
The compensatory growth response of the Atlantic cod: effects of nutritional history

Ti
MMa
b
c
a
ARRAA
KSRLMF
1
AceiatBvrfat(
M
F
h
h0
Behavioural Processes 119 (2015) 50–57
Contents lists available at ScienceDirect
Behavioural Processes
jo ur nal home p ag e: www.elsev ier .com/ locate /behavproc
he compensatory effect of regular exercise on long-term memorympairment in sleep deprived female rats
aryam Salari a,b, Vahid Sheibani a,b,∗, Hakimeh Saadati c,∗∗, Alimohammad Pourrahimi a,ohammad khaksarihadad a,b, Khadijeh Esmaeelpour a, Mehdi Khodamoradi a
Neuroscience Research Center, Kerman University of Medical Sciences, Kerman, IranDepartment of Physiology, School of Medicine, Kerman University of Medical Science, Kerman, IranFaculty of Medicine, Ardabil University of Medical Sciences, Ardabil, Iran
r t i c l e i n f o
rticle history:eceived 20 April 2015eceived in revised form 20 June 2015ccepted 25 June 2015vailable online 17 July 2015
eywords:
a b s t r a c t
Previous studies have been shown that exercise can improve short-term spatial learning, memory andsynaptic plasticity impairments in sleep deprived female rats. The aim of the present study was to inves-tigate the effects of treadmill exercise on sleep deprivation (SD) induced impairment in hippocampaldependent long-term memory in female rats. Intact and ovariectomized female rats were used in thecurrent study. Exercise protocol was 4 weeks treadmill running. Twenty four hour SD was induced byusing multiple platform apparatus after learning phase. Spatial learning and long-term memory was
leep deprivationegular exerciseong-term memoryorris water maze
emale rats
examined by using the Morris Water Maze (MWM) test. Our results indicated that sleep deprivationimpaired long term memory in the intact and ovariectomized female rats, regardless of reproductive sta-tus (p < 0.05) and treadmill exercise compensated this impairment (p < 0.05). In conclusion the results ofthe current study confirmed the negative effect of SD on cognitive functions and regular exercise seemsto protect rats from these factors, however more investigations need to be done.
. Introduction
Sleep plays an important role in normal physiological functions.lthough its functions remain elusive, sleep may be a fundamentalontributor of memory consolidation. Additionally, a key hypoth-sis is that post-training sleep facilitates consolidation of newnformation into long-term memory (Datta, 2010; Diekelmannnd Born, 2010; Walker and Stickgold, 2004) .Sleep also con-ributes to neuronal plasticity (Cai et al., 2009; Diekelmann andorn, 2010). For example, it has been shown that sleep depri-ation inhibits long-term potentiation (LTP) in the CA1 area ofat hippocampus (Davis et al., 2003). Sufficient sleep is necessaryor fostering of connections in neuronal networks (Diekelmann
nd Born, 2010; Gais et al., 2006). Previous studies have shownhat SD destroyed hippocampal dependent learning and memoryAleisa et al., 2011). SD is widespread in various occupations and∗ Corresponding author at: Neuroscience Research Center, Kerman University ofedical Sciences, Kerman, Iran. Fax: +98 341 2264198.
∗∗ Corresponding author at: Ardabil University of Medical Sciences, Ardabil, Iran.ax: +98 455510057.
E-mail addresses: [email protected], [email protected] (V. Sheibani),[email protected] (H. Saadati).
ttp://dx.doi.org/10.1016/j.beproc.2015.06.014376-6357/© 2015 Published by Elsevier B.V.
© 2015 Published by Elsevier B.V.
individuals, for example stressful occupations (Landrigan et al.,2004). Sleep disorders caused by various diseases and contributesto decrease in work/school efficiency in modern societies (Leconteand Bloch, 1970; Philip and Akerstedt, 2006). Most sleep stud-ies have been performed in males; however, the effects of SD onshort-term memory in females are investigated in a few studies(Alhola et al., 2005; Hajali et al., 2012). It is stated that post-menopausal women are vulnerable to cognitive deficiencies (Greenand Simpkins, 2000; Hogervorst et al., 2000). Hajali et al. also indi-cated that female rats are more susceptible to memory impairmentinduced by SD than male animals (Hajali et al., 2012). Hormonereplacement therapy (HRT) is a common way to treat menopausalsymptoms, such as cognitive dysfunction. However it has sideeffects, especially the increased incidence of uterine cancer andneoplasm of the breast (LeBlanc et al., 2001; Miquel et al., 2006).In addition, because of the fluctuations of female hormone levelsobserved during the estrus cycle, estrus cycle phase may be animportant factor when working with female animals (Colvin et al.,1968; Sell et al., 2000). Evidence indicated that physical activity,such aerobic exercise, has appeared as a promising low cost treat-
ment to improve neurocognitive function that is available to mostadults and is not involved by intolerable side effects mostly foundwith pharmaceutical treatments (Hillman et al., 2008). In the otherhand, numerous studies have shown that exercise has beneficial
ral Pro
eeb(mpamdtsp2isdtoohif
2
2
ff(ctsefooaswGo(
2
uamtf
2
hw0tbtc(
M. Salari et al. / Behaviou
ffects on memory and also on menopausal symptoms (Saadatit al., 2014a,b, 2015; Shangold, 1990). Regular physical activity haseneficial effects on cognitive function in both humans and rodentsBerchtold et al., 2005; Cotman and Berchtold, 2002). One of the
ost main effects of exercise is the effect on cognition, for exam-le, it raises learning and improves memory retention (Cotmannd Berchtold, 2002; Van Praag, 2009). Exercise training can incre-ent cerebral blood volume (Cotman and Berchtold, 2002; Van
er Borght et al., 2009). Several studies have reported the dele-erious effects of sleep deprivation on cognitive performances andignaling molecules in the hippocampus of male rats. Meanwhile,hysical exercise attenuates these impairments (Zagaar et al., 2013,012). Our previous studies indicated that regular exercise can
mprove spatial learning and memory (Saadati et al., 2015) andynaptic plasticity (Saadati et al., 2014b) impairments in sleepeprived female rats. In addition, our previous data demonstratedhat exercise can reverse the decreased effects of sleep deprivationn brain-derived neurotrophic factor levels in the hippocampus ofvariectomized female rats (Saadati et al., 2014a). Therefore, ourypothesis is regular exercise has preventive effects against SD-
nduced spatial long-term memory impairment in intact and OVXemale rats.
