
Dhole (Cuon alpinus) as a bone acumulator and new taphonomic agent. The case of the Noisetier Cave...
34
305 Article JTa138 All rights reserved. *E-mail: [email protected] 2012 Journal of Taphonomy PROMETHEUS PRESS/PALAEONTOLOGICAL NETWORK FOUNDATION (TERUEL) VOLUME 10 (ISSUE 3-4) Available online at www.journaltaphonomy.com Noisetier Cave (French Pyrenees) has yielded Mousterian artefacts associated with numerous faunal remains. The faunal spectrum is dominated by chamois and ibex followed by red deer and bovids. A previous taphonomic analysis underlined the occurrence of two distinct types of bone accumulations. The red deer, bovid and a part of the ibex remains have been accumulated by Neanderthal. We suspected that the bearded vultures were responsible for the chamois and some of the ibex remains. The study of the carnivore remains illustrated the abundance of teeth and to a lesser extent bones attributed to both Dhole (Cuon alpinus) as a Bone Accumulator and New Taphonomic Agent? The Case of the Noisetier Cave (French Pyrenees) Jean-Baptiste Mallye* Université Bordeaux 1, PACEA, UMR5199 Avenue des Facultés, 33405 Talence, France Sandrine Costamagno CNRS TRACES, UMR 5608, Université Toulouse 2 – Le Mirail Maison de la Recherche, 5 allées A. Machado, 31058 Toulouse cedex 9, France Myriam Boudadi-Maligne Université Bordeaux 1, PACEA, UMR5199 Avenue des Facultés, 33405 Talence, France Audrey Prucca Les loups du Gévaudan, Sainte-Lucie, 48100 Saint-Léger-de-Peyre, France Véronique Lauroulandie Université Bordeaux 1, PACEA, UMR5199 Avenue des Facultés, 33405 Talence, France Céline Thiébaut UMR 5608 –TRACES, Maison de la Recherche, 5 allées A. Machado, 31058, Toulouse cedex 9, France Vincent Mourre UMR 5608 - TRACES et Inrap - Direction Interrégionale Méditerranée, 561, rue Étienne Lenoir - KM Delta, 30900 Nîmes, France Journal of Taphonomy 10 (3-4) (2012), 305-338. Manuscript received 15 March 2012, revised manuscript accepted 15 November 2012.
-
Upload
univ-tlse2 -
Category
Documents
-
view
0 -
download
0
Transcript of Dhole (Cuon alpinus) as a bone acumulator and new taphonomic agent. The case of the Noisetier Cave...

305
Mallye et al.
Article JTa138 All rights reserved. *E-mail: [email protected]
2012
Journal of Taphonomy
PROMETHEUS PRESS/PALAEONTOLOGICAL NETWORK FOUNDATION (TERUEL)
VOLUME 10 (ISSUE 3-4)
Available online at www.journaltaphonomy.com
Noisetier Cave (French Pyrenees) has yielded Mousterian artefacts associated with numerous faunal
remains. The faunal spectrum is dominated by chamois and ibex followed by red deer and bovids. A
previous taphonomic analysis underlined the occurrence of two distinct types of bone accumulations.
The red deer, bovid and a part of the ibex remains have been accumulated by Neanderthal. We suspected
that the bearded vultures were responsible for the chamois and some of the ibex remains. The study of
the carnivore remains illustrated the abundance of teeth and to a lesser extent bones attributed to both
Dhole (Cuon alpinus) as a Bone Accumulator and
New Taphonomic Agent?
The Case of the Noisetier Cave
(French Pyrenees)
Jean-Baptiste Mallye* Université Bordeaux 1, PACEA, UMR5199 Avenue des Facultés, 33405 Talence, France
Sandrine Costamagno
CNRS TRACES, UMR 5608, Université Toulouse 2 – Le Mirail Maison de la Recherche, 5
allées A. Machado, 31058 Toulouse cedex 9, France
Myriam Boudadi-Maligne Université Bordeaux 1, PACEA, UMR5199 Avenue des Facultés, 33405 Talence, France
Audrey Prucca
Les loups du Gévaudan, Sainte-Lucie, 48100 Saint-Léger-de-Peyre, France
Véronique Lauroulandie Université Bordeaux 1, PACEA, UMR5199 Avenue des Facultés, 33405 Talence, France
Céline Thiébaut
UMR 5608 –TRACES, Maison de la Recherche, 5 allées A. Machado,
31058, Toulouse cedex 9, France
Vincent Mourre UMR 5608 - TRACES et Inrap - Direction Interrégionale Méditerranée, 561, rue Étienne
Lenoir - KM Delta, 30900 Nîmes, France
Journal of Taphonomy 10 (3-4) (2012), 305-338.
Manuscript received 15 March 2012, revised manuscript accepted 15 November 2012.

306
Dhole as a bone accumulator at Noisetier Cave
published that characterise the bone
assemblages created by various carnivores.
Fewer studies have discussed the role of
large canids in the formation and
modification of bone assemblages or their
modifications (Binford, 1981; Haynes, 1983;
Klippel et al., 1987; Stiner, 1994, 2004;
Castel, 2004; Fosse et al., 2004; Campmas
& Beauval, 2008; Castel et al., 2010;
Esteban-Nadal et al., 2010; Yravedra, 2011).
The studies on bone destruction of
large and medium ungulates by large
carnivores are mainly inferred from kill
sites or scavenging sites (Haynes, 1980;
Richardson, 1980; Brain, 1981;
Blumenschine, 1988; Hill, 1989; Fosse,
1994; Nasti, 2000; Prucca, 2003; Castel,
2004; Fosse et al., 2004; Campmas &
Beauval, 2008). Most of the taphonomic
studies devoted to coproscopic analysis of
carnivore scats deal with small vertebrate
remains (Andrews & Nesbit-Evans, 1983;
Payne & Munson, 1985; Denys et al., 1992;
Denys, 2011; Andrews, 1990; Schmitt &
Juell, 1994; Mondini, 2000; Matthews,
2006; Lloveras et al., 2008a, b; Montalvo et
Introduction
Caves and rock shelters could have
alternatively been occupied by hominids and
carnivores. Consequently, stone tools and
faunal remains associated with occupants
could be mixed by post-depositional
processes (e.g. Villa & Bartram, 1996; Villa
& Soressi, 2000). This is a potential source
of important admixtures leading to
misinterpretations for instance in terms of
hominine hunting strategies or the
recognition of site function. Ongoing
researches in taphonomy aim to characterise
the accumulation produced by each bone
collectors (carnivores, raptors, humans) in
order to interpret the archaeological record.
The literature concerning damage caused by
hyenas is extremely prolific: this is due to
their ability to modify bone assemblages and
to accumulate bones inside their dens
(Brain, 1981; Haynes, 1983; Blumenschine,
1988; Hill, 1989; Cruz-Uribe, 1991;
Capaldo & Blumenschine, 1994; Pokines &
Kerbis Peterhans, 2007). In the last decade,
numerous actualistic studies have been
young and adult Cuon alpinus individuals. The identification of shed milk teeth demonstrates that this
carnivore used the cave as a nursery den. According to several authors dholes never bring back carcasses to
their dens in order to protect their offspring from other carnivores. However they tend to select an area
inside their den to defecate. We analysed modern scats of wolf in order to constitute a taphonomic
referential. Our results strongly suggest that most of the digested remains from the Noisetier Cave come
from dhole scats. This carnivore can be considered, as Binford previously suggested, as a bone accumulator
and consequently as a new taphonomic agent. Given the numerous sites where the fossil remains of this
carnivore were identified we argue that the dholes could have biased the composition of faunal spectrums
and maybe our understanding on human subsistence.
Keywords: DHOLE, CUON, COPROCOENOSIS, TAPHONOMY, NOISETIER CAVE, BONE
ACCUMULATION, DIGESTION, LATE PLEISTOCENE.

307
Mallye et al.
the frequency of digested bones is quite
comparable with that of chamois, but
anthropogenic marks are abundant. The
body part representation for the ibex shows
an intermediate pattern between the red deer
and the bovids on the one hand, and the
chamois on the other hand (Costamagno et
al., 2008). In summary, red deer and bovids
remains are mostly the result of a hunt by
the Mousterian people even if some of them
were secondly scavenged by carnivores. A
part of the ibex assemblage seems to have
been accumulated by Mousterian people
whereas the other part was introduced by a
non human accumulator, presumably the
bearded vulture.
In this paper we address the question
of the relation between the occurrence of
digested bones and the presence of different
carnivores such as the wolf (Canis lupus),
the dhole (Cuon alpinus) and the leopard
(Panthera pardus). In the light of new
actualistic studies, we test the hypothesis
previously expounded by Costamagno and
colleagues (2008) on the origin of chamois
assemblage.
Sites and Material
Noisetier Cave
Noisetier Cave (French Pyrenees) is a small
cave dwelled at a height of 825m a.s.l. and
at 145m above the Aure Valley at the
bottom of which flows the river Neste
(Figure 1). It is of 20m deep and measures 3
-4m in width and 3-6m in height. M. Allard
al., 2008; Lloveras et al., 2011) while only a
few studies concern the characterization of
ungulates bone accumulations created by
faecal deposits of large carnivores (Binford,
1981; Payne & Munson, 1985; Klippel et
al., 1987; Barja-Nunez & Corona, 2007;
Esteban-Nadal et al., 2010; Stiner et al.,
2012). The results of these studies lead them
to the same conclusions regarding the small
size of the remains, the relatively high
percentage of partially digested remains and
the possible occurrence of tooth marks.
Since its first excavation at the
beginning of the 1990’s, Noisetier cave was
interpreted as a hunting camp specialized on
chamois (Rupicapra rupicapra) and ibex
(Capra ibex) hunt (Jaubert & Bismuth,
1993). The taphonomic analysis conducted
by Costamagno and colleagues (2008) led to
modify the initial assumption regarding the
function of the site. In this study, the authors
demonstrate that a large part of the chamois
remains as well as a significant percentage
of the ibex material were not accumulated
by Neanderthal. Because of the presence of
abundant digested remains, the low
percentage of tooth marks on bones, a large
amount of short bones and cancellous bone
portions, Costamagno and colleagues (2008)
proposed that the bearded vulture (Gypaetus
barbatus) could be one of the possible
accumulators of chamois bones in the cave.
In contrast, red deer (Cervus elaphus) and
bovids are mainly represented by shaft
fragments. Short bones and cancellous bone
portions are underrepresented for these taxa
and butchery marks are extremely abundant
while carnivore marks are rare. For the ibex,

