
A rare non-trilobite artiopodan from the Guzhangian (Cambrian Series 3) Weeks Formation...
12
A RARE NON-TRILOBITE ARTIOPODAN FROM THE GUZHANGIAN (CAMBRIAN SERIES 3) WEEKS FORMATION KONSERVAT-LAGERST € ATTE IN UTAH, USA by JAVIER ORTEGA-HERN ANDEZ 1,2 *, RUDY LEROSEY-AUBRIL 3 *, CARLO KIER 4 and ENRICO BONINO 4 1 Department of Earth Sciences, University of Cambridge, Downing Street, Cambridge, CB2 3EQ, UK; e-mail: [email protected] 2 Emmanuel College, University of Cambridge, St Andrews Street, Cambridge, CB2 3AP, UK 3 Laboratoire de G eologie de Lyon: Terre, Plan etes, Environnement (UMR 5276 CNRS), Universit e Claude Bernard Lyon 1, 2 rue Rapha€ el Dubois, 69622, Villeurbanne, France; e-mail: [email protected] 4 Back to the Past Museum, Carretera Canc un, Puerto Morelos, Quintana Roo 77580, M exico; e-mails: [email protected], [email protected] *Corresponding authors Typescript received 19 July 2014; accepted in revised form 26 September 2014 Abstract: We describe a weakly biomineralized non-trilo- bite artiopodan arthropod from the Guzhangian Weeks Formation of Utah. Falcatamacaris bellua gen. et sp. nov. is typified by a thin calcitic cuticle, broad cephalon without eyes or dorsal ecdysial sutures, an elongate trunk with distinctively sickle-shaped pleural spines and a long tailspine with a bifur- cate termination. The precise affinities of Falcatamacaris gen. nov. are problematic due to the presence of unique features within Artiopoda, such as the peculiar morphology of the pleural and posterior regions of the trunk. Possible affinities with aglaspidid-like arthropods and concilitergans are dis- cussed based on the possession of 11 trunk tergites, edge-to- edge articulations and overall body spinosity. The new taxon highlights the importance of the Weeks Formation Konservat- Lagerst€ atte for further understanding the diversity of extinct arthropod groups in the upper Cambrian. Key words: Arthropoda, non-trilobite Artiopoda, excep- tional preservation, Laurentia, biomineralization, Cambrian explosion. A RTIOPODA Hou and Bergstr€ om, 1997, is an important group of extinct arthropods that dominated the Palaeo- zoic Era due to their remarkable ecological success. By their abundance and diversity, trilobites are the most emblem- atic representatives of this clade; however, Artiopoda also includes numerous non-trilobite taxa described from Lower Palaeozoic deposits and particularly from sites of exceptional preservation around the world (Hendricks et al. 2008; Hendricks 2013). Non-trilobite artiopodans display a considerable degree of morphological variability, especially in terms of dorsal tagmosis (Edgecombe and Ramsk€ old 1999) and biramous limb construction (Ortega- Hern andez et al. 2013, fig. 4). Various non-trilobite Artiopoda have been successfully classified into relatively well-defined groups, which have also been supported by phylogenetic analyses (e.g. Xandarellida, Nektaspida, Con- ciliterga; see Hou and Bergstr€ om 1997; Paterson et al. 2012; Ortega-Hern andez et al. 2013). However, a few taxa are notorious for their unusual morphologies, which complicate their classification and precise phylogenetic affinities (e.g. Habelia optata Walcott, 1912; Phytophilaspis pergamena Ivanstov, 1999), or which occupy an unspeci- fied basal position relative to better defined groups (e.g. Retifacies abnormalis Hou et al. 1989; Squamacula clypeata Hou and Bergstr€ om 1997; see Paterson et al. 2012, Ortega-Hern andez et al. 2013, Stein et al. 2013). Here, we describe a new weakly biomineralized artiopodan from the late Guzhangian (Cambrian Series 3) Weeks Forma- tion of Utah (USA), which combines a mosaic of mor- phological characteristics as-yet observed in several artiopodan groups. The new taxon contributes towards the increasing knowledge on the diversity of late Cam- brian non-trilobite arthropods as informed by the Weeks Formation Konservat-Lagerst€ atte. GEOLOGICAL SETTING The Weeks Formation consists of a 300-m-thick sequence of thin-bedded lime mudstones, wackestones and grain- stones with variable amounts of shale. It crops out in the North Canyon and neighbouring areas only, in the © The Palaeontological Association doi: 10.1111/pala.12136 265 [Palaeontology, Vol. 58, Part 2, 2015, pp. 265–276]
Transcript of A rare non-trilobite artiopodan from the Guzhangian (Cambrian Series 3) Weeks Formation...

A RARE NON-TRILOBITE ARTIOPODAN FROM THE
GUZHANGIAN (CAMBRIAN SERIES 3) WEEKS
FORMATION KONSERVAT-LAGERST€ATTE IN UTAH,
USA
by JAVIER ORTEGA-HERN �ANDEZ1,2*, RUDY LEROSEY-AUBRIL3*,
CARLO KIER4 and ENRICO BONINO4
1Department of Earth Sciences, University of Cambridge, Downing Street, Cambridge, CB2 3EQ, UK; e-mail: [email protected] College, University of Cambridge, St Andrews Street, Cambridge, CB2 3AP, UK3Laboratoire de G�eologie de Lyon: Terre, Plan�etes, Environnement (UMR 5276 CNRS), Universit�e Claude Bernard Lyon 1, 2 rue Rapha€el Dubois, 69622,
Villeurbanne, France; e-mail: [email protected] to the Past Museum, Carretera Canc�un, Puerto Morelos, Quintana Roo 77580, M�exico; e-mails: [email protected], [email protected]
*Corresponding authors
Typescript received 19 July 2014; accepted in revised form 26 September 2014
Abstract: We describe a weakly biomineralized non-trilo-
bite artiopodan arthropod from the Guzhangian Weeks
Formation of Utah. Falcatamacaris bellua gen. et sp. nov. is
typified by a thin calcitic cuticle, broad cephalon without eyes
or dorsal ecdysial sutures, an elongate trunk with distinctively
sickle-shaped pleural spines and a long tailspine with a bifur-
cate termination. The precise affinities of Falcatamacaris gen.
nov. are problematic due to the presence of unique features
within Artiopoda, such as the peculiar morphology of the
pleural and posterior regions of the trunk. Possible affinities
with aglaspidid-like arthropods and concilitergans are dis-
cussed based on the possession of 11 trunk tergites, edge-to-
edge articulations and overall body spinosity. The new taxon
highlights the importance of the Weeks Formation Konservat-
Lagerst€atte for further understanding the diversity of extinct
arthropod groups in the upper Cambrian.
Key words: Arthropoda, non-trilobite Artiopoda, excep-
tional preservation, Laurentia, biomineralization, Cambrian
explosion.
ART IOPODA Hou and Bergstr€om, 1997, is an important
group of extinct arthropods that dominated the Palaeo-
zoic Era due to their remarkable ecological success. By their
abundance and diversity, trilobites are the most emblem-
atic representatives of this clade; however, Artiopoda also
includes numerous non-trilobite taxa described from
Lower Palaeozoic deposits and particularly from sites of
exceptional preservation around the world (Hendricks
et al. 2008; Hendricks 2013). Non-trilobite artiopodans
display a considerable degree of morphological variability,
especially in terms of dorsal tagmosis (Edgecombe and
Ramsk€old 1999) and biramous limb construction (Ortega-
Hern�andez et al. 2013, fig. 4). Various non-trilobite
Artiopoda have been successfully classified into relatively
well-defined groups, which have also been supported by
phylogenetic analyses (e.g. Xandarellida, Nektaspida, Con-
ciliterga; see Hou and Bergstr€om 1997; Paterson et al.
2012; Ortega-Hern�andez et al. 2013). However, a few taxa
are notorious for their unusual morphologies, which
complicate their classification and precise phylogenetic
affinities (e.g. Habelia optata Walcott, 1912; Phytophilaspis
pergamena Ivanstov, 1999), or which occupy an unspeci-
fied basal position relative to better defined groups (e.g.
