
Taphonomy of the middle Cambrian (Drumian) Ravens Throat River Lagerstätte, Rockslide Formation,...
20
Taphonomy of the middle Cambrian (Drumian) Ravens Throat River Lagerst € atte, Rockslide Formation, Mackenzie Mountains, Northwest Territories, Canada JULIEN KIMMIG AND BRIAN R. PRATT Kimmig J. & Pratt B.R. 2015: Taphonomy of the middle Cambrian (Drumian) Ravens Throat River Lagerst€ atte, Rockslide Formation, Mackenzie Mountains, Northwest Territories, Canada. Lethaia, DOI: 10.1111/let.12135. The Middle Cambrian (series 3, Drumian, Bolaspidella Biozone) Ravens Throat River Lagerst€ atte in the Rockslide Formation of the Mackenzie Mountains, northwestern Canada, contains a Burgess Shale-type biota of similar age to the Wheeler and Marjum formations of Utah. The Rockslide Formation is a unit of deep-water, mixed carbon- ate and siliciclastic facies deposited in a slope setting on the present-day northwestern margin of Laurentia. At the fossil-bearing locality, the unit is about 175 m thick and the lower part onlaps a fault scarp cutting lower Cambrian sandstones. It consists of a succession of shale, laminated to thin-bedded lime mudstone, debris-flow breccias, minor calcareous sandstone, greenish-coloured calcareous mudstone and dolomitic siltstone, overlain by shallow-water dolostones of the Broken Skull Formation, which indicates an overall progradational sequence. Two ~1-m-thick units of greenish calcar- eous mudstone in the upper part exhibit soft-bodied preservation, yielding a biota dominated by bivalved arthropods and macrophytic algae, along with hyoliths and tri- lobites. It represents a low-diversity in situ community. Most of the fossils occur in the lower unit, and only the more robust components are preserved. Branching burrows are present under the carapaces of some arthropods, and common millimetre-sized disruptions of laminae are interpreted as bioturbation. The fossiliferous planar-lami- nated calcareous mudstone consists of chlorite, illite, quartz silt, calcite and dolomite and is an anomalous facies in the succession. It was deposited via hemipelagic fallout of a mixture of platform-derived and terrestrial mud. Geochemical analysis and trace- element proxies indicate oxic bottom waters that only occasionally might have become dysoxic. Productivity in the water column was dominated by cyanobacteria. Frag- ments of microbial mats are common as carbonaceous seams. Complete decay of soft tissues was interrupted due to the specific sediment composition, providing support for the role of clay minerals, possibly chlorite, in the taphonomic process. □ Burgess Shale, Cambrian, Lagerst€ atte, Mackenzie Mountains, taphonomy. Julien Kimmig [[email protected]], and Brian R. Pratt [[email protected]], Depart- ment of Geological Sciences, University of Saskatchewan, 114 Science Place Saskatoon, SK S7N 5E2, Canada; manuscript received on 26/09/2014; manuscript accepted on 23/02/ 2015. Burgess Shale-type (BST) deposits are exceptional windows that reveal early Palaeozoic biodiversity because they preserve soft-bodied components of the biota. These are seen as two-dimensional com- pressed fossils that may contain remnants of inte- rior structures. In addition to continued study of the taxonomy and morphology of these fossils (e.g. Briggs et al. 1994; Butterfield 1995; Edgecombe & Legg 2013), the environmental and taphonomic aspects remain controversial. Most analyses sup- port oxygenated bottom waters in BST deposits (Powell et al. 2003; McKirdy et al. 2011), but the preservation of soft tissue is usually ascribed to anaerobic conditions, at least within the sediments (e.g. Allison 1988; Sagemann et al. 1999). These seemingly opposing factors have yet to be ade- quately reconciled (Gaines et al. 2008; Butterfield 2012). Sedimentological studies confirm that BST environments are low-energy settings in relatively deep water (Gabbott et al. 2008; Webster et al. 2008; Brett et al. 2009; Gaines et al. 2011). Several mechanisms have been suggested to explain the presumably sharp redox boundary between the oxygenated water column and the sediment, including rapid burial, clay mineral precipitation, organic matter enrichment and microbial mats (Butterfield 1995; Butterfield et al. 2007; Hall et al. 2011; Gaines et al. 2012a; Garson et al. 2012). The details of water-column productivity and benthic microbial activity are difficult to determine by bio- marker analysis because the signals are lost in most BST deposits because of diagenetic alteration and possibly incipient metamorphism. However, the presence of benthic microbial mats has been inter- preted based on organic seams (i.e. Butterfield 1990; McKirdy et al. 2011). DOI 10.1111/let.12135 © 2015 Lethaia Foundation. Published by John Wiley & Sons Ltd
Transcript of Taphonomy of the middle Cambrian (Drumian) Ravens Throat River Lagerstätte, Rockslide Formation,...

Taphonomy of the middle Cambrian (Drumian) RavensThroat River Lagerst€atte, Rockslide Formation, MackenzieMountains, Northwest Territories, Canada
JULIEN KIMMIG AND BRIAN R. PRATT
Kimmig J. & Pratt B.R. 2015: Taphonomy of the middle Cambrian (Drumian) RavensThroat River Lagerst€atte, Rockslide Formation, Mackenzie Mountains, NorthwestTerritories, Canada. Lethaia, DOI: 10.1111/let.12135.
The Middle Cambrian (series 3, Drumian, Bolaspidella Biozone) Ravens Throat RiverLagerst€atte in the Rockslide Formation of the Mackenzie Mountains, northwesternCanada, contains a Burgess Shale-type biota of similar age to the Wheeler and Marjumformations of Utah. The Rockslide Formation is a unit of deep-water, mixed carbon-ate and siliciclastic facies deposited in a slope setting on the present-day northwesternmargin of Laurentia. At the fossil-bearing locality, the unit is about 175 m thick andthe lower part onlaps a fault scarp cutting lower Cambrian sandstones. It consists of asuccession of shale, laminated to thin-bedded lime mudstone, debris-flow breccias,minor calcareous sandstone, greenish-coloured calcareous mudstone and dolomiticsiltstone, overlain by shallow-water dolostones of the Broken Skull Formation, whichindicates an overall progradational sequence. Two ~1-m-thick units of greenish calcar-eous mudstone in the upper part exhibit soft-bodied preservation, yielding a biotadominated by bivalved arthropods and macrophytic algae, along with hyoliths and tri-lobites. It represents a low-diversity in situ community. Most of the fossils occur in thelower unit, and only the more robust components are preserved. Branching burrowsare present under the carapaces of some arthropods, and common millimetre-sizeddisruptions of laminae are interpreted as bioturbation. The fossiliferous planar-lami-nated calcareous mudstone consists of chlorite, illite, quartz silt, calcite and dolomiteand is an anomalous facies in the succession. It was deposited via hemipelagic falloutof a mixture of platform-derived and terrestrial mud. Geochemical analysis and trace-element proxies indicate oxic bottom waters that only occasionally might have becomedysoxic. Productivity in the water column was dominated by cyanobacteria. Frag-ments of microbial mats are common as carbonaceous seams. Complete decay of softtissues was interrupted due to the specific sediment composition, providing supportfor the role of clay minerals, possibly chlorite, in the taphonomic process. □ BurgessShale, Cambrian, Lagerst€atte, Mackenzie Mountains, taphonomy.
Julien Kimmig [[email protected]], and Brian R. Pratt [[email protected]], Depart-ment of Geological Sciences, University of Saskatchewan, 114 Science Place Saskatoon, SKS7N 5E2, Canada; manuscript received on 26/09/2014; manuscript accepted on 23/02/2015.
Burgess Shale-type (BST) deposits are exceptionalwindows that reveal early Palaeozoic biodiversitybecause they preserve soft-bodied components ofthe biota. These are seen as two-dimensional com-pressed fossils that may contain remnants of inte-rior structures. In addition to continued study ofthe taxonomy and morphology of these fossils (e.g.Briggs et al. 1994; Butterfield 1995; Edgecombe &Legg 2013), the environmental and taphonomicaspects remain controversial. Most analyses sup-port oxygenated bottom waters in BST deposits(Powell et al. 2003; McKirdy et al. 2011), but thepreservation of soft tissue is usually ascribed toanaerobic conditions, at least within the sediments(e.g. Allison 1988; Sagemann et al. 1999). Theseseemingly opposing factors have yet to be ade-quately reconciled (Gaines et al. 2008; Butterfield2012). Sedimentological studies confirm that BST
environments are low-energy settings in relativelydeep water (Gabbott et al. 2008; Webster et al.2008; Brett et al. 2009; Gaines et al. 2011). Severalmechanisms have been suggested to explain thepresumably sharp redox boundary between theoxygenated water column and the sediment,including rapid burial, clay mineral precipitation,organic matter enrichment and microbial mats(Butterfield 1995; Butterfield et al. 2007; Hall et al.2011; Gaines et al. 2012a; Garson et al. 2012). Thedetails of water-column productivity and benthicmicrobial activity are difficult to determine by bio-marker analysis because the signals are lost in mostBST deposits because of diagenetic alteration andpossibly incipient metamorphism. However, thepresence of benthic microbial mats has been inter-preted based on organic seams (i.e. Butterfield1990; McKirdy et al. 2011).
DOI 10.1111/let.12135 © 2015 Lethaia Foundation. Published by John Wiley & Sons Ltd

The Ravens Throat River Lagerst€atte of Drumi-an (Middle Cambrian) age (Kimmig & Pratt2015) offers an opportunity to examine the envi-ronmental and taphonomic factors relating to BSTpreservation, or the lack of preservation, in a dee-per water slope or ramp setting. Occurring innon-metamorphosed strata, this is a low-diversitybiota comparable to the coeval Wheeler and Mar-jum formations of Utah. Besides analysis of thesedimentary facies in the field, a variety of labora-tory methods were employed, including opticalpetrography, X-ray diffractometry, backscatteredscanning electron microscopy and synchrotronX-ray fluorescence, along with determination oftrace-element composition, total organic carbon(TOC) content, biomarkers and organic carbonstable isotopes. The aim of this paper was to pres-ent a description of the fossil-bearing units andco-occurring facies and an interpretation of thedepositional environment especially as it pertainsto BST preservation.
Geological setting
The Rockslide Formation is a regionally extensive,mixed carbonate–siliciclastic unit of Middle Cam-brian (series 3, Plagiura–Poliella to late Bolaspidellaor Lejopyge laevigata Biozone) age in the MackenzieMountains of northwestern Canada (Fig. 1; Gabri-else et al. 1973; Fritz 1979; Pratt 1989). Thicknessescan exceed 500 m, and the rocks consist of fossilifer-ous argillaceous and silty lime mudstone, calcareoussiltstone and carbonate debris-flow breccias or con-glomerates. Previous investigations indicate deposi-tion in relatively deep water on a west-facing slopeflanking the Selwyn Basin that appears to have beenseaward of a mixed carbonate–siliciclastic platformon or rimming the Mackenzie Arch which has sincebeen planed off (Pyle et al. 2007). Mudmound reefsare locally present (Pratt 1989). BST fossils areknown only from the location named Ravens ThroatRiver Lagerst€atte (Kimmig & Pratt 2015). On the
A B
C
Fig. 1. Ravens Throat River Lagerst€atte. A, location north of the South Nahanni River (asterisk; 63°100N, 127°550W), Mackenzie Moun-tains, Northwest Territories, Canada. B, topographic map of Lagerst€atte area. C, panorama of the outcrop area near the headwaters ofthe Ravens Throat River looking west. Samples for geochemical analysis were collected at the north end of the cliff (arrow).
2 J. Kimmig & B. R. Pratt LETHAIA 10.1111/let.12135

