
Developmental Constraints Conserve Evolutionary Pattern in an Osteichthyan Dentition
8
Developmental Constraints Conserve Evolutionary Pattern in an Osteichthyan Dentition Moya Meredith Smith, 1 Natasha I. Krupina, 2 and Jean Joss 3 QUERY SHEET Q1: Au: Please spellout n.t. Q2: Au: Please update. Q3: Au: Please update. Q4: Au: Add pages please. 0
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Developmental Constraints Conserve Evolutionary Pattern in an Osteichthyan Dentition

Developmental Constraints Conserve Evolutionary Patternin an Osteichthyan Dentition
Moya Meredith Smith,1 Natasha I. Krupina,2 and Jean Joss3
QUERY SHEET
Q1: Au: Please spellout n.t.Q2: Au: Please update.Q3: Au: Please update.Q4: Au: Add pages please.
0

Connective Tissue Research, 43: 1–7, 2002Copyright c© 2002 Taylor and Francis0300-8207/02 $12.00 + .00
Developmental Constraints Conserve Evolutionary Patternin an Osteichthyan Dentition
Moya Meredith Smith,1 Natasha I. Krupina,2 and Jean Joss3
1Craniofacial Development, Dental Institute KCL, Guy’s Tower, London, SE1 9RT, United Kingdom2Paleontological Institute RAS, Moscow 117647, Russia3School of Biological Sciences, Macquarrie University, NSW 2109, Australia
5
The lungfish dentition is different from other osteichthyan fishbecause it has a characteristic and unique pattern of teeth arrangedas toothplates. Growth, addition of teeth, and retention as part ofa statodont dentition are determined by the initiation pattern. Inadult lungfish new teeth are only added laterally to each radial row15in the dentition. This is in marked contrast to marginal rows ofteeth with regular, alternating replacement in most osteichthyans.We analyze development from fossil hatchling forms of the LateDevonian dipnoan Andreyevichthys and compare with those ofNeoceratodus, the Australian lungfish. The specific pattern of de-20velopment, unique within lungfish, is also present in the transitory,marginal, anterior dentition in both, reflecting a strongly conserveddevelopmental pattern. These marginal teeth form but are thenlost in both, so that also this program of development is conservedwithin lungfish for 360 million years, from the earliest known form.25
Keywords Lungfish, Dentition, Development, Evolution, Tooth-plates, Constraints.
INTRODUCTIONConventionally in osteichthyan fish, linear rows of teeth form
the dentition on the marginal bones of both upper and lower30jaws (including tetrapods). All teeth are replaced throughoutlife by development of new teeth and tooth shedding in eachposition along the jaws, frequently from a lingual site. The pat-tern of replacement along the jaws is temporo-spatially relatedto loss of the old tooth by resorption. The regulation of tooth35resorption is currently unknown, however, the pattern of toothloss and replacement in osteichthyan fish has recently been re-viewed by Berkovitz [1]. In lungfish teeth are not replaced andthe arrangement of the sequentially initiated teeth is quite spe-cific, described as a radial pattern of tooth rows, or ridges. In40
Received 19 June 2001; accepted 28 November 2001.Address correspondence to Moya Meredith Smith, Craniofacial
Development, Dental Institute KCL, Guy’s Tower, London, SE1 9RT,UK. E-mail: [email protected]
fact, the most striking feature of adult lungfish, the closest liv-ing relatives of tetrapods, is their specialized dentition, in whichteeth and bones are absent from the jaw margins (those equiva-lent to the dentary, splenial, coronoid, premaxilla, and maxilla)in post-Devonian adult lungfish. Instead they have extensive, 45nonshedding (statodont) toothplates on bones of the palate (ec-topterygoids/pterygoids), and the inner side of the lower jaw(prearticulars),∗ and use these to occlude in crushing and grind-ing food. In the earliest and most primitive form, Diabolepis[2, 3], marginal bones do have separate teeth. Notably, separate 50teeth on the bones of the palate and inner (lingual) side of thelower jaw are arranged into radial rows, in a pattern that is uniquefor all post-Devonian dipnoans. The arrangement of toothplatesof this type evolved within dipnoans, because in the sister group,porolepiforms, there are separate teeth in linear rows on all up- 55per and lower marginal bones and inner bones of the palate andlower jaw [4].
Remarkably, one Devonian and one extant species developteeth along the jaw margins in the hatchling stages, which arefully functional but loose them in transition to the adult [5–7]. 60These teeth are also arranged in a radial pattern, not in linearrows, nor as irregular patches of teeth [8]. We cite this as anexample of a developmental pattern of tooth initiation and lossconserved for 360 million years (Myr). We further suggest thatthe lungfish signature for development of the dentition evolved 65by transcription from the inner palatal and lingual bones to thoseat the anterior margins [3], and occurred at some time in theDevonian period. Because we find the same pattern of develop-ment and loss in the Devonian fossil and the extant form, they
∗The exact homologues of the osteichthyan toothed dermal bonesare not known, as in later dipnoans dermal bones surrounding the jawsand rostral cartilages are absent. It is generally accepted that the toothedbones of the palate in the upper jaw are paired ectopterygoids, and pairedor a single vomer. Those of the lower jaw are on the inner, lingual side,and are paired prearticulars; sometimes there is a single symphysealtooth (as in Neoceratodus).