308
Dhole as a bone accumulator at Noisetier Cave
(Capreolus capreolus - 0.05%) and horse
(Equus caballus - 0.02%). Carnivores
represent less than 8% of number of
identified specimen for large and medium-
sized mammals. They are mostly
represented by foxes (Vulpes vulpes - 1.5%)
and bears (Ursus arctos - 3%). Weasel
(Mustela nivalis - 1%), dhole (Cuon alpinus
- 0.9%), leopard (Panthera pardus - 0.4%),
wolf (Canis lupus - 0.4%), polecat (Mustela
putorius - 0.05%) and Eurasian badger
(Meles meles – 0,02%) are represented by
fewer remains. Small games are mainly
composed of marmot (Marmotta marmotta -
3.6%) and hare (Lepus sp. - 1%).
Microvertebrates (rodents and birds)
remains are extremely abundant.
All coordinated remains and
anatomically and/or taxonomically identified
remains recovered after sieving operation
were analysed. The total number of identified
remains (NISP) at Noisetier cave is 4421
amongst which 1755 remains are attributed to
chamois. Inside the chamois assemblage, 482
remains were re-analysed in order to provide
a better characterization of the non-human
predation marks.
Les Loups du Gévaudan Park
Les Loups du Gévaudan Park is located in the
French department of Lozère (Figure 1). Since
2009, the authors have undertaken actualistic
taphonomic experimental studies there. The
aims of these studies are to characterise the
coprocoenosis generated by wolves from a
taphonomic and a micromorphologic
perspective. The wolves are fed with
first excavated the site in 1987 and 1992
(Allard, 1993, 1994). Since 2004 V.
Mourre, S. Costamagno and C. Thiébaut are
conducting new excavations and studies.
The excavation has yielded a Mousterian
industry with Discoïd and Levallois
debitage on local raw materials (Mourre et
al., 2008a,b; Thiébaut et al., 2012), bone
retouchers (Mallye et al., 2012) and human
remains (Maureille et al., 2007). Based on
AMS radiocarbon dates and
biochronological data (microfaunal and
macrofaunal remains), the archaeological
levels are attributed to a temperate phase of
the OIS 3 (see Mourre et al., 2008b for
details). The faunal spectrum is clearly
dominated by the chamois (Rupicapra
rupicapra - 64.4%), followed by the ibex
(Capra pyenaica - 12.7%), and to a lesser
extent by the red deer (Cervus elaphus -
9.3%), bovids (Bos / Bison - 1.5%), roe deer
Figure 1. Location of Noisetier cave and Les
Loups du Gévaudan Park.

309
Mallye et al.
In order to characterise the
coprocoenosis generated by wolves, 85
droppings of Mongolian wolf were
recovered. The scats were run through a
2mm mesh sieve. All skeletal and dental
elements as well as the residual fragments
of scats were collected and provide an
assemblage of 1,314 remains. In 12 scats,
no bone could be identified. The largest
number of remains recovered in a single
scat is 125 while the minimal number of
remains is 1. On average each scat yielded
15 remains. No microvertebrate remains
were identified. The percentage of identified
remains varies from 0 to 75% with an
average of 7.2%. Amongst the identified
remains, 105 were anatomically identified
and assigned to an ungulate class size.
At the Gévaudan, legs are rarely
given to the wolves. In order to characterize
the destruction pattern for these elements,
we decided to punctually feed the wolves
with ten sheep legs. All the legs were
composed of carpals / distal row of tarsals,
metapodials and phalanges with sesamoids.
After consumption the scats were
systematically sampled throughout the week
of experimentation. They were subsequently
run through a 2mm mesh sieve to collect the
bones. Out of the ten legs given to the
wolves, only 64 bones were recovered.
These remains represent only four legs.
The overall material collected at the
Gévaudan represents nearly 7,000 bones.
Most of the remains come from the
excavation of the dunging area (81,9%)
whereas the remaining part was recovered
after sieving of the scats.
ungulates carcasses (calf, beef, pork, lamb) cut
into quarters (about 1.8kg per wolf per day).
Head and leg ends (metapodials, phalanges
and sesamoids) are rarely given. Occasionally
the whole carcasses of pig, sheep or red deer
are distributed.
Two populations of wolves retained
our attention for taphonomic studies
undertaken on the dunging areas. The first
pack is composed of 31 individuals from
Siberia whose weight varies from 30 kg
(females, N=16) to 50 kg (males, N=29). They
occupy a fenced area of 1100 square metres.
We excavated a part of a dunging area of 1
square metre and 5cm deep. The sediment was
screened using a 2mm mesh sieve. This
operation yielded a total faunal assemblage of
1,285 specimens. The second sample comes
from an enclosure of 17,000 square metres
that include 3 packs composed of 45
individuals from Mongolia. The first pack is
composed of 14 males and 9 females, the
second is composed of 6 males and 7 females
and the last one comprises 3 males and 4
females. Average weights are identical to the
previous pack. We conducted an excavation of
2 square metres and 5cm deep. Using the
same screening method we collected 4,194
remains. Of the 5,479 bone specimens
recovered from the excavation of the two
dunging areas, a sample of 1,474 remains was
studied. This sample includes all identifiable
remains and/or longer than 1cm. Of this
sample, only 134 remains were identified
anatomically and/or taxonomically. The
species identified in this sample correspond to
those given during the feeding of wolves
except for the raven (Corvus corax).

310
Dhole as a bone accumulator at Noisetier Cave
bones, we used the criterion of dissolution
of the cortical to determine whether the
elements were digested or not.
In order to compare the chamois
sample from Noisetier cave with our
referential framework from Gévaudan, we
only took into account the anatomically and/
or taxonomically identified remains. This
quantification method unfortunately
prevents any comparison with data from
other studies on wolf scats (Esteban-Nadal
et al., 2010; Barja Núñez & Corona, 2007).
Concerning the identified bones
exhibiting traces of acid attacks, the intensity
of the digestion was assessed by considering
the proportion of bone attacked by the gastric
acids. Thus, codes ranging from 0 to 4 were
given based on the etched surface with respect
to the total surface area. The “0” value
corresponds to an intact remain, the “1” value
is given when up to 25% of the total surface is
modified by acid attacks, “2” value when 50%
is affected, “3” when 75% is affected and “4”
value when the entire surface is attacked.
For the spongy portions, we also
quantified the intensity of the dissolution of
the cortical bone. Stage 1 was assigned to
the remains with the cortical bone merely
dissolved whereas stage 2 corresponds to
the remains with a widening of the
cancellous bone.
The occurrence of tooth marks was
recorded according to the criteria described
by Binford (1981).
The occurrence of cupules on the bone
surfaces has also been recorded. These small
cupules are ellipsoid to rounded depressions
scattered on the surface of the bones.
Method
All dental and skeletal remains were measured
for maximum length (mm) to characterize the
fragmentation of the assemblage. Various
criteria were recorded for elements identifiable
and/or greater than 1cm.
The body part representation of the
chamois assemblage from Noisetier cave have
been compared with the data given by Robert
& Vigne (2002) for bearded vulture nests
using the percentage of representation defined
by Dodson & Wexlar (1979). Since some
anatomical parts were not present on the
carcasses given to the wolves at the Gévaudan
(e.g. heads or legs), the body part
representation could not be established on
these assemblages (scats and dunging area).
To fill this gap, we compared the anatomical
profile computed of the chamois assemblage
from Noisetier Cave with the results obtained
by Esteban-Nadal and colleagues (2010) on
wild wolf and by Stiner and colleagues (2012)
on Mountain lion using the percentage of
NISP for elements grouped per body parts:
skull (cranium + mandibule), teeth, vertebrae,
ribs (ribs + costal cartilage), girdles (scapula +
pelvis), long bones (humerus + radius + ulna +
femora + tibia), metapodial (metacarpals +
metatarsial + undefined metapodial), articular
bones (carpals + tarsals + patella + sesamoids)
and phalanges.
The criteria identified by d'Errico
and Villa (1997) were used during the
identification of digested bones. Digestion
is characterized by the appearance of holes
and by a slimming of the fracture edges and
general polishing of the bones. For spongy

311
Mallye et al.
Rates Mus spretus Crocidura russula Corrected pc/c 156.12 134.05
f+h/md+mx 54.55 37.84
t+r/f+h 78.21 64.29
Results
Noisetier Cave
1. Bone collectors in Noisetier Cave
A) Cuon
At Noisetier Cave, the reconsideration of
the faunal material led to the identification
of 48 remains of dhole including twenty
deciduous teeth (Figure 2). On several teeth
We used the categories of digestion
marks first described by Robert and Vigne
(2002) and later developed by Marin Arroyo
and colleagues (2009, 2011) to establish a
first quantification of the damages observed
on the phalanges.
In addition to this quantitative study
of acid attacks, we sought to highlight the
characteristic pattern of degradation for
various skeletal elements.
Figure 2. Noisetier cave. Deciduous teeth of dhole (Cuon alpinus). 1a, b, c, d: fourth upper deciduous
teeth. 2a, b, c: third upper deciduous teeth. 3: second lower deciduous incisor. 4: third lower deciduous
incisor. 5: lower deciduous canine. 6a, b: fourth lower deciduous teeth. 7a, b: third lower deciduous
teeth. 8a, b: second lower deciduous teeth.

312
Dhole as a bone accumulator at Noisetier Cave
compared to wolf deciduous teeth. The
proportion between the size of the metacone
and the size of the paracone is comparable to
what is observed on adult upper carnassial.
Furthermore, the small cusp located between
the paraconule and the protocone is less
pronounced on the teeth from Noisetier Cave
than on wolf deciduous teeth.
The shape of the Ud4 from Noisetier
Cave differs from the shape of wolf
deciduous teeth. The crown portion that
surrounds the metacone is very angular on
wolf Ud4, whereas it is more rounded on
the Ud4 from Noisetier Cave. On the wolf
Ud4, accessory cusps disposed on the mesio
-lingual edge of the crown can be observed,
while these additional cusps are absent on
the Ud4 from Noisetier Cave. On the wolf
Ud4, the mesial edge is straight, whereas it
is rounded on the Ud4 from Noisetier Cave.
A “V”-shaped depression, located in the
middle of the buccal edge, separates the
metacone from the paracone on wolf Ud4,
whereas it is absent on the Ud4 from
Noisetier Cave.
All these criteria/characteristics lead
us to attribute the complete assemblage of
canid deciduous teeth from Noisetier Cave
to the dhole (Cuon alpinus).
Carnivore marks are very rare on
dhole remains. Tooth marks were identified
on only one bone and a fragment of tooth
shows digestion marks (Table 1).
B) Wolf
At Noisetier Cave, the wolf is only
represented by adult remains. Only 23 remains
could be attributed to this carnivore and it is
(NISP = 16), a root resorption can be
identified due to exfoliation process. This
process is typical of shed deciduous teeth
(Hillson, 2005). We identified incisors
(Figure 2: 3 & 4), canines (Figure 2: 5),
second (figure 2: 8), third (Figure 2: 2 and
7) or fourth deciduous (Figure 2: 1 and 6)
teeth indicating various stages of
maturation.
The canid deciduous teeth identified
at Noisetier Cave were compared (1) to
modern wolf deciduous teeth from Italy and
Portugal (NI = 5), and (2) to upper
Pleistocene teeth of adult dhole from
Malarnaud Cave (Muséum d’Histoire
naturelle de Bordeaux). According to
Hillson (2005), in canids, the morphology
of deciduous teeth is similar to the
morphology of permanent teeth.
All the canid deciduous teeth from
Noisetier Cave are smaller but look more
robust than wolf milk teeth. From a
morphological point of view, several
differences in the shape of the teeth can be
observed on the fourth lower (Ld4), the
third upper (Ud3) and the fourth upper
deciduous teeth (Ud4).
The talonid of the Ld4 presents two
cusps in wolves, while a single cusp occurs
on the canid teeth from Noisetier Cave. The
occurrence of a single cusp on the talonid of
the lower carnassials of adults is a feature
that characterises the genus Cuon (Ewer,
1998). Thus, the morphology of the Ld4
from Noisetier Cave closely matches the
morphology of dhole lower carnassials.
The Ud3 in Noisetier Cave
assemblage have a small and lower metacone

