
Binding interactions between peptides and proteins of the class II Major Histocompatibility Complex

This article was downloaded by: [University of Waterloo]On: 22 April 2014, At: 19:17Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Transactions of the American Fisheries SocietyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/utaf20
Differentiation of Sympatric Arctic Char MorphotypesUsing Major Histocompatibility Class II GenesPablo Conejerosa, Anna Phanb, Michael Powerb, Michael O’Connellc, Sergey Alekseyevd, IvánSalinasa & Brian Dixonb
a Centro de Investigación y Gestión de Recursos Naturales, Facultad de Ciencias, Universidadde Valparaíso, Gran Bretaña 1111, Valparaíso, 2360102, Chileb Department of Biology, University of Waterloo, 200 University Avenue West, Waterloo,Ontario, N2L 3G1, Canadac Fisheries and Oceans Canada, Science Branch, Post Office Box 5667, St. John's,Newfoundland and Labrador, A1C 5X1, Canadad Kolzov Institute of Developmental Biology, Russian Academy of Sciences, Ulitsa Vavilova 26,Moscow, 119991, RussiaPublished online: 14 Apr 2014.
To cite this article: Pablo Conejeros, Anna Phan, Michael Power, Michael O’Connell, Sergey Alekseyev, Iván Salinas & BrianDixon (2014) Differentiation of Sympatric Arctic Char Morphotypes Using Major Histocompatibility Class II Genes, Transactionsof the American Fisheries Society, 143:3, 586-594
To link to this article: http://dx.doi.org/10.1080/00028487.2014.880734
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Transactions of the American Fisheries Society 143:586–594, 2014C© American Fisheries Society 2014ISSN: 0002-8487 print / 1548-8659 onlineDOI: 10.1080/00028487.2014.880734
NOTE
Differentiation of Sympatric Arctic Char MorphotypesUsing Major Histocompatibility Class II Genes
Pablo ConejerosCentro de Investigacion y Gestion de Recursos Naturales, Facultad de Ciencias,Universidad de Valparaıso, Gran Bretana 1111, Valparaıso 2360102, Chile
Anna Phan and Michael PowerDepartment of Biology, University of Waterloo, 200 University Avenue West, Waterloo,Ontario N2L 3G1, Canada
Michael O’ConnellFisheries and Oceans Canada, Science Branch, Post Office Box 5667, St. John’s,Newfoundland and Labrador A1C 5X1, Canada
Sergey AlekseyevKolzov Institute of Developmental Biology, Russian Academy of Sciences, Ulitsa Vavilova 26,Moscow 119991, Russia
Ivan SalinasCentro de Investigacion y Gestion de Recursos Naturales, Facultad de Ciencias,Universidad de Valparaıso, Gran Bretana 1111, Valparaıso 2360102, Chile
Brian Dixon*Department of Biology, University of Waterloo, 200 University Avenue West, Waterloo,Ontario N2L 3G1, Canada
AbstractArctic Char Salvelinus alpinus have colonized northern post-
glacial lakes within the last few thousand years. Divergent pop-ulations have adapted to thrive in the prevailing oligotrophicenvironments and thus have developed morphotypes with differ-ent ecological behaviors. The morphotypes usually differ in size,morphology, coloration, feeding ecology, and/or habitat occupancy.Although morphotypes that have very divergent spawning seasonsshould become genetically segregated, genetic differentiation, inmost cases, has been weak. Thus, results to date have suggestedthat Arctic Char morphotype separation has been driven largely bythe environmentally mediated phenotypic plasticity of the species,with differentiation between morphotypes having commenced toorecently to generate substantial genetic drift. Here we used themajor histocompatibility (MH) class II genes in an attempt to iso-late sympatric Arctic Char morphotypes known to be ecologicallydifferentiated. These morphotypes are from postglacial lakes in
*Corresponding author: [email protected] September 13, 2012; accepted November 19, 2013
both Siberia and eastern Canada, and differ in either diet, habitatoccupancy, or both. The MH Class II allelic polymorphism wassignificantly different between morphotypes. This suggested thereis differential heritable adaptation to the natural selection exertedby pathogens unique to each ecological niche within each lake.
Arctic Char Salvelinus alpinus are salmonid fish distributedthroughout the northern circumpolar region and are notedfor variability in morphology, coloration, and ecology. Theyare relatively recent occupants of many ecosystems and havecolonized many North American habitats after the retreat of theLaurentide Ice Sheet approximately 14,000–5,000 years ago(Legendre and Legendre 1984). As a result, char-occupied habi-tats tend to be oligotrophic and contain few other fish species(Knudsen et al. 2006). Recently deglaciated and species-poor
586
Dow
nloa
ded
by [
Uni
vers
ity o
f W
ater
loo]
at 1
9:17
22
Apr
il 20
14

NOTE 587
ecosystems are generally thought to facilitate developmentof polymorphic forms in Arctic Char, with the relaxation ofinterspecific competition and open niche availability being seenas essential (Skulason and Smith 1995). Thus, deep lakes withseparate littoral, pelagic, and profundal zones that offer uniqueforaging opportunities have been argued to be characteristicof systems where polymorphic populations of Arctic Char willdevelop (Jonsson and Jonsson 2001). Studies from northernQuebec (Power et al. 2009) and Russia (Alekseyev et al.2009a), however, contradict that view, as in both localesmorphologically distinctive forms of Arctic Char occur inshallow lakes (<30 m) containing diverse fish communities.
Diversity within and among populations is undoubtedly re-inforced by the fact that Arctic Char are habitat generalists andmay be found at appropriate times of the year in a variety ofaquatic habitats (Power et al. 2008). As a result, differences infeeding ecology, growth rate, size, age at maturity, and morphol-ogy of Arctic Char are often noted in lakes containing distinctiveArctic Char morphotypes (Parker et al. 2001). Correlations be-tween diet, morphology, and life history characteristics havebeen used to suggest that selection driven by differential trophicecology plays a role in phenotypic divergence and ecologicalspeciation (Schluter 2000). Nevertheless, the measured geneticdiversity within and between Arctic Char morphotypes gener-ally remains low (Jonsson and Jonsson 2001). Since geneticdivergence in many cases may be hampered by the presumablyshort interval of time that has been available in postglacial en-vironments, genetic differentiation, in the past, was not foundamong any of the morphotypes by using the available popu-lation markers (Hindar 1986; Magnusson and Ferguson 1987;Danzmann et al. 1991; Volpe and Ferguson 1996). Neverthe-less, some recent studies using microsatellites on Arctic Charmorphotypes with known differences in spawning seasons haveshown genetic differentiation (Gislason et al. 1999; Westgaardet al. 2004; Wilson et al. 2004; Gordeeva et al. 2010). Thesestudies have confirmed reproductive isolation and indicated thatphenotypic distinction has a genetic component (e.g., GomezUchida et al. 2008; Power et al. 2009).