. Methods
.1. Animals
Female Wistar rats with an average weight of 200 g were usedor the present study. Animals were caged in groups of four withree access to food and water. The temperature was controlled23 ± 1 ◦C) and they were also housed under a 12-h light–darkycle (lights on: 07:00–19:00 h). Two groups of intact and ovariec-omized (OVX) rats were accidentally selected, and the followingubgroups were formed: control (stayed in home cages), SD,xercise, exercise/SD, sham exercise and wide platform (sham plat-orm). A separate group of rats was also submitted as a sham surgeryr sham ovariectomy (submitted to surgery without removing thevaries) (n = 8 for each group). The rats of the OVX group underwent
bilateral ovariectomy. Estrus cycles of all the intact groups wereynchronized by maintaining in the same cages. All proceduresere performed inconformity with the National Research Council’suide for the care and use of laboratory animals and on approvalf the Ethics Committee of Kerman Neuroscience Research CenterEthics Code: KNRC-91-33).
.2. Surgical procedures
All of the operations were performed under general anesthesiasing a mixture of ketamine and xylazine (60 mg/kg, i.p.ketaminend 10 mg/kg, i.p. xylazine). Both ovaries were eliminated by a smallid-abdominal cutting under aseptic conditions. All of the ovariec-
omized rats and sham surgery groups were put in a special roomor one month after operation (Saadati et al., 2014b, 2015).
.3. Treadmill exercise
For four weeks from saturday to wednesday, the exercise groupsad force exercise sessions (at 0◦inclination) during the light cyclehich started from 9:00 till 14:30 (they received a mild shock,
.25 MA, while they stopped running). They were allowed to adapto treadmill environment for 30 min during 2 successive days
efore the onset of the exercise protocol, this was to eliminatehe possible stress of the new environment. The exercise proto-ol included the following steps: 30 min for the first two weeksat 10 m/min speed), 45 min for the third week and 60 min for thecesses 119 (2015) 50–57 51
fourth week (both at 15 m/min speed). Every 15 min during eachsession, the animals were given a 5 min break (Zagaar et al., 2012).
2.4. Induction of sleep deprivation
We used a multiple platform apparatus to induce SD(Zagaar et al., 2012; Hajali et al., 2015a,b). This apparatus(90 cm × 50 cm × 50 cm) included 10 columns (10 cm high,7 cmdiameter located 2 cm above the surface of the water) which wereordered in two rows and spaced 10 cm separate (edge to edge),this was to permit rats to jump from one platform to another. Thecage mates (4 rats) were put each other in a chamber to maintainsocial stableness. The rats had free access to clean water bottles,and food pellet baskets were always hanging from the top of thechamber. In the current research, SD was induced for 24 h. Animalswere kept under standard conditions [12:12-h light–dark cycle at acontrolled temperature (23 ± 1 ◦C)] in the sleep deprivation period(24 h). We carried out the SD sample for 24 h after performing thelast exercise session in the exercise/SD groups. The possible effectsof new environmental stress were assessed by putting the control(sham platform or wide platform) groups in a similar chamber butwith wider platforms (10 cm high, and 15 cm in diameter). The plat-forms were large enough so that the rats would not downfall intothe water during their sleep period.
2.5. Morris water maze (MWM)
The MWM included of a black circular pool, 160 cm diameter,80 cm height filled with water held at room temperature to a depthof 40 cm. The pool was geographically divided into four quadrantsof equal size and starting points were designated at each quadrantas N, S, E, and W. A square platform (10 cm diameter) was invis-ible just below (1.5 cm) the surface of the water in the center ofthe northeast quadrant. The experiments were carry out in a dimlylight room with different and fixed extra maze geometric images(e.g., circles, squares or triangles) attached at different points onthe walls around the maze. Performances were recorded by a smartvideo tracing system (NoldusEthovision® system, version 5, USA)and animals could be tracked on the screen of a computer.
2.6. Spatial learning and memory
In a single training protocol each rat accomplished three blocksseparated by a 30-min resting period before sleep deprivation. Eachblock consisted of four successive trials with 60 s duration andabout 60 s inter-trial intervals. All of the experimental groups weretested ∼30 min after the end of the SD period, during the lights onphase between 8:30 and 12:00 am. On each trial, rats were ran-domly released into the water from one of the four quadrants ofthe maze with its face toward the wall of the quadrant where it wasreleased. Each rat had 4 different releasing points. During acquisi-tion, the location of the platform remained constant and rats wereallowed to swim to the hidden escape platform. After the animalfound the platform, it was allowed to remain there for 20–30 s andwere then located in an animal cage to wait 20–30 s before the startof the next trial. While a rat failed to find the platform in 60 s, theexperimenter guided it toward the platform. The time and distanceto find the hidden platform were collected and analyzed later.
