
My Visits to the Hmong in the Triangle of Guizhou, Sichuan ...

ARTICLE IN PRESS
Quaternary International 126–128 (2005) 271–282
*Correspondin
E-mail addres
sarah@stoutami
(D.A. Bekken).
1040-6182/$ - see
doi:10.1016/j.qua
Stegodon orientalis from Panxian Dadong, a Middle Pleistocenearchaeological site in Guizhou, South China: taphonomy, population
structure and evidence for human interactions
L.A. Schepartza,*, S. Stoutamirea, D.A. Bekkenb
aDepartment of Anthropology, University of Cincinnati, P.O. Box 210380, Cincinnati, OH 45221-0380, USAbField Museum of Natural History, Roosevelt Rd. at Lakeshore Drive, Chicago, IL 60605-2496, USA
Available online 11 June 2004
Abstract
Panxian Dadong is a large karstic cave in the mountains of western Guizhou province. Archaeological excavations at Dadong
have yielded a rich Ailuropoda-Stegodon faunal sample in association with stone artifacts and human remains dating to 130–300 ka.
Stegodon orientalis is an important component of the faunal sample, which is dominated by large-bodied mammals. This study
describes the sample of stegodont dental and skeletal material, provides an age at death profile for the stegodont sample, and
evaluates the results in terms of taphonomy, site formation processes, and comparisons with other Middle Pleistocene
archaeological sites with evidence for human use of proboscideans.
The Dadong stegodont sample consists primarily of dental remains of young individuals less than 12 years of age with a minimum
of 12 stegodonts represented. Based on the available archaeological, faunal and taphonomic data, the stegodont faunal assemblage
at Dadong is probably a result of hominid activities (either hunting or scavenging) and the natural accumulation of young
stegodonts from accidental deaths. This study supports the results of other analyses (Schepartz et al., 2003) that suggest humans
played an important role in the formation of the Dadong faunal assemblage.
r 2004 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
This paper describes the analysis of Middle Pleisto-cene Stegodon orientalis material excavated from Pan-xian Dadong cave in Guizhou province, southern China.Stegodonts are commonly found in the Pleistocene cavedeposits of southern China, but the process by whichthey became elements of faunal assemblages is rarelystudied. Generally, they are just identified and includedin faunal lists that describe paleontological localities andarchaeological sites. The clear presence of humans atPanxian Dadong is documented by five isolated teeth,numerous stone tools made of limestone, chert andbasalt (Miller-Antonio et al., 2004), tools made ofRhinoceros sinensis teeth (Miller-Antonio et al., 2000),and cutmarks, percussion damage and burning on
g author:.
ses: [email protected] (L.A. Schepartz),
re.com (S. Stoutamire), [email protected]
front matter r 2004 Elsevier Ltd and INQUA. All rights
int.2004.04.026
faunal material (Schepartz et al., 2003). This evidencemakes it appropriate to evaluate the possible role playedby humans in the formation of the stegodont sample.
Our interest in the stegodonts at Panxian Dadong ispart of a larger faunal analysis designed to (1) provideinformation on the paleoenvironment of upland south-ern China during the Middle Pleistocene, (2) investigatethe interactions between prehistoric humans and mam-malian species, and (3) clarify the role of humans,carnivores and natural geological processes in theformation of the Panxian Dadong faunal assemblage.Our research incorporates paleontological, geological,taphonomic and archaeological data.
Stegodonts were present from the late Pliocenethrough the Pleistocene of Asia, and they were closelyrelated to the ancestors of the contemporary elephantgenera Loxodonta and Elephas. Stegodon is the mostcommon genus, with S. orientalis having the greatesttemporal range and geographical distribution (Zong, 1995;Saegusa, 1996, Fig. 4 in Takahashi and Namatsu, 2000).The importance of Stegodon in Chinese mammalian
reserved.
ARTICLE IN PRESSL.A. Schepartz et al. / Quaternary International 126–128 (2005) 271–282272
faunas is reflected in the term ‘Ailuropoda-Stegodon
assemblage’ that is commonly used to refer to southernChinese faunas of the Pleistocene. There are two co-existing species of Stegodon known from the MiddlePleistocene caves of South China. In addition to S.
