
Taphonomy of the Irregular Echinoid Clypeaster humilis from the Red Sea: Implications for taxonomic...
15
Figure 6.1. Location of study area. 114 Nebelsick
-
Upload
uni-tuebingen -
Category
Documents
-
view
5 -
download
0
Transcript of Taphonomy of the Irregular Echinoid Clypeaster humilis from the Red Sea: Implications for taxonomic...
Figure 6.1. Location of study area.
114 Nebelsick

TAPHONOMY OF THE IRREGULAR
ECHINOID CLYPEASTER HUMILIS
FROM THE RED SEA: IMPLICATIONS
FOR TAXONOMIC RESOLUTION
ALONG TAPHONOMIC GRADES
James H. Nebelsick
Taphonomy desc r ibes t he t rans i t ion o f skele ta l r e m a i n s f rom o n c e l iv ing
organisms to t he preserved r e m a i n s tha t pa leon to log i s t s u l t ima te ly r ecove r
a t the ou tc rop and store in the i r c o l l e c t i o n s . U n d e r s t a n d i n g t a p h o n o m i c
gradients is of essen t ia l i m p o r t a n c e for pa leon to log i s t s w h o use fossil re
mains to m a k e s t a t emen t s c o n c e r n i n g s u c h th ings a s a spec t s o f eco logy ,
diversity, and evolu t ion . A c t u a l i s t i c ( observa t ions c a n m a k e f u n d a m e n t a l
contr ibut ions to unde r s t and ing t a p h o n o m i c gradients for t he s imple reason
that the c o m p l e x e c o l o g i c a l i n t e rac t ions tha t af fect t a p h o n o m i c p rocesses
and features c a n be d i rec t ly obse rved and m e a s u r e d .
E c h i n o d e r m s no t on ly have an i l lus t r ious past bu t a lso f lour i sh today
in diverse m a r i n e e n v i r o n m e n t s . T h u s , we c a n c o n d u c t ac tua l i s t i c observa
tions o f e c h i n o d e r m s and d i rec t ly invest igate those e c o l o g i c a l factors tha t
affect no t on ly the i r d is t r ibut ion bu t a lso t he preserva t ion o f the i r r e m a i n s
after death . A l t h o u g h e c h i n o i d s c o m m o n l y d isar t icu la te rapidly d e p e n d i n g
on a n u m b e r o f factors (see Kie r , 1977 ; Al l i son , 1 9 9 0 ; Kidwel l and B a u -
mil ler , 1 9 9 0 ; D o n o v a n , 1 9 9 1 ; G r e e n s t e i n , 1 9 9 3 ) , c lypeas te ro ids are c o m
paratively robus t and stay c o m p l e t e l ong e n o u g h for t a p h o n o m i c factors to
be obse rved on t he sur face of t he test. E c h i n o d e r m t a p h o n o m y is a b road
subjec t and has b e e n t h e s u b j e c t o f a n u m b e r o f reviews by L e w i s ( 1 9 8 0 ) ,
D o n o v a n ( 1 9 9 1 ) , B r e t t e t al . ( 1 9 9 7 ) , A u s i c h ( 2 0 0 1 ) , and m o s t r e c e n t l y N e b
elsick ( 2 0 0 4 ) .
C lypeas t e ro ids b e l o n g to t he younges t g roup o f e c h i n o d e r m s to have
evolved, o r i g i n a t i n g in t h e P a l e o g e n e f rom cass idu lo id a n c e s t o r s (see
S m i t h , 2 0 0 1 ) . T h e y have a n u m b e r o f s p e c i a l i z e d fea tures , i n c l u d i n g the i r
cha rac te r i s t i c f l a t t ened shape , sp ine m i n i a t u r i z a t i o n and d i f fe rent ia t ion ,
c o n c e n t r a t i o n o f respiratory pod ia i n t he pe ta l s on t he abora l sur face , s e c
ondary un ipo res no t res t r ic ted to a m b u l a c r a , food grooves on t he oral sur
face , l unu l e s in t h e test of m a n y sand dol lars , and a s p e c i a l i z e d Aris tot le 's
l an te rn ( D u r h a m , 1955 , 1 9 6 6 ; M o o i , 1 9 8 9 ) . S tab i l i ty to t he test i s g iven by
in te rna l supports i n m o s t t axa , w h i c h c o n n e c t t he oral and abora l sides o f
the test. I n t e r l o c k i n g plates fu r ther s t r e n g t h e n t h e test. T h e s e fea tures
m a k e c lypeas te ro ids a m o n g t he m o s t robus t o f al l e c h i n o d e r m s , a l l o w i n g
t h e m to survive even in h i g h - e n e r g y sho re face e n v i r o n m e n t s . As of ten
Taphonomy of the Irregular Echinoid Clypeaster humulis 115
Introduction
6

Figure 6.2. Bottom to
pography of the study
area with main collection
sites of complete
Clypeaster humilis as
well as the location of
bulk sediment samples
containing Clypeaster
remains.
no ted , t h e s e s t ab i l i z ing skele ta l fea tures c o r r e s p o n d i n g l y i n c r e a s e their
p rese rva t ion po t en t i a l ( S e i l a c h e r , 1 9 7 9 ; N e b e l s i c k a n d K a m p f e r , 1994;
B r e t t e t al . , 1997 ; Mof fa t and B o t t j e r , 1 9 9 9 ; N e b e l s i c k , 1 9 9 9 ) .
T h e genus Clypeaster L a m a r c k , 1 8 0 1 , l ives today in m a n y tropical
m a r i n e e n v i r o n m e n t s ( G h i o l d and H o f f m a n n , 1 9 8 4 , 1 9 8 6 ) . T h e y have a
wide m o r p h o l o g i c a l var ie ty a n d i n c l u d e f l a t t ened i n f auna l a nd domed
ep i fauna l forms. T h e la t ter i s r ep resen ted by t he type spec ies Clypeaster
rosaceous ( L i n n a e u s , 1758 ) , wel l k n o w n f rom sha l low C a r i b b e a n waters.
Clypeaster i s an i m p o r t a n t o r g a n i s m in t h e s e e n v i r o n m e n t s . F o r example ,
116 Nebelsick

they can i n f l u e n c e s e d i m e n t pa r t i c l e s ize w h e n f eed ing by c r u s h i n g sand-
sized par t ic les and p r o d u c i n g f ine r s e d i m e n t s ( K a m p f e r and T e r t s c h n i g ,
1992).
Clypeaster i s also o n e of the mos t c o m m o n ech ino ids in C e n o z o i c shal
low-water sed iments . Hundreds of n o m i n a l fossil species have b e e n desc r ibed
(Durham, 1 9 6 6 ) , and n u m e r o u s s p e c i m e n s are k n o w n from fossil e c h i n o i d
assemblages. C o m m o n l y , different spec ies of Clypeaster o c c u p y different
habitats showing co r respond ing variat ions in size and m o r p h o l o g y (i.e., B o g -
gild and Rose , 1 9 8 4 ; Poddubiuk and R o s e , 1 9 8 4 ; Ne raudeau e t a l , 2 0 0 1 ;
Kroh and Nebe l s i ck , 2 0 0 3 ) .