east side of the Mackenzie Arch and in the Macken-zie River valley, the lower Rockslide Formation cor-relates with intrabasinal shales of the Mount CapFormation (Dixon & Stasiuk 1998; MacNaughtonet al. 2013). These shales yield rare examples of BSTpreservation (Butterfield 1994; Butterfield & Nicho-las 1996). To the north, the Rockslide Formationgrades into a thick succession of deep-water sedi-ments belonging to the Hess River Formation in theMisty Creek embayment (Cecile 1982; Chevrier &Turner 2013). To the west, the Rockslide Formationis overlain by deep-water limestones of the Rabbit-kettle Formation (Pratt 1992; Gordey & Anderson1993).
The Lagerst€atte is located near the headwatersof the Ravens Throat River (127°550W, 63°100N).About 175 m of the lower Rockslide Formationare exposed on a mountainside above anunnamed lake. It consists of a succession of siltyshale that grades upwards into interbedded shaleand lime mudstone, and then into thin-beddedlime mudstone (Fig. 2). The Rockslide Formationoverlies a thin limestone and dolomite unitbelonging to the lower Cambrian Sekwi Forma-tion, which itself rests abruptly on shallow-marinesandstones of the Ediacaran to lower CambrianBackbone Ranges Formation. The base of theRockslide Formation yields trilobites probablybelonging to the Albertella Biozone (Pratt, B.R.personal observation). About 750 m to the south,the upper Rockslide Formation continues withabout 50 m of thin-bedded dolomitic limestonesoverlain by 50 m of medium-bedded dolostone.Shallow-water dolostone belonging to the BrokenSkull Formation overlies the Rockslide Formationat this location (section 19 of Gabrielse et al.1973). The lower 50 m of the Broken Skull For-mation consist of dolomitic sandstone and sandydolomite that more recent stratigraphic studies inthe region would assign to the Avalanche Forma-tion (Gordey & Anderson 1993).
The Sekwi Formation and lower 60 m of theRockslide Formation at the Ravens Throat Riverlocality are in fault contact against the upper Back-bone Ranges Formation. This is the southern marginof a syndepositional fault scarp that can be tracednorthwards across the valley to the next ridge. Inplanar view, the fault appears to be broadly U-shaped; the fault plane was relatively gently inclinedbut steepened markedly at its southern margin. Achange in dip between the Sekwi Formation–lower-most Rockslide Formation interval (140°/56°W) andthe rest of the Rockslide Formation (125°/64°W)implies a second phase of movement. The fault islikely listric in nature and probably a local feature,
less steep than the high-angle Cathedral and Eldonescarpments of the southern Rocky Mountains (Col-lom et al. 2009; Johnston et al. 2009b).
Fig. 2. Generalized stratigraphy of the Rockslide Formation inthe Ravens Throat river area. Grey-coloured intervals containBurgess Shale-type fossils, mostly in the lower one.
LETHAIA 10.1111/let.12135 Middle Cambrian taphonomy 3

Two ~1–m-thick units of distinctive greenish limemudstone exhibit soft-bodied preservation in inter-vals 134–139 and 162–166 m above the Sekwi For-mation. The lower unit, at +135 m, contains almostall the fossils, whereas the upper one, at about+164 m, contains only rare fragments of Margaretiadorus. Trilobites and agnostoid arthropods indicatethat the fossil-bearing interval belongs to theBolaspidella Biozone and is therefore of similar ageas the Wheeler and Marjum formations of Utah. TheBurgess Shale by contrast is slightly older (Bathyuris-cus–Elrathina Biozone), but shares some taxa incommon with the Ravens Throat River Lagerst€atte(Kimmig & Pratt 2015). Because the outcrop is asteep cliff, most fossil specimens were collected fromthe scree.
Methodology
The fossil specimens are housed in the collections ofthe Royal Tyrrell Museum (TMP) in Drumheller,Alberta.
A suite of 55 polished thin sections of the variousRockslide lithofacies and the lowest 39 ‘beds’ of thelower fossil-bearing unit were examined with a Ni-kon Optiphot-Pol microscope for grain size, sedi-mentary structures and organic residues. Mineralogywas determined using a JEOL JXA-8600 Superprobewith backscattered scanning electron microscope.Mineralogical composition was determined by X-raydiffractometer (XRD) analysis of whole-rock powderusing a PANalytical Powder XRD operating at 40 kVand 45 mA with monochromated Co Kalpha radia-tion (100.9650 secs/0.0170° step).
Samples for trace-element analysis were powderedusing a Siebtechnik disc mill, then stored in 50-mlcentrifuge tubes. The samples were analysed by theSaskatchewan Research Council, Saskatoon, usingPerkinElmer Optima 4300DV or Optima 5300DVICP-MS instruments. An aliquot of pulp wasdigested to dryness in a hot block digestion systemusing a mixture of ultra-pure concentrated acids HF,HNO3 and HClO4. The residue was dissolved andmade up to volume using deionized water prior toanalysis. Total digestion and analysis were per-formed on samples to determine the dilutionrequired prior to analysis.
Enrichment factors (EF) were determined by nor-malizing selected trace elements to Al, which repre-sents a proxy for the detrital fraction, and comparingthose ratios to ‘average shale’. The EF is equal to (Ele-ment/Al)sample/(Element/Al)shale, where the ratio inthe numerator is that for the shale in question, and
the ratio in the denominator is that for a ‘typical’shale (Tribovillard et al. 2006; using data from Ture-kian & Wedepohl 1961; Wedepohl 1971).
Total organic carbon (TOC) and d13C analyses ofcarbonate-bearing samples were measured on sam-ples that were first acidified to remove carbonatematerial. The residue was then homogenized andloaded into tin capsules. Stable isotope values wereobtained using a Thermo Finnigan Flash 1112 EAcoupled to a Thermo Finnigan Delta Plus XLthrough a Conflo III in the Saskatchewan IsotopeLaboratory. Samples were dropped under heliuminto an oxidation furnace packed with chromium(VI) oxide and silvered cobaltic/cobaltous oxide (toremove any halogens) at 1000°C. Organic materialwas oxidized to CO2, various nitrogen-bearing gasesand water. This gas was then passed through areduction furnace packed with elemental copper at680 °C to reduce all nitrogen-bearing compounds topure gaseous nitrogen. The resulting gases were thenpassed through a water trap to eliminate moisture. AGC column at 50 °C then followed to unmix thecarbon dioxide and nitrogen gases for analysis in themass spectrometer. Data were blank-corrected. Car-bon isotope ratios were corrected for 17O contribu-tion using the Craig correction and reported in permil notation relative to the VPDB scale. Carbon datawere calibrated against the international standardsL-SVEC (d13C = �46.6& VPDB) and IAEA-CH6(d13C = �10.45& VPDB). IAEA-CH7, an interme-diate international standard, gave the followingresult during calibration of the in-house standards:d13C = �32.15 � 0.04& VPDB (n = 11), com-pared with the accepted value ofd13C = �32.15 � 0.10& VPDB. d13C values of car-bonate are reported with respect to VPDB with aprecision of �0.12& (n = 18). The %C measure-ments are reported by mass and have a precision of�10% of the reported percentage.
Observations
Lithofacies
The Rockslide Formation at the Ravens Throat Riverlocation is subdivided into five facies: (1) blackshale; (2) lime mudstone; (3) debris-flow breccia;(4) calcareous mudstone (containing the BST pres-ervation); and (5) dolomitic lime mudstone(Figs 3A–D, 4A–E). The Rockslide Formation at thislocation is thinner than at the type section to the east(section 19; Gabrielse et al. 1973) and is the onlylocation known where BST preservation occurs.
4 J. Kimmig & B. R. Pratt LETHAIA 10.1111/let.12135

Facies 1: Shale. – The base of the Rockslide Forma-tion consists of 20 m of thin-bedded, dark-grey toblack calcareous shale, which overlies the Sekwi For-mation abruptly. Planar laminae are variably rich insilt and rounded very fine sand (Fig. 4A). The shale(one sample) is composed of calcite (49%), quartz(22%), chlorite (clinochlore, 22%), and illite (7%)(Table 3). The basal beds are rich in disarticulatedtrilobite sclerites belonging to Ogygopsis klotzi andother ptychoparioid and eodiscoid trilobites, alongwith lingulate brachiopods.
The facies is interpreted to have been deposited ina deep-water environment, involving suspension set-tling of clay, silt and lime mud. The trilobite andbrachiopod fauna indicates sporadic benthic coloni-zation. However, the trace-element compositionindicates a dominance of anoxic bottom waters (see‘Geochemistry’ below).
Facies 2: Lime mudstone. – Thin- to medium-bed-ded, wavy and lenticular beds of silty lime mudstonecomprise the bulk of the Rockslide Formation. Theyconsist of intercalated layers of laminated silty mi-crite and uniform micrite without lamination or gra-dation (Fig. 4B,D). In places, the lime mudstone isinterlayered with dolomitic micrite laminae, produc-ing the ‘ribbon limestone’ fabric commonly encoun-
tered in Cambrian carbonate units (Fig. 3B).Trilobite and agnostoid sclerites and lingulate bra-chiopod valves occur rarely, mostly in a convex-upward orientation. A coquina is present 133 mabove the base, just below the lowest interval offacies 4. Lime mudstone (one sample from the mid-dle interval) is composed of calcite (73%) and quartz(17%), with minor dolomite (7%) and illite (3%);133 m above base, a bed containing neomorphicbladed calcite is composed of calcite (60%) andquartz (32%), with minor amounts of chlorite, illiteand pyrite (Table 3). The lime mudstone is variablyinterbedded with black shale between about 20 and40 m above base. Intervals of lime mudstone areseparated by a number of debris-flow breccia beds(facies 3).
This facies reflects deep-water conditions, involv-ing suspension fall-out of lime mud, silt and clay.Fossils represent sparse in situ populations thatestablished themselves from time to time; low-veloc-ity bottom currents oriented the sclerites into thestable position. Dysoxic to anoxic conditions areindicated by trace-element composition (see ‘Geo-chemistry’ below).
Facies 3: Debris-flow breccia. – Erosive-based, thick-bedded breccias range from 0.5 to 3 m thick and
A B
DC
Fig. 3. Photographs of the different lithofacies of the Rockslide Formation at the Ravens Throat River Lagerst€atte. A, lime mudstone(‘ribbon limestone’) at +110 m. B, debris-flow breccia at +73 m. C, lower portion of the calcareous mudstone unit at +135–137 m. D,calcareous mudstone with interbedded lime mudstone beds and lenses at +136 m. Scale bar is 10 cm for A, B and D.
LETHAIA 10.1111/let.12135 Middle Cambrian taphonomy 5