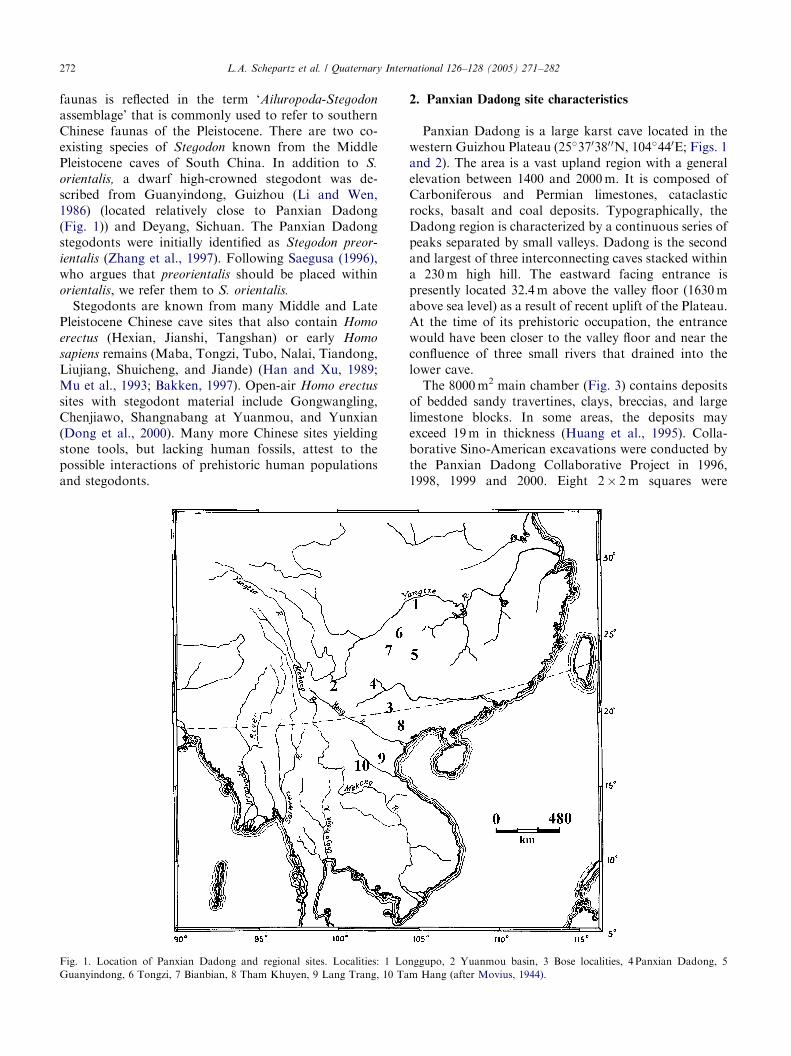
orientalis, a dwarf high-crowned stegodont was de-scribed from Guanyindong, Guizhou (Li and Wen,1986) (located relatively close to Panxian Dadong(Fig. 1)) and Deyang, Sichuan. The Panxian Dadongstegodonts were initially identified as Stegodon preor-
ientalis (Zhang et al., 1997). Following Saegusa (1996),who argues that preorientalis should be placed withinorientalis, we refer them to S. orientalis.
Stegodonts are known from many Middle and LatePleistocene Chinese cave sites that also contain Homo
erectus (Hexian, Jianshi, Tangshan) or early Homo
sapiens remains (Maba, Tongzi, Tubo, Nalai, Tiandong,Liujiang, Shuicheng, and Jiande) (Han and Xu, 1989;Mu et al., 1993; Bakken, 1997). Open-air Homo erectus
sites with stegodont material include Gongwangling,Chenjiawo, Shangnabang at Yuanmou, and Yunxian(Dong et al., 2000). Many more Chinese sites yieldingstone tools, but lacking human fossils, attest to thepossible interactions of prehistoric human populationsand stegodonts.
Fig. 1. Location of Panxian Dadong and regional sites. Localities: 1 Lo
Guanyindong, 6 Tongzi, 7 Bianbian, 8 Tham Khuyen, 9 Lang Trang, 10 Ta
2. Panxian Dadong site characteristics
Panxian Dadong is a large karst cave located in thewestern Guizhou Plateau (25�3703800N, 104�440E; Figs. 1and 2). The area is a vast upland region with a generalelevation between 1400 and 2000m. It is composed ofCarboniferous and Permian limestones, cataclasticrocks, basalt and coal deposits. Typographically, theDadong region is characterized by a continuous series ofpeaks separated by small valleys. Dadong is the secondand largest of three interconnecting caves stacked withina 230m high hill. The eastward facing entrance ispresently located 32.4m above the valley floor (1630mabove sea level) as a result of recent uplift of the Plateau.At the time of its prehistoric occupation, the entrancewould have been closer to the valley floor and near theconfluence of three small rivers that drained into thelower cave.
The 8000m2 main chamber (Fig. 3) contains depositsof bedded sandy travertines, clays, breccias, and largelimestone blocks. In some areas, the deposits mayexceed 19m in thickness (Huang et al., 1995). Colla-borative Sino-American excavations were conducted bythe Panxian Dadong Collaborative Project in 1996,1998, 1999 and 2000. Eight 2� 2m squares were
nggupo, 2 Yuanmou basin, 3 Bose localities, 4 Panxian Dadong, 5
m Hang (after Movius, 1944).

ARTICLE IN PRESS
Fig. 2. Panxian Dadong environment. The cave as viewed from a distance of approximately 10 km. This mountainous karst landscape is typical of
the Guizhou Plateau.
Fig. 3. Panxian Dadong main chamber excavations.
L.A. Schepartz et al. / Quaternary International 126–128 (2005) 271–282 273
excavated to a maximum depth of over 5m (Fig. 3inset). The stratigraphic profile in the excavation areaconsists of three major layers (Fig. 4). The top layer is a
thick cap of travertine and clay. No artifacts or fossilswere recovered from this layer. The second layer is abrown breccia with a low density of artifacts and animal

ARTICLE IN PRESS
Fig. 4. Panxian Dadong Stratigraphy. There are three major layers: a top layer of capping travertine and clay (app. 97.55–96.0), a second layer of
breccia (app. 96.0–95.0), and a third layer of clays with abundant stone artifacts and fauna (app. 95.0 and below). Weathered crusts represent
depositional hiatuses. Figure provided by P. Karkanas.
L.A. Schepartz et al. / Quaternary International 126–128 (2005) 271–282274
bone. Most of the stone artifacts and bone wererecovered from a third layer of dense brownish-redclay. Stratigraphic and micromorphological studies ofthe deposits document paleoclimatic changesthrough the sequence, as well as freeze-thaw activityand depositional hiatuses. Uranium-series dates onspeleothems and travertines (Shen et al., 1997) andelectron spin resonance (ESR) dates of tooth enamel(Rink et al., 2003) suggest that most of the investigatedDadong archaeological deposits are between 130–350 kain age.