C L Y P E A S T E R H U M I L I S I N T H E S T U D Y A R E A . Clypeaster humilis ( L e s k e ,
1778) is a m e d i u m - s i z e d spec ies tha t is d is t r ibuted t h r o u g h o u t t h e I n d o -
West Paci f ic r e a l m (C la rk and R o w e , 1971) . T h e ske le tons o f C . humilis
examined he r e o r ig ina te f rom the N o r t h e r n B a y o f S a f a g a , R e d S e a , E g y p t
(Figs. 6 . 1 , 6 . 2 ) . T h i s is a c o m p l e x a rea abou t 10 by 8 km l o n g wi th coas t a l
indentat ions, is lands, and bas ins separa ted f rom o n e a n o t h e r and f rom the
open R e d S e a by s u b m a r i n e r idges (P i l l e r and Pervesler , 1 9 8 9 ; P i l l e r a n d
Mansour , 1 9 9 0 ) . T h e N o r t h e r n B a y o f Safaga i s a shal low-water , c a r b o n a t e
env i ronmen t and i s c h a r a c t e r i z e d by a wide r ange of b o t t o m fac ies . S h a l
lower areas o f t he bay are d o m i n a t e d by c o a r s e sands , sea grass m e a d o w s ,
sands with sea grass, and sands wi th co ra l pa t ches . C o r a l reefs are a lso
present and are a c c o m p a n i e d by r e e f f la t s . However , m o s t co ra l cove r i s
represented by a co ra l c a r p e t tha t o c c u p i e s ex tens ive areas of t h e seafloor.
D e e p e r bas ins are o c c u p i e d by c a r b o n a c e o u s m u d d y and sandy m u d d y
sediments (F ig . 6 . 2 ) . B o t h regu la r and i r regula r e c h i n o i d s are c o m m o n
macrofauna l e l e m e n t s o f th is shal low-water c a r b o n a t e sys tem a n d have
been the s u b j e c t o f a n u m b e r o f ac tua l i s t i c s tudies c o n c e r n i n g the i r e c o l
ogy and t a p h o n o m y (e.g., N e b e l s i c k , 1 9 9 2 a , 1 9 9 2 b , 1 9 9 9 ; N e b e l s i c k and
Kampfer , 1 9 9 4 ; N e b e l s i c k and Kowalewsk i , 1 9 9 9 ) .
Clypeaster i s a m o n g t he m o s t c o m m o n e c h i n o i d s in t he N o r t h e r n B a y
o f Safaga and i s found as l iv ing s p e c i m e n s , dead tests, a n d f r agmen t s . Of
the four sympa t r i c spec ies in the study area , Clypeaster humilis is by far t he
most c o m m o n and total ly d o m i n a t e s t he c o l l e c t e d s p e c i m e n s . I n a l l , 4 7
living s p e c i m e n s and 86 dead s p e c i m e n s were c o l l e c t e d . Clypeaster fervens
Koeh le r , 1 9 2 2 , Clypeaster reticulatus ( L i n n a e u s , 1 7 5 8 ) , a n d Clypeaster
rarispinus de M e i j e r e , 1 9 0 2 , are m u c h less c o m m o n , and on ly a few spec i
mens o f e a c h spec i e s were c o l l e c t e d ( N e b e l s i c k , 1 9 9 2 b , 1 9 9 9 ) . S p e c i e s dif
ferent iat ion i s based on shape and size re la t ionships of test s ize , p e t a l o d i u m
length , a m b i t u s shape , and t he form o f t he frontal a m b u l a c r u m (C la rk and
Rowe, 1971) .
C o m p l e t e Clypeaster humilis s p e c i m e n s were found in shal low-water
areas of t h e bay wi th sandy substrates (F ig . 6 .1) . L i v i n g Clypeaster were
found bu r i ed just u n d e r n e a t h the s e d i m e n t sur face , rarely b e i n g just visible
as an o u t l i n e b e l o w t h e sur face , a t m a x i m u m dens i t ies o f two individuals
per square m e t e r ( N e b e l s i c k , 1 9 9 2 a ) . D e a d e c h i n o i d tests were d i scovered
on the s e d i m e n t sur face , par t ia l ly bu r i ed , o r u n d e r n e a t h t he sur face . S o m e
s p e c i m e n s were ex t r ac t ed by rak ing t he s e d i m e n t wi th a s tout rake p e n e
trat ing to a dep th of approx ima te ly 5 cm into the s e d i m e n t .
Taphonomy of the Irregular Echinoid Clypeaster humulis 117
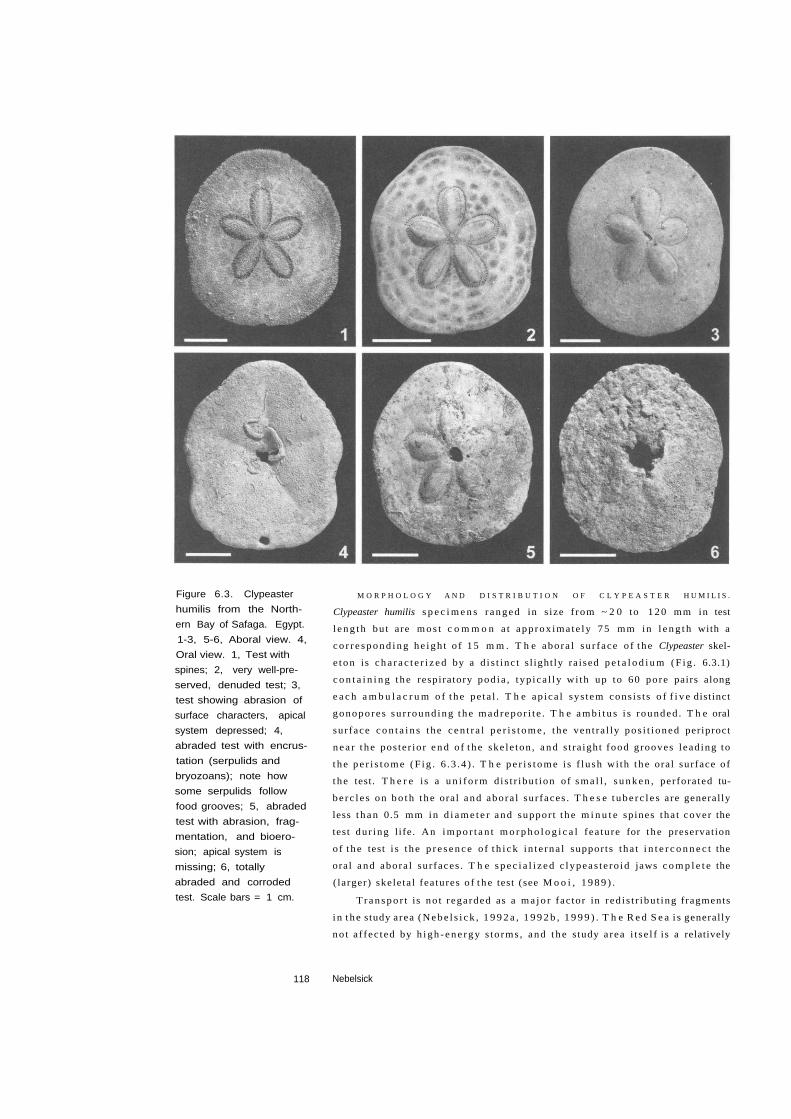
Figure 6.3. Clypeaster
humilis from the North
ern Bay of Safaga. Egypt.
1-3, 5-6, Aboral view. 4,
Oral view. 1, Test with
spines; 2, very well-pre
served, denuded test; 3,
test showing abrasion of
surface characters, apical
system depressed; 4,
abraded test with encrus
tation (serpulids and
bryozoans); note how
some serpulids follow
food grooves; 5, abraded
test with abrasion, frag
mentation, and bioero-
sion; apical system is
missing; 6, totally
abraded and corroded
test. Scale bars = 1 cm.
M O R P H O L O G Y A N D D I S T R I B U T I O N O F C L Y P E A S T E R H U M I L I S .