punctuate intervals of lime mudstone (facies 2).Clasts range from 1 mm to about 10 cm across andare subangular, tabular, equant or irregular in shape(Fig. 4C). Clasts include lime mudstone and sandypeloidal and oolitic grainstone. Both clast- andmatrix-supported fabrics are present; the matrix issilty and sandy lime mudstone or calcareous sand-stone with rare trilobite bioclasts.
These beds represent sporadic debris flows causedby collapse of beds of lime mud and weakly cemen-ted sediments further upslope. Because most clastsare lime mudstone, similar to the host intervals,transport distance was probably small, but the pres-ence of admixed sand in some beds suggestsconcomitant input from shallow-water areas. Theseevents might have been triggered by storms and/ortectonic activity.
Facies 4: Calcareous mudstone. – This facies isgreenish in colour and hosts the BST preservation. Itconsists of planar-laminated, variably silty claystonecontaining scattered pyrite crystals up to several mil-limetres across, and rare lenses up to 100 lm widecontaining small framboids. In thin section, mud-stone laminae are variably distinct, commonlygraded, and locally show deformation in the form oftiny folds or sheared domains a few millimetres insize; it is likely that some of these are millimetre-sized burrows (Fig. 5B,E; see below). Discontinuousplanar seams and flakes of organic matter are com-mon. Lenticular, very thin laminae and dispersedsilt-sized calcite crystals and angular silt grains arelocally present. In general, the laminae vary in theirproportion of clay, organic matter and calcite. Themudstone (six samples) is composed of quartz(~36%), illite (~26%) and chlorite (~24%), plus cal-cite and dolomite (~14%) and pyrite (Table 3). Int-erbedded are lighter grey-coloured laminae, thinbeds and concretionary lenses of lime mudstone(facies 2) containing less quartz (~25%) and morecarbonate minerals (~50%). The calcite is locally mi-crospar from recrystallization of a micrite precursor.Large pyrite crystals commonly replace larger calcitespar crystals.
Ptychoparioid trilobites and agnostoids are pres-ent and preserved in three dimensions, whereas thesoft-bodied fossils show little to no relief, and onlythe more robust parts are preserved (Kimmig &Pratt 2015).
This facies is interbedded with facies 2 and wastherefore likely deposited under similar deep-waterconditions.
Facies 5: Dolomitic siltstone. – Overlying the BSTdeposits at the top of the exposed section is a >10-
A
B
D
E
C
Fig. 4. Thin-section photomicrographs of lithofacies in theRockslide Formation at the Ravens Throat River Lagerst€atte. A,shale at +7 m consisting of plane-laminated claystone erosivelyoverlain by indistinctly cross- and plane-laminated silty clay-stone. B, lime mudstone with planar lamination at +35 m. C,debris-flow breccia at +102 m consisting of angular granules ofpeloidal grainstone (pelsparite) in lime mudstone matrix withtrilobite sclerite (arrow). D, lime mudstone at +133 m, the baseof the Burgess Shale-type unit; diagenetic pseudospar–microspar(arrow) between the limestone and the mudstone. E, dolomiticlimestone at +190 m (top of cliff). Scale bars are 1 cm.
6 J. Kimmig & B. R. Pratt LETHAIA 10.1111/let.12135

m-thick unit of thin-bedded reddish-stained, buff-weathering, variably dolomitic argillaceous siltstoneand silty lime mudstone (Fig. 4E) composed (onesample) of quartz (35%), calcite (27%), dolomite
(15%), K-feldspar (10%), illite (8%), chlorite (5%)and local pyrite cubes. The beds show flaggy weath-ering with irregular surfaces preferentially dolomi-tized. The beds are internally non-laminated and
AB
D
E F
G
H
C
Fig. 5. Thin-section photomicrographs (A–G) and SEM photomicrograph (H) of the lower fossiliferous interval, Rockslide Formation atthe Ravens Throat River Lagerst€atte. A, calcareous mudstone with burrow (white arrow) and trilobite sclerite (black arrow) at +134.4 m.B, close-up of A with possible burrow. C, close-up of A with trilobite sclerite. D, lime mudstone interlayer at +134.7 m. E, calcareousmudstone with pyrite crystals and scattered microspar crystals (possible bioclasts; white arrows) at +134.9 m. F, close-up of E with possi-ble burrow. G, calcareous mudstone with burrow (white arrow) at +134.5 m. H, calcareous mudstone showing pyrite cube (upper right).Scale bars are 10 mm for A, D, E and G, 1 mm for B, C and F, and 100 lm for H.
LETHAIA 10.1111/let.12135 Middle Cambrian taphonomy 7

appear bioturbated, and contain scattered trilobitesclerites and lingulate brachiopod valves.
Fossil preservation
The biota in the lower fossiliferous calcareousmudstone consists of bivalved arthropods, trilo-bites, agnostoids, worms, hyoliths and macrophyticalgae (Fig. 6). The arthropods are the most diversecomponent of the biota, while most specimensrecovered belong to the hyolith Haplophrentis car-
inatus and the macrophytic alga M. dorus. Thebivalved arthropods, worms and algae are pre-served mostly as dark-grey to black carbonaceousfilms. Most bivalved arthropods are represented bysingle valves, but a few are preserved in a ‘butter-flied’ fashion. The margins of a minority of thevalves (12 specimens) are not smooth but, rather,are cuspate, which may be scavenging traces, whileothers are only partially preserved, suggesting par-tial decomposition, perhaps after scavenging(Fig. 7A–F). Most specimens are flattened (~85%),
A B
D
E
F G H
C
Fig. 6. Ravens Throat River Lagerst€atte fossils: A, Perspicaris? dilatus in lateral view (TMP 2013.101.0005). Scratches on valve surface arean artefact of damage from lying on the scree slope where the specimen was collected. B, branching specimen of Margaretia dorus (TMP2013.101.0132). C, Haplophrentis carinatus with conch, operculum and helens (TMP 2013.101.0058). D, arthropod indet in lateral view(TMP 2013.101.0035). E, Isoxys mackensiensis in lateral view (TMP 2013.101.0038). F, partial branch of Margaretia dorus (TMP2013.101. 0141). G, H. carinatus (arrow) on coprolite (TMP 2013.101.110). H, coiled specimen of a priapulid, genus and species indet.(TMP 2013.101.0051). Scale bars are 10 mm for A–F and 5 mm for G and H.
8 J. Kimmig & B. R. Pratt LETHAIA 10.1111/let.12135

which generated concentric wrinkles. Some hyo-liths are complete, but about 80% are missing thehelens and about 40% are represented only by theconch without the operculum. Most worms aredecomposed and cannot be identified to phylumlevel. Most trilobites and agnostoids are articulated
and most are uncompacted. Margaretia dorus thal-lus fragments in the upper fossiliferous calcareousmudstone at about +165 m, where they comprisemost of the biota, are preserved with some posi-tive relief showing the characteristic network oflozenge-shaped indentations.
A
B
D
E
FC
Fig. 7. Photographs of partially decomposed fossils of the Ravens Throat River Lagerst€atte. A, Perspicaris? dilatus, lateral view of valve(TMP 2013.101.0016). B, arthropod indet. fragment (TMP 2013.101.0020). C, Perspicaris? dilatus, lateral view of valve (TMP2013.101.0017). D, Perspicaris? dilatus, lateral view of valve (TMP 2013.101.0018). E, Perspicaris? dilatus, lateral view of valve (TMP2013.101.0023) F, Branchiocaris? sp., lateral views of two disarticulated valves (TMP 2013.101.0021). Scale bars are 5 mm.
LETHAIA 10.1111/let.12135 Middle Cambrian taphonomy 9

Ichnology
The lime mudstone facies lacks burrows and otherevidence of bioturbation. The dolomitic siltstoneoverlying the Lagerst€atte-bearing interval containsfine sand- and silt-filled simple burrows 2 mm indiameter that meander horizontally and vertically.They are assigned to Palaeophycus.
The fossiliferous calcareous mudstone yields onlya few larger feeding and burrowing traces, includingvery rare Rhizocorallium-like burrows about 3 cmlong and 1.5 cm wide (Fig. 8D). These burrows areabout 0.5 mm wide, and the Spreiten are marked byfine arcuate organic lines.
Under two of the more than 50 non-biominer-alized arthropod valves collected are groove-shaped, simple burrows between 0.5 and 1 mmwide and several centimetres long (Fig. 8A,B).Apparent bifurcation is likely due to intersectionrather than true branching. They resemble thestructures described from below arthropod valves
in other BST deposits (Zhang et al. 2007;M�angano 2011; M�angano et al. 2012), but theyare somewhat wider and longer. The grooveshape indicates that the burrows remained openand the carapace was compacted and deformedinwards.
Laminae deformed at the millimetre scale arefairly common in the calcareous mudstone. Someexamples are elliptical to lenticular in cross-section,while others show domains where the laminae areoverthrust and pulled apart (Fig. 5F). Most appearto be from bioturbation, but some may representsmall soft-sediment deformation features triggeredby seismic activity under shallow burial before sig-nificant compaction.
Geochemistry
Redox-sensitive trace elements. – Aluminium fallsbetween 5% and 9% and is similar to ‘average shale’composite standards (Turekian & Wedepohl 1961).
A
B
D
C
Fig. 8. Trace fossils of the Ravens Throat River Lagerst€atte: A, Perspicaris? dilatus, lateral view of valve with apparent branching burrowbeneath, preserved as negative relief (latex cast dusted with ammonium chloride sublimate; TMP 2013.101.0002). B, close-up of A. C,Perspicaris? dilatus, lateral view of valve with burrows beneath (arrow) (TMP 2013.101.0033). D, Rhizocorallium? sp. (TMP2013.101.0303). Scale bar is 10 mm.
10 J. Kimmig & B. R. Pratt LETHAIA 10.1111/let.12135

