
Seasonal spatial patterns of two sympatric frogs: California Red-legged Frog and American Bullfrog
Upload
independentCategory
view
7download
0
Journal of Fish Biology (1998) 53 (Supplement A), 37–55Article No. jb980812
Direct mate choice maintains diversity among sympatriccichlids in Lake Victoria
O. S*, F. W, J. J. M. A N. B
Institute of Evolutionary and Ecological Sciences, University of Leiden, P.O. Box 9516,NL-2300 Leiden, The Netherlands
Mate choice may play an important role in animal speciation. The haplochromine cichlids ofLake Victoria are suitable to test this hypothesis. Diversity in ecology, coloration and anatomyevolved in these fish faster than postzygotic barriers to gene flow, and little is known about howthis diversity is maintained. It was tested whether recognizable forms are selection-maintainedmorphs or reproductively isolated species by investigating in the field reproductive timing,location of spawning sites, and mate choice behaviour. There was a large interspecific overlapin timing of breeding and location of spawning sites, which was largest in members of the samegenus. Behavioural mate choice of such closely related taxa was highly assortative, such that itis likely that they are sexually isolated species and that direct mate choice is the major force thatdirects gene flow and maintains form diversity. The results differ from what is known aboutrecent radiations of other lacustrine fish groups where speciation seems to be driven bydiverging microhabitat preferences or diverging timing of reproduction, but are in agreementwith predictions from models of speciation by diverging mate preferences.
? 1998 The Fisheries Society of the British Isles
Key words: adaptive radiation; mate recognition; polymorphism; reproductive isolation; sexualselection; speciation.
*Author to whom correspondence should be addressed. Tel.: +31-71-5274916; fax: +31-71-5274900;email: [email protected]
INTRODUCTION
Ecosystems that produce and accommodate high species numbers areparticularly interesting for ecologists and evolutionists trying to understand theorigin and maintenance of biodiversity. The East African Great Lakes containthe most diverse fish species assemblages in inland water bodies. Lake Victoria’sspecies flock of endemic haplochromine cichlids is considered an outstandingexample of explosive evolution and adaptive radiation in vertebrates(Greenwood, 1981; Futuyma, 1986). Though the lake basin may be as old as750 000 years (Beadle, 1981), recent evidence suggests that a pleistocene droughtperiod rendered the lake entirely dry 12 400 years ago: the persistence of satellitelakes during this drought is considered to be unlikely, leading to the suggestionthat the vast majority of the over 500 putative species have evolved since then(Johnson et al., 1996). Their evolutionary youth is reflected in geneticalcompatibility (Crapon de Caprona, 1986; Crapon de Caprona & Fritzsch,1984; Seehausen et al., 1997a) and very little molecular genetical divergence(Meyer et al., 1990) among the diverse morphotypes. Though most of the formsthat had been studied in the field over several years or decades retained theirspecific appearance over many generations, there are also examples wherephenotypic appearance of natural populations changed, possibly as a
37
0022–1112/98/53A037+19 $30.00/0 ? 1998 The Fisheries Society of the British Isles

38 . .
consequence of hybridization (Seehausen et al., 1997a,b), and there is little hardevidence to corroborate the assertion of taxonomists and ecologists that therecognizable forms are maintained by reproductive isolation. Alternativelyseveral forms might represent selection-maintained morphs of a smaller numberof species (Liem & Kaufman, 1984; Meyer, 1987). In the absence of molecularmarkers, there are two approaches to test these alternatives: studying reproduc-tion in the natural environment (Holzberg, 1978; Marsh et al., 1981), andbreeding in captivity to study the genetic basis of phenotypic divergence(Seehausen et al., 1997a). The present study used the first approach.
Most of the haplochromine cichlids of Lake Victoria exhibit conspicuoussexual dimorphism with brightly coloured males and cryptically colouredfemales. Such sexual dimorphism is common in animal groups that haveundergone rapid diversification, indicating a role of sexual selection in speciation(Coyne, 1992). It has been shown experimentally that male breeding color-ation can play an important role in female mate choice of Lake Victoriahaplochromines (Seehausen & van Alphen, 1998). However, it had not beentested whether sympatric forms with different male coloration constitute sexuallyisolated species in their natural environment, and how important mate choice isto the maintenance of any isolation. Mating behaviour in nature had hithertonot been studied in Lake Victoria because of its relatively turbid waters.
This study tested under natural conditions the null hypothesis that morpho-logically and ecologically defined sympatric forms, putative species, arereproductively not isolated; and what, if anything, maintains reproductiveisolation. Knowledge on how diversity is maintained in a system in the absenceof postzygotic isolation can shed some light on probabilities of alternative waysof speciation.
MATERIALS AND METHODS
The haplochromine cichlids of Lake Victoria are polygynous female mouthbrooders.Males occupy territories from which they exclude other males, and into which theyattempt to attract ripe females. Courtship and spawning take place in these territories.Females, quiescent males and subadults live and forage in small groups or larger shoals.Temporal, spatial and behavioural aspects of reproduction were studied at the rockyMakobe Island, a flat reef in the entrance to the Speke Gulf of Lake Victoria, 5 km off themainland shore. Makobe Island harbours more than 20 sympatric putative species(different in ecology, anatomy and coloration) and several putatively conspecific colourmorphs (different only in coloration) in an extensive shallow water zone which, withrelatively clear waters, provide suitable conditions for scuba diving (Seehausen & Bouton,1997).
TEMPORAL PATTERNS OF REPRODUCTIONTo test for temporal isolation (reproductive allochrony), developmental stages of
gonads were studied in two sample series: a 12-month series of three putative species, anda 3-month series of more than 10 putative species. Samples of the three, in shallow waternumerically dominant putative species, one of which comprises three colour morphs,were taken in monthly intervals from June 1990 to July 1991, to test for reproductiveseasonality. To obtain the samples, gillnets of mesh-sizes 31·8, 38·1 and 44·5 mmstretched mesh were set between 0830 and 1230 hours in inshore shallow water (1·5–2·5 mdepth). It was attempted to collect at least 10 adult males of each species, and as manyas possible of the more difficult to collect females.

