
Anti-inflammatory properties of interleukin-10 administration in hapten-induced colitis
Upload
independentCategory
view
1download
0
TEo
FSGa*M
BmamachwugauacmdescrppsmcdhBMtcgb
Cca
GASTROENTEROLOGY 2005;129:1991–2008
argeted Deletion of Metalloproteinase 9 Attenuatesxperimental Colitis in Mice: Central Rolef Epithelial-Derived MMP
LORENCIA E. CASTANEDA,* BALJIT WALIA,* MATAM VIJAY–KUMAR,‡ NEAL R. PATEL,*USANNE ROSER,§ VASANTHA L. KOLACHALA,* MAURICIO ROJAS,§ LIXIN WANG,*ABRIELA OPREA,‡ PALLAVI GARG,* ANDREW T. GEWIRTZ,‡ JESSE ROMAN,§ DIDIER MERLIN,*nd SHANTHI V. SITARAMAN‡
Division of Digestive Diseases; ‡Department of Pathology; and §Department of Medicine, Division of Pulmonary, Allergy and Critical Care
edicine, Emory University, Atlanta, GeorgiartisldpphhtapacAiaita
gsatcme
eHMt
ackground & Aims: There is mounting evidence thatatrix metalloproteinases are the predominant protein-
ses expressed in the gut mucosa during active inflam-atory bowel disease. We investigated the role of met-
lloproteinase 9 (MMP-9), a secreted gelatinase that isonsistently up-regulated in both animal models anduman inflammatory bowel disease and is associatedith disease severity, in the pathogenesis of colitis bysing mice containing a targeted deletion of the MMP-9ene. Methods: Dextran sodium sulfate–induced colitisnd Salmonella typhimurium–induced enterocolitis weresed as animal models to study colitis. Results: MMP-9ctivity and protein expression were absent from normalolonic mucosa but were up-regulated during experi-ental colitis. MMP-9�/� mice exposed to dextran so-
ium sulfate or salmonella had a significantly reducedxtent and severity of colitis. Immunohistochemicaltudies showed that MMP-9 was localized to epithelialells and granulocytes during active colitis. The immuneesponse to systemic administration of Salmonella ty-himurium was not affected in MMP-9�/� mice. Neutro-hil transmigration studies and bone marrow chimerashowed that neutrophil MMP-9 is neither required for itsigration nor sufficient to induce tissue damage during
olitis and that epithelial MMP-9 is important for tissueamage. MMP-9 inhibited cell attachment and woundealing in the model intestinal epithelial cell line, Caco2-BE. Conclusions: Taken together, our data suggest thatMP-9 expressed by epithelial cells may play an impor-
ant role in the development of colitis by modulatingell–matrix interaction and wound healing. Thus, strate-ies to inhibit MMP-9 may be of potential therapeuticenefit.
hronic inflammatory bowel diseases (IBD) of theintestine, including Crohn’s disease and ulcerative
olitis, result in considerable morbidity and are associ-
ted with an increased risk of cancer of the colon andectum.1 IBD is defined as chronic, relapsing inflamma-ion of the intestinal tract of unknown origin character-zed by inflammatory cell infiltration, epithelial cell de-truction, connective tissue defects, and ulceration of theining of the colon and intestine resulting in a markedisruption of the mucosal architecture.1 Although lym-hocytes play a central role in the chronic inflammatoryrocess, neutrophil accumulation is considered a keyistological feature of acute flares of the disease.2–4 Theseistological features underlie the clinical symptoms ofhe disease, which include bloody diarrhea, weight loss,nd abdominal pain. The etiopathogenesis of IBD isoorly understood and involves multiple environmentalnd genetic factors involving the immune system, mi-robial factors, and the intestinal–epithelial barrier.2–4
ccumulating data from clinical and experimental stud-es have shown that matrix metalloproteinases (MMPs)re the predominant proteinases in the gut mucosa dur-ng active IBD, and they are thought to play an impor-ant role in the tissue remodeling and ulceration associ-ted with the disease.5–10
MMPs, which include the collagenases, stromelysins,elatinases, and membrane-type MMPs, are a family oftructurally related zinc-dependent proteases that medi-te cellular infiltration, cytokine activation, cell migra-ion, tissue damage, remodeling, and repair.11,12 They areonsidered to be both effectors and regulators of inflam-atory response and are central to the regulation of
xtracellular matrix (ECM) turnover, having the ability
Abbreviations used in this paper: DSS, dextran sodium sulfate; ECM,xtracellular matrix; fMLP, n-formyl methionine leucine phenylalanine;BSS, Hanks’ balanced salt solution; MMP, matrix metalloproteinase;PO, myeloperoxidase; PMN, polymorphonuclear neutrophil; t1/2, half
ime; WT, wild type.© 2005 by the American Gastroenterological Association
0016-5085/05/$30.00
doi:10.1053/j.gastro.2005.09.017
tpupmatcmammMtprrt
ImrbmbfMIaMcutwhcMMdoMoDdu
Am
dpafffdshht6sMuhoaGoTAwsweld(
moBlMMw
dtbtudcftowga
1992 CASTANEDA ET AL GASTROENTEROLOGY Vol. 129, No. 6
o cleave most ECM proteins. Although MMPs likelylay important roles in normal tissue remodeling, dereg-lated expression of MMPs has been implicated in severalathological processes, such as arthritis, atherosclerosis,yocardial infarction, colorectal cancer, tumor invasion,
nd IBD.13–16 MMPs are secreted by a wide variety of cellypes, such as fibroblasts, myofibroblasts, smooth muscleells, epithelial cells, and endothelial cells, as well as byature leukocytes, such as neutrophils, macrophages,
nd lymphocytes.11,12,17 Neutrophils, which account forost of the inflammatory cells in the acute colitic tissue,ainly secrete 2 isoforms: MMP-9 and MMP-8.18
MP-9 is considered the terminal member of the pro-ease cascade that leads to matrix degradation. The ca-acity for gelatinase production is thought to play a keyole in leukocytes trafficking across ECM barriers toeach their target tissues at sites of inflammation throughhe degradation of basement membrane components.