Clypeaster humilis s p e c i m e n s r a n g e d in s ize f rom ~ 2 0 to 1 2 0 mm in test
l e n g t h bu t are m o s t c o m m o n a t app rox ima te ly 75 mm in l e n g t h with a
c o r r e s p o n d i n g h e i g h t o f 15 m m . T h e abora l su r face o f t he Clypeaster skel
e ton is c h a r a c t e r i z e d by a d i s t inc t s l ight ly ra ised p e t a l o d i u m (F ig . 6.3.1)
c o n t a i n i n g t he respiratory pod ia , typ ica l ly wi th up to 60 pore pairs along
e a c h a m b u l a c r u m o f t he pe ta l . T h e ap ica l sys tem cons is t s o f f i v e distinct
gonopores su r round ing t he madrepor i t e . T h e a m b i t u s i s rounded . T h e oral
sur face c o n t a i n s the c e n t r a l p e r i s t o m e , t he vent ra l ly pos i t i oned periproct
n e a r t he pos ter ior e n d of t he ske le ton , a n d s t ra ight food grooves l ead ing to
t he p e r i s t o m e (F ig . 6 .3 .4 ) . T h e p e r i s t o m e i s f lush wi th t h e oral surface o f
t he test. T h e r e is a u n i f o r m d is t r ibu t ion of s m a l l , s u n k e n , perforated tu
be r c l e s on b o t h t he oral and abora l sur faces . T h e s e t u b e r c l e s are generally
less t h a n 0.5 mm in d i a m e t e r and suppor t t h e m i n u t e spines tha t cover the
test dur ing l ife. An i m p o r t a n t m o r p h o l o g i c a l fea ture for t he preservation
o f t he test i s t he p r e s e n c e o f t h i c k in t e rna l suppor ts tha t i n t e r c o n n e c t the
oral and abora l sur faces . T h e s p e c i a l i z e d c lypeas te ro id jaws c o m p l e t e the
( larger) skele ta l features o f t h e test (see M o o i , 1 9 8 9 ) .
T r a n s p o r t is no t regarded as a m a j o r fac tor in red i s t r ibu t ing fragments
i n t he study area ( N e b e l s i c k , 1 9 9 2 a , 1 9 9 2 b , 1 9 9 9 ) . T h e R e d S e a i s generally
no t a f fec ted by h i g h - e n e r g y s to rms , and t h e study a rea i t se l f is a relatively
118 Nebelsick

protected e n v i r o n m e n t separa ted f rom the o p e n sea by is lands and under
water swells . T h e h i g h d i f ferent ia t ion o f e c h i n o i d f r agmen t s w i th in b u l k
samples and the i r c lose co r re l a t ion to t he hab i ta t o f c o r r e s p o n d i n g l iv ing
echinoids also suggest tha t t ranspor t i s no t an i m p o r t a n t t a p h o n o m i c factor
a t the baywide sca l e of inves t iga t ion (F ig . 6 . 2 ) . S m a l l - s c a l e t ranspor t i s dis
cussed in N e b e l s i c k ( 1 9 9 2 b ) and inc ludes t ranspor t f rom co ra l p a t c h e s a n d
coral c a r p e t to t h e su r round ing s e d i m e n t as well as f rom t h e reefs across
the r e e f f l a t s a n d to t h e r e e f s lopes . T h e s e pe r t a in mos t ly t o r egu la r e c h i
noids l iv ing in m o r e e x p o s e d e n v i r o n m e n t s .
P R E V I O U S S T U D I E S C O N C E R N I N G T H E T A P H O N O M Y O F C L Y P E A S T E R
H U M I L I S . T w o previous inves t iga t ions involving Clypeaster humilis f rom
the study a rea are o f i m p o r t a n c e in t he in te rpre ta t ion o f t a p h o n o m i c gra
dients p resen ted he re . N e b e l s i c k and K a m p f e r ( 1 9 9 4 ) s tudied shor t - te rm
t a p h o n o m i c p rocesses a f fec t ing test p reserva t ion of Clypeaster humilis a n d
Echinodiscus auritus L e s k e , 1 7 7 8 , by us ing an e x p e r i m e n t a l array of under
water c a g e d dead s p e c i m e n s over a t i m e pe r iod of 1.5 weeks . S p i n e disar
t i cu la t ion in b o t h spec i e s c o m m e n c e d wi th in 18 hours and was f in i shed
by 91 hours after t h e start o f t h e e x p e r i m e n t . After sp ine d i sa r t i cu la t ion ,
mos t tests r e m a i n e d s table , bu t o n c e plate d i saggrega t ion was in i t i a ted (in
th ree o f 20 cases ) , t h e tests rapidly d i sa r t i cu la ted in to larger f r agmen t s and
individual plates .
N e b e l s i c k ( 1 9 9 9 ) s tud ied t h e t a p h o n o m y o f Clypeaster f r a g m e n t s
t h r o u g h o u t t h e study area . F r a g m e n t s c o u l d on ly be ident i f ied to g e n u s
level b e c a u s e o f t h e l ack o f cha rac t e r i s t i c test fea tures n e e d e d for spec i e s
ident i f ica t ion . Mul t i va r i a t e s ta t is t ical analysis o f sur face preserva t ion fea
tures ( i nc lud ing abras ion , e n c r u s t a t i o n , and sur faces marks) of Clypeaster
f ragments led to t h e des igna t ion of four different t aphofac ies in t h e study
area ( N e b e l s i c k , 1 9 9 9 ) . T h e s e t aphofac ies were re la ted to dif ferent ia l e x p o
sure a n d s e d i m e n t a t i o n rates, e c o l o g i c a l factors tha t are no t readi ly dis
c e r n e d by t h e analysis o f diversity and m o r p h o l o g i c a l fea tures a l o n e .
F ive different p reserva t ion states a l o n g a t a p h o n o m i c g rad ien t were differ
en t i a t ed by us ing t he qual i ta t ive analysis o f t he sur face c h a r a c t e r s (F ig s .
6 . 3 - 6 . 5 ) . T h e basis o f this g rad ien t i s t he preserva t ion o f t h e fo l lowing sur
face cha rac t e r s : sp ines , t he ap ica l sys tem, gonopore s , m a d r e p o r i t e , p e t a l o -
d i u m , pore pairs , plate b o u n d a r i e s , t u b e r c l e s , and t he a m b i t u s (F ig . 6 . 5 ) .
T h i s s tage co r r e sponds to those a n i m a l s tha t have just b e e n k i l l ed , and a
few dead s p e c i m e n s were r ecove red wi th spines still a t t a c h e d by e p i t h e
l i u m ( F i g . 6 .3 .1 ) . T h e c o l o r o f t h e a n i m a l s , r a n g i n g f rom l igh t t o dark
b r o w n , i s sti l l r e c o g n i z a b l e . E v e n p e d i c e l l a r i a are ne s t ed b e t w e e n t h e
spines . T h e i n t e g u m e n t covers t he per iproc t . T h e t ee th are i n p l a c e wi th
t h e jaws still a r t i cu la ted . I t i s c l e a r tha t dead s p e c i m e n s of th is s tage c a n
k e e p the i r sp ines for on ly a few days as a resul t of t he d e c a y of soft t issue
and d i sa r t i cu la t ion o f t he spines ( N e b e l s i c k and K a m p f e r , 1 9 9 4 ) .
Taphonomic
Gradient of
Clypeaster humilis
Tests
Stage 1: Specimens
with Spines
Taphonomy of the Irregular Echinoid Clypeaster humulis 119

Figure 6.4. Fragment
preservation of
Clypeaster sp. 1, 3, 5,
Successive enlargements
of a single, well-preserved
plate. 2, 4, 6, Successive
enlargements of two
joined plates showing
high rates of abrasion and
encrustation of a serpulid
worm tube and encrust
ing foraminifera. Note
that the serpulid worm
tube has been broken
along the plate boundary,
suggesting encrustation
of a complete test and
subsequent fragmenta
tion. Scale bars: 1, 2=1
mm; 3,4 = 400 µm; 5, 6
= 100 µm
C H A R A C T E R P R E S E R V A T I O N . T h e ap ica l sys tem is c o m p l e t e (F ig . 6.3.2) .
