
The origin of conodonts and of vertebrate mineralized skeletons
Upload
independentCategory
view
0download
0
Placoderm Fishes, Pharyngeal Denticles, and theVertebrate DentitionZerina Johanson1* and Moya M. Smith2
1Palaeontology, Australian Museum, Sydney, NSW 2010, Australia2Department of Craniofacial Development, Dental Institute, Kings College London, Guy’s Campus,London SE1 9RT, UK
ABSTRACT The correlation of the origin of teeth withjaws in vertebrate history has recently been challengedwith an alternative to the canonical view of teeth derivingfrom separate skin denticles. This alternative proposesthat organized denticle whorls on the pharyngeal (gill)arches in the fossil jawless fish Loganellia are precursorsto tooth families developing from a dental lamina alongthe jaw, such as those occurring in sharks, acanthodians,and bony fishes. This not only indicates that homologs oftooth families were present, but also illustrates that theypossessed the relevant developmental controls, prior tothe evolution of jaws. However, in the Placodermi, a phy-logenetically basal group of jawed fishes, the state of pha-ryngeal denticles is poorly known, tooth whorls are ab-sent, and the presence of teeth homologous to those inextant jawed fishes (Chondrichthyes � Osteichthyes) iscontroversial. Thus, placoderms would seem to providelittle evidence for the early evolution of dentitions, or ofdenticle whorls, or tooth families, at the base of the cladeof jawed fishes. However, organized denticles do occur atthe rear of the placoderm gill chamber, but are associatedwith the postbranchial lamina of the anterior trunkshield,assumed to be part of the dermal cover. Significantly,these denticles have a different organization and morphol-ogy relative to the external dermal trunkshield tubercles.We propose that they represent a denticulate part of thevisceral skeleton, under the influence of pharyngeal pat-terning controls comparable to those for pharyngeal den-ticles in other jawed vertebrates and Loganellia. J. Mor-phol. 257:289–307, 2003. © 2003 Wiley-Liss, Inc.
KEY WORDS: Placodermi; jawed fishes; denticles; teeth;endoderm; neural crest
The Placodermi (Fig. 1) branch from the basalnode in the clade of jawed fishes (Young, 1986; Jan-vier, 1996, 2001) and are therefore crucial in inter-preting the phylogenetic development of jaws andteeth. However, the origins and developmental rela-tionship between teeth and jaws in vertebrate his-tory has recently been questioned, as has the axiomthat oral and pharyngeal denticles evolved via apositional shift of the external skin denticles into theoropharyngeal cavity (Smith and Coates, 1998,2000, 2001). One particular example of a mineral-ized oropharyngeal feeding arrangement in an ag-nathan (jawless) vertebrate is the conodont appara-tus, with enamel and dentine homologs (Sansom et
al., 1992). This apparatus consists of symmetricallyarranged composite structures used as biting ele-ments (Briggs et al., 1983; Aldridge and Purnell,1996; Donoghue et al., 1998; Smith and Coates,2000), while the conodont animal itself lacks anymineralized dermal covering. Thus, the conodontapparatus could not have been derived from exter-nal skin denticles (Aldridge et al., 1986; Smith andCoates, 1998, 2000, 2001).
An alternative theory to the classical view thatoropharyngeal denticles were derived secondarilyfrom external skin denticles moving into the mouthsuggests they evolved independently (Smith andCoates, 1998). In this theory, sets of denticles on thepharyngeal (gill) arches are proposed as precursorsof the organized tooth families developing within aspecialized dental epithelium (dental lamina) alongthe jaw margin (Reif, 1982; Smith and Hall, 1993;Smith et al., 1996; Smith and Coates, 1998, 2000,2001). This alternative theory was based on obser-vations of homologous arrays of denticle whorls oc-curring within the pharyngeal region of jawlessfishes, such as the thelodont Loganellia (van derBrugghen and Janvier, 1993) and comparisons totooth families of early chondrichthyans (Smith andCoates, 1998, 2000, 2001) (Fig. 2). These denticlewhorls showed several of the characteristics of toothfamilies found at the jaw margins of a variety ofsharks and of tooth whorls in the symphyseal regionof acanthodians and bony fishes (osteichthyans).Further, Smith and Coates (2001:224) proposed thatthe developmental processes producing the orga-nized denticles on the gill arches of agnathans suchas Loganellia were “co-opted” for the mandibulararch to produce the replacement tooth families.
Contract grant sponsors: Australian Research Council Fellowship(to ZJ), Royal Society research and travel grants (to MMS).
*Correspondence to: Zerina Johanson, Palaeontology, AustralianMuseum, 6 College Street, Sydney, NSW 2010, Australia.E-mail: [email protected]
DOI: 10.1002/jmor.10124
JOURNAL OF MORPHOLOGY 257:289–307 (2003)
© 2003 WILEY-LISS, INC.

These tooth families are characteristic of all chon-drichthyans, as are the statodont (nonreplacing)sets in tooth whorls of acanthodians and bony fishes.Thus, this prepattern for the dentition was indepen-dent of the presence of jaws, since it was alreadypresent in jawless fishes.
Phylogenetically, placoderms are the first verte-brates to possess jaws (Janvier, 1996), but althoughthere is very good fossil material of the gnathal
bones bearing denticles or teeth (O� rvig, 1973, 1980),the nature of the placoderm dentition is controver-sial. This includes whether or not the group pos-sessed true teeth developed within a dental lamina(Reif, 1982; Denison, 1978; Young, 1986; Smith andCoates, 1998; Donoghue et al., 2000; Donoghue andSmith, 2001; Young et al., 2001). Furthermore, thereare very few fossils with the pharyngeal gill archespreserved, so this part of the visceral skeleton is
Fig. 1. Cladogram showingphylogenetic relationships withinthe Placodermi (adapted fromGoujet, 2001), with examples ofdenticulation/tubercles on thepostbranchial lamina of variousgroups, including the Acantho-thoraci (A: Weejasperaspis [Fig.10]; B: Brindabellaspis [Fig.6D,E]; C: Romundina [Fig. 8]),Ptyctodontida (Fig. 6A), Actinol-epida (D: Aethaspis [Denison,1958]), and the Brachythoraci(Fig. 4B,C). Ordered rows of den-ticles also occur in the Rhe-nanida (Lelievre et al., 1995)and phylogenetically basal anti-archs (Zhang, 1980).
290 Z. JOHANSON AND M.M. SMITH
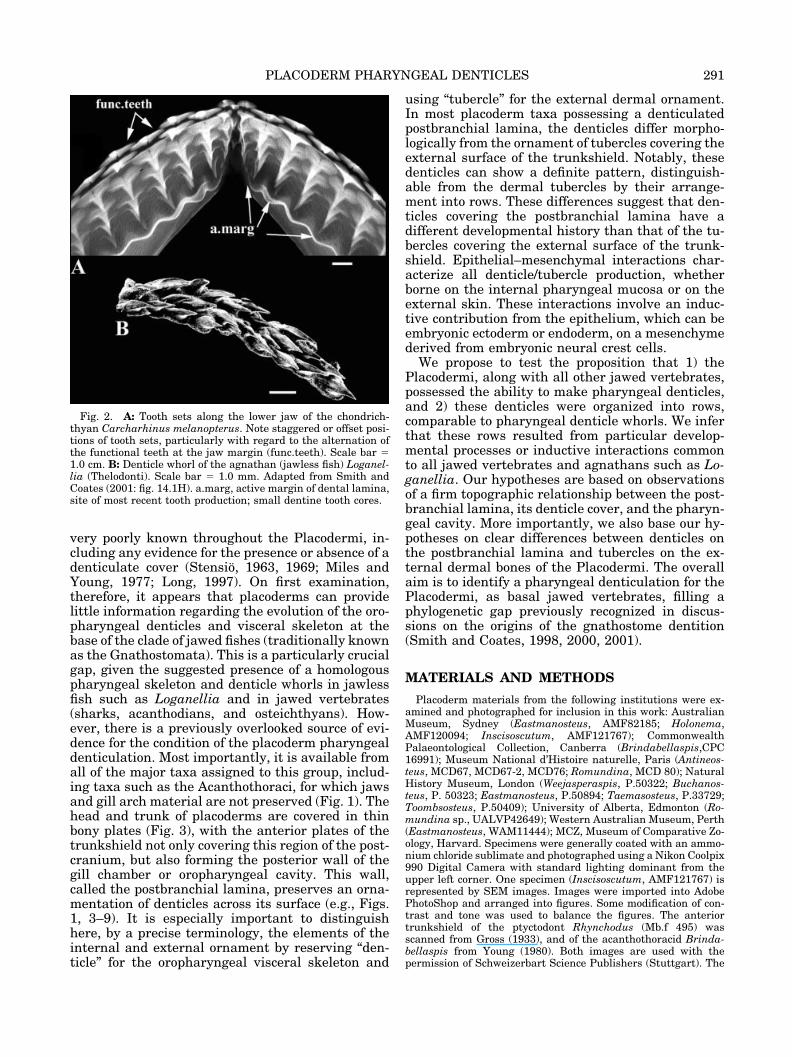
very poorly known throughout the Placodermi, in-cluding any evidence for the presence or absence of adenticulate cover (Stensio, 1963, 1969; Miles andYoung, 1977; Long, 1997). On first examination,therefore, it appears that placoderms can providelittle information regarding the evolution of the oro-pharyngeal denticles and visceral skeleton at thebase of the clade of jawed fishes (traditionally knownas the Gnathostomata). This is a particularly crucialgap, given the suggested presence of a homologouspharyngeal skeleton and denticle whorls in jawlessfish such as Loganellia and in jawed vertebrates(sharks, acanthodians, and osteichthyans). How-ever, there is a previously overlooked source of evi-dence for the condition of the placoderm pharyngealdenticulation. Most importantly, it is available fromall of the major taxa assigned to this group, includ-ing taxa such as the Acanthothoraci, for which jawsand gill arch material are not preserved (Fig. 1). Thehead and trunk of placoderms are covered in thinbony plates (Fig. 3), with the anterior plates of thetrunkshield not only covering this region of the post-cranium, but also forming the posterior wall of thegill chamber or oropharyngeal cavity. This wall,called the postbranchial lamina, preserves an orna-mentation of denticles across its surface (e.g., Figs.1, 3–9). It is especially important to distinguishhere, by a precise terminology, the elements of theinternal and external ornament by reserving “den-ticle” for the oropharyngeal visceral skeleton and
using “tubercle” for the external dermal ornament.In most placoderm taxa possessing a denticulatedpostbranchial lamina, the denticles differ morpho-logically from the ornament of tubercles covering theexternal surface of the trunkshield. Notably, thesedenticles can show a definite pattern, distinguish-able from the dermal tubercles by their arrange-ment into rows. These differences suggest that den-ticles covering the postbranchial lamina have adifferent developmental history than that of the tu-bercles covering the external surface of the trunk-shield. Epithelial–mesenchymal interactions char-acterize all denticle/tubercle production, whetherborne on the internal pharyngeal mucosa or on theexternal skin. These interactions involve an induc-tive contribution from the epithelium, which can beembryonic ectoderm or endoderm, on a mesenchymederived from embryonic neural crest cells.
