
Lake Trout (Salvelinus namaycush) Populations in Lake Superior and Their Restoration in 1959–1993

Relationships between lake ecology and morphologicalcharacters in Icelandic Arctic charr, Salvelinus alpinus
BJARNI K. KRISTJÁNSSON1*, HILMAR J. MALMQUIST2, FINNUR INGIMARSSON2,THÓRÓLFUR ANTONSSON3, SIGURD̄UR S. SNORRASON4 and SKÚLI SKÚLASON1
1Hólar University College, Háeyri 1, 550 Sauðárkrókur, Iceland2Natural History Museum of Kópavogur, Hamraborg 6a, 200 Kópavogur, Iceland3Institute of Freshwater Fisheries, Keldnaholti, 112 Reykjavik, Iceland4University of Iceland, Institute of Biology, Sturlugata 7, 101 Reykjavik, Iceland
Received 29 November 2010; revised 28 February 2011; accepted for publication 28 February 2011bij_1670 761..771
The common occurrence of parallel phenotypic patterns suggests that a strong relationship exists betweenecological dynamics and micro-evolution. Comparative studies from a large number of populations under vary-ing sets of ecological drivers could contribute to a better understanding of this relationship. We used data onmorphology of arctic charr (Salvelinus alpinus) and ecological factors from 35 Icelandic lakes to test the hypothesisthat morphological patterns among monomorphic charr populations from different lakes are related to interlakevariation in ecological characteristics. There is extensive phenotypic diversity among populations of Icelandic charr,and populations are easily distinguished based on overall body morphology. The results obtained in the presentstudy showed that the morphological diversity of charr was related to large-scale diversity in lake ecology. Variationin charr morphology was related to water origin (e.g. spring fed versus run-off), bedrock age, and fish communitystructure. The present study shows how various ecological factors can shape the biological diversity that weobserve. © 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, 103, 761–771.
ADDITIONAL KEYWORDS: adaptation – biodiversity – ESIL project – fish diversity – limnology –micro-evolution – morphometrics – natural selection – phenotypic plasticity.
INTRODUCTION
Biological diversity is commonly equated with speciesdiversity, which is a misconception because diver-sity does occur at multiple levels, including intraspe-cific diversity. Intraspecific phenotypic diversity isthe product of genes and the environment and theinteraction of these factors. Intraspecific biologicaldiversity is influenced by evolutionary history amongpopulations and species, which results in differencesin phenotypes, even though they are found insimilar environments (Langerhans & DeWitt, 2004).However, species or population may show morphologi-cal adaptations as a reaction to differences in theenvironment through phenotypic plasticity or throughlocal adaptation by natural selection. Both processes
would result in parallel patterns in phenotypes undersimilar environmental conditions (Nagel et al., 1998;Schluter, 2000; Schluter et al., 2004).
An important factor of adaptive radiation and par-allel evolution is the relationship between particularphenotype and specific niche, often referred to asphenotype–environment correlation (Schluter, 2000).Most commonly, this has been observed where feedingstructure and behaviour is correlated with a specificdiet niche (Wimberger, 1994; Schluter, 2000; Price,2008). For example, this can be clearly seen in thedivergence of beak morphology of Darwin’s groundfinches (Geospiza spp.), where beak morphology andfeeding behaviour can be related to feeding on specificseeds (Grant, 1999). An example of this is the evolu-tion of sympatric benthic/limnetic morphs seen in theadaptive radiation of Northern freshwater of fishes(Skúlason, Snorrason & Jónsson, 1999; Robinson &Schluter, 2000; Schluter, 2000).*Corresponding author. E-mail: [email protected]
Biological Journal of the Linnean Society, 2011, 103, 761–771. With 4 figures
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, 103, 761–771 761

Parallel evolution and phenotype–environmentcorrelations have been exclusively studied in sym-patric populations. However, do ecological factors,through local adaptation, shape phenotypic diversityof monomorphic populations in a similar way as thatobserved among sympatric populations? Reimchen& Nosil (2006) compared 40 populations of fresh-water threespine stickleback (Gasterosteus aculeatus)from British Columbia, Canada, and found that mor-phology of sticklebacks could be related to their habi-tats. Sticklebacks in smaller lakes or ponds had lessdefence structure and more ‘benthic’ trophic mor-phology than stickleback in larger lakes. Comparingpopulations and their habitats can provide valuableinformation on the ecological drivers of repeated evo-lutionary trajectories. However, this can be difficultto examine in complex ecosystems. Thus, studies onrelatively simple ecosystems, such as areas that haverecently become available or isolated islands, canprove valuable in this respect.
Northern freshwater systems are relatively youngand have only been accessible to fishes since the endof the last glaciation, approximately 10 000 years ago(Skúlason et al., 1999; Snorrason & Skúlason, 2004).These systems are generally species poor and coloniz-ing fish species have been presented with a diversityof vacant niches. Thus, they have evolved greatintraspecific diversity, where phenotypically similarmorphs or species are often found to occupy similarniches in different lakes (Skúlason et al., 1992;Snorrason & Skúlason, 2004). This has been observedin many unrelated fish species, including salmonids(Salmo, Oncorhynchus and Salvelinus spp.), sunfish(Lepomis spp.), smelt (Osmerus spp.), whitefish (Pros-pium, Coregonus spp.), and threespine stickleback(Wimberger, 1994; Skúlason et al., 1999; Robinson &Schluter, 2000) Arctic charr (Salvelinus alpinus) offergreat opportunities for comparative studies on eco-logical divergence. The species is widespread in lakesand rivers and occupies a range of different habitats(Klemetsen, 2010). Arctic charr have long been recog-nized for their extensive phenotypic variationand lakes that host sympatric morphs, frequentlylimnetic (planktonic and fish eating) versus benthicfeeders, are relatively common throughout theirrange of distribution (Skúlason et al., 1992, 1999;Snorrason & Skúlason, 2004; Klemetsen, 2010).Such morphs have been most extensively studiedin Loch Rannoch, Scotland (Adams et al., 1998),Fjellfröstvatn, Norway (Knudsen et al., 2006), andThingvallavatn, Iceland (Skúlason et al., 1989a;Skúlason, Noakes & Snorrason, 1989b). In Thingval-lavatn, four morphs of charr have been observed, andsympatric morphs have been documented in numer-ous other Icelandic lakes (Skúlason et al., 1992). Phe-notypic diversity among allopatric charr populations
is also extensive in Iceland (Skúlason et al., 1992). Astriking aspect of this diversity is the common occur-rence of small benthic charr, associated with lava andgroundwater spring habitats (Sigursteinsdóttir &Kristjánsson, 2005; Kristjánsson, 2008, Klemetsen,2010). Genetic studies suggest that Icelandic Arcticcharr populations, both sympatric and allopatric,have evolved independently, and that genetic mixingof populations after colonization has been limited(Gíslason et al., 1999; Wilson et al., 2004; Kapralovaet al., 2011). This offers important opportunity tostudy the effects of local ecology on evolutionary pat-terns and processes.