1
2 M. M. SMITH ET AL.
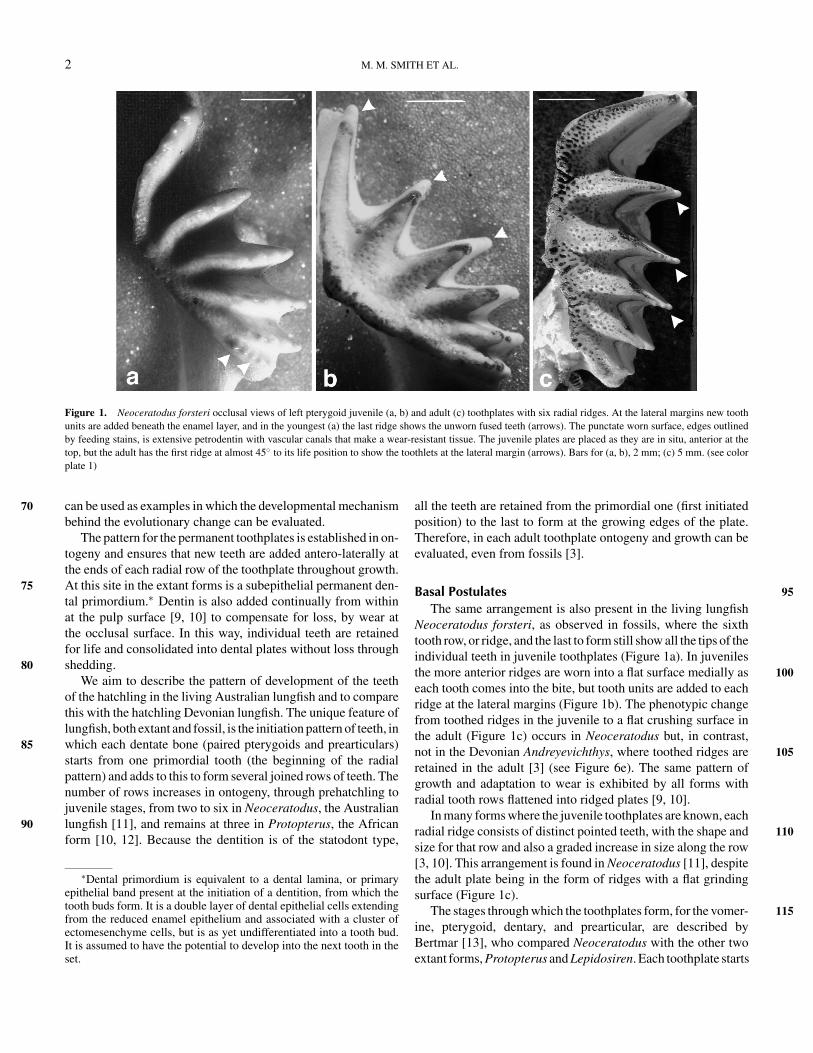
Figure 1. Neoceratodus forsteri occlusal views of left pterygoid juvenile (a, b) and adult (c) toothplates with six radial ridges. At the lateral margins new toothunits are added beneath the enamel layer, and in the youngest (a) the last ridge shows the unworn fused teeth (arrows). The punctate worn surface, edges outlinedby feeding stains, is extensive petrodentin with vascular canals that make a wear-resistant tissue. The juvenile plates are placed as they are in situ, anterior at thetop, but the adult has the first ridge at almost 45◦ to its life position to show the toothlets at the lateral margin (arrows). Bars for (a, b), 2 mm; (c) 5 mm. (see colorplate 1)
can be used as examples in which the developmental mechanism70behind the evolutionary change can be evaluated.
The pattern for the permanent toothplates is established in on-togeny and ensures that new teeth are added antero-laterally atthe ends of each radial row of the toothplate throughout growth.At this site in the extant forms is a subepithelial permanent den-75tal primordium.∗ Dentin is also added continually from withinat the pulp surface [9, 10] to compensate for loss, by wear atthe occlusal surface. In this way, individual teeth are retainedfor life and consolidated into dental plates without loss throughshedding.80
We aim to describe the pattern of development of the teethof the hatchling in the living Australian lungfish and to comparethis with the hatchling Devonian lungfish. The unique feature oflungfish, both extant and fossil, is the initiation pattern of teeth, inwhich each dentate bone (paired pterygoids and prearticulars)85starts from one primordial tooth (the beginning of the radialpattern) and adds to this to form several joined rows of teeth. Thenumber of rows increases in ontogeny, through prehatchling tojuvenile stages, from two to six in Neoceratodus, the Australianlungfish [11], and remains at three in Protopterus, the African90form [10, 12]. Because the dentition is of the statodont type,
∗Dental primordium is equivalent to a dental lamina, or primaryepithelial band present at the initiation of a dentition, from which thetooth buds form. It is a double layer of dental epithelial cells extendingfrom the reduced enamel epithelium and associated with a cluster ofectomesenchyme cells, but is as yet undifferentiated into a tooth bud.It is assumed to have the potential to develop into the next tooth in theset.
all the teeth are retained from the primordial one (first initiatedposition) to the last to form at the growing edges of the plate.Therefore, in each adult toothplate ontogeny and growth can beevaluated, even from fossils [3].