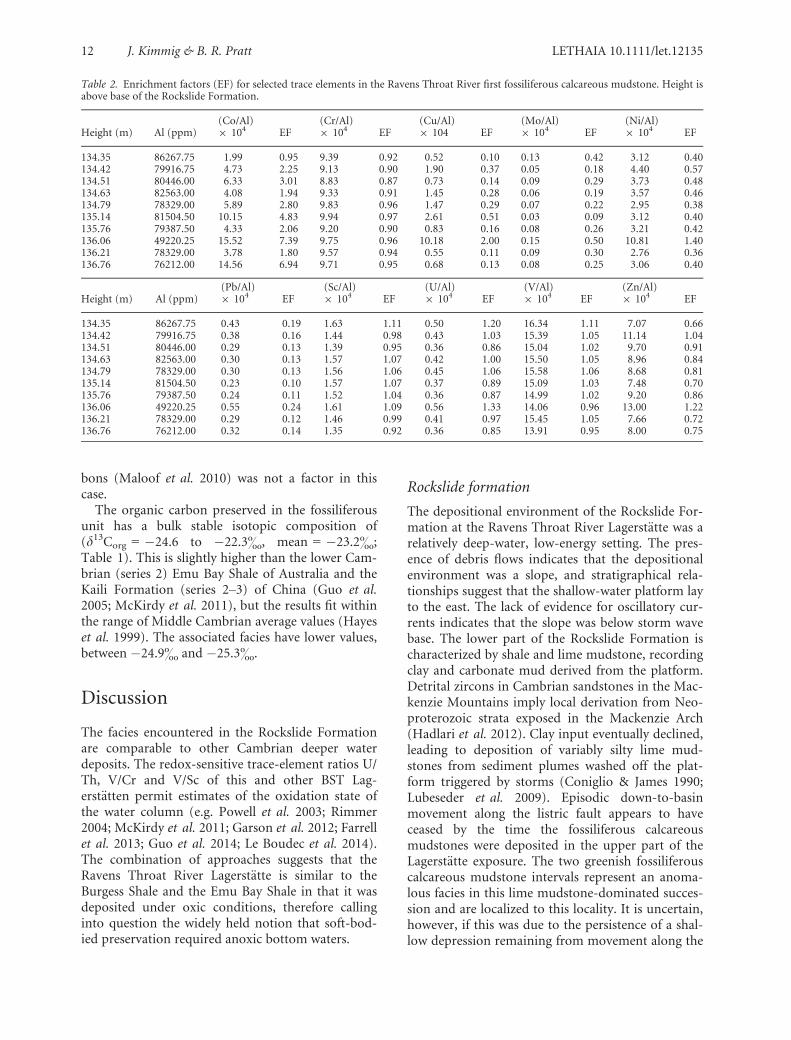
The trace-element data for the fossil beds and theother lithofacies (Table 1), in the context of the EFto account for the detrital bias (Table 2), show vari-able enrichment of most elements, but neither of themajor redox-sensitive metals (U and V) shows a cor-relation with Al, suggesting that they are not ofdetrital origin. The shales of the calcareous mud-stone are slightly enriched in Co (average EF = 3.4),but two beds above the lower fossiliferous unit at+136.06 m and +136.76 m show higher enrichment(EF of 7.39 and 6.94, respectively). On the otherhand, the calcareous mudstone overall is variablydepleted in Mo (avg. EF = 0.27), Cu (avg.EF = 0.41), Ni (avg. EF = 0.53), Pb (avg. EF = 0.14)and Zn (avg. EF = 0.85). Cr, Sc, U and V are at‘average shale’ level (Wedepohl 1971). Uraniumenrichment generally happens during redox condi-tions typical of sulphate reduction in marine envi-ronments, while V enrichment occurs underconditions of nitrate reduction (Jones & Manning1994; Piper & Isaacs 1995; Maloof et al. 2010). Noobvious co-variation with organic matter is exhib-ited (Fig. 9), but this might be due to the narrowspread of the observed TOC values (McKirdy et al.2011). The bed at +136.06 m is enriched in all traceelements compared to other beds (Table 1). Theseenrichments are minor, but do not necessarilyindicate a dysoxic water column, because mosttrace elements are depleted compared with ‘averageshale’.
Carbon. – Dolomite is the dominant carbonate inthe fossiliferous unit. TOC values are low with amean of 0.16% (0.10–0.22%; Table 1).
TOC values are similar to the Burgess Shale(0.11–0.42%; Butterfield 1990; Powell et al. 2003;Powell 2009) and the age-equivalent Marjum For-mation (~0.14%; Smith 2007). The thermal alter-ation of the Ravens Throat River Lagerst€atte appearsto be lower than that of the Burgess Shale, suggestingthat the primary TOC value was around 1.5% (Pow-ell 2003; McKirdy et al. 2011). That TOC content istoo low to have accumulated under stable anoxic ordysoxic conditions, even after accounting for a pri-mary TOC level of 1.5% (e.g. Algeo & Maynard2004). Sedimentary organic matter is preferentiallypreserved in oxygen-depleted bottom waters (De-maison & Moore 1980), although increased organicmatter can also be related to enhanced productivityin the photic zone (Calvert 1987), and suitable sedi-mentation rates.
Samples from the fossiliferous unit show no cor-relation between d13Corg and TOC (Fig. 10) suggest-ing that porosity- and permeability-dependentcontamination associated with migrating hydrocar-T
able
1.Trace-elementandorganiccarbondataofselected
bedsoftheRockslideForm
ationattheRavensThroat
River
Lagerst€ atte.H
eightisabove
baseoftheRockslideForm
ation.Thesam-
plesbetween+134.35
and+136.76
marefrom
thelower
fossiliferousinterval,w
hiletheoneat+164m
isfrom
theupper
fossiliferousinterval.
Facies
Height
Totalorganic
carbon
d13C
org
ScV
UTh
Pb
Mo
Cu
Co
Ni
Zn
Cr
V/Cr
V/Sc
U/Th
m%
&(V
PDB)
ppm
ppm
ppm
ppm
ppm
ppm
ppm
ppm
ppm
ppm
ppm
Shale
50.54
�24.8
7.2
234
6.32
4.24
136.07
16.3
2233.6
141
445.32
32.50
1.49
Lim
emudstone
600.44
�24.8
2.8
169
2.03
2.05
5.69
1.85
17.2
18.1
19.8
3615
11.27
60.36
0.99
Debris-flowbreccia
790.26
�24.8
5.2
67.2
2.44
4.3
11.6
2.16
1034.8
18.5
7027
2.49
12.92
0.57
Lim
emudstone
133
0.09
�24.3
4.2
36.9
2.14
1.64
5.39
0.88
14.4
68.4
19.8
1911
3.35
8.79
1.30
Calcareousmudstone
134.35
0.15
�23.5
14.1
141
4.34
12.1
3.74
1.08
4.5
17.2
26.9
6181
1.74
10.00
0.36
Calcareousmudstone
134.42
0.22
�24.6
11.5
123
3.46
10.3
3.01
0.42
15.2
37.8
35.2
8973
1.68
10.70
0.34
Calcareousmudstone
134.51
0.13
�22.7
11.2
121
2.89
9.58
2.32
0.71
5.9
50.9
3078
711.70
10.80
0.30
Calcareousmudstone
134.63
0.15
�22.7
13128
3.46
10.8
2.5
0.47
1233.7
29.5
7477
1.66
9.85
0.32
Calcareousmudstone
134.79
0.15
�22.3
12.2
122
3.5
10.7
2.36
0.52
11.5
46.1
23.1
6877
1.58
10.00
0.33
Calcareousmudstone
135.14
0.14
�23.0
12.8
123
3.03
10.3
1.85
0.22
21.3
82.7
25.4
6181
1.52
9.61
0.29
Calcareousmudstone
135.76
0.15
�22.9
12.1
119
2.89
101.93
0.63
6.6
34.4
25.5
7373
1.63
9.83
0.29
Calcareousmudstone
136.06
0.10
�23.6
7.9
69.2
2.75
5.74
2.69
0.74
50.1
76.4
53.2
6448
8.76
0.48
1.44
Calcareousmudstone
136.21
0.20
�22.8
11.4
121
3.19
11.4
2.25
0.71
4.3
29.6
21.6
6075
10.61
0.28
1.61
Calcareousmudstone
136.76
0.19
�23.7
10.3
106
2.73
9.8
2.41
0.58
5.2
111
23.3
6174
10.29
0.28
1.43
Calcareousmudstone
164
0.24
�23.1
12.5
107
2.27
9.12
4.6
0.43
8.2
31.9
25.3
6462
8.56
0.25
1.73
Dolomiticsiltstone
190
0.21
�24.9
7.2
72.8
4.38
6.63
14.8
1.56
18.2
64.2
26.5
4340
1.82
10.11
0.66
LETHAIA 10.1111/let.12135 Middle Cambrian taphonomy 11
bons (Maloof et al. 2010) was not a factor in thiscase.
The organic carbon preserved in the fossiliferousunit has a bulk stable isotopic composition of(d13Corg = �24.6 to �22.3&, mean = �23.2&;Table 1). This is slightly higher than the lower Cam-brian (series 2) Emu Bay Shale of Australia and theKaili Formation (series 2–3) of China (Guo et al.2005; McKirdy et al. 2011), but the results fit withinthe range of Middle Cambrian average values (Hayeset al. 1999). The associated facies have lower values,between �24.9& and �25.3&.
Discussion
The facies encountered in the Rockslide Formationare comparable to other Cambrian deeper waterdeposits. The redox-sensitive trace-element ratios U/Th, V/Cr and V/Sc of this and other BST Lag-erst€atten permit estimates of the oxidation state ofthe water column (e.g. Powell et al. 2003; Rimmer2004; McKirdy et al. 2011; Garson et al. 2012; Farrellet al. 2013; Guo et al. 2014; Le Boudec et al. 2014).The combination of approaches suggests that theRavens Throat River Lagerst€atte is similar to theBurgess Shale and the Emu Bay Shale in that it wasdeposited under oxic conditions, therefore callinginto question the widely held notion that soft-bod-ied preservation required anoxic bottom waters.
Rockslide formation
The depositional environment of the Rockslide For-mation at the Ravens Throat River Lagerst€atte was arelatively deep-water, low-energy setting. The pres-ence of debris flows indicates that the depositionalenvironment was a slope, and stratigraphical rela-tionships suggest that the shallow-water platform layto the east. The lack of evidence for oscillatory cur-rents indicates that the slope was below storm wavebase. The lower part of the Rockslide Formation ischaracterized by shale and lime mudstone, recordingclay and carbonate mud derived from the platform.Detrital zircons in Cambrian sandstones in the Mac-kenzie Mountains imply local derivation from Neo-proterozoic strata exposed in the Mackenzie Arch(Hadlari et al. 2012). Clay input eventually declined,leading to deposition of variably silty lime mud-stones from sediment plumes washed off the plat-form triggered by storms (Coniglio & James 1990;Lubeseder et al. 2009). Episodic down-to-basinmovement along the listric fault appears to haveceased by the time the fossiliferous calcareousmudstones were deposited in the upper part of theLagerst€atte exposure. The two greenish fossiliferouscalcareous mudstone intervals represent an anoma-lous facies in this lime mudstone-dominated succes-sion and are localized to this locality. It is uncertain,however, if this was due to the persistence of a shal-low depression remaining from movement along the
Table 2. Enrichment factors (EF) for selected trace elements in the Ravens Throat River first fossiliferous calcareous mudstone. Height isabove base of the Rockslide Formation.
Height (m) Al (ppm)(Co/Al)9 104 EF
(Cr/Al)9 104 EF
(Cu/Al)9 104 EF
(Mo/Al)9 104 EF
(Ni/Al)9 104 EF
134.35 86267.75 1.99 0.95 9.39 0.92 0.52 0.10 0.13 0.42 3.12 0.40134.42 79916.75 4.73 2.25 9.13 0.90 1.90 0.37 0.05 0.18 4.40 0.57134.51 80446.00 6.33 3.01 8.83 0.87 0.73 0.14 0.09 0.29 3.73 0.48134.63 82563.00 4.08 1.94 9.33 0.91 1.45 0.28 0.06 0.19 3.57 0.46134.79 78329.00 5.89 2.80 9.83 0.96 1.47 0.29 0.07 0.22 2.95 0.38135.14 81504.50 10.15 4.83 9.94 0.97 2.61 0.51 0.03 0.09 3.12 0.40135.76 79387.50 4.33 2.06 9.20 0.90 0.83 0.16 0.08 0.26 3.21 0.42136.06 49220.25 15.52 7.39 9.75 0.96 10.18 2.00 0.15 0.50 10.81 1.40136.21 78329.00 3.78 1.80 9.57 0.94 0.55 0.11 0.09 0.30 2.76 0.36136.76 76212.00 14.56 6.94 9.71 0.95 0.68 0.13 0.08 0.25 3.06 0.40
Height (m) Al (ppm)(Pb/Al)9 104 EF
(Sc/Al)9 104 EF
(U/Al)9 104 EF
(V/Al)9 104 EF
(Zn/Al)9 104 EF
134.35 86267.75 0.43 0.19 1.63 1.11 0.50 1.20 16.34 1.11 7.07 0.66134.42 79916.75 0.38 0.16 1.44 0.98 0.43 1.03 15.39 1.05 11.14 1.04134.51 80446.00 0.29 0.13 1.39 0.95 0.36 0.86 15.04 1.02 9.70 0.91134.63 82563.00 0.30 0.13 1.57 1.07 0.42 1.00 15.50 1.05 8.96 0.84134.79 78329.00 0.30 0.13 1.56 1.06 0.45 1.06 15.58 1.06 8.68 0.81135.14 81504.50 0.23 0.10 1.57 1.07 0.37 0.89 15.09 1.03 7.48 0.70135.76 79387.50 0.24 0.11 1.52 1.04 0.36 0.87 14.99 1.02 9.20 0.86136.06 49220.25 0.55 0.24 1.61 1.09 0.56 1.33 14.06 0.96 13.00 1.22136.21 78329.00 0.29 0.12 1.46 0.99 0.41 0.97 15.45 1.05 7.66 0.72136.76 76212.00 0.32 0.14 1.35 0.92 0.36 0.85 13.91 0.95 8.00 0.75
12 J. Kimmig & B. R. Pratt LETHAIA 10.1111/let.12135