3. The Panxian Dadong faunal sample
The Dadong fauna has a variety of species character-istic of the Ailuropoda-Stegodon faunas of southernChina (Pan and Yuan, 1997; Zhang et al., 1997;Schepartz et al., 2003). It is dominated by large-bodiedmammals, including animals that would not ordinarily
inhabit caves, such as Stegodon, Rhinoceros, and thegiant tapir Megatapirus. Other forms representedare primates (macaques, colobines and hominids), arange of ungulates (cervids, small and large bovids,pigs), and carnivores (mustelids, foxes, hyenas, tigers,leopards). This diversity of species suggests thatthe Pleistocene environment was a mixed woodland,as indicated by the presence of giant tapir, muskdeer, barking deer and rhinoceros. The occurrenceof pandas, bamboo rats and colobine monkeyssuggest some densely forested areas including bamboostands.
The mammalian faunal sample (N ¼ 7045) consists of3.3% cranial fragments, 12.5% dental elements and84% postcranial elements. The most common identifiedspecies is Rhinoceros sinensis, which comprises 24% ofthe total elements identifiable to taxon, followed byStegodon at 13%. Carnivores are quite rare, andrepresent less than 4% of the specimens identified totaxon (Schepartz et al., 2003).

ARTICLE IN PRESSL.A. Schepartz et al. / Quaternary International 126–128 (2005) 271–282 275
4. The stegodont component
The stegodont sample consists of 215 isolated orfragmentary teeth, four skull fragments, and 45 post-cranial elements (Fig. 5) yielding an NISP of 264 ofwhich over 81% is dentition. The postcrania include 10axial elements and 35 appendicular elements. Foot andankle elements are the most numerous category. Four ofthe elements are clearly from juvenile individuals (threeshaft fragments lacking epiphyses, and a sternum).Based on the number and representation of skeletalelements (the most numerous single bone is the cranium)and their stratigraphic positions, the minimum numberof stegodonts preserved at Dadong is three. The numberand diversity of dental remains suggests that a fargreater number of individuals are represented.
5. Stegodont dental terminology
Stegodonts had 24 cheek teeth with 6 in each jawquadrant. They also had small deciduous tusks andlarge permanent tusks (of which only small fragmentsare preserved at Dadong). During most of the life ofproboscideans, two teeth are usually in use at any time.
Fig. 5. Stegodon skeletal element representation.
Fig. 6. Crest and Enamel Measurements. Crest measurements shown on t
Maximum height, 2 Base to ridge at midpoint, 3 Crest length, 4 Width at
fragmentary specimen (the occlusal surface toward the top of the page): 6 E
Teeth are shed as they are replaced by the successivetooth in the jaw. Functional wear reduces the maximumlength of the tooth as the anterior crests eventually wearaway. Worn teeth are eventually swallowed or drop outof the mouth (Haynes, 1991).
Tassy (1996) has written on the complex nomencla-ture used to describe proboscidean teeth. The termsused in this paper are very general so that they maybe applied systematically to fragmentary material(see Fig. 6). The tooth class designations dp2, dp3,dp4, M1, M2, and M3 are used. Examples of thedifferent tooth classes are provided in Fig. 7. As inother proboscideans, Stegodon teeth of differentclasses can be distinguished by crest number becausethe tooth classes have increasingly greater numbersof crests. There is also a progressive increase intooth size, although the M1 and M2 are similar inmesiodistal length, which results in a decrease in crestsize between the M1 and M2 (Colbert and Hooijer,1953).
6. Identifying fragmentary dental specimens
The fragmentary nature of the Dadong stegodontdental remains complicates their identification to specifictooth classes. Only 21 of the 215 specimens havecomplete crowns with all crests represented. To assistin tooth class assignment, the Dadong material iscompared with the larger Yanjinggou sample fromSichuan province, an important collection of Ailuropo-
da-Stegodon fauna (Colbert and Hooijer, 1953).Specimens were initially separated into two groups:
those identifiable to class (Group 1, N ¼ 41; with156 crests complete enough for analysis) and those
he 3rd crest of a dp3 (occlusal view to the left, buccal at center): 1
lateral end, 5 Width at midpoint. Enamel measurements shown on a
namel at apex of crest, 7 Enamel at center of crest face.

ARTICLE IN PRESS
Fig. 7. Examples of Tooth Classes. Unless otherwise noted, views are
of the occlusal surface with the mesial aspect oriented toward the top
of the page. (A) maxillary dp2, (B) lateral view of mandibular dp2,
occlusal surface toward top of page; (C) maxillary dp3, (D)
mandibular dp3, (E) mandibular dp4, (F) maxillary M3.
L.A. Schepartz et al. / Quaternary International 126–128 (2005) 271–282276
unidentified to class (Group 2, N ¼ 174; with 207 crestsanalyzed). The majority of deciduous premolars inGroup 1 were identifiable to quadrant, although it wasdifficult to assign molars to class and jaw position. Formost purposes in this paper, they are grouped togetheras ‘Ms’.
Each Group 1 specimen was described, drawn,photographed, and measured (Fig. 6). The results werethen used to evaluate Group 2. Variables used included:maximum (mesiodistal) tooth length, crown height,maximum crest height, base to crest along midlineheight, crest length (buccolingual diameter of eachcrest), crest width at lateral edges, crest width atmidpoint, enamel thickness at crest apex, enamelthickness at midpoint of crest face, number of conelets,crest occlusal wear score, dentin exposure score, andcalculus thickness score.