It has been suggested that the great diversity ofIcelandic Arctic charr can be related to the diversity offreshwater habitats in Iceland (Malmquist et al., 1992;Skúlason et al., 1999; Snorrason & Skúlason, 2004),which can be linked to geological, hydrological, andgeochemical features of the island (Garðarsson, 1979;Malmquist et al., 2000; Karst-Riddoch, Malmquist &Smol, 2009). Long-term ‘plate-forming’ tectonic activ-ity has created an age gradient in the bedrock ofIceland. The youngest rocks are within the activeneo-volcanic zone, sometimes being only a few monthsold, whereas the oldest rocks (10–16 million years old)are furthest to the west and east (Pétursson &Jónsson, 2004) (Fig. 1). Older bedrock areas havebeen shaped by glaciers, are relatively impermeableto water, and are dominated by direct run-offwater bodies, characterized by a variable water regimeand temperature and low concentrations ofnutrients (Garðarsson, 1979; Malmquist et al., 2000;Karst-Riddoch et al., 2009). The neo-volcanic zone is,however, characterized by post-glacial lava fields. Thisbedrock is porous, which makes it rich in groundwaterand springs are common. Spring-fed water bodies arecharacterized by a relatively stable water regime andtemperature and also are rich in nutrients (Cantonati,Gerecke & Bertuzzi, 2006; Karst-Riddoch et al.,2009). Furthermore, lava provides a complex three-dimensional substrate, creating unique benthic habi-tats for fishes. Thus, invertebrates and fish haveextensive hiding places and are often found in highdensities (Sandlund et al., 1992; Snorrason et al., 1994;Malmquist et al., 2000; Kristjánsson, 2008).
Despite the geographic diversity of Icelandic fresh-water systems, their biological communities arerelatively simple. The short time since glaciation,together with geographic isolation, may explain whyonly six freshwater fish species are found on theisland (Sæmundsson, 1926; Albert, Jónsson & Ber-natchez, 2006). Arctic charr was one of the first fishspecies to colonize water bodies in Iceland, which tookplace rapidly after the ice retreated (Wilson et al.,2004; Kapralova et al., 2011). In many drainagesystems, charr must have colonized before isostatic
762 B. K. KRISTJÁNSSON ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, 103, 761–771

rebound and the formation of waterfalls closed offinland habitats from fish passage. The apparent colo-nization histories have resulted in a variable speciescomposition in Icelandic lakes, which could haveprofound effect on lake ecology.
In general, ecological drivers of parallel evolutionamong animal species are not well documented. Thesefactors are not easily approached by experimentalstudies and comparative studies require data from alarge number of populations. Icelandic Arctic charrpopulations are ideal for such a study because numer-ous populations are easily accessible and data onecological factors in their respective habitats havealready been collected by The Ecological Surveyof Icelandic Lakes (ESIL Project) (Malmquist et al.,2000; Karst-Riddoch et al., 2009). This project hascreated a comprehensive database that includesthe chemical and physiological characteristics oflakes, invertebrate species numbers, and densities inpelagic and benthic habitats, as well as biological
information on fish populations. The ESIL databaseprovides considerable analytical power for assessingthe relationship between phenotypic diversity ofArctic charr and the ecological and physical factorsof their habitat. In the present study, we test thehypothesis that morphological patterns among mono-morphic Arctic charr populations from different lakesin Iceland are related to interlake variation inecological characteristics.
MATERIAL AND METHODS
The lakes in the present study were all included in theESIL Project. Detailed information on the sampling inthe ESIL Project is given by Jónsdóttir et al. (1998),Malmquist et al. (2000) and Karst-Riddoch et al.(2009). For the present study, it was necessary toidentify lakes that had monomorphic charr popula-tions. This was carried out by examining publishedrecords of charr in Iceland, by talking to fisheries
Ónefnt vatn
Hlíðarvatn
Vatnsholtsvatn Oddastaðavatn
Hítarvatn
Geitabergsvatn
GlammastaðavatnEyrarvatn
ElliðavatnVífilsstaðavatnApavatn
Vesturhópsvatn
V-Friðmundarvatn
ArnarvatnÚlfsvatn
Mjóavatn
Ölvesvatn
Selvatn
Ásbjarnarvatn Syðra
Kötluvatn
SigurðarstaðavatnHraunhafnarfatn
Y-Deildarvatn
Nýpslón
Þuríðarvatn
Sænautavatn
Ánavatn
Urriðavatn
Heiðarvatn
Skriðuvatn
Eiðavatn
Langisjór
Langavatn
Frostastaðavatn
E-Gíslholtsvatn
Figure 1. Lakes sampled in a study on the relationship between ecological characters and the morphology of monomor-phic Arctic charr populations. The greyscale image shows the age of the bedrock; light grey, furthest to east and westupper Tertiary (> 3.3 Myr); mid-grey, upper Pliocene and lower Pleistocene (0.8–3.3 Myr); dark grey, upper Pleistocene(0.01–0.8 Myr); and darkest (nearest to the center), postglacial (0–0.01 Myr).
LAKE ECOLOGY AND MORPHOLOGY IN CHARR 763
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, 103, 761–771

specialists, and by examination of the age and lengthrelationships in relation to sexual maturation, photo-graphs of fish, and differences in diet among fish inthose lakes that did not have records of morphs. In thepresent study, we used only lakes where there wereclear indications of only one morph. Out of the 59lakes surveyed by the ESIL project, we used 35 lakesfor the analysis (Fig. 1) because other lakes either hadtoo small sample size or appeared to have more thanone morph (18 lakes). For each lake, either all theArctic charr, or a random sample from the catch toobtain a minimum of 30 fish, were used for furtherstudy. Each fish was photographed in the field (on theleft side), using a film camera (a few different modelswere used), under natural light conditions. Forklength (to the nearest 0.1 cm) and wet weight (to thenearest 0.5 g) were measured for all fish and stomachcontents were preserved for diet analysis. Otolithswere removed for age determination, and fish weredissected to determine sex and gonad maturationstatus. Stomach contents were stored in 5% bufferedformalin for subsequent identification and analysis.
Photographs of fish were scanned in high resolution(Epson Perfection V700 Photo; Bergvík). Using theTPSDIG software (developed by F. James Rohlf;http://life.bio.sunysb.edu/morph), 22 landmarks weremarked on each digital image. Six of these landmarkswere sliding landmarks, and the remainder werefixed landmarks (Fig. 2). The sliding landmarks com-prise landmarks that are allowed to slide to the left orright along a curve to minimize the shape changebetween the procrustes average of all the specimensand each specimen. Before analyzing morphology,caudal up and down bending of specimens was cor-rected for using the ‘unbend’ module in TPSUTILsoftware. The software creates a line between land-marks on the snout, at the end of caudal peduncle,and in the fork of the caudal fin. When the fish arebent, this line is curved. The procedure recalculatesthe position of landmarks on each fish so that the lineis straightened. Relative warp analysis in TPS-RELWwas used to analyze differences in morphology, at thesame time as controlling for geometric body size. Thisanalysis scales the landmarks from each fish to acentroid configuration (mean shape), position, and
rotation. The software then defines principal warpsfrom the centroid configuration, which are axes alongwhich shape variation can occur. Partial warps andtwo uniform components are then calculated andinclude a score for each fish (weight matrix) thatdescribes the realized amount of bending and stretch-ing necessary for the configuration of an individual tofit the centroid configuration. The weight matrix wasused in further analyses. TPSREGR and TPSSPLINEwere used to visualize morphological changes.