We propose to test the proposition that 1) thePlacodermi, along with all other jawed vertebrates,possessed the ability to make pharyngeal denticles,and 2) these denticles were organized into rows,comparable to pharyngeal denticle whorls. We inferthat these rows resulted from particular develop-mental processes or inductive interactions commonto all jawed vertebrates and agnathans such as Lo-ganellia. Our hypotheses are based on observationsof a firm topographic relationship between the post-branchial lamina, its denticle cover, and the pharyn-geal cavity. More importantly, we also base our hy-potheses on clear differences between denticles onthe postbranchial lamina and tubercles on the ex-ternal dermal bones of the Placodermi. The overallaim is to identify a pharyngeal denticulation for thePlacodermi, as basal jawed vertebrates, filling aphylogenetic gap previously recognized in discus-sions on the origins of the gnathostome dentition(Smith and Coates, 1998, 2000, 2001).
MATERIALS AND METHODS
Placoderm materials from the following institutions were ex-amined and photographed for inclusion in this work: AustralianMuseum, Sydney (Eastmanosteus, AMF82185; Holonema,AMF120094; Inscisoscutum, AMF121767); CommonwealthPalaeontological Collection, Canberra (Brindabellaspis,CPC16991); Museum National d’Histoire naturelle, Paris (Antineos-teus, MCD67, MCD67-2, MCD76; Romundina, MCD 80); NaturalHistory Museum, London (Weejasperaspis, P.50322; Buchanos-teus, P. 50323; Eastmanosteus, P.50894; Taemasosteus, P.33729;Toombsosteus, P.50409); University of Alberta, Edmonton (Ro-mundina sp., UALVP42649); Western Australian Museum, Perth(Eastmanosteus, WAM11444); MCZ, Museum of Comparative Zo-ology, Harvard. Specimens were generally coated with an ammo-nium chloride sublimate and photographed using a Nikon Coolpix990 Digital Camera with standard lighting dominant from theupper left corner. One specimen (Inscisoscutum, AMF121767) isrepresented by SEM images. Images were imported into AdobePhotoShop and arranged into figures. Some modification of con-trast and tone was used to balance the figures. The anteriortrunkshield of the ptyctodont Rhynchodus (Mb.f 495) wasscanned from Gross (1933), and of the acanthothoracid Brinda-bellaspis from Young (1980). Both images are used with thepermission of Schweizerbart Science Publishers (Stuttgart). The
Fig. 2. A: Tooth sets along the lower jaw of the chondrich-thyan Carcharhinus melanopterus. Note staggered or offset posi-tions of tooth sets, particularly with regard to the alternation ofthe functional teeth at the jaw margin (func.teeth). Scale bar �1.0 cm. B: Denticle whorl of the agnathan (jawless fish) Loganel-lia (Thelodonti). Scale bar � 1.0 mm. Adapted from Smith andCoates (2001: fig. 14.1H). a.marg, active margin of dental lamina,site of most recent tooth production; small dentine tooth cores.
291PLACODERM PHARYNGEAL DENTICLES

radiograph of a shark jaw and a branchial denticle whorl ofLoganellia was previously published by Smith and Coates (2001),reproduced here with permission of Taylor and Francis, London.
Two images of the placoderm Eastmanosteus (Fig. 3A,C) arereduced in size and repeated in subsequent figures to aid invisualizing the orientation of the isolated postbranchial laminaeillustrated in these figures. The phylogenetic relationships withinthe Placodermi, as recently described (Goujet and Young, 1995;Goujet, 2001), provide the framework for our observations (Fig.1). Particular emphasis is placed on the basal group Acanthotho-raci, as well as the more derived group Arthrodira, as taxa illus-trating a range of postbranchial lamina denticle morphologies.
RESULTS
The placoderm interolateral is a trunkshield platepossessing both a ventral external lamina as well asa dorsomedial flange, the latter separating the gillchamber from the internal regions of the trunk-shield (IL (pbl), trunk, Fig. 3). A medial flange fromthe more dorsal anterior lateral plate can also con-tribute to the separation of the gill chamber from theinternal trunkshield (AL (pbl), Fig. 6A). These
Fig. 3. A,B: Lateral (A) and anterior (B) views of the head and trunkshield of Eastmanosteus (Arthrodira), AMF82185. Blackarrows (A) indicate posterior opening into gill chamber between the headshield (headsh) and the trunkshield (trunk). In B, the gillchamber is located in the region between the infragnathals (Ifg) and the interolateral plate and associated postbranchial lamina(IL(pbl)). C: WAM11444, anterior view of Eastmanosteus trunkshield, with headshield, as shown in B, removed. ADL, anteriordorsolateral plate; AL, anterior lateral plate; orb, orbit; trunk(visceral), visceral or internal region of trunkshield. Asterisk indicatesdorsomedial margin of the postbranchial lamina and the location of the ‘active margin’ of denticle addition. Scale bar � 1.0 cm.
292 Z. JOHANSON AND M.M. SMITH

flanges together form the placoderm postbranchiallamina. The external trunkshield surfaces are orna-mented by tubercles, or bony ridges in nearly allplacoderms, while the postbranchial lamina is den-ticulated to varying degrees in all groups.
The denticulation on the postbranchial wall of thegill chamber varies throughout the placoderm clade,but in most taxa it is morphologically different fromtubercles on the external surface of the trunkshield(Figs. 3–9; large white arrows in Figs. 4A,C, 5B, 7C,tub). The postbranchial lamina denticles possess avery similar morphology, generally triangular inshape, with the apex of the triangle pointing in auniform direction, ventrally (Figs. 4, 5, 7, 9) or pos-terolaterally (Figs. 6D,E, 8). These denticles can bearranged in patterns of ordered rows and examplesof this morphology occur in every placoderm group(Fig. 1); Acanthothoraci (Figs. 6D,E, 8, 9A,C); Anti-archi (Zhang, 1980; Zhu, 1996); Rhenanida (Lelievreet al., 1995); Ptyctodontida (Fig. 6A); Arthrodira(Figs. 3–5, 7; also Denison [1958; fig. 1D] and John-son et al. [2000] for actinolepid arthrodires; Goujet[1984] for phlyctaeniid arthrodires). The denticlerows extend transversely across the postbranchiallamina and can be either ordered into rows of stag-gered or offset denticles (Figs. 4A,B, 7A,B, 8, 9), orarranged in separated rows (Figs. 4C, 5, 6A). Thesize of denticles in the rows often varies across thepostbranchial lamina (Figs. 4A,C, 6A, white arrows),with smaller denticles occurring where the post-branchial lamina joins the external lamina, andlarger denticles dorsally. This distribution is relatedto the growth of the individual as the size of thepostbranchial lamina increases. The denticle rowsare oriented parallel to the dorsomedial margin ofthe interolateral/anterior lateral plates (Figs. 4, 5,7A,B, 8, 9), and in the arthrodire Holonema (Fig. 5A)denticle rows are steeper and shorter near the baseof the lamina. These become longer and straightertowards the dorsomedial margin, showing how theshape of the postbranchial lamina has changedthrough growth.