As an alternative to molecular techniques that use neutralpopulation markers, we studied the polymorphism of the majorhistocompatibility (MH) class II receptor. Major histocompati-bility complex genes in mammals, indeed all vertebrates exceptteleost fish, are so named because they are linked on a singlechromosome in a genetic complex. In teleost fish, however,they are not linked in a genetic complex (Bingulac-Popovicet al. 1997) and thus are referred to as simply MH genes (Dixonand Stet 2001). The MH polymorphism patterns, unlike neutralmarkers, besides representing reproductive isolation representadaptive divergence (Hess and Edwards 2002; Blais et al. 2007)that should be structured by the distinct environment that eachArctic Char morphotype inhabits. The MH protein exposespathogen-derived peptides to T-lymphocytes, which determinewhether the particles are self or nonself and initiate the necessaryimmune response (Steinmetz and Hood 1983). The MH recep-
tors are polymorphic at the population level, which increasesthe possibilities of initiating a successful immune response toan ever-growing diversity of pathogens (Dawkins et al. 1999;Lohm et al. 2002; Messaoudi et al. 2002). However, MH allelesthat confront locally specific pathogens are rapidly selected,either positively or negatively, such that stable populationssharing a common habitat tend to have common, environmen-tally determined patterns in their MH polymorphism (Lamont1998; Bernatchez and Landry 2003; Bonneaud et al. 2006).
In most jawed vertebrates, there are two classes of MH re-ceptors present: MH class II receptors differentiate betweenrecently separated populations (Hardee et al. 1995; Landry andBernatchez 2001; Miller et al. 2001), and MH class I recep-tors show deep lineages that stretch back to the ancestors ofmany modern groups of fish (van Erp et al. 1996). These recep-tors are composed of an α and β subunit encoded by differentgenes. The second exon of each gene encodes the α1 and β1domains, respectively, that bind the antigen to be presented toT-lymphocytes (peptide binding region) and are the highly vari-able area of the molecule (Steinmetz and Hood 1983).
Sympatric morphotypes often differ ecologically and behav-iorally: they encounter and consume different prey (Adamset al. 1998; McCarthy et al. 2004; Power et al. 2009), mayuse differing thermal environments (Power et al. 2012), and cancontain different parasitic loads (Knudsen et al. 1997). Link-ages between environment, use of a habitat, prey selection, andpathogen loads should have produced differential immunologi-cal resistance, which ought to be associated with adaptation offish MH receptors and could provide a readily available meansof separating genetic from ecological factors as determinants ofthe observed patterns of morphological diversity in Arctic Char.Accordingly, in this study, MH diversity was correlated withdata that indicated there is a distinction of sympatric morpho-types (habitat use: littoral or profundal) as well as prey resourceuse (trophic level), which is represented by isotope data.
METHODSFish samples.—Sympatric Arctic Char morphotypes resid-
ing in distinctive lake environments were obtained from GanderLake, Newfoundland and Labrador, (48◦55′N, 54◦35′W) usingLundgren multiple-mesh nylon monofilament experimentalgill nets set in a combination of benthic and pelagic nets asdescribed for previous Newfoundland applications by Hammarand Filipsson (1985) and O’Connell and Dempson (2002).The lake is 113.2 km2, 288 m deep, and contains distinctivethermal habitats in the epilimnion and hypolimnion (O’Connellet al. 2005), which are occupied, respectively, by “dark” and“pale” Arctic Char morphotypes (O’Connell and Dempson2002). They have distinctive morphologies (O’Connell andDempson 2002), are separable into two gene pools (GomezUchida et al. 2008), and can have distinctive feeding ecologies(Power et al. 2005) and habitat occupancies (Power et al. 2012).Substructuring within the morphotype groupings related to
Dow
nloa
ded
by [
Uni
vers
ity o
f W
ater
loo]
at 1
9:17
22
Apr
il 20
14

588 CONEJEROS ET AL.
depth in the pale group (shallow versus deep) and size in thedark group (large versus small) have also been noted (Poweret al. 2005). To avoid possible confounding influences asso-ciated with ecological substructuring, samples were randomlyselected from the morphotype groupings known to consistentlyuse distinctive habitat types (Power et al. 2009). The palemorphotype is known to use profundal habitats (>70 m) andthe small, dark morphotype is known to use the littoral zone ofthe lake and feed at a similar trophic position to the profundalpale morphotype (Power et al. 2005).
Sympatric Arctic Char morphotypes residing in nondistinc-tive lake environments were similarly obtained using Lund-gren monofilament experimental gill nets from Lakes Kiryalta-3(57◦8.5′N, 119◦27′E) and Kiryalta-4 (57◦6.5′N, 119◦27′E) in themountainous northern Transbaikalia region of Siberia, northeastof Lake Baikal, Russia. Lake Kiryalta is geographically dividedinto two water bodies connected by a 300-m canal that allowsfish movement from the upstream Lake Kiryalta-3 to the down-stream Lake Kiryalta-4 (the canal is steep so movement in theopposite direction is difficult). Given exchange of genetic mate-rial is possible, it was considered as a single site for this study.Both lakes are small (Lake Kiryalta-3: length, 2 km; width,0.25 km. Lake Kiryalta-4: length, 3.6 km; width, 0.6 km) andshallow (Lake Kiryalta-3: 20 m; Lake Kiryalta-4: 34 m). Thelakes contain two resident Arctic Char morphotypes separableon the basis of morphology and size (Alekseyev et al. 2002).They differ in feeding (trophic) ecology, with the smaller mor-photype in each lake specializing in zooplankton and the largermorphotypes specializing in fish (Alekseyev et al. 2009b). As inGander Lake, randomly selected samples of each morphotypewere used in the analyses, in which the separation into morpho-type groupings had been completed and confirmed followingAlekseyev et al. (2002). Sample numbers for both locations aregiven in Table 1.
