
The eagle owl ( Bubo bubo ) as a leporid remains accumulator: taphonomic analysis of modern rabbit...
20
The Eagle Owl (Bubo bubo) as a Leporid Remains Accumulator: Taphonomic Analysis of Modern Rabbit Remains Recovered from Nests of this Predator L. LLOVERAS a,b , M. MORENO-GARCI ´ A b AND J. NADAL a a SERP, Departament de Prehisto`ria, Histo`ria Antiga i Arqueologia, Facultat de Geografia i Histo`ria, Universitat de Barcelona, Montalegre 6, 08001 Barcelona, Spain b Laborato´rio de Arqueozoologia, IGESPAR, I. P., Avenida da I ´ ndia 136, 1300-300 Lisbon, Portugal ABSTRACT The eagle owl (Bubo bubo) is the main leporid predator among nocturnal raptors. Although its role as an accumulator of leporid remains in archaeological sites has already been demon- strated, the taphonomic signature of this predator has not been properly characterised. Here we present the analysis of two samples of modern leporid remains recovered from eagle owls’ nests with the aim of contributing new data to this subject. Results show that although variability occurs, the observed anatomical representation, breakage and digestion patterns allow us to distinguish this raptor from other rabbit predators. Finally, several characteristic features are noted to distinguish eagle owls as agents of leporid bone accumulations in the fossil record. Copyright ß 2008 John Wiley & Sons, Ltd. Key words: taphonomic analysis; leporid remains; eagle owl; European rabbit; nocturnal raptors; digestion; breakage Introduction The eagle owl (Bubo bubo), the largest owl in Europe, is widely distributed not only across this continent but also in Asia and North Africa (Mikkola, 1994). It inhabits a large variety of habitats from boreal coniferous and mixed deciduous forests, Mediterranean scrub and steppes, to rocky and sandy deserts (Mikkola, 1994). The eagle owl is considered a generalist predator because it can feed on a wide prey spectrum (Mikkola, 1983); it is an opportunistic hunter that feeds on rabbit, hare, mice, rats, squirrels, birds, and any prey that is not larger than itself. However, in Mediterranean areas the owl depends heavily on leporids, specifically on the European rabbit (Oryctolagus cuniculus) (Ser- rano, 1998). This turns the eagle owl, together with the Spanish Imperial eagle (Aquila adalberti), into the most specialised rabbit predator among large European raptors (Delibes & Hiraldo, 1981; Donazar, 1987). Studies on the eagle owl diet are numerous (Hiraldo et al., 1975; Pe ´rez Mellado, 1980; Mikkola, 1983; Donazar, 1989; Amo & Gonza ´ lez, 1998; Serrano, 1998; Lourenc ¸o, 2006). They show that, although variations could be important depending on season of the year, geographical area or rabbit/hare abundance, leporids always constitute between 23% and International Journal of Osteoarchaeology Int. J. Osteoarchaeol. (2008) Published online in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/oa.995 * Correspondence to: SERP, Departament de Prehisto ` ria, Histo ` ria Antiga i Arqueologia, Facultat de Geografia i Histo ` ria, University of Barcelona, Montalegre 6, 08001 Barcelona, Spain. e-mail: [email protected] Copyright # 2008 John Wiley & Sons, Ltd. Received 30 November 2007 Revised 14 March 2008 Accepted 4 April 2008
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of The eagle owl ( Bubo bubo ) as a leporid remains accumulator: taphonomic analysis of modern rabbit...

International Journal of OsteoarchaeologyInt. J. Osteoarchaeol. (2008)Published online in Wiley InterScience
95
(www.interscience.wiley.com) DOI: 10.1002/oa.9* Correspondence to: SERP,Antiga i Arqueologia, FacultatBarcelona, Montalegre 6, 080e-mail: [email protected]
Copyright # 2008 Joh
The Eagle Owl (Bubo bubo) as aLeporid Remains Accumulator:Taphonomic Analysis of ModernRabbit Remains Recovered fromNests of this Predator
L. LLOVERASa,b�, M. MORENO-GARCIAb AND J. NADALa
a SERP, Departament de Prehistoria, Historia Antiga i Arqueologia, Facultat de Geografia i
Historia, Universitat de Barcelona, Montalegre 6, 08001 Barcelona, Spainb Laboratorio de Arqueozoologia, IGESPAR, I. P., Avenida da India 136, 1300-300 Lisbon,
Portugal
ABSTRACT The eagle owl (Bubo bubo) is the main leporid predator among nocturnal raptors. Although itsrole as an accumulator of leporid remains in archaeological sites has already been demon-strated, the taphonomic signature of this predator has not been properly characterised. Herewe present the analysis of two samples of modern leporid remains recovered from eagle owls’nests with the aim of contributing new data to this subject. Results show that althoughvariability occurs, the observed anatomical representation, breakage and digestion patternsallow us to distinguish this raptor from other rabbit predators. Finally, several characteristicfeatures are noted to distinguish eagle owls as agents of leporid bone accumulations in thefossil record. Copyright � 2008 John Wiley & Sons, Ltd.
Key words: taphonomic analysis; leporid remains; eagle owl; European rabbit; nocturnal
raptors; digestion; breakage
Introduction
The eagle owl (Bubo bubo), the largest owl inEurope, is widely distributed not only across thiscontinent but also in Asia and North Africa(Mikkola, 1994). It inhabits a large variety ofhabitats from boreal coniferous and mixeddeciduous forests, Mediterranean scrub andsteppes, to rocky and sandy deserts (Mikkola,1994). The eagle owl is considered a generalistpredator because it can feed on a wide preyspectrum (Mikkola, 1983); it is an opportunistic
Departament de Prehistoria, Historiade Geografia i Historia, University of01 Barcelona, Spain.
n Wiley & Sons, Ltd.
hunter that feeds on rabbit, hare, mice, rats,squirrels, birds, and any prey that is not largerthan itself. However, in Mediterranean areas theowl depends heavily on leporids, specifically onthe European rabbit (Oryctolagus cuniculus) (Ser-rano, 1998). This turns the eagle owl, togetherwith the Spanish Imperial eagle (Aquila adalberti),into the most specialised rabbit predator amonglarge European raptors (Delibes & Hiraldo, 1981;Donazar, 1987). Studies on the eagle owl diet arenumerous (Hiraldo et al., 1975; Perez Mellado,1980; Mikkola, 1983; Donazar, 1989; Amo &Gonzalez, 1998; Serrano, 1998; Lourenco, 2006).They show that, although variations could beimportant depending on season of the year,geographical area or rabbit/hare abundance,leporids always constitute between 23% and
Received 30 November 2007Revised 14 March 2008Accepted 4 April 2008

L. Lloveras, M. Moreno-Garcıa and J. Nadal
62% of the prey in their diet, being a resourceexploited all year around.This nocturnal bird of prey nests on the
ground, on ledges in cliffs, on fissures on rockfaces or in small caves. Whenever possible, itswallows its food whole. If prey are too large to beingested complete, as is the case with rabbits,carcasses are dismembered to be swallowed insmaller portions. Fur or feathers, bone and meatare also ingested. To discard what its body doesnot require, it then regurgitates pellets thataccumulate on the surface of the nest and thesurrounding area, contributing in this way to thecreation of important bone assemblages ofleporid and other species.In most Palaeolithic and Epipalaeolithic Iber-
ian sites, leporids, and in particular the Europeanrabbit, are the dominant taxon among faunalarchaeological assemblages.Recently, several studies have demonstrated
the importance that small game exploitation(i.e. leporids) would have had for the daily dietof prehistoric people, their relevance beingused to explain mobility patterns and an increaseof populations in the transition from thePleistocene to the Holocene (Villaverde et al.,1996; Stiner et al., 1999, 2000; Hockett & Bicho,2000; Hockett & Haws, 2002). At the sametime, taphonomic studies have alerted us to thefact that human activity would not be the onlyagent responsible for some of these archaeo-logical bone assemblages, but that they may berelated to the consumption habits of competinganimal predators (Hockett, 1989, 1991, 1995,1996; Fernandez-Jalvo & Andrews, 1992; PerezRipoll, 1993, 2004; Schmitt & Juell, 1994;Schmitt, 1995; Cruz-Uribe & Klein, 1998;Mondini, 2002; Lloveras et al., 2008a,b).Following this line of research, modern boneassemblages produced by eagle owl have beenanalysed focusing on the taphonomic charac-teristics visible on micromammals (Dodson &Wexlar, 1979; Hoffman, 1988; Andrews, 1990),birds (Bochenski et al., 1993; Bochenski &Tomek, 1997; Laroulandie, 2000, 2002), andalso leporid remains (Guillem & Martınez-Valle,1991; Hockett, 1995; Sanchis, 2000; Cochard,2004; Yravedra, 2004).Some of these works note that there is not a
straightforward, recognisable pattern in skeletal
Copyright # 2008 John Wiley & Sons, Ltd.
remains accumulations created by the eagle owl,but that variability may occur as a result ofthe origin of the assemblage (Hockett, 1995;Cochard, 2008). Thus, differences in anato-mical representation, fragmentation and diges-tion corrosion patterns are expected between:
(1) a
ssemblages created on the nest, whereremains from pellets, non-ingested skeletalelements and debris are accumulated bychicks and adults; and(2) p
ellets concentrated around roosting plat-forms by regurgitation of adults out of thebreeding season or by non-reproductive indi-viduals.Unfortunately, most of the taphonomic studiescarried out with leporid remains accumulated byeagle owls do not state clearly the origin of theaccumulation, and sometimes remains of mixedorigin are treated together (Sanchis, 2000;Yravedra, 2004). Also, on some occasions thesamples are so small that results cannot beconsidered significant (Yravedra, 2004). Con-sequently, it appears that a characteristic tapho-nomic signature of this nocturnal raptor onleporid bones, which can be successfully recog-nised in archaeological deposits, has not beenproperly registered yet. Thus, our aim is toanalyse in detail the taphonomic patterns left onrabbit remains accumulated at the nests of eagleowls by setting up a series of criteria that can beused in terms of comparison with assemblagesderived from other means or originated by otherpredators.
Material and methods
The leporid remains analysed come from twonests of eagle owl located in the Natural ParkMassıs del Garraf in Barcelona (northeast Spain;see Figure 1). Both samples were collected by staffof the park in July 2006, at the end of thebreeding season. The nests were situated in aprotected shelter, and remains accumulated didnot display any sign of weathering or having beendisturbed by scavengers. Samples consist ofdegraded pellets, unswallowed bones and debrissuch as feathers, eggshells and pebbles accumu-lated on the surface of the nests (Figure 2). Due to
Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa

Figure 1. Map of the Iberian Peninsula showing the location of Natural Park Massıs del Garraf (Barcelona, Spain).