Data on ecological characters in each lake wereobtained from the ESIL project (Malmquist et al.,2000; Karst-Riddoch et al., 2009). Lakes were classi-fied into four groups (water origin, bedrock, salmonidcommunity, stickleback presence) based on noncon-tinuous variables. The first group was based on theorigin of the water, where 1 indicates spring fed, 2indicates direct run off, 3 indicates a mixture of1 and 2, and 4 indicates coastal. The second groupwas based on the age of predominant bedrock inthe lake catchments, where 1 indicates historic[< 0.0011 Myr)], 2 indicates postglacial (0.0011–0.01 Myr), 3 indicates upper Pleistocene (0.01–0.8 Myr), 4 indicates upper Pliocene and lowerPleistocene (0.8–3.3 Myr), and 5 indicates Upper Ter-tiary (> 3.3. Myr). The third group was based on thediversity of salmonids in each lake where category 1indicates lakes with only Arctic charr, 2 indicateslakes with Arctic charr and brown trout, and 3 indi-cates lakes with Arctic charr, brown trout and Atlan-tic salmon. The fourth group was lakes with orwithout sticklebacks. Information of continuous vari-ables expressing physical characters of each lakewere also used (Table 1).
Discriminant function analysis (DFA) on the weightmatrix was used to determine whether fish could besorted based on the lake of their origin. We also usedDFA to determine whether fish from different lakegroups could be separated. Multivariate analysisof variance (MANOVA) on the weight matrix wasused to determine whether there were morphologicaldifferences among Arctic charr coming from differentlake groups (see above). The average morphologywithin each population was calculated from theweight matrix and the distribution of populationswas examined using nonmetric scaling (NMS)(Kruskal, 1964), applying Sörensen distance measureand random starting configuration, in the softwarePCORD 5 (McCune & Mefford, 2006). The numbers ofaxis were not pre-assigned but allocated by the soft-ware based on calculated stress and instability. Thesoftware ran the analysis 50 times using the datasetand 50 times with random data from the dataset.Environmental variables were correlated to theNMS ordination axes using Pearson correlation andoverlaid on the ordination plots.
Figure 2. Landmarks digitized on the left side of Arcticcharr in the present study. Five landmarks were slidinglandmarks, shown in grey.
764 B. K. KRISTJÁNSSON ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, 103, 761–771
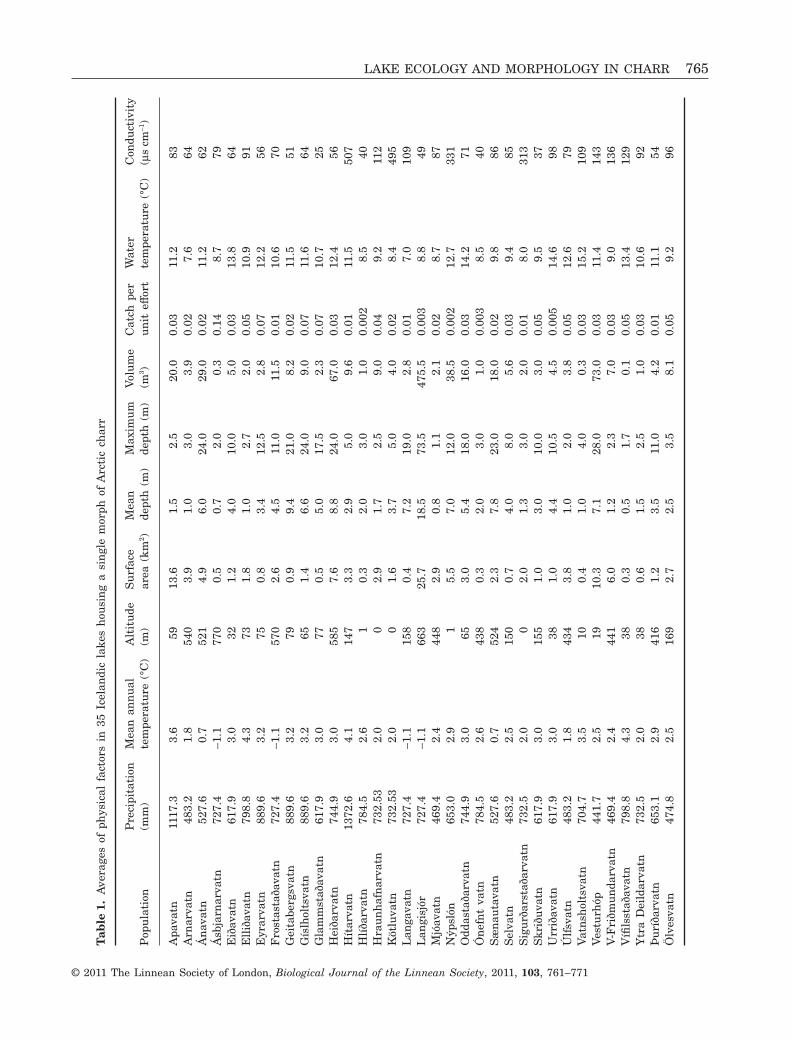
Tab
le1.
Ave
rage
sof
phys
ical
fact
ors
in35
Icel
andi
cla
kes
hou
sin
ga
sin
gle
mor
phof
Arc
tic
char
r
Pop
ula
tion
Pre
cipi
tati
on(m
m)
Mea
nan
nu
alte
mpe
ratu
re(°
C)
Alt
itu
de(m
)S
urf
ace
area
(km
2 )M
ean
dept
h(m
)M
axim
um
dept
h(m
)V
olu
me
(m3 )
Cat
chpe
ru
nit
effo
rtW
ater
tem
pera
ture
(°C
)C
ondu
ctiv
ity
(ms
cm–1
)
Apa
vatn
1117
.33.
659
13.6
1.5
2.5
20.0
0.03
11.2
83A
rnar
vatn
483.
21.
854
03.
91.
03.
03.
90.
027.
664
Án
avat
n52
7.6
0.7
521
4.9
6.0
24.0
29.0
0.02
11.2
62Á
sbja
rnar
vatn
727.
4-1
.177
00.
50.
72.
00.
30.
148.
779
Eið
avat
n61
7.9
3.0
321.
24.
010
.05.
00.
0313
.864
Ell
iðav
atn
798.
84.
373
1.8
1.0
2.7
2.0
0.05
10.9
91E
yrar
vatn
889.
63.
275
0.8
3.4
12.5
2.8
0.07
12.2
56F
rost
asta
ðava
tn72
7.4
-1.1
570
2.6
4.5
11.0
11.5
0.01
10.6
70G
eita
berg
svat
n88
9.6
3.2
790.
99.
421
.08.
20.
