
Temporal stability of niche use exposes sympatric Arctic charr to alternative selection pressures
16
ORIGINAL PAPER Temporal stability of niche use exposes sympatric Arctic charr to alternative selection pressures Rune Knudsen • Anna Siwertsson • Colin E. Adams • Monica Gardun ˜ o-Paz • Jason Newton • Per-Arne Amundsen Received: 8 March 2010 / Accepted: 10 November 2010 / Published online: 24 November 2010 Ó Springer Science+Business Media B.V. 2010 Abstract There is now strong evidence that foraging niche specialisation plays a critical role in the very early stages of resource driven speciation. Here we test critical elements of models defining this process using a known polymorphic population of Arctic charr from subarctic Norway. We test the long-term stability of niche specialisation amongst foraging predators and discuss the possibility that contrasting foraging specialists are exposed to differing selection regimes. Inter-individual foraging niche stability was measured by combining two time-integrated ecological tracers of the foraging niche (each individual’s d 13 C and d 15 N stable isotope (SI) signatures and their food borne parasite fauna) with a short-term measure of foraging niche use (stomach contents composition). Three dietary subgroups of predators were identified, including zooplankton, gammarid and benthivore specialists foragers. Zooplanktivorous specialists had muscle low in d 13 C, a high abun- dance of parasites transmitted from pelagic copepods, a smaller head, longer snout and a more slender body-form than gammaridivorous specialist individuals which had muscle more enriched in d 13 C and high abundance of parasites transmitted from benthic Gammarus. Benthivorous individuals were intermediate between the other two foraging groups according to muscle SI-signals (d 13 C) and loadings of parasites transmitted from Electronic supplementary material The online version of this article (doi:10.1007/s10682-010-9451-9) contains supplementary material, which is available to authorized users. R. Knudsen (&) A. Siwertsson C. E. Adams P.-A. Amundsen Department of Arctic and Marine Biology, University of Tromsø, 9037 Tromsø, Norway e-mail: [email protected] C. E. Adams Scottish Centre for Ecology and the Natural Environment, University of Glasgow, Rowardennan, Glasgow G63 0AW, Scotland, UK M. Gardun ˜o-Paz Facultad de Ciencias, Universidad Auto ´noma del Estado de Me ´xico, Instituto Literario No. 100, Toluca, Estado de Me ´xico C.P. 50150, Mexico J. Newton NERC Life Sciences Mass Spectrometry Facility, SUERC, East Kilbride, Glasgow G75 0QF, Scotland, UK 123 Evol Ecol (2011) 25:589–604 DOI 10.1007/s10682-010-9451-9
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Temporal stability of niche use exposes sympatric Arctic charr to alternative selection pressures

ORI GIN AL PA PER
Temporal stability of niche use exposes sympatric Arcticcharr to alternative selection pressures
Rune Knudsen • Anna Siwertsson • Colin E. Adams • Monica Garduno-Paz •
Jason Newton • Per-Arne Amundsen
Received: 8 March 2010 / Accepted: 10 November 2010 / Published online: 24 November 2010� Springer Science+Business Media B.V. 2010
Abstract There is now strong evidence that foraging niche specialisation plays a critical
role in the very early stages of resource driven speciation. Here we test critical elements of
models defining this process using a known polymorphic population of Arctic charr from
subarctic Norway. We test the long-term stability of niche specialisation amongst foraging
predators and discuss the possibility that contrasting foraging specialists are exposed to
differing selection regimes. Inter-individual foraging niche stability was measured by
combining two time-integrated ecological tracers of the foraging niche (each individual’s
d13C and d15N stable isotope (SI) signatures and their food borne parasite fauna) with a
short-term measure of foraging niche use (stomach contents composition). Three dietary
subgroups of predators were identified, including zooplankton, gammarid and benthivore
specialists foragers. Zooplanktivorous specialists had muscle low in d 13C, a high abun-
dance of parasites transmitted from pelagic copepods, a smaller head, longer snout and a
more slender body-form than gammaridivorous specialist individuals which had muscle
more enriched in d 13C and high abundance of parasites transmitted from benthic
Gammarus. Benthivorous individuals were intermediate between the other two foraging
groups according to muscle SI-signals (d13C) and loadings of parasites transmitted from
Electronic supplementary material The online version of this article (doi:10.1007/s10682-010-9451-9)contains supplementary material, which is available to authorized users.
R. Knudsen (&) � A. Siwertsson � C. E. Adams � P.-A. AmundsenDepartment of Arctic and Marine Biology, University of Tromsø, 9037 Tromsø, Norwaye-mail: [email protected]
C. E. AdamsScottish Centre for Ecology and the Natural Environment, University of Glasgow, Rowardennan,Glasgow G63 0AW, Scotland, UK
M. Garduno-PazFacultad de Ciencias, Universidad Autonoma del Estado de Mexico, Instituto Literario No. 100,Toluca, Estado de Mexico C.P. 50150, Mexico
J. NewtonNERC Life Sciences Mass Spectrometry Facility, SUERC, East Kilbride, GlasgowG75 0QF, Scotland, UK
123
Evol Ecol (2011) 25:589–604DOI 10.1007/s10682-010-9451-9

both copepods and Gammarus. The close relationship between subgroups identified by
stomach contents, time-integrated tracers of niche use (SI and parasites) and functional
trophic morphology (niche adaptations) demonstrate a long-term temporally stable niche
use of each individual predator. Differential habitat use and contrasting parasite commu-
nities and loadings, show differential exposure to different suites of selection pressures for
different foraging specialists. Results also show that individual specialisation in trophic
behaviour and thus exposure to different suites of selection pressures are stable over time,
and thus provide a platform for disruptive selection to operate within this sympatric
system.
Keywords Stable isotopes � Food web transmitted parasites � Functional morphology �Temporal niche stability � Salvelinus alpinus
Introduction
There is now compelling theoretical and empirical evidence that ecological processes can
play a significant role in the early stages of speciation (Schluter 1996, 2001). Models
describing this process typically invoke a behavioural specialisation on alternative
resources by individuals in a population; most frequently thought of in this context are
foraging resources (West-Eberhard 1989, 2005; Dieckmann and Doebeli 1999; Skulason
et al. 1999). For species that have the capacity for phenotypic plasticity, this may result in
the expression of alternative phenotypes through exposure of foraging specialists to
alternative environments (Adams et al. 2003; Snowberg and Bolnick 2008; Garduno-Paz
and Adams 2010). If this stage is reached, individuals expressing alternative phenotypes
and adopting different ecological specialisms may then be subject to disruptive selection
related directly to their use of different resources, or as a result of differential expression of
alternative morphological or behavioural phenotypes (West-Eberhard 1989, 2005; Skula-
son et al. 1999; Rueffler et al. 2007; Duckworth 2009). Thus both phenotypic plasticity and
genetic components play an important role in the process of divergence (Schluter and
Conte 2009; Pfennig et al. 2010) also in young species flocks (Losos 2010). Although the
process is far from fully understood, empirical evidence is now mounting, particularly for
fish inhabiting post-glacial lakes (but also for other animal groups, see e.g. Herrel et al.
2008; Funk 2010) that the above sequence of events has occurred, and is still occurring, in
a number of species (Bolnick et al. 2003).
Amongst freshwater fishes, lacustrine Arctic charr (Salvelinus alpinus (L.)), show clear
evidence of clear differential resource use (usually foraging resources) frequently associ-
ated with the expression of alternative phenotypes (resource polymorphisms sensuSkulason and Smith 1995) with a functional role (Adams and Huntingford 2002a;
Klemetsen et al. 2002, 2006; Knudsen et al. 2006). Alternative phenotypes are frequently
found in sympatry (Klemetsen 2010) and often take the form of a specialist zooplankton
feeders foraging in the limnetic zone and a specialist macro-invertebrate feeders, foraging
in the benthic zone of lakes (reviewed by Robinson and Parsons 2002; Hendry 2009).