Taphonomy of Rabbit Remains from Eagle Owl Nests
the relatively high percentage of bones (over 65%in both nests) that showed evidence of digestion(Table 7) and the low percentage of articulatedelements (see below), it can be assumed that alarge number of the rabbit remains analysedderived from degraded pellets and not fromuneaten leftovers.Material was carefully dry-sieved through a
1.5mm mesh in order to recover all prey remains.Leporid bones and teeth were selected and storedapart (Figure 3).
Figure 2. Material accumulated on the surfaces of Nest 1occurrence of bone remains from decomposed pellets andvegetation.
Copyright # 2008 John Wiley & Sons, Ltd.
The methodological analysis followed wasdesigned by us and has already been applied tothe study of modern rabbit remains accumu-lations created by the Spanish Imperial eagle andthe Iberian lynx (Lloveras et al., 2008a,b).
Anatomical representation
Numbers of identified skeletal elements (N)were counted and relative abundance was also
and Nest 2 as it arrived to us to be analysed. Note theuneaten food, feathers and debris such as pebbles and
Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa

Figure 3. Rabbit remains recovered from Nest 2 after dry-sieving the accumulated material.
L. Lloveras, M. Moreno-Garcıa and J. Nadal
calculated (Dodson & Wexlar, 1979; Andrews,1990) using the formula:
RAi ¼ MNEi=MNI� Ei
where RAi is the relative abundance of element i,MNEi is the minimum number of skeletonelement i, MNI is the minimum number ofindividuals based on the highest number of anysingle element in the assemblage, and Ei is thenumber of element i in the prey skeleton.In addition, proportions of skeletal elements
were evaluated using three indices for postcraniain relation to crania (Andrews, 1990):
(a) P
Co
CRT/CR: total numbers of postcranial (limbelements, vertebrae and ribs) compared withtotal numbers of cranial elements (mandibles,maxillae and teeth);
(b) P
CRAP/CR: limb elements (long bones, sca-pulae, innominate, patellae, metapodials, car-pyright # 2008 John Wiley & Sons, Ltd.
pals, tarsals and phalanges) compared withcranial elements (mandibles, maxillae andteeth);
(c) P
CRLB/CR: postcranial long bones (humerus,radius, ulna, femur and tibia) compared withcranial elements (mandibles and maxillae).Loss of distal limb elements was shown by twoindices (Lloveras et al., 2008a):
(a) A
UT/ZE: autopodium elements (metapodials,carpals, tarsals and phalanges) compared withzigopodium and stylopodium (tibia, radius,ulna, humerus, femur and patellae);(b) Z
/E: zigopodium elements (tibia, radius andulna) compared with stylopodium (femur andhumerus).A further index compared anterior to posteriorlimb elements – AN/PO: humerus, radius, ulnaand metacarpals compared with femur, tibia andmetatarsals.
Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa

Taphonomy of Rabbit Remains from Eagle Owl Nests
Breakage
The breakage pattern was described by themaximum length of all the identified skeletalelements. Additionally, Student’s t-test wasapplied to the maximum length of the determinedremains to assess any differences in the size offragments between the two nests. Percentages ofcomplete elements, isolated teeth and articulatedelements were also calculated. Different breakagecategories were used depending on bone type:
� P
C
atellae, carpals, tarsals and ribs were classifiedas complete (C) or fragmented (F).
� P
halanges were recorded as complete (C), prox-imal (P) or distal (D) fragments. When dis-tinguishing between proximal or distal was notpossible, they were recorded just as fragment (F).� V
ertebrae were registered as complete (C),vertebral body (VB), vertebral epiphysis (VE)or spinous process (SP).� B
reakage of teeth was calculated separately forisolated and in situ elements (Fernandez-Jalvo &Andrews, 1992), and they were classified ascomplete (C) or fragmented (F).� B
reakage categories used for long bones, meta-podials, mandibles, cranium, scapula and inno-minate are presented in Table 3. They were fullydescribed and illustrated in Lloveras et al. (2008a).Digestion
Damage to the bone surfaces was observed undera light microscope (�10 to �40). Based on themethodology used for micromammals byAndrews (1990) and Fernandez-Jalvo & Andrews(1992), different categories of digestion damagewere applied to leporid bones and teeth (Lloveraset al., 2008a,b). Five degrees of digestion weredistinguished: null (0), light (1), moderate (2),heavy (3) and extreme (4). These were evaluatedseparately for bones and dental remains.
Puncture marks
Finally, damage to the bone surfaces caused bythe predator beak or talons was also noted andcounted.
opyright # 2008 John Wiley & Sons, Ltd.
Results
Anatomical representationThe number of leporid bones and teeth, identifiedto skeletal element, was 1808 for Nest 1 and 1932for Nest 2. All leporid remains identified wererabbits (Figure 3), and taking into account thefusion stage of long bone and metapodialepiphyses, scapula, calcaneum and innominate,the percentage of adult individuals was 50% inboth nests. Based on calcaneum counts, theminimum number of individuals (MNI) estimatedwas 19 for Nest 1 and 26 for Nest 2.Table 1 shows the anatomical composition of
the identified remains for each nest. The entireskeleton was represented – vertebrae, ribs,phalanges, metatarsals and tibia/femur were themost frequent elements in both samples (N%).The main difference concerns cranial remainsthat were more abundant in Nest 2 than in Nest 1.Relative abundances of skeletal elements (RA%)are also shown in Table 1 and Figure 4. The meanvalues (45.2% for Nest 1 and 36.1% for Nest 2)indicate a significant loss of prey bones in bothnest assemblages.The best represented elements were those of the
hindlimb (Nest 1: patella, calcaneum, innominateand femur; Nest 2: calcaneum, innominate andtibia). Metatarsal bones and astragalus also showedhigh percentages in both samples, and long boneswere well represented only for tibia and femur(Figure 4). Relative proportions of skeletalelements are shown in Table 2. Results show that:
� I
ndices that compare postcranial to cranialelements (PCRT/CR; PCRAP/CR) point to adeficiency in the numbers of crania for Nest 1(171.3 and 183.6, that means 1.7 and 1.8 timesmore postcranial than cranial remains), whereasfor Nest 2 postcranial to cranial proportionswere similar (98.2 and 94.7).� W
hen cranial elements are compared with longbones (PCRLB/CR), both samples show adeficiency of crania (233.2 and 127.4).� C
omparisons of lower to upper limb elements(AUT/ZE and Z/E) show loss of the former in agreater proportion for both nests.� T
he AN/PO index reveals much better survivalof posterior limb elements than anterior limbbones in both nests.Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa
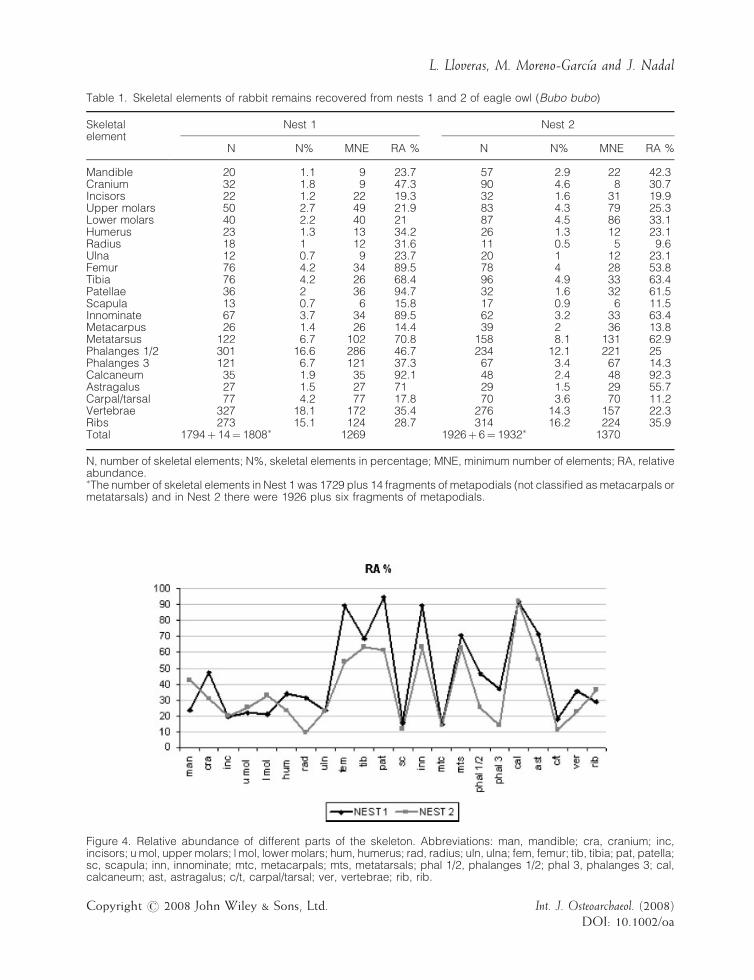
Table 1. Skeletal elements of rabbit remains recovered from nests 1 and 2 of eagle owl (Bubo bubo)
Skeletalelement
Nest 1 Nest 2
N N% MNE RA % N N% MNE RA %
Mandible 20 1.1 9 23.7 57 2.9 22 42.3Cranium 32 1.8 9 47.3 90 4.6 8 30.7Incisors 22 1.2 22 19.3 32 1.6 31 19.9Upper molars 50 2.7 49 21.9 83 4.3 79 25.3Lower molars 40 2.2 40 21 87 4.5 86 33.1Humerus 23 1.3 13 34.2 26 1.3 12 23.1Radius 18 1 12 31.6 11 0.5 5 9.6Ulna 12 0.7 9 23.7 20 1 12 23.1Femur 76 4.2 34 89.5 78 4 28 53.8Tibia 76 4.2 26 68.4 96 4.9 33 63.4Patellae 36 2 36 94.7 32 1.6 32 61.5Scapula 13 0.7 6 15.8 17 0.9 6 11.5Innominate 67 3.7 34 89.5 62 3.2 33 63.4Metacarpus 26 1.4 26 14.4 39 2 36 13.8Metatarsus 122 6.7 102 70.8 158 8.1 131 62.9Phalanges 1/2 301 16.6 286 46.7 234 12.1 221 25Phalanges 3 121 6.7 121 37.3 67 3.4 67 14.3Calcaneum 35 1.9 35 92.1 48 2.4 48 92.3Astragalus 27 1.5 27 71 29 1.5 29 55.7Carpal/tarsal 77 4.2 77 17.8 70 3.6 70 11.2Vertebrae 327 18.1 172 35.4 276 14.3 157 22.3Ribs 273 15.1 124 28.7 314 16.2 224 35.9Total 1794þ 14¼ 1808� 1269 1926þ 6¼ 1932� 1370
N, number of skeletal elements; N%, skeletal elements in percentage; MNE, minimum number of elements; RA, relativeabundance.�The number of skeletal elements in Nest 1 was 1729 plus 14 fragments of metapodials (not classified as metacarpals ormetatarsals) and in Nest 2 there were 1926 plus six fragments of metapodials.