0211
.551
Gís
lhol
tsva
tn88
9.6
3.2
651.
46.
624
.09.
00.
0711
.664
Gla
mm
stað
avat
n61
7.9
3.0
770.
55.
017
.52.
30.
0710
.725
Hei
ðarv
atn
744.
93.
058
57.
68.
824
.067
.00.
0312
.456
Hít
arva
tn13
72.6
4.1
147
3.3
2.9
5.0
9.6
0.01
11.5
507
Hlí
ðarv
atn
784.
52.
61
0.3
2.0
3.0
1.0
0.00
28.
540
Hra
un
haf
nar
vatn
732.
532.
00
2.9
1.7
2.5
9.0
0.04
9.2
112
Köt
luva
tn73
2.53
2.0
01.
63.
75.
04.
00.
028.
449
5L
anga
vatn
727.
4-1
.115
80.
47.
219
.02.
80.
017.
010
9L
angi
sjór
727.
4-1
.166
325
.718
.573
.547
5.5
0.00
38.
849
Mjó
avat
n46
9.4
2.4
448
2.9
0.8
1.1
2.1
0.02
8.7
87N
ýpsl
ón65
3.0
2.9
15.
57.
012
.038
.50.
002
12.7
331
Odd
asta
ðarv
atn
744.
93.
065
3.0
5.4
18.0
16.0
0.03
14.2
71Ó
nef
nt
vatn
784.
52.
643
80.
32.
03.
01.
00.
003
8.5
40S
æn
auta
vatn
527.
60.
752
42.
37.
823
.018
.00.
029.
886
Sel
vatn
483.
22.
515
00.
74.
08.
05.
60.
039.
485
Sig
urð
arst
aðar
vatn
732.
52.
00
2.0
1.3
3.0
2.0
0.01
8.0
313
Skr
iðu
vatn
617.
93.
015
51.
03.
010
.03.
00.
059.
537
Urr
iðav
atn
617.
93.
038
1.0
4.4
10.5
4.5
0.00
514
.698
Úlf
svat
n48
3.2
1.8
434
3.8
1.0
2.0
3.8
0.05
12.6
79V
atn
shol
tsva
tn70
4.7
3.5
100.
41.
04.
00.
30.
0315
.210
9V
estu
rhóp
441.
72.
519
10.3
7.1
28.0
73.0
0.03
11.4
143
V-F
riðm
un
darv
atn
469.
42.
444
16.
01.
22.
37.
00.
039.
013
6V
ífils
stað
avat
n79
8.8
4.3
380.
30.
51.
70.
10.
0513
.412
9Y
tra
Dei
ldar
vatn
732.
52.
038
0.6
1.5
2.5
1.0
0.03
10.6
92Þ
urí
ðarv
atn
653.
12.
941
61.
23.
511
.04.
20.
0111
.154
Ölv
esva
tn47
4.8
2.5
169
2.7
2.5
3.5
8.1
0.05
9.2
96
LAKE ECOLOGY AND MORPHOLOGY IN CHARR 765
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, 103, 761–771
RESULTS
Landmarks were digitized on 1220 pictures of Arcticcharr from 35 populations (Fig. 1, Table 1). The DFAwas significant (Wilks lambda � 0.0001, c2
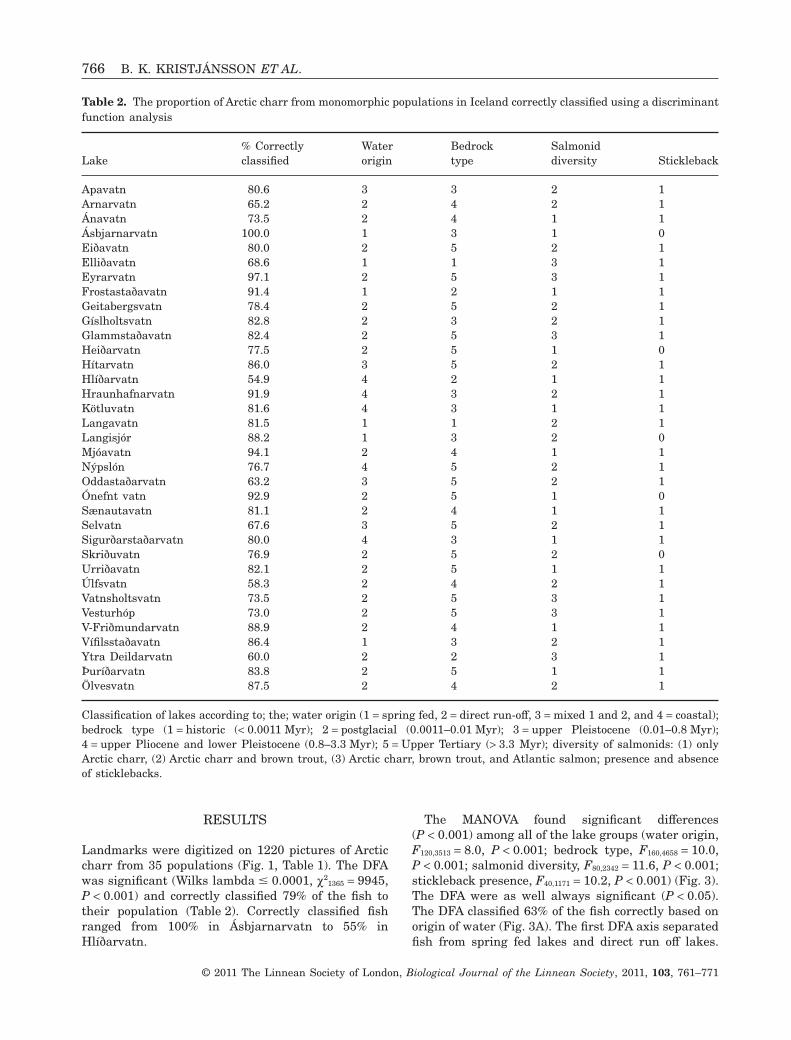
1365 = 9945,P < 0.001) and correctly classified 79% of the fish totheir population (Table 2). Correctly classified fishranged from 100% in Ásbjarnarvatn to 55% inHlíðarvatn.
The MANOVA found significant differences(P < 0.001) among all of the lake groups (water origin,F120,3513 = 8.0, P < 0.001; bedrock type, F160,4658 = 10.0,P < 0.001; salmonid diversity, F80,2342 = 11.6, P < 0.001;stickleback presence, F40,1171 = 10.2, P < 0.001) (Fig. 3).The DFA were as well always significant (P < 0.05).The DFA classified 63% of the fish correctly based onorigin of water (Fig. 3A). The first DFA axis separatedfish from spring fed lakes and direct run off lakes.