Retifacies abnormalis Hou et al. 1989; Squamacula clypeata
Hou and Bergstr€om 1997; see Paterson et al. 2012,
Ortega-Hern�andez et al. 2013, Stein et al. 2013). Here, we
describe a new weakly biomineralized artiopodan from
the late Guzhangian (Cambrian Series 3) Weeks Forma-
tion of Utah (USA), which combines a mosaic of mor-
phological characteristics as-yet observed in several
artiopodan groups. The new taxon contributes towards
the increasing knowledge on the diversity of late Cam-
brian non-trilobite arthropods as informed by the Weeks
Formation Konservat-Lagerst€atte.
GEOLOGICAL SETTING
The Weeks Formation consists of a 300-m-thick sequence
of thin-bedded lime mudstones, wackestones and grain-
stones with variable amounts of shale. It crops out in
the North Canyon and neighbouring areas only, in the
© The Palaeontological Association doi: 10.1111/pala.12136 265
[Palaeontology, Vol. 58, Part 2, 2015, pp. 265–276]

vicinity of Notch Peak (Central House Range, Utah,
USA). The Weeks Formation conformably overlies the
shales and argillaceous carbonates of the Marjum For-
mation, which represents relatively deep-water deposits,
and it is conformably overlain by the shallow water car-
bonates of the Orr Formation. The upper part of the
Weeks Formation actually records a substantial trans-
gressive succession to a shallower depositional environ-
ment (Beebe 1990); it marks the end of the House
Range embayment, a fault-controlled trough that devel-
oped within the carbonate platform during Cambrian
Stage 5, as a locus for deep-water sedimentation (Miller
et al. 2012). This upper part of the Weeks Formation
has yielded a rather diverse biomineralized fauna, domi-
nated by trilobites (Robison and Babcock 2011; Lerosey-
Aubril et al. 2012) and inarticulate brachiopods, as well
as exceptionally preserved fossils. These ‘soft-bodied’ or
weakly biomineralized organisms are predominantly
represented by arthropods (Lerosey-Aubril et al. 2013a,
2014; Lerosey-Aubril 2014) and worms (e.g. palaeosco-
lecids), although sponges and a possible ctenophore (i.e.
comb jelly) also occur. Trilobites (e.g. Cedaria minor
Walcott, 1916) indicate that the upper part of the
Weeks Formation was deposited in an open-shelf marine
environment during the late Guzhangian (i.e. late Cam-
brian Epoch 3, formerly early late Cambrian; Robison
and Babcock 2011). A more detailed account of the
structural and environmental contexts of the deposition
of the Weeks Formation is given in Lerosey-Aubril et al.
(2014).
MATERIAL AND METHODS
The studied material consists of the part and counter-
part (BPM 1022a and BPM 1022b, respectively) of a
single individual preserved as a dorsoventral compres-
sion in lime mudstone. Photographs of the dry speci-
mens were taken with a Nikon D3X digital camera
equipped with a Micro-Nikkor AF 60 mm f/2.8 D
macro lens, using low-angle cross-polarized light. The
preservation of the specimens was explored using SEM
(JEOL 310 JSM-6490LV and FEI Verios 460) equipped
with energy dispersive X-ray (EDX) modules (EDAX).
This material is housed at the Back to the Past
Museum (Canc�un, Mexico; BPM). Photographs of a
second specimen, unfortunately held in a private collec-
tion, were also examined. This specimen is fundamen-
tally identical to BPM 1022, except for the presence of
a long tailspine distinguished by a distal bifurcation.
Although this feature is not visible on the material
described herein due to the incomplete preservation of
the posterior part of the body, this aspect of the mor-
phology is briefly mentioned in the description of the
new taxon.
Terminology
The shape of the posterior margin of the cephalon is
described from sagittal axis to genal angle, using the
terms ‘procurved’ and ‘opisthocurved’ when it curved an-
terolaterally and posterolaterally, respectively (Dunlop
and Selden 1997; Lerosey-Aubril et al. 2013b; Lerosey-
Aubril 2014). Abbreviations: exs., exsagittal; sag., sagittal;
tr., transverse; T1–11, trunk tergites 1 to 11.
Preservation
The part (BPM 1022a) and counterpart (BPM 1022b) of
the specimen represent the dorsal exoskeleton of an
almost complete individual associated with possible
remains of internal organs and appendages (Figs 1, 2).
Both parts are almost flat, but the pattern of medial over-
lap of adjacent tergites suggests that the dorsal exoskele-
ton is exposed from the visceral side (i.e. upside-down
position). The specimen is mostly preserved as internal
(BPM 1022a?) or external (BPM 1022b?) moulds of the
dorsal exoskeleton, which exhibit a dark grey to red col-
ouration. The moulds are still partially covered by a thin
layer of light pink material in the medial region of the
F IG . 1 . Falcatamacaris bellua gen. et sp. nov. from the upper part of the Weeks Formation, late Guzhangian (Cedaria Zone), House
Range, Utah, USA. A–I, holotype (BPM 1022a, b), part and counterpart of the dorsal exoskeleton of an almost complete individual
associated with possible remains of internal organs and appendages; photographs of the dry specimens under cross-polarized light with
anterior end facing to the top. A, C–F, I, part (BPM 1022a); A, general view; C, three-dimensional patches of calcium carbonate (black
arrowheads) and iron oxides (white arrowheads) located medially under posterior part of cephalon and trunk tergites T1–3; thesematerials possibly represent the remains of digestive structures; D, three-dimensional patches of calcium phosphate located medially
under trunk tergites T6 and T7; E, left pleural regions of trunk tergites T1 and T2 showing probable remains of cuticle composed of
calcium carbonate; F, detail of the cephalic region showing possible remains of appendages (ap?); I, detail of the posterior trunk
region; note the particular morphology of trunk tergites T9–11, with their abaxial portions strongly curved backwards, mostly consist-
ing of pleural spines. B, G, H, counterpart (BPM 1022b); B, general view; G, detail of the cephalic region showing possible remains of
appendages (ap?); H, detail of trunk tergites T4–6 (right halves), note their straight transverse outline and hook-shaped pleural spines.
Scale bars represent 5 mm (C–E) and 1 cm (A, B, F–I). (Colour online)
266 PALAEONTOLOGY , VOLUME 58

A B C
D
E
F H
I
G
ORTEGA-HERN �ANDEZ ET AL . : NON-TR ILOB ITE ARTIOPODAN FROM THE WEEKS FORMATION 267
body (especially in BPM 1022b) that we interpret as
cuticular remains (Fig. 1E). EDX analyses reveal that this
material is predominantly composed of C, O and Ca
(with trace amounts of Si and Al), and thus probably
represents calcium carbonate. These results are surpris-
ing, for previous EDX analyses of the exoskeletons of
non-trilobite arthropods from the Weeks Formation
have shown that they are preserved as iron oxides,
sometimes associated with a cover of an undetermined
material rich in O, Si, Al and Mg (Lerosey-Aubril et al.
2014). The only known exception to date is the phos-
phatic cuticle of the aglaspidid-like arthropod Beckwithia
typa Resser, 1931, and the aglaspidid Tremaglaspis
vanroyi Lerosey-Aubril, Ortega-Hern�andez et al., 2013a;
however, it is likely that this biomineralization repre-
sents a particularity of Aglaspidida sensu lato (Lerosey-
Aubril et al. 2013a, b). Furthermore, the trilobites from
the Weeks Formation mostly retain their original calcitic
composition, although many specimens show traces of
diagenetic silicification (Adrain et al. 2009). Considering
these observations, we believe that the new arthropod
probably possessed a lightly biomineralized (calcitic?)
dorsal exoskeleton.