Stable isotope testing.—Dorsal muscle samples were cutfrom each specimen and immersed in 70% ethanol or DNApreserving solution (25 mM C6H5NaO7, 10 mM EDTA and4 M [NH4]2SO4) and shipped to the laboratory at Universityof Waterloo, Ontario, for genetic analyses. Duplicate, nonpre-served samples were obtained for stable isotope analyses (SIA).Samples of dorsal muscle tissue for SIA were treated follow-ing standard protocols (e.g., Power et al. 2005, 2009) and ana-lyzed using a Carlo Erba elemental analyser (Carlo Erba, Milan,Italy) connected to a Micromass VG Isochrom continuous-flowisotope ratio mass spectrometer (GV Instruments [Micromass],Manchester, UK) with an analytical precision for δ13C and δ15N,respectively, of 0.2‰ and 0.3‰ (Power et al. 2005). An ANOVAof muscle tissue stable isotope values were used to confirm thelake-specific, morphological- and ecological-based groupingsdescribed above and previously in the literature (e.g., Poweret al. 2005).
MH genes.—The DNA was extracted from the samples by in-cubation of the fish muscle samples with proteinase K and stan-dard phenol–chloroform purification. Primers were designed to
TABLE 1. Class II α and class II β divergence in Arctic Char morphotypes:sample number, number of alleles, and other divergence values for each mor-photype in Gander Lake and the Kiryalta lakes.
Gander Lake Lake Kiryalta
Class Parameter Pale Dark Small Large
Class II α Number ofsamples
9 12 15 17
C II α alleles 4 7 4 8Unique alleles 0 3 1 5Exon divergence 0.07 0.07 0.05 0.05Rate dn : ds 2.1 4.3 4.1 3.9Intron divergence 0.02 0.05 0.03 0.02
Class II β Number ofsamples
6 8 6 6
C II β alleles 3 6 3 5Unique alleles 2 5 2 4Exon divergence 0.02 0.05 0.03 0.07Rate dn : ds 9.4 2.1 2.3 2Intron divergence 0.03 0.03 0.04 0.04
amplify the second exon and the second intron in MH classII α and β genes, respectively. The primers were based on thefull length sequences of Arctic Char (Conejeros et al. 2012).The class II β PCR forward primer (SAALDABF: 5′ – GATACT CCT CAA AGG ACC TG – 3′) and PCR reverse primer(SAALDABR2: TAG TCA TAC TGT AAT GKC ACT AC), andclass II α PCR forward primer (SAALDAAF: 5′ – CTG GATGCA GTG ATT CAG ATG – 3′) and PCR reverse primer (SAAL-DAAR: 5′ – GAC GTG GCA GAT GAG AGT G – 3′) were usedrespectively under the same conditions for the PCR: initial de-naturation at 95◦C for 2 min, plus 35 cycles of denaturation at95◦C for 30 s, primer annealing at 57◦C for 40 s, and extensionat 72◦C for 1.5 min.
The PCR amplicons were cloned in pGEM-T vectors(Promega Corporation, Madison, Wisconsin) and transformedinto XL-blue competent strains of Escherichia coli (Sambrooket al. 2001). Eight to 12 white colonies were selected and grownin Luria-Bertani media. The bacteria were used for PCR usingthe regular forward primer and a reverse primer for denaturantgradient gel electrophoresis (DGGE), designed to match theend of the second exon (SAALDAADGGER: CGG GCG GGGGCG GCG GGC CGG GCG CGG GGC GCG GCG GGC GCTATT GTC TCT GGT GGG TYC for the class II α gene andSAALDABDGGER: CGG GCG GGG GCG GCG GGC CGGGCG CGG GGC GCG GCG GGC GCC AGT ACG GCG CTGTAG for the class II β gene). The amplicons were separatedin a denaturing gradient of 20–45% and 15–50% for class II α
and β genes, respectively. Only the alleles that differed betweensamples were selected for DNA sequencing.
Sequencing and data analysis.—For the analysis the pres-ence or absence of certain alleles in each population were
Dow
nloa
ded
by [
Uni
vers
ity o
f W
ater
loo]
at 1
9:17
22
Apr
il 20
14

NOTE 589
considered. The sequences obtained were aligned using thesoftware Muscle (Edgar 2004). An alignment of the previouslysequenced full length class II α and β complementary DNA(cDNA) sequences (Conejeros et al. 2012) were used as a guidefor the definition of the end of the second exon and the begin-ning of the intron. The intron and exon sequences were analyzedseparately. BioEdit (Hall 1999) was used to define the openreading frames and to translate them into protein sequences.The software MEGA 4 (Tamura et al. 2007) was used to findand correct sequences containing singleton errors, to calculatethe diversity of the sequences among and within populations,the ratios of nonsynonymous : synonymous substitutions, andto draw the phylogenetic trees using the Nei–Gojobori model(Nei and Gojobori 1986) for nonsynonymous substitutions inthe exon sequences. Arlequin 3.01 software (Excoffier et al.2005) was used to calculate the allelic frequencies and pairwiseF-statistics (FST). The distribution of alleles was also checkedby multivariate analysis using a presence–absence matrix ofindividuals versus alleles.
RESULTS
Stable Isotope TestingIn both Gander Lake and the Kiryalta lakes, Arctic Char
morphotype dorsal muscle tissues were differentiated on thebasis of stable isotope values (Figure 1). The pale and darkmorphotype groupings from Gander Lake differed significantlyin δ13C (ANOVA: F1, 18 = 178.76, P < 0.001) and small andlarge morphotype groupings from Lake Kiryalta differed signifi-cantly in both δ13C (ANOVA: F1, 18 = 6.71, P = 0.018) and δ15N
FIGURE 1. Stable isotope values for muscle tissue from Arctic Char mor-photypes examined in Gander Lake and the Kiryalta lakes. Gander dark andpale morphotypes are plotted, respectively, as open and filled boxes, whereasKiryalta large and small morphotypes are plotted, respectively, as open and filledcircles. Gander Lake morphotypes evidence a significant separation on the δ13Caxis related to differential habitat use (littoral versus profundal) and Kiryaltalakes morphotypes evidence significant separation on both isotope axes, butmost notably on the δ15N axis as a result of feeding at different tropic levels.