313
Mallye et al.
Bo
dy p
arts
Lar
ge
can
id
(Dh
ole
or
Wo
lf)
Dh
ole
W
olf
L
eop
ard
Tota
l N
ISP
T
ooth
mar
ks
Dig
esti
on
mar
ks
NIS
P
Tooth
mar
ks
Dig
esti
on
mar
ks
NIS
P
Tooth
mar
ks
Dig
esti
on
mar
ks
NIS
P
Tooth
mar
ks
Dig
esti
on
mar
ks
Sku
ll
- -
- -
- -
- -
- -
- -
-
Per
man
ent
teet
h
9
- 1
19
- 1
- -
- 1
0
- -
38
Dec
idu
ou
s te
eth
- -
- 2
0
- -
- -
- 1
- -
21
Ver
teb
rae
4
1
1
2
- -
2
- -
2
1
- 1
0
Rib
s 3
- -
- -
- -
- -
- -
- 3
Ste
rneb
rae
1
- -
- -
- -
- -
- -
- 1
Hu
mer
us
- -
- 1
1
- -
- -
- -
- 1
Rad
ius
- -
- -
- -
- -
- -
- -
-
Uln
a 1
1
- -
- -
- -
- -
- -
1
Car
pal
s -
- -
- -
- -
- -
- -
- -
Met
acar
pal
s -
- -
3
- -
2
- 1
- -
- 5
Pel
vis
-
- -
1
- -
- -
- -
- -
1
Fem
ora
-
- -
- -
- 1
1
- -
- -
1
Pat
ella
e 4
1
1
- -
- -
- -
- -
- 4
Fib
ula
1
- -
- -
- 1
- -
- -
- 2
Tib
ia
- -
- -
- -
- -
- -
- -
-
Tar
sals
1
1
1
0
- -
3
- 1
1
- -
5
Met
atar
sals
-
- -
1
- -
2
- -
1
- -
4
Ph
alan
ges
6
- -
- -
- 1
1
1
1
5
2
- 2
2
Ses
amo
ids
3
- 1
1
- -
- -
- 4
- 1
8
Un
iden
tifi
ied
met
apo
dia
ls
- -
- -
- -
1
- -
- -
- 1
Tota
l 3
3
4
5
48
1
1
23
2
3
24
3
1
12
8
Ta
ble
1.
Nu
mb
er o
f id
enti
fied
rem
ain
s (N
ISP
), n
um
ber
of
too
th m
ark
s a
nd
nu
mb
er o
f d
iges
ted
ma
rks
for
each
ca
rniv
ore
fro
m N
ois
etie
r
cave
ass
emb
lag
e.

314
Dhole as a bone accumulator at Noisetier Cave
exceed 2cm long (Figure 4A). Among the
482 remains of chamois we re-examined,
85% present digestion marks (NISP = 410).
More than the three thirds of the digested
material are less than 2cm long and do not
exceed 4cm long (Figure 4B).
B) Body part representation
The large amount of compact bones and
phalanges was one of the main arguments used
to infer the major role played by the bearded
vulture in the chamois accumulation
(Costamagno et al., 2008). In addition chamois
long bones are only represented by epiphysis
fragments while the red deer, bovid and horse
long bones are mostly composed of shaft
fragments (Costamagno et al., 2008). When
comparing the body part profile for the
chamois from Noisetier Cave with that from
modern bearded vulture nests (Robert & Vigne,
2002) several differences can be noticed
(Figure 5). It is particularly relevant regarding
the proportion of short bones, scapulae, pelvis,
femurs and patellae (Figure 5).
difficult to discuss the body part profile (Table
1). It is however worth noting that elements of
the feet are abundant. Compared to the dhole,
neither shed milk teeth nor remains of
juveniles were identified. Tooth marks are
present on two bones and three digested
remains have been recognised namely a first
metacarpal, a talus and a first phalange.
C) Leopard
The occurrence of leopard in the
assemblage is attested by a few remains
(Table 1). They mostly consist of teeth and
foot bones. Only one deciduous tooth was
discovered indicating the presence of at
least one juvenile. The roots of the tooth are
still present which implies that this
individual died in the cave. Tooth marks
were identified on vertebrae and two
phalanges. A sesamoid is partially digested.
2. Coprolites
A large number of orange-white coloured
and parallelepiped remains with chalky and
dense texture were unearthed from the
sediment of the cave. Except for the colour,
which could be the consequence of post-
depositional processes, their aspect is closely
similar to that of coprolites (Figure 3).
3. Ungulates
A) Size of the remains
At Noisetier Cave, the overall studied
material is composed of 4421 remains
(taxonomically and/or anatomically
identified). Of this sample, 1994 remains
show digestion marks and nearly the three
quarters (74,5%) of the material do not
Figure 3. Noisetier cave: Fragments of coprolites.

315
Mallye et al.
Figure 4. Ungulate size remains according to size-classes. Digested remains in black, intact in grey. A:
complete Noisetier cave ungulate assemblage. B: Sample of chamois assemblage. C: Gévaudan sample.
Figure 5. Body part representation of chamois compared to that given by Robert & Vigne (2002) in
bearded vulture nest.
316
Dhole as a bone accumulator at Noisetier Cave
significant difference exists between the
two sample (Spearman’s Rho test R =
0,864, P<0,005).
C) Tooth marks
The frequency of tooth marks (Table 2
and Table 3) is significantly higher
(26.7%) in the chamois sample we
revaluated than the frequency results
obtained in the previous study (3%,
Costamagno et al., 2008). It should be
noted that this percentage is calculated by
dividing the number of chamois remains
bearing such marks (NISP=127) by the
total number of chamois remains
considering in this study and excluding
teeth (NISP=476).
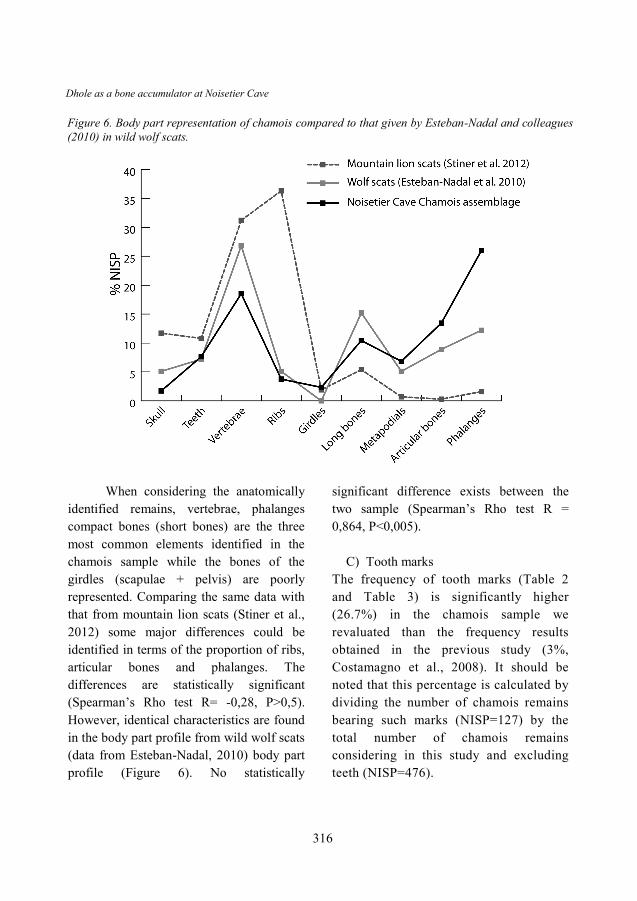
When considering the anatomically
identified remains, vertebrae, phalanges
compact bones (short bones) are the three
most common elements identified in the
chamois sample while the bones of the
girdles (scapulae + pelvis) are poorly
represented. Comparing the same data with
that from mountain lion scats (Stiner et al.,
2012) some major differences could be
identified in terms of the proportion of ribs,
articular bones and phalanges. The
differences are statistically significant
(Spearman’s Rho test R= -0,28, P>0,5).
However, identical characteristics are found
in the body part profile from wild wolf scats
(data from Esteban-Nadal, 2010) body part
profile (Figure 6). No statistically
Figure 6. Body part representation of chamois compared to that given by Esteban-Nadal and colleagues
(2010) in wild wolf scats.

317
Mallye et al.
Gévaudan sample Noisetier Cave
Scats Dunging area Chamois sample
Thinning 68 80 269
Cortical dissolution**
Stage 0 12 6 28
Stage 1 35 57 172
Stage 2 32 34 207
Digestion intensity
Stage 0 24 28 69 Stage 1 20 29 54 Stage 2 11 17 92 Stage 3 16 16 70 Stage 4 34 44 191
Tooth marks** Digested 10 16 109 Not digested 4 6 18
Cupule** 18 29 176
Table 2. Characteristic of chemical and carnivore attacks in Gévaudan sample and chamois sample from
Noisetier cave. ** implies that teeth are not counted.
Body part representation Noisetier Cave: Chamois sample
NISP Digestion
marks Tooth
marks Complete
remains
Skull 8 7 2 - Teeth 6 3 - 3 Vertebrae 45 35 7 - Ribs 7 4 1 - Costal cartilage - - - - Scapula 5 5 1 - Humerus 4 4 1 - Radius 2 1 1 - Ulna 6 5 1 - Carpals 32 31 7 11 Pelvis 7 6 1 - Femora 70 58 30 - Patellae 26 24 9 6 Tibia 19 10 5 - Metapodial 10 7 2 - Tarsal 45 41 19 7 Phalanges 147 129 35 72 Sesamoids 42 39 5 32 Long bones 1 1 - -
Total 482 410 127 131
Table 3. Studied sample of chamois remains from Noisetier Cave according to body parts, number of
digested remains, number of tooth marks and number of complete remains.

318
Dhole as a bone accumulator at Noisetier Cave
207). Among the 410 digested chamois
remains, 65.6% show a slimming of their
fracture edges and 42.9% present cupules
on their surface (Table 2).
E) Patterns of bone destruction
- Talus
On the talus, digestion marks are always
located near the articular surfaces (Figure
7). When the degree of digestion becomes
more intense, gastric acid attacks increase
and dig the bone around the sulcus tali.
While the general shape of the bone is
preserved, compact bone disappears and
cancellous bone begins to be dug therefore
creating punctures.
D) Digestion
More than eighty percent (85%) of the
chamois sample we analysed shows
indisputable digestion marks (Table and
Table 3). The analysis of chamois digested
bones from Noisetier Cave reveals that
63.6% of them are characterised by chemical
attacks affecting more than half of their
surface (Digestion intensity, stages 3 and 4).
Almost all of digested chamois
remains from Noisetier Cave assemblage
present a loss of cortical tissue (NISP =
379). Acid attacks have dissolved only the
compact bone (stage 1, NISP= 172) but in
most cases, they have also affected spongy
bone, creating perforations (stage 2 NISP =
Figure 7. Partially-digested talus from Noisetier Cave (left) and Gévaudan sample (right). Scale bar is 1cm.