However in some lakes, additional foraging specialisms have been identified in this species
(Snorrason et al. 1994; Adams et al. 1998; Knudsen et al. 2006). Similar parallel cases of
resource specialisation have also been recorded in other lacustrine fish species in post-
glacial systems (e.g. Kristjansson et al. 2002; Kahilainen and Østbye 2006). Arctic charr
have been shown to express, at least some, of the phenotypic variation described from
alternative phenotypes from the wild through ontogenetic plasticity (Adams and
590 Evol Ecol (2011) 25:589–604
123

Huntingford 2002a, 2004). For some populations, expressed alternative phenotypes also
have an inherited genetic component (Sandlund et al. 1992; Adams and Huntingford
2002b, 2004; Klemetsen et al. 2002, 2006). Thus, it is likely that across the species,
populations are exhibiting multiple stages on the route to evolutionary divergence
(Skulason et al. 1999; Losos 2010).
There are two critical pre-requisites for evolutionary divergence resulting from resource
specialisation to operate. Firstly, the alternative resource specialisms adopted by individ-
uals need to be stable over time. For the ontogenetic effect of phenotypic plasticity to
modify the expressed phenotype, individuals should retain their niche specialisms over a
period close to the generation time. For disruptive selection to result in evolutionary
divergence, resource specialisms need to be expressed in a population over multiple
generations (see Woo et al. 2008; Knudsen et al. 2010). Secondly, for disruptive selection
to operate, organisms occupying separate ecological niches need to be exposed to different
selection pressures, disruptive selection could result in alternative expressed phenotypes of
resource specialists (Abrams 2006; Doebeli et al. 2007). Here we test if these two pre-
requisites for resource-specialism mediated evolution exist, in a model system where
Arctic charr display discrete alternative phenotypes which show foraging specialisms and
which are presumed to be in the process for incipient speciation (Knudsen et al. 2007).
The Arctic charr in Fjellfrøsvatn, Northern Norway show discrete trophic specialisation
related to macro-habitat segregation (Amundsen et al. 2008; Knudsen et al. 2007, 2010).
This trophic specialisation exposes individuals with different specialisms to different
burdens of specific trophic transmitted parasite species (Knudsen et al. 2004), which could
act as strong parasite-mediated selection forces on the host populations (Maan et al. 2008;
Matthews et al. 2010). Here we, use an individual based approach to test three specific
hypotheses in this model system. First, we hypothesise that resource-use specialisation
persists over the life-time of the individual [i.e. that they do not fluctuate over short periods
(such as seasonally)]. To test this we examine short-term (stomach contents) and long term
(stable isotopes analysis and parasite fauna) integrators of diet. Secondly, we hypothesise
that the expression of resource specialisation in this model system remains temporally
stable over periods consistent with evolutionary divergence. This we test by comparing
resource use data in this study with historical data. Thirdly, we hypothesise that resource
specialisation are accompanied by functional morphological adaptations. We discuss our
result in a context that resource specialists are subject to differential selection pressures
consistent with divergent selection.
Materials and methods
Fjellfrøsvatn (69�050N, 19�200E, 125 m a.s.l) is a dimictic, oligotrophic lake situated in
subarctic Norway. It has a surface area of 6.5 km2, a maximum depth of 88 m and well
developed pelagic and littoral habitats. The littoral zone (\10 m) constitutes slightly less
than one-third of the lake surface area. The only fish species present are Arctic charr and
brown trout (Salmo trutta L.).
Mature Arctic charr develop secondary sexual traits (Skarstein and Folstad 1996). To
limit this source of variation in our study of functional morphological traits, we restricted
the analyses to immature individuals only. Arctic charr were sampled from two habitats in
October 1997, the littoral (using benthic gill-nets, set in 0–10 m water depth) and pelagic
(using offshore, floating, gill-nets, set above[30 m water depth). All nets used were multi-
mesh gill-nets (10, 12.5, 15, 18.5, 22, 26, 35 and 45 mm, knot to knot). In total 78 charr of
Evol Ecol (2011) 25:589–604 591
123

mean fork length 21.5 cm (±1.9 SD, range: 18.9–25.1 cm) and mean age 5.2 years (±0.7
SD, range: 4–7 years) were collected and used as a basis for all analyses described below.
Furthermore, historical data are included to provide an insight into long-term stability of
specialised trophic behaviour (see online Supplementary Fig. 2). The temporal stability of
resource use (diet and parasite infection; see details below) of the present Arctic charr
population was explored using a long-term data set (from 1992 to 2009; except the years
1993–1995, 2008; see online Supplementary Table 1 for the number of fish at each
sampling period) that were collected as described.
Prey items in the stomachs of individual fish were identified and their relative contri-
butions to the total stomach fullness estimated according to the technique of Amundsen
et al. (1996). Food items were categorised into two main groups (and five subgroups; A–E)
as either limnetic prey: A) zooplankton, or several different benthic prey: B) molluscs, C)
insect larvae, D) Gammarus lacustris L., and E) semi-pelagic chydorids. Individual charr
were categorised as either zooplanktivores or benthivores based on the dominance ([50%
abundance) of limnetic or benthic prey respectively in the stomach contents. Additionally,
the benthivorous fish were divided into benthic generalists and Gammarus-specialists, the
latter group defined by a dominance of G. lacustris (C50% abundance) in stomachs. For
historical data, detailed stomach analyses were performed only in 1992 (n = 237), 1997
(n = 78) and 2006 (n = 37). In the other years only course-grained diet data (i.e. abun-
dance of zooplankton, G. lacustris, and benthic prey) are available (see Supplementary
Table 1 for details).
Dietary specialisation was quantified by comparing each individual’s diet niche with the
population diet niche, and the proportional similarity was calculated according to Bolnick
et al. (2002):
PSi ¼X
j
min pij; qj
� �
where pij is the proportion (% abundance) of prey type j (see the diet categories above) in
the diet of individual i, and qj is the proportion of prey type j in the whole population (%
abundance is estimated according to Amundsen et al. (1996)). Values of PSi varies between
0 (no overlap between individual and population) and 1 (total overlap). The lower the PSi-
value, the more specialised the individuals are compared to the overall population (see
Bolnick et al. 2003; Sargeant 2007). Differences in mean PSi-values between dietary
groups were tested using t-tests (SYSTAT, v. 13, 2009, SYSTAT Software, Inc.).
For stable isotope analysis, a sample of muscle tissue was taken from below the dorsal
fin from each individual fish (e.g. Pinnegar and Polunin 1999). Stable isotope ratios of a
consumer reflect the isotopic signatures of the prey consumed, during the time period that
the tissue was synthesised. Muscle tissue is commonly used to determine long-term diet, in
temperate fish, typically reflecting a summer-period of somatic growth (Perga and Ger-
deaux 2005). Zooplankton and four different groups of benthic prey (molluscs, G. lacus-tris, insect larvae and chydorids) were collected and analysed for stable isotopes. To ensure
sufficient material for analysis, each analysis was conducted on several whole animals
combined. Samples were oven dried at 60�C for 24 h and pulverised using a mortar and
pestle, before subsamples were weighed into tin capsules. Isotopic analyses of carbon and
nitrogen were carried out at the NERC Life Sciences Mass Spectrometry Facility, East
Kilbride, Scotland, by continuous flow isotope ratio mass spectrometry (CF-IRMS), using
a Costech ECS 4010 elemental analyser interfaced with a ThermoFisher Scientific Delta
XPPlus IRMS. Stable isotope ratios are expressed conventionally as parts per thousand
592 Evol Ecol (2011) 25:589–604
123

(%) delta values (d13C and d15N), in relation to the international standards for carbon
(PeeDee Belemnite) and nitrogen (atmospheric nitrogen). Precision, obtained from repli-
cate analyses of internal gelatin and alanine standards, was \0.1% (carbon) and \0.2 %(nitrogen). To get information about the relative contribution of different food sources
based on stable isotope composition we used a recently developed Bayesian mixing model
(SIAR, Parnell et al. 2010), which allows for incorporation of uncertainties in both prey
isotopic estimates and trophic fractionation. We used trophic fractionation factor values
from the literature, mean (±SD) 0.4 (±1.3) % for d13C and 3.4 (±1.0) % for d15N (Post
2002). Correlations between posterior distributions of proportions of different prey showed
that the model had problems differentiating between zooplankton and chydorids (corre-
lation coefficient: -0.78) and between G. lacustris and molluscs (-0.59). Thus, com-
parisons between the three different diet groups of charr were performed for the three prey
categories: (1) zooplankton/chydorids, (2) insect larvae and (3) G. lacustris/molluscs. For
each prey category, probability values for differences between any two diet groups were
obtained by the proportion of the total number of simulations (30,000) where one group
had a higher contribution of the prey than the other. The Bayesian mixing model was fitted
using R software (R Development Core Team 2009) and the package SIAR (version 4.0, by
Andrew Parnell and Andrew Jackson).