39
After collection, the fishes were stored on ice, and their standard length (LS) wasmeasured with digital calipers to the nearest 0·5 mm. In the 12-month study, thedevelopmental stages of gonads were recorded on formalin preserved fishes. In the3-month study, they were recorded in fresh specimens, between 3 and 5 h after capture.Scores between 1 and 6 were given for ovaries and testes (gonads not visible; immature;early ripening or mature but inactive; late ripening; ripe; spent). For each species and sexthe gonadal stage was plotted against standard length and the size at 50% maturitydetermined as the size at which 50% of the individuals were in stage 4 or higher. Sinceruns tests (Sokal & Rohlf, 1981) for seasonality in size (LS) were not significant, onlyindividuals larger than size at 50% maturity were considered in further data analysis.
Runs tests for above and below the median (Sokal & Rohlf, 1981) were performed onmonthly mean gonad stages and on the monthly proportion of ripe individuals to test theH0 that mean gonad stage and the proportion of ripe fishes do not exhibit seasonality.Runs tests can detect temporal variation only when seasonal trends last long enough toextend over several (three or more) sample time units. To test whether the observedvariation in gonad stages was explained by shorter time effects, ANOVAs were calculatedwith gonad stage as dependent variable, and month as effect. Analyses were done forfemales and males separately.
Between December 1995 and February 1996 samples of as many as possible putativespecies were taken monthly in the way described above. Twelve species in sample sizesof at least five fish of the same sex (above size at 50% maturity) were obtained in at least1 month, representative of the community (three Neochromis, two Mbipia, threePundamilia, one Lithochromis, two Paralabidochromis, one ‘ Haplochromis ’). In thiscase, gillnets were set at two water depth ranges: in inshore shallow water (1·5–2·5 mdepth) and in offshore water (4–6 m depth). It was attempted to collect at least 10 adultindividuals of each sex, species and depth range. In Neochromis rufocaudalis Seehausen& Bouton, Mbipia mbipi Seehausen, Lippitsch & Bouton, and ‘ Haplochromis ’ cyaneusSeehausen, Bouton & Zwennes the proportion of sexually active individuals wassignificantly higher in shallow water (1·5–2·5 m) than in deep water (4–6 m) (Fisher’sexact probability test: P<0·0001 in rufocaudalis and mbipi, P=0·02 in cyaneus). As <10%of the populations of these species live at 4–6 m (Seehausen & Bouton, 1997), and mostof these individuals were immature, the analysis of breeding seasonality was confined tosamples from depths of <4 m. In these cases, it was attempted to take more individualsfrom the dominant depth range to have similar overall sample sizes of different species.
Indices of similarity in timing of reproduction (ST) were calculated separately for the12- and 3-month studies using the similarity index of Schoener (1968) as
ST=1"0·5ÓPpi"qiP=Ómin(pi, qi)
where pi is the proportional contribution of month i to the sum of all monthlyproportions of stage 5 in a species, and qi is the proportional contribution of the samemonth to the sum of all monthly proportions of stage 5 in another species. Males wereused for the calculation of ST because of the larger sample sizes. Gonad maturation maybe less seasonal in males than in females, and an investigation of males mightunderestimate possible seasonal trends. However, since continuous availability of onesex in sexually active stage should be sufficient to break down any isolation effects ofasynchronous timing of reproduction in the other sex, the expectedly less seasonal sexshould be investigated to test whether differences in timing might contribute to isolationof putative species.
SPATIAL PATTERNS OF REPRODUCTIONTo test for spatial isolation of putative species, the distribution of male territories was
mapped in a study depth transect at the westward slope of Makobe Island (Seehausen &Bouton, 1997). Quadrats of 4#4 m were established on the rocky bottom, with a sinkingrope. A diver swam twice (with an interval of 15 min) two trajectories across eachquadrat with a distance of 2 m between trajectories. Territorial males within 1 m to eitherside of the diver were counted and their positions mapped with a pencil on a PVC sheet.

40 . .
Mapping of a quadrat was completed within 30 min. Usually four quadrats were placedadjacent to each other. On 7 days, 17 quadrats were mapped in 0·6–2 m depth, 11 in2–3·5 m depth, 12 in 3·5–5 m depth, and eight in 5–6·5 m depth.
Territorial densities were determined for each depth range. To compare spawning sitedistribution with foraging distribution, the distribution of males was determined for eachspecies by experimental gillnet fishing at water depths corresponding to each of the fourdepth ranges, between 100 and 300 m from the scuba study area. This could not be doneby scuba because the low water transparency in combination with the fast movementof nonterritorial fishes would have made this very difficult. Males were used because of10 times larger male than female sample sizes. The distribution of females in the nets,however, rules out that females occupy spatial niches that would not also be utilized bymales.
Assuming balanced sex ratios and similar proportions of sexually active individualsamong adults, indices of similarity in bathymetric distribution (SB) of reproductivelyactive parts of populations were calculated with Schoener’s (1968) formula as
SB=1"0·5ÓPpi"qiP=Ómin(pi, qi)
where pi is the proportion of a species’ breeding territories, located in the ith depth rangeand qi is the proportion of another species’ breeding territories, located in the same depthrange.
Though depth is within one habitat type probably the most important, it is not theonly spatial parameter by which breeding grounds can be isolated. To account forthe possibility of depth-independent spatial isolation, species-specific frequencies ofintra- and interspecific adjacency of territorial sites (T) were calculated as
where NTiis the number of males of species y, whose territories are adjacent to territories
of males of species x in the ith depth zone, Ni is the observed number of territorial malesof species y in the ith depth zone, and Nmi
"2 is the observed mean number of territorialmales per area size of species y in the ith depth zone.
COMBINED NON-BEHAVIOURAL ISOLATIONA combined estimate of pairwise similarity of reproductive activities in terms of time
and space (SR) was calculated as
SR=STSB
This was extended, by weighting SR with the abundances of two species, into an index
where NX is the relative abundance of the species under consideration, and NZ that of theeffect species, determined as the sums of their mean territorial densities in the four depthranges. From this, an estimate of the probability of encounter of sexually activeindividuals of the two species, relative to the probability of encounter of conspecifics, canbe calculated as
In the absence of direct behavioural mate choice PEn predicts the probability ofhybridization as the proportion of interspecific matings among all matings of species x.
One putative species, Neochromis omnicaeruleus Seehausen & Bouton, comprises threecolour morphs, named ‘ blue ’, ‘ orange blotched ’ (OB) and ‘ black and white ’ (BW)

41
(Seehausen, 1996). OB and BW morphs exist in both sexes but males are extremely rare(Table I). Females of the three morphs were approximately equally common. As asimilar analysis for these putatively conspecific colour morphs did not show anydifferences in depth distribution (Seehausen & Bouton, 1997), and no differences inreproductive seasonality, data of the morphs were pooled.
BEHAVIOURAL MATE CHOICETo test for behavioural isolation by active mate choice, courtship behaviour was
sampled by the focal-animal method (Altmann, 1974). Low water transparency made itdifficult to follow gravid females. Males were focused upon more easily because of theirterritoriality-related site fidelity and more conspicuous coloration. Their courtshipbehaviour consists of a sequence of gestures (lateral display, quiver, lead, circle; illustratedin Seehausen, 1996). Territorial males were observed predominantly and their rates ofbeing visited by, and their interaction with females of different putative species and colourmorphs were recorded. Forty territorial males of five species and five ripe females of twospecies were observed between December 1995 and February 1996. The species includedthe three numerically dominant species in the shallow waters, and Pundamilia nyererei(Witte-Maas & Witte) that is dominant at greater depth.
Each focal individual was observed, and its behaviour recorded, as long as possible butno longer than 10 min. Within this period each encounter with haplochromines of theopposite sex was recorded, noting the type (putative species or colour morph) of female(or male), and the behaviour displayed. An encounter was defined as cases in which afemale and male were within the territory of the male, and no further than 50 cm fromeach other. The cumulative focal times are given in Table I, recorded in about 12 diver-h.For each territorial male the water depth was measured on a dive computer.
The study concentrated on the shallow waters where the density of territories washighest. For analysis, the data were split into two depth ranges: <2 m and >2 m(maximum depth 5·8 m), and data from focal-male and focal-female observations wereanalysed separately. To test for pre-courtship female choice, the observed ratios offrequencies of conspecific over frequencies of heterospecific female visits to territorialfocal-males, and the ratios of frequencies of focal-female visits to conspecific overfrequencies of visits to heterospecific males were compared with ratios expected underthe assumption of random visiting behaviour. Expected ratios were derived separatelyfor the two depth ranges, from the relative abundance of the species in experimentalgillnet catches of females, made in the above mentioned nets, and from the density ofterritorial males determined by scuba as reported under spatial patterns. High mobilityof females and low water transparency made it impossible to estimate female abundancesby scuba.
OVERALL PROBABILITIES OF MATINGFinally, estimates of pairwise overall probabilities of mating between putative species
and colour morphs could be calculated. The abundance-weighted pairwise indices ofsimilarity in time and space were used, and the observed proportions of encounters inwhich courtship proceeded to lead-swimming to the spawning site (the highest frequentlyobserved level of courtship escalation), as
where PMax,z is an estimate for the overall probability that an individual of species xmates with an individual of species z.
RESULTS
TEMPORAL PATTERNS OF REPRODUCTIONSize at 50% maturity (rounded to the nearest mm) for 15 species is given
in Table I. The three numerically dominant shallow-water species showed