Figure 4. Relative abundance of different parts of the skeleton. Abbreviations: man, mandible; cra, cranium; inc,incisors; u mol, upper molars; l mol, lower molars; hum, humerus; rad, radius; uln, ulna; fem, femur; tib, tibia; pat, patella;sc, scapula; inn, innominate; mtc, metacarpals; mts, metatarsals; phal 1/2, phalanges 1/2; phal 3, phalanges 3; cal,calcaneum; ast, astragalus; c/t, carpal/tarsal; ver, vertebrae; rib, rib.
Copyright # 2008 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa
L. Lloveras, M. Moreno-Garcıa and J. Nadal
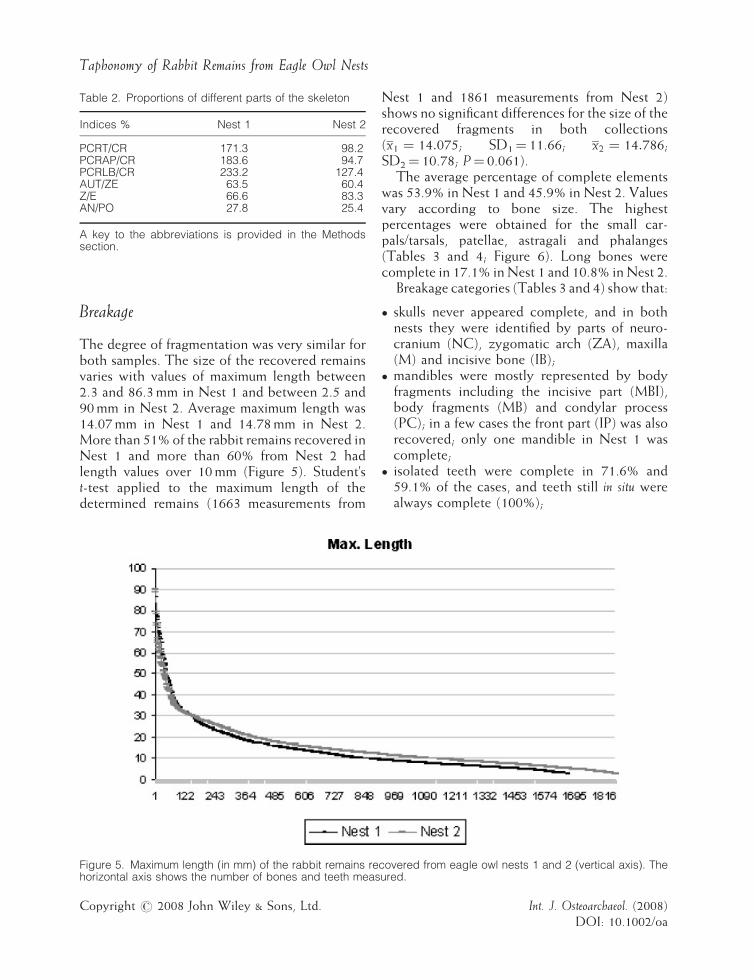
Table 2. Proportions of different parts of the skeleton
Indices % Nest 1 Nest 2
PCRT/CR 171.3 98.2PCRAP/CR 183.6 94.7PCRLB/CR 233.2 127.4AUT/ZE 63.5 60.4Z/E 66.6 83.3AN/PO 27.8 25.4
A key to the abbreviations is provided in the Methods
Taphonomy of Rabbit Remains from Eagle Owl Nests
section.
Breakage
The degree of fragmentation was very similar forboth samples. The size of the recovered remainsvaries with values of maximum length between2.3 and 86.3mm in Nest 1 and between 2.5 and90mm in Nest 2. Average maximum length was14.07mm in Nest 1 and 14.78mm in Nest 2.More than 51% of the rabbit remains recovered inNest 1 and more than 60% from Nest 2 hadlength values over 10mm (Figure 5). Student’st-test applied to the maximum length of thedetermined remains (1663 measurements from
Figure 5. Maximum length (in mm) of the rabbit remains rehorizontal axis shows the number of bones and teeth meas
Copyright # 2008 John Wiley & Sons, Ltd.
Nest 1 and 1861 measurements from Nest 2)shows no significant differences for the size of therecovered fragments in both collections(x1 ¼ 14:075; SD1¼ 11.66; x2 ¼ 14:786;SD2¼ 10.78; P¼ 0.061).The average percentage of complete elements
was 53.9% in Nest 1 and 45.9% in Nest 2. Valuesvary according to bone size. The highestpercentages were obtained for the small car-pals/tarsals, patellae, astragali and phalanges(Tables 3 and 4; Figure 6). Long bones werecomplete in 17.1% in Nest 1 and 10.8% in Nest 2.Breakage categories (Tables 3 and 4) show that:
� s
cour
kulls never appeared complete, and in bothnests they were identified by parts of neuro-cranium (NC), zygomatic arch (ZA), maxilla(M) and incisive bone (IB);
� m
andibles were mostly represented by bodyfragments including the incisive part (MBI),body fragments (MB) and condylar process(PC); in a few cases the front part (IP) was alsorecovered; only one mandible in Nest 1 wascomplete;� is
olated teeth were complete in 71.6% and59.1% of the cases, and teeth still in situ werealways complete (100%);vered from eagle owl nests 1 and 2 (vertical axis). Theed.
Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa
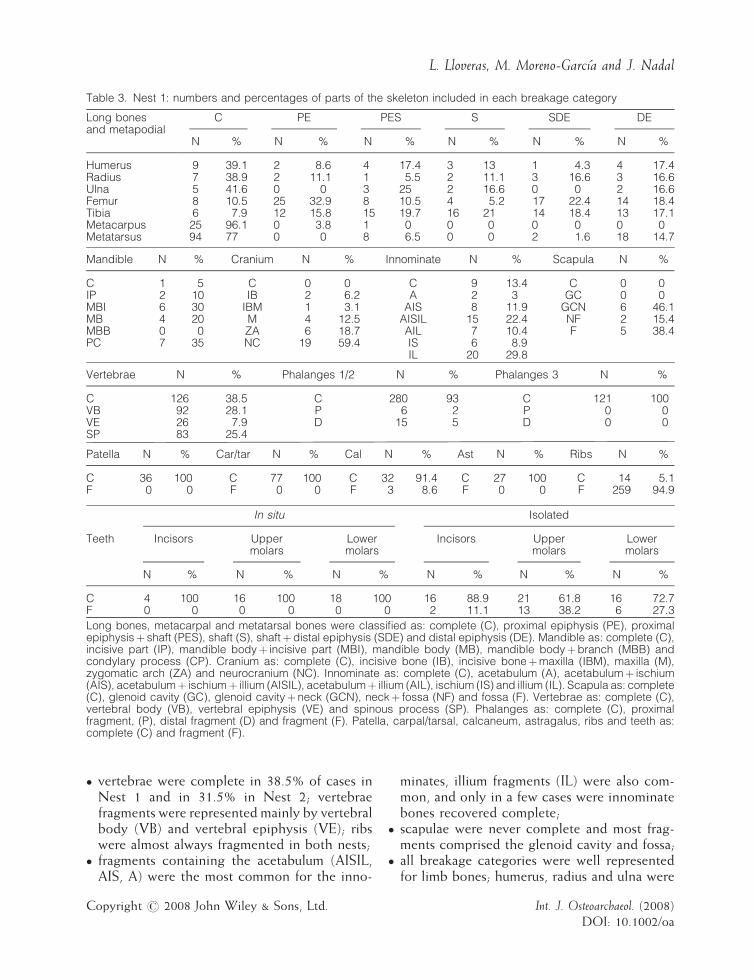
Table 3. Nest 1: numbers and percentages of parts of the skeleton included in each breakage category
Long bonesand metapodial
C PE PES S SDE DE
N % N % N % N % N % N %
Humerus 9 39.1 2 8.6 4 17.4 3 13 1 4.3 4 17.4Radius 7 38.9 2 11.1 1 5.5 2 11.1 3 16.6 3 16.6Ulna 5 41.6 0 0 3 25 2 16.6 0 0 2 16.6Femur 8 10.5 25 32.9 8 10.5 4 5.2 17 22.4 14 18.4Tibia 6 7.9 12 15.8 15 19.7 16 21 14 18.4 13 17.1Metacarpus 25 96.1 0 3.8 1 0 0 0 0 0 0 0Metatarsus 94 77 0 0 8 6.5 0 0 2 1.6 18 14.7
Mandible N % Cranium N % Innominate N % Scapula N %
C 1 5 C 0 0 C 9 13.4 C 0 0IP 2 10 IB 2 6.2 A 2 3 GC 0 0MBI 6 30 IBM 1 3.1 AIS 8 11.9 GCN 6 46.1MB 4 20 M 4 12.5 AISIL 15 22.4 NF 2 15.4MBB 0 0 ZA 6 18.7 AIL 7 10.4 F 5 38.4PC 7 35 NC 19 59.4 IS 6 8.9
IL 20 29.8
Vertebrae N % Phalanges 1/2 N % Phalanges 3 N %
C 126 38.5 C 280 93 C 121 100VB 92 28.1 P 6 2 P 0 0VE 26 7.9 D 15 5 D 0 0SP 83 25.4
Patella N % Car/tar N % Cal N % Ast N % Ribs N %
C 36 100 C 77 100 C 32 91.4 C 27 100 C 14 5.1F 0 0 F 0 0 F 3 8.6 F 0 0 F 259 94.9
In situ Isolated
Incisors Uppermolars
Lowermolars
Incisors Uppermolars
Lowermolars
Teeth
N % N % N % N % N % N %
C 4 100 16 100 18 100 16 88.9 21 61.8 16 72.7F 0 0 0 0 0 0 2 11.1 13 38.2 6 27.3
Long bones, metacarpal and metatarsal bones were classified as: complete (C), proximal epiphysis (PE), proximalepiphysisþ shaft (PES), shaft (S), shaftþdistal epiphysis (SDE) and distal epiphysis (DE). Mandible as: complete (C),incisive part (IP), mandible bodyþ incisive part (MBI), mandible body (MB), mandible bodyþbranch (MBB) andcondylary process (CP). Cranium as: complete (C), incisive bone (IB), incisive boneþmaxilla (IBM), maxilla (M),zygomatic arch (ZA) and neurocranium (NC). Innominate as: complete (C), acetabulum (A), acetabulumþ ischium(AIS), acetabulumþ ischiumþ illium (AISIL), acetabulumþ illium (AIL), ischium (IS) and illium (IL). Scapula as: complete(C), glenoid cavity (GC), glenoid cavityþ neck (GCN), neckþ fossa (NF) and fossa (F). Vertebrae as: complete (C),vertebral body (VB), vertebral epiphysis (VE) and spinous process (SP). Phalanges as: complete (C), proximalfragment, (P), distal fragment (D) and fragment (F). Patella, carpal/tarsal, calcaneum, astragalus, ribs and teeth as:complete (C) and fragment (F).
