionine wordt beschouwd als een cruciale factor om
het risico van neurolathyrisme te verlagen. Het draagt bij tot de verbetering van de
kwaliteit van het proteïne. Uit gamma-bestraalde zaden werden drie mutantlijnen
geselecteerd met een verschillend fenotype. De R4 regeneraties van de vermeende
mutantlijnen vertonen een verhoging tot 63% van de methionineconcentratie in de zaden
in vergelijking met de controle parentale lijn. Karakterisatie van de selecties toonde aan
dat ze minder rechtop staan met een medium biomassa, dat ze relatief kleinere en
glanzende groene bladeren hadden, medium zaadgrootte en een redelijke graanproductie.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
158
Ze werden gedefinieerd als diepgroen A-C (DGA, DGB, DGC) overeenkomstig met hun
eigen fenotype. Toch was het β-ODAP gehalte in de nieuwe mutanten niet verschillend
van het wildtype. Het methioninegehalte in de mutante lijnen zou de capaciteit van
zaailatyrus als voeding substantieel kunnen verbeteren en zijn nutritionele kwaliteit en
veiligheid bevorderen. Deze lijnen zouden in de toekomst kunnen gebruikt worden als
genetische bron voor verder onderzoek op de nutritionele kwaliteit van zaailathyrus.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
159
CURRICULUM VITAE
Full name: Asnake Fikre Woldemedhin Date of birth: 7/12/1972 Birth: Ethiopia, Arsi Citizenship: Ethiopian
Institution: Ethiopian Institute of Agricultural Research /Debre Zeit Center/ Training: Degree
University Duration (date/mon./year) Major field
B.Sc Alemaya Sep. 1, 1989- Sep. 5, 1992 Plant Science M.Sc Alemaya Sep. 1, 1997- July 10, 1999 Agronomy/Breeding PhD Ghent Oct. 2004- Sep. 2008 Applied biological Sciences Other trainings
Place Duration (date/mon./year)
Field of activity
Israel, Mashave
24Nov-15Dec 2004 Ecological Considerations for Sustainable Agricultural Development Projects
ICARDA, Syria
5-9, May, 2002
Specialized training course in pulse breeding and selection
EARO, Melkasa
24-29, Sep. 2001 Training workshop on research planning, monitoring and evaluation
DZARC Feb.-April (two months), 2001 Computer application program Work experience
Position Duration (month & year)
Field of work
Enumerator household survey Dec.-Feb., 1992 Survey on different livelihood aspects of the households in the rural areas data analysis
Junior expert Mar.1993-Oct.1995 Coffee agronomy Expert Dec. 1995- Jan.1997 Crop agronomy and protection Team leader in Mult. And Dissemination of horticultural crops
Feb.-Augst, 1998 Coordinating multiplication of horticultural crops & coffee seedlings
Head Zonal Planning and Economic Development department
March 1999-July 2000 Coordinating and leading integrated developmental activities among different sectors as per the TOR
Pulses breeder Aug 2000- Nov2002 Assistant researcher position design, plan and conducting experiments
National Lentil Project Coordinator and breeder (Asso. researcher)
Dec, 2001-Oct. 2004 Coordinating and leading the program at national level as per the TOR, and conducting breeding and agronomy experiments

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
160
Publications