Table 2. The proportion of Arctic charr from monomorphic populations in Iceland correctly classified using a discriminantfunction analysis
Lake% Correctlyclassified
Waterorigin
Bedrocktype
Salmoniddiversity Stickleback
Apavatn 80.6 3 3 2 1Arnarvatn 65.2 2 4 2 1Ánavatn 73.5 2 4 1 1Ásbjarnarvatn 100.0 1 3 1 0Eiðavatn 80.0 2 5 2 1Elliðavatn 68.6 1 1 3 1Eyrarvatn 97.1 2 5 3 1Frostastaðavatn 91.4 1 2 1 1Geitabergsvatn 78.4 2 5 2 1Gíslholtsvatn 82.8 2 3 2 1Glammstaðavatn 82.4 2 5 3 1Heiðarvatn 77.5 2 5 1 0Hítarvatn 86.0 3 5 2 1Hlíðarvatn 54.9 4 2 1 1Hraunhafnarvatn 91.9 4 3 2 1Kötluvatn 81.6 4 3 1 1Langavatn 81.5 1 1 2 1Langisjór 88.2 1 3 2 0Mjóavatn 94.1 2 4 1 1Nýpslón 76.7 4 5 2 1Oddastaðarvatn 63.2 3 5 2 1Ónefnt vatn 92.9 2 5 1 0Sænautavatn 81.1 2 4 1 1Selvatn 67.6 3 5 2 1Sigurðarstaðarvatn 80.0 4 3 1 1Skriðuvatn 76.9 2 5 2 0Urriðavatn 82.1 2 5 1 1Úlfsvatn 58.3 2 4 2 1Vatnsholtsvatn 73.5 2 5 3 1Vesturhóp 73.0 2 5 3 1V-Friðmundarvatn 88.9 2 4 1 1Vífilsstaðavatn 86.4 1 3 2 1Ytra Deildarvatn 60.0 2 2 3 1Þuríðarvatn 83.8 2 5 1 1Ölvesvatn 87.5 2 4 2 1
Classification of lakes according to; the; water origin (1 = spring fed, 2 = direct run-off, 3 = mixed 1 and 2, and 4 = coastal);bedrock type (1 = historic (< 0.0011 Myr); 2 = postglacial (0.0011–0.01 Myr); 3 = upper Pleistocene (0.01–0.8 Myr);4 = upper Pliocene and lower Pleistocene (0.8–3.3 Myr); 5 = Upper Tertiary (> 3.3 Myr); diversity of salmonids: (1) onlyArctic charr, (2) Arctic charr and brown trout, (3) Arctic charr, brown trout, and Atlantic salmon; presence and absenceof sticklebacks.
766 B. K. KRISTJÁNSSON ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, 103, 761–771

In general, fish from spring feed lakes were deeperbodied and had somewhat smaller heads. Axis 2 sepa-rated fish from coastal lakes from the other lakes. Incoastal lakes, fish were in general thinner bodied,with more straight pointing mouth. The DFA correctlyclassified 58% of the fish based on bedrock (Fig. 3B).DFA axis 1 roughly separated the fish based on age ofthe bedrock with fish from lakes in historic bedrockhaving the highest scores. Axis 2 separated fish fromlakes in postglacial bedrock from fish in lakes inupper Tertiary bedrock. The most pronounced body
shape differences seen here were in relation to headshape and mouth orientation, where fish from theyounger lakes had somewhat smaller heads andthinner bodies. The DFA correctly classified 67%of the fish correctly based on salmonid diversity(Fig. 3C). Axis 1 separated fish from lakes whereArctic charr was the only salmonid present (lowscores) from lakes that also had other salmonids.Arctic charr in lakes where there were other sal-monids had deeper bodies, short and thick caudalpeduncle, and somewhat smaller heads compared
Figure 3. Distribution of discriminant scores from an discriminant function analysis of Arctic charr from 35 monomor-phic Arctic charr populations in Iceland. The symbols denote population average and the bars denote 1 standard error.A, the discriminant analysis separated fish from lakes with different water origin: spring fed (open circles), direct run-off(filled triangles), mixture of spring and run-off (open diamonds), and coastal (filled squares). B, fish from lakes in bedrockof different age, the age were 1 (x) = historic (< 0.0011 Myr); 2 (filled squares) = postglacial (0.0011–0.01 Myr); 3 (opencircle) = upper Pleistocene (0.01–0.8 Myr); 4 (filled triangle) = upper Pliocene and lower Pleistocene (0.8–3.3 Myr); and 5(open diamond) = Upper Tertiary (> 3.3. Myr). C, the symbols denote population average and the bars denote one standarderror. The discriminant analysis separated fish from lakes having different salmonid species compositions: 1(diamonds) = only Arctic charr; 2 (triangles) = Arctic charr and brown trout; and 3 (squares) = Arctic charr, brown trout,and Atlantic salmon. D, the discriminant analysis separated fish from lakes with sticklebacks (open triangles) and thosewithout sticklebacks (filled triangles). Deformation grids show the morphology of an average fish in a given lake comparedto the average fish from all the lakes, shown at ¥3 magnification.
LAKE ECOLOGY AND MORPHOLOGY IN CHARR 767
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, 103, 761–771
to individuals in lakes where Arctic charr occur alone.Axis 2 roughly separated lakes where all threespecies of salmonids were present from lakes wherethere were either only Arctic charr or Arctic charr andbrown trout. Arctic charr in lakes where there weresalmon present had longer caudal fin. The DFA cor-rectly classified 82% of the fish based on the presenceor absence of sticklebacks (Fig. 3D). Arctic charr inlakes where there were no sticklebacks had narrowerbodies, and smaller heads with more subterminalmouth shape than those coming from lakes withsticklebacks.
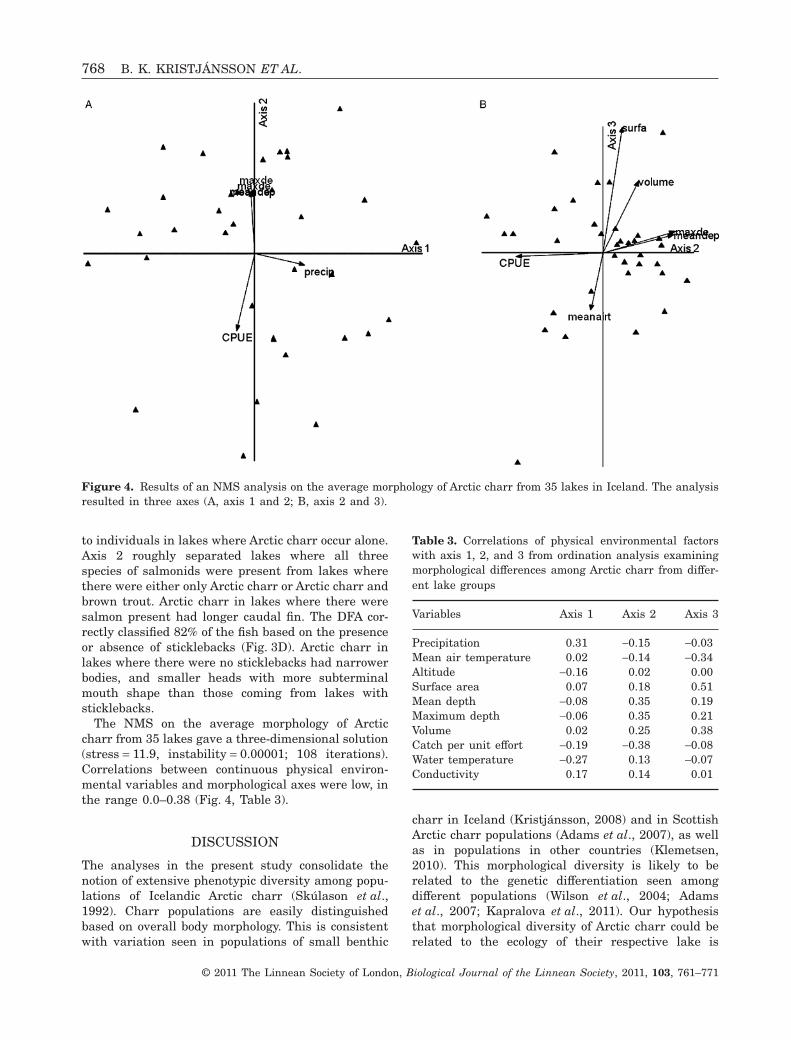
The NMS on the average morphology of Arcticcharr from 35 lakes gave a three-dimensional solution(stress = 11.9, instability = 0.00001; 108 iterations).Correlations between continuous physical environ-mental variables and morphological axes were low, inthe range 0.0–0.38 (Fig. 4, Table 3).