G o n o p o r e s are o p e n and t h e madrepo r i t e i s r e c o g n i z a b l e . T h e pe ta lod ium
i s d is t inc t , and pore pairs are free of s e d i m e n t . A m b u l a c r a l a n d in te rambu
lac ra ! plate b o u n d a r i e s c a n be readi ly d i s c e r n e d . T h e a m b i t u s i s perfectly
preserved. P r i s t ine sur face c h a r a c t e r s are p resen t a t t h e m i c r o s c o p i c scale
wi th c o n s p i c u o u s , s u n k e n , per fora ted t u b e r c l e s , a n d we l l -deve loped are-
oles (F ig . 6 .4 .1) . S e c o n d a r y t u b e r c l e s a n d sur face k n o b s c a n a lso be distin
gu i shed (F ig . 6 .4 .3 ) . D i f f e ren t i a l s t e r e o m is readi ly ev iden t (F ig . 6 .4 .5 ) .
T A P H O N O M I C P R O C E S S E S . D e c a y o f c o n n e c t i v e t issue has led to the
d i sa r t i cu la t ion o f sp ines f rom the sur face o f t h e test. T h e d e c a y o f ambu la
cra l t u b e feet resul t i n o p e n pores o f t h e p e t a l o d i u m and s e c o n d a r y uni-
pores . Jaws are n o w loose w i th in t he test. T h e ba re test i s n o w whi te . M o s t
s p e c i m e n s are still free o f in t e rna l s e d i m e n t .
T A X O N O M I C R E S O L U T I O N . S p e c i m e n s c a n readi ly b e ident i f ied at the
spec ies level . C o l o r var ia t ions are , however , no t d i s ce rn ib l e .
C H A R A C T E R P R E S E R V A T I O N . T h e ap i ca l sys tem c a n b e d a m a g e d (F ig .
6 .3 .3 , 6 .3 .4 ) . G o n o p o r e s , i f p resent , are f i l l e d wi th s e d i m e n t . T h e pe ta lo
d i u m is less d i s t inc t , and m a n y pore pairs are p lugged . A m b u l a c r a l and in
t e r a m b u l a c r a ! plate b o u n d a r i e s are hardly d i sce rned . T u b e r c l e s are abraded
wi th d a m a g e d c r e n u l a t i o n s and m a m m a l o n s . S t e r e o m di f ferent ia t ion i s
120 Nebelsick
Stage 2: Very Well-
preserved,
Denuded Tests
Stage 3a:
Abraded Tests

b e c o m i n g diff icul t to r e c o g n i z e a t t he sur face . T h e a m b i t u s i s still wel l
preserved.
T A P H O N O M I C P R O C E S S E S . Abras ion has led to loss o f sur face c h a r a c t e r
resolut ion. F i r s t s igns o f e n c r u s t a t i o n and b ioe ros ion are apparent . T h e test
has t aken a dul l co lor . T h e test c a n be f i l l ed wi th s e d i m e n t .
T A X O N O M I C R E S O L U T I O N . S p e c i m e n s c a n still b e ident i f ied at t he spe
cies level b e c a u s e shape and size re la t ionships o f t he test and p e t a l o d i u m
leng th , a m b i t u s shape , and frontal a m b u l a c r u m are still r e c o g n i z a b l e .
C H A R A C T E R PRESERVATION.The ap ica l sys tem i n c l u d i n g t h e g o n o p o r e s
and m a d r e p o r i t e c a n b e c o m p l e t e l y e roded (F ig . 6 . 3 .5 ) . T h e p e t a l o d i u m i s
less d i s t inc t , and m a n y pore pairs are p lugged . A m b u l a c r a l a n d i n t e r a m
bu lac ra l plate b o u n d a r i e s are hardly d i s c e r n e d . T h e a m b i t u s i s stil l wel l
preserved. A t t h e m i c r o s c o p i c s ca l e , e n c r u s t a t i o n cover sur face c h a r a c t e r s
(Fig. 6 . 4 . 2 ) , t ube rc l e s are abraded (F ig . 6 .4 .4 ) , and s t e r e o m di f fe rent ia t ion
b e c o m e s diff icul t (F ig . 6 . 4 . 6 ) .
T A P H O N O M I C P R O C E S S E S . E n c r u s t a t i o n is by u n i l a m i n a r b r y o z o a n s ,
serpulids, and e n c r u s t i n g fo ramin i fe ra . C o n s p i c u o u s ho les on t h e test sur
face m a y b e t h e resul t o f b o r i n g act ivi ty. T h e test c a n b e f i l l e d wi th
sed imen t .
T A X O N O M I C R E S O L U T I O N . T h e gross m o r p h o l o g y is stil l r e c o g n i z a b l e
s o spec i e s iden t i f i ca t ion c o u l d b e poss ib le . T h e d e g r e e o f e n c r u s t a t i o n
dictates i f o t h e r spec i e s -d i agnos t i c features , such as t he shape o f t he frontal
a m b u l a c r u m and pe ta l pores , are still r e c o g n i z a b l e .
C H A R A C T E R P R E S E R V A T I O N . C o r r o d e d test (F ig . 6 . 3 . 6 ) . Al l su r face c h a r a c
ters are c o m p l e t e l y miss ing . T h e a m b i t u s i s co r roded . T h e tests are n o t i c e
ably heav ie r and are f i l led wi th s e d i m e n t .
T A P H O N O M I C P R O C E S S E S . T h e s e tests are heavi ly co r roded . E a r l y dia-
genes is has l ead t o t h e f i l l i n g o f s t e r e o m and l i t h i f i ca t ion o f in f i l l ing
sed imen t .
T A X O N O M I C R E S O L U T I O N . S p e c i e s and genus iden t i f i ca t ion , as s u c h ,
i s no l o n g e r poss ib le wi thou t de ta i l ed k n o w l e d g e of t he c lypeas te ro id pres
ent in t he study area . G e n e r a l fo rm and h e i g h t are still d i s c e r n i b l e , bu t t he
co r roded a m b i t u s c h a n g e s t h e l e n g t h and width pa rame te r s o f t h e spec i
m e n . Al l deta i ls c o n c e r n i n g t h e p e t a l o d i u m and pore pairs are des t royed.
Sp ines d i sappear soon after dea th after t he d e c a y o f c o n n e c t i v e t issues a n d
musc l e s tha t c o n n e c t t h e spines t o t he test. T h e m i n u t e spines ( < 2 m m
long) are t h e n c o m m i t t e d t o t he s e d i m e n t . B e c a u s e t he analysis o f b u l k
s ed imen t samples was res t r ic ted to gra in sizes larger t h a n 2 mm ( N e b e l s i c k ,
1 9 9 2 a , 1 9 9 2 b ) , t he f r e q u e n c y o f d is t r ibut ion o f spines in t h e s e d i m e n t i s no t
recorded .
T h e apica l system with the gonopores and madrepor i t e shows a steady
degradat ion from stage 1 to 4. A l though perfect ly preserved in stage 2 (F ig .