The parasite community of each individual fish was described by a subset of endo-
parasite species. Adult trematodes (Crepidostomum spp.) and cestodes (Cyathocephalustruncatus (Pallas), Eubothrium salvelini (Schrank), Proteocephalus sp.) from the intes-
tines, and larval cestodes (Diphyllobothrium dendriticum (Nitzsch) and D. ditremum(Creplin)) were enumerated. The nematode Cystidicola farionis (Fisher) occurring in the
swim-bladder, was enumerated as L3-larvae (recruitment stage), preadults (L4-larvae) or
adults. All of these parasites are transmitted to the fish by species-specific intermediate
hosts such as pelagic copepods (E. salvelini, Proteocephalus sp., D. dendriticum, D. di-tremum), insect larvae (Crepidostomum spp.) and Gammarus lacustris (C. truncatus, C.farionis). The stability of parasite infection through time was explored by using long-term
data from 1992 to 2009 (see details above, n = 552) of the Gammarus-transmitted para-
sites (C. farionis and C. truncatus). A similar dataset for the copepod transmitted parasites
Diphyllobothrium spp. was only available for 1992 (n = 237), 1997 (n = 78) and 2006
(n = 37) when standardised sampling was performed (see Knudsen et al. 2004). The terms
prevalence and abundance of parasites are used according to Bush et al. (1997). Differ-
ences in parasite abundances between dietary groups was tested using either Mann–
Whitney U-tests or Kruskal–Wallis tests (SYSTAT, v. 13, 2009, SYSTAT Software, Inc.).
A Constrained Correspondence Analyses (CCA) followed by a Monte Carlo permuta-
tion test, was used to model relationships between stable isotope values, food borne par-
asites and resource use (stomach content) (Legendre and Legendre 1998). This analysis
was performed using the R software (R Development Core Team 2009) and the package
Vegan (version 1.8-8, by Jari Oksanen).
Landmark-based geometric morphometrics techniques were used to describe shape
variation of charr (Rohlf and Marcus 1993; Adams et al. 2004). The left side of each fish
was photographed under identical light conditions using Kodak 64 ASA EPR Ectachrome
film. The photographs were later digitised using the Kodak Photo CD-system. Shape-
analyses were performed separately for head and body, and in total 21 landmarks (see
online supplementary Fig. 1 for location of landmarks) were digitised using TPS Dig2
(Rohlf 2006). Landmark configurations were aligned, translated and rotated in a Gen-
eralized Procrustes Analysis (GPA) superimposition in order to remove position, orien-
tation and scale effects (Rohlf and Slice 1990). The GPA results in a data matrix with new
Evol Ecol (2011) 25:589–604 593
123
shape variables, Procrustes coordinates, which were used in statistical analyses. To test for
allometric effects of shape, shape variables were regressed on centroid size (body
P = 0.48; head P = 0.17). Centroid size in geometric morphometrics is defined as the
square root of the summed, square distance of all landmarks about their centroid (Zelditch
et al. 2004). There was no allometic effects over the size range of fish used in this study.
Shape differences between different diet groups were explored using Canonical Variates
Analyses (CVA) on Procrustes coordinates of body and head separately. Statistical
inference of group differences was made by pairwise permutation tests of the Mahalanobis
distances in shape between groups. MorphoJ 1.01a (Klingenberg 2008) was used for the
GPA and statistical analyses of shape.
Results
Dietary specialisation
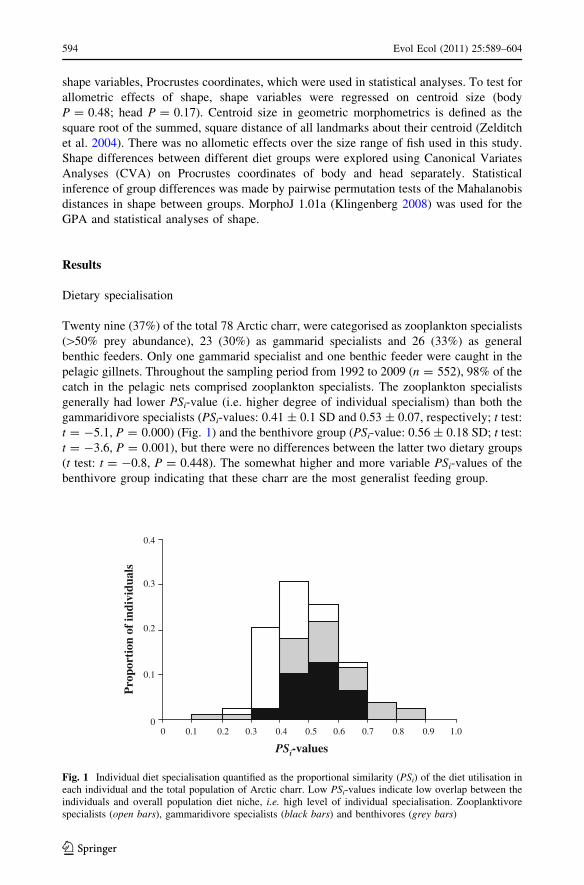
Twenty nine (37%) of the total 78 Arctic charr, were categorised as zooplankton specialists
([50% prey abundance), 23 (30%) as gammarid specialists and 26 (33%) as general
benthic feeders. Only one gammarid specialist and one benthic feeder were caught in the
pelagic gillnets. Throughout the sampling period from 1992 to 2009 (n = 552), 98% of the
catch in the pelagic nets comprised zooplankton specialists. The zooplankton specialists
generally had lower PSi-value (i.e. higher degree of individual specialism) than both the
gammaridivore specialists (PSi-values: 0.41 ± 0.1 SD and 0.53 ± 0.07, respectively; t test:
t = -5.1, P = 0.000) (Fig. 1) and the benthivore group (PSi-value: 0.56 ± 0.18 SD; t test:
t = -3.6, P = 0.001), but there were no differences between the latter two dietary groups
(t test: t = -0.8, P = 0.448). The somewhat higher and more variable PSi-values of the
benthivore group indicating that these charr are the most generalist feeding group.