It is important to recognize that a potential bias inusing crest measurements from fragmentary teeth maybe created by the loss of the anterior portions. Sincethose areas wear first and are more likely to bebroken off, posterior crests may be disproportionatelyrepresented in the record. In teeth that widen ornarrow posteriorly or have a range of diminishing crestsizes from front to back, the consistent loss of aparticular portion of the teeth affects the use of crestsize measurements to determine tooth classes. Inaddition, as Stegodon teeth may show some sexualdimorphism (Osborne, 1942), class identification bysize alone is problematic, particularly because theranges of measurements for particular classes overlapconsiderably.
7. Characteristics of the tooth classes at Panxian Dadong
Table 1 provides data for the Group 1 specimens andsome comparative data from Yanjinggou. The speci-mens from Panxian Dadong differ metrically from thosefound at Yanjinggou, as might be expected in differentpopulations of a wide-ranging species. The number ofcrests within each class of deciduous tooth is compar-able, but teeth from Panxian Dadong are generallysmaller in length and the penultimate crests of dp3s,dp4s, and Ms are not as high.
Penultimate ridge crest height, the measurementColbert and Hooijer (1953) used to characterize teethby class, shows progressive increase between successivetooth classes. Unfortunately, the ranges for thesemeasures on the three molar tooth classes overlap(Table 29, Colbert and Hooijer, 1953). For the Dadongsample, this variable (termed Median Penultimate CrestHeight in this study) is a fairly effective discriminatorbetween deciduous tooth classes, but preservation andidentification of the penultimate crest is infrequent. Itwas therefore necessary to use several measures toconfidently assign the fragmentary specimens to toothclass. Figs. 8 and 9 provide examples of bivariate plotsof Group 1 data that were used to classify fragmentaryteeth in Group 2.
8. Tooth class distributions
Table 2 is the tooth class distribution for both groups.Group 1 is dominated by deciduous premolars, whileGroup 2, although still dominated by deciduouspremolars, contains a greater number of molars. Forthose teeth that can be identified to a quadrant, thenumbers of maxillary and mandibular teeth areapproximately equal. Based on Table 2, the largestspecific class is dp4, with a total of 47 specimens. Thiswould mean an MNI of 12 stegodonts.

ARTICLE IN PRESS
Table 1
Quantitative measurements (mm) of teeth from Group 1 and Yanjinggou
dp2 dp3 dp4 molars
Maximum N 13 15 9 4
Maximum length N ¼ 1114:9223:8 (18.2)�
[N ¼ 1113:0225:0 (20.0)]
N ¼ 640:0258:0 (57.5)
[N ¼ 2344:0272:0 (61.0)]
N ¼ 475:2295:6 (87.2)
[N ¼ 1197:02130:0 (117)]
N ¼ 2140:02218:0 (179.0)
[N ¼ 13171:02360:0(217.8)]
Median penultimate
crest height
N ¼ 410:1214:8 (13.0)
[N ¼ 211:0212:0 (11.5)]
N ¼ 316:2217:7 (17.5)
[N ¼ 1516:0228:0 (21.4)]
N ¼ 222:3225:1 (23.7)
[N ¼ 1126:0235:0 (30.4)]
N ¼ 230:5237:8 (31.4)
[N ¼ 1240:0255:0 (47.0)]
# Conelets/crest 2–6 (4) 11 10–12 (11) 10–11 (11)
Max. crest height 9.7–17.8 (14.8) 14.3–27.7 (17.7) 12.4–29 (23.85) 30.0–47.8 (39.8)
Center enamel
thickness
1.0–1.5 (1.2) 1.1–1.8 (1.4) 1.1–3.3 (1.7) 4.2–4.8 (4.3)
Midpoint crest width 3.4–18 (8.7) 5.1–10.78 (10.3) 4.0–17.2 (13.9) 17.0–27.5 (22.0)
Crest length (BL width) 8.2–23.3 (14.5) 20.2–43.8 (32.9) 36.2–53.8 (45.9) 45.4–83.3 (74.6)
�Means are in brackets. Data for Yanjinggou from Colbert and Hooijer (1953) are in smaller font. Colbert and Hooijer did not specify how their
measures were taken; our measures should therefore be regarded as roughly comparable to theirs.
Fig. 8. Maximum crest height plotted against crest length for Group 1.
Fig. 9. Midpoint crest width plotted against crest length for Group 1.
Table 2
Tooth class distribution
Tooth Group 1 Group 2 Total
dp2 13 4 17
dp3 15 30 45
dp4 9 38 47
M 4 48 52
Tusk 0 1 1
Tooth germs 0 4 4
Undentified 0 49 49
41 174 215
Table 3
Generalized tooth class distribution
Tooth Group 1 Group 2 Total
dp 37 93 130
M 4 48 52
Unidentified 0 28 28
Tusk 0 1 1
Tooth germs 0 4 4
41 174 215
L.A. Schepartz et al. / Quaternary International 126–128 (2005) 271–282 277
As many specimens in Group 2 could only bedistinguished as deciduous premolars or molars, it isalso useful to look at a simplified distribution (Table 3).This increases the number of both dps and molarsidentified, although only to general class groupings. Thefar greater number of dps, evident in either table, but

ARTICLE IN PRESSL.A. Schepartz et al. / Quaternary International 126–128 (2005) 271–282278
especially in Table 3, suggests that the Dadong faunahas predominantly young stegodonts.