319
Mallye et al.
- Femora
Femur fragments are found in larger quantity in
Noisetier Cave assemblage. They are mostly
represented by heads which may include a shaft
fragment gradually tapering away from the
articular surface (Figure 10). This kind of
morphology (called “morphologie en clou”) has
been previously described in bearded vulture
nest accumulation (Robert & Vigne, 2002) and
observed by Costamagno and colleagues (2008).
On the distal parts of the femur, acid attacks are
located on around both sides of the condyle
surfaces. Acid attacks can also create small
perforations of the bone (Figure 11).
- Tibiae
Tibiae are often identified by the tibia tuberosity
to which is attached a portion of shaft. Acid
attacks are located around the tuberosity. On the
shaft portion a slimming of edge the fracture
edges can be observed (Figure 12).
- Calcaneus
At Noisetier Cave, two identifiable portions
of calcaneus were identified. The first is the
calcaneus head and the second, the posterior
face of the talus attached to a part of the
shaft (Figure 8). Both present acid attacks
and a slimming of the fracture edges.
- Patella
The patellae are found in large number
inside the Noisetier Cave assemblage. Thus,
the comparison between the least destroyed
patella and the more digested ones allows a
good understanding of the destruction
pattern. On the former, the digestion marks
are located on the periphery of the medial
and lateral articular surfaces. While the acid
attacks intensified, the areas located on the
periphery of the articular surfaces are
gradually removed. The patellae take then
the form of a dolly (Figure 9).
Figure 8. Comparison between partially-digested calcanei from Noisetier cave (left) and Gévaudan
sample (right). Scale bar is 1cm.

320
Dhole as a bone accumulator at Noisetier Cave
Figure 10. Partially digested femora
(proximal parts, “morphologie en clou”)
from Noisetier Cave (left) and Gévaudan
sample (right)
Figure 9. Partially digested patellae from Noisetier cave (left) and Gévaudan sample (right). Scale bar is
1cm.

321
Mallye et al.
Figure 11. Partially-digested femurs (distal parts) from Noisetier Cave (left) and Gévaudan sample
(right). Scale bar is 1cm.
Figure 12. Partially-digested tibias from Noisetier Cave (left) and Gévaudan sample (right). Scale bar
is 1cm.

322
Dhole as a bone accumulator at Noisetier Cave
Gévaudan sample
1. Coprolites
In the field (Figure 14), the scats become
quickly dry. Because of the weathering
and/or trampling effects they break up in
small fragments measuring less than 1cm
long releasing the faunal material they
contain and then get buried in the soil.
These fragments of scats were recovered
after sieving operation from both scats
and soil from the dunging area (Figure
15).
- Phalanges
In the Noisetier cave assemblage,
digestion marks observed on the first and
second phalanges are located on the shaft
and are associated with a slimming of the
fracture edges. In addition acid attacks
perforate the distal part of the first
phalanges (Costamagno et al., 2008).
Most of the third phalanges are
characterised by acid attacks located
around the articular surface. Digestion
marks are always recorded on broken
phalanges and code 3.6 (Robert & Vigne,
2002) or 36 (Marín-Arroyo et al., 2009) is
the most frequent (Figure 13).
Figure 13. Partially-digested phalanges from Noisetier Cave (left) and Gévaudan sample (right).
Scale bar is 1cm.

323
Mallye et al.
from the dunging area (3966/5479). In other
words, remains from the scats are longer
than those from the dunging area. In the
three samples, the remains have a maximum
length of 5cm. The average percentage of
complete elements is 1.9% in the scat
sample and 1.7% in the dunging area
sample. Of the 1314 remains from the scats
sample, only 18 were complete or almost
complete. Only 26 remains were complete
among the 1474 studied remains from the
dunging area sample.
3. Digestion
The analysis of digested remains from the
dunging area sample (NISP = 852) shows
that 56,6% of the remains is affected by
chemical attacks on more than the half of
the surface. This proportion reaches 61.7%
in the scat sample (Table 2).
Almost all the digested remains from
the Gévaudan assemblage (85.8% in
dunging area and 82.7% in scats) present a
2. Size of the remains
The small size of the remains recovered at
the Gévaudan testifies of their high
fragmentation state. More than 80% of the
remains measured less than 2cm long and
never exceed 5cm long (Figure 4). At the
Gévaudan, only 57.8% of the dunging area
sample (NISP = 1474) present digestion
marks. Among the 134 identified remains
from the dunging area excavation, 79.1%
present digestion marks and more than 85%
of these remains are smaller than 3cm. Of
the 105 identified remains from the scat
sample, 77.1% show clear digestion marks
and 97.5% measured less than 3cm. In both
sample, the size of the digested remains
does not exceed 5cm long. Remains smaller
than 1cm represent nearly 50% of the scat
sample (654/1314), while they constitute
more than 70% of the material excavated
Figure 14. Gévaudan: post-depositional evolution
of wolf scats.
Figure 15. Fragments of scats (=coprolites) re-
covered after sieving operation at Gévaudan.

324
Dhole as a bone accumulator at Noisetier Cave
- Femora
Femur heads are poorly represented in the
Gévaudan sample. Among the 7,000 bones
recovered only one femoral head was
identified (Figure 9). A remaining fragment
of shaft is still attached above it. Digestion
marks are visible around the articular surface
of the head and fracture edges are slim.
However, distal parts of the femur are well
represented in the Gévaudan sample and all
of them present digestion marks. These
attacks are located near the articular surfaces
of the condyles on both sides. The cortical
bone has disappeared and the acid attacks
have frequently perforated the spongy bone
creating small holes (Figure 10).
- Tibia
In the Gévaudan assemblage, fragment of
tibia tuberosity have also been identified.
Once again a portion of the shaft is present
and acid attacks located around the
tuberosity cause a slimming of the shaft on
the fracture edges (Figure 11).
- Sheep’s’ legs
The overall material collected is
fragmented; only twenty-five remains out of
64 are complete or almost complete –
mainly carpals, tarsals and sesamoids. All
identified remains show digestion marks.
The slimming on the fracture edges is
observed on 34 remains. On 46 remains, the
cortical bone is dug by acid attacks (code
2). We identified tooth marks on only four
remains mainly consisting of notches. Of all
the phalanges identified after consumption
by wolves, only a third phalange is
loss of cortical tissue. Slimming of the
fracture edges of bones is high in wolf scats
(84%) and on the remains from the dunging
area (75.4%). The occurrence of cupules on
remains varies from 17.1% in the wolf scats
to 21.6% in the dunging area (Table 2).
4. Patterns of bone destruction
- Talus
In the Gévaudan sample, the tali are complete
or almost complete. The same pattern of
destruction previously noticed in the Noisetier
Cave assemblage is encountered in the
Gévaudan sample (Figure 7).
- Calcaneus
No complete calcaneus was recovered in the
studied sample. Two identifiable portions
were identified. The first is the calcaneus
head and the second is the posterior face of
the talus attached to a part of the shaft. Both
present acid attacks and a slimming of the
fracture edges. Notches are identifiable on
both sides of the tubercular process
indicating that calcanei are crushed by
wolves (Figure 8).
- Patella
The patellae are found in large number in
the Gévaudan samples. On the least
destroyed specimens, the digestion marks
are located on the periphery of the medial
and lateral articular surfaces. Subsequently,
we noticed the loss of the apex. While the
acid attacks are rising, the areas located on
the periphery of the articular surfaces are
gradually removed. The patella then takes
the form of a dolly (Figure 9).

325
Mallye et al.
(size > 40mm). These large remains are
related to long bone shaft fragments
(splinters) of the red deer, bovid, horse and
to a lesser extent to the ibex. The
examination of the surfaces on these large
remains shows many anthropogenic marks
and a lack of carnivore marks as previously
noticed (Costamagno et al., 2008). This
indicates that they were introduced in the
cave by Neanderthals rather than by a non-
human predator.
The distribution of digested remains
in the chamois sample from Noisetier Cave
is quite similar to that observed in the
Gévaudan sample: bones larger than 40mm
are absent and bones between 10 and 20mm
are the most represented.
2. Digestion
The chamois digested remains from the
Noisetier Cave are smaller in size than the
anatomically or taxonomically remains
identified in the Gévaudan sample.
On only 57.8% of the 1474 remains
from the excavation of the dunging area
digestion marks have been recognised. The
high frequency of spongy bone fragments in
the sample (28%) could explain this low
percentage. As previously noticed by
Esteban-Nadal and colleagues (2010), the
structure of spongy bones restricts the
identification of digestion marks. When
taking into account only the identified
bones, the percentage of partially-digested
remains is quite similar across the different
samples. The lowest percentages are
observed on bones from the scat sample
(77%) and from the dunging area (79%)
complete. The other thirty-three specimens
of phalanges are fragmented. Finally, in
addition to slimming of the fracture edges,
we noticed that acid attacks begin to
perforate the distal part of the first and
second phalanges (Figure 12).
Discussion
Comparison between Noisetier Cave and
wolf accumulation
1. Size of the remains
The size of remains from the Gévaudan
scats is comparable to that observed by
Esteban-Nadal and colleagues (2010) in
wild wolf scats and to that observed by
Payne and Munson (1985) in dog faeces.
However, we demonstrated that the
maximal length of bones from the scats is
significantly longer than that observed on
remains from the dunging area. This can be
explained by post depositional
fragmentation due to trampling by wolves
as previously noticed on coprolites
fragments. It is highly probable that this
phenomenon could have affected the bones
in the past. Thus, the distribution by size
class observed for the bone recovered from
the dunging area is a better reflection of
what can be found in the fossil record than
the maximum length of the remains from
the scats. The distribution of remains by
size classes in the Gévaudan sample is quite
similar to that observed in Noisetier Cave
for the total ungulates sample. However, in
the latter larger remains have been recorded

326
Dhole as a bone accumulator at Noisetier Cave
75.3% in scats sample and 75.4% in
dropping area). In the sample of Iberian
wolf scats (Esteban-Nadal et al., 2010) the
slimming of edges is more limited (29.5%)
but the quantification methods applied by
these authors are different. They calculate
the proportions from whole assemblage,
including the unidentified remains whereas
in our study only the identified remains
were considered.
3. Tooth marks
Tooth marks have been recognised on
26.7% of the chamois bones from
Noisetier Cave. This percentage is higher
than the percentage previously observed
by Costamagno and colleagues (2008).
Lower but similar frequencies on
identified remains have been noticed on
the Gévaudan sample, ranging from
13.3% in the scats sample to 16.4% in the
dunging area sample. These frequencies
are higher than frequencies described in
other studies (2.4% Esteban-Nadal et al.,
2010; 7.9% Barja Núñez & Corona,
2007). However, in these studies, the
occurrence of tooth marks is calculated
according to the total number of remains,
including unidentified remains.
4. Bone completeness
In the Gévaudan samples, the percentages
of complete bones are very low: 1.8% in the
dunging area and only 1.34% in the scat
sample. Both are lower than percentages
calculated by Esteban-Nadal and colleagues
(2010) in their sample (2.2%). This
difference could be related to the variability
whereas 84.7% of the chamois remains
from Noisetier Cave show digestion marks.
All these percentages are nonetheless
smaller than those obtained by Esteban-
Nadal and colleagues (2010) [92.7%] or
Barja Núñez and Corona (2007) [99.1%] in
their study of wolf scats.
Regarding the digested elements,
about the two-thirds (65.6%) of chamois
remains in Noisetier Cave are characterised
by chemical attacks that affect more than
50% of the total surface (Stage 3 and 4).
Similar proportions are found in the
Gévaudan sample (61.7% in scats sample
and 56.6% in dunging area). Concerning the
proportions of the different codes, there is
no significant difference between the two
samples of digested bones from Gévaudan
(chi-square calc = 1,676, ddl = 4, P<0,1).
However, the comparison between the
Noisetier cave assemblage and the two
samples from Gévaudan highlights some
differences. In the bone sample from
Noisetier Cave, the intensity of digestion
was further coded as 2 or 4. In the
Gévaudan sample, the highest percentages
are found for bones coded 1 and 4.
Almost all of the digested bones
from Noisetier cave and Gévaudan showed
evidence of cortical dissolution (sensu
porosity by Esteban-Nadal et al. 2010). In
both cases, this kind of attack is substantial
since not only it affects the cortical bone but
also digs into the spongy bone.
Among the 410 digested chamois
remains, 65.6% shows a slimming of edges
fracture. This proportion is slightly higher
in the samples from Gévaudan (respectively