Fikre A ., Korbu L., Kuo Y.-H., Lambein F. (2008) The contents of the neuro-excitatory amino acid ß-ODAP (β-N-oxalyl-L-α,β-diaminopropionic acid), and other free and protein amino acids in the seeds of different genotypes of grass pea (Lathyrus sativus L.). Food Chemistry 110: 422-427. Fikre, A , F. Lambein, F, and Gheysen, G. 2006. A life saving food plant producing more neurotoxin under environmental stresses. Comm. Appl. Biol. Sci. Ghent University, Vol 71(1) 79-82. Fikre, A . 2007. Grass pea. A research review handbook. Ethiopian Institute of Agricultural Research, Addis, Ababa. Fikre, A. and Bejiga, G. Breeding lentil for wider adaptation. In Ali K, Keneni G., Ahmed S., Malhautra R., Bainwal S., Makkouk K and Halila M.H. (eds). 2006. Food and forage legumes of Ethiopia; progress and prospects. Proceeding of the workshop on food and forage legumes22-23 sep. 2003. Addis Ababa, Ethiopia. Fikre, A.,Sarker, A., Ahmed, S.,Ali, K., Halila, H. and Erskine, W. 2006. Science for the poor: an example of lentil research for the Ethiopian (in press), Syria, Aleppo. Getachew A., Asnake F. and Ayalew B. Soil fertility and crop management in highland pulses in central highland of Ethiopia. In Ali K, Keneni G., Ahmed S., Malhautra R., Bainwal S., Makkouk K and Halila M.H. (eds). 2006. Food and forage legumes of Ethiopia; progress and prospects. Proceeding of the workshop on food and forage legumes22-23 sep. 2003. Addis Ababa, Ethiopia. Legesse D., Senait R., Asnake F and Demisie M., 2005. Adoption of chickpea varieties in the central high lands of Ethiopia. Research reports no 62. EARO, Addis Ababa. Senait R., Legesse D., Demisie M., Asnake F and Aden A.H. 2005. Impact of research and technologies in selected lentil-growing areas of Ethiopia. Research reports no 67. EARO, Addis Ababa. Malhotra, R.S., Bejiga, G., Anbessa, Y., Eshete, M., Taddesse, N., Daba, K., Fikre, A ., Ahmed, S. and Khalaf, G. 2007. Registration of 'Teji' a kabuli Chick pea. Journal of plant registration. Crop Science Society of America, USA 1:111. Malhotra, R.S., Bejiga, G., Anbessa, Y., Eshete, M., Taddesse, N., Daba, K., Fikre, A., Ahmed, S. and Khalaf, G. 2007. Registration of 'Ejere' a kabuli Chick pea. Journal of plant registration. Crop Science Society of America, USA 1:112.

Im proving the nutritional quality of grass pea (L athyrus sativus L.)
161
Poster presentations and abstracts Lambein, F., Diasolua, D.N, Fikre, A , Banea Mayambu, J.-P and Kuo Y-H. 2008. Nutritional improvement of grass pea and cassava to prevent lathyrism and konzo. Science with Africa, March 3-7, 2008 Addis Ababa, Ethiopia. Abstract. Lambein, F., Fikre , A. Van Moorhem, M. and Kou, Y.-H. 2007. Lathyrus sativus (grass pea), from toxic plant to wonder crop. Integrating legume biology for sustainable agriculture. Lisbon, Portugal. Abstract. Fikre, A., Lambein, F., and Gheysen, G. 2006. A life saving food plant producing more neurotoxin under environmental stresses. Comm. Appl. Biol. Sci. Ghent University, Vol 71(1) 79-82. Poster presentation. Fikre, A . 2007. Intervention, gap analysis and applicability with regard to grass pea utilization in Ethiopian context. Seminar presentation. DZARC, Ethiopia, Aug 18-22, 2007. Fikre, A., Ahmad, S., Lambein, F. and Gheysen, G. 2007. The incidence of neurolathyrism is strongly linked to age, gender and grass pea food type in Ethiopia. First GAP Symposium: Heritage and/as reproduction in Africa: outcomes and limits, Ghent University. Poster presentation and abstract Fikre, A., Yami, A., Kuo, Y.-H. and Lambein, F. 2007. Protective effect of methionine on acute neurolathyrism in chicks. First GAP Symposium: Heritage and/as reproduction in Africa: outcomes and limits, Ghent University. Poster presentation and abstract. Van Moorhem, M., Fikre, A ., Lambein, F. and Leybaert, L. 2008. Lathyrus sativus (grass pea), from toxic plant to wonder crop. Science with Africa, March 3-7, 2008 Addis Ababa, Ethiopia. Abstract.
Copyright © 2022 FDOKUMEN




