A single probe trial was given 24 h after the last training trial totest the spatial memory in the water maze after sleep deprivation.In this trial the platform was removed and rat was allowed to swimfor 60 s. The time and distance spent in the target quadrant (quad-
rant 4) were analyzed as a measure of spatial memory retention.Following the probe trial, rats completed a visible platform testto determine any possibility of SD interference with sensory andmotor coordination or motivation. In this test, the animals ability

52 M. Salari et al. / Behavioural Processes 119 (2015) 50–57
F rning
b
tw(b
2
ndtoucppncn
ct(t
ig. 1. The mean path length swum to find the hidden platform during spatial leaefore sleep deprivation (SD). Each blocks consisted of four training trials.
o escape to a visible platform (platform was raised 2 cm above theater level and became visible with aluminum foil) was evaluated
Hajali et al., 2012). During the procedure, the experimenter waslind to the group membership of the rats.
.7. Vaginal cytology
To determine the estrus cycle phase of each female rat, vagi-al smears were taken between 08:00 and 09:00 AM on all testingays. A plastic smear loop was inserted in the vaginal opening, gen-ly rotated and withdrawn. The smear loop was immediately rollednto a glass slide and allowed to air dry. Slides were examinednder a light microscope for the presence of nucleated epithelialells, cornified epithelial cells, leukocytes, and mucus. Estrus cyclehase was determined using the following criteria; (1) pro-estrus:redominantly nucleated epithelial cells; (2) estrus: predomi-antly cornified epithelial cells; (3) met-estrus: cornified epithelialells and leukocytes; (4) di-estrus: predominantly leukocytes, someucleated epithelial cells and mucus.
Female rats in pro-estrus and estrus formed one group (pro/es;
haracterized by higher levels of estradiol and progesterone), andhe other group consisted of female rats in met-estrus and di-estrusmet/di; characterized by lower levels of estradiol and proges-erone). Similar subdivisions have been described in other studyin the Morris water maze test in the intact (A) and ovariectomized (B) female rats
investigating the role of estrus cycle in behavior (Van Goethemet al., 2012).
2.8. Data analysis
The escape latency and path length to find the hidden platformin the MWM training in the acquisition phase were analyzed usinga two-way analysis of variance (ANOVA) along with repeated mea-sures to find out the differences of the learning rates of the groups.All of the collected data from the MWM probe trials and swim speedlevels were analyzed by a one way and two-way analysis of variance(ANOVA). When statistical significance was found between groups,Tukey’s post hoc multiple comparison tests were performed todetermine points of significant difference. The data were expressedas means ± standard error of mean (SEM) and P < 0.05 was consid-ered statistically significant.
3. Results
3.1. Spatial learning
Before sleep deprivation, all groups were trained in the Morriswater maze for evaluation of spatial learning before sleep depriva-tion. As indicated in Figs. 1 and 2A and B, all subgroups of intact and

M. Salari et al. / Behavioural Processes 119 (2015) 50–57 53
F in thes
Opttissms
3
lot
Fa
ig. 2. The mean escape latency to find the hidden platform during spatial learningleep deprivation(SD). Each blocks consisted of four training trials.
VX rats groups showed decreases in escape latency (Fig. 1A, B) andath length (Fig. 2A,B) required to find the hidden platform duringhe learning phase in three blocks. Animals in all groups learnedo find the location of the hidden platform in the MWM test dur-ng learning phase. As well as, physical exercise did not lead to anyignificant change in the average block values of exercised femaleubgroups. Therefore, all subgroups of intact and OVX female ani-als have similar performance in the Morris water maze during
patial learning before sleep deprivation.
