
Numerical modelling of the benthic-pelagic coupling in coastal ...
Upload
independentCategory
view
1download
0
280
Temperature Sensitivity of Cardiac Function in Pelagic Fishes with
Different Vertical Mobilities: Yellowfin Tuna (Thunnus albacares),
Bigeye Tuna (Thunnus obesus), Mahimahi (Coryphaena hippurus),
and Swordfish (Xiphias gladius)
* Corresponding author. Present address: Land and Food Systems, University
of British Columbia, 2357 Main Mall, Vancouver, British Columbia V6T 1Z4,
Canada; e-mail: [email protected].
Physiological and Biochemical Zoology 82(3):280–290. 2009. � 2009 by TheUniversity of Chicago. All rights reserved. 1522-2152/2009/8203-8056$15.00DOI: 10.1086/597484
Gina L. J. Galli1,*Holly A. Shiels1
Richard W. Brill2
1Faculty of Life Sciences, University of Manchester, CoreTechnology Facility, 46 Grafton Street, Manchester M13 9NT,United Kingdom; 2Cooperative Marine Education andResearch Program, Northeast Fisheries Science Center,National Marine Fisheries Service, Virginia Institute ofMarine Science, P.O. Box 1346, Gloucester Point, Virginia23062
Accepted 6/19/2008; Electronically Published 3/13/2009
ABSTRACT
We measured the temperature sensitivity, adrenergic sensitivity,and dependence on sarcoplasmic reticulum (SR) Ca2� of ven-tricular muscle from pelagic fishes with different vertical mo-bility patterns: bigeye tuna (Thunnus obesus), yellowfin tuna(Thunnus albacares), and mahimahi (Coryphaena hippurus) anda single specimen from swordfish (Xiphias gladius). Ventricularmuscle from the bigeye tuna and mahimahi exhibited a biphasicresponse to an acute decrease in temperature (from 26� to 7�C);twitch force and kinetic parameters initially increased and thendeclined. The magnitude of this response was larger in thebigeye tuna than in the mahimahi. Under steady state condi-tions at 26�C, inhibition of SR Ca2� release and reuptake withryanodine and thapsigargin decreased twitch force and kineticparameters, respectively, in the bigeye tuna only. However, theinitial inotropy associated with decreasing temperature wasabolished by SR inhibition in both the bigeye tuna and themahimahi. Application of adrenaline completely reversed theeffects of ryanodine and thapsigargin, but this effect was di-minished at cold temperatures. In the yellowfin tuna, temper-ature and SR inhibition had minor effects on twitch force andkinetics, while adrenaline significantly increased these param-eters. Limited data suggest that swordfish ventricular muscle
responds to acute temperature reduction, SR inhibition, andadrenergic stimulation in a manner similar to that of bigeyetuna ventricular muscle. In aggregate, our results show that thetemperature sensitivity, SR dependence, and adrenergic sensi-tivity of pelagic fish hearts are species specific and that thesedifferences reflect species-specific vertical mobility patterns.
Introduction
Ultrasonic telemetry and archival tagging studies have shownthat the daily vertical movements and thermal range of largepelagic fishes (e.g., tunas, billfishes, and sharks) are speciesspecific (Brill et al. 1999; Block et al. 2001, 2005; Gunn andBlock 2001; Musyl et al. 2003). For example, yellowfin tuna(Thunnus albacares) and mahimahi (Coryphaena hippurus)rarely inhabit depths below 300 m, corresponding to a tem-perature range between 17� and 30�C (Schuck 1951; Carey andRobison 1981; Brill 1994; Block et al. 1997; Brill et al. 1999),while bigeye tuna (Thunnus obesus) and swordfish (Xiphiasgladius) routinely reach depths of 700 m, where they encounterand remain in water temperatures as low as 2.8�C (Carey andRobison 1981; Hanamoto 1987; Carey 1990; Holland et al.1990; Musyl et al. 2003).
For those pelagic fish that forage at depths in excess of 300m, their cardiorespiratory system must be able to function overa broad thermal range (2.8�–30�C; Holland et al. 1990; Brill1994; Brill and Bushnell 2001; Musyl et al. 2003; Brill et al.2005). It is likely, therefore, that pelagic fish that routinelyencounter broad thermal environments have cardiac speciali-zations to sustain function at cold temperatures. Some pelagicfish, such as tuna, swordfish, and lamnid sharks, have evolvedcountercurrent heat exchangers that conserve metabolic heatin the slow-twitch muscle, viscera, brain, and eyes and maintaintemperatures of these organs above ambient water temperatures(Graham and Dickson 2001). However, the hearts of these fishesare near the ventral body wall, outside areas maintained atelevated temperatures, and on the ‘‘downstream’’ side of thevascular heat exchangers (Carey et al. 1971; Carey and Gibson1983; Graham and Dickson 2001). Furthermore, the blood sup-ply to the coronary circulation arises directly from the gills andis always at ambient temperature (Carey and Gibson 1983;Carey et al. 1984). Thus, the hearts of even the largest tunas,billfishes, and pelagic sharks operate near (within 1�C) ambient

Temperature Sensitivity of Cardiac Muscle in Pelagic Fish 281
water temperatures while supplying blood to metabolicallyheated tissues (Graham 1983).
The factors enabling large pelagic fishes to maintain cardiacfunction over broad thermal gradients remain largely unknown.Several studies have pointed to enhanced specializations in ex-citation-contraction (E-C) coupling as a possible contributingfactor. For example, the L-type Ca2� channel current density(ICa) is larger and inactivation kinetics are faster in bluefin tuna(Thunnus thynnus orientalis) cardiac myocytes (a tuna specieswith a wide tolerance of ambient temperatures) than in thePacific mackerel (Scomber japonicus; Shiels et al. 2004). Fur-thermore, enhanced expression of E-C coupling proteins re-sponsible for sarcoplasmic reticulum (SR) Ca2� uptake havebeen demonstrated in bluefin tuna compared with yellowfintuna and albacore (Thunnus alalunga; Landeira-Fernandez etal. 2007). This is also the case in salmon sharks (Lamna ditropis)compared with blue sharks (Prionace glauca; Weng et al. 2005).The former routinely occupy cold north Pacific waters nearAlaska (Weng et al. 2005) whereas the latter are more limitedin their tolerance of cold temperatures (Carey and Sharold1990). The correlation between SR contribution and cold tol-erance has been further demonstrated in the myocardium ofhibernating mammals (Belke et al. 1991). Last, the sensitivityof the L-type Ca2� channel to adrenergic stimulation is en-hanced at cold temperatures in the rainbow trout (Oncorhyn-chus mykiss; Shiels et al. 2003), suggesting that adrenaline mayplay an important role in cold tolerance of cardiac function.Thus, enhanced cardiac Ca2� cycling may provide a means tosustain heart rate and cardiac performance in cold water.