New denticles originate along this dorsomedialmargin of the postbranchial lamina (asterisk, Fig.3C; a.marg, Figs. 4–8). Entire rows of denticles arenot added in one generative event, but instead areadded separately (Figs. 4A,B, 6, 7A,B, 8, 9) or as partof a shortened row (Figs. 4C, 5). This is clear indorsal views of the lamina margin, showing the mostrecent denticles added to the bone (Figs. 5D, 7B).When new denticles are added separately, each oneis added to the end of a diagonal row (Fig. 7B, largerwhite arrows) such that they occur in staggered oralternating positions relative to denticles of the pre-vious rows (Figs. 4, 7, 8). When added in shortgroups, the denticles are eventually incorporated tothe row extending across the lamina (Figs. 4C,5B,C). There is a distinct new contact surface be-tween the bone of the lamina and the denticles (Fig.5D, black arrowheads). Also, when denticles are lost
in specimen preparation this illustrates their sepa-rate formation from the remaining smooth surface ofthe underlying lamina bone, as clearly indicated inthe acanthothoracid Romundina (Fig. 9A, black ar-rows) and the arthrodire Antineosteus (Fig. 9B,black/white arrows). A thin layer of bone forms withthe denticles and is lost with them, representing thebone of attachment associated with teeth or den-ticles attaching to basal bone. This is represented inRomundina by a shallow depression (Fig. 9C,b.att(dep)), while the bone itself can be seen sur-rounding denticles remaining on the postbranchial.The margins of the attachment bone are visible (Fig.9C, b.att, black arrows), and the bone is pierced bysmall canal openings, differentiated by these fromthe basal supporting bone (Fig. 5D).
There is significant variation in denticle patternwithin the Placodermi, particularly in the taxa nearthe base of the clade (Goujet and Young, 1995; Gou-jet, 2001). For example, the postbranchial lamina ofthe Acanthothoraci is well known and preserves aninteresting range of denticle morphologies (Fig. 1A–C). The acanthothoracids are represented by a vari-ety of taxa, including Romundina sp. (O� rvig, 1975;Figs. 8, 9A,C), Palaeacanthaspis (Stensio, 1944,1969), Brindabellaspis (Young, 1980: fig. 19, pl. 2.3–2.5) (Fig. 6D,E) and Weejasperaspis (Fig. 10), amongothers (e.g., Radotina and Kosoroaspis [Gross,1959]).
The anterior plates of the trunkshield are gener-ally fused in these acanthothoracid taxa and thepostbranchial lamina is extensive. The post-branchial lamina is covered in denticles in Ro-mundina, Palaeacanthaspis, and Brindabellaspis;these are triangular in shape with serrated edges(Figs. 6D,E, 8, 9A,C) (Stensio, 1944; White, 1952;O� rvig, 1975; Young, 1980), and again markedly dif-ferent from star-shaped tubercles on the externalsurface of the trunkshield (Fig. 8B,D,G) (O� rvig,1975; Young, 1980). In a previously unfigured trunk-shield assigned to Romundina sp., the lamina showsordered offset or staggered rows of denticles, inwhich each denticle is positioned between two in theprevious row (Fig. 8: this specimen is UALVP 42649from the Early Devonian MOTH locality, northernCanada, currently being described by CR Hermusand MVH Wilson, University of Alberta). The trian-gular denticles have a common orientation (postero-laterally directed) and are packed into the availablespace on the postbranchial lamina, with newer onesforming at the dorsomedial edge of the lamina (Fig.8C, a.marg). This may also be seen in the acantho-thoracid Palaeacanthaspis (Stensio, 1959, 1969: fig.189A) and is more comparable to the packed denticlerows seen in the arthrodires Buchanosteus, East-manosteus (Fig. 4A,B) and Antineosteus (Figs. 7A,B,9B; Lelievre, 1984), rather than the spaced rowsseen in the arthrodires Taemasosteus (Fig. 4C),Toombsosteus or Holonema (Fig. 5A–C).
293PLACODERM PHARYNGEAL DENTICLES
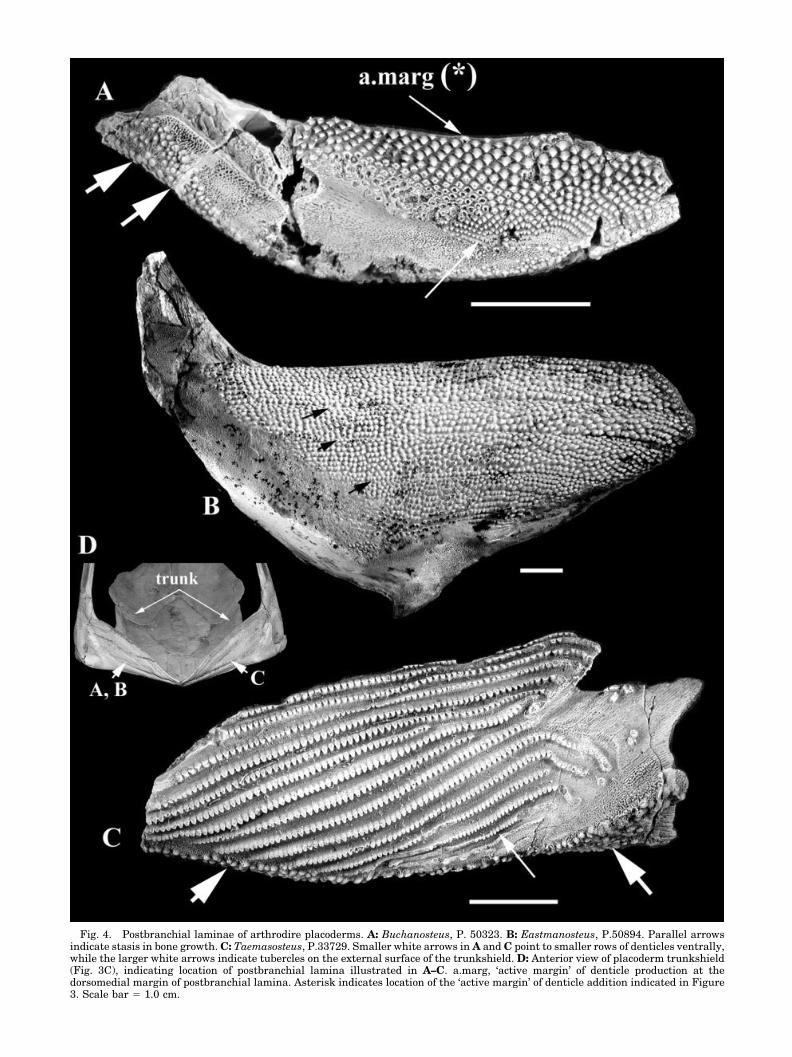
Fig. 4. Postbranchial laminae of arthrodire placoderms. A: Buchanosteus, P. 50323. B: Eastmanosteus, P.50894. Parallel arrowsindicate stasis in bone growth. C: Taemasosteus, P.33729. Smaller white arrows in A and C point to smaller rows of denticles ventrally,while the larger white arrows indicate tubercles on the external surface of the trunkshield. D: Anterior view of placoderm trunkshield(Fig. 3C), indicating location of postbranchial lamina illustrated in A–C. a.marg, ‘active margin’ of denticle production at thedorsomedial margin of postbranchial lamina. Asterisk indicates location of the ‘active margin’ of denticle addition indicated in Figure3. Scale bar � 1.0 cm.

In the acanthothoracid Brindabellaspis, the trian-gular denticles also possess a common posterolateralorientation on the postbranchial lamina (Young,1980) (Fig. 6D,E), but by contrast are sparsely dis-tributed and lack a clear arrangement or organiza-tion into rows. However, the boundaries of the
postbranchial lamina are recognizable in Brindabel-laspis, due to the morphological differences betweenthe more dorsal and anterior denticles representingthose of the postbranchial lamina (Fig. 6D), and thelateral and ventral tubercles representing the exter-nal trunkshield surface (Young, 1980). Otherwise,
Fig. 5. Postbranchial lami-nae of various arthrodire placo-derms, illustrating the positionof the dorsomedial ‘active mar-gin’ as the proposed location ofdenticle production. A,C,D: Ho-lonema, AMF120094. A: Post-branchial lamina, white andblack arrow indicating most re-cent positions of ‘active margin’(a.marg). Scale bar � 1.0 cm.C: Close-up of postbranchiallamina. Paired arrows indicate‘active margin’ and most recentdenticles added here, denticlerow marked by right arrow themost recent, and row marked byleft arrow representing the pre-vious denticle generating event.Arrow labeled ‘D’ indicates theview in D. Scale bar � 0.25 cm.D: Dorsal view of dorsomedial‘active margin’ showing den-ticles (dent) added in most re-cent denticle-generating event.Black arrowheads indicate con-tact between bone of post-branchial lamina (pbl) anddenticles. B: Toombsosteus,P.50409. Smaller white arrowsindicate ‘active margin’; largewhite arrow indicates location oftrunkshield tubercles. Scalebar � 1.0 cm. E,F: Inscisoscu-tum, AMF121767. E: Scalebar � 0.5 cm. F: SEM photo-graph of denticle addition topostbranchial lamina. Scalebar � 500 mm. G: Anterior viewof placoderm trunkshield (Fig.3C), indicating location of post-branchial lamina shown in A–C,E–F.