L. Lloveras, M. Moreno-Garcıa and J. Nadal
� v
C
ertebrae were complete in 38.5% of cases inNest 1 and in 31.5% in Nest 2; vertebraefragments were represented mainly by vertebralbody (VB) and vertebral epiphysis (VE); ribswere almost always fragmented in both nests;
� fr
agments containing the acetabulum (AISIL,AIS, A) were the most common for the inno-opyright # 2008 John Wiley & Sons, Ltd.
minates, illium fragments (IL) were also com-mon, and only in a few cases were innominatebones recovered complete;
� s
capulae were never complete and most frag-ments comprised the glenoid cavity and fossa;� a
ll breakage categories were well representedfor limb bones; humerus, radius and ulna wereInt. J. Osteoarchaeol. (2008)DOI: 10.1002/oa
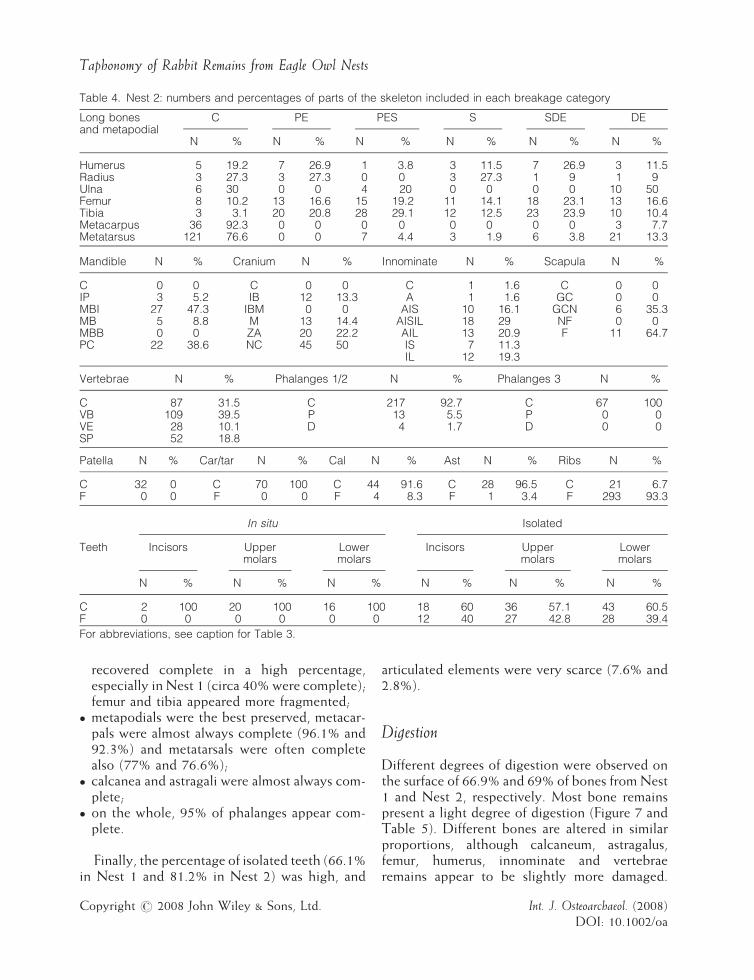
Table 4. Nest 2: numbers and percentages of parts of the skeleton included in each breakage category
Long bonesand metapodial
C PE PES S SDE DE
N % N % N % N % N % N %
Humerus 5 19.2 7 26.9 1 3.8 3 11.5 7 26.9 3 11.5Radius 3 27.3 3 27.3 0 0 3 27.3 1 9 1 9Ulna 6 30 0 0 4 20 0 0 0 0 10 50Femur 8 10.2 13 16.6 15 19.2 11 14.1 18 23.1 13 16.6Tibia 3 3.1 20 20.8 28 29.1 12 12.5 23 23.9 10 10.4Metacarpus 36 92.3 0 0 0 0 0 0 0 0 3 7.7Metatarsus 121 76.6 0 0 7 4.4 3 1.9 6 3.8 21 13.3
Mandible N % Cranium N % Innominate N % Scapula N %
C 0 0 C 0 0 C 1 1.6 C 0 0IP 3 5.2 IB 12 13.3 A 1 1.6 GC 0 0MBI 27 47.3 IBM 0 0 AIS 10 16.1 GCN 6 35.3MB 5 8.8 M 13 14.4 AISIL 18 29 NF 0 0MBB 0 0 ZA 20 22.2 AIL 13 20.9 F 11 64.7PC 22 38.6 NC 45 50 IS 7 11.3
IL 12 19.3
Vertebrae N % Phalanges 1/2 N % Phalanges 3 N %
C 87 31.5 C 217 92.7 C 67 100VB 109 39.5 P 13 5.5 P 0 0VE 28 10.1 D 4 1.7 D 0 0SP 52 18.8
Patella N % Car/tar N % Cal N % Ast N % Ribs N %
C 32 0 C 70 100 C 44 91.6 C 28 96.5 C 21 6.7F 0 0 F 0 0 F 4 8.3 F 1 3.4 F 293 93.3
In situ Isolated
Incisors Uppermolars
Lowermolars
Incisors Uppermolars
Lowermolars
Teeth
N % N % N % N % N % N %
C 2 100 20 100 16 100 18 60 36 57.1 43 60.5F 0 0 0 0 0 0 12 40 27 42.8 28 39.4
For abbreviations, see caption for Table 3.
C
Taphonomy of Rabbit Remains from Eagle Owl Nests
recovered complete in a high percentage,especially in Nest 1 (circa 40% were complete);femur and tibia appeared more fragmented;
� m
etapodials were the best preserved, metacar-pals were almost always complete (96.1% and92.3%) and metatarsals were often completealso (77% and 76.6%);� c
alcanea and astragali were almost always com-plete;� o
n the whole, 95% of phalanges appear com-plete.Finally, the percentage of isolated teeth (66.1%in Nest 1 and 81.2% in Nest 2) was high, and
opyright # 2008 John Wiley & Sons, Ltd.
articulated elements were very scarce (7.6% and2.8%).
Digestion
Different degrees of digestion were observed onthe surface of 66.9% and 69% of bones from Nest1 and Nest 2, respectively. Most bone remainspresent a light degree of digestion (Figure 7 andTable 5). Different bones are altered in similarproportions, although calcaneum, astragalus,femur, humerus, innominate and vertebraeremains appear to be slightly more damaged.
Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa

Figure 6. Percentage of complete rabbit remains from nests 1 and 2. For abbreviations, see Figure 4 caption.
L. Lloveras, M. Moreno-Garcıa and J. Nadal
Phalanges, metapodials, crania, patellae andcarpals/tarsals were the best preserved. Normally,the surface of bones was partially affected bydigestion corrosion, the articulation ends beingthe most altered. Teeth show a bias towards lesserdegrees of digestion corrosion (Figure 8, Table 6),with 42.7% and 64.6% of dental remains in Nests1 and 2 not altered by the effects of digestion.Teeth from Nest 1 present more digestion
Copyright # 2008 John Wiley & Sons, Ltd.
corrosion than those from Nest 2. No significantdifferences between incisors, upper and lowermolars were observed.
Puncture marks
Other traces caused by the beak of eagle owl onbone surfaces were observed on 35 bones (1.93%)
Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa
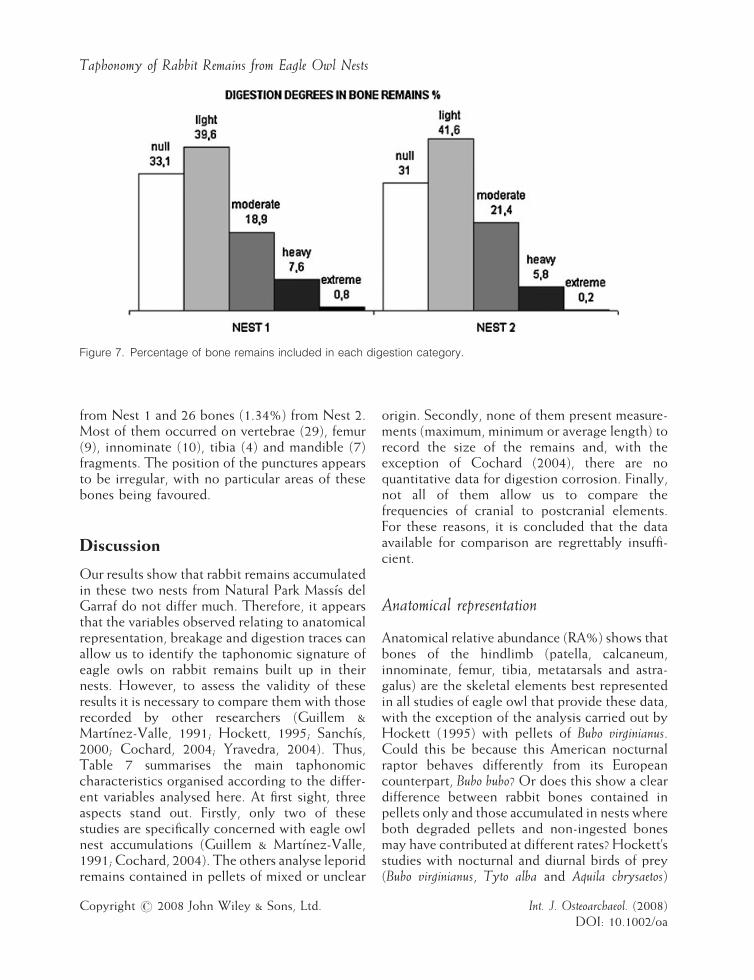
Figure 7. Percentage of bone remains included in each digestion category.
Taphonomy of Rabbit Remains from Eagle Owl Nests
from Nest 1 and 26 bones (1.34%) from Nest 2.Most of them occurred on vertebrae (29), femur(9), innominate (10), tibia (4) and mandible (7)fragments. The position of the punctures appearsto be irregular, with no particular areas of thesebones being favoured.