Our experiments were designed to test the hypothesis thatpelagic fish that routinely encounter broad thermal environ-ments (bigeye tuna and swordfish) have cardiac specializationsthat allow their hearts to function at cold temperatures. Wepropose that the SR and elevated adrenergic stimulation maycontribute to sustaining cardiac function in cold waters. To testthese hypotheses, we measured the temperature sensitivity, SRdependence, and adrenergic sensitivity of isolated ventricularmuscle from three pelagic species with different vertical move-ment patterns: bigeye tuna, yellowfin tuna, and mahimahi. Inaddition, we present preliminary data from one swordfish,whose cardiac contractile parameters have never before beenmeasured.
Material and Methods
Location and Experimental Animals
Operations were conducted at sea aboard the National Oceanicand Atmospheric Administration research vessel Oscar EltonSette in the central Pacific near the main Hawaiian Islands. Allexperimental fish (ranging between 1 and 3.5 kg) were caughtusing standard recreational trolling and conventional longlinegear. Three species were studied, chosen according to theirthermal range (Schuck 1951; Block et al. 1997; Brill et al. 1999,2005; Musyl et al. 2003): bigeye tuna ( ), yellowfin tunan p 4( ), and mahimahi ( ). Additionally, we obtained datan p 5 n p 5
from one swordfish, whose thermal range is similar to that ofthe bigeye tuna.
Experimental Protocols
Each animal was pithed to fully destroy neural connections,and the heart was quickly excised and transferred into oxy-genated physiological saline for further dissection. The con-stituents of the physiological saline were the same for eachspecies (mmol L�1): NaCl, 175; KCl, 7; MgCl2, 1.1; HEPES, 10;CaCl2, 3.0; glucose, 10; with a pH of 7.7, adjusted with NaOH.Four longitudinal myocardial strips were prepared from theventricle. Where possible, strips were dissected out of trabeculaebundles to promote similar fiber orientation.
Strips were hung vertically from force transducers in a four-channel Myobath system (World Precision Instruments, Sara-sota, FL). The tissue baths were filled with physiological saline,continually bubbled with 100% oxygen and maintained at de-sired temperatures by a circulating water bath. Contraction wasinitiated via platanium stimulation electrodes delivering squarestimulation pulses (pulse duration 10 ms, voltage 2# that elic-iting maximal response; Grass Instruments SD 9 stimulator,Qunicy, MA). The signals from the force transducers were am-plified using a four-channel Transbridge 4M bridge amplifier(WPI Instruments) and digitized (at 100 Hz sampling fre-quency) using an IOTech DaqBook 120 (Cleveland, OH). Thetemperature of physiological saline in one tissue bath was si-multaneously recorded using a thermocouple module (IOTechDBK 52) connected to the DaqBook 120. The force and tem-perature data (see Fig. 1) were displayed and recorded on alaptop computer using custom software developed by the Dasy-lab graphical programming system (version 9.0; National In-struments, Austin, TX).
After mounting, the ventricular strips were paced at 0.5 Hzfor 30 min at 26�C to allow them to stabilize. They were thenstretched to Lmax (length at which twitch force is maximal) andleft to stabilize for at least 15 min before experimentation. Forcalculation of absolute force values, the width of each musclestrip was measured at the end of experimentation so that cross-sectional area could be estimated (assuming that the strip wascylindrical and using the equation for cross-sectional area, pr2).
Contractile Performance and TemperatureSensitivity of Cardiac Tissue
Our experimental protocol was designed to determine the tem-perature sensitivity of ventricular tissue from each species andto test the hypothesis that enhanced SR Ca2� cycling and/oradrenergic stimulation sustains cardiac function at low tem-peratures. SR dependence was assessed using ryanodine (10mmol L�1), a specific blocker of the SR Ca2� release channel(Rousseau et al. 1987), and thapsigargin (2 mmol L�1), a specificinhibitor of the SR Ca2� ATPase pump (Sagara and Inesi 1991;Shiels and White 2005). Because these agents were used to-gether, both Ca2� release and Ca2� reuptake into the SR wereblocked simultaneously. Because adrenaline is present at na-
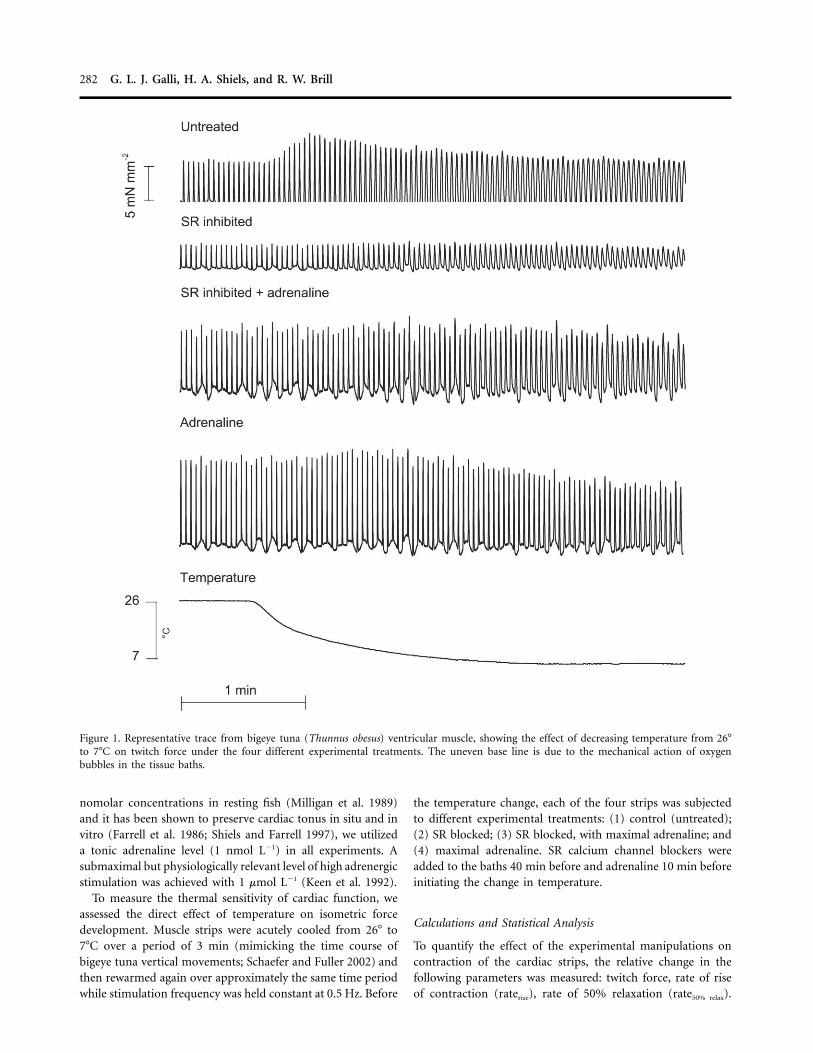
282 G. L. J. Galli, H. A. Shiels, and R. W. Brill
Figure 1. Representative trace from bigeye tuna (Thunnus obesus) ventricular muscle, showing the effect of decreasing temperature from 26�to 7�C on twitch force under the four different experimental treatments. The uneven base line is due to the mechanical action of oxygenbubbles in the tissue baths.