fault. Lime mud-depositing events, however, contin-ued during these intervals because they recorded bythin interbeds and nodules of lime mudstone. Thein situ nature of the fauna is indicated by articulatedhyoliths, agnostoids and trilobites. The fact that soft-bodied fossil preservation is not present throughoutthe calcareous mudstone could be due to either dif-ferences in decay rate and burial, or that the biotawas absent.
Sedimentation was discontinuous, as shown bythe lamination, varying lithology and the evidencefor scavenging of arthropod carapaces while theywere exposed on the seafloor. The planar-lami-nated mixtures of clay, silt and lime mud, com-prising the fossil-bearing host beds, indicates low-energy depositional events, from either hemipela-gic fall-out from sediment plumes, turbidity cur-rents or contour currents. If the laminaerepresent Bouma E divisions, there should be evi-dence for more complete turbidites in the succes-sion. However, none has yet been observed.Contour currents might have existed in the Sel-
Fig. 9. Relationship between total organic carbon content (TOC) and the abundance of selected redox-sensitive trace elements in thelower calcareous mudstone interval 134–137 m, Rockslide Formation at the Ravens Throat River Lagerst€atte.
Fig. 10. Relationship of total organic carbon content to theabundance of d13Corg in the lower calcareous mudstone interval134–137 m, Rockslide Formation at the Ravens Throat RiverLagerst€atte.
LETHAIA 10.1111/let.12135 Middle Cambrian taphonomy 13

wyn Basin. The silty and muddy contourite types(Stow et al. 2002) are similar to the calcareousmudstone facies. However, contour currents rep-resent long-lived conditions of oceanic circulationand therefore do not explain the limited extent ofthis facies.
The variable amounts of organic matter, especiallyas discontinuous carbonaceous seams and flakes,imply a combination of water-column productivityand benthic microbial mats. The lack of bioturbationin the lime mudstone interbeds likely reflects rapidsedimentation rates or possible dysoxic conditions.
Trace elements in the fossiliferous calcareousmudstone are mostly depleted or close to ‘averageshale’ values (e.g. Rudnick & Gao 2005; Tribovillardet al. 2006). The element ratios for U/Th and V/Crboth imply continuously oxygenated bottom waters,whereas V/Sc indicates suboxic conditions (Fig. 11).This suggests that the bottom waters were mostlyoxygenated with periods of dysoxia. Assuming thatthe V/Sc ratio was not disturbed by rapid sedimenta-tion events (Tribovillard et al. 2006), it is possiblethat episodic dysoxia led to a lower faunal diversityin the Ravens Throat River Lagerst€atte, comparedwith most other Cambrian BST examples. The oxic/anoxic boundary must have been at, or just beneath,the sediment–water interface.
Comparison
The fossils in the Burgess Shale were initially thoughtto represent a fauna that was transported fromshallower water (e.g. Whittington 1985). However,subsequent taphonomic analysis suggests that thebiota is in situ and represents a primary community(Caron & Jackson 2006; O’Brien & Caron 2012).Investigations of other BST deposits suggest thatthey also host in situ biotas: Emu Bay (Gehling et al.2011); Wheeler and Marjum formations (Briggset al. 2008); Sirius Passet (Babcock & Peel 2007);and Chengjiang (Zhang et al. 2001).
Earlier interpretations of the Burgess Shale con-cluded that the sediments consist of intermittentturbidites (Piper 1972), thus supporting thehypothesis that it was rapid burial that led toexceptional preservation (Piper 1972; Allison &Brett 1995). This interpretation has since beenchallenged. Gabbott et al. (2008) proposed thatrapid sediment deposition was from mud-richsemi-continuous density currents. Similar to theBurgess Shale, most BST deposits have been depos-ited as laminated mudstones that do not showindicators of turbidites, especially ripples and dis-tinct graded bedding. The Emu Bay Shale hassome evidence of syndepositional deformation, but
Fig. 11. Chemostratigraphic profiles of redox-sensitive trace-element ratios for the Burgess Shale-type unit of the Rockslide Formationat the Ravens Throat River Lagerst€atte. Vertical subdivisions represent the redox zones of Jones & Manning (1994) and Kimura & Watan-abe (2001). Height is above base of the lower calcareous mudstone at 134 m.
14 J. Kimmig & B. R. Pratt LETHAIA 10.1111/let.12135

Table
3.Comparisonofmineralogicalcompositionofselected
Laurentian
Cam
brian
shales
both
withandwithoutBurgessSh
ale-type(BST
)preservation.Samplesfrom
theRockslideForm
a-tionnotfrom
theRavensThroat
River
Lagerst€ attewerecollectedfrom
thetypesection(section19;G
abrielse
etal.1973).T
hesamplefrom
northernMontanawas
collectedfrom
theLittle
RockyMountainswheretheDeadwoodForm
ationiskn
ownas
theEmersonForm
ation.
Form
ation
Age
Biozone
Location
BST
Rock
type
Quartz
(%)
Carbonate
(%)
Chlorite
(%)
Illite
(%)
K-spar
(%)
Other
(%)
Other
References
Mural
Lower
Cam
brian
Bonnia–
Olenellus
Rocky
Mountains
YMudstone
300
268
00
–Balthasar
&Butterfield(2009)
Eager
Lower
Cam
brian
Bonnia–
Olenellus
Rocky
Mountains
YMudstone
240
2749
00
–Copeland(1993)
KaneBasin
Lower
Cam
brian
Bonnia–
Olenellus
Ellesmere
Island
NMudstone
160
1514
550
–Dew
inget
al.
(2008)
Quyuk
Lower
Cam
brian
Bonnia–
Olenellus
Victoria
Island
NSiltyshale
00
04
4254
Montm
orillonite
Dew
ing,K.D.,
Hadlari,
T.andPratt,
B.R.(in
preparation)
Mount
Phayre
Middle
Cam
brian
Plagiura–
Poliella
Victoria
Island
NSh
ale
615
00
3346
Montm
orillonite
Dew
ing,K.D.,
Hadlari,T.and
Pratt,B.R.
(inpreparation)
Burgess
Shale
Middle
Cam
brian
Bathyuriscus–
Elrathina
Rocky
Mountains
YMudstone
2812
2435
00
–Johnston
etal.(2009a)
Rockslide
Middle
Cam
brian
Plagiura–
Poliella
Mackenzie
Mountains
NSiltyshale
305
3629
00
–Gabrielse
etal.(1973)
Rockslide
Middle
Cam
brian
Albertella
Mackenzie
Mountains
NSiltyshale
2324
044
90
–Gabrielse
etal.(1973)
Rockslide
Middle
Cam
brian
Albertella
Mackenzie
Mountains
NSiltyshale
195
3542
00
–Gabrielse
etal.(1973)
Rockslide
Middle
Cam
brian
Albertella
Mackenzie
Mountains
NSh
ale
2249
227
00
–Thispaper
Rockslide
Middle
Cam
brian
Albertella
Mackenzie
Mountains
NSiltyshale
2249
227
00
–Thispaper
Rockslide
Middle
Cam
brian
Bolaspidella
Mackenzie
Mountains
NLim
emudstone
1780
03
00
–Thispaper
Rockslide
Middle
Cam
brian
Bolaspidella
Mackenzie
Mountains
NLim
emudstone
3260
53
00
–Thispaper
Rockslide
Middle
Cam
brian
Bolaspidella
Mackenzie
Mountains
YCalcareous
mudstone
4421
1223
00
–Thispaper
Rockslide
Middle
Cam
brian
Bolaspidella
Mackenzie
Mountains
YCalcareous
mudstone
3313
3915
00
–Thispaper
Rockslide
Middle
Cam
brian
Bolaspidella
Mackenzie
Mountains
YCalcareous
mudstone
3410
1838
00
–Thispaper
Rockslide
Middle
Cam
brian
Bolaspidella
Mackenzie
Mountains
YCalcareous
mudstone
370
3429
00
–Thispaper
Rockslide
Middle
Cam
brian
Bolaspidella
Mackenzie
Mountains
YCalcareous
mudstone
490
2427
00
–Thispaper
Rockslide
Middle
Cam
brian
Bolaspidella
Mackenzie
Mountains
YCalcareous
mudstone
2038
1626
00
–Thispaper
LETHAIA 10.1111/let.12135 Middle Cambrian taphonomy 15