DISCUSSION
The analyses in the present study consolidate thenotion of extensive phenotypic diversity among popu-lations of Icelandic Arctic charr (Skúlason et al.,1992). Charr populations are easily distinguishedbased on overall body morphology. This is consistentwith variation seen in populations of small benthic
charr in Iceland (Kristjánsson, 2008) and in ScottishArctic charr populations (Adams et al., 2007), as wellas in populations in other countries (Klemetsen,2010). This morphological diversity is likely to berelated to the genetic differentiation seen amongdifferent populations (Wilson et al., 2004; Adamset al., 2007; Kapralova et al., 2011). Our hypothesisthat morphological diversity of Arctic charr could berelated to the ecology of their respective lake is
Table 3. Correlations of physical environmental factorswith axis 1, 2, and 3 from ordination analysis examiningmorphological differences among Arctic charr from differ-ent lake groups
Variables Axis 1 Axis 2 Axis 3
Precipitation 0.31 -0.15 -0.03Mean air temperature 0.02 -0.14 -0.34Altitude -0.16 0.02 0.00Surface area 0.07 0.18 0.51Mean depth -0.08 0.35 0.19Maximum depth -0.06 0.35 0.21Volume 0.02 0.25 0.38Catch per unit effort -0.19 -0.38 -0.08Water temperature -0.27 0.13 -0.07Conductivity 0.17 0.14 0.01
Figure 4. Results of an NMS analysis on the average morphology of Arctic charr from 35 lakes in Iceland. The analysisresulted in three axes (A, axis 1 and 2; B, axis 2 and 3).
768 B. K. KRISTJÁNSSON ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, 103, 761–771

supported. Diverse morphology of Arctic charr fromdifferent lakes was associated with water origin,age of the surrounding bedrock, and the respectivefish communities. Strongest relationships were foundwith structure of fish communities. However, whenwe related continuous ecological characters with mor-phology, the results were much less clear, probablybecause these variables show considerable variationamong lakes within the same lake group.
In general, the relationship between the fish mor-phology and lake ecology suggests that ecologicalcharacters contribute to morphological evolution. Therelatively strong relationship between charr morphol-ogy and structure of fish communities in lakes sug-gests that competition and/or predation may play animportant role in shaping the observed morphologicaldiversity, as has been observed in many fish popu-lations (Schluter, 2000; Eklöv & Svanbäck, 2006),including charr (Knudsen, Amundsen & Klemetsen,2010). Morphological differences were seen in headshape and traits related to locomotion, such as theshape of the caudal peduncle, as well as size andposition of fins. In lakes where Arctic charr was theonly fish species, individuals had relatively shallowerbodies and smaller heads than in other lakes, whichindicates relatively poorer somatic condition. Thiscould result from larger population size of charr insuch lakes and thus high intraspecific competition(Klemetsen et al., 2002). In lakes where charr occurwith brown trout, competition with brown trout maycause habitat shift in charr, resulting in the observedphenotypic diversity. Studies have shown thatbrown trout may displace Arctic charr of similar sizefrom more rewarding areas in shallow waters in thelittoral zone to deeper waters (Hegge et al., 1989;Langeland et al., 1991; Jansen et al., 2002). Similarly,charr might compete with threespine stickleback forresources, which might result in the observed mor-phological differences. An alternative explanation isthat in lakes with other salmonids species deeper-bodied charr may show plastic response to predation,most likely by brown trout (Brönmark & Miner, 1992;Brönmark & Pettersson, 1994).
There were clear morphological differences amongcharr from lakes on different aged bedrock and lakesthat differed in water origin. These two ecologicalgroups are obviously not independent (Garðarsson,1979; Sigurðsson, 1993). Older bedrock is less per-meable to water and direct run-off systems are morecommon there than in areas of younger bedrockwhere spring-fed systems are prominent. Lakes situ-ated in areas of younger bedrock appear to generate amore ‘benthic’ morphotype. These charr have deeperbodies, a shorter and thicker caudal peduncle, andshorter heads, which are morphological differencesthat can be related to more benthic foraging (Malm-
quist, 1992; Malmquist et al., 1992; Skúlason et al.,1999). These lakes are often characterized by exten-sive benthic lava habitats, offering high densities ofbenthic invertebrate prey (Malmquist et al., 2000),and are ideal environments for juvenile charr withample spaces for hiding and avoiding predation (Snor-rason & Skúlason, 2004)
It is likely that a considerable part of the observedmorphological diversity has been caused by indepen-dent evolutionary processes in each lake (Gíslasonet al., 1999; Wilson et al., 2004; Kapralova et al., 2011),where factors such as founder effect and genetic driftmight be involved (Langerhans & DeWitt, 2004).However, we observed direct relationships betweencharr morphology and the ecological characteristics ofresepective lakes. We propose two mechanisms toexplain these patterns. The first is a result of adaptivedivergence where similar ecological factors promotesimilar evolutionary responses in different popula-tions. The other mechanism reflects a process wheremorphological patterns are shaped by repeated plasticresponses in similar phenotypic characters. It is wellestablished that fish can respond in a plastic way todiet (Day, Pritchard & Schluter, 1994) and, in somecases, substantial morphological differences betweenmorphs or even species are almost entirely the result ofdiet-induced plasticity (Wimberger, 1992, 1994). Thediverse diet of the Arctic charr populations in thepresent study is currently being analyzed in detail(Woods P. J. pers.observ) allowing for a future study ofthe relationship between morphology and dietary-induced plasticity. Plastic responses to predation arealso well known. For example, fish exposed to water-borne cues from predators develop deeper bodies. Thishas been well established in crucian carp (Carassiuscarassius) preyed on by pike (Esox lucius) (Brönmark& Miner, 1992; Brönmark & Pettersson, 1994). Arcticcharr is known to be very plastic (Skúlason et al., 1999;Snorrason & Skúlason, 2004), and studies have alsodemonstrated that populations differ in the level ofplastic responses (Skúlason et al., 1989a, b; Adams &Huntingford, 2002, 2004; Klemetsen et al., 2002; Snor-rason & Skúlason, 2004; Kristjánsson, 2008; Parsons,2008; Parsons, Skúlason & Ferguson, 2010). Thus, thelevel of plasticity in small benthic Arctic charr hasbeen found to be related to ecological factors in springs(Kristjánsson, 2008). Therefore, the two potentialmechanisms (i.e. selection and plasticity) contributingto parallel morphological patterns in Arctic charr arenot mutually exclusive and are likely additive.