nomolar concentrations in resting fish (Milligan et al. 1989)and it has been shown to preserve cardiac tonus in situ and invitro (Farrell et al. 1986; Shiels and Farrell 1997), we utilizeda tonic adrenaline level (1 nmol L�1) in all experiments. Asubmaximal but physiologically relevant level of high adrenergicstimulation was achieved with 1 mmol L�1 (Keen et al. 1992).
To measure the thermal sensitivity of cardiac function, weassessed the direct effect of temperature on isometric forcedevelopment. Muscle strips were acutely cooled from 26� to7�C over a period of 3 min (mimicking the time course ofbigeye tuna vertical movements; Schaefer and Fuller 2002) andthen rewarmed again over approximately the same time periodwhile stimulation frequency was held constant at 0.5 Hz. Before
the temperature change, each of the four strips was subjectedto different experimental treatments: (1) control (untreated);(2) SR blocked; (3) SR blocked, with maximal adrenaline; and(4) maximal adrenaline. SR calcium channel blockers wereadded to the baths 40 min before and adrenaline 10 min beforeinitiating the change in temperature.
Calculations and Statistical Analysis
To quantify the effect of the experimental manipulations oncontraction of the cardiac strips, the relative change in thefollowing parameters was measured: twitch force, rate of riseof contraction (raterise), rate of 50% relaxation (rate50% relax).

Temperature Sensitivity of Cardiac Muscle in Pelagic Fish 283
Table 1: Force and kinetic parameters for ventricular muscle from pelagic fish
Twitch Force(mN mm�2) TTP (ms) TT50% relax (ms)
Raterise
(mN mm�1 s�1)Rate50% relax
(mN mm�1 s�1)
Bigeye tuna 4.3 � .8a 308.8 � 9.5 143.8 � 7.5 14.0 � 2.5 16.0 � 3.7Mahimahi 3.0 � .6 295.9 � 13.6 111.2 � 6.4 11.3 � 2.8 14.7 � 3.4Yellowfin tuna 4.7 � .7b 287.1 � 11.4 150.0 � 7.7b 16.5 � 2.9b 16.3 � 3.0
Note. Absolute values for twitch force, time to peak force (TTP), time to 50% relaxation (TT50% relax), rate of rise of contraction
(raterise), and rate of 50% relaxation (rate50% relax) in ventricular muscle at 26�C from bigeye tuna (Thunnus obesus; ), mahimahin p 4
(Coryphaena hippurus; ), and yellowfin tuna (Thunnus albacares; ). Data are means � SEM. Differences were consideredn p 4 n p 5
significant when .P ! 0.05a Significant difference between bigeye tuna and mahimahi.b Significant difference between mahimahi and yellowfin tuna.
Measurements were taken before and after any pharmacologicalagents were used and at every 2�C drop in temperature duringthe cooling period of the temperature challenge. Twitch forcewas measured as the difference between peak and resting force,raterise was calculated by dividing twitch force by the time takento reach peak force from stimulation, and rate50% relax was cal-culated by dividing half twitch force with the time taken toreach 50% relaxation. Absolute values for twitch force are pre-sented in Table 1. For comparative purposes, all other data arepresented in relative terms, where data were normalized to theinitial value recorded at 26�C before both the temperature chal-lenge and any drug treatment. Significant ( ) changesP ! 0.05in relative force or kinetics were assessed using a one-way re-peated-measure ANOVA on ranks. Statistical tests were per-formed only when . Preliminary data from one swordfishn ≥ 4are presented separately from those from the other species atthe end of each section.
Results
Twitch force was significantly greater in the bigeye tuna andyellowfin tuna than in the mahimahi. All other absolute valueswere similar between species, with the exception of raterise,which was significantly larger in the yellowfin tuna than in themahimahi (Table 1).
Effect of Temperature on Untreated Ventricular Muscle
Exemplary results obtained with ventricular strips from a bigeyetuna (Fig. 1) demonstrate the effects of an acute reduction intemperature and the interplay between temperature and drugtreatment on ventricular muscle function. A reduction in tem-perature led to an initial increase in contractile force in bigeyetuna and mahimahi ventricular muscle, followed by a decline(Fig. 2A, 2B). The magnitude of the initial response was sig-nificantly larger in the bigeye tuna than in the mahimahi. Fur-thermore, the temperature at which peak twitch force occurredwas significantly lower in the bigeye tuna ( ) than14.3� � 0.1�Cin the mahimahi ( ). In the yellowfin tuna, chang-18.2� � 0.1�Cing temperature had no significant influence on twitch force(Fig. 2C). Raterise and rate50% relax in bigeye tuna ventricular mus-cle initially increased with decreasing temperature and thensteadily declined (Fig. 3A, 3D). In the mahimahi and yellowfin
tuna, however, no significant differences were observed in ki-netic parameters at any of the temperatures tested (Fig. 3B, 3C,3E, 3F). The lowest temperature at which ventricular musclemaintained regular contractions was significantly species spe-cific: bigeye tuna ( ) ! mahimahi ( ) !7.3� � 0.3�C 12.2� � 0.1�Cyellowfin tuna ( ).14.2� � 0.1�C
Effects of Temperature on Ventricular MuscleTreated with SR Inhibitors
At 26�C before a temperature challenge, SR blockade signifi-cantly reduced contractile force in the bigeye tuna only (Table2). In this species, the initial increase in contractile force andkinetic parameters associated with a reduction in temperaturewas entirely abolished by pretreatment with SR inhibitors (Figs.2A, 3A). Furthermore, the lowest temperature at which ven-tricular muscle could maintain regular contractions was sig-nificantly higher in ryanodine- and thapsigargin-treated muscle( ) than in untreated muscle ( ). In the13.5� � 0.6�C 7.3� � 0.3�Cmahimahi, SR inhibition also led to an abolition of the initialincrease in twitch force associated with decreasing temperature,while all other parameters were similar to those for untreatedmuscle (Fig. 2B; Fig. 3B, 3E; Table 2). In the yellowfin tuna,no significant differences were observed in untreated versusSR-inhibited ventricular muscle (Figs. 2C, 3C; Table 2).