The differing tooth class profiles for Groups 1 and 2reflect two taphonomic biases that are associated withuse of fragmentary material. Different elements mayhave a greater possibility of being preserved in relativelycomplete form. Fragmentation may result in theanalytical absence of an element (Lyman, 1994), and ifa particular element is more likely to be fragmentedbeyond recognition than others, the counts of that classwill under-represent the original presence of the element.Since stegodont teeth generally fragment along valleys,teeth with larger numbers of crests are likely to berepresented by a greater number of fragments, and to bedisproportionately represented in the sample. If frag-mentary elements are not included in the analysis, thenthe bigger molars would be under-represented; if allelements are included, molars may be over-represented.At Dadong it appears that the small, dense dp2s have agreater possibility of being recovered in relativelycomplete form, and that teeth with greater numbers ofcrests are more likely to be recovered as fragments. Inthe Group 1 sample, 92% of the dp2s are complete,while only 40% of the dp3s, 33% of the dp4s, andnone of the molars are complete. The high proportionof complete dp2s may be contrasted to the small numberof dp2s identified in Group 2. Why are there fewfragmentary dp2s? With their thinner enamel (Table 1),dp2s might not survive as well once fragmentationoccurs. However, they also have a compact morphologywith fewer valleys that could be areas for fragmentation.These smaller, rounder teeth may have been more likelyto be expelled in an intact form from the mouth of livingstegodonts, or they may have been swallowed whole andpreserved in dung.
Fig. 10. Age of dental remains profile.
9. Determination of dental age and construction of a
dental age profile
Haynes (1991) advocates using evidence of toothdevelopment in living proboscideans to determine age infossil animals, citing the similar timing of epiphysealfusion and tooth eruption in living and fossil probosci-deans. Sixty-five years is a generally agreed upon upperage limit for Elephas and Loxodonta, and may also beused for Mammuthus and Mammut (Haynes, 1991).Here, we use these standards to model developmentalstages for Stegodon. Following Haynes, we assignspecimens to one of five 12-year age categories.
There are several factors complicating the construc-tion of a dental age profile for this sample. First, sincedeciduous teeth were shed in succession, the presence ofisolated teeth does not necessarily indicate death of theindividual (although the sample does contain someunworn teeth that could only come from carcasses). The
dental age profile, therefore, represents the develop-mental age of source individuals. Because some of therecord may be made up of teeth shed naturally (unlikely,but not impossible in this cave faunal assemblage), it isnot really an age at death profile. It is an age of dentalremains profile. Second, the lack of distinction amongmolars means specimens that potentially belong to olderage categories cannot be aged, and are absent in theprofile. Third, each specimen that cannot be refit orassociated with other teeth must be considered torepresent an individual. Lastly, we examine the sampleas a whole without spatial or temporal divisions. Thishas important ramifications in the interpretation of theage at death profile, but the use of an unstratified sampleprobably does not introduce additional bias to thisstudy because of time-averaging effects that are char-acteristic of faunal assemblages from prehistoric sites(Haynes, 1991).
If all the Dadong teeth identifiable by class areassigned to an age cohort in Haynes’s system, all buttwo fall into the first category (0–12 years). Thisyoungest cohort encompasses the ages when dp2s,dp3s, dp4s and some M1s are in use (Haynes, 1991;compiled from several proboscidean developmentalstudies). There are clearly some other adult individuals,represented by fragmentary molars and two fairlycomplete M3s, but it is not possible to assign them toa specific cohort of animals over 12 years of age.Hypothetically, these teeth could belong to any of theother age cohorts, as shown in Fig. 10. If all theunassigned molars were in the same cohort, the profilewould have one tall bar at 0–12, and a second, middleheight bar at either 13–24, 25–36, 37–48 or 49–60. Thiswould be the most extreme situation, creating a U-shaped profile. Even if this were the distribution atDadong, there would still be a predominance of sub-adults. More probably, the molars represent individualsof various older ages, and the 48 specimens would bedistributed among the adult cohorts. It is important toemphasize that the true number of larger molars, and

ARTICLE IN PRESSL.A. Schepartz et al. / Quaternary International 126–128 (2005) 271–282 279
therefore adults, might actually be less than 48 if theinclusion of highly fragmentable molars has inflatedtheir representation in the sample.
10. Mortality profiles
There are two basic mortality profiles that archae-ozoologists use to model population age structure:catastrophic and attritional. In a catastrophic profile,each successive age class contains fewer individuals.Such a profile represents a population’s age structure atthe time of death as the result of a non-selectivecatastrophic event. An attritional (normal or cemetery)profile is bimodal, with the youngest and oldest ageclasses containing the majority of individuals, whileintermediate classes are less well represented (Klein,1982; Lyman, 1994). Both profiles assume the popula-tions are stable and stationary, i.e. lacking multiplebirths and not exhibiting dramatic size fluctuations.
Haynes (1991; Fig. 6.4) refines this classification,recognizing four main types of age profiles in living andextinct proboscidean assemblages. In the Haynesclassification, Type A shows the greatest portion ofindividuals in the first category of subadults with eachsuccessive category represented by decreasing propor-tions. This pattern results from non-selective mortalityin stable populations. Type B is bimodal. Althoughsubadults predominate in this type as in Type A, thereare few prime age adults. This type of profile resultsfrom selective mortality affecting mixed herds. Type C isdominated by prime-age adults. This results fromselective mortality affecting males only (for example,killings by hunters interested in ivory), or from non-selective mortality affecting declining populations. TypeD is patternless, resulting from various causes, includinginsufficient sample size.
The Dadong sample, with its greater preservation ofdps, would resemble either Type A or B. Even if it werepossible to assign all the molars to age cohorts, asdiscussed above, the sample would still have a distribu-tion that has predominantly younger individuals. Thisassumes that no portion of the sample has beenpreferentially removed through excessive breakage orother taphonomic factors.
11. Accumulation, transport and evidence for human
activity
Several activities could produce the stegodont assem-blage at Panxian Dadong. The material may haveaccumulated through the natural loss of deciduousteeth, the natural death of animals, water transport,transport by porcupines, or predation by large carni-vores or humans. It is therefore important to examine
each of these factors in terms of what appears to be adifferential representation of younger stegodonts.