Understanding the origin and maintenance ofbiological diversity is a key question in biology.In recent years, scientists have increasingly realizedthat there is a strong relationship between ecolo-gical dynamics and micro-evolutionary processes.Evolutionary changes in populations can affect the
LAKE ECOLOGY AND MORPHOLOGY IN CHARR 769
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, 103, 761–771

ecosystems they live in, and ecological changes cansimilarly affect the evolutionary processes of popula-tions (Fussmann, Loreau & Abrams, 2007). Studies ofthe relationship between ecological factors and evolu-tionary changes, comparing traits in related popula-tions, provide a powerful way of examining parallelevolution and its mechanisms. However, the strengthof this approach is directly related to the number ofpopulations compared. Therefore, studies of this kindare relatively few, especially when comparing largevertebrates such as fish. In the present study, wehave used the availability of detailed ecological andmorphological information from the Icelandic ESILdatabase to uncover relationships between morpho-logical diversity of Arctic charr in Icelandic lakes andtheir ecology. The findings of study show that parallelevolution and natural selection may be important forthe origin of intraspecific biological diversity. Thesefindings set the stage for continuing studies thataddress more detailed questions on the nature of thisdiversity and its origin.
ACKNOWLEDGMENTS
We wish to thank the numerous people that helpedwith gathering of data in the ESIL project. The manu-script has been improved by valuable comments byDavid L. G. Noakes, Joseph D. Ackerman, PamelaWoods, Camille Leblanc, and three anonymous refer-ees. The ESIL project was funded by a grant fromRannís. This paper is a part of the PhD study ofBjarni K. Kristjánsson at the University of Guelph,Canada. Bjarni K. Kristjánsson was funded by agraduate student grant from Rannís, Hólar Univer-sity College and by the Brock Doctoral Scholarship atthe University of Guelph.
REFERENCES
Adams CE, Huntingford FA. 2002. The functional signifi-cance of inherited differences in feeding morphology in asympatric polymorphic population of Arctic charr. Evolu-tionary Ecology 16: 15–25.
Adams CE, Huntingford FA. 2004. Incipient speciationdriven by phenotypic plasticity? Evidence from sympa-tric populations of Arctic charr. Biological Journal of theLinnean Society 81: 611–618.
Adams CE, Fraser D, Huntingford FA, Greer R, AskewCM, Walker A. 1998. Trophic polymorphism among arcticcharr from Loch Rannoch, Scotland. Journal of Fish Biology52: 1259–1271.
Adams CE, Fraser D, Wilson AJ, Alexander G, FergusonMM, Skulason S. 2007. Patterns of phenotypic and geneticvariability show hidden diversity in Scottish Arctic charr.Ecology of Freshwater Fish 16: 78–86.
Albert V, Jónsson B, Bernatchez L. 2006. Natural hybridsin Atlantic eels (Anguilla anguilla, A. rostrata): evidence for
successful reproduction and fluctuating abundance in spaceand time. Molecular Ecology 15: 1903–1916.
Brönmark C, Miner JG. 1992. Predator-induced phenotypi-cal change in body morphology in crucian carp. Science 258:1348–1350.
Brönmark C, Pettersson LB. 1994. Chemical cues frompiscivores induce a change in morphology in crucian carp.Oikos 70: 396–402.
Cantonati M, Gerecke R, Bertuzzi E. 2006. Springs ofthe Alps – sensitive ecosystems to environmental change:from biodiversity assessments to long-term studies.Hydrobiologia 562: 59–96.
Day T, Pritchard J, Schluter D. 1994. A comparison of twostickleback. Evolution 48: 1723–1734.
Eklöv P, Svanbäck R. 2006. Predation risk influences adap-tive morphological variation in freshwater fish populations.American Naturalist 167: 440–452.
Fussmann GF, Loreau M, Abrams PA. 2007. Eco-evolutionary dynamics of communities and ecosystems.Functional Ecology 21: 465–477.
Garðarsson A. 1979. Vistfræðileg flokkun íslenskra vatna.[Ecological classification of Icelandic lakes]. Týli 9: 1–10.
Gíslason D, Ferguson MM, Skúlason S, Snorrason SS.1999. Rapid and coupled phenotypic and genetic divergencein Icelandic Arctic charr (Salvelinus alpinus). CanadianJournal of Fisheries and Aquatic Sciences 56: 2229–2234.
Grant PR. 1999. Ecology and evolution of Darwin’s finches.Princeton, NJ: Princeton Uni-Versity Press.
Hegge O, Dervo BK, Skurdal J, Hessen DO. 1989. Habitatutilization by sympatric arctic charr Salvelinus alpinus L.and brown trout Salmo trutta L. in Lake Atnsjo, south-eastNorway. Freshwater Biology 22: 143–152.
Jansen PA, Slettvold H, Finstad AG, Langeland A. 2002.Niche segregation between Arctic char (Salvelinus alpinus)and brown trout (Salmo trutta): an experimental study ofmechanisms. Canadian Journal of Fisheries and AquaticSciences 59: 6–11.
Jónsdóttir H, Malmquist HJ, Snorrason SS,Guðbergsson G, Guðmundsdóttir S. 1998. Epidemiologyof Renibacterium salmoninarum in wild Arctic charr andbrown trout in Iceland. Journal of Fish Biology 53: 322–339.
Kapralova KH, Morrissey MB, Kristjánsson BK, Ólafs-dóttir GA, Snorrason SS, Ferguson MM. 2011. Evolutionof adaptive diversity and genetic connectivity in Arctic charr(Salvelinus alpinus) in Iceland. Hereditity 106: 472–487.
Karst-Riddoch TL, Malmquist HJ, Smol JP. 2009.Relationships between freshwater sedimentary dia-toms and environmental variables in Subarctic Icelandiclakes. Fundamental and Applied Limnology (Archive furHydrobiologie) 175: 1–28.
Klemetsen A. 2010. The charr problem revisited: exceptionalphenotypic plasticity promotes ecological speciation in post-glacial lakes. Freshwater Reviews 3: 49–74.
Klemetsen A, Amundsen P, Grotnes PE, Knudsen R,Kristoffersen R, Svenning M-A. 2002. Takvatn through20 years: long-term effects of an experimental mass removalof Arctic charr, Salvelinus alpinus, from a subarctic lake.Enviromental Biology of Fishes 64: 39–47.