Effects of Temperature on Ventricular Muscle Treated withAdrenaline and Adrenaline Plus SR Inhibitors
Application of maximal adrenergic stimulation (1 mmol L�1)at 26�C significantly increased contractile force and kinetics ofcontraction/relaxation in all species studied (Table 2; Figs. 2,3). This positive inotropy was apparent at all temperaturestested (Figs. 2, 3), but the effect was diminished at cold tem-peratures. The magnitude of the positive inotropy was largestin the bigeye tuna, while values in the mahimahi and yellowfintuna were similar (Table 2). When adrenaline was applied toventricular muscle pretreated with SR inhibitors, twitch forceand the kinetics of contraction were significantly higher in thebigeye tuna and mahimahi than in untreated muscle (Figs. 2,3A). However, this effect was diminished at cold temperatures.In the mahimahi, significant effects of adrenaline in muscle

284 G. L. J. Galli, H. A. Shiels, and R. W. Brill
Figure 2. Relative change in twitch force when temperature is decreasedfrom 26� to 7�C in ventricular muscle under different drug treatmentsfrom bigeye tuna (Thunnus obesus [A]; ), mahimahi (Coryphaenan p 4hippurus [B]; ), and yellowfin tuna (Thunnus albacares [C];n p 5
). Data are means � SEM. a p significant difference in un-n p 5treated muscle twitch force compared with initial value; b p significantdifference between control and sarcoplasmic reticulum (SR)-blocked(ryanodine, 10 mm L�1; thapsigargin, 2 mm L�1) muscle; c p significantdifference between control and SR-blocked � adrenaline-treated (1mm L�1) muscle; d p significant difference between control and adren-aline-treated (1 mm L�1) muscle ( )..P ! 0.05
pretreated with SR inhibitors were observed only in kineticparameters recorded at temperatures above 22�C (Fig. 3C).
Swordfish
Data were opportunistically obtained from one swordfish. Sim-ilar to the other three species studied, a reduction in temper-
ature led to a biphasic response, with twitch force initially in-creasing and then declining at the colder temperatures (Fig. 4).The magnitude of this response and the lowest attainable tem-perature were similar to those of bigeye tuna. The kinetics ofcontraction remained relatively constant throughout the tem-perature challenge (Fig. 4B, 4C).
SR inhibition led to a 32.5% reduction in twitch force inswordfish ventricular muscle at 26�C (Fig. 4A), similar to bigeyetuna hearts. Furthermore, SR blockade decreased the capacityof the muscle to develop force and reduced the rate of bothcontraction and relaxation (Fig. 4). Adrenergic stimulation in-creased twitch force and raterise at all temperatures tested, whilerate50% relax was unaffected (Fig. 4C). When adrenaline was givento swordfish ventricular muscle pretreated with SR Ca2� chan-nel inhibitors, the relative change in twitch force and raterise
increased at all temperatures tested, while rate50% relax was un-affected (Fig. 4).
Discussion
We present the first measurements of contractile force/kineticsin ventricular muscle from bigeye tuna, yellowfin tuna, ma-himahi, and swordfish. Furthermore, this is the first study toinvestigate the effects of acute temperature change, SR inhi-bition, and adrenergic sensitivity in ventricular muscle fromany of these species. Our results clearly demonstrate that thetemperature sensitivity of ventricular muscle from pelagic fishis species specific and that maintenance of contractility over awide temperature range may require the recruitment of intra-cellular Ca2� from the SR, the ability to augment force throughthe adrenergic signaling cascade, or both.
Effect of Temperature on Contractile Force and Kinetics
All three pelagic fish species exhibited robust twitch force at26�C, which is the temperature of the surface water from whichthey were caught. The absolute force production of ventricularstrips from pelagic species (Table 1) is comparable to that foundin atrial muscle from the yellowfin tuna at 25�C (Shiels et al.1999) and that found in the ectothermic sister taxa of Thunnus,Pacific mackerel (Scomber japonicus; Shiels and Farrell 2000),at 20�C. Twitch force and kinetics of contraction in ventricularmuscle from the bigeye tuna and mahimahi exhibited a biphasicresponse when temperature was reduced from 26� to 7�C, whilethese parameters were unaffected in the yellowfin tuna. Theinitial positive inotropy associated with acute reductions intemperature is a common response observed in a number ofteleost species, such as rainbow trout (Shiels and Farrell 1997),burbot (Tiitu and Vornanen 2001), carp (Vornanen 1989),mackerel (Shiels and Farrell 2000), and yellowfin tuna atrialmuscle (Shiels et al. 1999), and is likely to promote exploitationinto cold waters. In this study, the magnitude of this responsewas larger in the bigeye tuna than in the mahimahi. Further-more, the lowest temperature at which regular contractionswere maintained was significantly lower in the bigeye tuna thanin the other species. These findings correlate with the com-

Temperature Sensitivity of Cardiac Muscle in Pelagic Fish 285
Figure 3. Relative change in rate of rise of contraction (A–C) and rate of 50% relaxation (C–F) when temperature is decreased from 26� to7�C in bigeye tuna (Thunnus obesus [A, D]; ), mahimahi (Coryphaena hippurus [B, E]; ), and yellowfin tuna (Thunnus albacaresn p 4 n p 5[C, F]; ) ventricular muscle. Data are means � SEM. a p significant difference in untreated muscle twitch force compared with initialn p 5value; b p significant difference between control and sarcoplasmic reticulum (SR)-blocked (ryanodine, 10 mm L�1; thapsigargin, 2 mm L�1)muscle; c p significant difference between control and SR-blocked � adrenaline-treated (1 mm L�1) muscle; d p significant difference betweencontrol and adrenaline-treated (1 mm L�1) muscle ( ).P ! 0.05
paratively broader thermal range of the bigeye tuna, ∼3�–30�Ccompared with 17�–30�C in the yellowfin tuna or mahimahi(Schuck 1951; Brill 1994, 1999; Block et al. 1997).