Three-dimensional patches of yellowish, dark blue and
dark red materials present in the medial region of
the body may represent remains of internal organs
A B
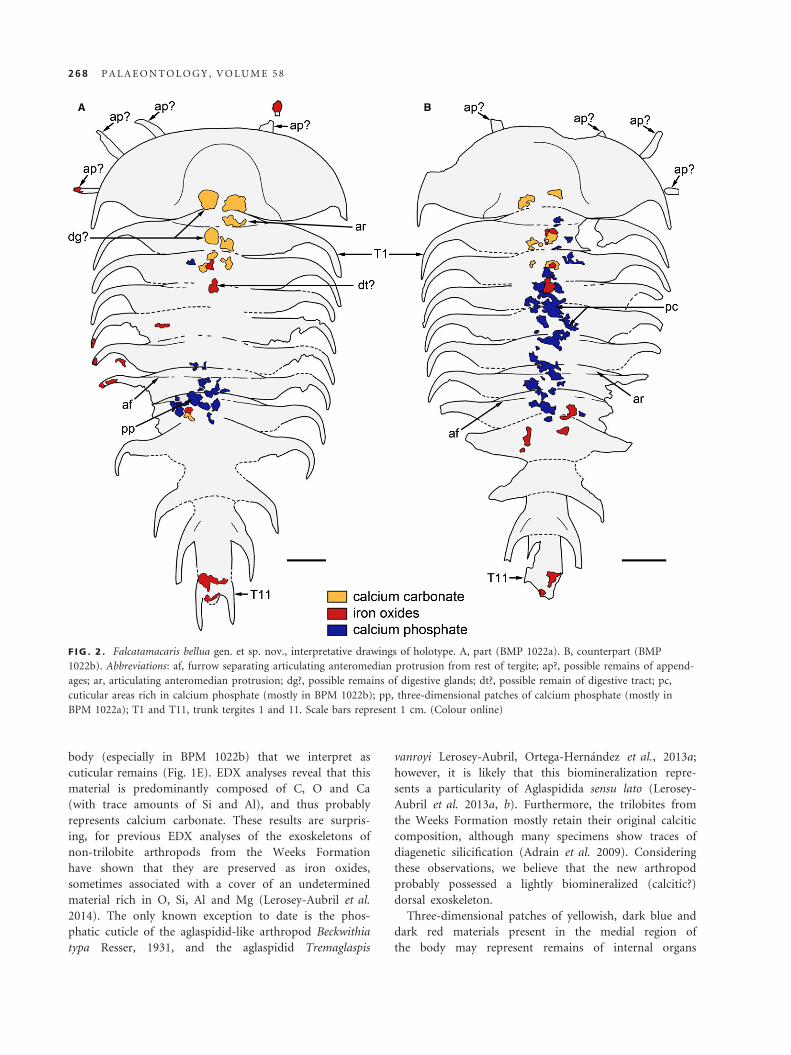
F IG . 2 . Falcatamacaris bellua gen. et sp. nov., interpretative drawings of holotype. A, part (BMP 1022a). B, counterpart (BMP
1022b). Abbreviations: af, furrow separating articulating anteromedian protrusion from rest of tergite; ap?, possible remains of append-
ages; ar, articulating anteromedian protrusion; dg?, possible remains of digestive glands; dt?, possible remain of digestive tract; pc,
cuticular areas rich in calcium phosphate (mostly in BPM 1022b); pp, three-dimensional patches of calcium phosphate (mostly in
BPM 1022a); T1 and T11, trunk tergites 1 and 11. Scale bars represent 1 cm. (Colour online)
268 PALAEONTOLOGY , VOLUME 58

(Figs 1A–D, 2A–B). EDX analyses suggest that the yellow-
ish (high peaks of C, O and Ca), dark blue (high peaks of
C, O, P and Ca; minor peaks of Si, F and Al) and the
dark red (strong peaks of C, O and Fe; minor peak of Si)
are mostly composed of calcium carbonate, calcium phos-
phate and iron oxides, respectively. The distribution and
significance of these three-dimensional patches are
addressed below. On BPM 1022b (Figs 1B, 2B), the puta-
tive cuticle displays large areas with a dark blue (rather
than light pink) colouration medially, suggestive of a local
enrichment in calcium phosphate (confirmed by EDX
analyses). The change in colour is gradual, and we believe
that this local phosphatization of the cuticle is secondary
and probably related to the early diagenetic phosphatiza-
tion of internal organs, some of which are now repre-
sented by the three-dimensional patches of calcium
phosphate. Lastly, the matrix is predominantly composed
of C, O, Si and Al (with trace amounts of K, Ca and Mg)
according to EDX analyses, and therefore, it is easily
differentiated from the different materials composing the
fossil.
SYSTEMATIC PALAEONTOLOGY
This published work and the nomenclatural acts it con-
tains have been registered in Zoobank: http://zoobank.
org/References/C27275AC-ED4D-43E0-AE49-75C19FB32C95
Phylum ARTHROPODA von Siebold, 1848
Class ARTIOPODA Hou and Bergstr€om, 1997
Remarks. The new taxon is tentatively assigned to the
Artiopoda based on the presence of a broad cephalon
with genal spines and trunk tergites with well-developed
pleurae and pleural spines, which give it a characteristic
dorsoventrally flattened trilobite-like appearance (see Hou
and Bergstr€om 1997, p. 43). Stein and Selden (2012)
recently proposed a more stringent diagnosis that uses
appendicular characteristics as definitive indicators of art-
iopodan affinities; furthermore, they emphasized that
expanded pleurae in the trunk region occur in several
non-artiopodan groups of Palaeozoic arthropods that
occupy diverse phylogenetic positions (e.g. fuxianhuiids,
Hou and Bergstr€om 1997; Yang et al. 2013; leanchoiliids,
Edgecombe et al. 2011; Haug et al. 2012) and should
therefore be regarded as a plesiomorphic feature.
Although we agree with this definition of Artiopoda,
and despite the paucity of appendicular data, we believe
that the combination of characters exhibited by the
new arthropod from the Weeks Formation supports its
phylogenetic position within this group (see detailed
comparisons below).
Genus FALCATAMACARIS gen. nov.
LSID. urn:lsid:zoobank.org:act:4F40D0F6-C285-48A8-8365-
AF67B7261968
Type species. Falcatamacaris bellua sp. nov. (by monotypy).
Derivation of name. From the Latin ‘falcatam’, meaning sickle
shaped, in reference to the morphology of the genal and pleural
spines, and ‘caris’, meaning crab, a suffix commonly used for
marine arthropods.
Diagnosis. Large and weakly biomineralized arthropod
exhibiting the following unique combination of charac-
ters: cephalon wide (tr.), with weakly defined glabellar
region, genal spines inserting laterally, abaxially procurved
posterior margin, and apparently no dorsal eyes or ecdy-
sial sutures; 11 trunk tergites, T1–8 with edge-to-edge
articulations and sickle-shaped pleural spines inserting an-
terolaterally, T9–11 characterized by an abrupt change in
tapering and reduction of the pleural regions into long,
backward-facing pleural spines.
Falcatamacaris bellua sp. nov.
Figures 1–3
LSID. urn:lsid:zoobank.org:act:AC5499E1-B907-453F-A793-
72C44451F59D
Derivation of name. From the latin ‘bellua’, meaning beast or
large animal.
Diagnosis. As for the genus.
Description. Holotype (BPM 1022a, b) consists of a dorsoven-
trally flattened articulated individual with a total length of
126 mm (Fig. 1A–B). Cephalon short (c. 20% total body length;
sag.), but wide (tr.), with length (sag.)/maximum width (tr.) ratio
of c. 0.5. Anterior margin broadly elliptical (Fig. 1F–G). Genalspines insert laterally, approximately at mid-length (exs.) of ceph-
alon, and project posteriorly slightly beyond posterior margin of
head. Posterolateral margin of cephalon possibly opisthocurved
medially, but procurved abaxially, except on a short distance close
to insertion sites of genal spines where it runs laterally only. Sur-
face of cephalon even and apparently devoid of any discrete fea-
tures, such as eyes, furrows or ecdysial sutures. Presence of an
axial region suggested by dark stained area medially, which seems
bilaterally symmetrical (Figs 1F, 2A); this putative axial region is
tongue shaped on its anterior half, but strongly widens (tr.) pos-
teriorly (from approximately a third to a half of maximum cepha-
lic width), and it is separated from anterior cephalic margin by a
distance equivalent to c. 16.5% of cephalic length (sag.). There is
no evidence for the presence of a hypostome on the ventral side.