Preservation of
Surface Features
along the
Taphonomic
Gradient
Taphonomy of the Irregular Echinoid Clypeaster humulis 121
Stage 3b:
Encrusted—
Bioeroded Tests
Stage 4:
Corroded Tests

Taphonomic gradient >
1 2 3a 3b 4
Character preservation Apical system 0 0 1 2 2 Gonopores 0 0 1 2 2 Madreporite 0 0 1 2 2 Petalodium 0 0 1 0-1 2 Pore pairs 0 0 1 0-1 2 Plate boundaries 0 0 2 1 2 Tubercles 0 0 1 1 2 Ambitus 0 0 0 0 1
Taphonomic processes Denudation 0 2 2 2 2 Abrasion 0 0 1 1 2 Encrustation 0 0 1 2 0 Bioerosion 0 0 1 2 0 Corrosion 0 0 0 0 2 Diagenesis 0 0 1 1 2
Taxonomic resolution
species species species species/ genus
genus/ family
Characters: Taphonomic features: 0 = complete/well preserved/free 0 = not present 1 = fragmented/partially preserved/partially plugged 1 = present 2 = destroyed/missing/completely plugged 2 = dominant
Figure 6.5. Taphonomic
gradient character pres
ervation, taphonomic
features, and taxonomic
resolution for Clypeaster
humilis test from the
Northern Bay of Safaga,
Red Sea, Egypt.
6 .3 .2 ) , i t c a n comple te ly disappear by stage 3 and 4 . T h e gonopores are open
in stage 2 but b e c o m e plugged by s e d i m e n t or c e m e n t by stage 3 (F ig . 6.3.3).
T h e apical system is obviously not as stable as the rest of t he test as observed
in the c o n t i n u o u s degradat ion from F igu re s 6 .3 .2 to 6.3.3 to 6.3.5 to 6 .3 .6 .
T h e p e t a l o d i u m is d i s t inc t in s tage 1 a n d e spec i a l l y in s tage 2 spec i
m e n s , as t he pore pairs are free of s e d i m e n t and thus resul t in dark holes
in a stark con t ra s t to t he b r igh t pla te sur faces on t h e abora l sur face of the
Clypeaster ske le ton . By s tage 3 , however , t h e p e t a l o d i u m b e c o m e s less
p r o m i n e n t as t he a m b u l a c r a l pore pairs are p lugged by s e d i m e n t o r c e m e n t
(F ig . 6 . 2 . 3 ) . T h e m i n u t e a m b u l a c r a l s m a l l pores a t t he base o f the peta ls
n e x t to t he ap ica l sys tem also b e c o m e progressively ind i s t inc t (F ig . 6 .3 .3) .
B e c a u s e they represen t depress ion on t h e uppe r side o f t h e test , t he pore
122 Nebelsick

2 2 2 2 2 2 2 1
2 2 0 0 2 2
genus/ family |
ic features: ssent t
int
>nopores are open
tage 3 (Fig. 6.3.3).
ie test as observed
to 6.3.5 to 6.3.6.
in stage 2 speci -
iult in dark holes
ral surface of the
m becomes less
iment or c e m e n t
ase of the petals
tinct (Fig. 6 .3 .3) .
he test, the pore
pair rows are c o m m o n l y preferred sites o f e n c r u s t a t i o n , w h i c h t h e n m a s k
the pore pairs (F ig . 6 . 3 .5 ) . T h e s l ight ou t l i ne o f t he pe ta l s i s bare ly r ecog
n izab le in t h e s tage 4 s p e c i m e n s (F ig . 6 . 3 . 6 ) .
A l t h o u g h plate b o u n d a r i e s are readi ly vis ible in l iv ing a n d s tage 1
s p e c i m e n s , t h e y b e c o m e even m o r e d i s t inc t i n t h e d e n u d e d s tage 2 e x a m
ples. P la te b o u n d a r i e s are n o t i c e a b l y l ighter in co lor , whereas t h e in ter ior
of t he plates i s darker, l e ad ing to a d i s t inc t pa t te rn on t h e sur face of t he
skele ton (F ig . 6 . 3 . 2 ) . P la te b o u n d a r i e s b e c o m e ind i s t inc t by s tage 3 , al
t h o u g h they c a n just b e r e c o g n i z e d . F o r e x a m p l e , i n F i g u r e 6 .3 .5 , t he plate
b o u n d a r i e s are s l ight ly depressed c o m p a r e d to t he plate cen te r s . T h e pro
gressive degrada t ion o f t he test sur face by abras ion a n d co r ros ion p rec lude
any r e c o g n i t i o n of pla te b o u n d a r i e s in s tage 4 (F ig . 6 . 3 . 6 ) .
T h e fate o f t h e t ube rc l e s c a n bare ly be d i s c e r n e d a t a m a c r o s c o p i c
sca le . T h e y are still o c c u p i e d by spines in s tage 1 . In s tage 2 , t h e t u b e r c l e s
are wel l def ined and d is t inc t ; by s tage 3 , t hey start b e c o m i n g ind i s t inc t ,
and they d i sappear total ly by s tage 4 . T h e s e c h a n g e s c a n bes t be s e e n a t a
m i c r o s c o p i c level in F i g u r e s 6 .4 .1 , 6 .4 .3 , and 6 .4 .5 . We l l -p re se rved test sur
faces have d i s t i nc t s u n k e n t u b e r c l e s wi th d e e p e n e d a r eo l e s , a d i s t i nc t
c r e n u l a t e d boss , a n d p r o m i n e n t , pe r fora ted m a m m e l o n s . T h e s e t h r e e
s t ruc tures are d i s t inc t b e c a u s e t hey are c o n s t r u c t e d o f different s t e r e o m
types , i n c l u d i n g c o a r s e l a b y r i n t h i c s t e r eom o f t he s u n k e n a reo le a n d gal -
le r ied s t e r e o m c o r r e s p o n d i n g to t h e m u s c l e a t t a c h m e n t areas . As i l lus
trated in F i g u r e 6 .4 .5 , t he m a m m e l o n i s c o n s t r u c t e d of a sparsely per fo
rated d e n s e s t e reom. T h e a rea b e t w e e n t he t u b e r c l e s i s c h a r a c t e r i z e d by
s e c o n d a r y t ube rc l e s a nd p r o t u b e r a n c e s . T h e s t e r e o m i s c o m p l e t e l y free o f
s e d i m e n t a n d c e m e n t a t i o n .
An abraded and e n c r u s t e d s p e c i m e n i s dep ic t ed in F i g u r e s 6 . 4 . 2 , 6 .4 .4 ,
and 6 . 4 . 6 . T h e t u b e r c l e s are no t on ly abraded bu t a lso cove red by an en
c rus t ing serpul id w o r m t u b e ( i t se l f ab raded) a n d e n c r u s t i n g fo ramin i fe ra .
A l t h o u g h p r i m a r y t ube rc l e s are abraded , p r o t u b e r a n c e s b e t w e e n t h e tu
be rc l e s c a n still be r e c o g n i z e d (F ig . 6 . 3 . 2 ) . As s e e n in F i g u r e 6 .4 .4 , t h e pri
m a r y t u b e r c l e s are less d i s t inc t , and t h e a reo les are par t ia l ly f i l l ed wi th
s e d i m e n t par t i c les . Abras ion o f t he t u b e r c l e i s typ ica l ly a c c o m p a n i e d by
the s h e a r i n g o f f o f t h e m a m m e l o n . T h e pore space b e t w e e n t he s t e r e o m
b e c o m e s r e d u c e d due probably t o d i a g e n e t i c a c c r e t i o n o f c e m e n t o n t h e
struts o f t h e s t e r e o m (F ig . 6 . 3 . 6 ) .