327
Mallye et al.
& Corona’s study (2007) on Iberian wild
wolf scats.
The morphology of tibial crest is quite
similar in both Noisetier Cave and the
Gévaudan assemblages. Similarly, a
comparison between the patellas identified at
Noisetier Cave and in the Gévaudan sample
shows the same patterns of alteration.
Chemical attacks recorded on phalanges in
Noisetier cave resemble closely to chemical
attacks observed on phalanges digested by
bearded vulture described by Robert & Vigne
(2002). This is especially true if we take into
account the perforation located at the distal
end. However, this pattern have also been
recognised by Klippel and colleagues (1987)
in their study on wolf scats samples and by
Chase (1990) on coyote scat sample. To date,
our sample of phalanges consumed by wolves
is too small to consider a statistical study of
the destruction pattern as it has been
conducted for other assemblages (Robert &
Vigne, 2002; Marin Arroyo et al., 2009; Marin
Arroyo & Margalida, 2011).
Bearded vulture nest or carnivore den?
The numerous patellae identified both at
Noisetier cave and in the Gévaudan sample
could provide information about the
accumulator of chamois. In mammals, the
extremities of the main thigh muscles are
located on the patella. This bone is thus
encased in a large muscle mass and is
articulated with the femur and the tibia by
ligaments. It is therefore likely to be
swallowed on fresh carcasses. Consequently,
we suggest that the patella is more accessible
between the different referential
frameworks. Comparing to Noisetier Cave
(37%), the number of anatomically and/or
taxonomically bones that are complete in
Gévaudan sample is lower (17-19%). This
implies that chamois remains at Noisetier
cave are less fragmented than remains
identified in wolf scats.
5. Cupules
Cupules have been identified in both the
Gévaudan sample (17-21%) and the
chamois remains from Noisetier Cave
(42,9%). It is difficult to establish clearly
their origin. Cupules can be created by the
action of gastric acids or be the result of
gnawing marks that are latter modified
during the digestion process as previously
noticed by Stiner and colleagues (2012).
6. Patterns of bone destruction
To complete our quantitative study of
digestion marks left on the ungulates
remains we compared the destruction of the
most frequently identified remains such as
carpals and tarsals but also phalanges,
patellae and tibia tuberosities in Gévaudan
sample and chamois remains from Noisetier
Cave. The remains presented in this study
belong primarily to small ungulates such as
sheep and chamois. These remains are very
small, and so can easily be ingested by
wolves.
The tali from the Noisetier Cave and
the Gévaudan sample show the same pattern
of degradation caused by carnivore teeth
and by chemical attacks. Identical patterns
of degradation are described in Barja Núñez

328
Dhole as a bone accumulator at Noisetier Cave
1981; Lyman, 1994; Mondini, 1995;
Mondini & Muñoz, 2008): residual
assemblage that results from a kill site and
in-situ consumption, transported assemblage
that corresponds to an accumulation of
bones from the kill site and scatological
assemblage which is the “result of the by-
products of bone ingestion and are
deposited within faeces” (Mondini &
Muñoz, 2008: 54). At Noisetier Cave, the
chamois assemblage is characterised by the
abundance of cancellous bone portions and
by the lack of long bones shaft fragments
(see Costamagno et al., 2008 for details and
proportions). In addition there is a large
amount of coprolites fragments and most of
the bones are digested. Consequently it is
highly probable that the chamois
assemblage derives from a scatological
assemblage. This assertion is reinforced by
the similarities observed between the body
part representations of the chamois sample
on the one hand and of the remains
identified in carnivores’ scats on the other
hand. In the last case the predators
unintentionally deposit faunal remains via
their scats. At Noisetier cave, the large
quantity of digested bones could be the
result of abundant scats indicating a long-
term use of the cavity by carnivores.
Which carnivore?
Eight species of carnivores have been
identified at Noisetier Cave. The smaller
taxa (weasel and polecat) cannot be
responsible of the chamois accumulation in
the cave as they prey mostly on small
to primary consumers such as terrestrial
carnivores rather than to scavengers such as
bearded vultures. In addition, the involvement
of a terrestrial carnivore in the constitution of
the chamois assemblage of Noisetier cave is
further proven by the high percentage of tooth
marks. We also demonstrate that the body part
representation of chamois assemblage of
Noisetier Cave is more closely similar to the
one provided by Esteban-Nadal and
colleagues (2010) in wolf scats than that given
by Robert and Vigne (2002) for bearded
vulture. The occurrence of coprolites has been
used to attest the occupation of the cave by the
bearded vulture (Costamagno et al., 2008).
Identical fragments were identified after the
excavation of the dunging area of wolves. In
such context the fragmentation is caused by
post-depositional processes such as
weathering or trampling by wolves for
instance. In the case of Noisetier Cave it is
likely that coprolite fragments derived from
the frequentation of the cave by inhabitants.
As previously noticed for other sites (e.g.
Stiner, 2004), the numerous fragments of
coprolites identified in Noisetier Cave indicate
long-term use of the cave as a den by
carnivores. An analysis of the geochemical
composition of these fragments compared to
modern coprolites of different species (wolf,
bearded vulture, dhole) would fully confirm
their origin.
Which kind of bone assemblage for chamois
in Noisetier cave?
Three kinds of accumulations can be related
to carnivores feeding activity (Binford,

329
Mallye et al.
marks and digestion marks. The body part
representation could hardly be discussed
due to the small size of the sample.
However, it is important to notice that some
of the undetermined canid remains belongs
to small individuals that could be related to
dholes.
Wolf is only represented by adult
remains. Only one milk tooth has been
attributed to the Leopard at Noisetier Cave.
The roots of this tooth are fully developed
without exfoliation process. If the cave was
used as a den by one of the carnivore over a
long period, remains of cubs should be
found in greater quantity as it is noticed in
cave used by bears in the context of
hibernation (e.g. David, 2002) or in hyaena
dens (e.g. Fosse, 1994). This is not the case
at Noisetier Cave.
The identification of shed milk teeth
of dhole leads us to make several
observations. To our knowledge, this is the
first time that dhole milk teeth are described
in the fossil record. According to the tooth
eruption wear pattern given by Hillson
(2005: 242) for canid, deciduous teeth are in
place at one to two months and permanent
teeth begins to erupt at five to six months to
be finally in place at six months. Few data
are available for dhole teething. However,
Sosnovskii (1967: 122) provides information
on dental eruption sequence from dholes kept
at the Moscow Zoo: “Teething begins on
Days 15 to 16, when the incisors of the upper
jaw appear. They are followed on Days 20-
21 by those of the lower jaw; canines appear
on Days 21-25; molars on Days 25-30; and
premolar on Days 30-35.” Thus, it appears
vertebrates. Large ungulate remains have
been identified in fox and badger burrows
as the result of scavenging habits of these
two carnivores (Mallye et al., 2008; Castel
et al., 2011). In these cases, such remains
are found in low quantities compared to
small vertebrates and are rarely digested. At
Noisetier Cave, cut marks have been
recorded on the only badger remain and
attest of human consumption. Traces of
digestion as well as tooth marks have been
observed on some of the fox remains and
attest of a non-human accumulation. The
large amount of bear deciduous teeth
indicate that the cave was occupied by bear
during hibernation.
Thus, three carnivores could be
responsible of chamois accumulation: the
dhole, the wolf and the leopard.
The comparison between the
chamois sample and the data provided on
wolf scats (Esteban-Nadal et al., 2010) and
mountain lion (Stiner et al., 2012) shows
greater similarities with canid scats than that
from lion. This is especially true when
looking at the proportion of ribs (higher in
lion scats) and that of short bones and
phalanges (higher in wolf scats).
On the other hand, the morphology
of digested chamois bones is quite similar to
what is observed in the Gévaudan
assemblage. Thus it is very probable that a
canid has created the chamois bone
assemblage in the cave through the
accumulation of its scats.
The analysis conducted on the
remains of three carnivores allowed us to
identify predation marks namely tooth

330
Dhole as a bone accumulator at Noisetier Cave
Fox, 1984). However scats are frequently
deposited in latrines and could be located in
the den (Fox, 1984; Johnsingh, 1982). Of
the 66 scats analysed, Borah and colleagues
(2009) report that bones are identified in
43.9% of the scats analysed. Then, the
accumulation of scats can produce a
coprocoenosis. Based on these ethological
data, it is very likely that the dholes used the
cave as a den and accidentally accumulated
ungulate bones by defecating.
The dhole as a new taphonomic agent
Up to now extensive research has been
conducted to characterise the bone
accumulations produced by large carnivores
such as hyaenas. Several factors led us to
underestimate the ability of dhole to create
notable bone accumulations in fossil context.
In some sites, it is possible that the remains
of dhole were not identified or were
mistaken with wolf remains. Recent studies
have identified the remains of fossil dholes
in fossil record and finally contributed to fill
this gap in our knowledge (Boudadi-
Maligne, 2010; Perez-Ripoll et al., 2010;
Pionnier-Capitan et al., 2011).
Few authors among which Simons
& Ettel (1970) for Southeast Asian sites
but also Binford (1988) in the case of
Vaufrey cave (layer VIII) have suggested
that dholes played a significant role in site
formation processes. In the latter case,
this hypothesis has been challenged
(Grayson & Delpech, 1994). However, the
work undertaken on Leporid remains
(Cochard, 2004, 2007) from the same
that tooth eruption pattern of dholes is quite
similar to that of other canids. Dhole are
known to occupy caves, rocky shelters or
burrow of their own digging to mate, give
birth and to breed the cubs (Davidar, 1974;
Johnsingh, 1982; Fox, 1984). The birthing
season occurs between November and March
with a peak in December (Davidar, 1975).
The nursing period lasts about 50 to 60 days
and thereafter adults regurgitate meat to feed
the cubs inside the den. Cubs help the adults
to kill prey at 6 to 8 months old. The
occurrence of shed milk teeth at Noisetier
Cave indicates that dhole have occupied the
cave. Based on sequences of tooth eruption,
and according to ethological data we may
conclude that this occupation may have
spanned for a long period, probably over 5
months of the year during the birthing and
weaning seasons.
Dholes are medium-size carnivores
that are widespread in South East Asia from
India through China to southern Java
(Durbin et al., 2004). Body weight for adult
males averages 20kg and 15kg for adult
females. This canid lives in pack ranging
from 5 to 11 individuals (Johnsingh, 1982)
but up to 40 individuals have been recorded
(Fox, 1984). Dholes hunt in pack and can
kill medium and large ungulates weighting
more than 200kg (Karanth & Sunquist,
1995; Venkataraman et al., 1995; Wang &
MacDonald, 2009; Thinley et al., 2011) as
well as smaller preys such as hare, birds and
rodents (Cohen, 1978; Venkataraman et al.,
1995; Borah et al. 2009).
Only occasionally dholes bring back
carcasses to their den (Johnsingh, 1982;