is considered to have been deposited under contin-uous sediment supply without significant influenceof bottom currents (Gehling et al. 2011). However,similar to the Burgess Shale, the fossils in the Mar-jum and Wheeler formations are preserved inmostly non-bioturbated finely laminated shales andwere likely deposited by turbidites (Gaines et al.2005; Brett et al. 2009). The host sediments of theChengjiang biota are greenish-grey mudstones inunweathered samples that have been interpretedvariously as episodic turbidity currents (Lindstr€om1995), storm beds (Babcock et al. 2001) or sedi-ments from rapid deposition of suspended mud(Zhu et al. 2001, 2005).
It has long been assumed that bioturbation isinimical to soft-bodied BST preservation (Allison &Briggs 1991). It is now recognized that burrows arepresent in such units, including, for example, theChengjiang and Sirius Passet Lagerst€atten (Zhanget al. 2007; M�angano 2011; Gaines et al. 2012b;M�angano et al. 2012). In the Ravens Throat RiverLagerst€atte, the fossiliferous calcareous mudstoneappears to show common millimetre-sized bioturba-tion and two arthropod valves preserve burrowsbelow the carapaces, whereas the unfossiliferous limemudstone beds are unburrowed. Thus, it seems thatthe presence or absence of bioturbation does notnecessarily correspond to oxic versus anoxic condi-tions in the water column or in the sediment, as pre-viously suggested (e.g. Caron & Jackson 2006), butmay reflect local sedimentation rate, food availabilityand other factors (Buatois & M�angano 2011, p. 38).
The fossiliferous calcareous mudstone of theRavens Throat River Lagerst€atte is similar to theBurgess Shale in that it is composed of graded, pla-nar-laminated, illite, chlorite, quartz and carbonatein similar proportions (Table 3), along with pyritelenses as in the ‘phyllopod bed’. It is also dominatedby clay-sized particles. The fossils imply a similardegree of compaction (~85%; Whittington 1985).The sediments presented an unusual medium thatwas conducive to BST preservation (Wilson & But-terfield 2014). It was not due to sediment mineral-ogy alone, however, as three samples of theRockslide Formation at its type section, while siltier,have a similar composition but do not show BSTpreservation, although trilobites are relatively com-mon. By contrast, samples from the correlativeMount Cap Formation farther to the east, where itdoes not exhibit BST either, have much less chloriteand illite but a significant amount of kaolinite. Onesample from an unnamed early middle Cambrianshale from western Victoria Island lacks chlorite andillite but is correspondingly rich in montmorilloniteand K-feldspar, and also lacks BST preservation.T
able3.
(continued)
Form
ation
Age
Biozone
Location
BST
Rock
type
Quartz
(%)
Carbonate
(%)
Chlorite
(%)
Illite
(%)
K-spar
(%)
Other
(%)
Other
References
Rockslide
Middle
Cam
brian
Bolaspidella
Mackenzie
Mountains
NDolomitic
siltstone
3542
58
100
–Thispaper
MountCap
Middle
Cam
brian
Albertella
Mackenzie
Mountains
NSiltyshale
6310
37
017
Kaolinite
Harvey&
Butterfield
(2011)
MountCap
Middle
Cam
brian
Glossopleura
Mackenzie
Mountains
NSh
ale
668
59
012
Fe-oxide
MacNaughton
etal.(2013)
McK
ayFurongian
Elvina
Rocky
Mountains
NClaystone
3420
312
120
–Chatterton&
Ludvigsen
(1998)
Deadwood
Furongian
Uncertain
Northern
Montana
NSiltyshale
157
111
660
–Lochman
(1950)
16 J. Kimmig & B. R. Pratt LETHAIA 10.1111/let.12135

Gaines et al. (2012a) suggested that early carbon-ate cementation was a key part of the process of soft-bodied preservation because they believed it sealedthe sediment and fossils from decay. However, in theRavens Throat River Lagerst€atte, the laminae arephysically compacted, indicating that near-surfacelithification did not occur. Early diagenetic carbon-ate cements have not been observed in the fossilifer-ous layers. Calcium carbonate cementation tookplace preferentially in concretionary lime mudstoneinterbeds and lenses not close to the sediment–waterinterface but during shallow burial (Westphal et al.2008). These do not host soft-tissue preservation.Furthermore, Butterfield (2012) argued that there isno consistent association of bed-capping carbonateswith BST biotas, and the best BST preservation actu-ally occurs in beds without a significant carbonatecomponent. Slump folds in the Burgess Shale at TheMonarch, and small-scale synsedimentary deforma-tion on Mount Field (B. R. Pratt, unpublished obser-vations) also refute the mechanism of diageneticsealing by carbonate cementation.
In common with other Middle Cambrian mud-stones (Hayes et al. 1999; Guo et al. 2005; Kouchin-sky et al. 2012), the d13Corg values are around�20& and show only minor variation between thecalcareous mudstone and other facies in the succes-sion (Table 1). McKirdy et al. (2011) interpreted thed13Corg signature in the Emu Bay Shale and otherAustralian Lagerst€atten as an indicator of cyanobac-terial origin, with values similar to those of moderncyanobacteria (Brady et al. 2013). The minor differ-ence in values likely are due to the up to ~10% dolo-mite in the calcareous mudstones (Hayes et al.1999). Thus, the majority of the Corg remaining thecalcareous mudstone is interpreted to be of cyano-bacterial origin, mostly from water-column produc-tivity. No biomarkers or aromatic hydrocarbonsremain, however, that would have elucidated thecontributions of various groups, especially addi-tional elements such as sponges.
Clays might have played a role in preservation offossils in the Burgess Shale (Butterfield et al. 2007;Wilson & Butterfield 2014), as the host rock containsabout 50% illite and chlorite. Butterfield (1995)argued that Fe-rich clays might be involved in deac-tivating degradative enzymes, but they might repre-sent evidence for enhanced Fe2+ availability in EarlyPaleozoic seas, which stabilized the substrates andincreased the preservation potential (Wilson & But-terfield 2014). The lack of most soft-tissue preserva-tion in the Ravens Throat River Lagerst€atte indicatesthat this mechanism, if viable, did not halt decay tothe same degree as seen in the Burgess Shale oroccurred at a later stage in the decomposition pro-
cess. Clays are predominant in most BST deposits(Table 3) and thus likely contributed at least to adegree in the preservation of soft tissue, were likelynot the sole contributor to the preservation process.
The high proportion of chlorite in the calcareousmudstone, as well as the Burgess Shale, in contrastto some other Cambrian shales that do not exhibitBST preservation (Table 3), is still suggestive of ataphonomic influence of this clay mineral. Powell(2003) assumed that the origin of the chlorite in theBurgess Shale was due to metamorphic overprinting.On the other hand, illite and chlorite can also beformed by hydrothermal fluids (e.g. Hermann & Hill2001; Meunier & Velde 2004) or can be derived fromweathering of older strata or bedrock. However, thehigh illite content (~30%) of the Ravens ThroatRiver Lagerst€atte BST sediments, as well as a lack ofmica, indicates that lower greenschist metamor-phism was not achieved. There is no evidence forhydrothermal fluid entry into this locality, eventhough there was synsedimentary fault activity.Thus, the chlorite in the calcareous mudstone islikely sedimentary. Considering that volcanic depos-its of Cambrian age have been reported in the Sel-wyn Basin region (Gordey & Anderson 1993;Moynihan 2014), chlorite could have originatedfrom weathering of basalts in those areas. It is alsopossible that much of it could be from erosion ofNeoproterozoic igneous rocks belonging to theFranklin sills, well to the north of the Ravens ThroatRiver Lagerst€atte. On the other hand, on VictoriaIsland clays in Cambrian shales which are in closeproximity to the sills are rich in montmorillonite.The illite component was likely derived from erosionof underlying Proterozoic sedimentary successionscomprising the Mackenzie Mountains Supergroupand older units, as well as the exposed PrecambrianShield (cf. Weaver 1989, p. 563).
Conclusions
The Middle Cambrian (series 3; stage 5 and Drumi-an) Rockslide Formation in the Mackenzie Moun-tains of northwestern Canada is a relatively deep-water succession of shales, mudstones and lime-stones deposited on a slope seaward of a mixed silic-iclastic–carbonate platform. In a single localityoverlying a submarine listric fault scarp, 2-m-thickunits of greenish-coloured, laminated calcareousmudstone yield a low-diversity biota that includesbivalved arthropods, trilobites, agnostoids, worms,hyoliths and macrophytic algae. Preservation ofnon-mineralized components places these beds intothe family of BST Lagerst€atten, and the fossil-bearing
LETHAIA 10.1111/let.12135 Middle Cambrian taphonomy 17