TABLE 2. Class II α and class II β divergence in Arctic Char morphotypes:interpopulation divergence values between the morphotypes for both genes forGander Lake and the Kiryalta lakes, including the interpopulation diversityand the interpopulation ratio of nonsynonymous : synonymous substitutions(dn : ds).
Interpopulation InterpopulationClass Lake diversity dn : ds ratio
Class II α Gander 0.007 59.133Kiryalta <0.001 <0.001
Class II β Gander 0.006 2.442Kiryalta 0.011 2.852
(ANOVA: F1, 18 = 146.43, P < 0.001). Morphotype groupingsfrom Gander Lake did not differ significantly in δ15N (ANOVA:F1, 18 = 0.54, P = 0.472).
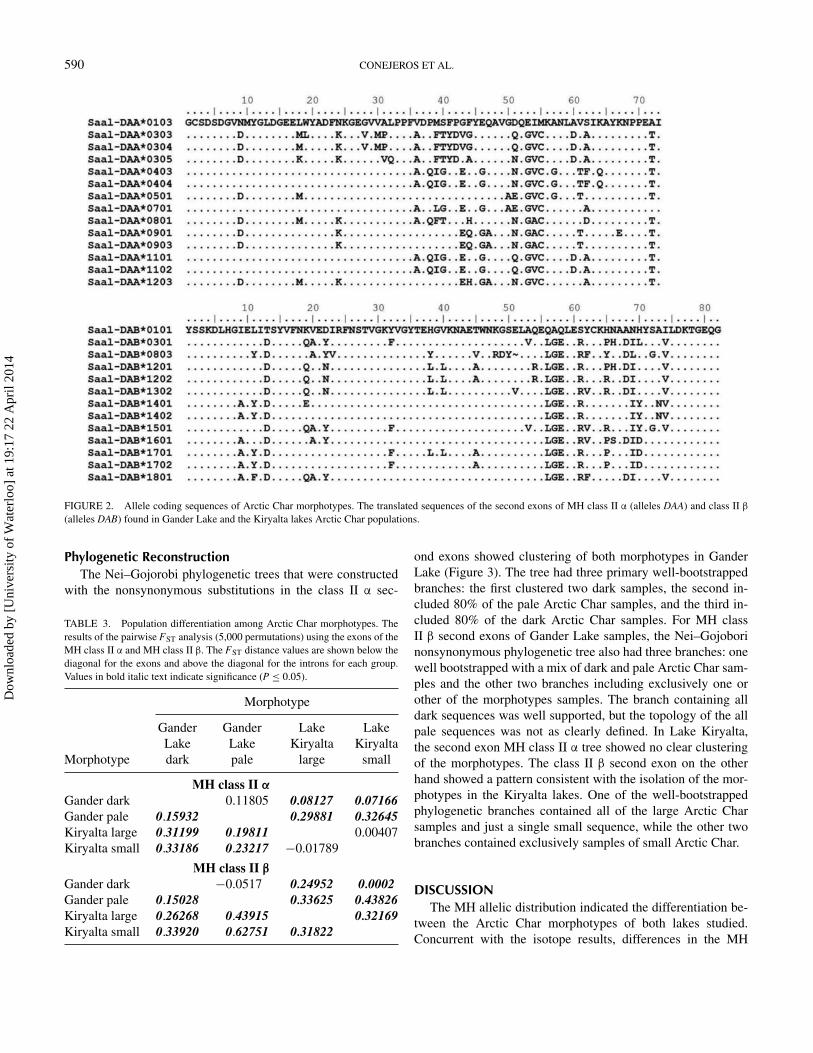
MH Class II AllelesIn total, 14 MH class II α1 and 13 MH class II β1 allele
sequences were identified from all of the samples in this study(Figure 2). No MH class II β alleles were shared between the fishfrom the two lake systems. For MH class II α allele sequences,only two alleles were shared. The most common alpha allele,Saal-DAA*0501, was found in five fish from Gander Lake andeight fish from Lake Kiryalta. The DAA*0303 allele was foundin four fish from Gander Lake and one fish from Lake Kiryalta.Divergence values as well as other relevant data for the allelesobtained of MH class II α and β are shown in Tables 1 and 2.
Population DifferentiationPairwise F-statistics (FST) between Gander Lake morpho-
type class II α second exon sequences indicated separation ofthe morphotypes (Table 3). When considering the second in-trons of the class II α gene, no isolation of the morphotypepopulations was found. The FST applied to the second exon ofthe class II β gene indicated morphotype isolation in GanderLake. Unlike the MH class II α intron analysis, the FST analysiswas significant between both morphotypes when the class II β
intron sequence was examined. On the other hand, pairwise FST
of the second exon sequences of MH class II α did not suggestthe isolation of the morphotypes within the Kiryalta lakes. Thesame result was obtained when comparing the MH class II α
second introns. However, FST estimates, when applied over theMH class II β second exon sequences, did suggest the isolationof the morphotypes. The isolation of large and small Arctic Charwas also supported by the FST analysis of the second introns ofthe MH class II β gene. The interpopulation ratio of nonsyn-onymous : synonymous substitutions (dn : ds) values alwaysindicated positive selection in the exons. Interpopulation dn : dsalso indicated positive selection in all cases, except that of theclass II α gene in Lake Kiryalta.
Dow
nloa
ded
by [
Uni
vers
ity o
f W
ater
loo]
at 1
9:17
22
Apr
il 20
14
590 CONEJEROS ET AL.
FIGURE 2. Allele coding sequences of Arctic Char morphotypes. The translated sequences of the second exons of MH class II α (alleles DAA) and class II β
(alleles DAB) found in Gander Lake and the Kiryalta lakes Arctic Char populations.