295PLACODERM PHARYNGEAL DENTICLES
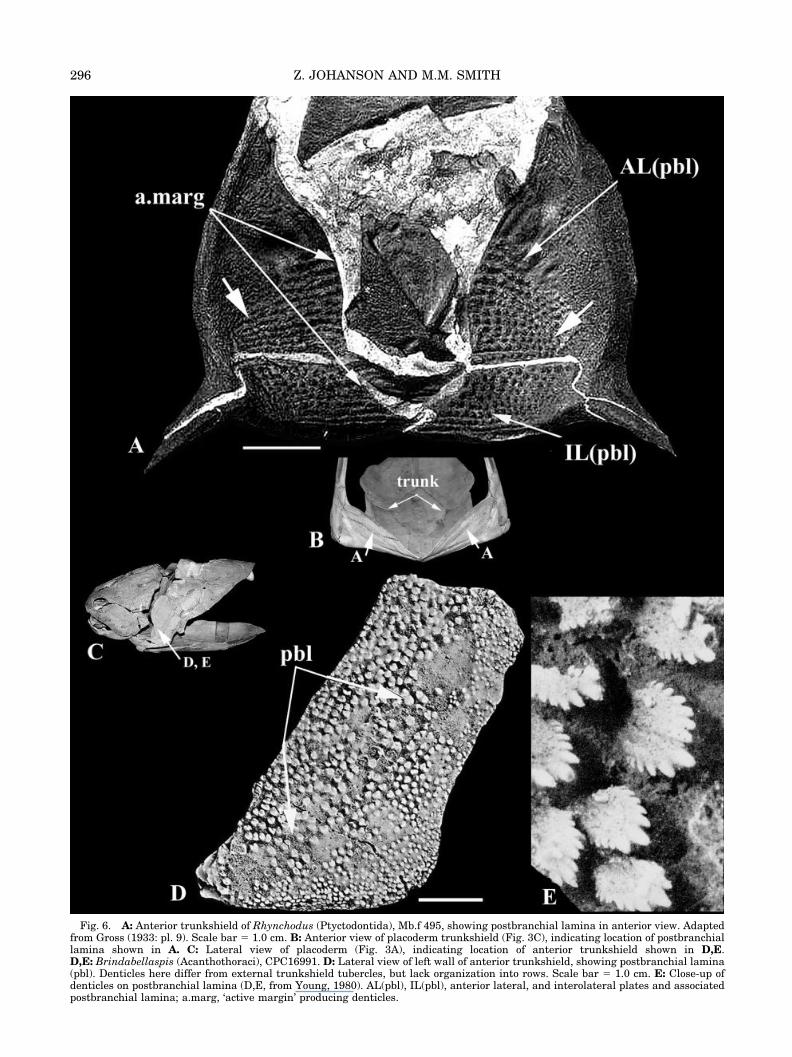
Fig. 6. A: Anterior trunkshield of Rhynchodus (Ptyctodontida), Mb.f 495, showing postbranchial lamina in anterior view. Adaptedfrom Gross (1933: pl. 9). Scale bar � 1.0 cm. B: Anterior view of placoderm trunkshield (Fig. 3C), indicating location of postbranchiallamina shown in A. C: Lateral view of placoderm (Fig. 3A), indicating location of anterior trunkshield shown in D,E.D,E: Brindabellaspis (Acanthothoraci), CPC16991. D: Lateral view of left wall of anterior trunkshield, showing postbranchial lamina(pbl). Denticles here differ from external trunkshield tubercles, but lack organization into rows. Scale bar � 1.0 cm. E: Close-up ofdenticles on postbranchial lamina (D,E, from Young, 1980). AL(pbl), IL(pbl), anterior lateral, and interolateral plates and associatedpostbranchial lamina; a.marg, ‘active margin’ producing denticles.
296 Z. JOHANSON AND M.M. SMITH

the postbranchial lamina appears to lie in the sameplane as the rest of the cleithrum. In the acantho-thoracid Weejasperaspis, the dorsomedial regions ofthe postbranchial lamina lack denticles (Fig. 10E).The scattering of tubercles on the more lateral parts
of the postbranchial lamina shows two morpholo-gies, and both may represent different tubercleforms also found on the external dermal trunk-shield. The first tubercle morphology, representedby the larger white arrows in Figure 10E, illustrates
Fig. 7. A–C: Antineosteus(Arthrodira). A,B: MCD 67.A: Interolateral plate showingpostbranchial lamina (pbl) in an-terior view. Parallel arrows indi-cate stasis in bone growth.B: Close-up of denticles on post-branchial lamina in A. Large ar-rows show new individual den-ticles being added at end ofdiagonal rows, offset betweentwo denticles of previous row.C: MCD 76, ventral view show-ing external surface of interolat-eral plate covered in sparse, ran-domly positioned tubercles (tub),and postbranchial lamina cov-ered in triangular denticles(dent). Note line of denticlesmarking the margin separatingthese surfaces. a.marg, ‘activemargin’ of denticle production atthe dorsomedial margin of post-branchial lamina. D: Anteriorview of placoderm trunkshield(Fig. 3C), indicating location ofpostbranchial lamina shown inA–C (the smaller arrow in D as-sociated with ‘C(tub)’ refers tothe ventral, external surface ofthe trunkshield). Scale bar � 1.0cm (A), � 1 mm (B,C).
297PLACODERM PHARYNGEAL DENTICLES

Fig. 8. A–G: Romundina sp. (Acanthothoraci), UALVP42649, external view of left anteriortrunkshield. Scale bar in central image � 0.5 cm. a.marg, ‘active margin’ of denticle production atthe dorsomedial margin of postbranchial lamina. H: Lateral view of placoderm (Fig. 3A), indicat-ing location of anterior trunkshield shown in A–G.
298 Z. JOHANSON AND M.M. SMITH

elongate tubercles that appear to have “wrappedaround” the lateral edge of the postbranchial lam-ina. Also present on the external face of the trunk-shield are small, star-shaped tubercles representingthe second morphology (small white arrow, Fig. 10B;
also White, 1978: pl. 3). These may represent earlierstages in the morphological development of the tu-bercular ornament in Weejasperaspis, later formingthe elongate types. These smaller, star-shapedforms are also present on the lateral portion of the
Fig. 9. A,C: Romundina (Acanthothoraci), MCD 80. A: Anterior view of postbranchial lamina; black arrows indicate smooth bonesurfaces where denticles have been lost during preparation of the specimen. Scale bar � 0.5 cm. C: Close-up of denticles showing boneof attachment (b.att) associated with each denticle (dent). Black arrows point to edges of bone. The bone itself is pierced by small canalopenings. Note difference in size between these and pores in laminar bone. Also, a shallow depression (b.att(dep)) results from the lossof the bone attachment associated with lost denticle. B: Antineosteus (Arthrodira), MCD 67-2, close-up of denticles covering post-branchial lamina. Black/white arrows show bare smooth bone where denticles have been lost. D: Anterior view of placodermtrunkshield (Fig. 3C), indicating location of postbranchial lamina shown in A–C. Scale bar � 1.0 cm.
299PLACODERM PHARYNGEAL DENTICLES
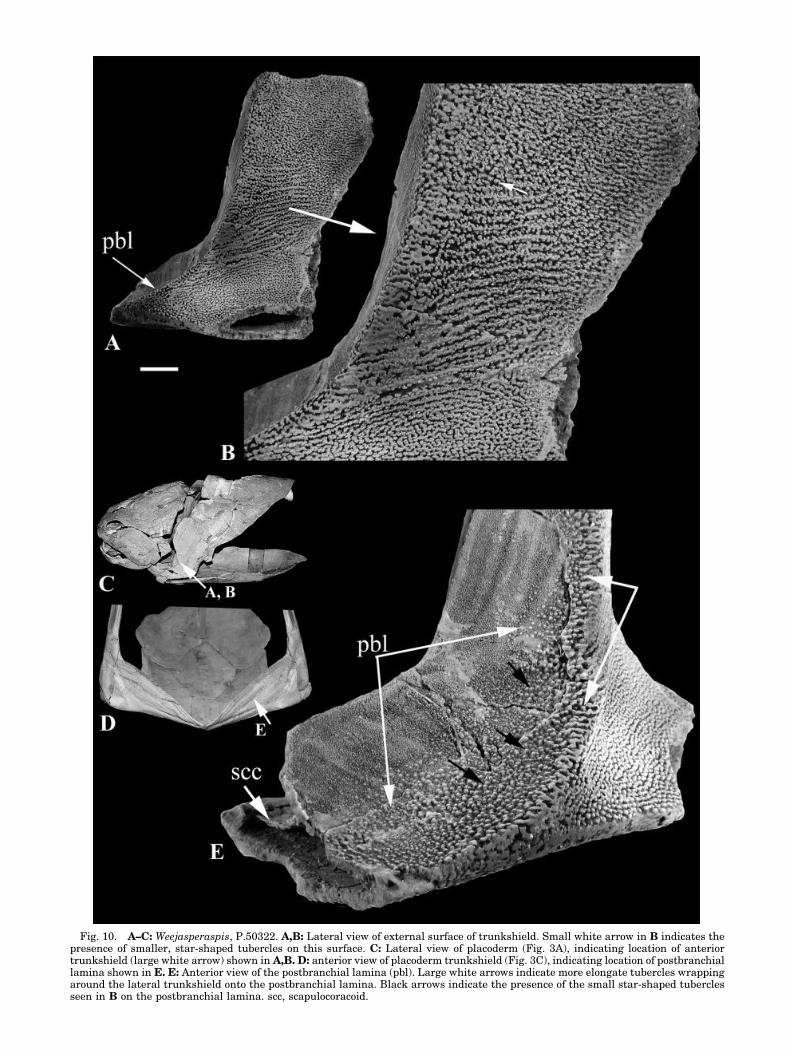
Fig. 10. A–C: Weejasperaspis, P.50322. A,B: Lateral view of external surface of trunkshield. Small white arrow in B indicates thepresence of smaller, star-shaped tubercles on this surface. C: Lateral view of placoderm (Fig. 3A), indicating location of anteriortrunkshield (large white arrow) shown in A,B. D: anterior view of placoderm trunkshield (Fig. 3C), indicating location of postbranchiallamina shown in E. E: Anterior view of the postbranchial lamina (pbl). Large white arrows indicate more elongate tubercles wrappingaround the lateral trunkshield onto the postbranchial lamina. Black arrows indicate the presence of the small star-shaped tuberclesseen in B on the postbranchial lamina. scc, scapulocoracoid.