0
0.1
0.2
0.3
0.4
0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
PSi-values
Pro
port
ion
of in
divi
dual
s
Fig. 1 Individual diet specialisation quantified as the proportional similarity (PSi) of the diet utilisation ineach individual and the total population of Arctic charr. Low PSi-values indicate low overlap between theindividuals and overall population diet niche, i.e. high level of individual specialisation. Zooplanktivorespecialists (open bars), gammaridivore specialists (black bars) and benthivores (grey bars)
594 Evol Ecol (2011) 25:589–604
123
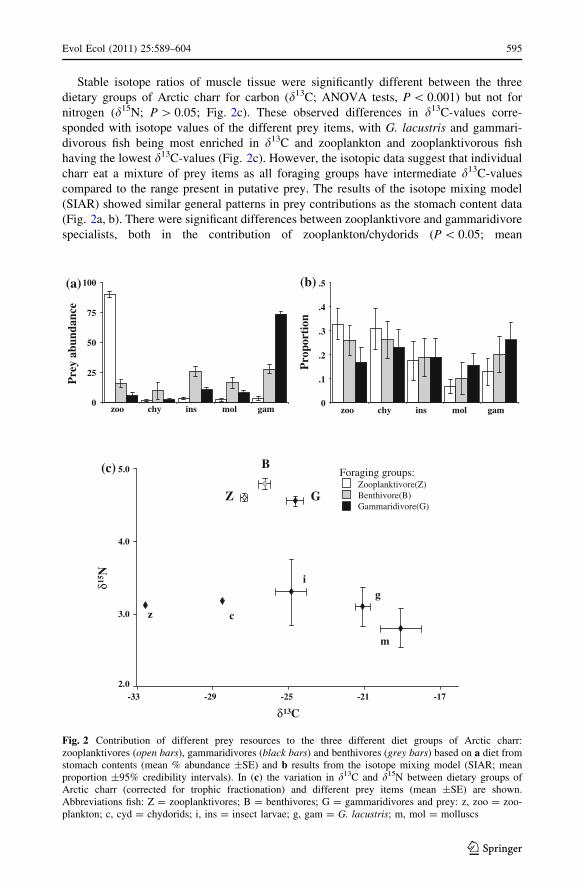
Stable isotope ratios of muscle tissue were significantly different between the three
dietary groups of Arctic charr for carbon (d13C; ANOVA tests, P \ 0.001) but not for
nitrogen (d15N; P [ 0.05; Fig. 2c). These observed differences in d13C-values corre-
sponded with isotope values of the different prey items, with G. lacustris and gammari-
divorous fish being most enriched in d13C and zooplankton and zooplanktivorous fish
having the lowest d13C-values (Fig. 2c). However, the isotopic data suggest that individual
charr eat a mixture of prey items as all foraging groups have intermediate d13C-values
compared to the range present in putative prey. The results of the isotope mixing model
(SIAR) showed similar general patterns in prey contributions as the stomach content data
(Fig. 2a, b). There were significant differences between zooplanktivore and gammaridivore
specialists, both in the contribution of zooplankton/chydorids (P \ 0.05; mean
(b)(a)
Pre
y ab
unda
nce
Pro
port
ion
0
25
50
75
100
0
.1
.2
.3
.4
.5
(c)
δ13C
2.0
3.0
4.0
zoo chy ins mol gamzoo chy ins mol gam
-33 -29 -25 -21 -17
δ15N
5.0
Z G
B
m
gi
cz
Foraging groups:Zooplanktivore(Z)Benthivore(B)Gammaridivore(G)
Fig. 2 Contribution of different prey resources to the three different diet groups of Arctic charr:zooplanktivores (open bars), gammaridivores (black bars) and benthivores (grey bars) based on a diet fromstomach contents (mean % abundance ±SE) and b results from the isotope mixing model (SIAR; meanproportion ±95% credibility intervals). In (c) the variation in d13C and d15N between dietary groups ofArctic charr (corrected for trophic fractionation) and different prey items (mean ±SE) are shown.Abbreviations fish: Z = zooplanktivores; B = benthivores; G = gammaridivores and prey: z, zoo = zoo-plankton; c, cyd = chydorids; i, ins = insect larvae; g, gam = G. lacustris; m, mol = molluscs
Evol Ecol (2011) 25:589–604 595
123
contribution for zooplanktivores: 63%, gammaridivores: 40%) and G. lacustris/molluscs
(P \ 0.05; zooplanktivores: 20%, gammaridivores: 42%). In benthivorous fish, the con-
tribution of these prey groups was intermediate to the two specialist groups. On the
individual level, the stomach contents corresponded nicely with the isotopic data (d13C) as
there was a positive correlation between the d13C-values and the abundance of G. lacustrisin the stomachs but a negative correlation with the abundance of zooplankton (Spearman
rank: 0.61 and -0.50, respectively; P \ 0.01).
Parasite loadings
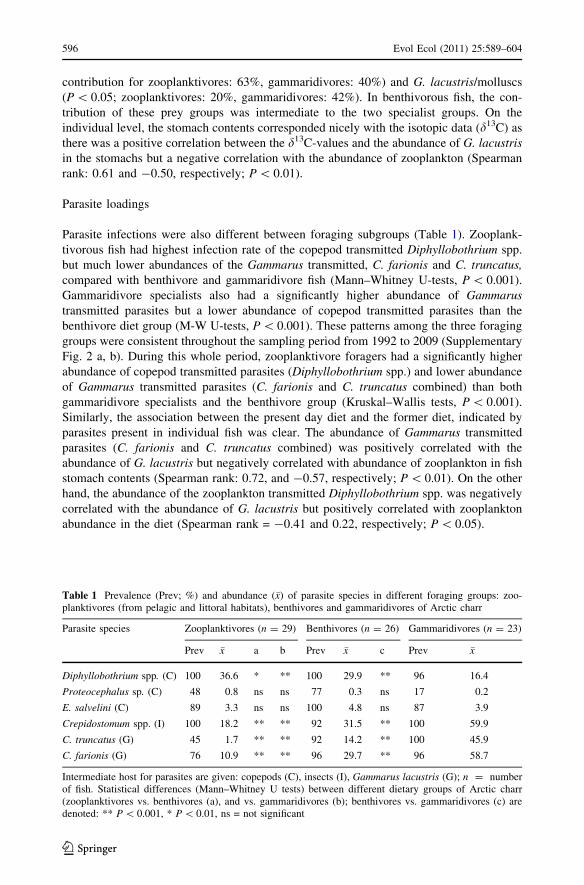
Parasite infections were also different between foraging subgroups (Table 1). Zooplank-
tivorous fish had highest infection rate of the copepod transmitted Diphyllobothrium spp.
but much lower abundances of the Gammarus transmitted, C. farionis and C. truncatus,compared with benthivore and gammaridivore fish (Mann–Whitney U-tests, P \ 0.001).
Gammaridivore specialists also had a significantly higher abundance of Gammarustransmitted parasites but a lower abundance of copepod transmitted parasites than the
benthivore diet group (M-W U-tests, P \ 0.001). These patterns among the three foraging
groups were consistent throughout the sampling period from 1992 to 2009 (Supplementary
Fig. 2 a, b). During this whole period, zooplanktivore foragers had a significantly higher
abundance of copepod transmitted parasites (Diphyllobothrium spp.) and lower abundance
of Gammarus transmitted parasites (C. farionis and C. truncatus combined) than both
gammaridivore specialists and the benthivore group (Kruskal–Wallis tests, P \ 0.001).
Similarly, the association between the present day diet and the former diet, indicated by
parasites present in individual fish was clear. The abundance of Gammarus transmitted
parasites (C. farionis and C. truncatus combined) was positively correlated with the
abundance of G. lacustris but negatively correlated with abundance of zooplankton in fish
stomach contents (Spearman rank: 0.72, and -0.57, respectively; P \ 0.01). On the other
hand, the abundance of the zooplankton transmitted Diphyllobothrium spp. was negatively
correlated with the abundance of G. lacustris but positively correlated with zooplankton
abundance in the diet (Spearman rank = -0.41 and 0.22, respectively; P \ 0.05).
Table 1 Prevalence (Prev; %) and abundance (�x) of parasite species in different foraging groups: zoo-planktivores (from pelagic and littoral habitats), benthivores and gammaridivores of Arctic charr
Parasite species Zooplanktivores (n = 29) Benthivores (n = 26) Gammaridivores (n = 23)
Prev �x a b Prev �x c Prev �x
Diphyllobothrium spp. (C) 100 36.6 * ** 100 29.9 ** 96 16.4
Proteocephalus sp. (C) 48 0.8 ns ns 77 0.3 ns 17 0.2
E. salvelini (C) 89 3.3 ns ns 100 4.8 ns 87 3.9
Crepidostomum spp. (I) 100 18.2 ** ** 92 31.5 ** 100 59.9
C. truncatus (G) 45 1.7 ** ** 92 14.2 ** 100 45.9
C. farionis (G) 76 10.9 ** ** 96 29.7 ** 96 58.7
Intermediate host for parasites are given: copepods (C), insects (I), Gammarus lacustris (G); n = numberof fish. Statistical differences (Mann–Whitney U tests) between different dietary groups of Arctic charr(zooplanktivores vs. benthivores (a), and vs. gammaridivores (b); benthivores vs. gammaridivores (c) aredenoted: ** P \ 0.001, * P \ 0.01, ns = not significant
596 Evol Ecol (2011) 25:589–604
123
Relationships between dietary specialisations, stable isotopes and food transmitted
parasites
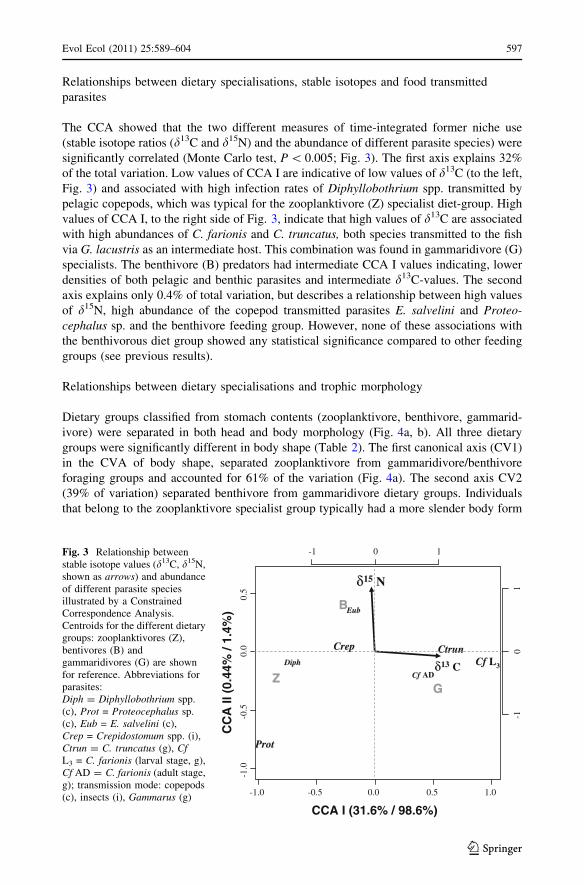
The CCA showed that the two different measures of time-integrated former niche use
(stable isotope ratios (d13C and d15N) and the abundance of different parasite species) were
significantly correlated (Monte Carlo test, P \ 0.005; Fig. 3). The first axis explains 32%
of the total variation. Low values of CCA I are indicative of low values of d13C (to the left,
Fig. 3) and associated with high infection rates of Diphyllobothrium spp. transmitted by
pelagic copepods, which was typical for the zooplanktivore (Z) specialist diet-group. High
values of CCA I, to the right side of Fig. 3, indicate that high values of d13C are associated
with high abundances of C. farionis and C. truncatus, both species transmitted to the fish
via G. lacustris as an intermediate host. This combination was found in gammaridivore (G)
specialists. The benthivore (B) predators had intermediate CCA I values indicating, lower
densities of both pelagic and benthic parasites and intermediate d13C-values. The second
axis explains only 0.4% of total variation, but describes a relationship between high values
of d15N, high abundance of the copepod transmitted parasites E. salvelini and Proteo-cephalus sp. and the benthivore feeding group. However, none of these associations with
the benthivorous diet group showed any statistical significance compared to other feeding
groups (see previous results).