T
I.C
ichl
idsp
ecie
sre
cord
edat
Mak
obe
Isla
nd,
wit
hsu
rvey
info
rmat
ion
colle
cted
Spec
ies
Size
at50
%m
atur
ity
Dat
aon
gona
dst
ages
*(m
onth
:ob
s.st
age)
No.
offo
cal
wat
ches
(M/F
)ti
me
(min
)
Loc
atio
nof
mal
ete
rrit
orie
s*†
(dep
thin
m)
Not
es12
-mon
thsu
rvey
,M
/F(m
m)
3-m
onth
surv
ey,
M(m
m)
Neo
chro
mis
rufo
caud
alis
Seeh
ause
n&
Bou
ton
83/6
087
Fig
s1,
212
/1(3
5)F
ig.
3N
eoch
rom
isom
nica
erul
eus
Seeh
ause
n&
Bou
ton
87/6
386
Fig
s1,
2(F
)12
/4(7
2)B
lue
mor
phF
igs
1,2
(M)
Fig
.3
OB
mor
phF
eb.:
4,M
ay:
5,Ju
l.:5
(M)
1·5
(1–4
)V
ery
rare
WB
mor
phJa
n.:
4(M
)(2
–4)
Ver
yra
reN
eoch
rom
isgi
gas
Seeh
ause
n&
Lip
pits
ch10
3F
ig.
22/
–(1
7)F
ig.
3N
eoch
rom
is‘u
nicu
spid
scra
per
’?
Feb
.:4
(3M
)?
Rar
eN
eoch
rom
is‘l
arge
eye
nigr
ican
s’
?(<
2)V
ery
rare
Mbi
pia
mbi
piSe
ehau
sen,
Lip
pits
ch&
Bou
ton
83/6
388
Fig
s1,
210
/–(4
1)F
ig.
3M
bipi
alu
tea
Seeh
ause
n&
Bou
ton
>12
3F
ig.
2(F
)F
eb.:
2(M
)<
1‡R
are
Pun
dam
ilia
nyer
erei
(Wit
te-M
ass
&W
itte
)79
Fig
.2
4/–
(14)
Fig
.3
Pun
dam
ilia
pund
amili
aSe
ehau
sen
&B
outo
n82
Fig
.2
1–3·
5‡C
rypt
icP
unda
mili
a‘p
ink
anal
’77
Fig
.2
Fig
.3
Lit
hoch
rom
is‘y
ello
wch
in’
77F
ig.
2F
ig.
3P
aral
abid
ochr
omis
chilo
tes
(Bou
leng
er19
11)
130
Fig
.2
(F)
Dec
.:4
(M)
nt?
(1–6
·5)
At
othe
ris
land
sno
n-te
rrit
oria
lP
aral
abid
ochr
omis
‘roc
kkri
bens
is’
87F
ig.
2(M
)Ja
n.:
5(2
F)
Fig
.3
Par
alab
idoc
hrom
is‘s
hort
snou
tsc
rape
r’
>93
?nt
?(1
–5)
Rar
e‘H
aplo
chro
mis
’cy
aneu
sSe
ehau
sen,
Bou
ton
&Z
wen
nes
90F
ig.
21·
5–3·
5‡O
nly
very
larg
em
ales
terr
itor
ial
‘Hap
loch
rom
is’
flavu
sSe
ehau
sen,
Zw
enne
s&
Lip
pits
ch>
93Ja
n.:
3,4
(M)
Feb
.:3
(M)
1·5–
3·5‡
Onl
yve
ryla
rge
mal
este
rrit
oria
lL
ipoc
hrom
ism
elan
opte
rus
(Tre
wav
as19
28)
107
(<2)
Hap
loch
rom
is‘b
lue
obliq
uide
ns’
?F
eb.:
5,6
(F)
<1
Hap
loch
rom
is‘p
urpl
eye
llow
’?
<1‡
Fem
ale
size
sat
50%
mat
urit
yw
ere
not
dete
rmin
edin
the
3-m
onth
sst
udy
beca
use
ofpa
rtic
ular
lysm
all
sam
ple
size
s.M
/F=
mal
es/f
emal
es.
*Ava
ilabl
eda
taon
rare
spec
ies
are
give
nhe
re,r
efer
ence
tofig
ures
ism
ade
for
the
quan
tita
tive
lyst
udie
dsp
ecie
s.†N
etca
tch
dete
rmin
edde
pth
rang
esof
mat
ure
fishe
sw
here
terr
itor
ial
mal
esha
veno
tor
rare
lybe
enlo
cate
dun
der
wat
erar
esh
own
inpa
rent
hese
s(S
eeha
usen
etal
.,19
98).
nt,
Mal
esno
tte
rrit
oria
l;‡n
ote
rrit
oria
lm
ales
loca
ted
inth
isst
udy
but
onot
her
occa
sion
s(S
eeha
usen
etal
.,19
98).

43
100
Mbi
pia
mbi
pi (
%)
Jul.
1990
0
20
40
60
80
10
Au
g. 1
990
11
Sep
t. 1
990
11
Oct
. 199
0
11
Nov
. 199
0
8
Dec
. 199
0
11 9
Feb
. 199
1
10
Mar
. 199
1
4
Apr
. 199
1
11
May
199
1
9
Jun
. 199
1
7100
Jul.
1990
0
20
40
60
80
5
Au
g. 1
990
5S
ept.
199
07
Oct
. 199
013
Nov
. 199
0
6
Dec
. 199
0
5 5
Feb
. 199
1
10
Mar
. 199
1
11
Apr
. 199
1
12
May
199
1
6
Jun
. 199
1
6
100
N. o
mn
icae
rule
us
(%)
0
20
40
60
80
10 10 6 8 11 10 13 13 11 13 12 5100
0
20
40
60
80
10 8 7 5 9 5 4 14 4 (7) (2) 9
100
N. r
ufo
cau
dali
s (%
)
0
20
40
60
80
16 11 11 11 12 10 4 11 10 11 7 11100
0
20
40
60
80
5 7 9 2 5 (1) 4 8 2 (1)
(a) (b)
Jul.
1991
Jul.
1991
2
F. 1. Developmental stages of gonads of the three, in shallow-water most abundant putative speciesfrom Makobe Island over a 12-month study period: (a) males, (b) females. On four occasions asingle female was available (N. rufocaudalis Dec. 1990, stage 3; May 1991, stage 4; N. omnicaeruleusMay 1991, stage 4; July 1991, stage 3). ., Stage 2; , stage 3; , stage 4; /, stage 5; , spent.Sample sizes are given above each month bar.
year-round reproductive activity (Fig. 1). There was no evidence for seasonalityamong females, where all runs tests for mean monthly gonad stage, monthlyproportions of stage 5 fishes, and of stage 4+5+6 fishes pooled yielded P valuesabove 0·1. Patterns in males were suggestive of slight seasonality. There was inN. rufocaudalis significant seasonality in the proportion of stage five males(P<0·05). A similar trend could be detected in N. omnicaeruleus (P<0·1). Aftercorrection of P values by sequential Bonferroni method (Rice, 1989), only theN. rufocaudalis result remained significant.
Month had no significant effect on gonad stage in adult females (all ANOVAsP>0·2). Month had significant effect on gonad stage in adult males of allthree species (ANOVAs, N. rufocaudalis: F=2·74, 11, d.f.=125, P=0·003;