Phylogenetic ReconstructionThe Nei–Gojorobi phylogenetic trees that were constructed
with the nonsynonymous substitutions in the class II α sec-
TABLE 3. Population differentiation among Arctic Char morphotypes. Theresults of the pairwise FST analysis (5,000 permutations) using the exons of theMH class II α and MH class II β. The FST distance values are shown below thediagonal for the exons and above the diagonal for the introns for each group.Values in bold italic text indicate significance (P ≤ 0.05).
Morphotype
Gander Gander Lake LakeLake Lake Kiryalta Kiryalta
Morphotype dark pale large small
MH class II α
Gander dark 0.11805 0.08127 0.07166Gander pale 0.15932 0.29881 0.32645Kiryalta large 0.31199 0.19811 0.00407Kiryalta small 0.33186 0.23217 −0.01789
MH class II β
Gander dark −0.0517 0.24952 0.0002Gander pale 0.15028 0.33625 0.43826Kiryalta large 0.26268 0.43915 0.32169Kiryalta small 0.33920 0.62751 0.31822
ond exons showed clustering of both morphotypes in GanderLake (Figure 3). The tree had three primary well-bootstrappedbranches: the first clustered two dark samples, the second in-cluded 80% of the pale Arctic Char samples, and the third in-cluded 80% of the dark Arctic Char samples. For MH classII β second exons of Gander Lake samples, the Nei–Gojoborinonsynonymous phylogenetic tree also had three branches: onewell bootstrapped with a mix of dark and pale Arctic Char sam-ples and the other two branches including exclusively one orother of the morphotypes samples. The branch containing alldark sequences was well supported, but the topology of the allpale sequences was not as clearly defined. In Lake Kiryalta,the second exon MH class II α tree showed no clear clusteringof the morphotypes. The class II β second exon on the otherhand showed a pattern consistent with the isolation of the mor-photypes in the Kiryalta lakes. One of the well-bootstrappedphylogenetic branches contained all of the large Arctic Charsamples and just a single small sequence, while the other twobranches contained exclusively samples of small Arctic Char.
DISCUSSIONThe MH allelic distribution indicated the differentiation be-
tween the Arctic Char morphotypes of both lakes studied.Concurrent with the isotope results, differences in the MH
Dow
nloa
ded
by [
Uni
vers
ity o
f W
ater
loo]
at 1
9:17
22
Apr
il 20
14

NOTE 591
FIGURE 3. The phylogenetic trees constructed from the exon nucleotide sequences with the Nei–Gojobori model, considering only nonsynonymous mutations.(A) and (B) are the trees constructed from the data obtained for Gander Lake and (C) and (D) are the trees for the Kiryalta lakes. (A) and (C) show the phylogenetictrees based on class II α genes and (B) and (D) show the phylogenetic trees based on class II β genes. Dark and large Arctic Char morphotype samples are shaded,while pale and small morphotype samples are not shaded.
allele composition also varied, with MH-based separation beingstronger in Gander Lake, where morphotypes occupied differenthabitats, than in the Kiryalta lakes, where ecological separationbetween the morphs was based mostly on feeding ecology.
MH Class II � and MH Class II � PolymorphismThe high divergence of MH class II α and MH class II β genes
in Arctic Char from Gander Lake and the Kiryalta lakes wasmostly caused by intrapopulation variation. This indicated thatonly a small percentage of the observed variation had occurredseparately within each morphotype stock, consistent with ourprevious observations (Conejeros et al. 2008). Nevertheless, theinterpopulation diversity seen here reaches high rates of nonsyn-onymous : synonymous substitutions suggesting that many ofthe changes were related to adaptation within the separate eco-logical niches. Considering the lower number of alleles found inthe pale or small Arctic Char, they seem to have have lost vari-ability compared with the larger forms during the adaptation.An equivalent loss of variability of intron sequences indicated
that, as opposed to single exon changes, complete alleles mighthave been lost during the adaptation process.
Larger Arctic Char resident in the littoral zone display morediverse trophic behavior than the smaller or profundal mor-photypes (Power et al. 2005), which is often related to a highernumber of parasite infections (Knudsen 1995; Lafferty et al.2006; Seilacher et al. 2007). Therefore, consistent with reportsin Norwegian lakes (Knudsen et al. 1997), it is possible that darkArctic Char from Gander Lake and the larger Arctic Char inthe Kiryalta lakes are infected by a significantly greater numberof parasite species than are the pale and small morphotypes inthose locations. Indeed, one of us has data on parasites present in20 dark (not separated into large versus small) and 20 pale (notseparated into shallow versus deep) Gander Lake char showingthat all 20 of the dark char were parasitized compared withonly 5 out of 20 for the pale char (M. O’Connell, unpublisheddata); the mean parasite load for dark char was 10.5 comparedwith 1.5 for pale char. The main parasites were cestodes (meanload of 6.3 for dark char and 0.4 for pale char) but trematodes,
Dow
nloa
ded
by [
Uni
vers
ity o
f W
ater
loo]
at 1
9:17
22
Apr
il 20
14

592 CONEJEROS ET AL.
nematodes, and Acanthocephala were also present. Conse-quently, the loss of MH allele diversity in the pale and smallmorphotypes may be due to selection for a lower number of MHalleles because these fish confront a lower variety of pathogensin the profundal zone or restricted feeding niches. Conversely,in Lake Kiryalta the increased parasite loads in the larger formsmay be because, as piscivores, they ingest more parasites. Ineither case, the parasite data correspond with the relationshipbetween isotopic separation of morphotypes and MH alleledifferentiation. Fish that live in different habitats have moredivergence than fish that are separated by feeding ecology alone.
Arctic Char MorphotypesIn most population studies the separation of Arctic Char
morphotypes living in sympatry has been weak (Hindar 1986;Magnusson and Ferguson 1987; Danzmann et al. 1991; Volpeand Ferguson 1996; Gislason et al. 1999; Wilson et al. 2004),which obstructs the understanding of the biological significanceof the morphotype segregation. Although some reports haveshown the heritability of the characteristics specific to the smallor large morphotypes (Svedang 1990; Westgaard et al. 2004;Gomez Uchida et al. 2008), the genetic results to date havereinforced the argument that the segregation in most cases has itsorigin in phenotypic plasticity acted on by varying environments(Skulason et al. 1996; Jonsson and Jonsson 2001).