Relationships between dietary specialisations and trophic morphology
Dietary groups classified from stomach contents (zooplanktivore, benthivore, gammarid-
ivore) were separated in both head and body morphology (Fig. 4a, b). All three dietary
groups were significantly different in body shape (Table 2). The first canonical axis (CV1)
in the CVA of body shape, separated zooplanktivore from gammaridivore/benthivore
foraging groups and accounted for 61% of the variation (Fig. 4a). The second axis CV2
(39% of variation) separated benthivore from gammaridivore dietary groups. Individuals
that belong to the zooplanktivore specialist group typically had a more slender body form
ProtProt
DiphDiph
EubEub
CrepCrep
CfCf ADAD
CtrunCtrunCfCf LL33
ZG
B
δδ1515 NN
δδ1313 CC
-1.0
-0.5
0.0
0.5
CCA I (31.6% / 98.6%)
CC
A II
(0.
44%
/ 1.
4%)
-10
1
-1.0 -0.5 0.0 0.5 1.0
-1 0 1Fig. 3 Relationship betweenstable isotope values (d13C, d15N,shown as arrows) and abundanceof different parasite speciesillustrated by a ConstrainedCorrespondence Analysis.Centroids for the different dietarygroups: zooplanktivores (Z),bentivores (B) andgammaridivores (G) are shownfor reference. Abbreviations forparasites:Diph = Diphyllobothrium spp.(c), Prot = Proteocephalus sp.(c), Eub = E. salvelini (c),Crep = Crepidostomum spp. (i),Ctrun = C. truncatus (g), CfL3 = C. farionis (larval stage, g),Cf AD = C. farionis (adult stage,g); transmission mode: copepods(c), insects (i), Gammarus (g)
Evol Ecol (2011) 25:589–604 597
123
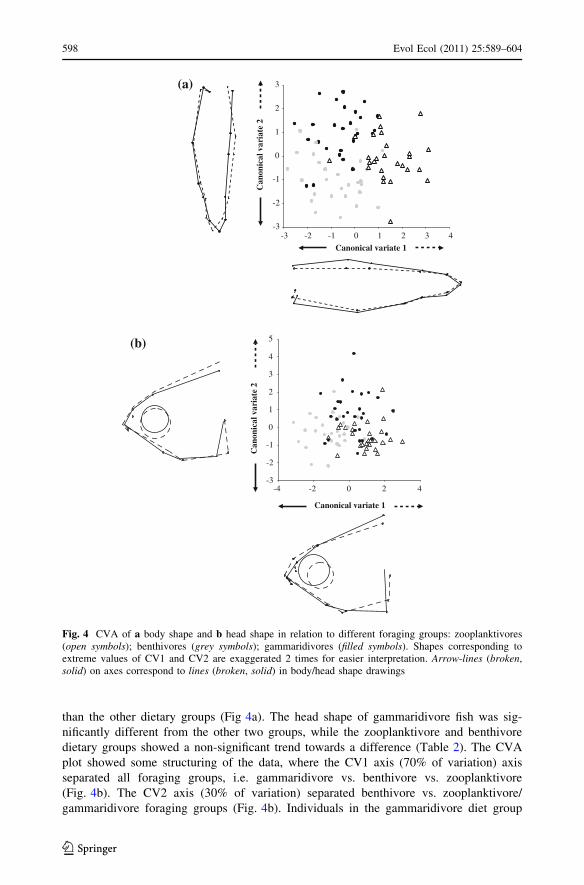
than the other dietary groups (Fig 4a). The head shape of gammaridivore fish was sig-
nificantly different from the other two groups, while the zooplanktivore and benthivore
dietary groups showed a non-significant trend towards a difference (Table 2). The CVA
plot showed some structuring of the data, where the CV1 axis (70% of variation) axis
separated all foraging groups, i.e. gammaridivore vs. benthivore vs. zooplanktivore
(Fig. 4b). The CV2 axis (30% of variation) separated benthivore vs. zooplanktivore/
gammaridivore foraging groups (Fig. 4b). Individuals in the gammaridivore diet group
(a)
-3
-2
-1
0
1
2
3
-3 -2 -1 0 1 2 3 4
Canonical variate 1C
anon
ical
var
iate
2
-3
-2
-1
0
1
2
3
4
5
-4 -2 0 2 4
Canonical variate 1
Can
onic
al v
aria
te2
(b)
Fig. 4 CVA of a body shape and b head shape in relation to different foraging groups: zooplanktivores(open symbols); benthivores (grey symbols); gammaridivores (filled symbols). Shapes corresponding toextreme values of CV1 and CV2 are exaggerated 2 times for easier interpretation. Arrow-lines (broken,solid) on axes correspond to lines (broken, solid) in body/head shape drawings
598 Evol Ecol (2011) 25:589–604
123

typically had blunter snouts and more robust heads, while zooplanktivore individuals had
longer snouts and lower head depths compared to the two other feeding groups.
Discussion
In this study we tested 3 hypotheses directly related to resource use and its role in eco-
logically driven evolutionary divergence. Phenotypic plasticity is known to result in epi-
genetic modification of traits in charr when individuals are exposed to differing resources
over periods which are broadly similar to an individual’s lifetime. Here we show that
individual Arctic charr demonstrating foraging specialisms, identified by measures that
integrate over very short temporal periods (hours) showed clear evidence of persistent
foraging specialisation over much longer temporal periods (months and years). In Fjell-
frøsvatn, Arctic charr individuals exhibiting dietary specialism, defined by their stomach
contents, providing a temporally restricted measure of diet over the preceding hours only,
also showed significant differences in two trophic tracers with a much longer integration
period. Muscle stable isotopes (SI) signatures integrate over months (Perga and Gerdeaux
2005) and parasite fauna integrates over months and years (Knudsen et al. 2004). Thus,
individuals identified as zooplanktivores based on their stomach contents, had depleted
d13C values in muscle tissue, a high number of copepod transmitted parasites (CTP) and a
low abundance of Gammarus-transmitted parasites (GTP). In contrast, individuals with
stomach contents dominated by macrobenthos, exhibited enriched d13C values, a high
number of GTP and a low abundance of CTP. Individuals categorised as gammarid spe-
cialists on the basis of their stomach contents, showed the most enriched muscle d13C and
had the highest abundance of Gammarus-transmitted parasites. Despite that foraging
behaviour is often regarded as a highly labile trait (e.g. West-Eberhardt 2005; Duckworth
2009), we conclude that individual foraging specialisms identified from stomach contents,
do persist in Fjellfrøsvatn charr over periods that are sufficient to induce ontogenetic
plastic responses in individuals (e.g. Day and McPhail 1996; Adams et al. 2003; Alexander
and Adams 2004; Garduno-Paz et al. 2010) that is, broadly equivalent to the life-time of an
individual. This finding is consistent with models where alternative resource specialism
drives phenotypic plasticity in the earliest stages of divergence (Skulason et al. 1999).