O t h e r features i nc lude the a m b i t u s o f t he c lypeas te ro id skele ta l , w h i c h
r e m a i n s s table t h r o u g h o u t t he t a p h o n o m i c grad ien t unt i l s tage 5 , w h e n t he
p e r i m e t e r o f t he ske le ton starts t o b e c o m e co r roded . T h e oral su r face has
the s a m e gene ra l t e n d e n c i e s a s t h e abora l sur face . In teres t ingly , t h e serpu
lid w o r m t u b e s fol low t h e food grooves in s tage 3a ( F i g . 6 .3 .4 ) . By s tage 4 ,
t he food grooves b e c o m e i n d i s t i n g u i s h a b l e , a n d t h e b o u n d a r i e s o f t h e
pe r ip roc t and p e r i s t o m e also b e c o m e ind i s t inc t .
T h e r e are obvious ly two e n d m e m b e r s o f t h e t a p h o n o m i c g rad ien t de
sc r ibed he re , wi th sp ine -cove red skele tons on t he o n e h a n d and tota l ly
c o r r o d e d s p e c i m e n s on t h e o ther . T h e s e a lso r ep resen t a t e m p o r a l se -
Discussion
Taphonomy of the Irregular Echinoid Clypeaster humulis 123

q u e n c e wi th a c l e a r b e g i n n i n g and an end . S p i n e loss is ub iqu i tous in the
we l l -oxygena ted , shal low, ag i ta ted waters o f t h e study area . T h e loss o f sur
face cha rac t e r s t h r o u g h abras ion is obvious ly l inear . However , encrusted
and b i o e r o d e d s p e c i m e n s c a n have wel l -preserved sur face cha rac te r s , but
converse ly , ab raded s p e c i m e n s show no o r l i t t le e n c r u s t a t i o n . T h u s , there
i s a m o s a i c d e v e l o p m e n t of t a p h o n o m i c features p resen t on t he test surface
for s tage 3 (F ig . 6 . 5 ) . T o w a r d t he e n d of t h e t a p h o n o m i c pathway, all fea
tures p e r t a i n i n g to t he test su r face are c o m p l e t e l y des t royed, eradicating
al l e v i d e n c e o f e n c r u s t a t i o n a n d b i o e r o s i o n . T h e g e n e r a l s h a p e o f the
who le ske le ton as such , however , r e m a i n s r emarkab ly in tac t .
T h e t a p h o n o m i c g rad ien t c o n s t r u e d he r e represents a m i x t u r e o f de
s t ruct ive and cons t ruc t i ve t a p h o n o m i c p rocesses tha t are ac t ive a t different
s tages o f t he g rad ien t (F ig . 6 . 5 ) . D e c a y o c c u r s shor t ly after dea th . Abrasion
i s c o n t i n u o u s and b e c o m e s m o r e ev iden t a l o n g t he gradient . Encrus ta t ion
and b ioe ros ion a lso i n c r e a s e a l o n g t h e g rad ien t to s tage 3b bu t i s eroded
away by s tage 4 . D i a g e n e s i s c a n set in by s tage 3 , e s p e c i a l l y in th is tropical
m a r i n e e n v i r o n m e n t where p r i m a r y m a r i n e c e m e n t a t i o n i s prevalent and
c o n t i n u e s t h r o u g h s tage 4 . C o r r o s i o n , t h e last p rocess to a c t on the skele
tons , d e t e r m i n e s t he a p p e a r a n c e o f t he last s tage o f t h e gradient .
D e c a y i s destruct ive, caus ing the d isar t icu la t ion of spines, disaggrega
t ion of t he jaws, and a l oosen ing of plate boundar i e s . Abras ion is also obvi
ously destructive, as is b ioerosion. However, encrus ta t ion and early diagenesis
c a n serve to s t r eng then the test. For e x a m p l e , enc rus t i ng serpulids and bryo-
zoans cross plate boundar ies . Ea r ly diagenesis also f i l l s in the s te reom. Thus ,
stage 4 skeletons waste away and do no t readily b reak up into f ragments .
A l t h o u g h the re i s a t e m p o r a l s u c c e s s i o n of events , no s t a t emen t s can
b e m a d e a b o u t t h e absolu te t i m i n g . S tud i e s o n t h e ages o f differentially
preserved bivalves in shal low-water se t t ings have shown surpr is ingly larger
age d i sc repanc ie s a m o n g differently preserved s p e c i m e n s (e.g., Kowalewski
e t al . , 1 9 9 8 ) . T h i s shou ld a lso be e x p e c t e d for e c h i n o d e r m r e m a i n s , al
t h o u g h t o m y k n o w l e d g e , n o s u c h s tudies have b e e n m a d e o n ech ino -
de rms . Var ia t ions in t he in tensi ty of t a p h o n o m i c p rocesses such as different
rates o f abras ion , b ioe ros ion , a n d e n c r u s t a t i o n c a n be e x p e c t e d wi th in dif
ferent facies . T h e l e n g t h o f r e s i d e n c e t i m e o n t he s e d i m e n t sur face will
a lso a f fec t t he rate o f t a p h o n o m i c p rocesses a f f ec t ing t h e e c h i n o i d skele
ton. T h e s e d i f fe rences have b e e n s h o w n to o c c u r for Clypeaster f ragments
r ecove red f rom bu lk samples ( N e b e l s i c k , 1 9 9 9 ) s u c h tha t 1 ) low-energy
e n v i r o n m e n t s and low s e d i m e n t a t i o n rates (wh ich e q u a l l o n g sur face resi
d e n c e t imes ) lead to good sur face preserva t ion and h i g h e n c r u s t a t i o n rates;
and 2 ) h igh-ene rgy , shal low-water e n v i r o n m e n t s wi th h i g h e r sed imenta
t ion rates l ead to h igh ly abraded s p e c i m e n s wi th low e n c r u s t a t i o n rates.
However , m a n y m o r e s p e c i m e n s of Clypeaster humilis f rom different facies
would be n e e d e d in order to d i s ce rn s u c h pa t te rns a m o n g c o m p l e t e tests.
T h i s study i s res t r ic ted to c o m p l e t e s p e c i m e n s , a l t h o u g h they are miss
ing the i r spines . O n c e t he s p e c i m e n s are f r a g m e n t e d , di f ferent patterns
e m e r g e (see N e b e l s i c k , 1 9 9 9 ) . F o r e x a m p l e , t he ba re e c h i n o i d test offers a
relat ively large sur face for e n c r u s t i n g o r g a n i s m s in o the rwi se uns tab le en
v i r o n m e n t s ( N e b e l s i c k e t al . , 1 9 9 7 ) . T h e r e f o r e , i t i s no t surpr is ing tha t en-
Nebelsick 124

c rus t a t ion c a n be relat ively h i g h in shal low-water e n v i r o n m e n t s . However ,
f r agmen t s in th is e n v i r o n m e n t are mos t ly h igh ly abraded a n d free o f e n
c rus ta t ion . T h i s m o s t l ikely has to do wi th t he different h y d r o d y n a m i c
proper t ies o f large tests on t he o n e h a n d and f r agmen t s on t he o ther . F rag
m e n t s are m o r e l ikely to be m o r e h igh ly abraded in h ighe r - ene rgy env i ron
m e n t s b e c a u s e o f the i r s m a l l e r s ize and the i r e n t r a i n m e n t i n wave m o v e
m e n t . P r i m a r y enc rus t a t ion on f ragments (as shown by encrus te rs o c c u r r i n g
d i rec t ly on t h e d i sa r t i cu la ted plate bounda r i e s ) a lso o c c u r s bu t i s largely
res t r ic ted to deeper , quie t -water se t t ings (in t he W e s t Area ; see F ig . 1) wi th
low s e d i m e n t a t i o n rates (see N e b e l s i c k , 1 9 9 9 ) . T h i s a l lows for l o n g sur face
r e s i d e n c e t i m e s wi th l i t t le d i s t u r b a n c e tha t wou ld be c o n d u c i v e to t h e set
t l e m e n t o f a n e n c r u s t i n g ep i fauna .