facies is closely similar. Trace-element data indicatethat the bottom waters were oxygenated duringdeposition, but there may have been occasional dys-oxic episodes. Despite the presence of bioturbationand probably scavenging, at least some of the morerobust components of the soft tissues surviveddecomposition and were preserved. Thus, burrow-ing, by itself, likely did not have a primary effect onBST preservation. Lithology and the lack of correla-tion with layers cemented by calcium carbonateunder shallow burial indicate that early carbonatecementation cannot be invoked as a means to sealthe sediment and prevent further decay, at least inthis example.
Consequently, it is fortuitous combination ofenvironmental and ecological factors and the nat-ure of the host sediment that hold the key to BSTpreservation. The specific mixture of lime mud,clay and silt along with clay mineralogy – chloriteand illite – seem to have presented a unique med-ium that slowed decay of buried carcasses. Exactlyhow this combination operated, however, is stillunclear.
Acknowledgements. – We thank M.G. M�angano for help withthe trace fossils, C. West, K. Gerein and B. Uzelman for assistancein the field, B. Novakovski for thin-section preparation, T. Bonlifor XRD, D. Besic (Saskatchewan Isotope Laboratory) forisotopic analyses, R. Millar (Saskatchewan Research Council) fortrace-element analyses, and V. Thiel and J.-P. Duda (Universityof G€ottingen) for biomarker analyses. Specimens have been col-lected under Aurora Institute Research Licenses delivered toBRP. The expeditions to the Mackenzie Mountains were possiblethanks to support of the Natural Sciences and EngineeringResearch Council grants to BRP. J. Schiffbauer (University ofMissouri) and an anonymous reviewer are thanked for reviewingthe manuscript. This paper is part of J. Kimmig’s PhD research.
References
Algeo, T.J. & Maynard, J.B. 2004: Trace-element behavior andredox facies in core shales of Upper Pennsylvanian Kansas-type cyclothems. Chemical Geology 206, 289–318.
Allison, P.A. 1988: The role of anoxia in the decay and minerali-zation of proteinaceous macrofossils. Paleobiology 14, 139–154.
Allison, P.A. & Brett, C.E. 1995: In situ benthos and paleo-oxy-genation in the Middle Cambrian Burgess Shale, BritishColumbia, Canada. Geology 23, 1079–1082.
Allison, P.A. & Briggs, D.E.G. 1991: Taphonomy of nonmineral-ized tissues. In Allison, P.A. & Briggs, D.E.G. (eds): Taphon-omy: Releasing the Data Locked in the Fossil Record, 25–70.Plenum, New York.
Babcock, L.E. & Peel, J.S. 2007: Palaeobiology, taphonomy andstratigraphic significance of the trilobite Buenellus from theSirius Passet Biota, Cambrian of North Greenland. In LaurieJ.R. & Paterson J.R. (eds): Papers in honour of John H. Sher-gold 1938–2006. Memoirs of the Australasian Association ofPalaeontologists, 34, 401–418.
Babcock, L.E., Zhang, W.-T. & Leslie, S.A. 2001: The Chengji-ang Biota: record of the Early Cambrian diversification oflife and clues to exceptional preservation of fossils. GSAToday 11, 4–9.
Balthasar, U. & Butterfield, N.J. 2009: Early Cambrian ‘soft–shelled’ brachiopods as possible stem–group phoronids. ActaPalaeontologica Polonica 54, 307–314.
Brady, A.L., Druschel, G., Leoni, L., Lim, D.S.S. & Slater, G.F.2013: Isotopic biosignatures in carbonate-rich, cyanobacteria-dominated microbial mats of the Cariboo Plateau, B.C. Geobi-ology 11, 437–456.
Brett, C.E., Allison, P.A., DeSantis, M.K., Liddell, W.D. & Kra-mer, A. 2009: Sequence stratigraphy, cyclic facies, and lag-erst€atten in the Middle Cambrian Wheeler and Marjumformations, Great Basin, Utah. Palaeogeography, Palaeoclima-tology, Palaeoecology 277, 9–33.
Briggs, D.E.G., Erwin, D.H. & Collier, F.J. 1994: The Fossils of theBurgess Shale, 238 pp. Smithsonian Institution Press, Washing-ton.
Briggs, D.E.G., Lieberman, B.S., Hendricks, J.H., Halgedahl, S.L.& Jarrard, R.D. 2008: Middle Cambrian arthropods fromUtah. Journal of Paleontology 82, 238–254.
Buatois, L. & M�angano, M.G. 2011: Ichnology. Organism-Sub-strate Interactions in Space and Time, 358 pp. Cambridge Uni-versity Press, Cambridge.
Butterfield, N.J. 1990: Organic preservation of non-mineralizingorganisms and the taphonomy of the Burgess Shale. Paleobiol-ogy 16, 272–286.
Butterfield, N.J. 1994: Burgess Shale-type fossils from a LowerCambrian shallow-shelf sequence in northwestern Canada.Nature 369, 477–479.
Butterfield, N.J. 1995: Secular distribution of Burgess-Shale-typepreservation. Lethaia 28, 1–13.
Butterfield, N.J. 2012: Does cement-induced sulfate limitationaccount for Burgess Shale-type preservation? Proceedings of theNational Academy of Sciences 109, E1901.
Butterfield, N.J. & Nicholas, C.J. 1996: Burgess Shale-type preser-vation of both non-mineralizing and ‘shelly’ Cambrian organ-isms from the Mackenzie Mountains, northwestern Canada.Journal of Paleontology 70, 893–899.
Butterfield, N.J., Balthasar, U. & Wilson, L.A. 2007: Fossil dia-genesis in the Burgess Shale. Palaeontology 50, 537–543.
Calvert, S.E. 1987: Oceanographic controls on the accumulationof organic matter in marine sediments. In Brooks, J. & Fleet,A.J. (eds): Marine Petroleum Source Rocks, 137–151. GeologicalSociety of London Special Publication 26, Oxford.
Caron, J.B. & Jackson, D.A. 2006: Taphonomy of the greaterphyllopod bed community, Burgess Shale. Palaios 21, 451–465.
Cecile, M.P. 1982: The lower Paleozoic Misty Creek embayment,Selwyn Basin, Yukon and Northwest Territories. GeologicalSurvey of Canada, Bulletin 335, 1–78.
Chatterton B. D. E. & Ludvigsen, R. 1998: Upper Steptoean(Upper Cambrian) trilobites from the McKay Group of south-eastern British Columbia, Canada. The Paleontological SocietyMemoir 49, 43 pp.
Chevrier, T.S. & Turner, E.C. 2013: Lithostratigraphy of deep-water Paleozoic strata in the central misty creek embayment,Mackenzie Mountains, Northwest Territories. Geological Sur-vey of Canada, Current Research 2013–2014, 1–21.
Collom, C.J., Johnston, P.A. & Powell, W.G. 2009: Reinterpreta-tion of ‘Middle’ Cambrian stratigraphy of the rifted westernLaurentian margin: Burgess Shale Formation and contiguousunits (Sauk II Megasequence); Rocky Mountains, Canada. Pal-aeogeography, Palaeoclimatology, Palaeoecology 277, 63–85.
Coniglio, M. & James, N.P. 1990: Origin of fine-grained carbon-ate and siliciclastic sediments in an Early Palaeozoic slopesequence, Cow Head Group, western Newfoundland. Sedimen-tology 37, 215–230.
Copeland, M.J. 1993: Anomalocaris (of unknown affinity) andTuzoia (a possible arthropod) from the Lower Cambrian EagerFormation near Cranbrook, British Columbia. Geological Sur-vey of Canada 444, 1–5.
Demaison, G.J. & Moore, G.T. 1980: Anoxic environments andoil source bed genesis. Organic Geochemistry 2, 9–31.
Dewing, K., Mayr, U., Harrison, J.C. & de Freitas, T. 2008: Upperneoproterozoic to lower devonian stratigraphy of northeast
18 J. Kimmig & B. R. Pratt LETHAIA 10.1111/let.12135

Ellesmere Island. In Mayr, U. (ed.): Geology of Northeast Elles-mere Island Adjacent to Kane Basin and Kennedy Channel,Nunavut. Geological Survey of Canada Bulletin 592, 31–108.
Dixon, J. & Stasiuk, L.D. 1998: Stratigraphy and hydrocarbonpotential of Cambrian strata, Northern Interior Plains, North-west Territories. Bulletin of Canadian Petroleum Geology 46,445–470.
Edgecombe, G.D. & Legg, D.A. 2013: The arthropod fossilrecord. In Minelli, A., Boxshall, G. & Fusco, G. (eds): Arthro-pod Biology and Evolution, Molecules, Development, Morphol-ogy, 393–415. Springer, Berlin.
Farrell, �U.C., Briggs, D.E., Hammarlund, E.U., Sperling, E.A. &Gaines, R.R. 2013: Paleoredox and pyritization of soft-bodiedfossils in the Ordovician Frankfort Shale of New York. Ameri-can Journal of Science 313, 452–489.
Fritz, W.H. 1979: Cambrian stratigraphic section between SouthNahanni and Broken Skull Rivers, southern Mackenzie Moun-tains. Geological Survey of Canada Paper, 79-1B, 121–125.
Gabbott, S.E., Zalasiewicz, J. & Collins, D. 2008: Sedimentationof the phyllopod bed within the Cambrian Burgess Shale For-mation of British Columbia. Journal of the Geological Society,London 165, 307–818.
Gabrielse, H., Blusson, S.L. & Roddick, J.A. 1973: Geology of FlatRiver, Glacier Lake, and Wrigley Lake map-areas, District ofMackenzie and Yukon Territory. Geological Survey of Canada,Memoir 366, 1–153.
Gaines, R.R., Kennedy, M.J. & Droser, M.L. 2005: A new hypoth-esis for organic preservation of Burgess Shale taxa in the Mid-dle Cambrian Wheeler Formation, House Range, Utah.Palaeogeography, Palaeoclimatology, Palaeoecology 220, 193–205.
Gaines, R.R., Briggs, D.E.G. & Zhao, Y. 2008: Cambrian BurgessShale-type deposits share a common mode of fossilization.Geology 36, 755–758.
Gaines, R.R., Mering, J.A., Zhao, Y. & Peng, J. 2011: Stratigraphicand microfacies analysis of the Kaili Formation, a candidateGSSP for the Cambrian Series 2–Series 3 boundary. Palaeoge-ography, Palaeoclimatology, Palaeoecology 311, 171–183.
Gaines, R.R., Hammarlund, E.U., Hou, X., Qi, C., Gabbott, S.E.,Zhao, Y., Peng, J. & Canfield, D.E. 2012a: Mechanism for Bur-gess Shale-type preservation. Proceedings of the National Acad-emy of Sciences 109, 5180–5184.
Gaines, R.R., Droser, M.L., Orr, P.J., Garson, D., Hammarlund,E., Qi, C. & Canfield, D.E. 2012b: Burgess shale-type biota’swere not entirely burrowed away. Geology 40, 283–286.
Garson, D.E., Gaines, R.R., Droser, M.L., Liddell, W.D. & Sap-penfield, A. 2012: Dynamic palaeoredox and exceptional pres-ervation in the Cambrian Spence Shale of Utah. Lethaia 45,164–177.
Gehling, J.G., Jago, J.B., Paterson, J.R., Garc�ıa-Bellido, D.C. &Edgecombe, G.D. 2011: The geological context of the lowerCambrian (Series 2) Emu Bay Shale Lagerst€atte and adjacentstratigraphic units, Kangaroo Island, South Australia. Austra-lian Journal of Earth Sciences 58, 243–257.
Gordey, S.P. & Anderson, J.G. 1993: Evolution of the northernCordilleran miogeocline, Nahanni map area (105I), Yukonand Northwest Territories. Geological Survey Canada Memoir428, 1–214.
Guo, Q.-J., Strauss, H., Liu, C.-Q., Zhao, Y.-L., Pi, D.-H., Fu, P.-Q., Zhu, L.-J. & Yang, R.-D. 2005: Carbon and oxygen isotopiccomposition of Lower to Middle Cambrian sediments at Taiji-ang, Guizhou Province, China. Geological Magazine 142, 723–733.
Guo, Q.-J., Strauss, H., Zhao, Y., Yang, X., Peng, J., Yang, Y. &Deng, Y. 2014: Reconstructing marine redox conditions forthe transition between Cambrian Series 2 and Cambrian Series3, Kaili area, Yangtze Platform: evidence from biogenic sulfurand degree of pyritization. Palaeogeography, Palaeoclimatology,Palaeoecology 398, 144–153.
Hadlari, T., Davis, W.J., Dewing, K., Heaman, L.M., Lemieux, Y.,Ootes, L., Pratt, B.R. & Pyle, L.J. 2012: Two detrital zircon sig-natures for the Cambrian passive margin of northern Lauren-
tia highlighted by new U-Pb results from northern Canada.Geological Society of America Bulletin 124, 1155–1168.
Hall, P.A., McKirdy, D.M., Halverson, G.P., Jago, J.B. & Gehling,J.G. 2011: Biomarker and isotopic signatures of an early Cam-brian Lagerst€atte in the Stansbury Basin, South Australia.Organic Geochemistry 42, 1324–1330.
Harvey, T.H.P. & Butterfield, N.J. 2011: Great CanadianLagerst€atten 2. Macro- and Microfossils of the Mount CapFormation (Early and Middle Cambrian, Northwest Territo-ries). Geoscience Canada 38, 165–173.
Hayes, J.M., Strauss, H. & Kaufman, A.J. 1999: The abundance of13C in marine organic matter and isotopic fractionation in theglobal biogeochemical cycle of carbon during the past 800 Ma.Chemical Geology 161, 103–125.
Hermann, W. & Hill, A.P. 2001: The origin of chlorite–tremo-lite–carbonate rocks associated with the Thalanga volcanic-hosted massive sulfide deposit, noth Queensland, Australia.Economic Geology 96, 1149–1173.
Johnston, P.A., Johnston, K.J., Collom, C.J., Powell, W.G. & Pol-lock, R.J. 2009a: Palaeontology and depositional environmentsof ancient brine seeps in the Middle Cambrian Burgess Shaleat The Monarch, British Columbia, Canada. Palaeogeography,Palaeoclimatology, Palaeoecology 277, 86–105.
Johnston, K.J., Johnston, P.A. & Powell, W.G. 2009b: A new,Middle Cambrian, Burgess Shale-type biota, BolaspidellaZone, Chancellor Basin, southeastern British Columbia.Palaeogeography, Palaeoclimatology, Palaeoecology 277, 106–126.
Jones, B. & Manning, D.A.C. 1994: Comparison of geochemicalindices used for the interpretation of paleoredox conditions inancient sediments. Chemical Geology 111, 111–119.
Kimmig, J. & Pratt, B.P. 2015: Soft-bodied biota from the middleCambrian (Drumian) Rockslide Formation, Mackenzie Moun-tains, northwestern Canada. Journal of Paleontology 89, 51–71.
Kimura, H. & Watanabe, Y. 2001: Ocean anoxia at the Precam-brian-Cambrian boundary. Geology 29, 995–998.
Kouchinsky, A., Bengston, S., Runnegar, B., Skovsted, C., Steiner,M. & Vendrasco, M. 2012: Chronology of early Cambrian bio-mineralization. Geological Magazine 149, 221–251.
Le Boudec, A., Ineson, J., Rosing, M., Døssing, L., Martineau, F.,L�ecuyer, C. & Albar�ede, F. 2014: Geochemistry of the Cam-brian Sirius Passet Lagerst€atte, northern Greenland. Geochem-istry, Geophysics, Geosystems 15, 886–904.
Lindstr€om, M. 1995: The environment of the Early CambrianChengjiang fauna. In Chen, J.Y., Edgecombe, G. & Ramsk€old,L. (eds): International Cambrian Explosion Symposium, Pro-gramme and Abstracts, 17.
Lochman, C. 1950: Upper Cambrian faunas of the Little RockyMountains, Montana. Journal of Paleontology 24, 32–349.
Lubeseder, S., Redfern, J. & Boutib, L. 2009: Mixed siliciclas-tic-carbonate shelf sedimentation – Lower Devoniansequences of the SW Anti-Atlas, Morocco. SedimentaryGeology 215, 13–32.
MacNaughton, R.B., Pratt, B.R. & Fallas, K.M. 2013: Observa-tions on Cambrian stratigraphy in the eastern MackenzieMountains, Northwest Territories. Geological Survey of CanadaCurrent Research 2013–10, 1–7.
Maloof, A.C., Porter, S.M., Moore, J.L., Dud�as, F.€O., Bowring,S.A., Higgins, J.A., Fike, D.A. & Eddy, M.P. 2010: Theearliest Cambrian record of animals and ocean geochemicalchange. Geological Society of America Bulletin 122, 1731–1774.
M�angano, M.G. 2011: Trace-fossil assemblages in a BurgessShale-type deposit from the Stephen Formation at StanleyGlacier, Canadian Rocky Mountains: unraveling ecologic andevolutionary controls. Palaeontographica Canadiana 31,89–118.
M�angano, M.G., Bromley, R.G., Harper, D.A.T., Nielsen, A.T.,Smith, M.P. & Vinther, J. 2012: Nonbiomineralized carapacesin Cambrian seafloor landscapes (Sirius Passet, Greenland):opening a new window into early Phanerozoic benthic ecol-ogy. Geology 40, 519–522.
LETHAIA 10.1111/let.12135 Middle Cambrian taphonomy 19