Secondly we show, at the level of the population, stability in foraging specialisms that
persist over a period of almost two decades. The expression of alternative foraging spe-
cialisms that persists over multiple generations is a key requirement for ecologically driven
evolution through resource specialisation (Dieckmann and Doebeli 1999; Schluter 2000).
Thirdly we show that individuals demonstrating foraging specialisations also showed
morphological adaptations with a functional significance related to foraging. Thus the
gammaridivore specialists had blunter snout and more robust head and body compared
with zooplanktivore specialists. Benthivorous individuals were intermediate between the
Table 2 Pairwise comparisons (P-values from permutation tests of Mahalanobis distances in shape) ofdifferences in trophic morphology (body and head shape) between the three dietary groups of Arctic charr
Body shape Head shape
Zooplanktivores vs. gammaridivores \0.001 \0.001
Zooplanktivores vs. benthivores \0.001 0.066
Benthivores vs. gammaridivores \0.001 0.003
Evol Ecol (2011) 25:589–604 599
123

two other foraging groups according to both head shape and body shape. Similar patterns
of functional body shape differences between zooplanktivore and benthivore specialists
have been widely reported amongst a number of postglacial fishes in general, and spe-
cifically in Arctic charr elsewhere (Robinson and Parsons 2002; Knudsen et al. 2007;
Hendry 2009).
Despite high levels of flexibility in foraging behaviour (e.g. West-Eberhard 2005;
Duckworth 2009), variation in behavioural traits relating to foraging and prey selection in
Arctic charr has been shown in some populations, to have at least some genetic component
to its expression (Adams and Huntingford 2002a,b; Klemetsen et al. 2002, 2006; Albertson
et al. 2005; Roger and Bernatchez 2007). Similarly, expression of alternative head mor-
phologies associated with foraging specialisms, has recently been linked to ecologically
based, disruptive selection in fish and seems to be genetically controlled by a very few loci
(Albertson et al. 2005; Tripathi et al. 2009) but has also been shown to be highly plastic
(Adams et al. 2003; Alexander and Adams 2004; Adams and Huntingford 2004). The
ultimate mechanisms regulating the expression of resource specialisation and morpho-
logical traits are not known for the model system investigated here. However it is most
likely that expression of the variation in foraging behaviour and morphology shown here is
the result of an interplay between genes and the environment (Schluter 2000; Adams and
Huntingford 2004; Abrams 2006; Rueffler et al. 2007).
One important outcome of this study in the context of evolutionary divergence, is that
the foraging specialists described here are subject to differential selection pressures.
Individuals adopting differing foraging specialisms in this study occupied very different
and contrasting habitat types. Thus the zooplankton specialists were predominantly col-
lected from, and clearly foraged in, the pelagic zone of the lake. In contrast the macro-
benthos and Gammarus foraging specialists were mainly collected from, and foraged in,
the littoral zone of the lake. These two environments differ substantially across a wide
range of physical and biotic characteristics (e.g. light penetration, habitat heterogeneity,
predation vulnerability, wave energy, etc.) strongly suggesting that specialists occupying
these alternative habitats would be subject to different suite of selection pressures. In
addition, the predator characteristics (e.g. the behavioural search image and capture
techniques) required of successful specialists foraging on the three different prey resources
in this study are very different (Day and McPhail 1996; Fraser et al. 2008).
Parasites have been shown to result in selection pressures with evolutionary conse-
quences for the host population (e.g. Maan et al. 2008; Blanchet et al. 2009; MacColl
2009). The foraging specialists in this study exhibited very different parasite fauna and
infection rates. This is also likely to have a significant impact on the selection pressures to
which each foraging specialist was exposed. Several of the endoparasites observed in the
current study are transmitted by foraging on specific intermediate hosts. In this study
zooplankton specialists had a higher abundance of Diphyllobothrium spp., and a lower
abundance of Crepidostomum spp., C. truncatus and C. farionis parasites than both
macrobenthos and gammarus specialists. The macrobenthos specialists had a higher par-
asite abundance of Diphyllobothrium spp., and a lower abundance of Crepidostomum spp.,
C. truncatus and C. farionis parasites than gammarus specialists. C. farionis, C. truncatus,
and Diphyllobothrium spp. have been shown to place direct fitness costs on their fish host
(see e.g. Curtis 1984; Knudsen et al. 2004, 2008). In sticklebacks limnetic and benthic
morph pairs show differential immune responses (Matthews et al. 2010) probably in order
to be able to specifically deal with the contrasting parasite faunas often found among
trophically diverged species pairs (Knudsen et al. 2003; MacColl 2009). Parasites often
have a dose dependent effect on their host, where the most parasitised individuals run the
600 Evol Ecol (2011) 25:589–604
123

highest risk of being affected. Thus, dietary specialists run a high risk of suffering from
parasite infections as they accumulate high densities of a restricted group of parasites
(Amundsen et al. 2003; Knudsen et al. 2004), but may also benefit from a reduced
investment in a costly immune defence (Moret and Schmid-Hempel 2000). By concen-
trating the immune responses against a few parasite species, specialist foragers may be able
to respond effectively to manage infection levels. An ability to withstand the effects of a
specific, restricted group of parasite larvae may then allow a specialist predator to exploit a
potentially infected prey group without accumulating the high costs associated with
defense against a more diverse range of parasite types (Lafferty et al. 2000). Thus it is
highly likely that parasites are causing parasite-mediated differential selection pressures
amongst the three specialist charr foragers in Fjellfrøsvatn and that these and other iden-
tified differential selection pressures have remained consistent for at least 2 decades (see
also Knudsen 2010).
In the early process of ecologically driven speciation, theoretical models typically
include resource specialisation by individuals in a population adapted specifically to dif-
ferential resource usage in alternative environments (Dieckmann and Doebeli 1999; Sch-
luter 2000; Rueffler et al. 2007). The importance of dietary specialisation in sustaining
isolated morphs was recently highlighted in a collapsing divergent morph pair of stick-
lebacks (Behm et al. 2010). In the present study, where multiple co-occurring behavioural
specialists, adapted to a variety of distinct ecological niches, may have arisen as a result of
disruptive selection initiating the speciation process. Populations may appear at different
stages within the evolutionary diversification process, but not all diverging populations will
result in speciation (Hendry 2009; Nosil et al. 2009). If ecologically significant traits are
directly linked to mate choice (magic traits; sensu Gavrilets 2004), positive assortative
mating could be an important mechanism in promoting the likelihood of reproductive
isolation. Adaptive ecological traits related to visual cues and signals like trophic behav-
iour, body size and colouration (e.g. dietary-based carotenoid red ornamentation) seem
important in mate recognition and assortative mating of both reproductively isolated
postglacial fishes and in unimodal populations that are adapted to different dietary niches
(Snowberg and Bolnick 2008; Head et al. 2009; Stelkens and Seehausen 2009). Thus, the
behaviour and/or morphological traits related to alternative niche adaptation observed in
this study could eventually be involved in reducing gene flow among distinct phenotypes.
The observed incidence of multiple co-occurring specialist ecotypes adapted to a variety
of distinct ecological niches, has support in recent model studies, highlighting that adaptive
radiation may result in stable multiple morphs (e.g. Abrams 2006; Bolnick 2006; Doebeli
et al. 2007). In Fjellfrøsvatn, a deep-water morph has been found to be reproductively
isolated and, genetically and ecologically distinct from the ecotypes examined here
(Klemetsen et al. 1997; Westgaard et al. 2004; Amundsen et al. 2008) with specific
behavioural and morphological adaptations allowing the utilisation of a narrow profundal
soft bottom dietary niche (Klemetsen et al. 2002, 2006; Knudsen et al. 2006). Other studies
have also highlighted that locally adapted benthic morphs may live in sympatry (e.g.