Discussion
Our results show that rabbit remains accumulatedin these two nests from Natural Park Massıs delGarraf do not differ much. Therefore, it appearsthat the variables observed relating to anatomicalrepresentation, breakage and digestion traces canallow us to identify the taphonomic signature ofeagle owls on rabbit remains built up in theirnests. However, to assess the validity of theseresults it is necessary to compare them with thoserecorded by other researchers (Guillem &Martınez-Valle, 1991; Hockett, 1995; Sanchıs,2000; Cochard, 2004; Yravedra, 2004). Thus,Table 7 summarises the main taphonomiccharacteristics organised according to the differ-ent variables analysed here. At first sight, threeaspects stand out. Firstly, only two of thesestudies are specifically concerned with eagle owlnest accumulations (Guillem & Martınez-Valle,1991; Cochard, 2004). The others analyse leporidremains contained in pellets of mixed or unclear
Copyright # 2008 John Wiley & Sons, Ltd.
origin. Secondly, none of them present measure-ments (maximum, minimum or average length) torecord the size of the remains and, with theexception of Cochard (2004), there are noquantitative data for digestion corrosion. Finally,not all of them allow us to compare thefrequencies of cranial to postcranial elements.For these reasons, it is concluded that the dataavailable for comparison are regrettably insuffi-cient.
Anatomical representation
Anatomical relative abundance (RA%) shows thatbones of the hindlimb (patella, calcaneum,innominate, femur, tibia, metatarsals and astra-galus) are the skeletal elements best representedin all studies of eagle owl that provide these data,with the exception of the analysis carried out byHockett (1995) with pellets of Bubo virginianus.Could this be because this American nocturnalraptor behaves differently from its Europeancounterpart, Bubo bubo? Or does this show a cleardifference between rabbit bones contained inpellets only and those accumulated in nests whereboth degraded pellets and non-ingested bonesmay have contributed at different rates? Hockett’sstudies with nocturnal and diurnal birds of prey(Bubo virginianus, Tyto alba and Aquila chrysaetos)
Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa
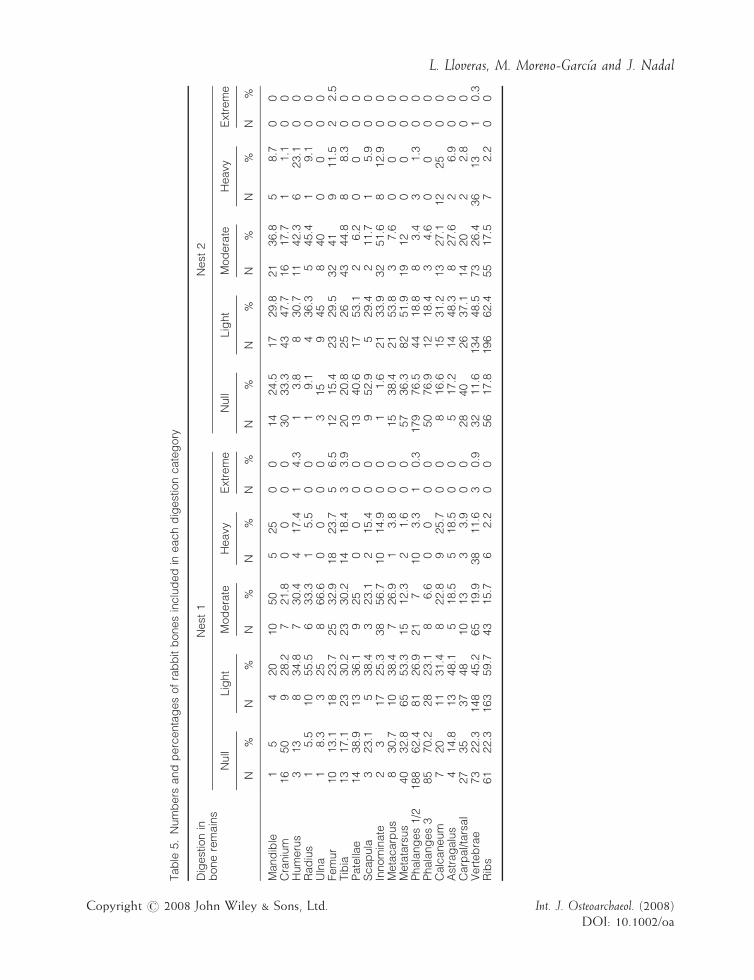
Tab
le5.
Num
bers
and
perc
enta
ges
of
rab
bit
bones
inclu
ded
ineach
dig
estion
cate
gory
Dig
estion
inb
one
rem
ain
sN
est
1N
est
2
Null
Lig
ht
Mod
era
teH
eavy
Extr
em
eN
ull
Lig
ht
Mod
era
teH
eavy
Extr
em
e
N%
N%
N%
N%
N%
N%
N%
N%
N%
N%
Mand
ible
15
420
10
50
525
00
14
24.5
17
29.8
21
36.8
58.7
00
Cra
niu
m16
50
928.2
721.8
00
00
30
33.3
43
47.7
16
17.7
11.1
00
Hum
eru
s3
13
834.8
730.4
417.4
14.3
13.8
830.7
11
42.3
623.1
00
Rad
ius
15.5
10
55.5
633.3
15.5
00
19.1
436.3
545.4
19.1
00
Uln
a1
8.3
325
866.6
00
00
315
945
840
00
00
Fem
ur
10
13.1
18
23.7
25
32.9
18
23.7
56.5
12
15.4
23
29.5
32
41
911.5
22.5
Tib
ia13
17.1
23
30.2
23
30.2
14
18.4
33.9
20
20.8
25
26
43
44.8
88.3
00
Pate
llae
14
38.9
13
36.1
925
00
00
13
40.6
17
53.1
26.2
00
00
Scap
ula
323.1
538.4
323.1
215.4
00
952.9
529.4
211.7
15.9
00
Innom
inate
23
17
25.3
38
56.7
10
14.9
00
11.6
21
33.9
32
51.6
812.9
00
Meta
carp
us
830.7
10
38.4
726.9
13.8
00
15
38.4
21
53.8
37.6
00
00
Meta
tars
us
40
32.8
65
53.3
15
12.3
21.6
00
57
36.3
82
51.9
19
12
00
00
Phala
ng
es
1/2
188
62.4
81
26.9
21
710
3.3
10.3
179
76.5
44
18.8
83.4
31.3
00
Phala
ng
es
385
70.2
28
23.1
86.6
00
00
50
76.9
12
18.4
34.6
00
00
Calc
aneum
720
11
31.4
822.8
925.7
00
816.6
15
31.2
13
27.1
12
25
00
Astr
ag
alu
s4
14.8
13
48.1
518.5
518.5
00
517.2
14
48.3
827.6
26.9
00
Carp
al/ta
rsal
27
35
37
48
10
13
33.9
00
28
40
26
37.1
14
20
22.8
00
Vert
eb
rae
73
22.3
148
45.2
65
19.9
38
11.6
30.9
32
11.6
134
48.5
73
26.4
36
13
10.3
Rib
s61
22.3
163
59.7
43
15.7
62.2
00
56
17.8
196
62.4
55
17.5
72.2
00
Copyright # 2008 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa
L. Lloveras, M. Moreno-Garcıa and J. Nadal
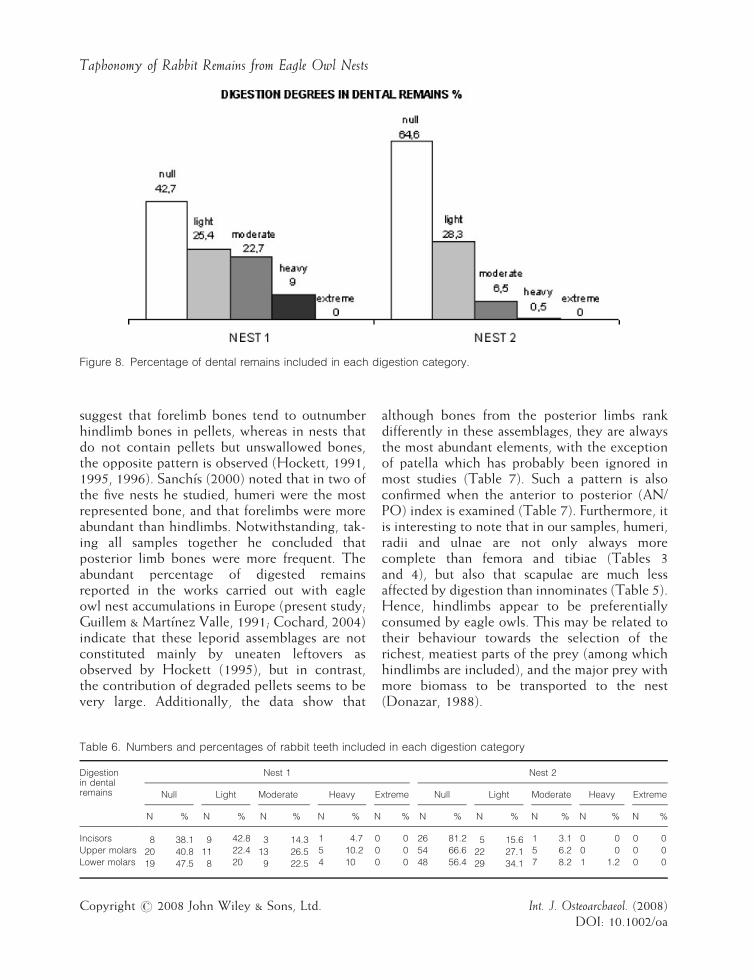
Figure 8. Percentage of dental remains included in each digestion category.