331
Mallye et al.
the bone accumulation process. The
constitution of a coproscopic referential
framework for this canid becomes
imperative. The continuation of such studies
will improve our understanding on human
paleoecology or human/carnivore
interactions as well as site formation
processes and site functions.
Acknowledgments
The authors would like to thank Jordi Rosell,
Enrique Baquedano, Ruth Blasco and
Edgard Camaros for organising the
International congress “Hominid Carnivore
Interaction during the Pleistocene” in Salou
where this paper was originally presented.
This research is granted by La Fondation des
Treilles project. Excavation at Le Noisetier
Cave is supported by the Ministère de la
culture et de la Communication and also the
Conseil Général des Hautes-Pyrénées. We
also would like to thank Jessica Lacarrière
who provides us supplementary data that
have stimulated our research, Walter E.
Klippel who provides us data on digested
bones. We want to thank Melvin E.
Sunquist, Leon Durbin, Lonnie I. Grassman,
Kate Jenks, Jan Kamler and Jimmy Borah
for their helpful information on Dhole scats.
Thanks to Luca Sitzia for his help on figure
16, many thanks to Le Parc des Loups du
Gévaudan to welcome us, with special
mention to Sylvain Macchi. Thanks to the
two anonymous reviewers for their helpful
comments that improve a previous version
of the paper.
layer tends to support the hypothesis of
Binford (1988). Based on recent works
(Brugal & Boudadi-Maligne, 2011; Perez-
Ripoll et al., 2010) and additional
bibliographic data, we can notice that the
dhole is widely distributed in western and
central Europe on the edge of the
mountain ranges (Figure 16). In several
sites, anthropogenic marks have been
identified on dhole remains (Perez-Ripoll
et al., 2010) attesting of human origin. In
other sites, such as Amalda cave (Altuna
et al., 1990) or Boquete de Zafarraya
(Barroso Ruiz, 2003; Barroso & de
Lumley, 2005) or Gabasa (Blasco-Sancho,
1995) dhole remains are interestingly
associated with ungulates that are
identical in size (e.g. chamois and ibex) to
those identified in Noisetier cave. Hence,
bone accumulations identical to the one
described in this paper could possibly be
identified in these sites.
Conclusion
The results obtained from both Noisetier
Cave and the Gévaudan assemblage offer
new research perspectives in taphonomy
concerning the role of carnivores in the site
formation processes. We suggest that
medium-sized carnivores such as dholes
could have had a real impact on site
formation processes and that dhole could
now be considered as a potential
taphonomic agent. This carnivore has been
recognised in many Pleistocene
assemblages and could have contributed to

332
Dhole as a bone accumulator at Noisetier Cave
Fig
ure
16
. P
ale
og
eog
rap
hic
dis
trib
uti
on o
f dh
ole
s d
uri
ng
th
e P
leis
toce
ne
in W
este
rn E
uro
pe.

333
Mallye et al.
Arambourg, C. (1958). Les gros mammiferes des couches tayaciennes. In (Alimen, H., Arambourg,
C. & Schreuder, A., eds.) La grotte de
Fontéchevade. Paris: Archive de l'Institut de Paléontologie Humaine, pp.185-229.
Argant, A. (1991). Carnivores quaternaires de
Bourgogne. Document du laboratoire de Géologie
de Lyon, Lyon.
Barja Núñez, I. & Corona-M, E. (2007). El análysis de
excretas desde la etología y la arqueozoología. El caso del lobo ibérico. In (Corona-M., E. & Arroyo-
Carales, J., eds.) Human and Faunal Relationships
Reviewed: An Archaeozoological Approach. BAR International Series, pp.113-121.
Barroso Ruiz, C. (2003). El Pleistoceno superior de la
Cueva del Boquete de Zafarraya. Arqueología Monographías, Junta da Andalucía, Consejería de
Cultura, Sevilla.
Barroso Ruiz, C. & de Lumley, H. (2006). La Grotte du Boquete de Zafarraya (Málaga, Andalousie).
Junta de Andalucía, Sevilla.
Baryshnikov, G. F. & Tsoukala, E. (2010). New analysis of the Pleistocene carnivores from
Petralona Cave (Macedonia, Greece) based on the
collection of the Thessaloniki Aristotle University. Geobios, 43: 389-402.
Binford, L. R. (1981). Bones: Ancient Men and Modern Myths. Academic Press, New-York.
Binford, L. R. (1988). Etude taphonomique des restes
fauniques de la grotte Vaufrey, Couche VIII. In (Rigaud, J.-P., ed.) La grotte Vaufrey:
Paleoenvironnement - Chronologie - Activité
humaines. Mémoires de la Société Préhistorique Française, pp.535-563.
Blasco Sancho, F. (1995). Hombres, fieras y presas.
Estudio arqueozoológico y tafonómico del yacimiento del Paleolítico Medio de la Cueva de Gabasa 1
(Huesca). Monografías Arqueológicas, Zaragoza.
Blumenschine, R. J. (1988). An experimental model of the timing of Hominid and carnivore influence on
archaeological bone assemblages. Journal of
Archaeological Science, 15: 483-502. Bonifay, M.-F. (1971). Carnivores quaternaires du Sud
-Est de la France. Mémoire du Muséum National
d'Histoire Naturelle, Paris. Borah, J., Deka, K., Dookia, S. & Prasad Gupta, R.
(2009). Food habits of dholes (Cuon alpinus) in
Saptura Tiger Reserve, Madhya Pradesh, India. Mammalia, 73: 85-88.
Bouchud, J. (1951). Etude paléontologique de la faune
d'Isturitz. Mammalia, 15(4): 184-203.
Bouchud, P. & Bouchud, J. (1953). La faune des
grottes des Orciers et de Cottier. Bulletin de la
Société Préhistorique Française, 50: 444-457.
References
Thiollay, J.M. (1978). Les rapaces d’une zone de
contact savane-forêt en Côte d’Ivoire :
spécialisations alimentaires. Alauda, 46: 147-170.
Winkler, A. J., Denys, C. & Avery, M. (2010).
Rodentia. In: Werdelin, L., Sanders, W. J. (Eds.):
Cenozoic Mammals of Africa. University of
California Press, Berkeley, Los Angeles, London. Chap. 17: 263-304.
Adam, K. D. (1959). Mittelpleistozäne Caniden aus
dem Heppenloch bei Gutenberg (Württemberg). Stuttgarter Beiträge zur Naturkunde, 27: 1-46.
Allard, M. (1993). Fréchet-Aure, Grotte du Noisetier.
In Bilan Scientifique 1992 de la Direction Régionale de Affaires Culturelles de Midi
Pyrénées. Ministère de la Culture, pp.113-114.
Allard, M. (1994). Fréchet-Aure. Grotte de Peyrère 1 (du Noisetier). In Bilan Scientifique 1993 de la
Direction Régionale de Affaires Culturelles de
Midi Pyrénées. Ministère de la Culture, pp.156. Altuna, J. (1973). Fauna de mamíferos del yacimiento
prehistórico de Los Casares (Guadalajara). In
(Barandiarán, I., ed.) La Cueva de Los Casares (en Riba de Saelice, Gudalajara). Madrid: Excavaciones
Arqueológicas en España, pp.97–116.
Altuna, J. (1981). Restos óseos deI yacimiento prehistórico deI Rascaño (Santander). In (González
Echegaray, F. & Barandiarán, I., eds.) El
Paleolitico superior de la cueva del Rascaño (Santander). Centro de Investigacion y Museo de
Altamira, pp.223-269.
Altuna, J. (1983). Hallazgo de un cuón (Cuon alpinus Pallas) en Obarreta, Gorbea (Vizcaya). Kobie,
XIII: 141-158.
Altuna, J. (1986). The mammalian faunas from the prehistoric site of La Riera. In (Straus, L. G. &
Clark, G., eds.) La Riera Cave. Stone Age Hunter–
Gatherer Adaptations in Northern Spain. Tempe:
Anthropological Research Papers, pp.237-274.
Altuna, J., Baldeón, A. & Mariezkurrena, K. (1990). La
Cueva de Amalda (Zestoa, País Vasco): Ocupaciones Paleolíticas y Postpaleolíticas. Fundación José
Miguel de Barandiarán, San Sebastián.
Andrews, P. & Nesbit Evans, E. M. (1983). Small mammal bone accumulations produced by
mammalian carnivores. Paleobiology, 9(3): 289-307.
Andrews, P. J. (1990). Owls, caves and fossils: predation, preservation and accumulation of small
mammal bones in caves, with an analysis of Pleistocene cave faunas from Westbury-sub-
Mendip, Somerset, UK. University of Chicago
Press, Chicago.