Snorrason et al. 1994; Kahilainen and Østbye 2006). These results suggest that local
adaptations (Ravigne et al. 2009; Losos 2010) to alternative niches can occur beyond the
traditional view of divergence into littoral-pelagic morph-pairs which has been commonly
observed among postglacial fishes (reviewed by Robinson and Parsons 2002).
We conclude that these results suggest that differential selection pressures from several,
separate ecological processes are involved in shaping the stable foraging niches of several
different co-occurring ecotypes. The present lake ecosystem with stable resource spe-
cialists represents a promising candidate for complementary studies addressing how
Evol Ecol (2011) 25:589–604 601
123

plasticity and heritability of traits are involved in the early process of speciation and which
definitive mechanisms are involved in the reproductive isolation process. Our findings are
thus particularly important in respect to understanding the mechanisms of incipient spe-
ciation, which appear to be related to a subtle interplay between ecological and evolu-
tionary processes at the individual and population level.
Acknowledgments We thank Laina Dalsbø, Jan Evjen and Pal Sørensen for field and laboratory assis-tance, Raul Primicerio for statistical advice and Jennifer Dodd for grammar checking of the manuscript. Wealso thank two anonymous referees for their constructive criticisms.
References
Abrams PA (2006) The effects of switching behaviour on the evolutionary diversification of generalistconsumers. Am Nat 168:645–659
Adams CE, Huntingford FA (2002a) Inherited differences in head allometry in polymorphic charr fromLoch Rannoch, Scotland. J Fish Biol 60:515–520
Adams CE, Huntingford FA (2002b) The functional significance of inherited differences in feeding mor-phology in sympatric polymorphic population of Arctic charr. Evol Ecol 16:15–25
Adams CE, Huntingford FA (2004) Incipient speciation driven by phenotypic plasticity? Evidence fromsympatric populations of Arctic charr. Biol J Linn Soc 81:611–618
Adams CE, Fraser D, Huntingford FA, Greer RB, Askew CM, Walker AF (1998) Trophic polymorphismamongst Arctic charr from Loch Rannoch, Scotland. J Fish Biol 52:1259–1271
Adams CE, Woltering C, Alexander G (2003) Epigenetic regulation of trophic morphology through feedingbehaviour in Arctic charr, Salvelinus alpinus. J Fish Biol 78:43–49
Adams DC, Rohlf FJ, Slice DE (2004) Geometric morphometrics: ten years of progress following the‘revolution’. Ital. J. Zool. 71:5–16
Albertson RC, Streelman JT, Kocher TD, Yelick PC (2005) Integration and evolution of the cichlid man-dible: The molecular basis of alternate feeding strategies. PNAS 102:16287–16292
Alexander GD, Adams CE (2004) Exposure to a common environments erodes between-strain trophicmorphology differences in Arctic charr. J Fish Biol 64:254–258
Amundsen P-A, Gabler H-M, Staldvik FJ (1996) A new approach to graphical analysis of feeding strategyfrom stomach contents data–modification of the Costello (1990) method. J Fish Biol 48:607–614
Amundsen P-A, Knudsen R, Kuris AM, Kristoffersen R (2003) Seasonal and ontogenetic dynamics introphic transmission of parasites. Oikos 102:285–293
Amundsen P-A, Knudsen R, Klemetsen A (2008) Seasonal and ontogenetic variation in resource use by twosympatric Arctic charr populations. Environ Biol Fish 83:45–55
Behm JE, Ives AR, Boughman JW (2010) Breakdown in postmating isolation and the collapse of a speciespair through hybridization. Am Nat 175:11–26
Blanchet S, Rey O, Berthier P, Lek S, Loot G (2009) Evidence of parasite-mediated disruptive selection ongenetic diversity in a wild fish population. Mol Ecol 18:1112–1123
Bolnick DI (2006) Multi-species outcomes in a common model of sympatric speciation. J Theor Biol241:734–744
Bolnick DI, Yang LH, Fordyce JA, Davis JM, Svanback R (2002) Measuring individual-level resourcespecialization. Ecology 83:2936–2941
Bolnick DI, Svanback R, Fordyce JA, Yang LH, Davis JM, Hulsey CD, Forister ML (2003) The ecology ofindividuals: incidence and implications of individual specialisation. Am Nat 161:1–28
Bush AO, Lafferty KD, Lotz JM, Shostak AW (1997) Parasitology meets ecology on its own terms:Margolis et al. revisited. J Parasitol 83:575–583
Curtis MA (1984) Diphyllobothrium spp. and the Arctic charr: parasite acquisition and its effects on a lakeresident population. In: Johnson L, Burns BI (eds) Biology of the Arctic charr. Proceedings of theInternational Symposium on a Arctic charr, Winnipeg, Mannitoba, May 1981. University of ManitobaPress, Winnipeg, pp 395–411
Day T, McPhail JD (1996) The effect of behavioural and morphological plasticity on foraging efficiency inthe threespine stickleback (Gasterosteus sp.). Oecologia 108:380–388
Dieckmann U, Doebeli M (1999) On the origin of species by sympatric speciation. Nature 400:354–357Doebeli M, Block HJ, Leimar O, Dieckmann U (2007) Multimodal pattern formation in phenotype distri-
butions of sexual populations. Proc R Soc B 274:347–357
602 Evol Ecol (2011) 25:589–604
123

Duckworth RA (2009) The role of behaviour in evolution: a search for mechanisms. Evol Ecol 23:513–531Fraser D, Huntingford FA, Adams CE (2008) Foraging specialisms, prey size and life-history patterns: a test
of predictions using sympatric polymorphic Arctic charr (Salvelinus alpinus). Ecol Freshw Fish 17:1–9Funk DJ (2010) Does strong selection promote host specialisation and ecological speciation in insect
herbivores? Evidence from Neochlamisus leaf beetles. Ecol Entomol 35:41–53Garduno-Paz MV, Adams CE (2010) Discrete prey availability promotes foraging segregation and early
divergence in Arctic charr, Salvelinus alpinus. Hydrobiologia 650:15–26Garduno-Paz MV, Couderc S, Adams CE (2010) Habitat modulates phenotypic expression through
developmental plasticity in the three-spined stickleback. Biol J Linn Soc 100:407–413Gavrilets S (2004) Fitness landscapes and the origin of species. Princeton University Press, NJ, p 432Head ML, Price EA, Boughman JW (2009) Body size differences do not arise from divergent mate pref-
erences in a species pair of threespine stickleback. Biol Lett 5:517–520Hendry AP (2009) Ecological speciation! Or the lack thereof? Can J Fish Aquat Sci 66:1383–1398Herrel A, Huyghe K, Vanhooydonck B, Backeljau T, Breugelmans K, Grbac I, Van Damme R, Irschick DJ
(2008) Rapid large-scale evolutionary divergence in morphology and performance associated withexploitation of a different dietary resource. Proc Natl Acad Sci USA 105:4792–4795
Kahilainen KK, Østbye K (2006) Morphological differentiation and resource polymorphism in three sym-patric whitefish Coregonus lavaretus (L.) forms in a subarctic lake. J Fish Biol 68:63–79
Klemetsen A (2010) The charr problem revisited: exceptional phenotypic plasticity promotes ecologicalspeciation in postglacial lakes. Freshw Rev 3:49–74
Klemetsen A, Amundsen P-A, Knudsen R, Hermansen B (1997) A profundal, winter-spawning morph ofArctic charr Salvelinus alpinus (L.) in lake Fjellfrøsvatn, northern Norway. Nord J Freshw Res73:13–23
Klemetsen A, Elliot JM, Knudsen R, Sørensen P (2002) Evidence for genetic differences in the offspring oftwo sympatric morphs of Arctic charr. J Fish Biol 60:933–950
Klemetsen A, Knudsen R, Primicerio R, Amundsen P-A (2006) Divergent natural selection on the feedingbehaviour of two sympatric Arctic charr (Salvelinus alpinus) morphs. Ecol Freshw Fish 15:350–355