A n o t h e r factor tha t i s obvious ly i m p o r t a n t in t he t a p h o n o m y of t he
ske le tons and p r o d u c t i o n of f r agmen t s i s des t ruc t ive p reda t ion events (see
N e b e l s i c k , 1 9 9 9 ; Kowalewsk i and N e b e l s i c k , 2 0 0 3 ) . F i s h p reda t ion pro
duces a gapp ing w o u n d wi th j agged borders on t he oral sur face . T h e role
o f p reda t ion in t he t a p h o n o m i c s c e n a r i o de sc r i bed above i s t he s u b j e c t o f
c o n t i n u e d inves t iga t ion tha t shou ld add fur ther ins ights in to t he c o m p l e x
ity o f e c h i n o d e r m preserva t ion .
T h e differential stages o f preservat ion a long the t a p h o n o m i c gradient
will affect the t a x o n o m i c resolut ion o f ident if icat ion o f fossil s p e c i m e n s . T h e
level o f t a x o n o m i c ident i f icat ions depends on the loss o f d iagnos t ic t axo
n o m i c charac te rs a long the t a p h o n o m i c gradient . In stages 1 and 2 , spec ies
ident i f icat ion is readily identifiable. Ident i f icat ion at stage 3a and 3b depends
on w h i c h charac te rs h a p p e n to be preserved (i.e., no t destroyed by abrasion
or b ioeros ion) or exposed (i.e., no t covered by encrus ta t ion) . I t also depends
on h o w well the species from the study area are k n o w n with respec t to the i r
m o r p h o l o g i c features and pheno typ ic variat ions. T h e preservat ion o f stage
4 ech ino ids (F ig . 6 .3 .6 ) obviously precludes d i rec t ident i f icat ion at t he spe
cies and even genus level, bu t the order Clypeas te rero ida c a n still be identi
f ied. No o ther e c h i n o d e r m s have such a f la t tened form with in ternal sup
ports, so tha t they c a n be r e c o g n i z e d in even the mos t co r roded s p e c i m e n s .
However , i t will be difficult to m a k e any c loser de t e rmina t ion a t t he fami ly
and suborder levels. E v e n if the test is totally cor roded , the i r des igna t ion as
e c h i n o d e r m s i s possible b e c a u s e o f the u n i q u e s t ruc ture o f t he e c h i n o d e r m
s te reom at a m i c r o s c o p i c level ( S m i t h , 1 9 8 0 , 1 9 8 4 , 1 9 9 0 ) .
T h i s study demons t r a t e s tha t fossil ske le tons have to be r e m o v e d f rom
the t a p h o n o m i c c y c l e in order to be preserved. T h e o r e t i c a l l y , a l l t he s tud
ied s p e c i m e n s wou ld have e n d e d up as s tage 4 ske le tons i f t hey r e m a i n e d
o n t he sur face l ong e n o u g h , a n d even t h e n they c o u l d c o r r o d e c o m p l e t e l y
away. T h i s study a lso rei terates t h e c o m p l e x i t y and in terplay o f dif ferent
factors a f f ec t ing t he preserva t ion o f e c h i n o d e r m skele tons . A l t h o u g h the re
are def ini te e n d m e m b e r s , t h e t a p h o n o m i c p rocess i n b e t w e e n t h e s e e n d
m e m b e r s c a n dif ferent ia l ly af fec t t he preserva t ion o f t he ske le tons . I t has
b e e n a rgued tha t us ing f r agmen t s c a n i n c r e a s e ou r k n o w l e d g e o f t h e dis
t r ibu t ion o f e c h i n o d e r m s ( G o r d o n and D o n o v a n , 1 9 9 2 ; N e b e l s i c k , 1 9 9 2 a ,
1 9 9 2 b ; D o n o v a n , 2 0 0 1 , 2 0 0 3 ; K r o h , 2 0 0 5 ) : t he f r agment s c a n m o r e c lose ly
preserve t h e l ong - t e rm s e t t l e m e n t pa t te rns b e c a u s e l iv ing e c h i n o i d distri-
Taphonomy of the Irregular Echinoid Clypeaster humulis 725

bu t ions are notor ious ly patchy. T h i s s tudy shows tha t b o t h c o m p l e t e skele
tons and f r agment s shou ld a lso be i n c l u d e d in t a p h o n o m i c analysis in or
der to ob t a in a m o r e c o m p l e t e p i c tu r e of t he e c o l o g i c a l factors affecting
preserva t ion a t different sca les o f obse rva t ion .
I t h a n k t he s taf f o f t he Ins t i tu te o f G e o s c i e n c e s Univers i ty o f T u b i n g e n for
the i r support , e spec i a l l y W. G e r b e r for t he pho tographs . I s ince re ly thank
W. I . Aus i ch and C . S c h n e i d e r for the i r ca re fu l reviews.
Allison, P. A. 1990. Variation in rates and decay and disarticulation of Ech ino-
dermata: implications for the application of actualistic data. Palaios,
5 :432-330.
Ausich, W. I. 2001 . Echinoderm taphonomy, p. 171-227. In M. Jangoux and J.
M. Lawrence (eds.), Echinoderm Studies 6. Balkema, Lisse.
Boggild, G. R., and E. P. F. Rose. 1984. Mid-Tertiary echinoid biofacies as pal-
aeoenvironmental indices. Annales Geologique des Pays Hellenique,
22 :57-67 .
Brett, C. E . , H. A. Moffat, and W. L. Taylor. 1997. Echinoderm taphonomy, ta-
phofacies, and Lagerstatten, p. 147-190 . In J. A. Waters and C. G. Maples
(eds.), Geobiology of Echinoderms. Paleontological Society Papers, 3.
Clark, A. M. , and F. W E. Rowe. 1971. Monograph of the Shallow-Water Indo-
West Pacific Echinoderms. British Museum, London, 238 p.
De Meijere, J. C. H. 1902. Vorlaufige Beschreibung der neuen, durch die Si-
boga-Expedition gesammelten Echiniden. Tijdschrift van de Nederlansche
Dierkundige Vereeniging Leiden, 2 ( 8 ) 4 - 1 6 .
Donovan, S. K. 1991. T h e taphonomy of echinoderms: calcareous multi-ele
ment skeletons in the marine environment, p. 2 4 1 - 2 6 9 . In S. K. Donovan
(ed.), T h e Processes of Fossilisation. Belhaven Press, London.
Donovan, S. K. 2001 . Evolution of Car ibbean echinoderms during the C e n o -
zoic: moving towards a complete picture using all of the fossils. Palaeogeog
raphy, Palaeoclimatology, Palaeoecology, 166:177-192 .
Donovan, S. K. 2003 . Completeness of a fossil record: the Pleistocene echinoids
of the Antilles. Lethaia, 36:1-7.
Durham, J. W. 1955. Classification of clypeasteroid echinoids. California Uni
versity Publications in Geological Sc ience , 31 :73-198 .
Durham, J. W. 1966. Clypeasteroids, p. U 4 5 0 - U 4 9 1 . In R. C. Moore (ed.), Trea
tise on Invertebrate Paleontology. Pt. U. Echinodermata 3. Geological Soci
ety of America and University of Kansas Press, Lawrence.
Ghiold, J . , and A. Hoffman. 1984. Clypeasteroid echinoids and historical bioge-
ography. Neues Jahrbuch fiir Geologie und Palaontologie Monatshefte,
1984 :529 -538 .
Ghiold, J . , and A. Hoffman. 1986. Biogeography and biogeographic history of
clypeasteroid echinoids. Journal of Biogeography, 13 :183-206 .