Trunk composed of 11 tergites (Figs 1A–B, 2A–B). T1 rela-
tively long (approximately a third of sagittal length of ceph-
ORTEGA-HERN �ANDEZ ET AL . : NON-TR ILOB ITE ARTIOPODAN FROM THE WEEKS FORMATION 269
alon), as wide (tr.) as cephalon, medially protruding above
cephalon, but transverse abaxially; hook-shaped pleural spines,
projecting posteriorly from anterior half of tergite up to mid-
length (exs.) of T2; posterior margin concealed under T2
medially, but otherwise transverse, except for most abaxial portions
that run anterolaterally to form broad notches with posterior
edges of pleural spines. T2–8 essentially similar to T1 (Fig. 1H),
except for moderate and progressive decrease in width (tr.;
c. 75% of maximum width of cephalon only in T8) and gradual
increase in degree of curvature of pleural spines rearwards
(more pronounced sickle shape; Figs 1A–B, 2A–B). T4–8 proba-
bly slightly displaced backwards during flattening of specimen,
resulting in gaps between abaxial parts of tergites; however,
articulations between tergites probably similar from T1 to T8,
consisting of partial overlap of one tergite by an anteromedian
protrusion of the tergite immediately following it (when
observed from the visceral side) and edge-to-edge contacts abax-
ially, as illustrated by T1–T3. T8 slightly differs from more ante-
rior tergites by its longer (sag.) and subtriangular anteromedian
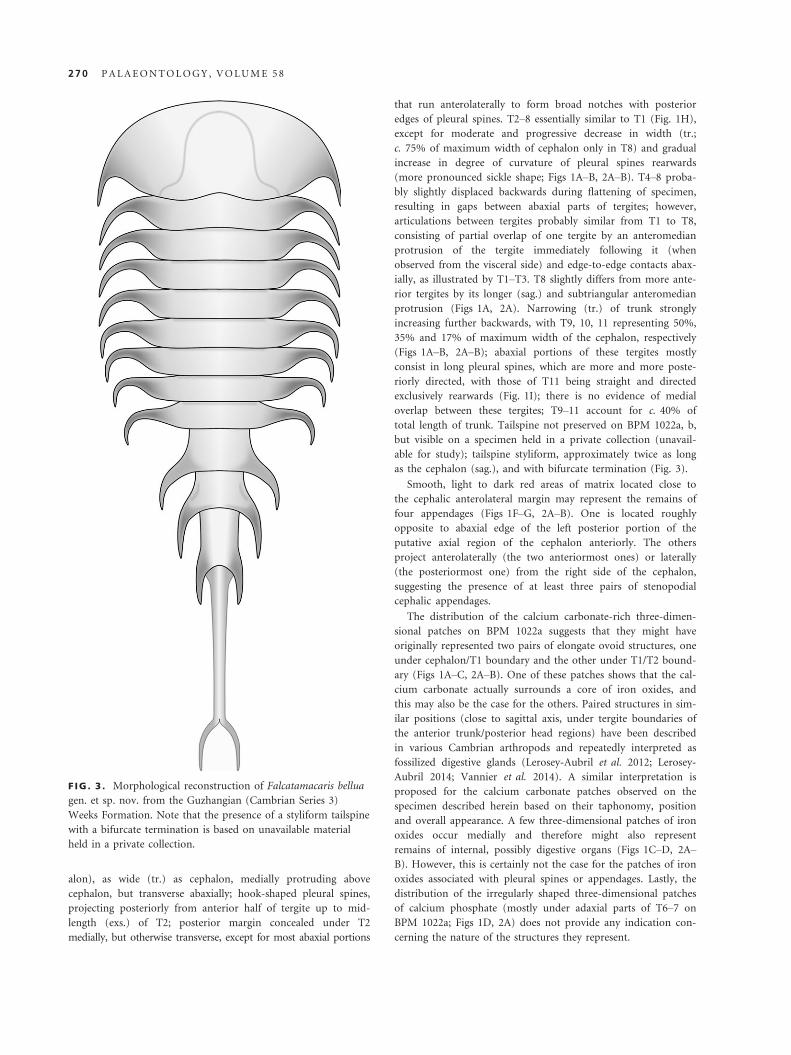
protrusion (Figs 1A, 2A). Narrowing (tr.) of trunk strongly
increasing further backwards, with T9, 10, 11 representing 50%,
35% and 17% of maximum width of the cephalon, respectively
(Figs 1A–B, 2A–B); abaxial portions of these tergites mostly
consist in long pleural spines, which are more and more poste-
riorly directed, with those of T11 being straight and directed
exclusively rearwards (Fig. 1I); there is no evidence of medial
overlap between these tergites; T9–11 account for c. 40% of
total length of trunk. Tailspine not preserved on BPM 1022a, b,
but visible on a specimen held in a private collection (unavail-
able for study); tailspine styliform, approximately twice as long
as the cephalon (sag.), and with bifurcate termination (Fig. 3).
Smooth, light to dark red areas of matrix located close to
the cephalic anterolateral margin may represent the remains of
four appendages (Figs 1F–G, 2A–B). One is located roughly
opposite to abaxial edge of the left posterior portion of the
putative axial region of the cephalon anteriorly. The others
project anterolaterally (the two anteriormost ones) or laterally
(the posteriormost one) from the right side of the cephalon,
suggesting the presence of at least three pairs of stenopodial
cephalic appendages.
The distribution of the calcium carbonate-rich three-dimen-
sional patches on BPM 1022a suggests that they might have
originally represented two pairs of elongate ovoid structures, one
under cephalon/T1 boundary and the other under T1/T2 bound-
ary (Figs 1A–C, 2A–B). One of these patches shows that the cal-
cium carbonate actually surrounds a core of iron oxides, and
this may also be the case for the others. Paired structures in sim-
ilar positions (close to sagittal axis, under tergite boundaries of
the anterior trunk/posterior head regions) have been described
in various Cambrian arthropods and repeatedly interpreted as
fossilized digestive glands (Lerosey-Aubril et al. 2012; Lerosey-
Aubril 2014; Vannier et al. 2014). A similar interpretation is
proposed for the calcium carbonate patches observed on the
specimen described herein based on their taphonomy, position
and overall appearance. A few three-dimensional patches of iron
oxides occur medially and therefore might also represent
remains of internal, possibly digestive organs (Figs 1C–D, 2A–B). However, this is certainly not the case for the patches of iron
oxides associated with pleural spines or appendages. Lastly, the
distribution of the irregularly shaped three-dimensional patches
of calcium phosphate (mostly under adaxial parts of T6–7 on
BPM 1022a; Figs 1D, 2A) does not provide any indication con-
cerning the nature of the structures they represent.
F IG . 3 . Morphological reconstruction of Falcatamacaris bellua
gen. et sp. nov. from the Guzhangian (Cambrian Series 3)
Weeks Formation. Note that the presence of a styliform tailspine
with a bifurcate termination is based on unavailable material
held in a private collection.
270 PALAEONTOLOGY , VOLUME 58

Morphological comparisons with Palaeozoic Arthropoda
Cephalic region. A wide (tr.) cephalon with a broadly
elliptical anterolateral margin is a common feature within
Artiopoda (e.g. most aglaspidids, trilobitomorphs and
trilobites; Edgecombe and Ramsk€old 1999), but is also
found in more phylogenetically distant groups (e.g.
xiphosurids; Lamsdell 2013). The overall cephalic shape
of Falcatamacaris gen. nov., however, differs from most
Cambrian arthropods in the presence of an abaxially pro-
curved posterolateral cephalic margin (Fig. 3). Within
Artiopoda, a similar character is also present in represen-
tatives of Conciliterga with freely articulating tergites (e.g.
Helmetia, Kuamaia; see Hou and Bergstr€om 1997; Pater-
son et al. 2012), the aglaspidid-like arthropod Kwanyina-
spis (Zhang and Shu 2005), and also members of the
post-Cambrian group Cheloniellida (Dunlop and Selden
1997). In all these taxa, however, the anteriormost trunk
tergites are anteriorly reflexed abaxially (see Ortega-
Hern�andez et al. 2013, character 50), while in Falcatamac-
aris gen. nov., they have a straight transverse outline;
moreover, almost all of these artiopodans have either
reduced genal spines or these structures are entirely
absent. Only the concilitergan Rhombicalvaria exhibits
well-developed genal spines (Hou 1987), which insert lat-
erally on the cephalon, similar to the new genus. A some-
what comparable condition can also be observed in
various trilobite orders (Redlichiida, e.g. Whittington
et al. 1997; Ptychopariida, e.g. Clarkson et al. 2004;
Odontopleurida, e.g. Whittington and Bohlin 1958). A
differentiated axial region on the cephalon is a widespread
feature among Palaeozoic arthropods (see Ortega-Hern�an-
dez et al. 2013, character 35), as best exemplified by the
glabella of trilobites. However, the weak expression of this
axial region in Falcatamacaris gen. nov. indicates that it
was not delimited by furrows as in trilobites, and unlike
the subtriangular glabellar region of many aglaspidids
(Hesselbo 1992; Ortega-Hern�andez et al. 2010), it is ton-
gue-shaped anteriorly. The lack of dorsal eyes in BPM
1022 offers two possibilities: either these were ventral (e.g.