334
Dhole as a bone accumulator at Noisetier Cave
Champagne, F. & Espitalié, R. (1981). Le Piage, Site préhistorique de Lot. Mémoire de la Société
Préhistorique Française, Paris.
Chase, P. G. (1990). Tool-Making Tools and Middle Paleolithic Behavior. Current Anthropology, 31(4):
443-447.
Clot, A. (1987). La grotte de la Carrière (Gerde,
Hautes-Pyrénées). Stratigraphie et paléontologie
des Carnivores. Société Ramond, Bagnère-de-
Bigorre. Cochard, D. (2004). Les léporidés dans la subsistance
paléolithique du Sud de la France. Unpublished
PhD thesis, Université Bordeaux 1. 354 p. Cochard, D. (2007). Caractérisation des apports de
Léporidés dans les sites paléolithiques et
application méthodologiques à la couche VIII de la grotte Vaufrey. In Congrès du centenaire : Un
siècle de discours scientifique en Préhistoire.
XXVIè congrès préhistorique de France - Avignon, 21-25 septembre 2004. Paris: Société Préhitorique
Française, pp.467-480.
Cohen, J. A. (1978). Cuon alpinus. Mammalian Species, 100: 1-3.
Cordy, J.-M. (1983). Découverte de Cuon alpinus
europaeus Bourguignat dans le quaternaire de Belgique. In (Poplin, F., ed.) La Faune et l'Homme
préhistoriques. Paris: Mémoire de la société préhistorique française, pp.49-54.
Costamagno, S., Robert, I., Laroulandie, V., Mourre,
V. & Thiébaut, C. (2008). Rôle du Gypaète barbu (Gypaetus barbatus) dans la constitution de
l'assemblage osseux de la grotte du Noisetier
(Fréchet-Aure, Hautes Pyrénées, France). Annales de Paléontologie, 94: 245-265.
Crégut-Bonnoure, E., Boulbes, N., Daujard, C.,
Fernandez, P. & Valensi, P. (2010). Nouvelles données sur la grande faune de l'Éémien dans le
sud-est de la France. Quaternaire, 21/3: 227-248.
Cruze-Uribe, K. (1991). Distinguish Hyena from Hominid Bone Accumulations. Journal of Field
Archaeology, 18(4): 467-486.
d'Errico, F. & Villa, P. (1997). Holes and grooves: the contribution of microscopy and taphonomy to the
problem of art origins. Journal of Human
Evolution, 33: 1-31. David, F. (2002). Les ours du Châtelperronien de la
grotte du Renne à Arcy-sur-Cure (Yonne). In
(Tillet, T. & Binford, L. R., eds.) L'Ours et l'Homme. Actes du colloque d'Auberives-en-
Royans, 1997. Liège: ERAUL, pp.185-192.
Davidar, E. R. C. (1974). Observations at the dens of
the dhole or Indian wild dog (Cuon alpinus).
Journal of the Bombay Natural History society, 71
(2): 183-187.
Boudadi-Maligne, M. (2010). Les Canis pléistocènes du Sud de la France : Approches biostratigraphique,
évolutive et biochronologique. Unpublished PhD
Thesis, Université Bordeaux 1. 446 p. Boule, M. & Villeneuve, L. d. (1927). La grotte de
l'Observatoire à Monaco. Archive de l'Institut de
paléontologie humaine, Paris.
Bourguignat, J.-R. (1875). Recherches sur les
ossements de Canidae constatés en France à l'état
fossile pendant la période quaternaire. Masson, Paris.
Brain, C. K. (1981). The hunters or the hunted? An
introduction to african cave taphonomy. The University of Chicago Press, Chicago & London.
Brugal, J.-P. & Boudadi-Maligne, M. (2011). Quaternary
small to large canids in europe: Taxonomic status and biochronological contribution. Quaternary
International, 243: 171-182.
Campmas, E. & Beauval, C. (2008). Consommation osseuse des carnivores : résultats de l'étude de
l'exploitation de carcasses de bieufs (Bos taurus)
par des loups captifs. Annales de Paléontologie, 94: 167-186.
Capaldo, S. D. & Blumenschine, R. J. (1994). A
quantitative diagnosis of notches made by hammerstone percussion and carnivore gnawing on
Bovid long bone. American Antiquity, 59(4): 724-748.
Cardoso, J. L. (1992). Présence de Cuon alpinus
europaeus Bourguignat, 1868 (Mammalia, Carnivora) dans le Pléistocène du Portugal.
Ciências da Terra (UNL), 11: 65-76.
Castaños, P. M. (1987). Los carnívoros prehistóricos de Vizcaya. Kobie (serie Paleoanthropología),
XVI: 7-38.
Castel, J.-C. (2004). L'influence des canidés sur la formation des ensembles archéologiques.
Caractérisation des destructions dues au Loup.
Revue de Paléobiologie, Genève, 23(2): 675-693. Castel, J.-C., Coumont, M.-P., Boudadi-Maligne, M. &
Prucca, A. (2010). Rôle des grands carnivores dans
les accumulations naturelles. Le cas des loups (Canis lupus) de l'Igue du Gral (Sauliac-sur-Célé,
Lot, France). Revue de Paléobiologie, Genève, 29
(2): 411-425. Castel, J.-C., Mallye, J.-B. & Oppliger, J. (2011). Les
petits carnivores dans leur abris temporaires: choix
des espèces et caractéristiques taphonomiques. Implications pour l'archéologie. In (Laroulandie,
V., Mallye, J.-B. & Denys, C., eds.) Taphonomie
des Petits vertébrés: Référentiels et Transferts aux fossiles. Actes de la Table Ronde du RTP
Taphonomie, Talence 20-21 octobre 2009. Oxford:
BAR International Series, pp.77-91.

335
Mallye et al.
Fosse, P. (1994). Taphonomie paléolithique: Les grands Mammifères de soleilhac (Haute-Loire) et
de Lunel-Viel (Hérault). Thèse de Doctorat de
l'université de Provence (Aix-Marseille I), Université de Provence - Aix-Marseille I. 257 p.
254 annexes.
Fosse, P., Laudet, F., Selva, N. & Warjac, A. (2004).
Premières observations néotaphonomiques sur des
assemblages osseux de Bialowieza (N. E.
Pologne): intérêts pour les gisements pléistocènes d'Europe. PALEO, 16: 91-116.
Fox, M. W. (1984). The Whistling Hunter. Field
Studies of the Asiatic Wild Dog (Cuon alpinus). State University of New York Press, Albany.
García, N. (2003). Osos y Otros Carnívoros de la
Sierra de Atapuerca. Fundacion Oso de Asturias, Ovideo.
García, N. & Arsuaga, J.-L. (1998). The carnivore
remains from the Hominid-bearing trinchera-Galería, Sierra de Atapureca, Middle Pleistocene
site (Spain). Geobios, 31: 659-674.
Grassman, L. I. J., Tewes, M. E., Silvy, N., J. & Kreetiyutanont, K. (2005). Spatial ecology and diet
of the dhole Cuon alpinus (Canidae, Carnivora) in
north central Thailand. Mammalia, 69(1): 11-20. Grayson, D. K. & Delpech, F. (1994). The evidence for
Middle Palaeolithic scavenging from Couche VIII, Grotte Vaufrey (Dordogne, France). Journal of
Archaeological Science, 21: 359-375.
Haynes, G. (1980). Prey bones and predators: Potential ecologic information from analysis of bone sites.
Ossa, 7: 75-97.
Haynes, G. (1983). A guide for differentiating mammalian carnivore taxa responsible for gnaw
damage to herbivore limb bones. Paleobiology, 9
(2): 164-172. Hill, A. (1989). Bone modification by Modern Spotted
Hyenas. In (Bonnichsen, R. & Sorg, M. H., eds.)
Bone modification. Orono, Maine: Center for the Study of the First Americans, pp.169-178.
Hillson, S. (2005). Teeth, 2nd edition. Cambridge
University Press, New York. Jánossy, D. (1986). Pleistocene Vertebrate Faunas of
Hungary. Developments in Palaeontology and
Stratigraphy. Elsevier Science, Amsterdam. Jaubert, J. & Bismuth, T. (1993). Le Paléolithique
moyen des Pyrénées centrales esquisse d'un
schéma chronologique et économique dans la perspective d'une étude comparative avec les
documents ibériques. In Actes du 118è Congrès
National des sociétés historiques et scientifiques.
Pau: CTHS, pp.9-26.
Jéquier, J.-P. (1975). Le Moustérien alpin. Révision
critique. Eburodunum Yverdon, 2: 7-130.
Davidar, E. R. C. (1975). 8. Ecology and Behavior of the Dhole or Indian Wild dog Cuon alpinus
(Pallas). In (Fox, M. W., ed.) The wild Canids.
Their Systematics, Behavioral Ecology and Evolution. Wenatchee: Dogwise Publishing,
pp.109-119.
Del Campana, D. (1923). Sopra un Cuon e una Mustela
del Quaternario di Equi (Alpi Apuane). Atti della
Reale Accademia Nazionale dei Lincei. Rendiconti
della Classe di scienze fisiche, matematiche e naturali, 32: 170-172.
Del Campana, D. (1954). Carnivori quaternari della
Tecchia e della Caverna di Equi nelle Alpi Apuane (Mustelidi, Canidi, Felidi). Palaeontographia
Italica, 44: 1-42.
Delpech, F. (1988). Les grands Mammifères, à l'exception des Ursidés. In (Rigaud, J.-P., ed.) La
Grotte Vaufrey Paléoenvironnement - Chronologie
- Activités Humaines. Mémoires de la Société Préhistorique Française, pp.213-289.
Denys, C. (2011). Des référentiels en taphonomie des
petits vertébrés: bilan et prespectives. In (Laroulandie, V., Mallye, J.-B. & Denys, C., eds.)
Taphonomie des Petits vertébrés: Référentiels et
Transferts aux fossiles. Actes de la Table Ronde du RTP Taphonomie, Talence 20-21 octobre 2009.
Oxford: BAR International Series, pp.7-22. Denys, C., Kowalski, K. & Dauphin, Y. (1992).
Mechanical and chemical alterations of skeletal
tissues in a recent Saharian accumulation of faeces from Vulpes rueppelli (Carnivora, Mammalia).
Acta Zoologica cracovensis, 35(2): 265-283.
Dodson, P. & Wexlar, D. (1979). Taphonomic investigations of owl pellets. Paleobiology, 5(3):
275-284.
Dufour, R. (1989). Les Carnivores pléistocènes de la caverne de Malarnaud (Ariège) : collection E.
Harlé, Museum d'Histoire Naturelle de
Bordeaux. Muséum d'Histoire Naturelle Bordeaux, Bordeaux.
Durbin, L. S., Venkataraman, A. B., Hedges, S. &
Duckworh, W. (2004). Dhole. Cuon alpinus (Pallas, 1811). In (Sillero-Zubiri, C., Hoffman, M.
& Macdonald, D. W., eds.) Canids: Foxes, Wolves,
Jackals and Dogs. 2004 Status Survey And Conservation Action Plan, IUCN/SSC Canid
Specialist Group. pp.210-219.
Esteban-Nadal, M., Cáceres, I. & Fosse, P. (2010). Characterization of a current coprogenic sample
originated by Canis lupus as a tool for identifying
a taphonomic agent. Journal of Archaeological Science, 37: 2959-2970.
Ewer, R. F. (1998). The carnivores. Cornell University
Press, Ithaca, New York.