Although MH molecule variation results in part from geneticdrift, a strong component of the polymorphism in a given popu-lation is established by the natural selection of the alleles (Bryjaet al. 2006; de Eyto et al. 2007). The MH alleles can be selectedout of, or fixed in, a population in an extremely short periodof time since the natural selection exerted by pathogens rapidlyeliminates the individuals that are not fit for the ecological nichethey occupy.
Accordingly, in this study the differentiation between themorphotypes in both lakes was much clearer when consideringMH exons rather than introns, reinforcing the idea of the adap-tive nature of the divergence. In both lakes, the morphotypeshave different life histories (i.e., age at maturity) and inhabit dif-ferent environments or different prey recourses. In the Kiryaltalakes, the small morphotype consumes mainly zooplankton andaquatic macroinvertebrates while the large morphotype is pis-civorous (Alekseyev et al. 2002). In Gander Lake, dark and paleArctic Char are consistently separated along niche resource(Power et al. 2005) and habitat occupancy axes (Power et al.2012). Such habitat and feeding differences should be reflectedby the pathogens to which the morphotypes are exposed, as hasbeen reported in other lakes where morphotypes exist (Knudsenet al. 1997). Specialization in trophic behavior can expose indi-viduals to different suites of selection pressures that are stableover time and provide a platform for disruptive selection to op-erate within sympatric systems (Knudsen et al. 2011). Thus, thefact that the Kiryalta lakes morphotypes showed differentiationonly with the class II β gene while Gander Lake morphotypesdisplayed both α and β gene differentiation indicates this may
be a consequence of the greater niche differentiation shownin Gander Lake (habitat and resource use) compared with theKiryalta lakes (resource use). On the other hand, while only theclass II β was significantly different between the Kiryalta lakesmorphotypes, the differentiation values (i.e., FST) were similarto those of the α and β genes in Gander Lake and may thensimply reflect the nature of the pathogens within the Kiryaltalakes system.
As seen in this study, Arctic Char morphotypes within Gan-der Lake and the Kiryalta lakes have adapted rapidly by main-taining population-specific class II α and/or class II β allelesthat have aided in their becoming separate populations withheritable genetic characters that facilitate environment-specificsurvival. The data shown here add complementary informationto the neutral data, presenting evidence of inherited functionaldifferentiation of the morphotypes. Each morphotype possessesexclusive MH alleles, which implies that movement from onestock (habitat) to another happens rarely, because most fish maynot have the alleles necessary for survival in the other habitat.Nevertheless, positive selection in MH genes forces the mainte-nance of ancestral alleles in the population (Klein 1987), as ob-served by the occurrence of alleles shared between morphotypesin both lakes. The sharing of alleles between the morphotypessuggests that they arose from a common ancestral stock, butthat sufficient differences have since accumulated for heritablefunctional differentiation to be detected. The presence of differ-entiation in Arctic Char studied in both lakes that have disparatelocations and environmental conditions suggests that rapid MHallele differentiation is common and will occur for most otherlakes where sympatric morphotypes exist.
ACKNOWLEDGMENTSSupport for the work was provided by Fisheries and Oceans
Canada, the Gander River Management Association, NaturalScience and Engineering Research Council of Canada Discov-ery Grant numbers 217529-2008 to B.D. and 155928-2010 toM.P., and the Russian Foundation for Basic Research (projectnumber 11-04-00109): Program of Fundamental Studies of thePresidium of Russian Academy of Sciences “Biodiversity” toS.A. We also thank A. Walsh, R. Noel, B. Gillingham, G.Furey, M. Bloom, M. Shears, N. Sinnatamby, V. Samusenok, A.Matveev, I. Knizhin, A. Yur’ev, and A. Vokin for field collectionand laboratory analyses of the samples. Nucleotide sequencedata reported are available in the GenBank databases underthe accession numbers: EF450348–EF450407 and EU159605–EU159630.
REFERENCESAdams, C. E., D. Fraser, F. A. Huntingford, R. B. Greer, C. M. Askew, and
A. F. Walker. 1998. Trophic polymorphism amongst Arctic Charr from LochRannoch, Scotland. Journal of Fish Biology 52:1259–1271.
Alekseyev, S. S., R. Bajno, N. V. Gordeevna, J. D. Reist, M. Power, A. F.Kirillov, V. P. Samusenok, and A. N. Matveev. 2009a. Phylogeography andsympatric differentiation of the Arctic Charr Salvelinus alpinus (L.) complex
Dow
nloa
ded
by [
Uni
vers
ity o
f W
ater
loo]
at 1
9:17
22
Apr
il 20
14

NOTE 593
in Siberia as revealed by mtDNA sequence analysis. Journal of Fish Biology75:368–392.
Alekseyev, S. S., M. V. Mina, E. M. Smirina, and A. A. Sokolov. 2009b. Lateontogeny growth acceleration and size form transformations in TransbaikalianArctic Charr, Salvelinus alpinus complex: evidence from fin ray cross sectiongrowth layers. Environmental Biology of Fishes 86:487–505.
Alekseyev, S. S., V. P. Samusenok, A. N. Matveev, and M. Y. Pichugin. 2002.Diversification, sympatric speciation, and trophic polymorphism of ArcticCharr, Salvelinus alpinus complex, in Transbaikalia. Environmental Biologyof Fishes 64:97–114.
Bernatchez, L., and C. Landry. 2003. MH studies in nonmodel vertebrates: whathave we learned about natural selection in 15 years? Journal of EvolutionaryBiology 16:363–377.
Bingulac-Popovic, J., F. Figueroa, A. Sato, W. S. Talbot, S. L. Johnson, M.Gates, J. H. Postletwaith, and J. Klein. 1997. Mapping of the class I andclass II region to different linkage groups in the zebrafish. Immunogenetics46:129–134.
Blais, J., C. Rico, C. van Oosterhout, J. Cable, G. F. Turner, and L. Bernatchez.2007. MH adaptive divergence between closely related and sympatric Africancichlids. PLoS One [online serial] 2(8):e734.