Nektaspida, Conciliterga, Leanchoiliidae) and not pre-
served in this specimen, or they were genuinely absent/
secondarily lost (e.g. Squamacula, Tremaglaspis). Without
additional data on the ventral morphology of Falcatamac-
aris gen. nov., it is not possible to decide between these
hypotheses. However, it should be noted that there are no
raised bulges that could accommodate a pair of ventral
eyes on the cephalon of Falcatamacaris gen. nov., as it is
typically observed in concilitergans (Hou and Bergstr€om
1997; Edgecombe and Ramsk€old 1999; Paterson et al.
2012). Finally, the absence of dorsal ecdysial sutures is
shared with all non-trilobite Palaeozoic arthropods
(Cotton and Braddy 2004, characters 28 and 29).
‘Anterior’ trunk region. A trunk composed of 11 freely
articulating tergites has been described in various Palaeo-
zoic arthropods. Among non-artiopodans, for instance,
this feature characterizes the megacheiran family Leanch-
oiliidae (Edgecombe et al. 2011; Haug et al. 2012) and
has even been considered as the probable ground pattern
of Xiphosura within Euchelicerata (Lamsdell 2013). In
Artiopoda, however, the presence of 11 trunk tergites is
more typical of the Aglaspidida sensu stricto (Van Roy
2006; Ortega-Hern�andez et al. 2013), being observed in
all but one species of this group (Fortey and Rushton
2009; see also Lerosey-Aubril et al. 2013a). Eleven free
trunk tergites are also found in the artiopodan Squamacula
buckorum Paterson, Garc�ıa-Bellido and Edgecombe, 2012,
and in the problematic Cambrian arthropod Nettapezoura
basilikos Briggs, Lieberman et al., 2008; however, unlike
Falcatamacaris gen. nov. and many aglaspidids, the latter
two species do not possess well-developed pleural spines.
The sickle-shaped pleural spines of the anterior tergites
(T1–8) in Falcatamacaris gen. nov. are particularly dis-
tinctive in that they project from the anterior portion of
each tergite (Fig. 3). This contrasts with the condition
observed in other artiopodans, such as trilobites, aglaspid-
ids or concilitergans (e.g. Rhombicalvaria Hou, 1987),
where the pleural spines normally extend either from the
entire pleurae or from their posterior portions. To our
knowledge, only a few trilobites (e.g. some Ctenopyge;
Clarkson et al. 2004, figs 17A, 19) exhibit a type of inser-
tion of pleural spines approximating that observed in the
new genus. Tergites of the anterior trunk of Falcatamac-
aris gen. nov. are provided with an anterior medial pro-
trusion apparently separated by a transverse furrow from
the main part of the tergite, which recalls the articulating
half-ring of trilobites. Articulation between two of these
tergites involves the overlap of this structure by the ter-
gite in front, while little- to no-overlap between tergites
occurs abaxially (Fig. 3). This organization deviates from
the typically wide tergite overlap found in most Palaeozo-
ic arthropods, but closely resembles the ‘edge-to-edge’
articulation defined by Edgecombe and Ramsk€old (1999,
character 18), which is primarily observed in the trunk of
concilitergans and trilobites, and to a lesser degree also in
some aglaspidids and aglaspidid-like arthropods (see Ort-
ega-Hern�andez et al. 2013, character 44). Among non-art-
iopodan taxa, a similar anterior extension of the medial
region of the trunk tergites is present in the synziphosu-
rine Willwerathia laticeps Størmer, 1969 (see Anderson
et al. 1998), and further developed into a pseudo-half
ring in the opisthosomal articulation of Xiphosura (see
Lamsdell 2013, fig. 9).
‘Posterior’ trunk region. The posterior exoskeletal mor-
phology of Falcatamacaris gen. nov. is uncommon among
ORTEGA-HERN �ANDEZ ET AL . : NON-TR ILOB ITE ARTIOPODAN FROM THE WEEKS FORMATION 271

Palaeozoic arthropods; the three posteriormost trunk
tergites (T9–T11) differ significantly from the anterior
ones, being narrower (tr.), longer (sag.) and having ‘pleu-
ral’ regions mostly consist of elongated spines (Fig. 3).
The absence of a pygidium (i.e. an ensemble of fused pos-
terior trunk tergites) is a widespread character within Art-
iopoda; however, in most species, the posterior trunk
shows a gradual change in the morphology of tergites,
which become narrower (tr.) and increasingly curved
rearwards (Hou and Bergstr€om 1997; Edgecombe and
Ramsk€old 1999). The notable and rather abrupt change
in the posterior tergite morphology of Falcatamacaris gen.
nov. has no direct equivalent among Artiopoda and prob-
ably represents a derived condition. There are, however,
some Cambrian arthropods that seem to show a superfi-
cial similarity to the posterior differentiation of Falcata-
macaris gen. nov. For example, the last three trunk
tergites in N. basilikos and Dicranocaris guntherorum
Briggs et al., 2008, are significantly narrower (tr.) than
the anterior ones (Briggs et al. 2008; Hendricks and
Lieberman 2008); the last trunk tergite of D. guntherorum
also bears a pair of well-developed posterior spines
(Briggs et al. 2008, fig. 5), as in Falcatamacaris gen. nov.
The xenopod Sidneyia inexpectans Walcott, 1911, also fea-
tures a discrete anteroposterior trunk differentiation; this
consists of anterior tergites with well-developed pleural
regions and much narrower (tr.) and cylindrical posterior
tergites (see discussion in Ortega-Hern�andez et al. 2013,
character 60). The recently described arthropod Notchia
weugi Lerosey-Aubril, 2014, is also typified by a morpho-
logically distinct posterior trunk region composed of four
tergites that are straight (tr.), rather than abaxially curved
backwards like the anteriormost ones.
Outside of Artiopoda, the peculiar morphology of the
trunk of Falcatamacaris gen. nov. recalls that of the syn-
ziphosurine W. laticeps (see Anderson et al. 1998, fig. 8).
In both taxa, the anterior trunk tergites are characterized
by straight transverse outlines, while the three posterior-
most ones are increasingly curved backwards. However,
the trunk of W. laticeps is composed of ten tergites only,
including a reduced anteriormost one, and its cephalic
shield differs significantly from that of the new genus.
The presence of a long tailspine is particularly common
among Palaeozoic arthropods (e.g. aglaspidids, Emeraldella,
xiphosurids; see Ortega-Hern�andez et al. 2013, character
70); however, a bifurcate termination is only known
in the aglaspidid Glypharthrus simplex (Raasch, 1939;
Hesselbo 1992, figs 11.1, 11.4) and the problematic
artiopodan Habelia optata (Whittington 1981, fig. 61).
Internal structures. The putative paired midgut glands in
Falcatamacaris gen. nov. draw comparisons with similar
structures in various Palaeozoic arthropods. Although the
preservation of BMP 1022 does not allow the fine details
of the morphology to be observed (e.g. radiating ridges
and grooves; Butterfield 2002), the paired arrangement of
the midgut glands is similar to that observed in various
trilobite species (Lerosey-Aubril et al. 2011, 2012; Fatka
et al. 2013) and non-trilobite artiopodans (e.g. Chen
et al. 1997; Lagebro et al. 2009; Paterson et al. 2012), par-
ticularly in their position under cephalon/T1 and T1/T2
articulations (Ortega-Hern�andez and Brena 2012).
DISCUSSION
Phylogenetic affinities
The affinities of Falcatamacaris gen. nov. are problematic
due to the scarcity of clear synapomorphic characters
shared with particular clades within Artiopoda (Fig. 4).
However, given the absence of fused body regions into
distinct tagmata (or at least pseudotagmata) and lack of
discrete cephalic structures, it is possible to reject close
affinities with phylogenetically derived groups of Palaeo-
zoic arthropods, such as euchelicerates. The overall mor-
phology of Falcatamacaris gen. nov. may suggest an
association with the Aglaspidida sensu lato (cf. Van Roy
2006), a poorly constrained group composed of the
Aglaspidida sensu stricto and a number of closely related
aglaspidid-like arthropods (Ortega-Hern�andez et al. 2013).