It should be noted, however, that our study did not take intoaccount the effect of frequency on contractile force becausemuscle was paced at 0.5 Hz at all temperatures. A reductionin temperature leads to bradycardia in teleosts (Korsmeyer etal. 1997a, 1997b; Overgaard et al. 2004; Sandblom and Axelsson2007), including tuna (Blank et al. 2002, 2004). In addition,most fish exhibit a negative force-frequency response, with con-
tractile force decreasing with increasing frequency over thephysiological range (Shiels et al. 2002a). Force-frequency re-lationships for ventricular muscle from the pelagic species inthis study have not yet been ascertained. However, assuming anegative force-frequency response and a cold bradycardia, ourdata may have underestimated the positive inotropic effect oflow temperatures on cardiac function. The cold-induced pos-itive inotropy is likely to afford protection in colder watersthrough increased cardiac pumping capacity (product of twitchforce and frequency).

286 G. L. J. Galli, H. A. Shiels, and R. W. Brill
Table 2: Percentage change in force for ventricular muscle from pelagic fishspecies at 26�C after sarcoplasmic reticulum (SR) inhibition or adrenalinestimulation
Bigeye Tuna Mahimahi Yellowfin Tuna
SR blockade (% decrease) 29.2 � 4.7a,b,c 1.0 � 1.0 6.1 � 3.6Adrenaline (% increase) 329.2 � 2.5a,b,c 239.1 � 35.8a 192.8 � 22.0a
Note. Percentage decrease in twitch force due to SR inhibition (ryanodine, 10 mmol L�1; thapsigargin,
2 mmol L�1) and percentage increase with adrenaline (1 mmol L�1) in ventricular muscle from the
bigeye tuna (Thunnus obesus; ), mahimahi (Coryphaena hippurus; ), and yellowfin tunan p 4 n p 4
(Thunnus albacares; ). Values are means � SEM. All preparations were paced at 0.5 Hz. Dif-n p 5
ferences were considered significant when .P ! 0.05a Significant change as a result of drug treatment from control value of 100%.b Significant difference between bigeye tuna and mahimahi.c Significant difference between bigeye tuna and yellowfin tuna.
Blockade of SR Ca2� Release and Reuptake
In bigeye tuna ventricular muscle under steady state conditionsat 26�C, SR inhibition with ryanodine and thapsigargin reducedcontractile force by , indicating that at this tem-29.2% � 4.8%perature, the SR contributes significantly to contraction andrelaxation on a beat-to-beat basis. Under identical conditions,however, there were no significant effects of SR inhibition inmahimahi and yellowfin tuna ventricular muscle.
SR dependence has been linked to many factors, includinghigh heart rates and blood pressures (Keen et al. 1992; Shielset al. 1999; Shiels and Farrell 2000; Galli et al. 2006), tissuetype (Keen et al. 1992; Aho and Vornanen 1999; Bers 2001),and tolerance of environmental stress (Belke et al. 1991; Vor-nanen and Paajanen 2004). In most teleosts, ventricular muscleis relatively SR independent, with contraction and relaxationbeing strongly dependent on sarolemmal Ca2� cycling (Driedzicand Gesser 1988; Hove-Madsen and Gesser 1989; Shiels andFarrell 1997; Vornanen 1997, 1998; Aho and Vornanen 1999;Harwood et al. 2000; Hove-Madsen and Tort 2001; Tiitu andVornanen 2001). However, in atrial muscle, the SR contributessignificant amounts of activator Ca2� for contraction, partic-ularly in the active species of fish of the Scombridae family(Keen et al. 1992; Shiels et al. 1999, 2001; Shiels and Farrell2000). In yellowfin and skipjack tuna atrial muscle paced at0.5 Hz at 25�C, ryanodine decreased contractile force by ∼50%and 30%, respectively (Keen et al. 1992; Shiels et al. 1999). Thishas led to the suggestion that high heart rates and blood pres-sures require intracellular Ca2� cycling to support contractionand relaxation in ectotherms (Keen et al. 1992; Shiels et al.1999; Shiels and Farrell 2000; Galli et al. 2006). Recent studieson Neotropical fish have suggested that SR involvement is de-pendent on phylogeny, with certain superorders such as Os-tariophysi relying more strongly on SR Ca2� than the super-order Acanthopterygii (Rivaroli et al. 2006). This hypothesiscould be extended to the family Scombridae because the Pacificmackerel, yellowfin, skipjack, bluefin, and bigeye tunas appearto be SR dependent (Keen et al. 1992; Shiels et al. 1999; Shielsand Farrell 2000; Landeira-Fernandez et al. 2007). In this study,only bigeye tuna ventricular muscle was sensitive to SR inhi-
bition under steady state conditions at 26�C, which is perhapsreflective of the high cardiac activity of this animal.
SR inhibition completely abolished the positive inotropy as-sociated with a reduction in temperature in bigeye tuna andmahimahi. This suggests that the SR plays an important rolein regulating contractility at cold temperatures. In mammals,acute reductions in temperature increase the open probabilityof the SR Ca2� release channel (Sitsapesan et al. 1991), withthe channel being locked in an open state at 0�–1�C (Bers 1987).In contrast, in multicellular preparations from burbot accli-mated and tested at 1�C, ryanodine reduced force by 32% inatrium and 16% in ventricle (Tiitu and Vornanen 2001). Infact, the SR Ca2� leak actually decreased with temperature from20� to 5�C in rainbow trout ventricular myocytes (Hove-Madsen et al. 1998). Thus, in stark contrast to mammals, theSR continues to function at cold temperatures in fish and islikely to play an important role in maintaining contractility.
The increased SR involvement with acute temperature re-duction may arise from a prolongation of the cardiac actionpotential at cold temperatures (Driedzic and Gesser 1994; Shielset al. 2002b; Vornanen et al. 2002). This would increase theamount of activator Ca2� entering the myocyte through the L-type Ca2� channel and could lead to greater Ca2� loading inthe SR (Bers 2001). However, our study did not take into ac-count the effect of frequency, which is known to modify actionpotential duration and SR loading in fish (Shiels et al. 2002b).Thus, future research on pelagic fish should be directed towardintegrating the effects of frequency, action potential duration,and temperature on contractile performance.