44 . .
T II. Indices of similarity of reproductive activities in time and space
Ruf Omn Mbi Pun Nye Pin Yel Rkr
3-month studyN. gigas (Gig), S 0·87 0·82 0·75 0·86 0·68 0·69 0·63 0·75 ST
1·00 0·49 0·64 — 0·00 0·00 0·00 0·50 SB0·87 0·40 0·48 0·00 0·00 0·00 0·38 SR
N. rufocaudalis (Ruf), S 0·73 0·84 0·83 0·71 0·80 0·71 0·75 ST0·49 0·64 — 0·00 0·00 0·00 0·50 SB0·36 0·54 0·00 0·00 0·00 0·38 SR
N. omnicaeruleus (Omn) 0·71 0·70 0·58 0·67 0·88 0·66 ST0·68 — 0·51 0·21 0·21 0·87 SB0·48 0·30 0·14 0·18 0·57 SR
M. mbipi (Mbi), S 0·93 0·87 0·96 0·59 0·92 ST— 0·12 0·00 0·00 0·75 SB
0·10 0·00 0·00 0·69 SRPu. pundamilia (Pun) 0·80 0·89 0·66 0·88 ST
— — — — SBSR
Pu. nyererei (Nye), D 0·88 0·45 0·91 ST0·36 0·36 0·37 SB0·32 0·16 0·34 SR
Pu. ‘ pink anal ’ (Pin), D 0·55 0·93 ST1·00 0·25 SB0·55 0·23 SR
L. ‘ yellow chin ’ (Yel), D 0·54 ST0·25 SB0·14 SR
ST12-month study
N. rufocaudalis/N. omnicaeruleus 0·76N. rufocaudalis/M. mbipi 0·90N. omnicaeruleus/M. mbipi 0·77
ST, Similarity in timing; SB, similarity in spawning site distribution; SR, combined similarity inreproductive activities in time and space (see Materials and Methods); Rkr, Pa. ‘ rockkribensis ’; S,shallow breeding; D, deep breeding (see text).
N. omnicaeruleus: F=1·92, 11, d.f.=114, P=0·044; M. mbipi: F=2·32, 11,d.f.=108, P=0·013), adding evidence for some temporal fluctuations of theproportion of individuals that are sexually active.
Similarity indices (ST), based on the 12-month study are given at the bottom ofTable II. In the three sampling months in 1995/1996 there was a largeinterspecific temporal overlap too (Table II). Provided five or more individualslarger than size-at-50%-maturity were caught, ripe ones were among them in allputative species at any month, except in one case (Fig. 2). Several species werecaught in smaller numbers (Table I). Temporal fluctuations in reproductiveactivity were somewhat more apparent in deep-living species than in shallow-living species (Fig. 2), and ST indices among some deep-living species were lowerthan those among shallow-living species (mean&.. 0·63&0·23 v. 0·82&0·06;Table II).

45
100
Feb
ruar
y
0
20
40
60
80
8
N
17
N
1
N
10
M
6
P
33
P
32
P
6
L
1
Pa
4
Pa
14
N
8
N
12
N
13
M
7
M
3
P
20
P
11
P
2
L
100
Jan
uar
y
20
40
60
80
13 22 2 13 7 23 20 15 5 17 9 8 9 5 16 5 5 4
100
Dec
embe
r
20
40
60
80
8 19 11 10 11 17 16 4 7 13 2 17 3 13 7 7 6 3 7
(a) (b)
N N M P P L Pa Pa
N N N M P P P Pa Pa
F. 2. Developmental stages of gonads of 12 putative species from Makobe Island that could be collectedin at least 1 month in five or more individuals of the same sex. (a) Males, (b) females. N,Neochromis (from left: N. rufocaudalis, N. omnicaeruleus, N. gigas); M, Mbipia (from left: M. mbipi,M. lutea); P, Pundamilia (from left: P. pundamilia, P. nyererei, P. ‘ pink anal ’); L, Lithochromis‘ yellow chin ’; Pa, Paralabidochromis (from left in males: ‘ H ’. cyaneus, Pa. ‘ rockkribensis ’; infemales: ‘ H ’. cyaneus, Pa. chilotes). Shading of bars as in Fig. 1. Sample sizes are given aboveeach species bar.
SPATIAL PATTERNS OF REPRODUCTIONThere was a higher density of territorial males in shallow (5–11, 7·4&1·7 per
16 m2 at <2 m) than in deep water (1–8·5, 3·9&2·5 per 16 m2 at 5–6·5 m depth;Fig. 3).

0.5–
2
Depth range (m) 2–3.
5
3.5–
5
5–6.
5 00
01
02
12
30
23
10
00
11
23
00
Nu
mbe
r of
mal
e te
rrit
orie
s 16
m–2
(b)
0.5–
2
2–3.
5
3.5–
5
5–6.
5 00
020
6040
8020
100
100
200
120
020
200 N
um
ber
of m
ales
30-
m n
et–1
(a) 'H
a.' '
blu
e ob
liqu
iden
s'N
e. g
igas
Ne.
ru
foca
ud
alis
060
4080
120
6040
800
4010
060
4080
120
020
Mbi
pia
mbi
piN
e. o
mn
icae
rule
us
Pu
.'p
ink
anal
'
Pa.
'roc
kkri
ben
sis'
Pu
. nye
rere
iL
i.'y
ello
w c
hin
'
F
.3.
Ove
rall
bath
ymet
ric
popu
lati
onan
dsp
awni
ngsi
tedi
stri
buti
onof
nine
spec
ies,
terr
itor
ies
ofw
hich
have
been
obse
rved
.(a
)Ove
rall
popu
lati
ondi
stri
buti
onba
sed
onm
ean
net
catc
hes
done
in19
95/1
996.
(b)
Spaw
ning
site
dist
ribu
tion
base
don
scub
am
appi
ngof
terr
itor
ial
mal
es(m
ean
num
ber
ofte
rrit
orie
spe
rar
ea).