.2. Spatial long-term memory
Probe test was done 24 h after acquisition phase to examineong- term spatial memory retention after sleep deprivation. Thebtained results included the mean percentage (%) for time, dis-
ance and the number of crossing in the target quadrant.Two-way ANOVA indicated a significant treatment effect [time:(5, 89) = 4.07, P = 0.003; effect size(partial eta- squared or �2) = 0.23nd distance: F(5, 89) = 3.2, P = 0.012; effect size(�2) = 0.2], but
Morris water maze task in the intact (A) and ovariectomized (B) female rats before
not for reproductive status (gonadally intact or ovariectomized)[time: F(1, 89) = 1.4, P = 0.22; effect size(�2) = 0.02 and distance:F(1, 89) = 1.02, P = 0.31; effect size(�2) = 0.016]. These results indi-cated that the sleep-deprived intact and OVX female rats spentless time and distance in the target quadrant compared to the con-trol, wide platform, exercise, sham exercise and exercise/SD groups(intact females: F(5,42) = 4.02, P = 0.004 for time and F(5,42) = 3.11,P = 0.024 for distance in the target quadrant; OVX females:F(5,42) = 8.31, P = 0.00 for time and F(5,42) = 6.6 for distance inthe target quadrant; Fig. 3A, B respectively), which demonstratedlong term memory impairment. However, this impairment wasprevented by 4 weeks of treadmill exercise in the exercise/sleepdeprived groups since they spent more time and distance in thetarget quadrant than the sleep deprived rats (Fig. 3A, B). All of theintact and OVX female rats presented a similar number of crossing
in the target quadrant in the MWM (Fig. 3A, B). There was no sig-nificant difference among the performance of the control, exercise,exercise/SD, wide platform and sham exercise groups of intact andOVX female rats in the long-term memory test (two-way ANOVA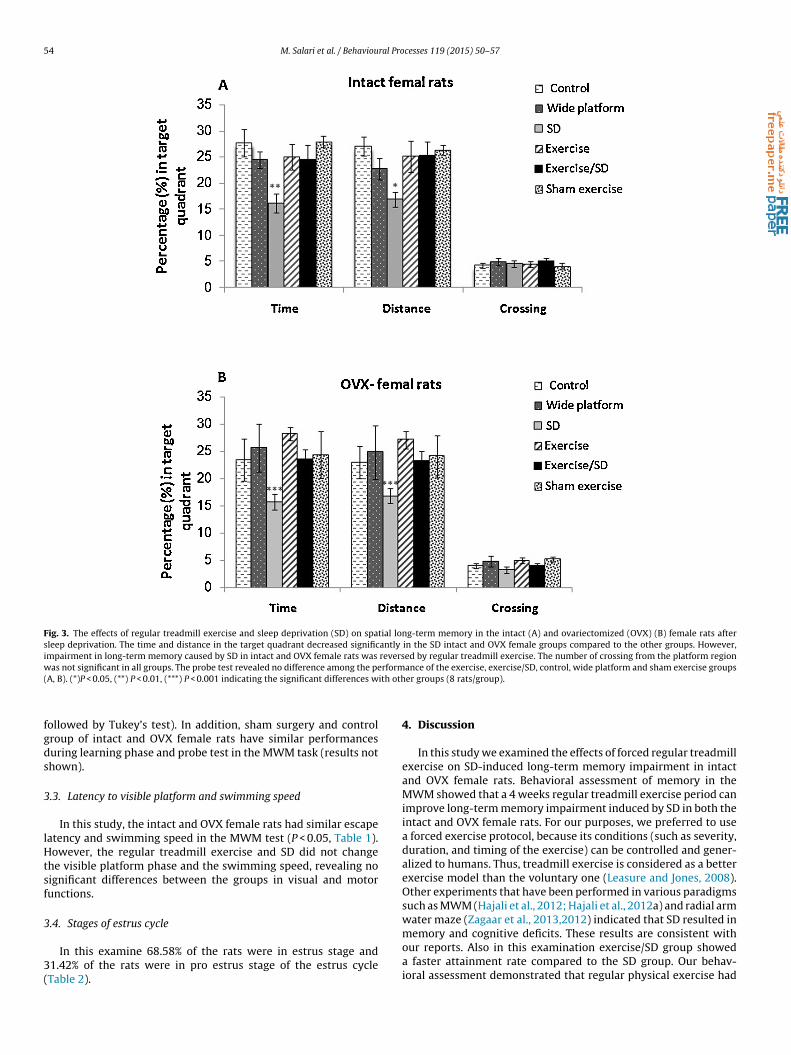
54 M. Salari et al. / Behavioural Processes 119 (2015) 50–57
Fig. 3. The effects of regular treadmill exercise and sleep deprivation (SD) on spatial long-term memory in the intact (A) and ovariectomized (OVX) (B) female rats aftersleep deprivation. The time and distance in the target quadrant decreased significantly in the SD intact and OVX female groups compared to the other groups. However,impairment in long-term memory caused by SD in intact and OVX female rats was reversed by regular treadmill exercise. The number of crossing from the platform regionw rform( ith oth
fgds
3
lHtsf
3
3(
as not significant in all groups. The probe test revealed no difference among the peA, B). (*)P < 0.05, (**) P < 0.01, (***) P < 0.001 indicating the significant differences w
ollowed by Tukey’s test). In addition, sham surgery and controlroup of intact and OVX female rats have similar performancesuring learning phase and probe test in the MWM task (results nothown).
.3. Latency to visible platform and swimming speed
In this study, the intact and OVX female rats had similar escapeatency and swimming speed in the MWM test (P < 0.05, Table 1).owever, the regular treadmill exercise and SD did not change
he visible platform phase and the swimming speed, revealing noignificant differences between the groups in visual and motorunctions.
.4. Stages of estrus cycle
In this examine 68.58% of the rats were in estrus stage and1.42% of the rats were in pro estrus stage of the estrus cycleTable 2).
ance of the exercise, exercise/SD, control, wide platform and sham exercise groupser groups (8 rats/group).
4. Discussion
In this study we examined the effects of forced regular treadmillexercise on SD-induced long-term memory impairment in intactand OVX female rats. Behavioral assessment of memory in theMWM showed that a 4 weeks regular treadmill exercise period canimprove long-term memory impairment induced by SD in both theintact and OVX female rats. For our purposes, we preferred to usea forced exercise protocol, because its conditions (such as severity,duration, and timing of the exercise) can be controlled and gener-alized to humans. Thus, treadmill exercise is considered as a betterexercise model than the voluntary one (Leasure and Jones, 2008).Other experiments that have been performed in various paradigmssuch as MWM (Hajali et al., 2012; Hajali et al., 2012a) and radial armwater maze (Zagaar et al., 2013,2012) indicated that SD resulted in
memory and cognitive deficits. These results are consistent withour reports. Also in this examination exercise/SD group showeda faster attainment rate compared to the SD group. Our behav-ioral assessment demonstrated that regular physical exercise had
M. Salari et al. / Behavioural Pro
Table 1Swimming speed and latency to escape onto the visible platform of intact andovariectomized female rats.