Effects of Adrenaline
Consistent with previous studies on fish, adrenaline increasedtwitch force in ventricular tissue from all species and at alltemperatures (Shiels and Farrell 1997, 2000; Shiels et al. 1999,1998, 2002a, 2003; Farrell et al. 2007; Rocha et al. 2007; Rytterand Gesser 2007). This classic positive inotropic response canbe attributed to a rise in cytosolic Ca2� concentration mediatedthrough the adrenergic signaling cascade (Reuter et al. 1983,

Temperature Sensitivity of Cardiac Muscle in Pelagic Fish 287
Figure 4. Change in twitch force (A), rate of rise of contraction (B),and rate of 50% relaxation (C) when temperature is decreased from26� to 7�C in cardiac muscle from one swordfish (Xiphias gladius)under different experimental treatments.
1986). In the bigeye tuna and mahimahi, adrenaline over-whelmed the negative effects of SR inhibition, suggesting thatin physiological situations where sympathetic tone is high, suchas exercise, the relative contribution of the SR is overshadowedby an adrenergically stimulated increase in sarcolemmal Ca2�
influx, which is sufficient to allow adequate Ca2� cycling. Thus,adrenergic stimulation offers another line of defense for fishexploiting cold water and may be an important modulator ofcontractility during cold foraging dives. This may be especiallyimportant if pelagic fish species show an acute cold-induced
enhancement of adrenergic sensitivity (Shiels et al. 2003). How-ever, the role of adrenaline in modulating inotropy during adive into cold water may be species specific and reflect aninterplay between sarcolemmal and SR Ca2� cycling. In themahimahi and yellowfin, adrenaline increased force above con-trol values across all temperatures in the presence and absenceof SR blockade (Fig. 2B, 2C). Thus, in species where SR uti-lization is less evident, adrenaline may be important for mo-bilizing cellular Ca2� flux. In the bigeye tuna, the adrenergicincrease in force (in the presence and absence of SR blockade)was dampened at low temperatures (Fig. 2A), suggesting thatSR Ca2� is vital for maintaining cardiac function in the cold.
Preliminary Data from the Swordfish
Data from the swordfish suggest that this species responds toacute temperature change in a manner similar to the bigeyetuna. Furthermore, SR inhibition led to a 32.5% reduction intwitch force under steady state conditions at 26�C and com-pletely abolished the positive inotropy associated with a re-duction in temperature. Similar to bigeye tuna, swordfish rou-tinely exploit depths of 700 m and encounter watertemperatures as low as 2.8�C (Carey and Robison 1981; Carey1990). Therefore, our preliminary data from the swordfish arein line with our hypothesis that the hearts of pelagic fish thatroutinely encounter broad thermal environments are relativelyinsensitive to temperature and rely strongly on SR Ca2� cyclingto support contraction at low temperatures.
Concluding Remarks
In order to exploit the large prey resources of the deep sound-scattering layer, certain pelagic species such as bigeye tuna andswordfish have evolved a host of physiological specializationsto tolerate the large fluctuations in temperature that accompanyvertical mobility in the sea. Our results suggest that one suchspecialization is cardiac muscle that increases force as temper-ature declines and that recruits Ca2� from the intracellularstores of the SR to support contraction/relaxation at cold tem-peratures. The absence or retardation of this specialization inother pelagic species such as yellowfin tuna and mahimahi maylimit the depth at which these species can forage. In this way,the thermal sensitivity of the fish heart may play a role invertical mobility and niche expansion.
Acknowledgments
This study was supported by the Biotechnology and BiologicalSciences Research Council, the Company of Biologists, the Fish-eries Society of the British Isles, and the U.S. National Oceanicand Atmospheric Administration. Special thanks to M. Musyl(University of Hawaii) and the crew and the scientists aboardthe Oscar Elton Sette for their help and advice.

288 G. L. J. Galli, H. A. Shiels, and R. W. Brill
Literature Cited
Aho E. and M. Vornanen. 1999. Contractile properties of atrialand ventricular myocardium of the heart of rainbow trout(Oncorhynchus mykiss): effects of thermal acclimation. J ExpBiol 202:2663–2677.
Belke D.D., R.E. Milner, and L.C. Wang. 1991. Seasonal vari-ations in the rate and capacity of cardiac SR calcium accu-mulation in a hibernating species. Cryobiology 28:354–363.
Bers D.M. 1987. Ryanodine and the calcium content of cardiacSR assessed by caffeine and rapid cooling contractures. AmJ Physiol 253:C408–C415.
———. 2001. Excitation-Contraction Coupling and Contrac-tile Force. Kluwer Academic, Dordecht.
Blank J.M., J.M. Morrissette, P.S. Davie, and B.A. Block. 2002.Effects of temperature, epinephrine and Ca2� on the heartsof yellowfin tuna (Thunnus albacares). J Exp Biol 205:1881–1888.
Blank J.M., J.M. Morrissette, A.M. Landeira-Fernandez, S.B.Blackwell, T.D. Williams, and B.A. Block. 2004. In situ cardiacperformance of Pacific bluefin tuna hearts in response toacute temperature change. J Exp Biol 207:881–890.
Block B.A., H. Dewar, S.B. Blackwell, T.D. Williams, E.D.Prince, C.J. Farwell, A. Boustany, et al. 2001. Migratorymovements, depth preferences, and thermal biology of At-lantic bluefin tuna. Science 293:1310–1314.
Block B.A., J.E. Keen, B. Castillo, H. Dewar, E.V. Freund, D.J.Marcinek, R.W. Brill, and C. Farwell. 1997. Environmentalpreferences of yellowfin tuna (Thunnus albacares) at thenorthern extent of its range. Mar Biol 130:119–132.
Block B.A., S.L. Teo, A. Walli, A. Boustany, M.J. Stokesbury,C.J. Farwell, K.C. Weng, H. Dewar, and T.D. Williams. 2005.Electronic tagging and population structure of Atlantic blue-fin tuna. Nature 434:1121–1127.
Brill R. 1994. A review of temperature and oxygen tolerancesstudies of tunas, pertinent to fisheries oceanography, move-ment models, and stock assessments. Fish Oceanogr 3:206–216.
Brill R.W., K.A. Bigelow, M.K. Musyl, K.A. Fritsches, and E.J.Warrant. 2005. Bigeye tuna behavior and physiology andtheir relevance to stock assessments and fishery biology. Col-lect Vol Sci Pap ICCAT 57:142–161.