Klingenberg CP (2008) MorphoJ. Faculty of Life Sciences, University of Manchester, UK. Available athttp://www.flywings.org.uk/MorphoJ_page.htm
Knudsen R, Amundsen P-A, Klemetsen A (2003) Inter-and intra-morph patterns in helminth communities ofsympatric whitefish morphs. J Fish Biol 62:847–859
Knudsen R, Curtis MA, Kristoffersen R (2004) Aggregation of helminths: the role of feeding behaviour offish hosts. J Parasitol 90:1–7
Knudsen R, Klemetsen A, Amundsen P-A, Hermansen B (2006) Incipient speciation through nicheexpansion: an example from the Arctic charr in a subarctic lake. Proc R Soc B 273:2291–2298
Knudsen R, Amundsen P-A, Primicerio R, Klemetsen A, Sørensen P (2007) Contrasting niche-based var-iation on trophic morphology within Arctic charr populations. Evol Ecol Res 9:1005–1021
Knudsen R, Jobling M, Amundsen P-A, Klemetsen A (2008) Differences in pyloric caeca morphologybetween Arctic charr ecotypes: adaptation to trophic specialisation or parasite-induced phenotypicmodifications? J Fish Biol 73:275–287
Knudsen R, Primicerio R, Amundsen P-A, Klemetsen A (2010) Temporal stability of individual feedingspecialization may promote speciation. J Anim Ecol 79:161–168
Kristjansson B, Skulason S, Noakes DL (2002) Morphological segregation of Icelandic threespine stick-leback (Gasterosteus aculeatus L). J Fish Biol 76:247–257
Lafferty KD, Thomas F, Skorping A (2000) Evolution of host phenotype manipulation by parasites and itsconsequences. Develop Anim Vet Sci 32:117–127
Legendre P, Legendre L (1998) Numerical ecology. Elsevier, AmsterdamLosos JB (2010) Adaptive radiation, ecological opportunity, and evolutionary determinism. Am Nat
175:623–639Maan ME, Van Rooijen AMC, Van Alphen JJM, Seehausen O (2008) Parasite-mediated sexual selection
and species divergence in Lake Victoria cichlid fish. Biol J Linn Soc 94:53–60MacColl ADC (2009) Parasite may contribute to ‘magic trait’ evolution in the adaptive radiation of three-
spined sticklebacks, Gasterosteus aculeatus (Gasterosteiformes: Gasterosteidae). Biol J Linn Soc96:425–433
Matthews B, Harmon LJ, M’Gonigle L, Marchinko KB, Schaschol H (2010) Sympatric and allopatricdivergence of MHC genes in threespined stickleback. Plos One 5:e10948
Moret Y, Schmid-Hempel P (2000) Survival for immunity: the price of immune system for activation ofbumblebee workers. Science 290:1166–1168
Nosil P, Harmon JL, Seehausen O (2009) Ecological explanations for (incomplete) speciation. Trends EcolEvol 24:145–146
Evol Ecol (2011) 25:589–604 603
123

Parnell AC, Inger R, Bearhop S, Jackson AL (2010) Source partitioning using stable isotopes: coping withtoo much variation. Plos One 5:e9672
Perga ME, Gerdeaux D (2005) ‘Are fish what they eat’ all year round? Oecologia 144:598–606Pfennig DW, Wund MA, Snell-Rodd EC, Cruickshank T, Schlicthing CD, Moczek AP (2010) Phenotypic
plasticity’s impacts on diversification and speciation. Trends Ecol Evol 25:459–467Pinnegar JK, Polunin NVC (1999) Differential fractionation of d13C and d15N among fish tissues: impli-
cations for the study of trophic interactions. Func Ecol 13:225–231Post DM (2002) Using stable isotopes to estimate trophic position: models, methods, and assumptions.
Ecology 83:703–718R Development Core Team (2009) R: a language and environment for statistical computing. R Foundation
for Statistical Computing, ViennaRavigne V, Dieckmann U, Olivieri I (2009) Live where you thrive: joint evolution of habitat choice and
local adaptation facilitates specialization and promotes diversity. Am Nat 174:E141–E169Robinson BW, Parsons KJ (2002) Changing times, spaces, and faces: tests and implications of adaptive
morphological plasticity in the fishes of northern postglacial lakes. Can J Fish Aquat Sci 59:1819–1833Roger SM, Bernatchez L (2007) The genetic architecture of ecological speciation and the association with
signatures of selection in natural lake whitefish (Coregonus sp. Salmonidae) species pairs. Mol BiolEvol 24:1423–1438
Rohlf FJ (2006) TPS Dig version 2.10. Deptartment of Ecology and Evolution, State University of NewYork at Stony Brook. Available at http://life.bio.sunysb.edu/morph
Rohlf FJ, Marcus LF (1993) A revolution in morphometrics. Trends Ecol Evol 8:129–132Rohlf FJ, Slice DE (1990) Extensions of the procrustes method for the optimal superimposition of land-
marks. Syst Zool 39:40–59Rueffler C, Van Dooren TJM, Metz AJ (2007) The interplay between behaviour and morphology in the
evolutionary dynamics of resource specialisation. Am Nat 169:E34–E52Sandlund OT, Gunnarsson K, Jonasson PM, Jonsson B, Lindem T, Magnusson KP, Malmquist HJ, Si-
gurjonsdottir H, Skulason S, Snorrason SS (1992) The Arctic charr Salvelinus alpinus in Thingva-llavatn. Oikos 64:305–335
Sargeant BL (2007) Individual foraging specialization: niche width versus niche overlap. Oikos116:1431–1437
Schluter D (1996) Ecological speciation in postglacial fishes. Phil Trans R Soc Lond B 351:807–814Schluter D (2000) The ecology of adaptive radiation. Oxford University Press, UKSchluter D (2001) The ecology and origin of species. Trends Ecol Evol 16:372–380Schluter D, Conte GL (2009) Genetics and ecological speciation. Proc Natl Acad Sci U S A 106:9955–9962Skarstein F, Folstad I (1996) Sexual dicromatism and the immunocompetence handicap: an observational
approach using Arctic charr. Oikos 76:359–367Skulason S, Smith TB (1995) Resource polymorphism in vertebrates. Trends Ecol Evol 10:366–370Skulason S, Snorrason SS, Jonsson B (1999) Sympatric morphs, populations and speciation in freshwater
fish with emphasis on Arctic charr. In: Magurran AE, May RM (eds) Evolution of biological diversity.Oxford University Press, Oxford, pp 70–92
Snorrason SS, Skulason S, Jonsson B, Malmquist HJ, Jonasson PM, Sandlund OT, Lindem T (1994) Trophicspecialization in Arctic charr Salvelinus alpinus (Pisces; Salmonidae): morphological divergence andontogenetic niche shifts. Biol J Linn Soc 52:1–18
Snowberg LK, Bolnick DI (2008) Assortative mating by diet in a phenotypically unimodal but ecologicalvariable population of stickleback. Am Nat 172:733–739
Stelkens RB, Seehausen O (2009) Phenotypic divergence but not genetic distance predicts assortativemating among species of a cichlid fish radiation. J Evol Biol 22:1679–1694
Tripathi N, Hoffmann M, Willing E-M, Lanz C, Weigel D, Dreyer C (2009) Genetic linkage map of theguppy, Poecilia reticulata, and quantitative trait loci analysis of male size and colour variation. Proc RSoc B 276:2195–2208
West-Eberhard MJ (1989) Phenotypic plasticity and the origins of diversity. Ann Rev Ecol Syst 20:249–278West-Eberhardt MJ (2005) Developmental plasticity and the origins of species differences. Proc Natl Acad
Sci U S A 102:6543–6549Westgaard JI, Klemetsen A, Knudsen R (2004) Genetic differences between two sympatric morphs of Arctic
charr Salvelinus alpinus (L.) confirmed by microsatellite DNA. J Fish Biol 65:1185–1191Woo KJ, Elliot KH, Davidson M, Gaston AJ, Davoren GK (2008) Individual specialization in diet by a
generalistic marine predator reflects specialisation in foraging behaviour. J Anim Ecol 77:1082–1091Zelditch ML, Swiderski DL, Sheets HD, Fink WL (2004) Geometric morphometrics for biologists: a primer.
Elsevier, New York
604 Evol Ecol (2011) 25:589–604
123




