postbranchial lamina (Fig. 10E, black arrows).Therefore, comparable tubercles are present on boththe external trunkshield and the postbranchial lam-ina of Weejasperaspis and, importantly, denticlesare absent. Moreover, the tubercles are associatedwith the ventrolateral sections of the lamina, sug-gesting production here at the dermal margin,rather than at the dorsomedial margin of the post-branchial lamina. This one example represents asignificant difference from the predominant ar-rangement of the denticles characteristic of mostplacoderm taxa (e.g., Figs. 3–9). These arrange-ments include morphologically distinct denticleswith a common orientation but no ordering into rows(e.g., Brindabellaspis), distinct denticles, uniformorientation, and ordering into staggered rows (Buch-anosteus, Antineosteus, Eastmanosteus) and order-ing into rows separated by bone of the postbranchiallamina (Holonema, Taemasosteus, Toombsosteus).
DISCUSSION
Although the pharyngeal region of the fossil jawedvertebrate group Placodermi is poorly known, den-ticles are present at the rear of the pharyngealcavity, associated with the anterior face of the post-branchial lamina. This medially inflected trunk-shield wall separates the pharyngeal cavity from thetrunk, and so the anterior face of the lamina formsthe rear wall to this cavity. All major placodermgroups possess a denticulated surface on the post-branchial lamina, although there is notable varia-tion. In most placoderms, these denticles differ mor-phologically, and in terms of organization, fromthose on the external surface of the trunkshield. Theremainder of this section compares these denticlesto patterned teeth/denticles in other jawed verte-brates and Loganellia, in terms of general morphol-ogy and the developmental processes responsible forthese morphologies. Although a fossil group, struc-tural patterning features within the Placodermi al-low us to make inferences regarding their proposeddevelopment.
Several structural characteristics of a tooth family(set) in jawed vertebrates, in which each tooth set isformed from a dental primordium, were identified bySmith and Coates (1998, 2000, 2001). The primor-dium is the generative organ for teeth, including aninductive oral epithelial infolding (dental lamina)together with a tooth-producing mesenchyme, situ-ated along the jaw margin (Reif, 1982). The struc-tural characteristics of these tooth sets were alsoobserved in denticle whorls along the gill arches inthe pharyngeal cavity of chondrichthyans and taxasuch as the agnathan Loganellia (Fig. 2), includingpolarity of shape, sequential timing, and unidirec-tionality of addition of teeth/denticles. The timing oftooth or denticle initiation in each set is controlledautonomously by cells of the dental primordium(Smith and Coates, 2001:236).
Some of these characteristics can also explain therows of denticles on the placoderm postbranchiallamina (Figs. 3–9). The structural features includepolarity of shape (resulting in a common orienta-tion), unidirectionality of addition, sequential initi-ation of each denticle (or groups of denticles), andspacing at specific positions along the dorsomedialmargins of the postbranchial lamina to form theuniform rows. In taxa such as Buchanosteus, East-manosteus (Fig. 4A,B), Antineosteus (Fig. 7A,B), andRomundina (Fig. 8), the patterning, or ordering, ofdenticle addition involves offset individual denticlesin closely adjacent rows (e.g., new denticle added tothe end of a diagonal row and positioned betweentwo in previous row; Fig. 7B). In taxa such as Tae-masosteus (Fig. 4C), Toombsosteus, Holonema, andIncisoscutum (Fig. 5), several denticles are added (asa group or short row) and incorporated into contin-uous rows separated from one another by the bone ofthe lamina. These morphologies imply a control toboth the location and timing of denticle addition, butare also dependent on bone growth as seen clearly inphases of addition and stasis in Eastmanosteus andAntineosteus (Figs. 4B, 7A, parallel arrows on lam-ina). Differences in denticle sizes from the base (Fig.4A, white arrow) of the postbranchial lamina to thedorsomedial edge (Figs. 3C, 4A, asterisk, a.marg)suggest that this denticle addition occurredthroughout the life of the individual. Importantly,by contrast with the ornament in the external skin,new denticles are not added randomly within previ-ously formed rows, nor are they superimposed, asmay happen in the tuberculate dermal ornament(Gross, 1957: fig. 9E). They also form with their ownbone of attachment separate from the bone of thepostbranchial lamina, as seen in accidental prepar-ative loss of patches of the denticulation (Fig. 9A,B).
All denticles and tubercles are formed fromepithelial–mesenchymal interactions (Schaeffer,1977; Reif, 1982; Smith and Hall, 1990, 1993)whether generated externally on the dermal bonesurface (tubercles) or internally in the oropharyn-geal cavity (denticles). The addition of new denticlesexclusively to the dorsomedial margins of the post-branchial lamina (e.g., Figs. 3, 4, 5, 7B, 8B) suggeststhat this was where the relevant primordial ordenticle-forming tissues were located in the soft tis-sue space at the bone margin. This is comparable tothe addition of new teeth from the dental primor-dium along the lingual margin of the shark dentallamina (Reif, 1982; Smith and Coates, 2000: fig.10B) (Fig. 2, a.marg). The dorsomedial margin of thepostbranchial lamina is termed an “active margin”(a.marg, Figs. 4–9) to reflect this ongoing process.Moreover, generative tissues at this margin (wellremoved from the external surface of the trunk-shield, even in younger, smaller individuals) notonly produce denticles differing morphologicallyfrom tubercles comprising the trunkshield orna-ment, but also provide denticles with an indepen-
301PLACODERM PHARYNGEAL DENTICLES

dent pattern relative to the trunkshield tubercles.These differences suggest that the tissues responsi-ble for denticle formation on the postbranchial lam-ina differ, in their embryonic origin (cell lineage),from those on the external surface producing tuber-cles on the head and trunkshield, as indeed do thoseproducing teeth in the oral cavity. The followingsection attempts to provide a developmental expla-nation for these differences, based on comparativemorphology of the denticles on the placoderm post-branchial lamina.
Developmental Processes
Discussion of putative developmental processeswith regards to the fossil group Placodermi can beextrapolated from the contrasting denticle morphol-ogy and arrangement on the postbranchial laminarelative to that of the tubercles on the external headand trunkshield surfaces (e.g., Antineosteus, Fig.7C). The unique denticle morphology and the pat-terning of these denticles into rows on the placodermpostbranchial lamina would have resulted from de-velopmental processes specific to this region, rela-tive to the external trunkshield. We infer thesedevelopmental processes associated with the placo-derm pharyngeal cavity, based on similarities be-tween the arrangement of denticles on the placo-derm postbranchial lamina and the tooth familiesand denticle whorls of other jawed vertebrates, interms of the “structural characteristics” describedabove.
Both oropharyngeal denticles and teeth are initi-ated as the result of inductive interactions betweenepithelium and ectomesenchyme (neural crest: e.g.,Schaeffer, 1977; Reif, 1982; Smith and Hall, 1990,1993). The contribution of an ectodermally derivedepithelium to this inductive interaction is assumedfor external skin tubercles, but the contribution ofan endodermal epithelium to the equivalent processinternally is less accepted, as has lately been em-phasized (Smith and Coates, 1998). The necessarycontribution of the endoderm to tooth induction wasoriginally noted for amphibians (Sellman, 1946; re-viewed in Smith and Hall, 1990, 1993; also Gravesonet al., 1997; Smith and Coates, 1998, 2000, 2001)and recent work has identified the involvement ofendoderm in the induction of mammalian teeth(Imai et al., 1998; see also review by Smith, 2003).Although mammalian teeth are restricted to the oralcavity, the functional teeth in pharyngognaths suchas the zebrafish occur exclusively on the last gillarch in the pharyngeal cavity. From the topographicposition of these gill arch teeth (Van der Heyden andHuysseune, 2000), it seems logical to propose theoperation of the same inductive endodermal source,as the inner surface of the pharyngeal cavity wasalmost certainly lined by endoderm. Additionally,one aspect emphasized in the research above wasthe initiation of tooth development where endoderm
and ectoderm meet (e.g., Imai et al., 1998; Smith andCoates, 2001). This boundary occurs between thepharyngeal pouches and gill slits, with internalendoderm and external ectoderm meeting in thisregion (Smith and Coates, 1998; Graham, 2001). Thelocation of jawed vertebrate pharyngeal denticlewhorls is also along the inner surface of the pharyn-geal cavity and the whorls can be associated withthe gill slits in various taxa (Nelson 1969: figs. 9,14–16). These show a comparable arrangement andpatterned addition relative to the zebrafish gill archteeth (Smith and Hall, 1993; Smith and Coates,1998, 2001).
Similarly, we suggest that the inductive influenceof endoderm could be involved in the initiation ofdenticles associated with the placoderm post-branchial lamina. In this hypothesis, the laminaitself, placed medially at the rear of the pharyngealcavity, would be covered by an epithelium at least inpart derived from endoderm, or on which the bound-ary between embryonic ectoderm and endoderm waslocated. The initial “active margin,” in the earlieststages, was located along the dorsomedial edge ofthe lamina and putatively represents this boundarybetween ectoderm and the inductive endodermalsource of denticle production. Most notably, theendodermal influence in denticle developmentwould be absent from the external surface of thehead/trunkshield plate, providing a potential expla-nation for differences in denticle morphologies andarrangement between these two surfaces.