47
The study failed to locate territorial males of three out of the 12 putativespecies in the temporal analysis of reproduction, and of one of the three putativecolour morphs (Table I). In all species, the breeding depth range was within thespecies’ foraging depth range (Seehausen & Bouton, 1997). In many species,both from shallow (N. rufocaudalis, M. mbipi) and deep water (Pu. ‘ pink anal ’,Pu. nyererei, L. ‘ yellow chin ’), the breeding range was narrower than theforaging range (Fig. 3, see also Fig. 2 in Seehausen & Bouton, 1997). Severalspecies’ spawning grounds were isolated by depth, even though their foraginggrounds overlapped. In many others the spawning grounds were not isolated bydepth either (Fig. 3, Table II). Complete isolation of spawning grounds by depthoccurred between the three shallow-breeding species H. ‘ blue obliquidens ’, N.gigas and N. rufocaudalis on the one hand, and the three deep-breeding speciesPu. nyererei, Pu. ‘ pink anal ’ and L. ‘ yellow chin ’ on the other hand, andbetween M. mbipi, and two deep-breeding species, Pu. ‘ pink anal ’ and L.‘ yellow chin ’.
Lack of interspecific territorial adjacency occurred in five more species pairs(bold print in Table III). Where both species are rare, this was most likely dueto small sample sizes (compare Fig. 3), but the lack of adjacency of Pu. nyerereiand L. ‘ yellow chin ’ territories, both being rather abundant at 5–6·5 m depth,may reflect some difference in microhabitat choice.
T III. Frequencies of territorial adjacencies, T
Bob Gig Ruf Omn Mbi Nye Pin Yel Rkr
H. ‘ blue obliquidens ’ (Bob) 0·00 0·00 1·00 0·00 1·00 0·00 0·00 0·00 0·00N. gigas (Gig) 0·00 0·00 1·00 1·00 1·00 0·00 0·00 0·00 0·00N. rufocaudalis (Ruf) 0·03 0·13 0·53 0·83 0·70 0·00 0·00 0·00 0·07N. omnicaeruleus (Omn) 0·00 0·05 0·34 0·75 0·25 0·25 0·18 0·03 0·07M. mbipi (Mbi) 0·16 0·14 0·26 0·36 0·89 0·00 0·00 0·00 0·08Pu. nyererei (Nye) 0·00 0·00 0·00 0·43 0·00 0·74 0·12 0·00 0·00Pu. ‘ pink anal ’ (Pin) 0·00 0·00 0·00 0·89 0·00 0·44 0·56 0·22 0·11L. ‘ yellow chin ’ (Yel) 0·00 0·00 0·00 1·00 0·00 0·00 1·00 0·00 1·00Pa. ‘ rockkribensis ’ (Rkr) 0·00 0·00 0·33 0·67 0·67 0·00 0·33 0·33 0·00
To be read row-wise. Cases of no adjacency, despite bathymetrically overlapping spawning grounds areshown in bold.
COMBINED NON-BEHAVIOURAL ISOLATIONIndices of combined similarity of reproductive activities in time and space are
given in Table II. Indices among deep-breeding species (0·34&0·20, n=3;marked with D in Table II) and between deep- and shallow-breeding species(0·01&0·03, n=9; marked with S in Table II) are smaller than those betweenshallow-breeding species (0·68&0·28, n=3), implying that shallow-breedingspecies are not only more similar to each other than to deep-breeding species, butalso more similar than are deep-breeding species to each other.
The probabilities with which sexually active individuals encounter sexuallyactive heterospecifics, rather than conspecifics, are given in Table IV. In theabsence of active mate choice, interspecific hybridization would account for

48 . .
>10% of the matings in 24 of 56 pair-wise species comparisons, and wouldaccount for >5% of the matings in an additional 9 pairwise comparisons.
T IV. Pairwise probabilities of interspecific encounter during reproduction, PEn
Gig Ruf Omn Mbi Nye Pin Yel Roc
N. gigas (Gig) 0·92 0·94 0·92 0·00 0·00 0·00 0·43N. rufocaudalis (Ruf) 0·07 0·52 0·51 0·00 0·00 0·00 0·06N. omnicaeruleus (Omn) 0·01 0·11 0·24 0·15 0·03 0·01 0·03M. mbipi (Mbi) 0·02 0·22 0·43 0·08 0·00 0·00 0·06Pu. nyererei (Nye) 0·00 0·00 0·34 0·10 0·10 0·01 0·03Pu. ‘ pink anal ’ (Pin) 0·00 0·00 0·43 0·00 0·49 0·09 0·07L. ‘ yellow chin ’ (Yel) 0·00 0·00 0·84 0·00 0·73 0·75 0·18Pa. ‘ rockkribensis ’ (Rkr) 0·16 0·70 0·91 0·89 0·78 0·44 0·08
To be read row-wise. Values are species pairwise probabilities (for species in rows) of encounter witha hetero-, instead of a conspecific sexually active individual. The probability of encounter with aconspecific is 1"(probability of encounter with a heterospecific). Values are species specific becauserelative abundance in species pairs is asymmetric.
BEHAVIOURAL MATE CHOICETo calculate expected female visitation rates, a total of 292 females was
obtained in the shallow depth range on four separate occasions, and a total of224 females in the deeper range on seven occasions. The observed conspecificover heterospecific visitation ratios were compared with expected ratios byFisher’s exact probability test. The observed rates of courtship engagementupon encounter of a territorial male by conspecific females were compared withthose upon encounter with heterospecific females as ratios of courtshipencounters/all encounters (Fisher’s exact probability test) on several levels ofinclusiveness: pooled over species, pooled per species, and for pairs of species.Data from focal-female and focal-male observations were used together. Thesame was done for three putative colour morphs of one species.
In some species, species recognition operates prior to the onset of courtship.Observed ratios of conspecific v. heterospecific female visits to Neochromis malesdiffered from those expected under random visiting behaviour. In contrast, theproportions of conspecific visitors to M. mbipi and Pundamilia nyererei maleswere not different from those expected under random visiting. Neochromismales patrol their territories and display openly in well-illuminated shallowwater, while territorial Mbipia and Pundamilia males are more cryptic in theirbehaviour, often leave the crevices between rocks only for a quick display(Mbipia), or live in less well-illuminated depth (Pundamilia).
On the highest level of inclusiveness, the pooled data showed that courtshipamong members of the same putative species is much more frequent than amongmembers of different putative species (Table V), and that the degree ofassortativeness increases with each step in the courtship sequence. The elementslead and circle that occur late in the sequence, were never observed betweenindividuals of different putative species. This suggests that species recognitionaccuracy improves in the course of the courtship interaction. On the other
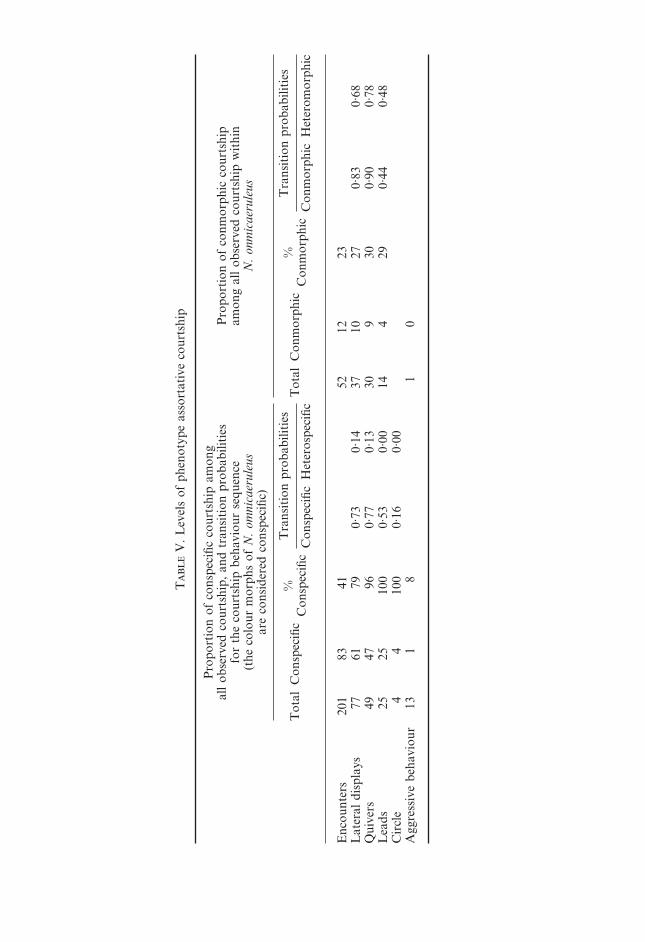
T
V.
Lev
els
ofph
enot
ype
asso
rtat
ive
cour
tshi
p
Pro
port
ion
ofco
nspe
cific
cour
tshi
pam
ong
all
obse
rved
cour
tshi
p,an
dtr
ansi
tion
prob
abili
ties
for
the
cour
tshi
pbe
havi
our
sequ
ence
(the
colo
urm
orph
sof
N.
omni
caer
uleu
sar
eco
nsid
ered
cons
peci
fic)
Pro
port
ion
ofco
nmor
phic
cour
tshi
pam
ong
all
obse
rved
cour
tshi
pw
ithi
nN
.on
mic
aeru
leus
Tot
alC
onsp
ecifi
c%
Con
spec
ific
Tra
nsit
ion
prob
abili
ties
Tot
alC
onm
orph
ic%
Con
mor
phic
Tra
nsit
ion
prob
abili
ties
Con
spec
ific
Het
eros
peci
ficC
onm
orph
icH
eter
omor
phic
Enc
ount
ers
201
8341
5212
23L
ater
aldi
spla
ys77
6179
0·73
0·14
3710
270·
830·
68Q
uive
rs49
4796
0·77
0·13
309
300·
900·
78L
eads
2525
100
0·53
0·00
144
290·
440·
48C
ircl
e4
410
00·
160·
00A
ggre
ssiv
ebe
havi
our
131
81
0