770 B. K. KRISTJÁNSSON ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, 103, 761–771

Knudsen R, Klemetsen A, Amundsen P-A, Hermansen B.2006. Incipient speciation through niche expansion: anexample from the arctic charr in a subarctic lake. Proceed-ings of the Royal Society of London Series B, BiologicalSciences 273: 2291–2298.
Knudsen R, Amundsen P-A, Klemetsen A. 2010. Arcticcharr in sympatry with burbot – ecological and evolutionaryconsequences. Hydrobiologica 650: 43–54.
Kristjánsson BK. 2008. Fine scale phenotypic diversity ofArctic charr (Salvelinus alpinus) in relation to ecologicalcharacters. PhD Thesis, University of Guelph.
Kruskal JB. 1964. Nonmetric multidimensional scaling: anumerical method. Psychometrika 29: 115–129.
Langeland A, L’Abée-Lund JH, Jonsson B, Jonsson N.1991. Resource partitioning and niche shift in Arctic charrSalvelinus alpinus and Brown trout Salmo trutta. Journalof Animal Ecology 60: 895–912.
Langerhans RB, DeWitt TJ. 2004. Shared and unique fea-tures of evolutionary diversification. American Naturalist164: 335–349.
Malmquist HJ. 1992. Phenotype-specific feeding behaviourof two Arctic charr Salvelinus alpinus morphs. Oecologia 3:354–361.
Malmquist HJ, Snorrason SS, Skúlason S, Sandlund OT,Jonsson B, Jónasson PM. 1992. Diet differentiationin polymorphic Arctic charr in Thingvallavatn, Iceland.Journal of Animal Ecology 61: 21–35.
Malmquist HJ, Antonsson T, Guðbergsson G, Skúlason S,Snorrason SS. 2000. Biodiversity of macroinvertebrates onrocky substrate in the surf zone of Icelandic lakes. Verhand-lungenInternational Vereinigung Limnology 27: 1–7.
McCune B, Mefford MJ. 2006. PC-ORD. Multivariate analy-sis of ecological data, Version 5.10. Gleneden Beach, OR:MjM Software.
Nagel L, Schluter D. 1998. Body size, natural selection, andspeciation in sticklebacks. Evolution 52: 209–218.
Parsons KJ. 2008. Phenotypic plasticity and the geneticbasis of adaptive divergence in Icelandic Arctic charr(Salvelinus alpinus). PhD Thesis, University of Guelph.
Parsons KJ, Skúlason S, Ferguson MM. 2010. Mor-phological variation over ontogeny and environments inresource polymorphic arctic charr (Salvelinus alpinus).Evolution and Development 12: 246–257.
Pétursson JI, Jónsson JG. 2004. Almenn jarðfræði. Iðnú:Reykjavík.
Price T. 2008. Speciation in birds. Greenwood Village, CO:Roberts and Company Publishers.
Reimchen TE, Nosil P. 2006. Replicated ecological land-scapes and the evolution of morphological diversity amongGasterosteus populations from an archipelago on the westcoast of Canada. Canadian Journal of Zoology 84: 643–654.
Robinson BW, Schluter D. 2000. Natural selection and theevolution of adaptive genetic variation in northern freshwa-ter fishes. In: Mosseau TA, Sinervo B, Endler JA, eds.Adaptive genetic variation in the wild. Oxford: OxfordUniversity Press.
Sæmundsson B. 1926. Fiskarnir (pisces islandiæ). Reykjavík:Bókaverslun Sigfúsar Eymundssonar.
Sandlund OT, Gunnarsson K, Jónasson PM, Jonsson B,Lindem T, Magnússon KP, Malmquist HJ, Sigurjóns-dóttir H, Skúlason S, Snorrason SS. 1992. The Arcticcharr Salvelinus alpinus in Thingvallavatn. Oikos 64: 305–351.
Schluter D. 2000. The ecology of adaptive radiation. Oxford:Oxford University Press.
Schluter D, Clifford EA, Nemethy M, McKinnon JS.2004. Parallel evolution and inheritance of quantitativetraits. The American Naturalist 163: 809–822.
Sigurðsson F. 1993. Groundwater chemistry and aquiferclassification in Iceland. In: Banks S, Banks D, eds. Hydrol-ogy of hard rocks. Oslo: International Association of Hydro-geologists Memories, XXIV. Part 1.
Sigursteinsdóttir RJ, Kristjánsson BK. 2005. Parallel evo-lution, not always so parallel: comparison of small benthiccharr, Salvelinus alpinus, from Grímsnes and Thingvallav-atn, Iceland. Environmental Biology of Fishes 74: 239–244.
Skúlason S, Snorrason SS, Noakes DLG, Ferguson MM,Malmquist HJ. 1989a. Segregation in spawning and earlylife history among polymorphic Arctic charr, Salvelinusalpinus, in Thingvallavatn, Iceland. Journal of Fish Biology35: 225–232.
Skúlason S, Noakes DLG, Snorrason SS. 1989b. Ontogenyof trophic morphology of four sympatric morphs of arcticcharr, Salvelinus alpinus in Thingvallavatn, Iceland. Bio-logical Journal of the Linnean Society 38: 281–301.
Skúlason S, Antonsson T, Guðbergsson G, MalmquistHJ, Snorrason SS. 1992. Variability in Icelandic Arcticcharr. Icelandic Agricultural Sciences 6: 143–153.
Skúlason S, Snorrason SS, Jónsson B. 1999. Sympatricmorphs, population and speciation in freshwater fish withemphasis on arctic charr. In: Magurran A, May R, eds.Evolution of biological diversity: from population to species.Oxford: Oxford University Press.
Snorrason SS, Skúlason S. 2004. Adaptive speciation innorthern freshwater fish – patterns and processes. In: Diek-mann U, Metz H, Doebeli M, Tautz D, eds. Adaptive spe-ciation. Cambridge: Cambridge University Press.
Snorrason SS, Skúlason S, Jonsson B, Malmquist HJ,Jónasson PM, Sandlund OT, Lindem T. 1994. Trophicspecialization in Arctic charr Salvelinus alpinus (Pisces:Salmonidae): morphological divergence and ontogeneticniche shifts. Biological Journal of the Linnean Society 52:1–18.
Wilson AJ, Gislason D, Skúlason S, Snorrason SS,Adams CE, Alexander G, Danzmann RG, FergusonMM. 2004. Population genetic structure of Arctic charr,Salvelinus alpinus from northwest Europe on large andsmall spatial scales. Molecular Ecology 13: 1129–1142.
Wimberger PH. 1992. Plasticity of fish body shape: the effectof diet, development, family and age in two species ofGeophagus (Pisces: Cichlidae). Biological Journal of theLinnean Society 45: 197–218.
Wimberger PH. 1994. Trophic polymorphism, plasticity, andspeciation in vertebrates. In: Stoner DJ, Fresh KL, FellerRJ, eds. Theory and application of fish feeding ecology. TheBelle W. Baruch Library in Marine Science series no 18.
LAKE ECOLOGY AND MORPHOLOGY IN CHARR 771
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, 103, 761–771
Copyright © 2022 FDOKUMEN


