Groups Swimming speed (cm/s) Escape latency (sec)
IntactControl 26.99 ± 3.03 39.70 ± 4.90Wide platform 24.36 ± 1.30 37.97 ± 3.85SDa 25.89 ± 1.25 38.30 ± 4.65Exercise 24.45 ± 1.09 35.52 ± 4.65Exercise/SD 27.05 ± 0.71 33.11 ± 3.04Sham exercise 22.24 ± 2.24 38 ± 4.77
OVXb
Control 22.10 ± 1.71 33.96 ± 4.30Wide platform 26.44 ± 1.93 36.69 ± 2.13SD 21.9 ± 1.26 35.92 ± 2.97Exercise 23.82 ± 1.01 30.23 ± 2.49Exercise/SD 24.49 ± 0.43 30.04 ± 1.90Sham exercise 20.01 ± 0.75 35.16 ± 4.80
The mean of swimming speed and latency to escape onto the visible platform inMorris water maze task in the intact and ovariectomized female rats. Data aremeans ± S.E.M. (8 rats/ group).
a Sleep deprivation.b Ovariectomized groups.
Table 2Percentages of estrus cycle stages.
Groups Pro-estrus (%) Estrus (%)
Control 20 80Wide platform 50 50SDa 75 25Exercise 33.33 66.66Exercise/SD 12.5 87.5Sham exercise 25 75
as
wepssttsoieb(ctdotati
oTtpffim
Total 31.42 68.58
a Sleep deprivation.
counteracting effect on long-term memory impairments of theleep deprived female rats.
Sleep deprivation was induced using multiple platform methodhich depends on the losing of muscle tone during REM (rapid
ye movement) sleep. Albeit this technique is effective in sup-ressing about 95% of REM sleep, it can also interfere with NREMleep (Machado et al., 2004). Thus, in the current study we inducedleep deprivation in female rats by multiple platform apparatushat depend on the loss of muscle tone during REM sleep. This pro-ocol also eliminates both the social isolation and restraint factorsuch as immobilization which accompanied with the other meth-ds (Suchecki and Tufik, 2000). Additionally, previous experiment
n our laboratory showed that the differences of corticosterone lev-ls and the performances in Morris water test were not significantetween the control and sham platform (wide platform) groupsSaadati et al., 2015). In addition the results of current study indi-ated that control and sham platform have similar performance inhe MWM test. It appeared that the cognitive impairment in sleep-eprived rats on the narrow platforms was principally the resultf sleep loss and not from stresses of the column in water appara-us. In addition to, an almost equal swimming speed and latency tottain visible platform in the experimental groups demonstratedhat the observed changes were not owing to the motor and visualmpairments in the sleep deprived rats.
Several investigations have demonstrated that sleep plays a piv-tal role in learning and memory (Diekelmann and Born, 2010).here are also some human studies reporting the increased sensi-ivity of postmenopausal women to the disadvantageous effects of
oor sleep on physical (Goldman et al., 2007) and cognitive per-ormances (Blackwell et al., 2006). According to above-mentionedndings, it seems that diminution of endogenous sex steroid hor-ones is the main candidate for mediating the more sensibility of
cesses 119 (2015) 50–57 55
OVX female rats to the harmful effects of sleep loss on cognitiveand synaptic functions.
Furthermore, there are well documented studies for sex dif-ference in many aspects of brain structure, learning/memory,neuroendocrine regulation and neurotransmitter systems (Cahill,2006). It has been previously demonstrated that estrogens havesome important profits on the aging brain (Wise et al., 2001). Post-menopausal estrogen replacement therapy can prolong diminishin cognitive function (LeBlanc et al., 2001) and reduces the risk ofAlzheimer’s disease (Zandi et al., 2002). Furthermore, brain derivedneurotrophic factor (BDNF) gene expression augments in responseto estrogen (Scharfman and MacLusky, 2005). Some evidences haveoffered that the level of estrogen may play a significant role inthe regulation of sleep (Manber and Armitage, 1999). The roleof female sex steroids in sleep regulation is specially obvious inpostmenopausal women, who have low levels of the estrogen incirculate(Dzaja et al., 2005; Manber and Armitage, 1999) and aremore vulnerable to the detrimental effects of poor sleep on cogni-tive performance(Hajali et al., 2012; Saadati et al., 2015). Albeit,human study demonstrated that differences in sleep,mood, andpublic health among premenopausal and menopausal women arenot significant(Jahangard et al., 2013).
Ovarian hormone elevations are associated with increasedlearning/memory (Gasbarri et al., 2012). Furthermore, females inpro-estrus/estrus demonstrated better performance compared tofemales in metestrus and di-estrus (Van Goethem et al., 2012).Commonly, female rodents and other mammals living in groupswere synchronized in the other experiment (McClintock, 1984), wealso synchronized the estrus cycles in the intact female rats in thepresent study. Researchers during another investigation put femalerats in pro-estrus and estrus formed one group (pro/es; charac-terized by higher levels of estradiol and progesterone), and theother group consisted of female rats in met-estrus and di-estrus(met/di; characterized by lower levels of estradiol and proges-terone) (Van Goethem et al., 2012); similar subdivisions have beendescribed in other studies investigating the role of estrus cycle inbehavior (Sell et al., 2000; Warren and Juraska, 1997). Estrogentreatment is reported to exert effects on memory, particularly thosetypes of memory in which the hippocampus plays a role and canenhance LTP (Foy et al., 2000) .In addition, aging female rats thathave low estrogen plasma levels in the “estropause” are reportedto perform significantly more poorly in a Morris water maze thanfemale rats with high estrogen levels (Hajali et al., 2012); thus weto synchronize intact rats. In total; estrogens exert complex andtime-dependent effects on cognitive function including learningand memory processes, in animals and humans (Gasbarri et al.,2012). The basal forebrain cholinergic system and hippocampusare two brain systems that appear to be involved in both attentionand memory effects of estrogens (Gibbs and Aggarwal, 1998).