McKirdy, D.M., Hall, P.A., Nedin, C., Halverson, G.P., Michael-sen, B.H., Jago, J.B., Gehling, J.G. & Jenkins, R.J.F. 2011: Pale-oredox status and thermal alteration of the lower Cambrian(Series 2) Emu Bay Shale Lagerst€atte, South Australia. Austra-lian Journal of Earth Sciences 58, 259–272.
Meunier, A. & Velde, B. 2004: Illite: Origins, Evolution and Meta-morphism, 286 pp. Springer, Heidelberg.
Moynihan, D. 2014: Bedrock Geology of NTS 106B/04, easternRackla Belt. In MacFarlane, K.E., Nordling, M.G. & Sack, P.J.(eds): Yukon Exploration and Geology 2013, 147–167. YukonGeological Survey, Whitehorse.
O’Brien, L.J. & Caron, J.-B. 2012: A new stalked filter-feederfrom the Middle Cambrian Burgess Shale, British Columbia,Canada. PLoS One 7, e29233.
Piper, D.J.W. 1972: Sediments of the Middle Cambrian BurgessShale. Lethaia 5, 169–175.
Piper, D.Z. & Isaacs, C.M. 1995: Minor elements in Quaternarysediment from the Sea of Japan: a record of surface-water pro-ductivity and intermediate-water redox conditions. GeologicalSociety of America Bulletin 107, 54–67.
Powell, W.G. 2003: Greenschist-facies metamorphism of the Bur-gess Shale and its implications for models of fossil formationand preservation. Canadian Journal of Earth Sciences 40, 13–25.
Powell, W.G. 2009: Comparison of geochemical anddistinctive mineralogical features associated with theKinzers and Burgess Shale formations and their associatedunits. Palaeogeography, Palaeoclimatology, Palaeoecology 277,127–140.
Powell, W.G., Johnston, P.A. & Collom, C.J. 2003: Geochemi-cal evidence for oxygenated bottom waters duringdeposition of fossiliferous strata of the Burgess Shale For-mation. Palaeogeography, Palaeoclimatology, Palaeoecology201, 249–268.
Pratt, B.R. 1989: Deep-water Girvanella-Epiphyton reef on a mid-Cambrian continental slope, Rockslide Formation, MackenzieMountains, Northwest Territories. In Geldsetzer, H.H.J.,James, N.P. & Tebbutt, G.E. (eds): Reefs, Canada and AdjacentAreas. Canadian Society of Petroleum Geologists, Memoir 13,161–164.
Pratt, B.R. 1992: Trilobites of the Marjuman and Steptoeanstages (Upper Cambrian), Rabbitkettle Formation, southernMackenzie Mountains, northwest Canada. PalaeontographicaCanadiana 9, 1–179.
Pyle, L.J., Allen, T.L., Fraser, T.A., Gal, L.P., Hadlari, T., Jones,A.L., Lemieux, Y. & Zantvoort, W.G. 2007: Field-based strati-graphic studies in the Peel Plateau, Peel Plain, and adjacentareas, Northwest Territories and Yukon. Geological Survey ofCanada Current Research 2007A-7, 1–11.
Rimmer, S.M. 2004: Geochemical paleoredox indicators in Devo-nian-Mississippian black shales, Central Appalachian Basin(USA). Chemical Geology 206, 373–391.
Rudnick, R.L. & Gao, S. 2005: Composition of the continentalcrust. In Holland, H.D. & Turekian, K.K. (eds): Treatise onGeochemistry – Volume 5, The Crust, 1–56. Elsevier, Amster-dam.
Sagemann, J., Bale, S.J., Briggs, D.E.G. & Parkes, R.J. 1999: Con-trols on the formation of authigenic minerals in associationwith decaying organic matter: an experimental approach. Geo-chimica et Cosmochimica Acta 63, 1083–1095.
Smith, D. 2007: Sequence Stratigraphy of the Middle CambrianMarjum Formation: Response of Sedimentary Facies and Biotato Sea-Level Fluctuations [unpublished Master’s thesis], 1–184.Utah State University, Utah.
Stow, D.A.V., Fauge‘res, J.-C., Howe, J.A., Pudsey, C.J. & Viana,A.R. 2002: Bottom currents, contourites and deep-sea sedi-ment drifts: current state-of-the- art. In Stow, D.A.V., Pudsey,C.J., Howe, J.A., Faug�eres, J.-C. & Viana, A.R. (eds): Deep-Water Contourite Systems; Modern Drifts and Ancient Series,Seismic and Sedimentary Characteristics, 7–20. Geological Soci-ety, London, Memoir, 22.
Tribovillard, N., Algeo, T.J., Lyons, T. & Riboulleau, A. 2006:Trace metals as paleoredox and paleoproductivity proxies: anupdate. Chemical Geology 232, 12–32.
Turekian, K.K. & Wedepohl, K.H. 1961: Distribution of the ele-ments in some major units of the Earth’s crust. Geological Soci-ety of America Bulletin 72, 175–192.
Weaver, C.E. 1989: Clays, Muds, and Shales. Developments in Sed-imentology, 819 pp. Elsevier, New York.
Webster, M., Gaines, R.R. & Hughes, N.C. 2008: Microstratigra-phy, trilobite biostratinomy, and depositional environmentof the ‘Lower Cambrian’ Ruin Wash Lagerst€atte, PiocheFormation, Nevada. Palaeogeography, Palaeoclimatology, Pal-aeoecology 264, 100–122.
Wedepohl, K.H. 1971: Environmental influences on the chemicalcomposition of shales and clays. In Ahrens, L.H., Press, F.,Runcorn, S.K. & Urey, H.C. (eds): Physics and Chemistry of theEarth, volume 8, 307–331. Pergamon, Oxford.
Westphal, H., Munnecke, A., Bohm, F. & Bornholdt, S. 2008:Limestone-marl alternations in epeiric sea settings – witnessesof environmental changes or diagenesis. Geological Associationof Canada Special Paper 48, 389–406.
Whittington, H.B. 1985: The Burgess Shale, 151 pp. Yale Univer-sity Press, New Haven.
Wilson, L.A. & Butterfield, N.J. 2014: Sediment effects on thepreservation of Burgess Shale-type compression fossils. Palaios29, 145–154.
Zhang, X., Shu, D., Li, Y. & Han, J. 2001: New sites of Chengjiangfossils: crucial windows on the Cambrian explosion. Journal ofthe Geological Society, London 158, 211–218.
Zhang, X., Bergstr€om, J., Bromley, R.G. & Hou, X. 2007: Dimin-utive trace fossils in the Chengjiang Lagerst€atte. Terra Nova 19,407–412.
Zhu, M.-Y., Zhang, J.-M. & Li, G.-X. 2001: Sedimentary environ-ments of the early Cambrian Chengjiang Biota: sedimentologyof the Yu’anshan Formation in Chengjiang County, easternYunnan. Acta Palaeontologica Sinica 40, 80–105.
Zhu, M., Babcock, L.E. & Steiner, M. 2005: Fossilization modesin the Chengjiang Lagerst€atte (Cambrian of China): testing theroles of organic preservation and diagenetic alteration inexceptional preservation. Palaeogeography, Palaeoclimatology,Palaeoecology 220, 31–46.
20 J. Kimmig & B. R. Pratt LETHAIA 10.1111/let.12135




