Basal Postulates 95The same arrangement is also present in the living lungfish
Neoceratodus forsteri, as observed in fossils, where the sixthtooth row, or ridge, and the last to form still show all the tips of theindividual teeth in juvenile toothplates (Figure 1a). In juvenilesthe more anterior ridges are worn into a flat surface medially as 100each tooth comes into the bite, but tooth units are added to eachridge at the lateral margins (Figure 1b). The phenotypic changefrom toothed ridges in the juvenile to a flat crushing surface inthe adult (Figure 1c) occurs in Neoceratodus but, in contrast,not in the Devonian Andreyevichthys, where toothed ridges are 105retained in the adult [3] (see Figure 6e). The same pattern ofgrowth and adaptation to wear is exhibited by all forms withradial tooth rows flattened into ridged plates [9, 10].
In many forms where the juvenile toothplates are known, eachradial ridge consists of distinct pointed teeth, with the shape and 110size for that row and also a graded increase in size along the row[3, 10]. This arrangement is found in Neoceratodus [11], despitethe adult plate being in the form of ridges with a flat grindingsurface (Figure 1c).
The stages through which the toothplates form, for the vomer- 115ine, pterygoid, dentary, and prearticular, are described byBertmar [13], who compared Neoceratodus with the other twoextant forms, Protopterus and Lepidosiren. Each toothplate starts
DEVELOPMENTAL CONSTRAINTS—DENTITION 3
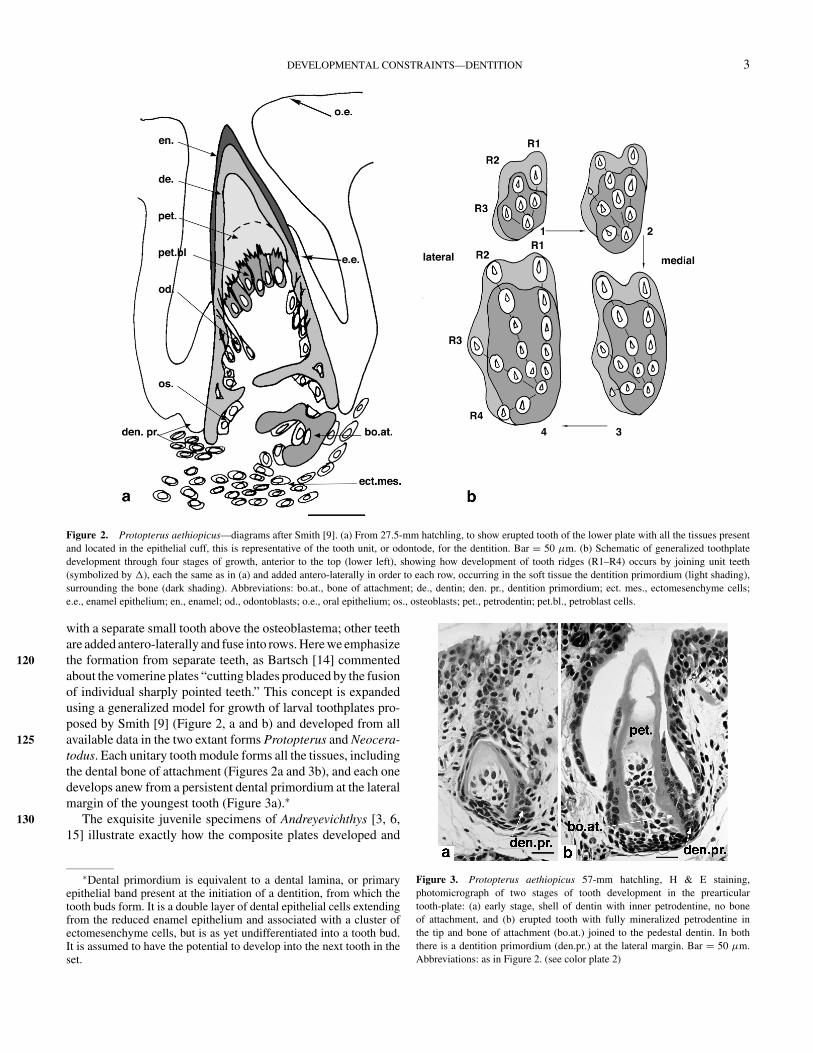
Figure 2. Protopterus aethiopicus—diagrams after Smith [9]. (a) From 27.5-mm hatchling, to show erupted tooth of the lower plate with all the tissues presentand located in the epithelial cuff, this is representative of the tooth unit, or odontode, for the dentition. Bar = 50 µm. (b) Schematic of generalized toothplatedevelopment through four stages of growth, anterior to the top (lower left), showing how development of tooth ridges (R1–R4) occurs by joining unit teeth(symbolized by �), each the same as in (a) and added antero-laterally in order to each row, occurring in the soft tissue the dentition primordium (light shading),surrounding the bone (dark shading). Abbreviations: bo.at., bone of attachment; de., dentin; den. pr., dentition primordium; ect. mes., ectomesenchyme cells;e.e., enamel epithelium; en., enamel; od., odontoblasts; o.e., oral epithelium; os., osteoblasts; pet., petrodentin; pet.bl., petroblast cells.