Our results indicated that treadmill exercise has beneficialeffects on long term memory in sleep deprived female rats. Wealso showed that regular exercise can improve memory deficitinduced by SD completely, because memory performance in theSD- exercise groups was similar to the control groups. Other stud-ies offered that treadmill exercise had neuroprotective effects ondetrimental behavioral, synaptic and molecular changes inducedby sleep deprivation (Fernandes et al., 2013; Zagaar et al., 2013,2012). These studies demonstrated that the benefits of exercise atthe cellular level likely due to its ability to increase the production ofBDNF and other signaling molecules in sleep-deprived male (Zagaaret al., 2012) and female (Saadati et al., 2014a) rats. HippocampalBDNF can mediate the effect of exercise on cognitive functions
(Vaynman et al., 2004). Therefore physical exercise could preventthe decreasing effect of SD in the OVX female rats.Of course we didnot focus on the role of signaling molecule in the present study. Itseems that further experiments are needed to examine the effect of

5 ral Pro
pn
ftfn
tbBhefmt
tt
4
ets
atepond
D
A
t
R
A
A
A
B
B
CC
C
C
D
6 M. Salari et al. / Behaviou
hysical exercise on the expression of BDNF mRNA and other sig-aling molecules in female rats following 24 h SD.
In addition, the exercise and control groups have similar per-ormance in the behavioral test. These findings assert the concepthat maybe forced exercise regimens exert positive effects in theace of an insult or impairment such as sleep loss, ischemia, oreurodegeneration.
However some experiments have demonstrated the consequen-ial promotion of cognitive performance, learning and memoryy physical activities (Ang and Gomez-Pinilla, 2007; Cotman anderchtold, 2002). Other studies, in agreement with our findings,ave revealed lack of improvement (Titterness et al., 2011; Zagaart al., 2012) .These conflicting findings may be because of the dif-erences in the length, type and intensity of the physical exercise
oreover stages of estrus cycle in female animals used as well ashe differences in age and strains of the experimental animals.
Therefore, alteration in methodologies, including the length andype of physical exercise and experimental protocol might causehe different results.
.1. Limitation
Surgical ovariectomy was not confirmed by measure of plasmastradiol concentrations or by checking of estrus cycle of OVX rats inhe current experiment. This issue was the limitation of the presenttudy.
In conclusion the results of the current study confirmed the neg-tive effect of SD on cognitive functions and regular exercise seemso have protective effects against SD-induced memory deficit, how-ver more investigations need to be done. The advantage of theresent study is to generalize to women especially menopausalne. Although these findings failed to indicate any differential cog-itive susceptibility of the two reproductive conditions to sleepeprivation.
eclaration of interest
The authors report no conflicts of interest.
cknowledgement
We would like to acknowledge the Neuroscience Research Cen-er of Kerman, Iran for financial support.
eferences
leisa, A.M., Alzoubi, K.H., Alkadhi, K.A., 2011. Post-learning REM sleep deprivationimpairs long-term memory: reversal by acute nicotine treatment. Neurosci.Lett. 499, 28–31.
lhola, P., Tallus, M., Kylmälä, M., Portin, R., Polo-Kantola, P., 2005. Sleepdeprivation, cognitive performance, and hormone therapy in postmenopausalwomen. Menopause 12, 149–155.
ng, E., Gomez-Pinilla, F., 2007. Potential therapeutic effects of exercise to thebrain. Curr. Med. Chem 14, 2564–2571.
erchtold, N.C., Chinn, G., Chou, M., Kesslak, J.P., Cotman, C.W., 2005. Exerciseprimes a molecular memory for brain-derived neurotrophic factor proteininduction in the rat hippocampus. Neuroscience 133, 853–861.
lackwell, T., Yaffe, K., Ancoli-Israel, S., Schneider, J.L., Cauley, J.A., Hillier, T.A., Fink,H.A., Stone, K.L., 2006. Poor sleep is associated with impaired cognitivefunction in older women: the study of osteoporotic fractures. J. Gerontol. ABiol. Sci. Med. Sci. 61, 405–410.
ahill, L., 2006. Why sex matters for neuroscience. Nat. Rev. Neurosci. 7, 477–484.ai, J.D., Shuman, T., Gorman, M.R., Sage, J.R., Anagnostaras, S.G., 2009. Sleep
selectively enhances hippocampus-dependent memory in mice. Behav.Neurosci. 123, 713.
olvin, G.B., Whitmoyer, D.I., Lisk, R.D., Walter, D.O., Sawyer, C.H., 1968. Changes insleep-wakefulness in female rats during circadian and estrous cycles. Brain
Res. 7, 173–181.otman, C.W., Berchtold, N.C., 2002. Exercise: a behavioral intervention to enhancebrain health and plasticity. Trends. Neurosci. 25, 295–301.
atta, S., et al., 2010. Sleep: learning and memory. In: Koob, G.F. (Ed.), Encyclopediaof Behavioral Neuroscience. Academic Press, Oxford, pp. 218–226.
cesses 119 (2015) 50–57
Davis, C.J., Harding, J.W., Wright, J.W., 2003. REM sleep deprivation-induced deficitsin the latency-to-peak induction and maintenance of long-term potentiationwithin the CA1 region of the hippocampus. Brain Res. 973, 293–297.