Taphonomy of Rabbit Remains from Eagle Owl Nests
suggest that forelimb bones tend to outnumberhindlimb bones in pellets, whereas in nests thatdo not contain pellets but unswallowed bones,the opposite pattern is observed (Hockett, 1991,1995, 1996). Sanchıs (2000) noted that in two ofthe five nests he studied, humeri were the mostrepresented bone, and that forelimbs were moreabundant than hindlimbs. Notwithstanding, tak-ing all samples together he concluded thatposterior limb bones were more frequent. Theabundant percentage of digested remainsreported in the works carried out with eagleowl nest accumulations in Europe (present study;Guillem & Martınez Valle, 1991; Cochard, 2004)indicate that these leporid assemblages are notconstituted mainly by uneaten leftovers asobserved by Hockett (1995), but in contrast,the contribution of degraded pellets seems to bevery large. Additionally, the data show that
Table 6. Numbers and percentages of rabbit teeth include
Digestionin dentalremains
Nest 1
Null Light Moderate Heavy E
N % N % N % N % N
Incisors 8 38.1 9 42.8 3 14.3 1 4.7 0
Upper molars 20 40.8 11 22.4 13 26.5 5 10.2 0
Lower molars 19 47.5 8 20 9 22.5 4 10 0
Copyright # 2008 John Wiley & Sons, Ltd.
although bones from the posterior limbs rankdifferently in these assemblages, they are alwaysthe most abundant elements, with the exceptionof patella which has probably been ignored inmost studies (Table 7). Such a pattern is alsoconfirmed when the anterior to posterior (AN/PO) index is examined (Table 7). Furthermore, itis interesting to note that in our samples, humeri,radii and ulnae are not only always morecomplete than femora and tibiae (Tables 3and 4), but also that scapulae are much lessaffected by digestion than innominates (Table 5).Hence, hindlimbs appear to be preferentiallyconsumed by eagle owls. This may be related totheir behaviour towards the selection of therichest, meatiest parts of the prey (among whichhindlimbs are included), and the major prey withmore biomass to be transported to the nest(Donazar, 1988).
d in each digestion category
Nest 2
xtreme Null Light Moderate Heavy Extreme
% N % N % N % N % N %
0 26 81.2 5 15.6 1 3.1 0 0 0 0
0 54 66.6 22 27.1 5 6.2 0 0 0 0
0 48 56.4 29 34.1 7 8.2 1 1.2 0 0
Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa

L. Lloveras, M. Moreno-Garcıa and J. Nadal
In all cases, independently of the origin ofthe remains, metacarpals and carpals/tarsals arealways the worst-represented elements. Thereseems to be an important loss of distal(specifically phalanges and carpals/tarsals) versusproximal limb elements (P/D).Finally, relative abundance values of postcra-
nial elements are greater than cranial values in allthe assemblages that provide this information – atrend also observed in the index that comparesproportions between long bones and cranialelements (PCRLB/CR). In the case of nestassemblages this is an expected result, since theeagle owl usually cuts and eats the head of theprey before bringing it to the nest (Donazar,1988). In spite of this fact, our Nest 2 registeredequal proportions between cranial and postcra-nial elements when the latter were quantified alltogether (PCRT/CR¼ 98.2; Table 2). This wasprobably due to a bias caused by greater loss ofdistal elements than in Nest 1, although othervariables not considered here may have influ-enced such a result and should be taken intoaccount in future studies. For example, would thenumber of chicks bred in a particular nest beworth controlling? Is it possible that more chicksmean less cranial remains, as prey would bethoroughly exploited? Also, would prey abun-dance affect consumption patterns?
Breakage pattern
The size of the leporid remains analysed here didnot present significant differences, but compari-sons with other assemblages accumulated by Bubobubo are not possible because this variable was notrecorded in any other published works. In fact,the mean value of maximum length (around15mm) found in our samples may be acharacteristic of eagle owl nest accumulations.As shown in Table 8, this value is clearly higherthan that obtained for other predators like diurnalraptors (Spanish Iberian eagle, 8.36mm: Lloveraset al., 2008a) and terrestrial carnivores (Iberianlynx, 7.1mm; Lloveras et al., 2008b). The same istrue for the percentage of bones smaller than10mm. Whereas they represent 40–49% in oureagle owl nests, they tend to comprise more than70% in bone assemblages originated from
Copyright # 2008 John Wiley & Sons, Ltd.
Spanish Imperial eagle pellets and from Iberianlynx and coyote scats (Table 8). These resultssuggest that eagle owls damage their prey bonesless than other predators.In relation to the frequency of complete
elements, two situations were assessed:
(a) t
he percentage of complete long bones; and (b) t he percentage of complete elements in thetotal sample.
No discrepancy occurs between Nest 1 andNest 2 in the present study, but our results appearto differ greatly from others, as far as the meanvalue of whole long bones is concerned (Table 7).For instance, contrary to complete long bonefrequencies of 14.6% and 10.8% (nests 1 and 2,respectively), Cochard (2004) recorded a value of58%, Sanchis (2000) noted a value of 48%, andYravedra (2004) found nearly 79% (Table 7).Guillem & Martınez-Valle (1991) were the onlyones to register a value close to our results (10%),although they only counted femur and tibia.Thus, one plausible explanation for the differ-ences encountered may be that under thecategory of complete long bones, each analystincludes different skeletal elements or poolstogether complete and ‘almost complete’ longbones (Cochard, 2004).
Another aspect to take into account is theorigin of the assemblage. As a result of thetrampling action of the chicks and adult owls,pellets that remain in the nest and non-ingesteddebris quickly become decomposed and broken(Andrews, 1990). Therefore, remains obtainedfrom nest accumulations tend to be morefragmented than those recovered from outsidethe nest. Considering materials of mixed origintogether (Sanchıs, 2000; Yravedra, 2004) hindersa clear picture of nest assemblages.
In all studies with eagle owls (Table 7), thefrequency of complete long bones is alwayssuperior to 10%, and the mean value of completeelements ranges from 45.9% to 75%. Incomparison with other predators (Table 8) suchas the Spanish Imperial eagle (0% and 27.9%;Lloveras et al., 2008a), the Iberian lynx (2.5% and43%; Lloveras et al., 2008b) and the coyote (0%and 7%; Schmitt & Juell, 1994), these percentagesare higher. A discrepancy is observed with regardto the frequency of complete long bones
Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa
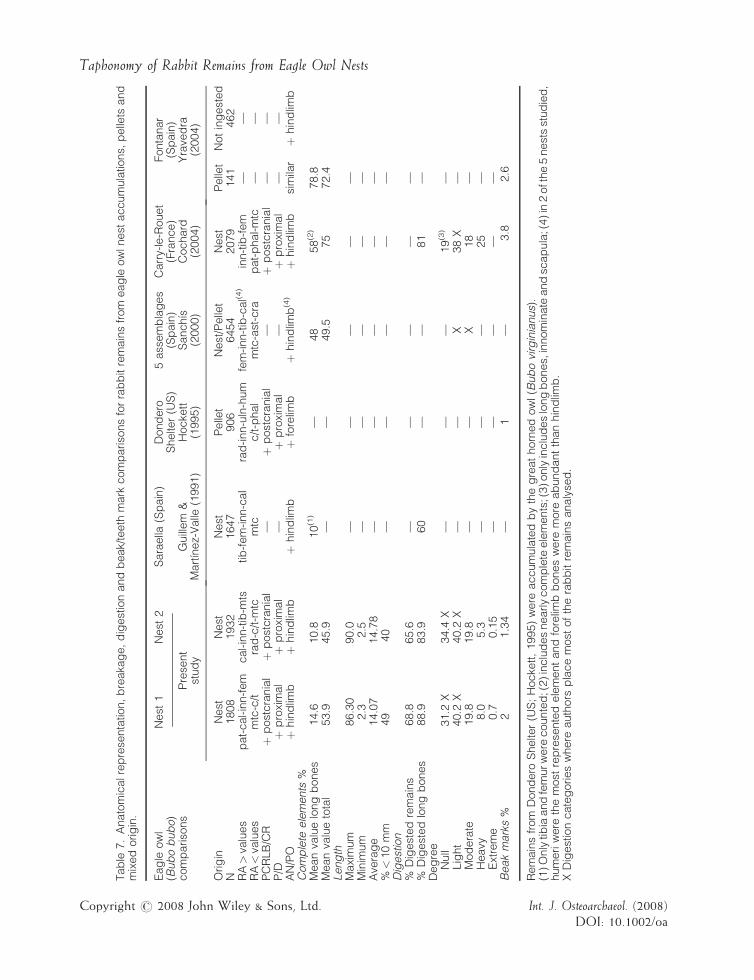
Tab
le7.
Anato
mic
alre
pre
senta
tion,
bre
akag
e,
dig
estion
and
beak/t
eeth
mark
com
parisons
for
rab
bit
rem
ain
sfr
om
eag
leow
lnest
accum
ula
tions,
pelle
tsand
mix
ed
orig
in.
Eag
leow
l(B
ubobubo)
com
parisons
Nest
1N
est
2S
ara
ella
(Sp
ain
)D
ond
ero
Shelter
(US
)5
assem
bla
ges
(Sp
ain
)C
arr
y-le-R
ouet
(Fra
nce)
Fonta
nar
(Sp
ain
)P
resent
stu
dy
Guill
em
&M
art
ınez-V
alle
(1991)
Hockett
(1995)
Sanchıs
(2000)
Cochard
(2004)
Yra
ved
ra(2
004)
Orig
inN
est
Nest
Nest
Pelle
tN
est/
Pelle
tN
est
Pelle
tN
ot
ing
este
dN
1808
1932
1647
906
6454
2079
141
462
RA>
valu
es
pat-
cal-in
n-f
em
cal-in
n-t
ib-m
tstib
-fem
-inn-c
al
rad
-inn-u
ln-h
um
fem
-inn-t
ib-c
al(4
)in
n-t
ib-f
em
——
RA<
valu
es
mtc
-c/t
rad
-c/t
-mtc
mtc
c/t
-phal
mtc
-ast-
cra
pat-
phal-m
tc—
—P
CR
LB
/CR
þp
ostc
rania
lþ
postc
rania
l—
þp
ostc
rania
l—
þp
ostc
rania
l—
—P
/Dþ
pro
xim
al
þp
roxim
al
—þ
pro
xim
al
—þ
pro
xim
al
——
AN
/PO
þhin
dlim
bþ
hin
dlim
bþ
hin
dlim
bþ
fore
limb
þhin
dlim
b(4
)þ
hin
dlim
bsim
ilar
þhin
dlim
bComplete
elements
%M
ean
valu
elo
ng
bones
14.6
10.8
10
(1)
—48
58
(2)
78.8
Mean
valu
eto
tal
53.9
45.9
——
49.5
75
72.4
Length
Maxim
um
86.3
090.0
——
——
—M
inim
um
2.3
2.5
——
——
—A
vera
ge
14.0
714.7
8—
——
——
%<
10
mm
49
40
——
——
—Digestion
%D
igeste
dre
main
s68.8
65.6
——
——
—%
Dig
este
dlo
ng
bones
88.9
83.9
60
——
81
—D
eg
ree
Null
31.2
X34.4
X—
——
19
(3)
—Lig
ht
40.2
X40.2
X—
—X
38
X—
Mod
era
te19.8
19.8
——
X18
—H
eavy
8.0
5.3
——
—25
—E
xtr
em
e0.7
0.1
5—
——
——
Beakmarks
%2
1.3
4—
1—
3.8
2.6
Rem
ain
sfr
om
Dond
ero
Shelter
(US
;H
ockett
,1995)
were
accum
ula
ted
by
the
gre
at
horn
ed
ow
l(B
ubovirginianus).