Gordon, C. M. , and S. K. Donovan. 1992. Disarticulated echinoid ossicles in
paleoecology and taphonomy: the last interglacial Falmouth Formation of
Jamaica. Palaios, 7 :157-166.
Greenstein, B. J. 1993. Is the fossil record of regular echinoids really so poor? A
comparison of living and subfossil assemblages. Palaios, 8 : 5 9 7 - 6 0 1 .
Kampfer, S., and W Tertschnig. 1992. Feeding biology of Clypeaster rosaceus
(Echinoidea, Clypeasteroidea) and its impact on shallow lagoon sediments,
126 Nebelsick
Acknowledgments
References

p. 1 9 7 - 2 0 0 . In L. Scalera-Liaci and C. Canicat t i (eds.), Ech inoderm Re
search 1991: Proceedings of the 3rd European Echinoderm Conference,
L e c c e , Italy, 1991. A. A. Balkema, Rotterdam.
Kidwell, S. M. , and T. Baumiller. 1990. Experimental disintegration of regular
echinoids: roles of temperature, oxygen and decay thresholds. Paleobiology,
16 :247-271 .
Kier, P. M. 1977. T h e poor fossil record of the regular echinoids. Paleobiology,
3 :168-174.
Koehler, R. 1922. Echinides du musee indien a Calcutta. 11. Clypeastrides et
Cassidulides. Echinoderma of the Indian Museum, part I X , Echinoidea
(II). Calcut ta , 161 p.
Kowalewski, M. , G. A. Goodfriend, and K. W. Flessa. 1998. T h e high-resolution
estimates of temporal mixing in shell beds: the evils and virtues of t ime-
averaging. Paleobiology, 2 4 : 2 8 7 - 3 0 4 .
Kowalewski, M. , and J. H. Nebelsick. 2003 . Predation on Recent and fossil echi
noids, p. 2 7 9 - 3 0 2 . In P. H. Kelley, M. Kowalewski, and T. A. Hansen (eds.),
Predator-Prey Interactions in the Fossil Record. Topics in Geobiology Se
ries, 20. Kluwer Academic/Plenum Publishers, New York.
Kroh, A. 2005 . Catalogus Fossilium Austriae. Band 2. Echinoidea neogenica.
Osterreichische Akademie der Wissenschaften, Vienna , 210 p.
Kroh, A., and J. H. Nebelsick. 2003 . Echinoid assemblages as a tool for palaeoen-
vironmental reconstruction—an example from the Early M i o c e n e of Egypt.
Palaeogeography, Palaeoclimatology, Palaeoecology, 201:157-177.
Lamarck, J. B. P. A. DE M. 1801. Systeme des animaux sans vertebres, ou table
general des classes, des ordres, et des genres et ces animaux. Paris, chez
Deterville, 432 p.
Leske, N. G. 1778. Additamenta ad Jacobi Theodori Klein Naturalem Disposi-
t ionem Echinodermatum et Lucubrat iunculam de Aculeis Ech inorum
Marinorum. Officina Gleditschiana, Lipsiae, 278 p.
Lewis, R. 1980. Taphonomy, p. 2 7 - 3 9 . In T. W. Broadhead and J. A. Waters
(eds.), Echinoderms: Notes for a Short Course. University of Tennessee
Studies in Geology, 3. University of Tennessee Press, Knoxville.
Linnaeus, C. 1758. Systema naturae (10th edition). Stockholm, Laurentii Salvii.
824 p.
Moffat, H. A., and D. J. Bottjer. 1999. Echinoid concentration beds: two exam
ples from the stratigraphic spectrum. Palaeogeography, Palaeoclimatology,
Palaeoecology, 1 4 9 : 3 2 9 - 3 4 8 .
Mooi , R. 1989. Living and fossil genera of the Clypeasteroida (Echinoidea: Ech i -
nodermata): an illustrated key and annotated checklist. Smithsonian Con
tributions to Zoology, 4 8 8 : 1 - 5 1 .
Nebelsick, J. H. 1992a. T h e Northern Bay of Safaga (Red Sea , Egypt): an actuo-
palaeontological approach. I l l Distribution of echinoids. Beitrage zur
Palaontologie von Osterreich, 17:5-79.
Nebelsick, J. H. 1992b. T h e use of fragments in deducing echinoid distribution
by fragment identification in Northern Bay of Safaga; Red Sea, Egypt.
Palaios, 7 :316-328 .
Nebelsick J. H. 1999. Taphonomic signatures and taphofacies distribution as
recorded by Clypeaster fragments from the Red Sea. Lethaia , 32 :241-252 .
Nebelsick, J. H. 2 0 0 4 . Taphonomy of echinoderms: introduction and outlook, p.
4 7 1 - 4 7 7 . In T. Heinzeller, and J. H. Nebelsick (eds.), Echinoderms
Miinchen: Proceedings of the 11th International Echinoderm Meeting.
Taylor and Francis, London.
Nebelsick, J. H., and S. Kampfer. 1994. Taphonomy of Clypeaster humilis and
Taphonomy of the Irregular Echinoid Clypeaster humulis 7 2 7

Echinodiscus auritus from the Red Sea , p. 8 0 3 - 8 0 8 . In B. David, A. Guil le,
J.-R Feral, and M. Roux (eds.), Echinoderms through T i m e . A. A. Balkema,
Rotterdam.
Nebelsick, J. H., and M. Kowalewski. 1999. Dril l ing predation on recent
Clypeasteroid echinoids from the Red Sea. Palaios, 14 :127-144 .
Nebelsick, J . H., B. Schmid , and M. Stachowitsch. 1997. T h e encrustation of
fossil and recent sea-urchin tests: ecological and taphonomic significance.
Lethaia, 3 0 : 2 7 1 - 2 8 4 .
Neraudeau, D., E. Goubert , D. Lacour, and J . M. Rouchy. 2001 . Changing bio
diversity of Mediterranean irregular echinoids from the Messinian to pres
ent-day. Palaeogeography, Palaeoclimatology, Palaeoecology, 1 7 5 : 4 3 - 6 0 .
Piller, W., and A. M. Mansour. 1990. T h e Northern Bay of Safaga (Red Sea,
Egypt): an actuopalaeontological approach, II. Sediment analysis and sedi
mentary facies. Beitrage zur Palaontologie von Osterreich, 16:1-102.
Piller, W., and P. Pervesler. 1989. T h e Northern Bay of Safaga (Red Sea, Egypt):
an actuopalaeontological approach, II. Topography and bottom facies. B e
itrage zur Palaontologie von Osterreich, 15:103-147.
Poddubiuk, R. H., and E. P. F. Rose. 1984. Relationships between mid-Tertiary
echinoid faunas from the Central Mediterranean and eastern Caribbean
and their palaeobiogeographic significance. Annales Geologique des Pays
Hellenique, 31:115-127.
Seilacher, A. 1979. Constructional morphology of sand dollars. Paleobiology,
5:191-221.
Smith, A. B. 1980. Stereom microstructure of the echinoid test. Special Papers
in Palaeontology, 25 :1 -81 .
Smith, A. B. 1984. Echinoid Palaeobiology. George Allen and Unwin, London,
191 p.
Smith, A. B. 1990. Biomineralization in echinoderms, p. 4 1 3 - 4 4 3 . In J. G.
Carter (ed.), Skeletal Biomineril ization. Patterns, Processes and Evolution
ary Trends, volume I. Van Nostrand Reinhold, New York.
Smith, A. B. 2001 . Probing the cassiduloid origins of clypeasteroid echinoids
using stratigraphically restricted parsimony analysis. Paleobiology,
2 7 : 3 9 2 - 4 0 4 .
Nebelsick 128




