336
Dhole as a bone accumulator at Noisetier Cave
Mallye, J.-B., Laroulandie, V. & Cochard, D. (2008). Accumulations osseuses en périphérie de terriers
de petits carnivores : les stigmates de prédation et
de fréquentation. Annales de Paléontologie, 94: 187-208.
Mallye, J.-B., Thiébaut, C., Mourre, V., Costamagno, S.,
Claud, E. & Weisbecker, P. (2012). The Mousterian
bone retouchers of Noisetier Cave: experimentation
and identification of marks. Journal of
Archaeological Science, 39(4): 1131-1142. Marín Arroyo, A. B., Fosse, P. & Vigne, J.-D. (2009).
Probable evidences of bone accumulation by
Pleistocene bearded vulture at the archaeological site of El Mirón Cave (Spain). Journal of
Archaeological Science, 36: 284-296.
Marín Arroyo, A. B. & Margalida, A. (2012). Distinguish Bearded Vulture activities within
Archaeological Contexts: Identification guidelines.
International Journal of Osteoarchaeology, 22(5): 563-576.
Matthews, T. (2006). Taphonomic Characteristics of
Micrommals Predated by Small Mammalian Carnivores in South Africa: Application to Fossil
Accumulations. Journal of Taphonomy, 4(3): 143-161.
Maureille, B., Bruxelles, L., Colonge, D., Costamagno, S., Cravinho, S., Jeannet, M., Laroulandie, V.,
Thiébaut, C. & Mourre, V. (2007). Nouveaux vestiges humains moustériens de la Grotte du
Noisetier (Fréchet-Aure, Hautes-Pyrénées).
1833ème Réunion scientifique de la société d'Anthropologie de Paris (23-25 janvier 2008),
Marseille. Bulletin et Mémoires de la société
d'Anthropologie de Paris. Moigne, A.-M., Palombo, M.-R., Belda, V., Heriech-
Briki, D., Kacimi, S., Lacombat, F., de Lumley, M.
-A., Moutoussamy, J., Rivals, F., Quilès, J. & Testu, A. (2006). Les faunes de grands
mammifères de la Caune de l'Arago (Tautavel)
dans le cadre biochronologique des faunes du Pléistocène moyen italien. L'Anthropologie, 110:
788-831.
Moncel, M.-H., Brugal, J.-P., Prucca, A. & Lhomme, G. (2008). Mixed occupation during the Middle
Palaeolithic: case study of a small pit-cave-site of
Les Pêcheurs (Ardèche, south-eastern France). Journal of Anrthropological Archaeology, 27: 382-
398.
Mondini, M. (1995). Artiodactyl prey transport by foxes in Puna rock shelters. Current Anthropology,
36(3): 520-524.
Mondini, M. (2000). Tafonomia de abrigos rocosos de
la Puna. Formación de conjuntos escatológicos por
zorros y sus implicaciones arqueológicas.
Archaeofauna, 9: 151-164.
Johnsingh, A. J. T. (1982). Reproductive and social behaviour of the Dhole, Cuon alpinus (Canidae).
Journal of Zoology, London, 198: 443-463.
Kahlke, H. D. (1961). Revision der Säugetierfaunen der klassischen deutschen Pleistozän-Fundestellen
von Süßenborn, Mosbach und Taubach. Geologie,
10: 493-532.
Karanth, U. K. & Sunquist, L. E. (1995). Prey
Selection by Tiger, Leopard and Dhole in Tropical
Forests. The Journal of Animal Ecology, 64(4): 439-450.
Kawanishi, K. & Sunquist, M. E. (2008). Food habits
and activity patterns of the Asiatic golden cat (Catopuma temminckii) and the dhole (Cuon
alpinus) in a primary rainforest of Peninsula
Malaysia. Mammal Study, 33: 173-177. Klippel, W. E., Snyder, L. M. & Parmalee, P. W.
(1987). Taphonomy and Archaeologically
recovered Mammal Bone from Southeast Missouri. Journal of Ethnobiology, 7(2): 155-169.
Kumaraguru, A., Saravanamuthu, R., Brinda, K. &
Asokan, S. (2011). Prey preference of large carnivores in Anamalai Tiger Reserve, India.
European Journal of Wildlife Research, 57: 627-
637. Lloveras, L., Moreno Garcia, M. & Nadal, J. (2008b).
Taphonomic study of leporid remains accumulated by the Spanish Imperial Eagle (Aquila adalberti).
Geobios, 41: 91-100.
Lloveras, L., Moreno Garcia, M. & Nadal, J. (2011). Feeding the Foxes: An experimental study to
assess their taphonomic signature on Leporid
R em a in s . In t e rn a t io n a l Jo u rn a l o f osteoarchaeology, 22(5): 577-590.
Lloveras, L., Moreno-García, M. & Nadal, J. (2008a).
Taphonomic analysis of leporid remains obtained from modern Iberian Lynx (Lynx pardinus) scats.
Journal of Archaeological Science, 35: 1-13.
Lyman, R. L. (1994). Vertebrate Taphonomy. Cambridge University Press, Cambridge.
Malez, M. (1974). Noviji Rezultati Istrazivanja
Paleolitika u Velikoj pecini, Veternici i Sandalji. - Neue Ergebnisse der Paläolithikum-Forschungen
in Velika Pecina, Veternica und Sandalja
(Kroatien). Jugoslavenske Akademija Znanosti i Umjetnosti: 7-44.
Malez, M. (1975). Die Höhle Vindija - Eine neue
Fundstelle fossiler Hominiden in Kroatien. Bulletin Scientifique, Section A, 20: 139-140.
Malez, M. & Malez, V. (1989). The Upper Pleistocene
Fauna from the Neanderthal Man Site in Krapina
(Croatia, Yugoslavia). Geoloski vjesnik instituta za
geoloska istrazivanja u zagrebu i hrvatskog
geoloskob drustva, 42: 49-57.

337
Mallye et al.
Pokines, J. T. & Kerbis Peterhans, J. C. (2007). Spotted hyena (Crocuta crocuta) den use and taphonomy
in the Masai Mara National Reserve, Kenya.
Journal of Archaeological Science, 34: 1914-1931. Prucca, A. (2003). Caractérisation de l'impact des
Loups sur les ossements d'Herbivores (Cerfs de
Virginie, Orignaux, Bisons) : Étude des
modifications infligées par des Loups captifs et
sauvages Nord-Américains. Unpublished Master
thesis, Université de Provence. 135 p. Richardson, P. R. K. (1980). Carnivore Damage to
Antelope Bones and its Archaeological Implications.
Palaeontologica Africana, 23: 109-125. Robert, I. & Vigne, J.-D. (2002). The Bearded Vulture
(Gypaetus barbatus) as an Accumulator of
archaeological Bones. Late Glacial Assembalges and Present-day Reference Data in Corsica
(Western Mediterranean). Journal of
Archaeological Science, 29: 763-777. Schmitt, D. N. & Juell, K. E. (1994). Toward the
Identification of coyote Scatological Faunal
accumulations in Archaeological Contexts. Journal of Archaeological Science, 21(2): 249-262.
Simons, E. L. & Ettel, P. C. (1970). Gigantopithecus.
Scientific American: 77-85. Sosnovskii, I. P. (1967). Bredding the Red dog or
dhole Cuon alpinus at Moscow Zoo. International Zoo Yearbook, 7: 120-122.
Stiner, M. C. (1994). Honor among Thieves. A
zooarchaeological study of Neandertal ecology. Princeton University Press, Princeton.
Stiner, M. C. (2004). Comparative ecology and
taphonomy of spotted hyenas, humans, and wolves in Pleistocene Italy. Revue de Paléobiologie,
Genève, 23(2): 771-785.
Stiner, M. C., Munro, N. D. & Sanz, M. (2012). Carcass damage and digested bone from mountain lions
(Felis concolor): implications for carcass persistence
on landscapes as a function of prey age. Journal of Archaeological Science, 39: 896-907.
Thenius, E. (1954). Die Caniden (Mammalia) aus dem
Altquartär von Hundsheim (Niederösterreich) nebst Bemerkungen zur Stammesgeschichte der
Gattung Cuon. Neues Jahrbuch für Geologie und
Paläontolologie, 99: 230-286. Thiébaut, C., Mourre, V., Chalard, P., Colonge, D.,
Coudenneau, A., Deschamps, M. & Sacco-
Sonador, A. (2012). Lithic technology of the final Mousterian on both sides of the Pyrenees.
Quaternary International, 247: 182-198.
Thinley, P., Kamler, J. F., Wang, S. W., Lham, K.,
Stenkewitz, U. & Macdonald, D. W. (2011). Seasonal
diet of dholes (Cuon alpinus) in northwestern Bhutan.
Mammalian Biology, 76: 518-520.
Montalvo, C. I., Pessino, M. E. M. & Bagatto, F. C. (2008). Taphonomy of the bones of rodents
consumed by Andean hog-nosed skunks
(Conepatus chinga, Carnivora, Mephitidae) in central Argentina. Journal of Archaeological
Science, 35: 1481-1488.
Mourre, V., Costamagno, S., Bruxelles, L., Colonge, D.,
Cravinho, S., Laroulandie, V., Maureille, B., Thiébaut,
C. & Viguier, J. (2008a). Exploitation du milieu
montagnard dans le Moustérien final: La grotte du Noisetier à Fréchet-Aure (Pyrénées centrales
françaises). In (Grimaldi, S., Perrin, T. & Guilaine, J.,
eds.) Mountains environments in Prehistoric Europe. BAR International Series, pp.1-10.
Mourre, V., Costamagno, S., Thiébaut, C., Allard, M.,
Bruxelles, L., Colonge, D., Cravinho, S., Jeannet, M., Juillard, F., Laroulandie, V. & Maureille, B.
(2008b). Le site moustérien de la Grotte du
Noisetier à Fréchet-Aure (Hautes-Pyrénées) - premiers résultats des nouvelles fouilles. In
(Jaubert, J., Bordes, J.-G. & Ortega, I., eds.) Les
sociétés du Paléolithique dans un Grand Sud-Ouest de la France : nouveaux gisements,
nouveaux résultats, nouvelles méthodes. Mémoire
de la Société préhistorique française, pp.189-202. Nasti, A. (2000). Modification of Vicuña carcasses in
high-altitude deserts. Current Anthropology, 41(2): 279-283.
Nehring, A. (1891). Diluviale Reste von Cuon, Ovis,
Saiga, Ibex und Rupicapra aus Mähren. Neues Jahrbuch fiir Mineralogie, Geologie und
Paleontologie, 2: 107-155.
Pailhaugue, N. (1995). La faune de la salle Monique, Grotte de la Vache (Alliat, Ariège). Bulletin de la
société Préhistorique de l'Ariège, L: 225-289.
Payne, S. & Munson, P. J. (1985). Ruby an dhow many Squirrels? The destruction of Bones by Dogs. In
(Fieller, N. R. J., Gilbertson, D. E. & Ralph, N. G.
A., eds.) Palaeobiological Investigations. Research Design Method and Data Analysis. BAR
international Series, pp.31-39.
Pérez Ripoll, M., Morales Pérez, J. V., Sanchis Serra, A., Aura Tortosa, J. E. & Sarrión Montañana, I.
(2010). Presence of the genus Cuon in upper
Pleistocene and initial Holocene sites of the Iberia Penninsula: new remains identified in
archaeological contexts of the Mediterranean
region. Journal of Archaeological Science, 37(3): 437-450.
Pionnier-Capitan, M., Bémilli, C., Bodu, P., Célérier,
G., Ferrié, J.-G., Fosse, P., Garcia, M. & Vigne, J.-
D. (2011). New evidence for Upper Palaeolithic
dogs in South-Western Europe. Journal of
Archaeological Science, 38: 2123-2140.

338
Dhole as a bone accumulator at Noisetier Cave
Trinkaus, E., Marks, A. E., Brugal, J.-P., Bailey, S. E., Rink, W. J. & Richter, D. (2003). Later Middle
Pleistocene human remains from the Almonda
Karstic system, Torres Novas, Portugal. Journal of Human evolution, 45: 219-226.
Venkataraman, A. B., Arumugam, R. & Sukumar, R.
(1995). The foraging ecology of dhole (Cuon
alpinus) in Mudumalai Sanctuary, southern India.
Journal of Zoology, London, 237: 543-561.
Villa, P. & Bartram, L. E. (1996). Flaked bone from hyena den. PALEO, 8: 143-159.
Villa, P. & Soressi, M. (2000). Stone tool in carnivore
sites: the case of Bois Roche. Journal of Archaeological Research, 56: 187-211.
Wang, S. W. & Macdonald, D. W. (2009). Feeding habits and niche partitioning in a predator guild
composed of tigers, leopards and dholes in a
temperate ecosystem in central Bhutan. Journal of Zoology, 277: 275-283.
Wetzel, R., Bosinski, G. & Filzer, P. (1969). Die
Bocksteinschmiede im Lonetal (Markung
Rammingen, Kreis Ulm). Veröffentlichungen des
Staatlichen Amtes für Denkmalpflege Stuttgart / A,
Vor- und Frühgeschichte, Stuttgart. Yravedra, J., Lagos, L. & Bárcena, F. (2011). A
taphonomic study of Wild Wolf (Canis lupus)
modification of horse bones in Northwestern Spain. Journal of Taphonomy, 9: 37-65.




