(1)O
nly
tib
iaand
fem
urw
ere
counte
d;(
2)in
clu
des
nearly
com
ple
teele
ments
;(3)only
inclu
des
long
bones,i
nnom
inate
and
scap
ula
;(4)in
2oft
he
5nests
stu
die
d,
hum
eri
were
the
most
rep
resente
dele
ment
and
fore
limb
bones
were
more
ab
und
ant
than
hin
dlim
b.
XD
igestion
cate
gories
where
auth
ors
pla
ce
most
of
the
rab
bit
rem
ain
sanaly
sed
.
Copyright # 2008 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa
Taphonomy of Rabbit Remains from Eagle Owl Nests

Tab
le8.
Anato
mic
alre
pre
senta
tion,
bre
akag
e,
dig
estion
and
beak/t
eeth
mark
com
parisons
for
rab
bit
rem
ain
saccum
ula
ted
by
eag
leow
l,d
iurn
alra
pto
rsand
terr
estr
ialcarn
ivore
s
Eag
leow
l(B
ubobubo)
and
oth
er
pre
dato
rsN
octu
rnalra
pto
rsD
iurn
alra
pto
rsTerr
estr
ial
carn
ivore
s
Eag
leow
lBubobubo
Pre
sent
stu
dy
Sp
anis
hIm
perial
eag
leAquila
adalberti
Gold
en
eag
leAquila
chrysaetos
Iberian
lynx
Lynxpardinus
Coyote
Canis
latrans
Nest
1N
est
2Llo
vera
setal.
(2008a)
Hockett
(1996)
Hockett
(1995)
Schm
itt
(1995)
Llo
vera
setal.
(2008b
)S
chm
itt
&Juell
(1994)
Orig
inN
est
Nest
Pelle
tP
elle
tN
est
Nest
Scat
Scat
N1808
1932
824
33
930
909
1522
840
RA>
valu
es
pat-
cal-in
n-f
em
cal-in
n-t
ib-m
tsp
hal3-
um
ol-
tib
—cal-ast-
tib
-fem
tib
-cal-fe
m-inn
man-t
eeth
-cra
—R
A<
valu
es
mtc
-c/t
rad
-c/t
-mtc
rib
-fem
-rad
-ver
—cra
-sc-h
um
cra
-sc-u
lnc/t
-ver-
rib
s—
PC
RLB
/CR
þp
ostc
rania
lþ
postc
rania
lþ
cra
nia
l—
þp
ostc
rania
lþ
postc
rania
lþ
postc
rania
l—
P/D
þp
roxim
al
þp
roxim
al
þd
ista
l—
þd
ista
leq
ual
þp
roxim
al
—A
N/P
Oþ
hin
dlim
bþ
hin
dlim
bþ
poste
rior
—þ
poste
rior
þp
oste
rior
þante
rior
—Complete
elements
%M
ean
valu
elo
ng
bones
14.6
10.8
066.6
—30.3
2.5
0M
ean
valu
eto
tal
53.9
45.9
27
——
35.1
�43
7Length
(inmm)
Maxim
um
86.3
90.0
36.1
80
——
30.1
35
Min
imum
2.3
2.5
1.8
——
—1.1
Avera
ge
14.0
714.7
88.3
6—
——
7.1
%<
10
mm
49
40
73
——
—80
75
Digestion
%D
igeste
dre
main
s68.8
65.6
98
Majo
rity
——
97.2
100
%D
igeste
dlo
ng
bones
88.9
83.9
100
——
100
100
Deg
ree
Null
31.2
X34.4
X2.0
——
—2.8
—Lig
ht
40.2
X40.2
X18.2
——
—12.0
—M
od
era
te19.8
19.8
46.8
X—
—22.0
X—
Heavy
8.0
5.3
27.4
X—
X43.8
X—
Extr
em
e0.7
0.1
55.6
——
—19.3
X—
Beak/teeth
marks
%2.0
1.3
40.5
——
0.3
0.2
60.9
� Teeth
,vert
eb
rae,
rib
s,
carp
al/ta
rsaland
phala
ng
es
not
inclu
ded
.X
Dig
estion
cate
gories
where
auth
ors
pla
ce
most
of
the
rab
bit
rem
ain
sanaly
sed
.
Copyright # 2008 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa
L. Lloveras, M. Moreno-Garcıa and J. Nadal
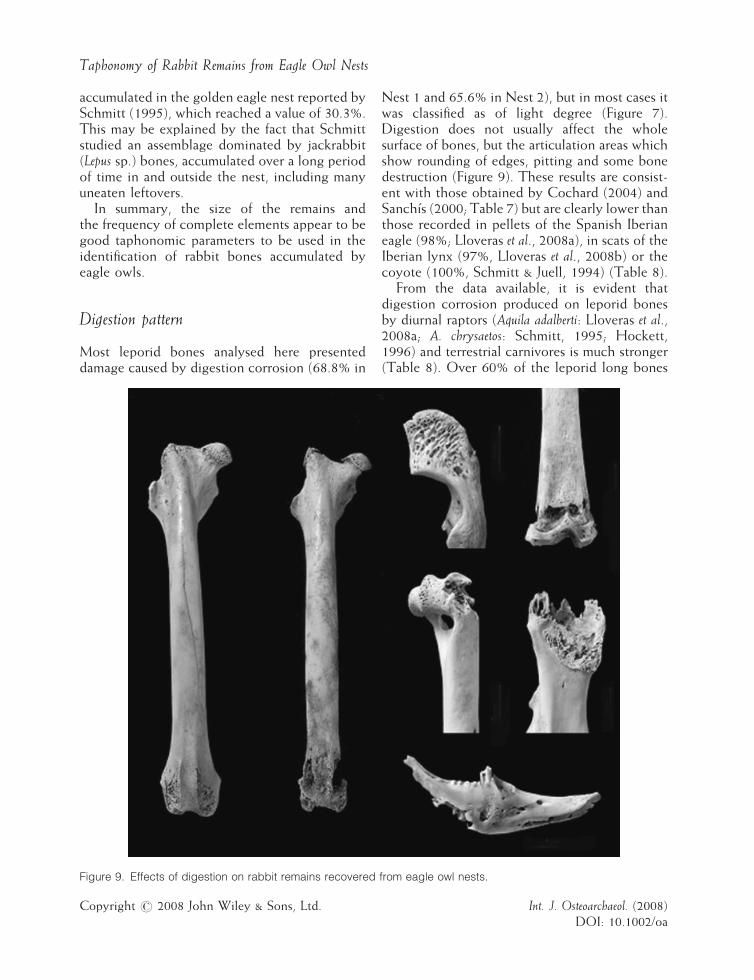
Taphonomy of Rabbit Remains from Eagle Owl Nests
accumulated in the golden eagle nest reported bySchmitt (1995), which reached a value of 30.3%.This may be explained by the fact that Schmittstudied an assemblage dominated by jackrabbit(Lepus sp.) bones, accumulated over a long periodof time in and outside the nest, including manyuneaten leftovers.In summary, the size of the remains and
the frequency of complete elements appear to begood taphonomic parameters to be used in theidentification of rabbit bones accumulated byeagle owls.
Digestion pattern
Most leporid bones analysed here presenteddamage caused by digestion corrosion (68.8% in
Figure 9. Effects of digestion on rabbit remains recovered
Copyright # 2008 John Wiley & Sons, Ltd.
Nest 1 and 65.6% in Nest 2), but in most cases itwas classified as of light degree (Figure 7).Digestion does not usually affect the wholesurface of bones, but the articulation areas whichshow rounding of edges, pitting and some bonedestruction (Figure 9). These results are consist-ent with those obtained by Cochard (2004) andSanchıs (2000; Table 7) but are clearly lower thanthose recorded in pellets of the Spanish Iberianeagle (98%; Lloveras et al., 2008a), in scats of theIberian lynx (97%, Lloveras et al., 2008b) or thecoyote (100%, Schmitt & Juell, 1994) (Table 8).From the data available, it is evident that
digestion corrosion produced on leporid bonesby diurnal raptors (Aquila adalberti: Lloveras et al.,2008a; A. chrysaetos: Schmitt, 1995; Hockett,1996) and terrestrial carnivores is much stronger(Table 8). Over 60% of the leporid long bones
from eagle owl nests.
Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa

L. Lloveras, M. Moreno-Garcıa and J. Nadal
digested by these predators are grouped aroundthe moderate and heavy degrees of corrosion.Thus, this seems a parameter worth recording todetermine different predator agents.
Puncture marks
It is also noteworthy that puncture marks causedby the beak of the eagle owl are relativelyabundant (Table 7), with a percentage alwayssuperior to 1% and with a mean value of 2.15%.Although references to these kinds of marksproduced by diurnal raptors are scarce, theirfrequency appears to be lower than thatregistered in leporid assemblages accumulatedby Bubo bubo (Table 8).
Conclusion
The analysis presented here has shown thatidentifying diagnostic features of eagle owls’leporid accumulations is not a straightforwardissue. The reasons are two-fold. Firstly, thenumber of works dealing with this topic is stillvery scarce. Samples are small, no systematicmethodology of analysis has been followed, anddata for other nocturnal and diurnal raptors oreven terrestrial carnivores do not allow trust-worthy comparisons. Secondly, most nestassemblages originate from a combination ofdegraded pellets and uneaten food remains.Since raptors may differentially consume certainprey body parts, knowing the contribution madeby each of these kinds of remains appears to bean important issue. Consequently, anatomicalrepresentation of prey in nest assemblages mayvary for the same predator according to thehigher or lower presence of ingested remains.Probably because this is a difficult issue to bequantified, it has not been taken into accountin most taphonomic studies. In our case,the situation was overcome by considering thecontribution of digested material. Hence weconcluded that our eagle owl nest accumulationscomprised a large proportion of degradedpellets, which may not stand in other cases.For these reasons, it seems that anatomical
Copyright # 2008 John Wiley & Sons, Ltd.
representation on its own is a very variableparameter, dependent upon a wide number ofissues such as availability and age of prey,number and age of chicks, and so on, that for thetime being have not been properly assessed,hampering the definition of a pattern charac-teristic of any particular predator.
According to our results, eagle owl nestingsites are characterised by high values forrelative abundance of posterior skeletal elements,more hindlimbs than forelimb bones, and morepostcranial than cranial elements. As shown inTable 8, these are features partially shared bydiurnal raptors, so they cannot be considered astruly diagnostic of eagle owls. In fact, it appearsthat breakage and degrees of digestive corrosionmay be more diagnostic factors to distinguishbetween different rabbit predators. Diurnalraptors and terrestrial carnivores tend to breakthe bones of their victims more than eagle owls(Table 8). Thus, their prey bone averagemaximum length is 15mm in contrast with valuesof 8.36mm and 7.1mm, recorded for Aquilaadalberti and Lynx pardinus respectively (Lloveraset al., 2008a,b). Such differences are more evidentif the degrees of digestive corrosion are con-sidered. While over 40% of digested remains ineagle owl nest assemblages were classified aslightly corroded, A. adalberti pellets showed thesame proportion of leporid remains moderatelydigested, and Iberian lynx scats registered thatpercentage as heavily corroded.