Diekelmann, S., Born, J., 2010. The memory function of sleep. Nat. Rev. Neurosci.11, 114–126.
Dzaja, A., Arber, S., Hislop, J., Kerkhofs, M., Kopp, C., Pollmächer, T., Polo-Kantola, P.,Skene, D.J., Stenuit, P., Tobler, I., 2005. Women’s sleep in health and disease. J.Psychiatr. Res. 39, 55–76.
Fernandes, J., Baliego, L.G.Z., Peixinho-Pena, L.F., de Almeida, A.A., Venancio, D.P.,Scorza, F.A., de Mello, M.T., Arida, R.M., 2013. Aerobic exercise attenuatesinhibitory avoidance memory deficit induced by paradoxical sleep deprivationin rats. Brain Res. 1529, 66–73.
Foy, M.R., Henderson, V.W., Berger, T.W., Thompson, R.F., 2000. Estrogen andneural plasticity. Curr. Dir. Psychol. Sci. 9, 148–152.
Gais, S., Lucas, B., Born, J., 2006. Sleep after learning aids memory recall. Learn.Mem. 13, 259–262.
Gasbarri, A., Tavares, M.C.H., Rodrigues, R.C., Tomaz, C., Pompili, A., 2012. Estrogen,cognitive functions and emotion: an overview on humans, non-humanprimates and rodents in reproductive years. Rev. Neurosci. 23, 587–606.
Gibbs, R.B., Aggarwal, P., 1998. Estrogen and basal forebrain cholinergic neurons:implications for brain aging and Alzheimer’s disease-related cognitive decline.Horm. Behav. 34, 98–111.
Goldman, S.E., Stone, K.L., Ancoli-Israel, S., Blackwell, T., Ewing, S.K., Boudreau, R.,Cauley, J.A., Hall, M., Matthews, K.A., Newman, A.B., 2007. Poor sleep isassociated with poorer physical performance and greater functionallimitations in older women. Sleep 30, 1317–1324.
Green, S.P., Simpkins, J.W., 2000. Neuroprotective effects of estrogens: potentialmechanisms of action. Int. J. Dev. Neurosci. 18, 347–358.
Hajali, V., Sheibani, V., Esmaeili-Mahani, S., Shabani, M., 2012. Female rats aremore susceptible to the deleterious effects of paradoxical sleep deprivation oncognitive performance. Behav. Brain Res. 228, 311–318.
Hajali, V., Sheibani, V., Esmaeili-Mahani, S., Shabani, M., Aliabadi, H.P., Saadati, H.,Esmaeilpour, K., 2015a. Ovariectomy does not exacerbate the negative effectsof sleep deprivation on synaptic plasticity in rats. Physiol. Behav. 144, 73–81.
Hajali, V., Sheibani, V., Ghazvini, H., Ghadiri, T., Valizadeh, T., Saadati, H., Shabani,M., 2015b. Effect of castration on the susceptibility of male rats to the sleepdeprivation-induced impairment of behavioral and synaptic plasticity.Neurobiol. Learn. Mem. 123, 140–148.
Hillman, C.H., Erickson, K.I., Kramer, A.F., 2008. Be smart, exercise your heart:exercise effects on brain and cognition. Nat. Rev. Neurosci. 9, 58–65.
Hogervorst, E., Williams, J., Budge, M., Riedel, W., Jolles, J., 2000. The nature of theeffect of female gonadal hormone replacement therapy on cognitive functionin post-menopausal women: a meta-analysis. Neuroscience 101, 485–512.
Jahangard, L., Haghighi, M., Bajoghli, H., Holsboer-Trachsler, E., Brand, S., 2013.Among a sample of Iranian pre-menopausal and menopausal womendifferences in mood, sleep and health quality are small. Int. J. Psychiatry Clin.Pract., 1–16.
Landrigan, C.P., Rothschild, J.M., Cronin, J.W., Kaushal, R., Burdick, E., Katz, J.T., Lilly,C.M., Stone, P.H., Lockley, S.W., Bates, D.W., Czeisler, C.A., 2004. Effect ofreducing interns’ work hours on serious medical errors in intensive care units.N. Engl. J. Med. 351, 1838–1848.
Leasure, J., Jones, M., 2008. Forced and voluntary exercise differentially affect brainand behavior. Neuroscience 156, 456–465.
LeBlanc, E.S., Janowsky, J., Chan, B.K., Nelson, H.D., 2001. Hormone replacementtherapy and cognition: systematic review and meta-analysis. JAMA 285,1489–1499.
Leconte, P., Bloch, V., 1970. Deficiency in retention of conditioning afterdeprivation of paradoxical sleep in rats. C. R. Acad. Sci. Hebd. Seances. Acad.Sci. D. 271, 226–229.
Machado, R.B., Hipolide, D.C., Benedito-Silva, A.A., Tufik, S., 2004. Sleep deprivationinduced by the modified multiple platform technique: quantification of sleeploss and recovery. Brain Res. 1004, 45–51.