with a separate small tooth above the osteoblastema; other teethare added antero-laterally and fuse into rows. Here we emphasizethe formation from separate teeth, as Bartsch [14] commented120about the vomerine plates “cutting blades produced by the fusionof individual sharply pointed teeth.” This concept is expandedusing a generalized model for growth of larval toothplates pro-posed by Smith [9] (Figure 2, a and b) and developed from allavailable data in the two extant forms Protopterus and Neocera-125todus. Each unitary tooth module forms all the tissues, includingthe dental bone of attachment (Figures 2a and 3b), and each onedevelops anew from a persistent dental primordium at the lateralmargin of the youngest tooth (Figure 3a).∗
The exquisite juvenile specimens of Andreyevichthys [3, 6,13015] illustrate exactly how the composite plates developed and
∗Dental primordium is equivalent to a dental lamina, or primaryepithelial band present at the initiation of a dentition, from which thetooth buds form. It is a double layer of dental epithelial cells extendingfrom the reduced enamel epithelium and associated with a cluster ofectomesenchyme cells, but is as yet undifferentiated into a tooth bud.It is assumed to have the potential to develop into the next tooth in theset.
Figure 3. Protopterus aethiopicus 57-mm hatchling, H & E staining,photomicrograph of two stages of tooth development in the prearticulartooth-plate: (a) early stage, shell of dentin with inner petrodentine, no boneof attachment, and (b) erupted tooth with fully mineralized petrodentine inthe tip and bone of attachment (bo.at.) joined to the pedestal dentin. In boththere is a dentition primordium (den.pr.) at the lateral margin. Bar = 50 µm.Abbreviations: as in Figure 2. (see color plate 2)

4 M. M. SMITH ET AL.
grew at the Devonian stage of evolution, because there are sev-eral ontogenetic stages available, and many specimens. BothAndreyevichthys and Neoceratodus, although different in theadult, start in the juvenile from radial rows of organised, incre-135mental, unitary teeth. Both forms develop teeth at the jaw mar-gin where the dentary would be, and in Andreyevichthys but notNeoceratodus, teeth also develop on the rostral region (probablypremaxilla), and these are lost in the change to adult in both [3].
OBSERVATIONS140
Neoceratodus—Hatchling DentitionGeneral Development
Detailed descriptions of precise stages in development of thedentition of Neoceratodus forsteri have been provided by Kemp[7, 11, 16, 17]. New observations have been made on whole-145mount cleared and stained preparations (alizarin red and al-cian blue), serial coronal sections stained with hematoxylin andeosin (H and E) and Mallory’s trichrome, and scanning electronmicroscopy (SEM) of the oral surfaces of critical-point-driedstaged embryos.150
Petrodentin DevelopmentThe ontogenetic development of specialized dentin tissue,
petrodentin, has been described in Protopterus aethiopicus [9]and equated with the descriptions by Kemp [7, 16], of centraltissue, or interdenteonal dentin in Neoceratodus forsteri. The155sequence of events is identical, as are the location and timingof its development. Significantly, petrodentin develops secon-darily, immediately inside the shell of dentine of the early teeth(Figures 2a and 3b) on the prearticular, pterygoid, and vomerinetoothplate. Petrodentine develops from a specialized set of pulp160cells (petroblasts) and grows into the expanding dentin cone ofthe tooth before it erupts and has fused to the basal bone ofthe toothplate (Figures 3b and 4d). This occurs in all the per-
Figure 4. Neoceratodus forsteri, photomicrographs of coronal sections through 2 months posthatchling jaws, H & E except (b), which is aniline blue andorange G: (a) dentary tooth shedding (arrow); (b) dentary tooth shedding and adjacent tooth with resorption of bone of attachment (arrow), fibrous connection toMeckel’s cartilage; (c) vomerine, prearticular and dentary teeth with petrodentine in the former but not in the smaller dentary tooth; (d) one tooth of prearticularwith petrodentine forming inside a shell of primary dentine, itself joined to bone of attachment. Abbreviations: d.t., dentary tooth; m.c., Meckel’s cartilage;pr.b., prearticular bone; pr.t., prearticular tooth; sym., symphyseal tissue; v.t., vomerine tooth. (see color plate 3)
manent toothplates to produce a wear-resistant grinding surface(Figure 1c). 165
The rudimentary, but functional teeth of the dentary, andthe single symphyseal tooth, do not develop a central core ofthis tissue (Figure 4, a and b), as seen by comparison in asection through both dentary, prearticular, and vomerine teeth(Figure 4c). It is perhaps because these transitory teeth are 170smaller and have a tiny pulp cavity (see also Kemp [7, 16],petrodentin referred to here as complex dentin). Other data showthat the primordial tooth of each of the prearticular, pterygoid,and vomerine toothplates in the medial position is rudimentary,without a core of petrodentin, and, as is true of the symphyseal 175and dentary teeth, this tooth is also resorbed.