Bonneaud, C., O. Chastel, P. Federici, H. Westerdahl, and G. Sorci. 2006.Complex Mhc-based mate choice in a wild passerine. Proceedings of theRoyal Society of London Series B Biological Sciences 273:1111–1116.
Bryja, J., M. Galan, N. Charbonnel, and J. F. Cosson. 2006. Duplication, balanc-ing selection and trans-species evolution explain the high levels of polymor-phism of the DQA MH class II gene in voles (Arvicolinae). Immunogenetics58:191–202.
Conejeros, P., A. Phan, M. Power, S. Alekseyev, M. O’Connell, B. Dempson, andB. Dixon. 2008. MH class IIa polymorphism in local and global adaptationof Arctic Charr (Salvelinus alpinus L.). Immunogenetics 60:325–337.
Conejeros, P., M. Power, S. Alekseyev, and B. Dixon. 2012. Global major his-tocompatibility class II β (mh-IIβ)-polymorphism in Arctic Charr Salvelinusalpinus. Journal of Fish Biology 81:1158–1174.
Danzmann, R., M. Ferguson, S. Skualson, S. Snorranson, and D. Noakes. 1991.Mitochondrial DNA diversity among four sympatric morphs of Arctic Charr,Salvelinus alpinus L, from Thingvallavatn, Iceland. Journal of Fish Biology39:649–659.
Dawkins, R., C. Leelayuwat, S. Gaudieri, G. Tay, J. Hui, S. Cattley, P. Martinez,and J. Kulski. 1999. Genomics of the major histocompatibility complex:haplotypes, duplication, retroviruses and disease. Immunological Reviews167:275–304.
de Eyto, E., P. McGinnity, S. Consuegra, J. Coughlan, J. Tufto, K. Farrell, H.-J.Megens, W. Jordan, T. Cross, and R. J. M. Stet. 2007. Natural selection actson Atlantic Salmon major histocompatibility (MH) variability in the wild.Proceedings of the Royal Society of London Series B Biological Sciences274:861–869.
Dixon, B., and R. J. M. Stet. 2001. The relationship between major histocom-patibility receptors and innate immunity in teleost fish. Developmental andComparative Immunology 25:683–699.
Edgar, R. C. 2004. MUSCLE: multiple sequence alignment with high accuracyand high throughput. Nucleic Acids Research 32:1792–1797.
Excoffier, L., G. Laval, and S. Schneider. 2005. Arlequin (version 3.0): an inte-grated software package for population genetics data analysis. EvolutionaryBioinformatics Online [online serial] 1:47–50.
Gislason, D., M. Ferguson, S. Skulason, and S. S. Snorrason. 1999. Rapidand coupled phenotypic and genetic divergence in Icelandic Arctic Char(Salvelinus alpinus). Canadian Journal of Fisheries and Aquatic Sciences56:2229–2234.
Gomez Uchida, D., K. P. Dunphy, M. F. O’Connell, and D. E. Ruzzante.2008. Genetic divergence between sympatric Arctic Charr Salvelinus alpinusmorphs in Gander Lake, Newfoundland: roles of migration, mutation andunequal effective population sizes. Journal of Fish Biology 73:2040–2057.
Gordeeva, N., A. Osinov, S. Alekseyev, A. Matveev, and V. Samusenok. 2010.Genetic differentiation of Arctic Charr Salvelinus alpinus complex from
Transbaikalia revealed by microsatellite markers. Journal of Ichthyology50:351–361.
Hall, T. A. 1999. BioEdit: a user-friendly biological sequence alignment editorand analysis program for Windows 95/98/NT. Nucleic Acids SymposiumSeries 41:95–98.
Hammar, J., and O. Filipsson. 1985. Ecological test fishing with the lundgrengillnets of multiple mesh size: the Drottningholm technique modified forNewfoundland Arctic Char populations. Institute of Freshwater ResearchDrottningholm Report 62:13–35.
Hardee, J. J., U. Godwin, R. Benedetto, and T. J. McConnell. 1995. Majorhistocompatibility complex class II A gene polymorphism in the StripedBass. Immunogenetics 41:229–238.
Hess, C. M., and S. V. Edwards. 2002. The evolution of the major histocompat-ibility complex in birds. Bioscience 52:423–431.
Hindar, K. 1986. Genetic differenciation among local populations and morpho-types of Arctic Charr, Salvelinus alpinus. Biologicl Journal of the LinneanSociety 27:269–285.
Jonsson, B., and N. Jonsson. 2001. Polymorphism and speciation in ArcticCharr. Journal of Fish Biology 58:605–638.
Klein, J. 1987. Origin of major histocompatibility complex polymor-phism: the trans-species hypothesis. Human Immunology 19:155–162.
Knudsen, R. 1995. Relationships between habitat, prey selection and parasiteinfection in Arctic Charr (Salvelinus alpinus). Nordic Journal of FreshwaterResearch 71:333–344.
Knudsen, R., A. Klemetsen, P. A. Amundsen, and B. Hermansen. 2006. Incip-ient speciation through niche expansion: an example from the Arctic Charrin a subarctic lake. Proceedings of the Royal Society of London Series BBiological Sciences 273:2291–2298.
Knudsen, R., R. Kristoffersen, and P. A. Amundsen. 1997. Parasite communitiesin two sympatric morphs of Arctic Charr, Salvelinus alpinus (L.), in northernNorway. Canadian Journal of Zoology 75:2003–2009.
Knudsen, R., A. Siwertsson, C. E. Adams, M. Garduno-Paz, J. Newton, andP.-A. Amundsen. 2011. Temporal stability of niche use exposes sympatricArctic Charr to alternative selection pressures. Evolutionary Ecology 25:589–604.
Lafferty, K., A. Dobson, and A. Kuris. 2006. Parasites dominate food web links.Proceedings of the National Academy of Sciences of the USA 103:11211–11216.
Lamont, S. J. 1998. Impact of genetics on disease resistance. Poultry Science77:1111–1118.
Landry, C., and L. Bernatchez. 2001. Comparative analysis of population struc-ture across environments and geographical scales at major histocompatibilitycomplex and microsatellite loci in Atlantic Salmon (Salmo salar). MolecularEcology 10 (:2525–2539.