We would suggest that the endoderm providedboth morphological and patterning information tothe denticles at the dorsomedial “active margin” ofthe placoderm postbranchial lamina. However, theacanthothoracid Brindabellaspis lacks an organiza-tion of denticles into rows, although importantly, thedenticles themselves differ morphologically from tu-bercles on the external surface of the trunkshield(Fig. 6D,E). Thus, an additional explanation may besought to explain patterning and morphological dif-ferences of the denticles. Two sources of neuralcrest-derived mesenchyme (ectomesenchyme) havebeen identified, and differences between denticlesand tubercles might be attributed to whether thesource of ectomesenchyme is cranial or trunk. Pla-coderm trunkshield plates are exoskeletal in originand formed of dermal bone which, in common withall exoskeletal structures, is generally believed tohave formed from ectomesenchyme (e.g., Smith andHall, 1990); however, whether the ectomesenchymeis derived from the cranial or trunk neural crest hasnever been established (Smith and Hall, 1993). Al-though research on the contribution of trunk neuralcrest to dermal bone in fish is limited, Smith et al.(1994) did establish a contribution of trunk crest-derived tissue to the caudal, dermal fin ray skeletonin zebrafish, with in vitro manipulation demonstrat-ing the potential of chick trunk crest to make bone(McGonnell and Graham, 2002), and this would cer-
302 Z. JOHANSON AND M.M. SMITH

tainly include the placoderms. All inferred thattrunk ectomesenchyme possessed the ability to formbone in the earliest vertebrates (also Graveson et al.,1997). If the placoderm trunkshield plates and thetubercles covering these plates were formed bytrunk ectomesenchyme, then the postbranchial lam-ina, especially the denticles, could by contrast havebeen produced by the cranial ectomesenchyme.
Cranial ectomesenchyme contributes to tooth andjaw formation (Chai et al., 2000) and to the forma-tion of the gill arches (Schilling and Kimmel, 1994;Piotrowski and Nusslein-Volhard, 2000; Grahamand Smith, 2001). We infer that it would also con-tribute to the teeth and denticles on these arches.Recent research has shown that cranial ectomesen-chyme can also contribute to structures posterior tothe gill arches. For example, Graveson et al. (1997)noted that with regard to the urodele amphibianneural crest, the tooth-producing potential of thecranial neural crest extended more posteriorly thanexpected, making it difficult to choose a criterion fora boundary between the cranial and trunk neuralcrest regions. This suggests that the tooth-producingcontribution of cranial neural crest need not be re-stricted but could continue posteriorly (e.g., onto thesurface of the rear wall of the gill chamber). Addi-tionally, Kontges and Lumsden (1996) noted thatthe connective tissue fascia of the cranial muscula-ture and the bone where this fascia attached to theskull were derived from the same cranial neuralcrest source. Certain of these cranial muscles attachto the pectoral girdle (rather than the skull), suchthat their attachment point on the girdle must alsobe derived from cranial neural crest (McGonnell,2001; McGonnell et al., 2001). This further indicatescranial neural crest involvement in epithelial–mesenchymal interactions posterior to the gillarches and in association with the postcranial skel-eton at the rear of the pharyngeal cavity.
Because placoderm fishes are known only fromfossils, the embryonic origins of epithelium and ec-tomesenchyme responsible for denticle generationon the postbranchial lamina cannot be directly ob-served. However, the placoderm postbranchial lam-ina, situated at the rear of the pharyngeal cavity, iscovered by denticles whose morphology and patternrequire explanation. The most salient point is thatin most placoderm taxa these denticles differ fromexternal trunkshield tubercles, suggesting a differ-ent embryonic derivation for both the epitheliumand mesenchyme involved in the denticle-formingepithelial–mesenchymal interactions. We suggestthat the laminar denticles in the majority of placo-derms would have been produced under the influ-ence of endoderm, inducing cranial ectomesenchyme(both accounting for differences between denticlesand tubercles). In other words, the placoderm post-branchial lamina denticles are produced internally,in the pharyngeal cavity, rather than externally aspart of the outer surfaces of the head and trunk-
shield plates. By comparison, the postbranchial lam-ina of the acanthothoracid placoderm Weejasperas-pis (Fig. 10) is covered with randomly scatteredtubercles (no patterning information) morphologi-cally similar to the tubercles on the external surfaceof the trunkshield. We suggest these tubercles, inthe absence of denticle formation on the post-branchial lamina, were derived from the sameepithelial–mesenchymal interaction responsible forthe dermal tubercles and bones in general, involvingectoderm and trunk ectomesenchyme. This may re-flect the absence of an established denticle-producing ectodermal-endodermal boundary inWeejasperaspis, which suggests that this boundarymay be under specific genetic controls. Given thepresence of these denticles through the placodermclade, we would suggest that this absence in Weejas-peraspis represents a loss of these controls withinthe group, rather than a plesiomorphic absence forthe Placodermi.
Comparisons to the Actinopterygii
Although the placoderm postbranchial lamina ispart of the trunkshield, we suggest that, develop-mentally, it is also part of the pharyngeal cavity, asare the denticles associated with this lamina. Whilenot common, other jawed vertebrates also possessorganized rows of denticles (as opposed to tubercles)associated with the pectoral girdle (cleithrum andclavicle) and the postbranchial lamina. Among bonyfishes, these denticles very rarely occur in the Sar-copterygii, in the fossil taxon Psarolepis (Zhu et al.,1999). These denticles are otherwise restricted tothe Actinopterygii and include the patches of multi-cusped denticles identified in the fossil teleost Pho-lidophorus at the edge of the postbranchial lamina,on the clavicle, and also on the urohyal, the latterassociated with the visceral skeleton (Patterson,1977). As well, in an unassigned palaeoniscid taxon(basal actinopterygian) illustrated by Poplin (1974:pl. 38), several elongate rows of denticles are posi-tioned dorsally, near the boundary between the post-branchial lamina and the external face of the cle-ithrum and onto the lamina itself. These denticlerows are markedly different from the ornament ofthe external surface, which is composed of fine,smooth ridges. Other actinopterygians with a denti-cle row in a comparable position on the border be-tween cleithrum and postbranchial lamina includethe fossil neopterygians Propterus (Bartram, 1977)and Semionotus (Olsen and McCune, 1991). Dermalornament appears to be otherwise lacking on theexternal surface of the cleithrum in these taxa. Thedenticle rows were said to be comparable to rowslocated on small bones positioned at the anteroven-tral end of the cleithrum; these small bones areidentified as the “serrated appendages” (Bartram,1977), described in detail for the extant neoptery-gian Amia.
303PLACODERM PHARYNGEAL DENTICLES

The serrated appendages in Amia are located an-terolateral to the cleithrum and consist of thin, elon-gate, bony plates crossed by a large number of bonyridges supporting denticles (Liem and Woods, 1973;Grande and Bemis, 1998), and have also been de-scribed from the fossil amiids Solnhofenamia, Ami-opsis, and the fossil caturiid Amblysemius (Grandeand Bemis, 1998: fig. 335). In Amia, the denticlesthemselves are narrow and pointed and are posi-tioned along triangular projections of the bonyridges. The serrated appendages were homologizedwith the clavicle because of their anterolateral posi-tion, the overlapping of the appendage onto the cle-ithrum, and suggested similarity between the den-ticle rows on the appendage and those located on theexternal cleithral surface of Amia (Liem and Woods,1973). Although this similarity would seem to con-tradict suggested differences between denticles onthe postbranchial lamina and external cleithrum,denticles on the latter are overlain by the operculum(M. Coates, pers. commun., 2002; pers. obs. MCZ153684, 2002), and could be considered part of thepharyngeal cavity.
Additionally, the serrated appendage (clavicle)has been described as a posterior branchial arch(Dumeril, 1870). Interestingly, in a juvenile of theamiid Solnhofenamia, denticles form short isolatedrows or whorls, situated one behind the other andreminiscent of the arrangement of gill arches in theoropharyngeal cavity (Grande and Bemis, 1998: fig.335). Two other rows are located dorsally, and one inparticular is positioned along a distinct ridge (mark-ing the separation of the postbranchial lamina andthe external cleithrum surface [Grande and Bemis,1998: fig. 334A]), in a similar position to denticlerows in the undescribed palaeoniscid (Poplin, 1974),Semionotus (Olsen and McCune, 1991) and Prop-terus (Bartram, 1977). In adults of Solnhofenamia,the rows become associated with bony plates (simi-lar to the “serrated appendages”), indicating an on-togenetic change from isolated rows in the juvenileform. These isolated whorls in the juvenile Solnhofe-namia suggest that the denticle whorls/rows developindependently (later fusing to bony plates in adultSolnhofenamia); it is important to note, however,that in Amia, the bony plate forms first, with thedenticles appearing later (Liem and Woods, 1973).
Despite uncertainties involving the homology ofthe clavicle in Amia and related fossil taxa relativeto other serrated appendages (Semionotus and Prop-terus) and to the more dorsal denticle whorls asso-ciated with the cleithrum (Gardiner et al., 1996;Grande and Bemis, 1998), there are features thatare of interest in these actinopterygians with re-spect to the denticles on the postbranchial lamina ofplacoderms. For example, the denticles in the acti-nopterygians occur on, or in association with, thepostbranchial lamina and differ from the ornamenton the external surface of the cleithrum. This is truefor the undescribed palaeoniscid noted above (Pop-
lin, 1974), but also for juvenile Solenhofenamia,Semionotus, and Propterus, where the cleithrum isunornamented. The multicusped denticles in Pholi-dophorus also appear to differ from the ornament ofthe cleithrum (Patterson, 1977: fig. 6B). As well, themore anteroventral denticles in the juvenile Solen-hofenamia form short, evenly spaced rows thatmimic the spacing of gill arches, although archesthemselves are absent and the denticle rows con-tinue up onto the cleithrum.