Brill R.W., B.A. Block, C.H. Boggs, K.A. Bigelow, E.V. Freund,and D.J. Marcinek. 1999. Horizontal movements and depthdistribution of large adult yellowfin tuna (Thunnus albacares)near the Hawaiian Islands, recorded using ultrasonic telem-etry: implications for the physiological ecology of pelagicfishes. Mar Biol 133:395–408.
Brill R.W. and P.G. Bushnell. 2001. The cardiovascular systemof tunas. Pp. 79–120 in B.A. Block and E.D. Stevens, eds.Tuna: Physiology, Ecology and Evolution. Academic Press,San Diego, CA.
Carey F.G. 1990. Further ultrasonic telemetry of swordfish. Pp.103–122 in R.H. Stroud, ed. Proceedings of the Second In-ternational Billfish Symposium. Pt. 2. Planning the Future
of Billfish. National Coalition for Marine Conservations, Sa-vannah, GA.
Carey F.G. and Q.H. Gibson. 1983. Heat and oxygen exchangein the rete mirable of the bluefin tuna, Thunnus thynnus.Comp Biochem Physiol A 74:333–342.
Carey F.G., J. Kanwisher, K. Lawson, and J.S. Beckett. 1971.Warm-bodied fish. Am Zool 11:137–145.
Carey F.G., J.W. Kanwisher, and E.D. Stevens. 1984. Bluefintuna warm their viscera during digestion. J Exp Biol 109:1–6.
Carey F.G. and B.H. Robison. 1981. Daily patterns in the ac-tivities of swordfish, Xiphias gladius, observed by acoustictelemetry. Fish Bull 79:277–292.
Carey F.G. and J.V. Sharold. 1990. Movements of blue sharks(Prionace glauca) in depth and course. Mar Biol 106:329–342.
Driedzic W.R. and H. Gesser. 1988. Differences in force fre-quency relationships and calcium dependency between elas-mobranch and teleost hearts. J Exp Biol 140:227–241.
———. 1994. Energy metabolism and contractility in ecto-thermic vertebrate hearts: hypoxia, acidosis, and low tem-perature. Physiol Rev 74:221–258.
Farrell A.P., M. Axelsson, J. Altimiras, E. Sandblom, and G.Claireaux. 2007. Maximum cardiac performance and adren-ergic sensitivity of the sea bass Dicentrarchus labrax at hightemperatures. J Exp Biol 210:1216–1224.
Farrell A.P., K.R. MacLeod, and B. Chancey. 1986. Intrinsicmechanical properties of the perfused rainbow trout heartand the effects of catecholamines and extracellular calciumunder control and acidotic conditions. J Exp Biol 125:319–345.
Galli G.L.J., H. Gesser, E.W. Taylor, H.A. Shiels, and T. Wang.2006. The role of the sarcoplasmic reticulum in the gener-ation of high heart rates and blood pressures in reptiles. JExp Biol 209:1956–1963.
Graham J.B. 1983. Heat transfer. Pp. 248–278 in P.W. Webband D. Weihs, eds. Fish Biomechanics. Praeger, New York.
Graham J.B. and K. Dickson. 2001. Anatomical and physio-logical specializations for endothermy. Pp. 121–165 in B.A.Block and E.D. Stevens, eds. Tuna: Physiology, Ecology andEvolution. Academic Press, San Diego, CA.
Gunn J. and B.A. Block. 2001. Acoustic, archival and pop-upsatellite tagging of tunas. Pages 167–224 in B.A. Block andE.D. Stevens, eds. Tuna: Physiology, Ecology and Evolution.Academic Press, San Diego, CA.
Hanamoto E. 1987. Effect of oceanographic environment onbigeye tuna distribution. Bull Jpn Soc Fish Oceanogr 51:203–216.
Harwood C.L., F.C. Howarth, J.D. Altringham, and E. White.2000. Rate-dependent changes in cell shortening: intracel-lular Ca2� levels and membrane potential in single isolatedrainbow trout (Oncorhynchus mykiss) ventricular myocytes.J Exp Biol 203:493–504.
Holland K.N., R. Brill, and R.K.C. Chang. 1990. Horizontaland vertical movements of tunas (Thunnus spp.) associatedwith fish aggregating devices. Fish Bull 88:493–507.

Temperature Sensitivity of Cardiac Muscle in Pelagic Fish 289
Hove-Madsen L. and H. Gesser. 1989. Force frequency relationin the myocardium of rainbow trout: effects of K� and adren-aline. J Comp Physiol B 159:61–69.
Hove-Madsen L., A. Llach, and L. Tort. 1998. Quantificationof Ca2� uptake in the sarcoplasmic reticulum of trout ven-tricular myocytes. Am J Physiol 44:R2070–R2080.
Hove-Madsen L. and L. Tort. 2001. Characterization of therelationship between Na�-Ca2� exchange rate and cytosoliccalcium in trout cardiac myocytes. Pflug Arch Eur J Physiol441:701–708.
Keen J.E., A.P. Farrell, G.F. Tibbits, and R.W. Brill. 1992. Cardiacphysiology in tunas. 2. Effect of ryanodine, calcium, andadrenaline on force frequency relationships in atrial stripsfrom skipjack tuna, Katsuwonus pelamis. Can J Zool 70:1211–1217.
Korsmeyer K.E., N.C. Lai, R.E. Shadwick, and J.B. Graham.1997a. Heart rate and stroke volume contribution to cardiacoutput in swimming yellowfin tuna: response to exercise andtemperature. J Exp Biol 200:1975–1986.
———. 1997b. Oxygen transport and cardiovascular responsesto exercise in the yellowfin tuna Thunnus albacares. J ExpBiol 200:1987–1997.
Landeira-Fernandez A., J.M. Morrisette, J.M. Blank, and B.A.Block. 2007. Temperature dependence of Ca2�-ATPase(SERCA2) in the ventricles of tuna and mackerel. Am J Phys-iol 286:R398–R404.
Milligan C.L., M.S. Graham, and A.P. Farrell. 1989. The re-sponse of trout red cells to adrenaline during seasonal ac-climation and changes in temperature. J Fish Biol 35:229–236.
Musyl M., R. Brill, C.H. Boggs, D.S. Curran, T.K. Kazama, andM.P. Seki. 2003. Vertical movements of bigeye tuna (Thunnusobesus) associated with islands, buoys, and seamounts of theHawaiian Archipelago from archival tagging data. FishOceanogr 12:152–169.