Legendre, P., and V. Legendre. 1984. Postglacial dispersal of freshwater fishesin the Quebec peninsula. Canadian Journal of Fisheries and Aquatic Sciences41:1781–1802.
Lohm, J., M. Grahn, Å. Langefors, Ø. Andersen, A. Storset, and T. von Schantz.2002. Experimental evidence for major histocompatibility complex-allele-specific resistance to a bacterial infection. Proceedings of the Royal Societyof London Series B Biological Sciences 269:2029–2033.
Magnusson, K., and M. Ferguson. 1987. Genetic analysis of four syrnpatricrnorphs of Arctic Charr, Salvelinus alpinus, from Thingvallavatn, Iceland.Environmental Biology of Fishes 20:67–73.
McCarthy, I., D. Fraser, S. Waldron, and C. Adams. 2004. A stable isotopeanalysis of trophic polymorphism among Arctic Charr from Loch Ericht,Scotland. Journal of Fish Biology 65:1435–1440.
Messaoudi, I., J. A. Guevara Patino, R. Dyall, J. LeMaoult, and J. Nikolich-Zugich. 2002. Direct link between mhc polymorphism, T cell avidity, anddiversity in immune defense. Science 298:1797–1800.
Miller, K. M., K. H. Kaukinen, T. D. Beacham, and R. E. Withler. 2001.Geographic heterogeneity in natural selection on an MH locus in sockeyesalmon. Genetica 111:237–257.
Dow
nloa
ded
by [
Uni
vers
ity o
f W
ater
loo]
at 1
9:17
22
Apr
il 20
14

594 CONEJEROS ET AL.
Nei, M., and T. Gojobori. 1986. Simple methods for estimating the numbers ofsynonymous and nonsynonymous nucleotide substitutions. Molecular Biol-ogy and Evolution 3:418–426.
O’Connell, F., and B. Dempson. 2002. The biology of Arctic Charr,Salvelinus alpinus, of Gander Lake, a large, deep, oligotrophic lakein Newfoundland, Canada. Environmental Biology of Fishes 64:115–126.
O’Connell, M. F., J. B. Dempson, and M. Power. 2005. Ecology and trophicrelationships of the fishes of Gander Lake, a large, deep, oligotrophic lakein Newfoundland, Canada. International Review of Hydrobiology 90:486–510.
Parker, H., E. Noonburg, and R. Nisbet. 2001. Models of alternative life-history strategies, population structure and potential speciation in salmonidfish stocks. Journal of Animal Ecology 70:260–272.
Power, M., F. O’Connell, and B. Dempson. 2005. Ecological segregation withinand among Arctic Char morphotypes in Gander Lake, Newfoundland. Envi-ronmental Biology of Fishes 73:263–274.
Power, M., M. F. O’Connell, and B. Dempson. 2012. Determining the consis-tency of thermal habitat segregation within and among Arctic Charr morpho-types in Gander Lake, Newfoundland. Ecology of Freshwater Fish 21:245–254.
Power, M., G. Power, J. D. Reist, and R. Bajno. 2009. Ecological and geneticdifferentiation among the Arctic Charr of Lake Aigneau, Northern Quebec.Ecology of Freshwater Fish 18:445–460.
Power, M., J. Reist, and B. Dempson. 2008. Fish in high altitude lakes. Pages249–268 in W. F. V. a. J. Laybourn-Parry, editor. Polar lakes and rivers:limnology of Arctic and Antarctic aquatic ecosystems. Oxford UniversityPress, Oxford, UK.
Sambrook, J., and D. W. Russell. 2001. Molecular cloning: a laboratory manual,3rd edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NewYork.
Schluter, D. 2000. The ecology of adaptive radiation. Oxford University Press,New York.
Seilacher, A., W. E. Reif, and P. Wenk. 2007. The parasite connection in ecosys-tems and macroevolution. Naturwissenschaften 94:155–169.
Skulason, S., and T. B. Smith. 1995. Resource polymorphisms in vertebrates.Trends in Ecology and Evolution 10:366–370.
Skulason, S., S. Snorrason, D. Noakes, and M. Ferguson. 1996. Genetic basisof life history variations among sympatric morphs of Arctic Char, Salveli-nus alpinus. Canadian Journal of Fisheries and Aquatic Sciences 53:1807–1813.
Steinmetz, M., and L. Hood. 1983. Genes of the major histocompatibility com-plex in mouse and man. Science 222:727–733.
Svedang, H. 1990. Genetic basis of life-history variation of dwarf and normalArctic Charr, Salvelinus alpinus (L.), in Stora Rosjon, central Sweden. Journalof Fish Biology 36:917–932.
Tamura, K., J. Dudley, M. Nei, and S. Kumar. 2007. MEGA4: molecular evolu-tionary genetics analysis (MEGA) software version 4.0. Molecular Biologyand Evolution 24:1596–1599.
van Erp, S. H. M., E. Egberts, and R. J. M. Stet. 1996. Evidence for multipledistinct major histocompatibility complex class I lineages in teleostean fish.International Journal of Immunogenetics 23:371–381.
Volpe, J. P., and M. M. Ferguson. 1996. Molecular genetic examination ofthe polymorphic Arctic Charr Salvelinus alpinus of Thingvallavatn, Iceland.Molecular Ecology 5:763–772.
Westgaard, J., A. Klemetsen, and R. Knudsen. 2004. Genetic differences be-tween two sympatric morphs of Arctic Charr confirmed by microsatelliteDNA. Journal of Fish Biology 65:1185–1191.
Wilson, A. J., D. Gislason, S. Skulason, S. S. Snorrason, C. E. Adams, G.Alexander, R. G. Danzmann, and M. M. Ferguson. 2004. Population geneticstructure of Arctic Charr, Salvelinus alpinus from northwest Europe on largeand small spatial scales. Molecular Ecology 13:1129–1142.
Dow
nloa
ded
by [
Uni
vers
ity o
f W
ater
loo]
at 1
9:17
22
Apr
il 20
14
Copyright © 2022 FDOKUMEN




