In summary, it is proposed that fossil assem-blages may be confidently attributed to eagleowls if most of the criteria described above aremet, particularly if breakage patterns are moder-ate and there is only a light degree of digestioncorrosion.
Despite this, it is clear that more detailed workneeds to be done with rabbit remains accumu-lated by nocturnal and diurnal avian raptors, tounderstand their taphonomic signatures fully. Inparticular, material from roosting sites and pelletsof adult individuals need to be researched. Toaccomplish such a task it has become evident thatthe analysis cannot be reduced to recording oneor two variables. Within the main topicsconcerned with anatomical representation,breakage and digestion patterns, a broad spec-trum of indices, measurements and quantitative
Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa

Taphonomy of Rabbit Remains from Eagle Owl Nests
data must be considered. Finally, in studies withmodern material, it would be worth under-standing birds’ ethology of killing and consump-tion as well as their breeding strategies.
Acknowledgements
We are very grateful to Santi Llacuna and Rodrigodel Amo from Parc del Garraf (Barcelona, Spain)who supplied us with the Iberian leporid remainsfrom eagle owl nests to carry out this study. Z.Bochenski is also acknowledged for kindly send-ing us copies of his papers. Thanks to Ze PauloRuas for the photos in Figure 9.We thank the tworeferees that reviewed this paper for their veryuseful comments, and particularly Bryan Hockettfor stimulating us to continue this line of research.A research grant (SFRH/32025/2006) from
Portuguese Fundacao para a Ciencia e a Tecno-logia (FCT) to Lluıs Lloveras, financial supportfrom research project HAR2008-00103 from theMinisterio de Educacion y Ciencia, and fromSGR2005-00299 from the Generalitat de Catalu-nya are gratefully acknowledged.
References
Amo R, Gonzalez D. 1998. L’alimentacio del duc (Bubobubo) al Massıs del Garraf. II Trobada d’estudiosos delGarraf, Monografies 26. Diputacio de Barcelona:Barcelona; 47–51.
Andrews P. 1990. Owls, Caves and Fossils. BritishMuseum of Natural History: London.
Bochenski ZM, Tomek T. 1997. Preservation of birdbones: erosion versus digestion by owls. InternationalJournal of Osteoarchaeology 7: 372–387.
Bochenski ZM, Tomek T, Boev Z, Mitev I. 1993.Patterns of bird bone fragmentation in pellets ofthe TawnyOwl (Strix aluco) and the Eagle Owl (Bubobubo) and their taphonomic implications. Acta Zool-ogica Cracoviensa 36: 313–328.
Cochard D. 2004. Les leporides dans la subsistance Paleo-lithique du sud de la France. PhD thesis, UniversiteBordeaux I.
Cochard D. 2008. Discussion sur la variabilite intrar-eferentiel d’accumulations osseuses de petits preda-teurs. Annales de Paleontologie 94: 89–101.
Copyright # 2008 John Wiley & Sons, Ltd.
Cruz-Uribe K, Klein RG. 1998. Hyrax and hare bonesfrom modern South African eagle roost and thedetection of eagle involvement in fossil boneassemblages. Journal of Archaeological Science 25:135–147.
Delibes M, Hiraldo F. 1981. The rabbit as a prey inthe Iberian Mediterranean ecosystem. In WorldLagomorph Conference, Myers K, MacInnes CD(eds). Guelph: Canada; (1979); 614–622.
Dodson P, Wexlar D. 1979. Taphonomic investi-gations of owl pellets. Paleobiology 5: 275–284.
Donazar JA. 1987. Geographic variation in the diet ofEagle Owls in western Mediterranean Europe. InBiology and Conservation of Northern Forest Owls: Sym-posium Proceedings, Nero RW (ed.). Winnipeg: Man-itoba, Canada; 220–224.
Donazar JA. 1988. Variaciones en la alimentacionentre adultos reproductores y pollos en el BuhoReal (Bubo bubo). Ardeola 35: 278–284.
Donazar JA. 1989. Variaciones geograficas y estacio-nales en la alimentacion del Buho Real (Bubo bubo) enNavarra. Ardeola 36: 25–39.
Fernandez-Jalvo Y, Andrews P. 1992. Small mammaltaphonomy of Gran Dolina, Atapuerca (Burgos),Spain. Journal of Archaeological Science 19: 407–428.
Guillem P, Martınez-Valle R. 1991. Estudio de laalimentacion de las rapaces nocturnas aplicado ala interpretacion del registro faunıstico arqueolo-gico. Saguntum 24: 23–34.
Hiraldo F, Andrada J, Parreno FF. 1975. Diet of theEagle Owl (Bubo bubo) in Mediterranean Spain. ActaVertebrata 2: 161–177.
Hockett BS. 1989. Archaeological significance of rab-bit–raptor interactions in Southern California.NorthAmerican Archaeologist 10: 123–139.
Hockett BS. 1991. Toward distinguishing human andraptor patterning on leporid bones. American Anti-quity 56: 667–679.
Hockett BS. 1995. Comparison of leporid bones inraptor pellets, raptor nests, and archaeological sitesin the Great Basin. North American Archaeologist 16:223–238.
Hockett BS. 1996. Corroded, thinned and polishedbones created by golden eagles (Aquila chrysaetos):taphonomic implications for archaeologicalinterpretations. Journal of Archaeological Science 23:587–591.
Hockett BS, Bicho NF. 2000. The rabbits of PicareiroCave: small mammal hunting during the late UpperPaleolithic in the Portuguese Estremadura. Journal ofArchaeological Science 27: 715–723.
Hockett BS, Haws JA. 2002. Taphonomic and meth-odological perspectives of leporid hunting duringthe Upper Paleolithic of the western Mediterranean
Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa

L. Lloveras, M. Moreno-Garcıa and J. Nadal
basin. Journal of Archaeological Method and Theory 9:269–302.
Hoffman R. 1988. The contribution of raptorial birdsto patterning in small mammal assemblages. Paleo-biology 14: 81–90.
Laroulandie V. 2000. Taphonomie et archeozoologie desoiseaux en Grotte: applications aux sites Paleolithiques duBois-Ragot (Vienne), de Combe Sauniere (Dordogne) et de laVache (Ariege). PhD thesis, Universite de Bordeaux I.
Laroulandie V. 2002. Damage to pigeon long bones inpellets of the Eagle Owl Bubo bubo and food remainsof peregrine falcon Falco peregrinus: zooarchaeologi-cal implications. Proceedings of the 4th Meeting ofthe ICAZ Bird Working Group Krakow, Poland,2001. Acta Zoologica Cracoviensia 45 (spec. issue):331–339.
Lloveras L, Moreno-Garcıa M, Nadal J. 2008a. Tapho-nomic study of leporid remains accumulated by theSpanish Imperial Eagle (Aquila adalberti). Geobios 41:91–100.
Lloveras L, Moreno-Garcıa M, Nadal J. 2008b. Tapho-nomic analysis of leporid remains obtained frommodern Iberian Lynx (Lynx pardinus) scats. Journal ofArchaeological Science 35: 1–13.
Lourenco R. 2006. The food habits of Eurasian Eagle-Owls in Southern Portugal. Journal of Raptor Research10: 297–300.
Mikkola H. 1983. Owls of Europe. T. and A.D. Poyser:Calton.
Mikkola H. 1994. Eagle owl. In Birds in Europe: theirConservation Status. BirdLife Conservation Series No. 3,Tucker GM, Heath MF (eds). Birdlife Inter-national: Cambridge; 326–327.
Mondini M. 2002. Carnivore taphonomy and the earlyhuman occupations in the Andes. Journal of Archae-ological Science 29: 791–801.
Perez Mellado V. 1980. Alimentacion del Buho Real(Bubo bubo) en Espana Central. Ardeola 25: 93–112.
Perez Ripoll M. 1993. Las marcas tafonomicas enhuesos de lagomorfos. In Estudios sobre el Cuaternario.Medios sedimentarios. Cambios ambientales. Habitat humano,Fumanal MP, Bernabeu J (eds). Universidad deValencia: Valencia; 227–231.
Copyright # 2008 John Wiley & Sons, Ltd.
Perez Ripoll M. 2004. La consommation humaine deslapins pendant le Paleolithique dans la region deValencia (Espagne) et l’etude des niveaux gravetiensde La Cova de les Cendres (Alicante). In Petitsanimaux et societes humaines, Brugal JP, Desse J(eds). Editions APDCA: Antibes; 191–206.
Sanchis A. 2000. Los restos de Oryctolagus cuniculus enlas tafocenosis de Bubo bubo y Vulpes vulpes y suaplicacion a la caracterizacion del registro faunısticoarqueologico. Saguntum 32: 31–50.
Schmitt DN. 1995. The taphonomy of Golden Eagleprey accumulations at Great Basin roosts. Journal ofEthnobiology 15: 237–256.
Schmitt DN, Juell KE. 1994. Toward the identificationof coyote scatological faunal accumulations inarchaeological contexts. Journal of ArchaeologicalScience 21: 249–262.
Serrano D. 1998. Diferencias interhabitat en la ali-mentacion del Buho Real (Bubo bubo) en el vallemedio del Ebro (NE de Espana): efecto de la dis-ponibilidad de conejo (Oryctolagus cuniculus). Ardeola45: 35–46.
Stiner MC, Munro ND, Surowell TA. 2000. Thetortoise and the hare: small-game use, the broad-spectrum revolution, and Paleolithic demography.Current Anthropology 41: 39–73.
Stiner MC, Munro ND, Surowell TA, Tchernov E,Bar-Yosef O. 1999. Paleolithic population growthpulses evidenced by small animal exploitation.Science 283: 190–194.
Villaverde V, Martınez-Valle R, Guillem P, FumanalM. 1996. Mobility and the role of small game in theMiddle Paleolithic of the central region of theSpanish Mediterranean: a comparison of CovaNegra with other Paleolithic deposits. In The LastNeandertals, the First Anatomically Modern Humans,Carbonell E (ed.). Cambridge University Press:Cambridge; 267–288.
Yravedra J. 2004. Implications taphonomiques desmodifications osseuses faites par les vrais hiboux(Bubo bubo) sur les lagomorphes. In Petits animaux etsocietes humaines, Brugal JP, Desse J (eds). EditionsAPDCA: Antibes; 321–324.
Int. J. Osteoarchaeol. (2008)DOI: 10.1002/oa




