Development and Loss of Marginal Dentition—Symphyseal andDentaryThe teeth in the position of a dentary form a single row on the
outer margin of each side of Meckel’s cartilage, to a maximum 180of 6 teeth between stages 42 and 55. New teeth are seriallyadded in posterolateral positions (Figure 5, b and c) remainingfrom stages 42–47/48. They form the most anterior part of thedentition (Figure 5a), with the single midline tooth (symphyseal)in a symmetrical position medial and posterior to the dentary 185teeth (Figure 5b), and above connective tissue between the twoprearticular bones, which is not cartilage (Figures 4c, 5b, and 5c).The single midline tooth and the paired toothplates (equivalentto the dentary teeth in other osteichthyan fish) are only supportedby cartilage of the lower jaw (Meckel’s, Figures 4c, 5b, and 5c); 190dermal bone does not form (Figure 5). The marginal teeth ofthe lower jaw are all lost after these early stages by a processof resorption of the bone of attachment (Figure 4b), with thetooth being shed through the epithelium (Figure 4, a and b). Itis notable that the process of resorption begins medially in the 195dentary row and spreads tooth by tooth along the row, with themidline (symphyseal) tooth starting the process (Figure 5c). At

DEVELOPMENTAL CONSTRAINTS—DENTITION 5
Figure 5. Neoceratodus forsteri whole mounts stained with alizarin red alone and with alcian blue: (a, b) stage 50, (c) stage 52. (a) Lateral view shows themarginal, anterior tooth plates (arrows) in relation to the large pterygoid and prearticular plates situated below the parasphenoid bone, with the cleithrum at theback of the gill chamber. (b) Five dentary teeth in a joined row (d.t.r.) and one symphyseal tooth (s.t.) supported by Meckel’s cartilage (m.c.); prearticular teeth(pr.t.) are added to the lateral margin. (c) Arrows show the sixth tooth to be added to the dentary plate, while the first tooth on the right side has been resorbed butnot on the left (black arrowheads), and the symphyseal tooth has also been lost (white arrowhead). (see color plate 4)
the time the last tooth in the dentary row is added, the first andmedial tooth in the row has been lost (Figure 5c).
This pattern to the process of formation of the dentarytoothplate is the same as that of the prearticular and pterygoid200toothplates, with unit teeth only added at one end of the rows; atthis stage these already have between three and six teeth. Toothdevelopment has also started on the vomer and these teeth forma single row on each bone; again, new teeth are added postero-laterally. It is highly significant that there is this pattern to the205process of formation and loss in the marginal dentition, as it re-flects what has happened in evolution to achieve the characteris-tic developmental pattern for dipnoans, and the loss of marginalbones. It emphasizes that the dermal bone can be lost indepen-dently of the tooth units, as separate modules in development210and in evolution.
Andreyevichthys Epitomus—Hatchling DentitionGeneral Upper and Lower Jaw
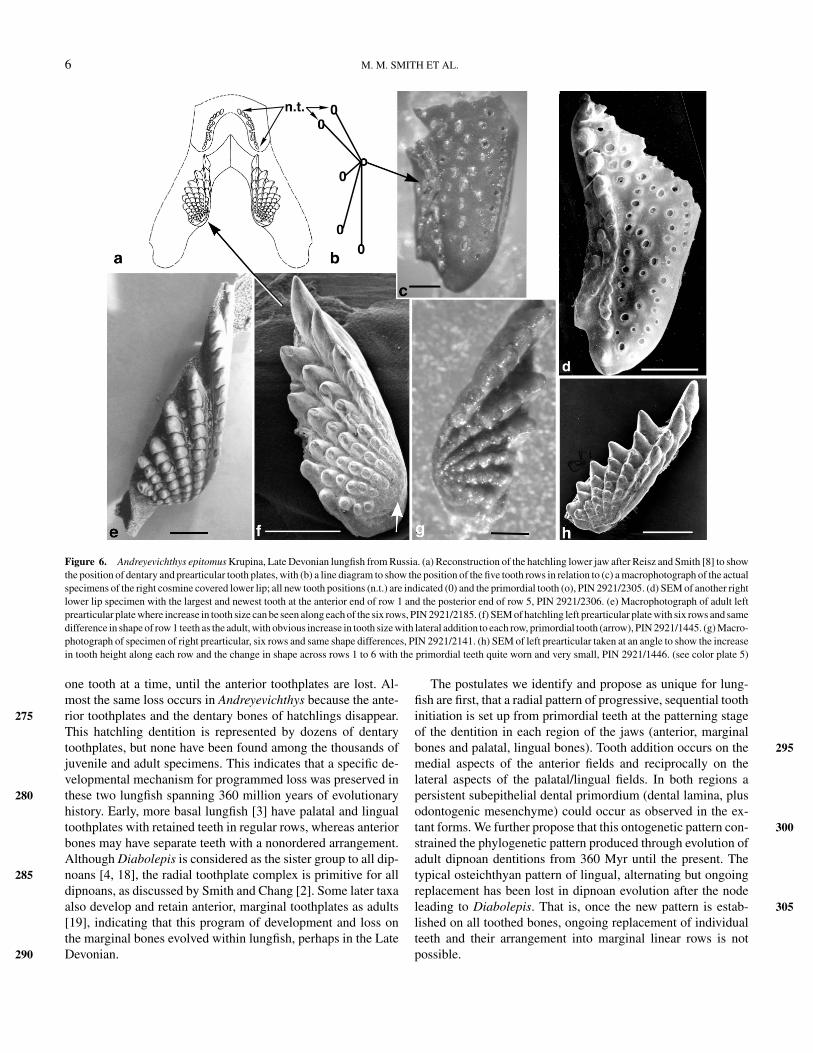