The lack of diagnostic aglaspidid features in Falcatam-
acaris gen. nov. (i.e. postventral plates, anterior tergal
processes), however, rule out any affinities with aglaspid-
ids sensu stricto. A position close to trilobitomorph
arthropods cannot be entirely discarded either, as
Falcatamacaris gen. nov. shares various characters (e.g.
advanced genal spines, pleural spinosity, tergite articula-
tion) with the concilitergan Rhombicalvaria (Hou 1987;
see also Edgecombe and Ramsk€old 1999). Despite the lack
of a fused pygidium and anteriorly flexed trunk tergites,
Falcatamacaris gen. nov. could potentially occupy a posi-
tion within Conciliterga, as the patterns of trunk tagmosis
can be extremely variable even within monophyletic trilo-
bitomorph clades (e.g. Australimicola in Conciliterga, see
Paterson et al. 2012; Luohuilinella in Xandarellida, see
Zhang et al. 2012). A third possibility is that Falcatam-
acaris gen. nov. may occupy an unresolved basal position
within Artiopoda as a whole, similar to other problematic
representatives such as Retifacies and Squamacula (see
Paterson et al. 2012; Ortega-Hern�andez et al. 2013; Stein
et al. 2013). The evolutionary significance of the position
of the gut diverticulae under the cephalon/T1 and T1/T2
articulations in the new arthropod is somewhat inconclu-
sive. Ortega-Hern�andez and Brena (2012) argued that
such organization in trilobites and nektaspidids is indica-
tive of a derived mode of trunk segmentation within Artio-
poda. However, a recent study by Vannier et al. (2014)
272 PALAEONTOLOGY , VOLUME 58

clearly showed that the gut diverticulae of lobopodians in
the stem-lineage of Euarthropoda (e.g. Jianshanopodia,
Pambdelurion) also occupies a similar inter-segmental
position that could suggest a plesiomorphic state. This
issue remains unresolved until more information on the
digestive structures of extinct Panarthropoda becomes
available. Finally, the weakly biomineralized cuticle of
Falcatamacaris gen. nov. is uncommon for non-trilobite
artiopodans and carries an uncertain phylogenetic signal.
The phosphatic cuticle of some aglaspidids (Raasch 1939;
Briggs and Fortey 1982; Hesselbo 1992; Lerosey-Aubril
et al. 2013a) or aglaspidid-like arthropods (Waggoner
2003) is arguably the best-known example of biomineral-
ization outside of Trilobita; a similar exoskeletal composi-
tion, however, is also known in the enigmatic arthropod
Phytophilaspis from the lower Cambrian of Siberia
(Ivanstov 1999; Lin et al. 2011). To our knowledge, the
only report of a calcitic cuticle in a non-trilobite artiopo-
dan concerns an undescribed aglaspidid from the Ordovi-
cian of Shanxi Province in China (Fortey and Theron
1994; Ortega-Hern�andez et al. 2013), even though no
direct evidence supporting this claim was provided
(Lerosey-Aubril et al. 2013a).
The non-trilobite arthropod fauna from the Weeks
Formation
The Weeks Formation has yielded a particularly diverse
trilobite fauna composed of more than 35 species, most
of which occur in the upper part of the formation where
exceptionally preserved fossils are found. Examination of
some 150 specimens of non-trilobite arthropod specimens
from the Weeks Formation suggests that they may also
constitute a rather diverse assemblage (c. 15 species).
However, only six species have been described to date:
Beckwithia typa Resser, 1931 (see also Raasch 1939;
Hesselbo 1989), Anabarochilina australis (Hinz-Schallre-
uter, 1993; see also Siveter and Williams 1997), Trema-
glaspis vanroyi Lerosey-Aubril, Ortega-Hern�andez et al.,
2013a, Anomalocaris aff. canadensis (Lerosey-Aubril et al.
2014), Notchia weugi Lerosey-Aubril, 2014 and now
F IG . 4 . Possible phylogenetic positions of Falcatamacaris bellua gen. et sp. nov. within Artiopoda; topology sensu Ortega-Hern�andez
et al. 2013, fig. 6b.
ORTEGA-HERN �ANDEZ ET AL . : NON-TR ILOB ITE ARTIOPODAN FROM THE WEEKS FORMATION 273

Falcatamacaris bellua gen. et sp. nov. All of these discov-
eries have resulted in new information about the palaeo-
geographical and stratigraphical distribution of these taxa.
For instance, the bradoriid A. australis is mostly known
from the lower Guzhangian, but its stratigraphical range
extents from the Cambrian Stage 5 to the upper Guzhan-
gian (the youngest occurrence being in the Weeks
Formation; Collette et al. 2011). This arthropod had a
worldwide palaeogeographical distribution, although pos-
sibly restricted to low latitudes, which suggests a pelagic
lifestyle (Collette et al. 2011). Anomalocaris aff. canadensis
has been recently described from isolated frontal append-
ages, which all exhibited particularly small sizes (< 25 mm
in length) compared to previously described frontal
appendages of Anomalocaris species. According to Lero-
sey-Aubril et al. (2014), they belong to a new species of
Anomalocaris phylogenetically close to A. canadensis
Whiteaves, 1892. This latter species is only known with
certainty from the Cambrian Stage 4 (e.g. Kinzers Forma-
tion, Latham Shale) to the Cambrian stage 5 (e.g. Burgess
Shale Formation) of Laurentia, but the genus Anomalo-
caris extends from the Cambrian Stage 3 to the upper
Guzhangian (the youngest occurrence being in the Weeks
Formation), with known occurrences in China (Hou et al.
1995), Australia (Daley et al. 2013) and North America
(Daley and Edgecombe 2014). Until its discovery in the
Cambrian of Utah, the aglaspidid Tremaglaspis was
regarded as a typically Ordovician arthropod, having been
reported from the Tremadocian of Wales (Fortey and
Rushton 2003, 2009) and Morocco (Van Roy et al. 2010)
exclusively. The description of Tremaglaspis vanroyi in the
upper Weeks Formation has extended the stratigraphical
range of the genus back to the Guzhangian and its palaeo-
geographical distribution to Laurentia (Lerosey-Aubril
et al. 2013a). It also illustrated that the Weeks Formation
fauna might provide important insights into the transi-
tion between Cambrian and Palaeozoic Evolutionary
Faunas (Sepkoski 1981).
Unlike Anabarochilina, Anomalocaris and Tremaglaspis,
Beckwithia and Notchia have limited stratigraphical and
palaeogeographical ranges. Beckwithia typa is only
confidently known from the upper part of the Weeks
Formation, where it represents one of the most common
non-trilobite arthropods. Beckwithia? major (Graham,
1931), the only other representative of the genus, was
described from disarticulated and fragmentary sclerites
from the Eau Claire Formation in Wisconsin (Graham
1931; Raasch 1939). However, this material does not per-
mit to distinguish B.? major from B. typa (Hesselbo 1989)
and therefore more likely documents a second occurrence
of B. typa, especially because the Eau Claire Formation and
the Weeks Formation are subcontemporaneous (Babcock
et al. 2014). Beckwithia strongly resembles Aglaspidida
sensu stricto, but a recent phylogenetic analysis has revealed
that it is close to, but definitely outside this clade (Ortega-
Hern�andez et al. 2013). The affinities of Notchia are more
problematical. Only known from the upper Weeks Forma-
tion, this arthropod exhibits a short (sag.) cephalon and a
non-styliform tailspine and, in this regard, somewhat
resembles other Cambrian arthropods, such as Paleomerus,
Strabops or Sidneyia (Lerosey-Aubril 2014). However, care-
ful examination of the only available specimen revealed
many differences compared to these three taxa, which led
Lerosey-Aubril (2014) to question a close phylogenetic
relationship between Notchia and any of them. The
description of Falcatamacaris gen. nov. further exemplifies
the uniqueness of the non-trilobite arthropod fauna from
the Weeks Formation and how it could contribute to a
potentially significant reassessment of the disparity of these
organisms in the late Cambrian.
Acknowledgements. JOH is supported by a Research Fellowship
in Emmanuel College, University of Cambridge (UK). This is a
contribution of the ANR project RALI 197 ‘Rise of Animal Life
(Cambrian–Ordovician) – organization and tempo: evidence
from exceptionally preserved biota’. We thank Jeremy Skepper
(University of Cambridge) for assistance with elemental analysis
of the material and James Lamsdell (Yale University) for discus-
sion on Willwerathia laticeps.