50 . .
T VI. Comparisons of the likelihood of courtship between and within putativespecies: female visiting behaviour
Observed Expected P value*
Visiting female (n visits)Neochromis rufocaudalis (5) Consp./het. 3/2 1/4 0·20Neochromis omnicaeruleus (27) Consp./het. 19/8 10/17 0·01
Visited male (n visits)Neochromis rufocaudalis (66) Consp./het. 10/56 4/62 0·09Neochromis omnicaeruleus (60) Consp./het. 36/24 6/54 <0·001Mbipia mbipi (26) Consp./het. 15/11 18/8 0·45Pundamilia nyererei (11) Consp./het. 2/9 1/10 0·53
*Fisher’s exact probability test.Observations on visiting females were all done in 1–2 m depth. Observations on visited males were done
in 1–6 m depth, where expected ratios were calculated for observations in the depth ranges 1–2 and 2–6 mseparately and subsequently pooled.
T VII. Comparisons of the likelihood of courtship between and within putativespecies and colour morphs: courtship engagement
Intraspecific Interspecific P value*
Pooled over speciesLd/En 66/90 15/115 <0·001Qu/En 47/85† 3/115 <0·0001
Ld-Qu transition probability 47/61† 3/15 0·03
Male Female Male FemalePooled per species
Neochromis rufocaudalis Ld/En 9/15 3/62 0/6 <0·0001 0·07Neochromis omnicaeruleus Ld/En 37/55 3/26 5/53 <0·001 <0·0001Mbipia mbipi Ld/En 17/18 1/14 5/38 <0·001 <0·0001Pundamilia nyererei Ld/En 2/2 3/9 2/4 0·35 —
Qu/En 2/2 1/9 0/4 0·10 —
Intramorphic IntermorphicPooled over morphs
Ld/En 10/13 27/42 0·71Qu/En 9/13 21/42 0·52
Proportions of intra- v. interspecific encounters (En) with courtship (Ld, lateral display; Qu, quiver).*Fisher’s exact probability test.†In five cases, courtship could not be recorded after Ld.
hand, antagonistic intersexual behaviours were observed predominantly amongindividuals of different putative species (Fisher’s exact test for the ratiosall/conspecific encounters v. all/conspecific antagonistic interactions P=0·07).
The analysis on species and interspecific levels for the four numericallydominant species Neochromis rufocaudalis, Neochromis omnicaeruleus, Mbipiambipi, and Pundamilia nyererei revealed a similar pattern (Tables VI and VII).

51
The rates of courtship engagement upon encounter were significantly higherin encounters between conspecifics than in encounters between heterospecifics inN. rufocaudalis, N. omnicaeruleus, and M. mbipi (Table VII). This assortative-ness in courtship occurred in all species–sex combinations (data not shown),indicating that direct mate choice contributes substantially to reproductiveisolation among these Neochromis and Mbipia species. Compared to theNeochromis and Mbipia species, N. omnicaeruleus and Pu. nyererei were poorlyisolated on the lateral display level of the behavioural sequence. Assortativenessincreased on the next level (quiver), but sample sizes were too small to drawconclusions (Table VII; see below).
Visit rates of differently coloured females of the putatively colour-polymorphicN. omnicaeruleus, and courtship engagement probabilities of blue males withthese females did not deviate from expectations under random mating. Bluemales were visited by brown and blotched (OB, BW) females and all threeelicited male courtship. However, no instance of courtship between an OB maleand brown, or BW females was observed.
The sensitivity of the results to sample size was examined by subsampling N.omnicaeruleus, the species with the largest number of observed encounters. Itwas subsampled by units of 10 encounters and then 10-encounter subsampleswere pooled randomly to larger subsamples until a P<0·1 was obtained in 95%of the comparisons of conspecific and heterospecific courtship rates. This wasreached at a subsample size of 40 encounters, suggesting that the results inTable VII are reliable except those for Pu. nyererei.
OVERALL PROBABILITIES OF MATINGDue to assortative mate choice, overall probabilities of mating (PMa) are zero
among the four putative species, indicating that they are reproductively isolated.Probabilities of mating are possibly reduced in some pairwise combinations ofthe three putative colour morphs, but in other combinations are as high as thosebetween individuals of the same colour morph, making it very likely that theseare not fully reproductively isolated.
DISCUSSION
Our data make it very likely that morphotypes of Lake Victoria cichlids thatdiffer in coloration, anatomy and ecology, and were putatively designated speciesstatus by field ecologists, are reproductively isolated species. The data showfurther that isolation is incomplete or lacking between colour morphs that differonly in coloration, and were putatively considered conspecific.
The major factor accounting for reproductive isolation of species is assortativemate choice, which makes up for between 49 and 94% of isolation amongmembers of the same genus (Table IV). Reproductive parapatry, e.g. isolationby depth, was found exclusively between species that belong to different genera,such as Neochromis and Pundamilia (Table II). By contrast, spawning groundsof closely related species generally overlapped broadly, with the smallestsimilarity index in spatial distribution of territories 0·36 between two species ofPundamilia in deep water. Reproductive allochrony was even less developed.