The Late Devonian dipnoan Andreyevichthys epitomus [5, 6]provides examples of fossil hatchling to juvenile toothplates with215remarkable preservation. These are able to reveal exactly how thetoothplates develop and grow in this fossil species. The patternin the prearticular and pterygoid toothplates is of sequentiallyadded teeth in rows diverging from a rudimentary initial tooth,situated in a postero-medial position. Growth points are at the220antero-lateral margin with the largest teeth, and teeth graduallydiminishing in size along each row, but maintenance of shapedifferences between rows one and six is striking (Figure 5, d–h).Also in the upper jaw there are paired rostral (premaxillary) andthree vomerine plates (one median, two lateral). Each is a pair225of single rows of teeth offset at a large angle, appearing to bea linear row, but with the oldest in the center and the newestat opposite ends, whereas the paired dentary toothplates havea complex of several radial rows [8]. Premaxillary, vomerineand pterygoid toothplates of the upper jaw are illustrated by230Smith and Krupina [3]. The core of specialized hypermineralizeddentin, petrodentin, is present in all teeth, even the smallest, andis obvious in etched specimens as well as in sections throughthe teeth.
Lower Jaw 235The most anterior part of the hatchling dentition of Andreye-
vichthys, probably equivalent to the dentary (Figure 6, c and d),provides a good comparison with that of Neoceratodus forsteri.In the fossil example, teeth are arranged along radial rows inwhich teeth diverge from a single primordial tooth. The line dia- 240gram (Figure 6b) shows the primordial tooth (o) from whichall five rows radiate, and the position of the latest teeth tobe added to each row (0). The reconstruction shows completedentary toothplates in situ together with the prearticular plates(Figure 6a), whereas actual specimens of the lower jaw (Figure 6, 245c and d) are incomplete at the anterior end. These anterior tooth-plates are present in the hatchling but not the adult, and the teeth,although much smaller than those on the prearticular toothplates,are arranged in diverging radial rows with increase in size alongthe row (Figure 6, a–d). The position of tooth addition is almost 250the reverse of the prearticular toothplates, being on the medialand anterior, or posterior ends (n.t. Figure 6, a and b). This is Q1assumed to be the only protected area, out of the bite, where thetooth germs could be located. Teeth on the prearticular increasein size along each row from the postero-medial site where the 255smallest primordial tooth is situated (Figure 6, f, g, and h). Ineach toothplate there are six rows as in the adult (Figure 6e),and the increase in height is most apparent in an oblique medialview (Figure 6h).
DISCUSSION AND CONCLUSIONS 260We have observed that in lungfish the mode of dentition pat-
terning is consistent in each tooth field, unique and specialized.Whether it is the marginal, anterior part, or the palatal, lingualdentition, it is specialized relative to all other osteichthyans. Re-markably, there is a pattern of programmed loss in reconstruction 265of the dentition from hatchlings to juveniles, through selectiveresorption of the teeth in the anterior dentition. This does notoccur in the pterygoid and prearticular toothplates. We presumethis must be regulated within the dentition autonomously, but themechanism is quite unknown. In Neoceratodus the sequence of 270loss starts medially with the single symphyseal tooth, and thenthose on the medial end of each dentary toothplate are resorbed,