Editor. Xi-Guang Zhang
REFERENCES
ADRAIN, J. M., PETERS, S. E. and WESTROP, S. R.
2009. The Marjuman trilobite Cedarina Lochman: thoracic
morphology, systematics, and new species from western Utah
and eastern Nevada, USA. Zootaxa, 2218, 35–58.ANDERSON, L. I., POSCHMANN, M. and BRAUCK-
MANN, C. 1998. On the Emsian (Lower Devonian)
arthropods of the Rhenish Slate Mountains: 2. The
synziphosurine Willwerathia. Pal€aontologische Zeitschrift, 72,
325–336.BABCOCK, L. E., BARANOSKI , M. T. and COOK, A. E.
2014. Cambrian (Guzhangian Stage) trilobites from Ohio,
USA, and modification of the Cedaria Zone as used in Laur-
entia. GFF, 136, 6–15.BEEBE, M. A. 1990. Trilobite faunas and depositional environ-
ments of the Weeks Formation (Cambrian), Utah. Unpublished
PhD Thesis. University of Kansas, Lawrence, KS, 103 pp.
BRIGGS, D. E. G. and FORTEY, R. A. 1982. The cuticle of
the aglaspidid arthropods, a red-herring in the early history of
the vertebrates. Lethaia, 15, 25–29.-LIEBERMAN, B. S., HENDRICKS, J. R., HALGE-
DAHL, S. L. and JARRARD, R. D. 2008. Soft-bodied
arthropods from the middle Cambrian of Utah. Journal of
Paleontology, 82, 238–254.BUTTERFIELD, N. J. 2002. Leanchoilia guts and the interpre-
tation of three-dimensional structures in Burgess Shale-type
fossils. Paleobiology, 28, 155–171.
274 PALAEONTOLOGY , VOLUME 58

CHEN, J. Y., EDGECOMBE, G. D. and RAMSK €OLD, L.
1997. Morphological and ecological disparity in naraoiids
(Arthropoda) from the Early Cambrian Chengjiang fauna,
China. Records of the Australian Museum, 49, 1–24.CLARKSON, E. N. K., AHLGREN, J. and TAYLOR, C.
2004. Ontogeny, structure and functional morphology of some
spiny Ctenopyge species (Trilobita) from the upper Cambrian
of V€asterg€otland, Sweden. Transactions of the Royal Society of
Edinburgh: Earth Sciences, 94, 115–143.COLLETTE, J. H., HUGHES, N. C. and PENG, S. 2011.
The first report of a himalayan bradoriid arthropod and the
paleogeographic significance of this form. Journal of Paleontol-
ogy, 85, 76–82.COTTON, T. J. and BRADDY, S. J. 2004. The phylogeny of
arachnomorph arthropods and the origin of Chelicerata.
Transactions of the Royal Society of Edinburgh: Earth Sciences,
94 (for 2003), 169–193.DALEY, A. C. and EDGECOMBE, G. D. 2014. Morphology
of Anomalocaris canadensis from the Burgess Shale. Journal of
Paleontology, 88, 68–91.-PATERSON, J. R., EDGECOMBE, G. D., GARC�IA-
BELLIDO, D. C. and JAGO, J. B. 2013. New anatomical
information on Anomalocaris from the Cambrian Emu Bay
Shale of South Australia and a reassessment of its inferred
predatory habits. Palaeontology, 56, 971–990.DUNLOP, J. A. and SELDEN, P. A. 1997. The early his-
tory and phylogeny of the chelicerates. 221–235. In FOR-
TEY, R. A. and THOMAS, R. H. (eds). Arthropod
relationships. Systematics Association Special Volume Series,
55, 383 pp.
EDGECOMBE, G. D. and RAMSK €OLD, L. 1999. Relation-
ships of Cambrian Arachnata and the systematic position of
Trilobita. Journal of Paleontology, 73, 263–287.-GARC�IA-BELLIDO, D. C. and PATERSON, J. R.
2011. A new leanchoiliid megacheiran arthropod from the
lower Cambrian Emu Bay Shale, South Australia. Acta Palae-
ontologica Polonica, 56, 385–400.FATKA, O., LEROSEY-AUBRIL , R., BUDIL, P. and
RAK, �S. 2013. Fossilised guts in trilobites from the Upper
Letn�a Formation (Prague Basin, Czech Republic). Bulletin of
Geosciences, 88, 95–104.FORTEY, R. A. and RUSHTON, A. W. 2003. A new aglaspi-
did arthropod from the Lower Ordovician of Wales. Palaeon-
tology, 46, 1031–1038.--2009. The Ordovician aglaspidid arthropod Tremagla-
spis reconsidered. Memoirs of the Association of Australasian
Palaeontologists, 37, 17–23.-and THERON, J. N. 1994. A new Ordovician arthropod,
Soomaspis, and the agnostid problem. Palaeontology, 37, 841–861.
GRAHAM, W. A. P. 1931. A new crustacean of the family
Aglaspidae from the Upper Mississippi Valley. Ohio Journal of
Science, 31, 127–128.HAUG, J. T., BRIGGS, D. E. G. and HAUG, C. 2012. Mor-
phology and function in the Cambrian Burgess Shale megac-
heiran arthropod Leanchoilia superlata and the application of
a descriptive matrix. BMC Evolutionary Biology, 12, 162.
HENDRICKS, J. R. 2013. Global distributional dynamics of
Cambrian clades as revealed by Burgess Shale-type deposits.
Memoirs of the Geological Society of London, 38, 35–43.-and LIEBERMAN, B. S. 2008. New phylogenetic insights
into the Cambrian radiation of arachnomorph arthropods.
Journal of Paleontology, 82, 585–594.--and STIGALL A. L. 2008. Using GIS to study palaeo-
biogeographic and macroevolutionary patterns in soft-bodied
Cambrian arthropods. Palaeogeography, Palaeoclimatology,
Palaeoecology, 264, 163–175.HESSELBO, S. P. 1989. The aglaspidid arthropod Beckwithia
from the Cambrian of Utah and Wisconsin. Journal of Paleon-
tology, 63, 635–642.-1992. Aglaspidida (Arthropoda) from the upper Cambrian
of Wisconsin. Journal of Paleontology, 66, 885–923.HINZ-SCHALLREUTER, I. 1993. Cambrian ostracodes
mainly from Baltoscandia and Morocco. Archiv fur Gescheibek-
unde, 1, 385–448.HOU, X.-G. 1987. Three new large arthropods from lower
Cambrian, Chengjiaing, eastern Yunnan. Acta Palaeontologica
Sinica, 26, 272–285.-and BERGSTR €OM, J. 1997. Arthropods of the lower
Cambrian Chengjiang fauna, southwest China. Fossils and
Strata, 45, 1–116.-CHEN, J.-Y. and LU, H.-Z. 1989. Early Cambrian new
arthropods from Chengjiang, Yunnan. Acta Palaeontologica
Sinica, 28, 42–57.-BERGSTR €OM, J. and AHLBERG, P. 1995. Anomalocaris
and other large animals in the Lower Cambrian Chengjiang
Fauna of southwest China. GFF, 117, 163–183.IVANSTOV, A. Y. 1999. Trilobite-like arthropod from the
lower Cambrian of the Siberian Platform. Acta Palaeontologica
Polonica, 44, 455–466.LAGEBRO, L., STEIN, M. and PEEL, J. S. 2009. A new ?
lamellipedian arthropod from the early Cambrian Sirius Passet
fauna of North Greenland. Journal of Paleontology, 83, 820–825.
LAMSDELL, J. C. 2013. Revised systematics of Palaeozoic
‘horseshoe crabs’ and the myth of monophyletic Xiphosura.
Zoological Journal of the Linnean Society, 167, 1–27.LEROSEY-AUBRIL , R. 2014. Notchia weugi gen. et sp. nov.,
a new short-headed arthropod from the Weeks Formation
Konservat-Lagerst€atte (Cambrian; Utah). Geological Magazine,
published online 31 July 2014. doi: 10.1017/
S0016756814000375
-HEGNA, T. A. and OLIVE, S. 2011. Inferring internal
anatomy from the trilobite exoskeleton: the relationship
between frontal auxiliary impressions and the digestive system.