Overgaard J., J.A. Stecyk, H. Gesser, T. Wang, and A.P. Farrell.2004. Effects of temperature and anoxia upon the perfor-mance of in situ perfused trout hearts. J Exp Biol 207:655–665.
Reuter H., A.B. Cachelin, J.E. De Peyer, and S. Kokubun. 1983.Modulation of calcium channels in cultured cardiac cells byisoproterenol and 8-bromo-cAMP. Cold Spring HarborSymp Quant Biol 48:193–200.
Reuter H., S. Kokubun, and B. Prod’hom. 1986. Properties andmodulation of cardiac calcium channels. J Exp Biol 124:191–201.
Rivaroli L., F.T. Rantin, and A.L. Kalinin. 2006. Cardiac functionof two ecologically distinct Neotropical freshwater fish: cur-imbata, Prochilodus lineatus (Teleostei, Prochilodontidae),and trahira, Hoplias malabaricus (Teleostei, Erythrinidae).Comp Biochem Physiol A 145:322–327.
Rocha M.L., F.T. Rantin, and A.L. Kalinin. 2007. Importanceof the sarcoplasmic reticulum and adrenergic stimulation onthe cardiac contractility of the Neotropical teleost Synbran-chus marmoratus under different thermal conditions. J CompPhysiol B 177:713–721.
Rousseau E., J.S. Smith, and G. Meissner. 1987. Ryanodinemodifies conductance and gating behavior of single Ca2�
release channel. Am J Physiol 253:C364–C368.Rytter D. and H. Gesser. 2007. The effect of adrenaline and
high Ca2� on the mechanical performance and oxygen con-sumption of the isolated perfused trout (Oncorhynchus my-kiss) heart. Comp Biochem Physiol A 147:1083–1089.
Sagara Y. and G. Inesi. 1991. Inhibition of the sarcoplasmicreticulum Ca2� transport ATPase by thapsigargin at subnano-molar concentrations. J Biol Chem 266:13503–13506.
Sandblom E. and M. Axelsson. 2007. Venous hemodynamicresponses to acute temperature increase in the rainbow trout(Oncorhynchus mykiss). Am J Physiol 292:R2292–R2298.
Schaefer K.M. and D.W. Fuller. 2002. Movements, behaviour,and habitat selection of bigeye tuna (Thunnus obesus) in theeastern equatorial Pacific, ascertained through archival tags.Fish Bull 100:765–788.
Schuck H.A. 1951. Notes on the dolphin (Coryphaena hippurus)in North Carolina waters. Copeia 1951:35–39.
Shiels H.A., J.M. Blank, A.P. Farrell, and B.A. Block. 2004.Electrophysiological properties of the L-type Ca2� current incardiomyocytes from bluefin tuna and Pacific mackerel. AmJ Physiol 286:R659–R668.
Shiels H.A. and A.P. Farrell. 1997. The effect of temperatureand adrenaline on the relative importance of the sarcoplas-mic reticulum in contributing Ca2� to force development inisolated ventricular trabeculae from rainbow trout. J Exp Biol200:1607–1621.
———. 2000. The effect of ryanodine on isometric tensiondevelopment in isolated ventricular trabeculae from Pacificmackerel (Scomber japonicus). Comp Biochem Physiol A 125:331–341.
Shiels H.A., E.V. Freund, A.P. Farrell, and B.A. Block. 1999.The sarcoplasmic reticulum plays a major role in isometriccontraction in atrial muscle of yellowfin tuna. J Exp Biol202:881–890.
Shiels H.A., E.D. Stevens, and A.P. Farrell. 1998. Effect of tem-perature, adrenaline and ryanodine on power production introut (Oncorhynchus mykiss) ventricular trabeculae. J ExpBiol 201:19–25.
Shiels H.A., M. Vornanen, and A.P. Farrell. 2001. Effect oftemperature on Ca2� cycling in rainbow trout myocytes. Bio-phys Soc Annu Meet Abstr 646a.
———. 2002a. The force-frequency relationship in fish hearts:a review. Comp Biochem Physiol A 132:811–826.
———. 2002b. Temperature dependence of cardiac sarcoplas-mic reticulum function in rainbow trout myocytes. J ExpBiol 205:3631–3639.
———. 2003. Acute temperature change modulates the re-sponse of I Ca to adrenergic stimulation in fish cardiomyo-cytes. Physiol Biochem Zool 76:816–824.
Shiels H.A. and E. White. 2005. Temporal and spatial propertiesof cellular Ca2� flux in trout ventricular myocytes. Am JPhysiol 288:R1756–R1766.
Sitsapesan R., R.A.P. Montgomery, K.T. Macleod, and A.J. Wil-liams. 1991. Sheep cardiac sarcoplasmic reticulum calcium-

290 G. L. J. Galli, H. A. Shiels, and R. W. Brill
release channels: modification of conductance and gating by
temperature. J Physiol 434:469–488.
Tiitu V. and M. Vornanen. 2001. Cold adaptation suppresses
the contractility of both atrial and ventricular muscle of the
crucian carp heart. J Fish Biol 59:141–156.
Vornanen M. 1989. Regulation of contractility of the fish (Car-
assius carassius L.) heart ventricle. Comp Biochem Physiol
C 94:477–483.
———. 1997. Sarcolemmal Ca influx through L-type Ca chan-
nels in ventricular myocytes of a teleost fish. Am J Physiol
41:R1432–R1440.
———. 1998. L-type Ca2�current in fish cardiac myocytes:
effects of thermal acclimation and beta-adrenergic stimula-tion. J Exp Biol 201:533–547.
Vornanen M. and V. Paajanen. 2004. Seasonality of dihydro-pyridine receptor binding in the heart of an anoxia-tolerantvertebrate, the crucian carp (Carassius carassius L.). Am JPhysiol 287:R1263–R1269.
Vornanen M., H.A. Shiels, and A.P. Farrell. 2002. Plasticity ofexcitation-contraction coupling in fish cardiac myocytes.Comp Biochem Physiol A 132:827–846.
Weng K.C., P.C. Castilho, J.M. Morrissette, A.M. Landeira-Fernandez, D.B. Holts, R.J. Schallert, K.J. Goldman, and B.A.Block. 2005. Satellite tagging and cardiac physiology revealniche expansion in salmon sharks. Science 310:104–106.
Copyright © 2022 FDOKUMEN




