We suggest that the denticle rows in these acti-nopterygians (as in the placoderm groups describedabove) illustrate the presence of pharyngeal den-ticles developing independently posterior to the gillarches and in association with the pectoral girdle.Although associated with the girdle, these denticlesare unique morphologically and/or in terms of theirarrangement relative to the external surface of thepectoral girdle.
CONCLUSIONS
We conclude that a pharyngeal denticulation ispresent in the Placodermi. Structural characteris-tics described for pharyngeal denticle whorls andtooth sets in the thelodont Loganellia and jawedvertebrates (Fig. 2) are identified in the denticles onthe placoderm postbranchial lamina. These den-ticles clearly differ from the external tubercular der-mal ornament covering the head and trunkshieldplates and we suggest that they are part of thepharyngeal denticulation, representing part of thevisceral skeleton but located posterior to the gillarches.
The placoderm postbranchial lamina representsthe posterior, internal margin of the gill chamberwhere various states of denticle morphology andorganization can be identified. In the first state,denticle morphology is different from the tubercleson that of external surface but organization intodistinct rows is lacking (Brindabellaspis). In theother two states, denticle morphology differs fromthe external trunk tubercles but the denticles arealso organized into rows on the lamina surface. Inthe second state, these rows are staggered, while inthe third state the rows are linear and often sepa-rated by undenticulated bone. The second and thirddenticle states are found in taxa from most placo-derm groups, while the first is restricted to the Ac-anthothoraci (Fig. 1). The polarity of these states isdifficult to resolve at present because of uncertain-ties with regard to the relationships within the Ac-anthothoraci. Although the position of the Acantho-thoraci at the base of the placoderm cladogram iswell established (Fig. 1), relationships within Acan-thothoraci are not, and both the first and secondstates of denticle organization are representedwithin the group. It is certain that a basic similarityexists between all denticle morphologies (triangularin shape) and their orientation and we suggest this
304 Z. JOHANSON AND M.M. SMITH

is plesiomorphic (i.e., the state described for Brinda-bellaspis). The condition of the spaced rows of den-ticles on the postbranchial lamina is absent from theAcanthothoraci and may represent the derived mor-phology for the Placodermi. However, the distribu-tion of these states needs to be tested within theframework of a cladistic analysis of the group as awhole. Despite these phylogenetic uncertainties, westress that the presence of morphologically uniquedenticles associated with the pharyngeal cavity, rel-ative to tubercles or placoid denticles of the dermalexternal skeleton, is shared among placoderms,other jawed vertebrates, and the agnathan Loganel-lia.
We infer specific developmental patterning pro-cesses involved in the production of denticles on theplacoderm postbranchial lamina because these den-ticles differ morphologically and in terms of theirorganization from tubercles on the external surface.These differences should relate to differences in thecell lineage of the epithelium and mesenchyme in-volved in denticle/tubercle formation. Tubercles onthe external surface of the placoderm head andtrunkshield would form from an inductive ectoder-mal epithelium in the absence of endoderm, whiledenticles on the postbranchial lamina in all otherplacoderms may have formed in the presence of aninductive endoderm as one part of the internal vis-ceral skeleton.
Posterior pharyngeal denticle whorls are also as-sociated with, but independent of, the pectoral girdlein the sarcopterygian Psarolepis, several fossil acti-nopterygians, and in the living taxon Amia. It wouldbe interesting to use cell lineage markers to tracethe embryonic origins of cells contributing to thedenticulation associated with the clavicle in Amia,to test our inferences regarding the identity of theepithelium and ectomesenchyme producing den-ticles in fossil placoderms.
Finally, our observations for the Placodermi arecompatible with recent suggestions that duringevolution, pharyngeal denticles and teeth are in-dependently derived relative to external skin tu-bercles, and that neither pharyngeal denticles,nor teeth, derive from the latter during the evolu-tion of jawed fishes (Smith and Coates, 1998, 2000,2001).
Nothing is yet known of the evolutionary origin ofthe placoderm dentition, and homology with that ofthe upper and lower jaws of other jawed fishes isdebated (e.g., Donoghue et al., 2000; Young et al.,2001, Donoghue and Smith, 2001). We do not knowif oral denticles, such as those on the placodermgnathal plates, evolved independently of pharyngealdenticles, or if they evolved as coopted structuresfrom the ordered state on the postbranchial lamina.This is a question we attempt to address in workthat is in preparation.
ACKNOWLEDGMENTS
We thank Daniel Goujet for many discussions,advice, and access to undescribed material of Ro-mundina sp., and Herve Lelievre for access to unde-scribed material of a Saudi Arabian brachythoracidarthrodire and for access to specimens in the collec-tions at the Museum National d’histoire Naturelle.We also thank Mark Wilson, Chelsea Hermus, andGavin Hanke (University of Alberta, Edmonton) forallowing us to use UALVP 42469 (Romundina sp.) inthis work and for photographing the specimen forus. We also thank Gloria Arratia, John Long, RobertJones, Karsten Hartel, and Sally Young for allowingus to study and photograph specimens in the Mu-seum fur Naturkunde (Berlin), Western AustralianMuseum (Perth), Australian Museum (Sydney), Mu-seum of Comparative Zoology, Harvard, and theNatural History Museum (London). Two anonymousreviewers greatly improved the quality of this arti-cle. Images of Brindabellaspis (Fig. 6D, E) andRhynchodus (Fig. 6A) are used with the permissionof Schweizerbart Science Publishers, Stuttgart(http://www.schweizerbart.de). The image of Lo-ganellia and the radiographs of the shark jaws (Fig.2) are reproduced with the permission of Taylor andFrancis, London.
LITERATURE CITED
Aldridge RJ, Purnell MA. 1996. The conodont controversies.TREE 11:463–468.
Aldridge RJ, Briggs DEG, Clarkson ENK, Smith PM. 1986. Theaffinities of conodonts—new evidence from the Carboniferous ofScotland. Lethaia 19:279–292.
Bartram AWH. 1977. The Macrosemiidae, a Mesozoic family ofholostean fishes. Bull Br Mus (Nat Hist) Geol 29:138–239.
Briggs DEG, Clarkson ENK, Aldridge RJ. 1983. The conodontanimal. Lethaia 16:1–92.
Chai Y, Jiang X, Ito Y, Bringas P, Han J, Rowitch DH, Soriano P,McMahon, AP, Sucov, HM. 2000. Fate of the mammalian cra-nial neural crest during tooth and mandibular morphogenesis.Development 127:1671–1679.
Denison RH. 1958. Early Devonian fishes from Utah. III. Arth-rodira. Fieldiana Geol 11:461–551.
Denison RH. 1978. Placodermi. In: Schultze H-P, editor. Hand-book of paleoichthyology, vol. 2. Stuttgart: Gustav Fischer Ver-lag.
Donoghue PCJ, Smith MP. 2001. The anatomy of Turinia pagei(Powrie), and the phylogenetic status of the Thelodonti. Trans RSoc Edinb Earth Sci 92:15–37.
Donoghue PCJ, Purnell MA, Aldridge RJ. 1998. Conodont anat-omy, chordate phylogeny and vertebrate classification. Lethaia31:211–220.
Donoghue PCJ, Forey PL, Aldridge RJ. 2000. Conodont affinityand chordate phylogeny. Biol Rev 75:191–251.
Dumeril A. 1870. Histoire naturelle des poissons ou ichthyologiegeneral. 11 (Ganoides, Dipnes, Lophobranches). Paris: Roret.p 1–625.
Gardiner BG, Maisey JG, Littlewood DTJ. 1996. Interrelation-ships of basal Neopterygians. In: Stiassny M, Parenti L, John-son GD, editors. Interrelationships of fishes. San Diego: Aca-demic Press. p 117–146.
Goujet D. 1984. Les poissons Placodermes du Spitsberg. Arthro-dires Dolichothoraci de la Formation de Wood Bay (Devonieninferieur). Paris: Editions Centre National Recherche Scienti-fique, cahiers de paleontologie.
305PLACODERM PHARYNGEAL DENTICLES

Goujet D. 2001. Placoderms and basal gnathostome apomorphies.In: Ahlberg PE, editor. Major events in early vertebrate evolu-tion. Syst Assoc Spec Vol Ser 61. London: Taylor ‘Francis’. p209–222.
Goujet DF, Young GC. 1995. Interrelationships of placodermsrevisited. Geobios Mem Spec 19:89–96.
Graham A. 2001. The development and evolution of the pharyn-geal arches. J Anat 199:133–141.
Graham A, Smith A. 2001. Patterning the pharyngeal arches.BioEssays 23:54–61.
Grande L, Bemis WE. 1998. A comprehensive phylogenetic studyof Amiid fishes (Amiidae) based on comparative skeletal anat-omy. An empirical search for interconnected patterns of naturalhistory. Soc Vert Paleontol Mem 4; J Vert Paleontol Suppl18:1–690.
Graveson AC, Smith MM, Hall BK. 1997. Neural crest potentialfor tooth development in a urodele amphibian: fevelopmentaland evolutionary significance. Dev Biol 188:34–42.