Lethaia, 44, 166–184.--KIER, C., BONINO, E., HABERSETZER, E.
and CARR�E, M. 2012. Controls on gut phosphatisation: the
trilobites from the Weeks Formation Lagerst€atte (Cambrian;
Utah). PLoS One, 7, e32934.
-ORTEGA-HERN �ANDEZ, J., KIER, C. and BONI-
NO, E. 2013a. Occurrence of the Ordovician-type aglaspidid
Tremaglaspis in the middle Cambrian Weeks Formation. Geo-
logical Magazine, 150, 945–951.
ORTEGA-HERN �ANDEZ ET AL . : NON-TR ILOB ITE ARTIOPODAN FROM THE WEEKS FORMATION 275

--and ZHU X.-J. 2013b. The first aglaspidid sensu
stricto from the Cambrian of China (Sandu Formation, Guan-
gxi). Geological Magazine, 150, 565–571.-HEGNA, T. A., BABCOCK, L. E., BONINO, E. and
KIER, C. 2014. Arthropod appendages from the Weeks For-
mation Konservat-Lagerst€atte: new occurrences of anomaloc-
aridids in the Cambrian of Utah, USA. Bulletin of Geosciences,
89, 269–282.LIN, J.-P., IVANSTOV, A. Y. and BRIGGS, D. E. G.
2011. The cuticle of the enigmatic arthropod Phytophilaspis
and biomineralization in Cambrian arthropods. Lethaia, 44,
344–349.MILLER, J. F., EVANS, K. R. and DATTILO, B. F. 2012.
The Great American Carbonate Bank in the miogeocline of
western central Utah: tectonic influences on sedimentation.
769–854. In DERBY, J. R., FRITZ, R., LONGACRE, S.
A., MORGAN, W. and STERNBACK, C. (eds). The great
American carbonate bank: the geology and economic resources of
the Cambro-Ordovician Sauk Sequence of Laurentia. American
Association of Petroleum Geologists Memoir, 98, 504 pp.
ORTEGA-HERN �ANDEZ, J. and BRENA, C. 2012. Ances-
tral patterning of tergite formation in a centipede suggests
derived mode of trunk segmentation in trilobites. PLoS One,
7, e52623.
-BRADDY, S. J., JAGO, J. B. and BAILLIE , P. W.
2010. A new aglaspidid arthropod from the upper Cambrian
of Tasmania. Palaeontology, 53, 1065–1076.-LEGG, D. A. and BRADDY, S. J. 2013. The phylogeny
of aglaspidid arthropods and the internal relationships within
Artiopoda. Cladistics, 29, 15–45.PATERSON, J. R., GARC�IA-BELLIDO, D. C. and
EDGECOMBE, G. D. 2012. New artiopodan arthropods
from the early Cambrian Emu Bay Shale Konservat-Lagerst€atte
of South Australia. Journal of Paleontology, 86, 340–357.RAASCH, G. O. 1939. Cambrian Merostomata. Geological Soci-
ety of America Special Paper, 16, 146 pp.
RESSER, C. E. 1931. A new Middle Cambrian merostome
crustacean. Proceedings of the United States National Museum,
79, article 33, 1–4.ROBISON, R. A. and BABCOCK, L. E. 2011. Systematics,
paleobiology, and taphonomy of some exceptionally preserved
trilobites from Cambrian Lagerst€atten of Utah. Paleontological
Contributions, 5, 1–47.SEPKOSKI , J. J. JR 1981. A factor analytical description of the
Phanerozoic marine fossil record. Paleobiology, 7, 36–53.S IEBOLD, C. T. VON. 1848. Lehrbuch der vergleichenden
Anatomie der Wirbellosen Thiere. In VON SIEBOLD, C. T.
and STANNIUS, H. (eds), Lehrbuch der vergleichenden
Anatomie. Verlag von Veit and Company, Berlin, 679 pp.
S IVETER, D. J. and WILLIAMS, M. 1997. Cambrian
bradoriid and phosphatocopid arthropods of North America.
Special Papers in Palaeontology, 57, 69 pp.
STEIN, M. and SELDEN, P. A. 2012. A restudy of the
Burgess Shale (Cambrian) arthropod Emeraldella brocki and reas-
sessment of its affinities. Journal of Systematic Palaeontology, 10,
361–383.
-BUDD, G. E., PEEL, J. S. and HARPER, D. A. T.
2013. Arthroaspis n. gen., a common element of the Sirius Pas-
set Lagerst€atte (Cambrian, North Greenland), sheds light on
trilobite ancestry. BMC Evolutionary Biology, 13, 99.
STØ RMER, L. 1969. Eurypterids from the Lower Devonian of
Willwerath, Eifel. Senckenbergiana Lethaea, 50, 21–35.VAN ROY, P. 2006. A new aglaspidid arthropod from the
Upper Ordovician of Morocco with remarks on the affinities
and limitations of Aglaspidida. Transactions of the Royal Soci-
ety of Edinburgh: Earth Sciences, 96, 327–350.-ORR, P., BOTTING, J., MUIR, L., VINTHER, J., LEF-
EBVRE, B., HARIRI , K. and BRIGGS, D. E. G. 2010.
Ordovician faunas of Burgess Shale type. Nature, 465, 215–218.VANNIER, J., L IU, J., LEROSEY-AUBRIL , R., VIN-
THER, J. and DALEY, A. C. 2014. Sophisticated digestive
systems in early arthropods. Nature Communications, 5, 3641.
WAGGONER, B. 2003. Non-trilobite arthropods from the Sil-
ver Peak Range, Nevada. Journal of Paleontology, 77, 706–720.WALCOTT, C. D. 1911. Cambrian geology and paleontology
II: Middle Cambrian Merostomata. Smithsonian Miscellaneous
Collections, 57, 17–40.-1912. Cambrian geology and paleontology II: Middle Cam-
brian Branchiopoda, Malacostraca, Trilobita and Merosto-
mata. Smithsonian Miscellaneous Collections, 57, 145–228.-1916. Cambrian geology and paleontology III, no. 5, Cam-
brian trilobites. Smithsonian Miscellaneous Collections, 64, 303–570.
WHITEAVES, J. F. 1892. Description of a new genus and spe-
cies of phyllocarid Crustacea from the Middle Cambrian of
Mount Stephen, B.C. Canadian Record of Science, 5, 205–208.WHITTINGTON, H. B. 1981. Rare arthropods from the Bur-
gess Shale, Middle Cambrian, British Columbia. Philosophical
Transactions of the Royal Society of London, Series B: Biological
Sciences, 292, 329–357.-and BOHLIN, B. 1958. New Lower Ordovician Odonto-
pleuridae (Trilobita) from €Oland. Bulletin of the Geological
Institutions of the University of Uppsala, 38, 37–45.-CHATTERTON, B. D. E., SPEYER, S. E., FORTEY,
R. A., OWENS, R. M., CHANG, W. T., DEAN, R. A.,
JELL, P. A., LAURIE, J. R., PALMER, A. R., REPINA,
L. N., RUSHTON, A. W. A., SHERGOLD, J. H.,
CLARKSON, E. N. K., WILMOT, N. V. and KELLY, S.
R. A. 1997. Trilobita. 1–530. In KAESLER, R. L. (ed.). Trea-
tise on invertebrate paleontology, Part O, Arthropoda 1 (revised).
Geological Society of America, Boulder, CO, and University of
Kansas Press, Lawrence, KS, 530 pp.
YANG, J., ORTEGA-HERN �ANDEZ, J., BUTTERFIELD,
N. J. and ZHANG, X.-G. 2013. Specialized appendages in fu-
xianhuiids and the head organization of early euarthropods.
Nature, 494, 468–471.ZHANG, X. and SHU, D. 2005. A new arthropod from the
Chengjiang Lagerst€atte, early Cambrian, southern China.
Alcheringa, 29, 185–194.-FU, D. and DAI , T. 2012. A new xandarellid arthropod
from the Chengjiang Lagerst€atte, lower Cambrian of South-
west China. Geobios, 45, 335–338.
276 PALAEONTOLOGY , VOLUME 58







![[Is two weeks the optimum duration criterion for major depression?]](https://static.fdokumen.com/doc/165x107/63366991242ed15b940db544/is-two-weeks-the-optimum-duration-criterion-for-major-depression.jpg)