Manber, R., Armitage, R., 1999. Sex steroids and sleep: a review. Sleep 22, 540–555.McClintock, M.K., 1984. Estrous synchrony: modulation of ovarian cycle length by
female pheromones. Physiol. Behav. 32, 701–705.Miquel, J., Ramirez-Bosca, A., Ramirez-Bosca, J.V., Alperi, J.D., 2006. Menopause: a
review on the role of oxygen stress and favorable effects of dietaryantioxidants. Arch. Gerontol. Geriatr. 42, 289–306.
Philip, P., Akerstedt, T., 2006. Transport and industrial safety, how are they affectedby sleepiness and sleep restriction. Sleep Med. Rev. 10, 347–356.
Saadati, H., Sheibani, V., Esmaeili-Mahani, S., Darvishzadeh-Mahani, F., Mazhari, S.,2014a. Prior regular exercise reverses the decreased effects of sleepdeprivation on brain-derived neurotrophic factor levels in the hippocampus ofovariectomized female rats. Regul. Pept. 194–195, 11–15.
Saadati, H., Sheibani, V., Esmaeili-Mahani, S., Hajali, V., Mazhari, S., 2014b. Priorregular exercise prevents synaptic plasticity impairment in sleep deprivedfemale rats. Brain Res. Bull. 108, 100–105.
Saadati, H., Esmaeili-Mahani, S., Esmaeilpour, K., Nazeri, M., Mazhari, S., Sheibani,V., 2015. Exercise improves learning and memory impairments in sleepdeprived female rats. Physiol. Behav. 138, 285–291.
Scharfman, H.E., MacLusky, N.J., 2005. Similarities between actions of estrogen and
BDNF in the hippocampus: coincidence or clue. Trends Neurosci. 28,79–85.
Sell, S.L., Scalzitti, J.M., Thomas, M.L., Cunningham, K.A., 2000. Influence of ovarianhormones and estrous cycle on the behavioral response to cocaine in femalerats. J. Pharmacol. Exp. Ther. 293, 879–886.

ral Pro
S
S
T
V
V
V
V
Neurobiol. Dis. 45, 1153–1162.
M. Salari et al. / Behaviou
hangold, M.M., 1990. Exercise in the menopausal woman. Obstet. Gynecol. 75,53S–58S.
uchecki, D., Tufik, S., 2000. Social stability attenuates the stress in the modifiedmultiple platform method for paradoxical sleep deprivation in the rat. Physiol.Behav. 68, 309–316.
itterness, A., Wiebe, E., Kwasnica, A., Keyes, G., Christie, B., 2011. Voluntaryexercise does not enhance long-term potentiation in the adolescent femaledentate gyrus. Neuroscience 183, 25–31.
an der Borght, K., Kóbor-Nyakas, D.É., Klauke, K., Eggen, B.J., Nyakas, C., Van derZee, E.A., Meerlo, P., 2009. Physical exercise leads to rapid adaptations inhippocampal vasculature: temporal dynamics and relationship to cellproliferation and neurogenesis. Hippocampus 19, 928–936.
an Goethem, N.P., Rutten, K., van der Staay, F.J., Jans, L.A., Akkerman, S.,Steinbusch, H.W., Blokland, A., van’t Klooster, J., Prickaerts, J., 2012. Objectrecognition testing: rodent species, strains, housing conditions, and estrouscycle. Behav. Brain Res. 232, 323–334.
an Praag, H., 2009. Exercise and the brain: something to chew on. TrendsNeurosci. 32, 283–290.
aynman, S., Ying, Z., Gomez-Pinilla, F., 2004. Hippocampal BDNF mediates theefficacy of exercise on synaptic plasticity and cognition. Eur. J. Neurosci. 20,2580–2590.
cesses 119 (2015) 50–57 57
Walker, M.P., Stickgold, R., 2004. Sleep-dependent learning and memoryconsolidation. Neuron 44, 121–133.
Warren, S.G., Juraska, J.M., 1997. Spatial and nonspatial learning across the ratestrous cycle. Behav. Neurosci. 111, 259.
Wise, P.M., Dubal, D.B., Wilson, M.E., Rau, S.W., Böttner, M., Rosewell, K.L., 2001.Estradiol is a protective factor in the adult and aging brain: understanding ofmechanisms derived from in vivo and in vitro studies. Brain. Res. Rev. 37,313–319.
Zagaar, M., Dao, A., Alhaider, I., Alkadhi, K., 2013. Regular treadmill exerciseprevents sleep deprivation-induced disruption of synaptic plasticity andassociated signaling cascade in the dentate gyrus. Mol. Cell. Neurosci. 56,375–383.
Zagaar, M., Alhaider, I., Dao, A., Levine, A., Alkarawi, A., Alzubaidy, M., Alkadhi, K.,2012. The beneficial effects of regular exercise on cognition in REM sleepdeprivation: behavioral, electrophysiological and molecular evidence.
Zandi, P.P., Carlson, M.C., Plassman, B.L., Welsh-Bohmer, K.A., Mayer, L.S., Steffens,D.C., Breitner, J.C., Investigators, C.C.M.S., 2002. Hormone replacement therapyand incidence of Alzheimer disease in older women: the Cache County Study.Jama 288, 2123–2129.
Copyright © 2022 FDOKUMEN




