Gross W. 1933. Die Wirbeltiere des rheinischen Devons. AbhandPreuss Geol Landes 154:1–83.
Gross W. 1957. Mundzahne und hautzahne der acanthodier undarthrodiren. Pal Abt A 109:40.
Gross W. 1959. Arthrodiren aus dem Obersilur der Prager Mulde.Pal Abt A 113:1–35.
Imai H, Osumi N, Eto K. 1998. Contribution of foregut endodermto tooth initiation of mandibular incisor in rat embryos. EurJ Oral Sci 106:19–23.
Janvier P. 1996. Early vertebrates. Oxford monographs on geol-ogy and geophysics 33. Oxford: Oxford Science Publications.
Janvier P. 2001. Ostracoderms and the shaping of the gnathos-tome characters In: Ahlberg PE, editor. Major events in earlyvertebrate evolution. Syst Assoc Spec Vol Ser 61. London: Tay-lor ‘Francis. p 172–186.
Johnson HG, Elliott DK, Wittke JH. 2000. A new actinolepidarthrodire (Class Placodermi) from the Lower Devonian SevyDolomite, East-Central Nevada. Zool J Linn Soc 129:241–266.
Kontges G, Lumsden A. 1996. Rhombencephalic neural crestsegmentation is preserved throughout craniofacial ontogeny.Development 122:3229–3242.
Lelievre H. 1984. Antineosteus lehmani, n.g., n.sp., nouveauBrachythoraci du Devonien inferieur du Maroc presaharien.Ann Paleont 70:115–158.
Lelievre H, Janvier P, Janjou D, Halawani M. 1995. Nefudinaqalibahensis nov. gen., nov. sp. un rhenanide (Vertebrata, Pla-codermi) du Devonien inferieur de la formation Jauf (Emsien)d’Arabie Saoudite. Geobios 18:109–115.
Liem KF, Woods LP. 1973. A probable homologue of the claviclein the holostean fish Amia calva. J Zool Lond 170:521–531.
Long JA. 1997. Ptyctodontid fishes (Vertebrata, Placodermi) fromthe Late Devonian Gogo Formation, Western Australia, with arevision of the European genus Ctenurella O� rvig 1960. Geodi-versitas 19:515–555.
McGonnell IM. 2001. The evolution of the pectoral girdle. J Anat199:189–194.
McGonnell IM, Graham A. 2002. Trunk neural crest has skeleto-genic potential. Curr Biol 12:767–771.
McGonnell IM, McKay IJ, Graham A. 2001. A population ofcaudally migrating cranial neural crest cells: functional andevolutionary implications. Dev Biol 236:54–363.
Miles RS, Young GC. 1977. Placoderm interrelationships recon-sidered in the light on new ptyctodontids from Gogo, WesternAustralia. In: Andrews SM, Miles RS, Walker AD, editors.Problems in vertebrate evolution. London: Academic Press. p123–198.
Nelson GJ. 1969. Gill arches and the phylogeny of fishes, withnotes on the classification of vertebrates. Bull Am Mus Nat Hist41:475–552.
Olsen PE, McCune AR. 1991. Morphology of the Semionotuselegans species group from the Early Jurassic part of the New-ark Supergroup of eastern North America with comments onthe Family Semionotidae (Neopterygii). J Vert Paleont 11:269–292.
O� rvig T. 1973. Acanthodian dentition and its bearing on therelationships of the group. Pal Abt A 143:119–150.
O� rvig T. 1975. Description, with special reference to the dermalskeleton, of a new radotinid arthrodire from the Gedinnian ofArctic Canada. In: Lehman JP, editor. Problemes actuels dePaleontologie: Evolution des Vertebres. Paris: Colloques Inter-nationaux du Centre National de la Recherche Scientifique). p41–72.
O� rvig T. 1980. Histological studies of ostracoderms, placodermsand fossil elasmobranchs. 4. Ptyctodontid tooth plates and theirbearing on holocephalian ancestry: the condition of Ctenurellaand Ptyctodus. Zool Scrip 14:55–79.
Patterson C. 1977. Cartilage bones, dermal bones and membranebones, or the exoskeleton versus the endoskeleton. In: AndrewsSM, Miles RS, Walker AD, editors. Problems in vertebrateevolution. London: Academic Press. p 77–121.
Piotrowski T, Nusslein-Volhard C. 2000. The endoderm plays animportant role in patterning the segmented pharyngeal regionin zebrafish (Danio rerio). Dev Biol 225:39–356.
Poplin C. 1974. Etude de Quelques paleoniscides Pennsylvaniensdu Kansas. Paris: Editions Centre National Recherche Scienti-fique, Cahiers de Paleontologie.
Reif WE. 1982. Evolution of dermal skeleton and dentition invertebrates: the odontode-regulation theory. Evol Biol 15:287–368.
Samson IJ, Smith MP, Armstrong HA, Smith MM. 1992. Pres-ence of the earliest vertebrate hard tissues in conodonts. Sci-ence 256:1308–1311.
Schaeffer B. 1977. The dermal skeleton in fishes. In: AndrewsSM, Miles RS, Walker AD, editors. Problems in vertebrateevolution. London: Academic Press. p 25–52.
Schilling TF, Kimmel CB. 1994. Segment and cell type lineagerestrictions during pharyngeal arch development in the ze-brafish embryo. Development 120:483–494.
Sellman S. 1946. Some experiments on the determination of thelarval tooth in Ambystoma mexicanum. Odontol Tids 54:1–28.
Smith MM. 2003. Vertebrate dentitions at the origin of jaws:When and how pattern evolved. Evol Dev 5.
Smith MM, Coates MI. 1998. Evolutionary origins of the verte-brate dentition: phylogenetic patterns and developmental evo-lution. Eur J Oral Sci 106:482–500.
Smith MM, Coates MI. 2000. Evolutionary origins of teeth andjaws: Developmental models and phylogenetic patterns. In:Teaford, MF, Smith, MM, Ferguson, MWJ, editors. Develop-ment, function and evolution of teeth. Cambridge, UK: Cam-bridge University Press. p 133–151.
Smith MM, Coates MI. 2001. The evolution of vertebrate denti-tions: phylogenetic pattern and developmental methods. In:Ahlberg PE, editor. Major events in early vertebrate evolution.Systematics Association Special Volume Series 61. London:Taylor ‘Francis. p 223–240.
Smith MM, Hall BK. 1990. Developmental and evolutionary ori-gins of vertebrate skeletogenic and odontogenic tissues. BiolRev 65:277–374.
Smith MM, Hall BK. 1993. A developmental model for the evolu-tion of the vertebrate exoskeleton and teeth. The role of cranialand trunk neural crest. Evol Biol 27:387–448.
Smith MM, Hickman A, Amanze D, Lumsden A, Thorogood P.1994. Trunk neural crest origin of caudal fin mesenchyme in thezebrafish Brachydanio rerio. Proc R Soc Lond B 256:137–145.
Smith MM, Samson IJ, Smith MP. 1996. ‘Teeth’ before armour:the earliest vertebrate mineralized tissues. Mod Geol 20:309–319.
Stensio E. 1944. Contributions to the knowledge of the vertebratefauna of the Silurian and Devonian of Western Podolia. II.Notes on two Arthrodires from the Downtonian of Podolia. ArkZool 35A:1–83.
Stensio E. 1959. On the pectoral fin and shoulder girdle of thearthrodires. K Svens Vetenskap handl 8:1–229.
Stensio E. 1963. Anatomical studies on the arthrodiran head. KSvens Vetenskap handl 9:1–419.
306 Z. JOHANSON AND M.M. SMITH

Stensio E. 1969. Placodermata; Arthrodires. In: Piveteau J, edi-tor. Traite de Paleontologie, vol. 4. Paris: Masson. p 71–692.
Van der Bruggen W, Janvier P. 1993. Denticles in thelodonts.Nature 364:107.
Van der Heyden C, Huysseune A. 2000. Dynamics of tooth for-mation and replacement in the zebrafish (Danio rerio) (Te-leostei, Cyprinidae). Dev Dyn 219:486–496
White E. 1952. Australian arthrodires. Bull Br Mus Nat Hist Geol1:249–304.
White E. 1978. The larger arthrodiran fishes from the area ofBurrinjuck Dam, N.S.W. Trans Zool Soc Lond 34:149–262.
Young GC. 1980. A new early Devonian placoderm from NewSouth Wales, Australia, with a discussion of placoderm phylog-eny. Pal Abt A 167:1–76.
Young GC. 1986. The relationships of placoderm fishes. Zool JLinn Soc Lond 88:1–57.
Young GC, Lelievre H, Goujet D. 2001. Primitive jaw structure inan articulated brachythoracid arthrodire (placoderm fish; EarlyDevonian) from southeastern Australia. J Vert Paleontol 21:670–678.
Zhang M-M. 1980. Preliminary note on a lower Devonian anti-arch from Yunnan, China. Vert PalAsiatica 18:179–190.
Zhu M. 1996. The phylogeny of the Antiarchi (Placodermi, Pi-sces), with the description of early Devonian antiarchs fromQujing, Yunnan, China. Bull Mus Nat Hist Nat Sect C 4e ser18:233–348.
Zhu M, Yu X, Janvier P. 1999. A primitive fossil fish sheds lighton the origin of bony fishes. Nature 397:607–610.
307PLACODERM PHARYNGEAL DENTICLES
Copyright © 2022 FDOKUMEN




