6 M. M. SMITH ET AL.
Figure 6. Andreyevichthys epitomus Krupina, Late Devonian lungfish from Russia. (a) Reconstruction of the hatchling lower jaw after Reisz and Smith [8] to showthe position of dentary and prearticular tooth plates, with (b) a line diagram to show the position of the five tooth rows in relation to (c) a macrophotograph of the actualspecimens of the right cosmine covered lower lip; all new tooth positions (n.t.) are indicated (0) and the primordial tooth (o), PIN 2921/2305. (d) SEM of another rightlower lip specimen with the largest and newest tooth at the anterior end of row 1 and the posterior end of row 5, PIN 2921/2306. (e) Macrophotograph of adult leftprearticular plate where increase in tooth size can be seen along each of the six rows, PIN 2921/2185. (f) SEM of hatchling left prearticular plate with six rows and samedifference in shape of row 1 teeth as the adult, with obvious increase in tooth size with lateral addition to each row, primordial tooth (arrow), PIN 2921/1445. (g) Macro-photograph of specimen of right prearticular, six rows and same shape differences, PIN 2921/2141. (h) SEM of left prearticular taken at an angle to show the increasein tooth height along each row and the change in shape across rows 1 to 6 with the primordial teeth quite worn and very small, PIN 2921/1446. (see color plate 5)
one tooth at a time, until the anterior toothplates are lost. Al-most the same loss occurs in Andreyevichthys because the ante-rior toothplates and the dentary bones of hatchlings disappear.275This hatchling dentition is represented by dozens of dentarytoothplates, but none have been found among the thousands ofjuvenile and adult specimens. This indicates that a specific de-velopmental mechanism for programmed loss was preserved inthese two lungfish spanning 360 million years of evolutionary280history. Early, more basal lungfish [3] have palatal and lingualtoothplates with retained teeth in regular rows, whereas anteriorbones may have separate teeth with a nonordered arrangement.Although Diabolepis is considered as the sister group to all dip-noans [4, 18], the radial toothplate complex is primitive for all285dipnoans, as discussed by Smith and Chang [2]. Some later taxaalso develop and retain anterior, marginal toothplates as adults[19], indicating that this program of development and loss onthe marginal bones evolved within lungfish, perhaps in the LateDevonian.290
The postulates we identify and propose as unique for lung-fish are first, that a radial pattern of progressive, sequential toothinitiation is set up from primordial teeth at the patterning stageof the dentition in each region of the jaws (anterior, marginalbones and palatal, lingual bones). Tooth addition occurs on the 295medial aspects of the anterior fields and reciprocally on thelateral aspects of the palatal/lingual fields. In both regions apersistent subepithelial dental primordium (dental lamina, plusodontogenic mesenchyme) could occur as observed in the ex-tant forms. We further propose that this ontogenetic pattern con- 300strained the phylogenetic pattern produced through evolution ofadult dipnoan dentitions from 360 Myr until the present. Thetypical osteichthyan pattern of lingual, alternating but ongoingreplacement has been lost in dipnoan evolution after the nodeleading to Diabolepis. That is, once the new pattern is estab- 305lished on all toothed bones, ongoing replacement of individualteeth and their arrangement into marginal linear rows is notpossible.

DEVELOPMENTAL CONSTRAINTS—DENTITION 7
The terminological confusion in descriptions of dipnoan den-titions, where cusps are described as the units on the toothplate310blastema, instead of teeth, was discussed previously [10]. It wasconcluded that there is a real and significant difference betweenteeth and cusps, with the latter being part of a morphogeneticevent in a tooth germ at one stage of development. Teeth areformed at the initiation stage as separate and sequential events,315later fusing into the composite dipnoan dental plates. The im-portance of the concept of a standard gnathostome tooth module(odontode) is emphasized by Smith and Hall [20], in whicheach module also forms its tooth-related bone of attachment.This bone can exist separately from that for the dermal bone of320the jaws, with genetic independence, classically seen in chon-drichthyans, and does so in the anterior teeth of Neoceratodus.The dentary teeth are joined together by the bone of attachmentof each tooth but are not supported by a dentary bone.
The primordial teeth are not competent to produce the spe-325cialized petrodentine tissue, and may be shed. Similarly, theearly tooth shards of the shark dentition [21, 22] establish thepattern for the dentition and are nonfunctional at this stage. Fora recent review of petrodentin as a specialized hypermineralizeddentin infilling primary dentin see Smith and Sansom [23]. In330the hatchling lower jaw in Neoceratodus there is a median singletooth, above the midline symphysis between the two preartic-ular bones; no others develop alongside and there is no der-mal bone beneath. Smith and Coates [22] have proposed that inbasal gnathostomes this tooth is essential to initiate pattern in-335formation, giving subsequent tooth germs left–right and antero-posterior positions. This may be an example in lungfish of similarpatterning controls derived from more basal gnathostomes andretained for the functional hatchling dentition.
The patterning of the dipnoan dentition provides a specific340example of intrinsic developmental constraints. In this situation,the only way in which new teeth can be added is at the ends ofdiverging radial rows and not normally in the interrow position.In each of these positions there is a dental primordium, located inthe lingual or palatal furrow. This suggests that a developmental345constraint operated in the patterning of the dentition over thislong period of evolution for 360 Myr. Both the Devonian formand the living lungfish retain part of the anterior dentition dur-ing early hatchling feeding stages, only to be transformed by itscomplete loss in the adult. There is no other example in oste-350ichthyan fish of this type of dental pattern. It is direct evidence ofan intrinsic developmental constraint operating throughout thephylogeny of dipnoans [8].
ACKNOWLEDGMENTSWe gratefully acknowledge Robert Reisz for continuing sup-355
port and discussion, and Jenny Norman of Macquarrie Univer-sity and the SEM unit at Guy’s Hospital for technical assistance.
REFERENCES[1] Berkovitz, B.K. (2000). “Tooth replacement patterns in non-mammalian
vertebrates.” In: Development, Function and Evolution of Teeth, M.F.360Teaford, M. Meredith Smith, and M.W.J. Ferguson (eds.), pp. 186–200.(Cambridge University Press, Cambridge).