52 . .
Members of the same genus overlapped with between 73 and 89% of theirsexual activity, as measured in males, and sampling through one year yieldedsimilarity indices very similar to those obtained in a 3-month study (Table II).The largest timing difference was found between two species of different genera(Pundamilia and Lithochromis) in deep water, the activity of which stilloverlapped to 45%.
Though it appeared that partial reproductive allochrony and partial reproduc-tive parapatry are slightly more developed at greater depth than in the shallows(compare Fig. 3 with Table II), the interspecific overlaps that we measured intiming and location of reproduction are of such magnitude, that in the absenceof assortative mate choice and postzygotic isolation, probably none of thespecies currently found at Makobe Island, could be maintained (Table IV). Thespecies with the lowest maximum hybridization probabilities would be N.omnicaeruleus and Pu. nyererei with probabilities of still 0·24 and 0·34.Postzygotic barriers to gene flow have not been found in haplochromine cichlidsof Lake Victoria (Crapon de Caprona & Fritzsch, 1984; Crapon de Caprona,1986; Seehausen et al., 1997a). Removing altogether the behavioural componentin mate choice would probably lead to introgression, a prediction supported byfield observations and laboratory experiments (Seehausen et al., 1997a). Ourdata on non-behavioural prezygotic isolation (Tables II–IV) indicate that ashallow-water and a deep-water population might be maintained at MakobeIsland by bathymetric isolation, and possibly some difference in timing ofreproduction which may reflect asynchronous fluctuations in abundance ofepibios and plankton, the principal foods in deep and shallow water speciesrespectively (Bouton et al., 1997).
The number of species that breed in full sympatry at Makobe Island decreaseswith increasing water depth (Seehausen et al., 1997a). This is shown only partlyin Fig. 3. The following numbers of additional species breed at the differentdepth ranges: 10 in <2 m depth, five between 2 and 3·5 m, two between 3·5 and5 m, and one in 5 to 6·5 m (Table I). An explanation for the negativerelationship between the number of syntopically reproducing species and waterdepth that is in concordance with results from this study, is that constraints onbehavioural mate recognition in the deeper and less well-illuminated habitat limitthe number of species that can be isolated reproductively there. Coloration is animportant cue in interspecific mate choice (Seehausen & van Alphen, 1998) butassessment of coloration depends on ambient light. Intensity and spectral widthof ambient light decrease with increasing water depth, reducing the potential forvariation in intra- and interspecific signal structure that can be perceived by thesignal receptor. The shifting of spawning grounds, relative to feeding grounds,towards shallower water, observed in males of several species (Fig. 3), could haveevolved in response to the intraspecific component of this constraint. Relatedobservations have been made on haplochromine cichlids in very differenthabitats: cichlids from poorly illuminated sublittoral regions of Lake Victoriareproduce at the time when water transparency is maximum (Witte et al., 1998).Behaviour evolved to utilize ambient light such that it enhances the effects ofcolourful courtship display has also been found in birds that use light spotsin forests as courtship arenas (Thery & Vehrencamp, 1995; Endler & Thery,1996).

53
What do the results say about likely ways of speciation? It is becomingincreasingly apparent that speciation is often driven by the evolution of traitsthat, by causing prezygotic isolation, break down gene flow between incipientpopulations, rather than by stochastic genetical divergence (Tregenza & Bridle,1997). Like other rapidly evolving animals (Butlin & Tregenza, 1998; Coyne,1998), haplochromine cichlid species are isolated prezygotically, and evidence forpostzygotic barriers is absent. A neglected field in speciation research is theidentification of those prezygotic traits that break down gene flow (traits ofspeciation). That the most closely related species of haplochromine cichlids areisolated primarily by direct behavioural mate choice is at odds with what hasbeen found in other recently and sympatrically radiated fish groups, and maysuggest that speciation mechanisms differ. While ecological speciation has beeninferred as the dominant mode among fish in postglacial water bodies (Smith &Todd, 1984; Schluter, 1996) and in some tropical lakes (Nagelkerke & Sibbing,1996), our data are in agreement with predictions from models of speciation bysexual selection (Fisher, 1930; Lande, 1981; Turner & Burrows, 1995; van Doornet al., 1998), and support the hypothesis that mate preference can be a trait ofspeciation in haplochromine cichlids (Dominey, 1984; Goldschmidt & de Visser,1990; McKaye, 1991; Turner, 1994; Deutsch, 1997; Seehausen et al., 1997a;Seehausen & van Alphen, 1998).
Temporal and spatial isolation of haplochromine cichlids at rocky shores ofLake Victoria may have evolved often after speciation, possibly as consequencesof adaptation. However, that bathymetric isolation alone can account forreproductive isolation between some of the species of Makobe Island, suggestspotential contributions of ecological speciation (e.g. Schluter, 1993, 1996) to theorigin of the haplochromine species diversity at rocky shores. Judging from ourdata, the most probable ecological speciation event at rocky shores appears to bea split into a shallow-dwelling benthos scraping and a deep-dwelling planktonsucking population, the kind of differentiation that would be facilitated by theversatile trophic anatomy of these cichlids (Galis & Drucker, 1996). Specieswhich are bathymetrically isolated and ecologically different (Bouton et al.,1997) belong to different genera, while there are many closely related speciesamong those that are only isolated behaviourally. This suggests, that the relativeimportance of different speciation mechanisms may have been different atdifferent phases in the radiation of rock-dwelling cichlids. Ecological speciation,initiated by strong divergent natural selection, is likely to be most importantwhere new niches with new functional demands are vacant. Speciation by matechoice, initiated by sexual selection, may be relatively more important whereniche space is being filled up, without the evolution of major functional andecological innovation.
We thank the Tanzanian Government and P. O. J. Bwathondi for research permits; theMwanza center of TAFIRI (E. F. B. Katunzi) for hospitality and support; M. Kayeba,R. Enoka, A. Samwel Terry, M. Haluna for their great assistance in the field; L.Schadhauser, A. Samwel Terry, H. Nielsen, R. Kiteri, H. G. Mbilinyi, C. Schaefer, G. DeLeeuw, R. Stawikowski, Mr Mapunda for logistic support; J. Endler, M. E. Knight andG. Turner for constructive advice on the manuscript. Support with diving equipmentfrom SCUBAPRO and SEAWAY, and financial support by the Dutch Foundation forTropical Research (WOTRO), the German Cichlid Association (DCG), TETRA andEHEIM is greatly acknowledged.