Accidental deaths of young stegodonts might natu-rally occur in karst environments where natural fissuretraps are formed through limestone dissolution. Thisappears to have been the case at Yanjinggou, which hasvery complete portions of predominantly young stego-donts. The inaccessible nature of the traps is thought tohave aided in the preservation of the fossils. The trapsseem also to have attracted many carnivores to thelocality, but there is no evidence for human activity(Colbert and Hooijer, 1953; Bakken, 1997). There mayhave been similar fissures in the Panxian Dadong regionduring the Middle Pleistocene. Dadong itself was alarge, high-ceilinged cavern by that time, based on thedating of a large stalacto-stalagmite that had formed inthe interior (Shen et al., 1997). Young stegodonts mayhave become trapped after wandering into the cave(J. Shoshani, per. comm. 2001), but this seems unlikelyduring the times when humans were active there. Thereis some question as to why stegodonts or otherproboscideans would naturally appear in a cave fauna.Haynes (1991) mentions dry cave sites in Utah contain-ing large amounts of Mammuthus columbi dung. Hesuggests the site was used seasonally over a long timespan, but he does not detail why mammoths would haveused caves. With abundant drainage systems on theGuizhou Plateau and in the Dadong valley, stegodontswould probably not have had to venture into the cave toobtain water, but it might have been a source of mineralsalts.
The Dadong stegodont dental specimens show a greatvariety of breakage and damage that suggests they weresubject to destructive or natural transport processes.The enamel of some specimens appears opaque andpartially dissolved, while other crowns have a degradedrough and opaque surface. These changes may reflectdiagenic processes as well as physical destruction. Lessthan 6% of the sample exhibits weathering or surfaceerosion that could result from exposure prior to burial.The low frequency of polish and rounding that can beattributed to water action does not provide support forthe idea that water transport was particularly importantin Dadong. Rounding is visible on crown interiors androots of approximately 8% of the teeth, and on two ofthe stegodont bones. Overall, most of the stegodontsample was not substantially altered by natural pro-cesses, suggesting these activities were not importantfactors in the formation of the assemblage. This is alsotrue for the total faunal sample (c.f. Schepartz et al.,2003).
Rodents often collect faunal elements that they gnawto maintain their incisor functioning. Porcupines andsmall rodents were active in Dadong, but their damageto bones and tooth roots is fairly limited, affectingapproximately 5% of the total sample (Schepartz et al.,

ARTICLE IN PRESSL.A. Schepartz et al. / Quaternary International 126–128 (2005) 271–282280
2003). Only 3% of the stegodont sample exhibitsgnawing.
Big and small cats, such as Chinese wild cats andtigers, hyenas, foxes and other small carnivores werepresent in Dadong. These species are rare (4% of thesample identifiable to taxon), and there are fewthat would have preyed upon animals the size ofstegodonts. Scavenging carnivores, such as hyenas,could have transported portions of younger and smallerstegodonts into the cave. This might have beenparticularly true in the case of elements that are left atkill sites, such as crania or lower limbs (c.f. Blumen-shine, 1986; Lyman, 1994). Hence, scavenging mightaccount for the high proportion of stegodont teeth in theassemblage, if scavengers were able to detach crania ormandibles. The numbers of foot elements might also beexplained.
There is limited evidence for carnivore damage on theDadong bones (although this probably under-representsthe possible extent of carnivore activity in the generationof bone assemblages (c.f. Lyman, 1994)). The possiblerole of carnivores in the generation of the sample canalso be evaluated by examining the ratio of carnivores toungulates and herbivores. Carnivore-generated assem-blages, such as den sites, ordinarily contain highfrequencies of carnivore bone (Brain, 1981). TheDadong faunal sample falls outside the range of valuesobserved for carnivore-generated den deposits (at 4.6%,or 55 carnivores to 1176 herbivores and ungulates, of theelements identified to taxon). The scarcity of carnivoresat Dadong is notable, as it is a cave with side chambersand crevices suitable for denning. It could be that thecurrent faunal sample represents an area of the cave thatwas not extensively used by the carnivores, or thatcarnivore presence in the cave alternated with humanactivity phases.
Although the above factors may have contributed tothe formation of the Dadong stegodont assemblage, wesuggest that none of them were the single or primarysource of accumulation. As the stegodont material isfound in association with stone tools and hominidfossils, humans probably played some role in theiraccumulation. No evidence of human processing hasbeen found on stegodont bones. However, some of theother bones have stone tool cutmarks, impact fractures,and burning that we attribute to hominid activities.Burning is the most common alteration, occurring on3% of the bones. Although it is impossible to rule outother explanations for the impact fractures and burning,the co-occurrence of these features with cutmarks (andthe lack of their co-occurrence with carnivore damage)strengthens our interpretation of them as the result ofhominid behavior.
Evaluating the hominid role is complicated, as theycould have acted as hunters or scavengers. The large sizeof proboscideans means that any processing of their
carcasses might entail different behaviors. Recenthunters spend considerable time and energy usingproboscidean resources at the kill site. They consumeon-site, or smoke and dry meat to decrease its carryingweight (Fisher, 1993). If the Dadong stegodonts died orwere killed away from the cave, which is highlyprobable, the problem of transport could result insimilar signatures for human hunting, human scaven-ging, and carnivore scavenging.