[2] Smith, M.M., and Chang, M.-M. (1990). The dentition of Diabolepis sper-atus Chang and Yu, with further considerations of its relationships and theprimitive dipnoan dentition. J. Vert. Paleont. 10:420–433. 365
[3] Smith, M.M., and Krupina, N.I. (2001). Conserved developmental pro-cesses constrain evolution of lungfish dentitions. J. Anat., in press. Q2
[4] Chang, M.M. (1995). Diabolepis and its bearing on the relationshipsbetween porolepiforms and dipnoans. Bull. Mus. Natl. Hist. Nat. Paris17:235–268. 370
[5] Krupina, N.I. (1987). A new dipnoan from the Upper Devonian of Tularegion. Paleont. J. 3:40–47 (in Russian).
[6] Krupina, N.I. (1992). “Some data on the ontogeny of Devonian dipnoans.”In: Fossil Fishes as Living Animals, E. Mark-Kurik (ed.), pp. 215–222.(Academia 1, Tallin). 375
[7] Kemp, A. (1995). Marginal tooth-bearing bones in the lower jaw ofthe recent Australian lungfish Neoceratodus forsteri Kreft (Osteichthys,Dipnoi). J. Morpholo. 225:345–355.
[8] Reisz, R.R., and Smith, M.M. (2001). Dental development in lungfish: Apattern conserved for 360 million years. Nature 411:548. 380
[9] Smith, M.M. (1985). The pattern of histogenesis and growth of tooth-platesin larval stages of extant lungfish. J. Anat. 140:627–643.
[10] Smith, M.M. (1988). “The dentition of Palaeozoic lungfishes: A consid-eration of the significance of teeth, denticles and tooth-plates for dipnoanphylogeny.” In: Teeth Revisited: Proceedings of the VIIth International 385Symposium on Dental Morphology, Paris 1986. D.E. Russel, J. P. Santoro,and D. Sigogneau-Russel (eds.), Mem. Mus. Natn. Hist. Nat., Paris (serieC), 53:179–194.
[11] Kemp, A. (1977). The pattern of tooth-plateformation in the Australianlungfish, Neoceratodus forsteri Kreft. Zool. J. Linn. Soc. 60:223–258. 390
[12] Smith, M.M. (1984). Petrodentine in extant and fossil dipnoan denti-tions: Microstructure, histogenesis and growth. Proc. Linn. Soc. N.S.W.107:367–407.
[13] Bertmar, G. (1966). The development of skeleton, blood vessels and nervesin the dipoan snout with a discussion on the homology of the dipnoan 395posterior nostrals. Acta Zool. 47:82–150.
[14] Bartsch, P. (1994). Development of the cranium of neoceratodus forsteri,with a discussion of the suspensorium and the opercular apparatus inDipnoi. Zoomorphology 114:1–31.
[15] Krupina, N.I., and Reisz, R.R. (1999). Reconstruction of the larval denti- 400tion of Andreyevichthys epitomus, a Late Devonian dipnoan from Russia.Mod. Geol. 24:99–108.
[16] Kemp, A. (1979). The histology of tooth formation in the Australian lung-fish, Neoceratodus forsteri Kreft. Zool. J. Linn. Soc. 66:251–287.
[17] Kemp, A. (1999). Ontogeny of the skull of the Australian lungfish 405Neoceratodus forsteri (Osteichthys: Dipnoi). J. Zool., Lond. 248:97–137.
[18] Chang, M.-M., and Yu, X.-B. (1984). Structure and phyogenetic signifi-cance of Diabolepis speratus gen. et sp. Nov., a new dipnoan-like formfrom the Lower Devonian of eastern Yunnan, China. Proc. Linn. Soc. 410N.S.W. 107:171–184.
[19] Krupina, N.I., Reisz, R.R., and Scott, D. (2001). The skull and tooth systemof Orlovichthys limnatis, a Late Devonian dipnoan from Russia. Can. J. Q3Earth Sci., in press.
[20] Smith, M.M., and Hall, B.K. (1993). A developmental model for evolution 415of the vertebrate exoskeleton and teeth: The role of cranial and trunk neuralcrest. Evol. Biol. 27:387–447.
[21] Reif, W.-E. (1984). “Pattern regulation in shark dentitions.” In: PatternFormation. A Primer in Developmental Biology, G. M. Malacinski, (ed.),pp. 603–621. (Macmillan, New York). 420
[22] Smith, M.M., and Coates, M.I. (2001). “The evolution of vertebrate denti-tions: Phylogenetic pattern and developmental models.” In: Major Events Q4in Early Vertebrate, P.E. Ahlberg (ed.), pp. (Taylor & Francis, London).
[23] Smith, M.M., and Sansom, I.J. (2000). “Evolutionary origins of dentinein the fossil record of early vertebrates: Diversity, development and func- 425tion.” In: Development, Function and Evolution of Teeth, M.F. Teaford,M. Meredith Smith, and M.W.J. Ferguson (eds.), pp. 133–151. (CambridgeUniversity Press, Cambridge).




