54 . .
References
Altmann, J. (1974). Observational study of behaviour: sampling methods. Behaviour 49,227–267.
Beadle, L. C. (1981). The Inland Waters of Tropical Africa, 2nd edn. London: Longman.Bouton, N., Seehausen, O. & van Alphen, J. J. M. (1997). Resource partitioning among
rock-dwelling haplochromines (Pisces: Cichlidae) from Lake Victoria. Ecology ofFreshwater Fish 6, 225–240.
Butlin, R. K. & Tregenza, T. (1998). Levels of genetic polymorphism: marker loci versusquantitative traits. Philosophical Transactions of the Royal Society of London B353, 187–198.
Coyne, J. A. (1992). Genetics and speciation. Nature 355, 511–515.Coyne, J. A. (1998). The evolutionary genetics of speciation. Philosophical Transactions
of the Royal Society of London B 353, 287–305.Crapon de Caprona, M. D. (1986). The use of fertile hybrids for the study of the
accuracy of species recognition in cichlids. Annales Musee Royal de l’AfriqueCentrale, Sciences Zoologiques 251, 117–120.
Crapon de Caprona, M. D. & Fritzsch, B. (1984). Interspecific fertile hybrids forhaplochromine Cichlidae (Teleostei) and their possible importance for speciation.Netherlands Journal of Zoology 34, 503–538.
Deutsch, J. C. (1997). Colour diversification in Malawi cichlids: evidence for adaptation,reinforcement or sexual selection? Biological Journal of the Linnean Society 62,1–14.
Dominey, W. (1984). Effects of sexual selection and life history on speciation: speciesflocks in African cichlids and Hawaiian Drosophila. In Evolution of Fish SpeciesFlocks (Echelle, A. A. & Kornfield, I., eds), pp. 231–249. Orono: University ofMaine Press.
van Doorn, G. S., Noest, A. J. & Hogeweg, P. (1998). Sympatric speciation andextinction by environment dependent sexual selection. Proceedings of the RoyalSociety of London B, in press.
Endler, J. A. & Thery, M. (1996). Interacting effects of lek placement, display behavior,ambient light, and color patterns in three neotropical forest-dwelling birds.American Naturalist 148, 421–452.
Fisher, R. A. (1930). The Genetic Theory of Natural Selection. Oxford: Clarendon.Futuyma, D. J. (1986). Evolutionary Biology, 2nd edn. Sunderland, MA: Sinauer.Galis, F. & Drucker, E. G. (1996). Pharyngeal biting mechanics in centrarchids and
cichlids: Insights into a key evolutionary innovation. Journal of EvolutionaryBiology 9, 641–670.
Goldschmidt, T. & de Visser, J. (1990). On the possible role of egg mimics in speciation.Acta Biotheoretica 38, 125–134.
Greenwood, P. H. (1981). Species flocks and explosive evolution. In Chance, Change andChallenge—The Evolving Biosphere (Greenwood, P. H. & Forey, P. L., eds),pp. 61–74. Cambridge: Cambridge University Press.
Holzberg, S. (1978). A field and laboratory study of the behaviour and ecology ofPseudotropheus zebra (Boulenger), an endemic cichlid of Lake Malawi (Pisces;Cichlidae). Zeitschrift fur Zoologie und Systematische Evolutionsforschung 16,171–187.
Johnson, T. C., Scholz, C. A., Talbot, M. R., Kelts, K., Ricketts, R. D., Ngobi, G.,Beuning, K., Ssemmanda, I. & McGill, J. W. (1996). Late Pleistocene desiccationof Lake Victoria and rapid evolution of cichlid fishes. Science 273, 1091–1093.
Lande, R. (1981). Models of speciation by sexual selection on polygenic traits.Proceedings of the National Academy of Science of the USA 78, 3721–3725.
Liem, K. F. & Kaufman, L. S. (1984). Intraspecific macroevolution: Functional biologyof the polymorphic cichlid species Cichlasoma minckleyi. In Evolution of FishSpecies Flocks (Echelle, A. A. & Kornfield, I., eds), pp. 203–215. Orono:University of Maine Press.

55
McKaye, K. R. (1991). Sexual selection and the evolution of the cichlid fishes of LakeMalawi, Africa. In Cichlid Fishes. Behaviour, Ecology and Evolution (Keenleyside,M. H. A., ed.), pp. 241–257. London: Chapman & Hall.
Marsh, A. C., Ribbink, A. J. & Marsh, B. A. (1981). Sibling species complexesin sympatric populations of Petrotilapia Trewaves (Cichlidae, Lake Malawi).Zoological Journal of the Linnean Society 71, 253–264.
Meyer, A. (1987). Phenotypic plasticity and heterochrony in Cichlasoma managuense(Pisces, Cichlidae) and their implications for speciation in cichlid fishes. Evolution41, 1357–1369.
Meyer, A., Kocher, T. D., Basasibwaki, P. & Wilson, A. C. (1990). Monophyletic originof Lake Victoria cichlid fishes suggested by mitochondrial DNA sequences.Nature 347, 550–553.
Nagelkerke, L. A. J. & Sibbing, F. A. (1996). Reproductive segregation among the largebarbs (Barbus intermedius complex) of Lake Tana, Ethiopia. An example ofintralacustrine speciation? Journal of Fish Biology 49, 1244–1266.
Rice, W. R. (1989). Analysing tables of statistical tests. Evolution 43, 223–225.Schluter, D. (1993). Adaptive radiation in stickebacks: size, shape, and habitat use
efficiency. Ecology 74, 699–709.Schluter, D. (1996). Ecological speciation in postglacial fishes. Philosophical Trans-
actions of the Royal Society of London B 351, 807–814.Schoener, T. W. (1968). The Anolis lizards of Bimini: resource partitioning in a complex
fauna. Ecology 49, 704–726.Seehausen, O. (1996). Lake Victoria Rock Chichlids—Taxonomy, Ecology, and
Distribution. Zevenhuizen, Netherlands: Verduijn Cichlids.Seehausen, O. & van Alphen, J. J. M. (1998). The effect of male coloration on female
mate choice in closely related Lake Victoria cichlids (Haplochromis nyerereicomplex). Behavioural Ecology and Sociobiology 42, 1–8.
Seehausen, O. & Bouton, N. (1997). Microdistribution and fluctuations in niche overlapin a rocky shore cichlid community in Lake Victoria. Ecology of Freshwater Fish6, 161–173.
Seehausen, O., van Alphen, J. J. M. & Witte, F. (1997a). Cichlid fish diversity threatenedby eutrophication that curbs sexual selection. Science 277, 1808–1811.
Seehausen, O., Lippitsch, E., Bouton, N. & Zwennes, H. (1998). Mbipi, the rock-dwelling cichlids of Lake Victoria: description of three new genera and fifteen newspecies (Teleostei). Ichthyological Exploration of Freshwaters 9, 1–100.
Seehausen, O., Witte, F., Katunzi, E. F., Smits, J. & Bouton, N. (1997b). Patterns of theremnant cichlid fauna in southern Lake Victoria. Conservation Biology 11,890–904.
Smith, G. R. & Todd, T. N. (1984). Evolution of species flocks of fishes in northtemperate lakes. In Evolution of Fish Species Flocks (Echelle, A. A. & Kornfield,I., eds), pp. 45–68. Orono: University of Maine Press.
Sokal, R. R. & Rohlf, F. J. (1981). Biometry: the Principles and Practice of Statistics inBiological Research, 2nd edn. San Francisco: W. H. Freeman.
Thery, M. & Vehrencamp, S. L. (1995). Light patterns as cues for mate choice in thelekking white-throated manakin (Corapipo gutturalis). Auk 112, 133–145.
Tregenza, T. & Bridle, J. R. (1997). The diversity of speciation. Trends in Ecology andEvolution 12, 382–383.
Turner, G. F. (1994). Speciation mechanisms in Lake Malawi cichlids: a critical review.Archiv fur Hydrobiologie Beihefte Limnologie 44, 139–160.
Turner, G. F. & Burrows, M. T. (1995). A model of sympatric speciation by sexualselection. Proceedings of the Royal Society of London B 260, 287–292.
Witte, F., Goudswaard, P. C., Katunzi, E. F. B., Mkumbo, O., Seehausen, O. & Wanink,J. H. (1998). Lake Victoria’s ecological changes and their relationships with theriparian societies. Lakes and People, in press. Shiga, Japan: Lake Biwa Museum.
Copyright © 2022 FDOKUMEN




