The age structure of the stegodont sample, with itsindications of possible selective mortality of youngstegodonts, provides additional information pertainingto the possible behavior of hominids at Dadong. Klein(1982), in a classic study of hunting vs. scavenging ofPelorovis (giant buffalo) at two archaeological sites inSouth Africa, suggested that a relatively high proportionof very young individuals in an age profile indicatesactive human hunting. A profile that is deficient in veryyoung individuals may be produced by the selectiveremoval of their less durable bones by destructiveagencies, such as carnivore feeding, before they areincorporated into a faunal assemblage. Conversely,scavenging could account for an abundance of veryyoung individuals if the hominids located carcassesbefore other predators used them, a circumstance thatKlein considered to be unlikely. If hominids are themajor agents in the formation of the stegodont sampleat Dadong, and if Klein’s (1982) arguments are appliedto the Dadong stegodont assemblage, the role ofhominids as either hunters or scavengers of youngstegodonts cannot be ruled out.
If Haynes’s (1991) interpretations of proboscideanmortality profiles are applied to the Dadong data, theneither Type A or B profiles could be accommodated bythe evidence. A Type A profile would have resulted fromnon-selective mortality in a stable stegodont population;in a Type B profile, the predominance of subadultswould represent selective killing or scavenging. It istempting to suggest that the Type B profile is a better‘fit’ with the full suite of Dadong data that incorporatesthe information discussed above on natural and non-human accumulation factors and the archaeologicalassociations of stone tools with stegodont remains.However, a Type A profile is to be expected in a time-averaged sample like the one being analyzed. Therefore,the assumption that there is no human role in theaccumulation of the stegodont remains is not necessarilydisproved by these data.
Other Middle Pleistocene archaeological sites, suchas Ehringsdorf (Taubach-Ehringsdorf and Weimar-Ehringsdorf), Mauer (the Palaeoloxodon sample) andLa Cotte de St. Brelade, are characterized by subadultdominated proboscidean assemblages (see discussionand references in Haynes, 1991). La Cotte de St. Breladein the Jersey Islands is a cave site with abundantevidence for human/proboscidean interactions that

ARTICLE IN PRESSL.A. Schepartz et al. / Quaternary International 126–128 (2005) 271–282 281
presents an interesting comparison with Dadong. Scott(1989) analyzed the remains of over 20 mammoths and 5rhinoceroses preserved within two distinct layers. Themost substantial portions of the skeleton, such as theskull, tusks and denser portions of the scapula andpelvis, were best represented. In contrast to the situationat Dadong, only three isolated teeth were found. Basedon the stacked and articulated orientation of theremains, Scott (1989) proposed that the animals weredriven over sheer cliffs surrounding the site. Animalseither fell directly into the site, or were transported intothe cave from the cliff base or nearby chasms.Interestingly, less weighty but meat-bearing portions ofthe skeleton (the lower limbs, feet and mandibles) areunder-represented, and might have been removed byhominids or carnivores. These are also the better-represented elements for the Dadong stegodonts. If thedata from La Cotte are analyzed according to thedefined layers, there are differences is the relativerepresentation of the different age cohorts, but olderadults are absent from all of them. This may representselective mortality or non-selective death in a veryyoung population. If the total sample is used toconstruct the age profile, the result is a typical time-averaged profile (Haynes, 1991).
12. Conclusions
Analysis of the Panxian Dadong stegodont dental andskeletal material illustrates that the sample consistsprimarily of younger individuals aged 0–12 years. Acombination of tooth measures and morphologicalcharacteristics was used to identify fragmentary speci-mens and to increase the analytical sample. Continuedresearch on the application of metric data for identifica-tion of single crests and small fragments is needed beforefragmentary molars can be confidently assigned to toothclass. As teeth with more crests are more likely tofragment and be disproportionately represented, it maynot be appropriate to compare the entire sample withother studies that do not take fragmentary material intoaccount.
The sample is likely to have accumulated as the resultof more than one collection agent, and to have beensubsequently subjected to destructive forces duringformation of the faunal assemblage. Based on theavailable archaeological, faunal and taphonomic data,the stegodont faunal assemblage at Dadong is probablya result of hominid activities (either hunting orscavenging) and the natural accumulation of youngstegodonts from accidental deaths. This study supportsthe results of other taphonomic work (Schepartz et al.,2003) that suggests humans played an important role inthe formation of the Dadong faunal assemblage.
Acknowledgements
This work was supported by the National ScienceFoundation, the L.S.B. Leakey Foundation, theWenner-Gren Foundation for Anthropological Re-search, the Henry Luce Foundation, the University ofCincinnati Research Council and the Charles P. TaftFund, and California State University, Stanislaus. Wethank Sari Miller-Antonio, Huang Weiwen, HouYamei, Wang Wei, Si Xinqiang and Liu Jun for theirsuggestions and contributions to the fieldwork atDadong.
References
Bakken, D, 1997. Taphonomic and faunal parameters of pleistocene
hominid sites in China. Indo Pacific Prehistory Association Bulletin
16 (3), 13–26.
Blumenshine, R.J., 1986. Carcass consumption sequences and the
archaeological distinction of scavenging and hunting. Journal of
Human Evolution 15, 639–659.
Brain, C.K., 1981. The Hunters or the Hunted? University of Chicago
Press, Chicago.
Colbert, E.H., Hooijer, D.A., 1953. Pleistocene mammals from the
limestone fissures of Szechwan, China. Bulletin of the American
Museum of Natural History 101, 1–135.
Dong, W., Jin, C.Z., Xu, Q.Q., Liu, J.Y., Tong, H.W., Zheng, L.T.,
2000. A comparative analysis on the mammalian faunas associated
with Homo erectus in China. Acta Anthropologica Sinica 19
(supplement), 246–256.
Fisher, Jr., J.W., 1993. Foragers and farmers: material expressions of
interaction at elephant processing sites in the ituri forest, zaire. In:
Hudson, J. (Ed.), From Bones to Behavior: Ethnoarchaeological
and Experimental Contributions to the Interpretation of Faunal
Remains. Center for Archaeological Investigations, Occasional
Paper No. 21, Carbondale, IL, pp. 247–262.
Han, D.F., Xu, C.H., 1989. Quaternary mammalian faunas and
environment of fossil humans in south China. In: Wu, R.K., Wu,
X.Z., Zhang, S.S. (Eds.), Early Humankind in China. Science
Press, Beijing, pp. 338–391.
Haynes, G., 1991. Mammoths, Mastodonts, and Elephants. Cam-
bridge University Press, Cambridge.
Huang, W.W., Si, X.Q., Hou, Y.M., Miller-Antonio, S., Schepartz,
L.A., 1995. Excavations at Panxian Dadong, Guizhou province,
southern China. Current Anthropology 36 (5), 844–846.
Klein, R.G., 1982. Age (mortality) profiles as a means of distinguishing
hunted species from scavenged ones in Stone Age archaeological
sites. Paleobiology 8, 151–158.
Li, Y.X., Wen, B.H., 1986. Guanyindong: a Lower Paleolithic Site at
Qianxi County, Guizhou Province. Cultural Relics Publishing
House, Beijing (in Chinese).
Lyman, R.L., 1994. Vertebrate Taphonomy. Cambridge Manuals in
Archaeology. Cambridge University Press, Cambridge.
Miller-Antonio, S., Schepartz, L.A., Bakken, D., 2000. Raw material
selection and evidence for rhinoceros tooth tools at Dadong Cave,
southern China. Antiquity 74, 372–379.
Miller-Antonio, S., Schepartz, L.A., Karkanas, P., Hou, Y., Huang,
W., Bekken, D. 2004. Lithic raw material use at Dadong. Asian
Perspectives, 43 (2).
Movius, H.L., 1944. Early man and Pleistocene stratigraphy in
southern and eastern Asia. Papers of the Peabody Museum,
Harvard University, XIX(3).

ARTICLE IN PRESSL.A. Schepartz et al. / Quaternary International 126–128 (2005) 271–282282
Mu, X.A., Xu, H.K., Mu, D.C., Zhong, S.L., Xu, Q.Q., Zhang, H.,
Zhang, Y.Y., 1993. Discovery of Homo erectus remains from
Tangshan, Nanjing and its significance. Acta Anthropologica
Sinica 32 (4), 393–398 (in Chinese with English abstract).
Osborne, H.F., 1942. Proboscidea, A Monograph of the Discovery,
Evolution, Migration and Extinction of the Mastodonts and
Elephants of the World, Vol. II Stegodontoidea Elephantoidea.
The American Museum Press, New York.
Pan, Y.R., Yuan, C.W., 1997. Pleistocene primates from Panxian
Dadong, Guizhou Province. Acta Anthropologica Sinica 16 (3),
201–208 (in Chinese with English abstract).
Rink, W.J., Schepartz, L.A., Miller-Antonio, S., Huang, W., Hou, Y.,
Bakken, D., Richter, D., and Jones, H.L., 2003. Electron spin
resonance (ESR) dating of tooth enamel at Panxian Dadong cave,
Guizhou, China. In: Shen, C., Keates, S.G. (Eds.), Current
Research in Chinese Pleistocene Archaeology. BAR Monograph
Series, Oxford, pp. 95–106.
Saegusa, H., 1996. Stegodontidae: evolutionary relationships. In:
Shoshani, J., Tassy, P. (Eds.), The Proboscidea. Evolution and
Palaeoecology of Elephants and Their Relatives. Oxford University
Press, Oxford, pp. 178–190.
Schepartz, L.A., Bakken, D., Miller-Antonio, S., Paraso, K.,
Karkanas, P., 2003. Faunal approaches to site formation processes
at Panxian Dadong. In: Shen, C., Keates, S.G. (Eds.), Current
Research in Chinese Pleistocene Archaeology. BAR Monograph
Series, Oxford, pp. 70–94.
Scott, K., 1989. Mammoth bones modified by humans: evidence from
La Cotte de St Brelade Jersey Channel Islands. In: Bonnichsen, R.,
Sorg, M.H. (Eds.), Bone Modification. Center for the Study of the
First Americans Institute for Quaternary Studies. Orono, ME,
pp. 335–346.
Shen, G.J., Liu, J., Jin, L.H., 1997. Preliminary results on U-series
dating of Panxian Dadong in S-W China’s Guizhou province. Acta
Anthropologica Sinica 16 (3), 1–8 (in Chinese with English
abstract).
Takahashi, K., Namatsu, K., 2000. Origin of the Japanese Proboscidea
in the Plio-Pleistocene. Earth Science 54, 257–267.
Tassy, P., 1996. Dental homologies and nomenclature in the
Proboscidea. In: Shoshani, J., Tassy, P. (Eds.), The Proboscidea.
Evolution and Palaeoecology of Elephants and Their Relatives.
Oxford University Press, Oxford, pp. 21–26.
Zhang, Z.H., Liu, J., Zhang, H.G., Yuan, C.W., 1997. A Pleistocene
mammalian fauna from Panxian Dadong, Guizhou Province. Acta
Anthropologica Sinica 16 (3), 207–220 (in Chinese with English
abstract).
Zong, G., 1995. On new material of Stegodon with recollections of the
classification of Stegodon in China. Vertebrata PalAsiatica 33,
216–230 (in Chinese with English abstract).
Copyright © 2022 FDOKUMEN




