Several studies have characterized MMP profiles inBD. MMPs including MMP-2, MMP-3, MMP-9, andembrane-type MMP-1 have been shown to be up-
egulated in IBD.5,6,19–22 Of the MMPs, MMP-9 haseen shown to be consistently increased in several animalodels of colitis, eg, T cell–mediated colitis, trinitro-
enzene sulfonic acid–induced and dextran sodium sul-ate (DSS)-induced colitis, and human colitis.7,8,23–26
MP-3 and MMP-9 expression correlate with fistulas inBD, and MMP-9 is associated with active inflammationssociated with fistulas.27 It is interesting to note thatMP-2 and MMP-9 activity have been detected in lower
rypt epithelial cells surrounding degraded matrix and innderlying ulcerations.8,10,23 Furthermore, serum andissue MMP-9 levels, but not MMP-2 levels, correlateith disease activity in ulcerative colitis.20,28 MMP in-ibitors ameliorate trinitrobenzene sulfonic acid and DSSolitis.29,30 Despite extensive characterization of theMP profile in IBD, little is known about the role ofMP in the pathogenesis of intestinal inflammation. To
irectly examine the role of MMP-9 in the pathogenesisf colitis, we compared mice genetically deficient inMP-9 with wild-type (WT) littermates for the devel-
pment of colitis in response to oral administration ofSS or Salmonella typhimurium. Furthermore, we ad-ressed the potential mechanism by which MMP-9 mod-lates intestinal inflammation.
Materials and Methods
Experimental Animals
The Animal Care Committee of the Emory University,tlanta, Georgia, approved all procedures performed on ani-
als. In addition, all experiments were conducted in accor- aance with the Guide for the Care and Use of Laboratory Animalsublished by the US Public Health Service. MMP-9�/� micend age- and sex-matched WT littermates were purchasedrom Jackson Laboratories (Bar Harbor, ME) and bred at ouracility. Mice lacking MMP-9 show a normal phenotype exceptor developmental defects in long bone formation caused by aelay in vascularization of the growth plate, which is compen-ated for by the time of birth resulting in normal bone.31 Theomozygous MMP-9–deficient mice used were progeny ofeterozygous breeding pairs of FVB background with disrup-ion of the MMP-9 gene that were backcrossed for more thangenerations. To confirm the absence of MMP-9 gene expres-
ion, genomic DNA was isolated, and the disruption of theMP-9 gene was confirmed via polymerase chain reaction by
sing primers designed to specifically detect heterozygote andomozygote mice.25 For MMP-9, we used a sense oligonucle-tide primer (5=-GCA TAC TTG TAC CGC TAT GG-3=) andn antisense primer (5=-TAA CCG GAG GTC CAA ACTG-3=). For the neomycin cassette, we also used a sense
ligonucleotide primer (5=-GAA GGG ACT GGC TGC TATG-3=) and an antisense primer (5=-AAT ATC ACG GGTGC CAA CG-3=). The lack of MMP-9 protein and activityas confirmed by Western blot and gelatin zymography, re-
pectively. WT and MMP-9�/� littermates used in the studyere between 6 and 8 weeks old at the beginning of the
xperimental protocol, were maintained on a 12-hour dark/ight cycle, and were allowed free access to pelleted nonpurifiediet and tap water under conditions of controlled temperatures25°C � 2°C).
Induction of Dextran Sodium Sulfate Colitis
Colitis was induced in 2 groups of age- and sex-atched male and female WT and MMP-9�/� littermates by
ral administration of DSS (molecular weight, 50000; ICNiomedicals, Aurora, OH) at 3% (wt/vol) in tap water ad
ibitum for 6 days. Age-matched male and female WT andMP-9�/� littermates receiving tap water served as controls.ice were observed daily and evaluated for changes in bodyeight and development of clinical symptoms.
Salmonella typhimurium Infection
Gut-restricted S typhimurium infection was induced asescribed previously.32 To prepare S typhimurium inocula, bac-eria (SL3201) were grown overnight at 37°C in 10 mL of LBroth in a 20-mL container with shaking (150 rpm) and werehen used to inoculate fresh medium (1:100) and were grownnder the same conditions for 2 to 3 hours until an opticalensity at 550 nm of 0.35 to 0.6 was reached. Bacterialultures were then diluted in normal saline, and the colony-orming units were enumerated by plating a dilution series ofhe inoculum. Water and food were withdrawn 4 hours beforeral treatment with 7.5 mg of streptomycin (75 �L of sterileater containing streptomycin or 75 �L of sterile water byavage). Afterward, animals were supplied with food and waterd libitum. At 20 hours after streptomycin treatment, food
nd water were withdrawn again for 4 hours before mice were
i�tohDauf
pSfdlwa12(pbatadmtoaf
wmcH(Csiicwewtssefw
ss
bpamchfmoau1e
frwCfecsrNTTniiiocltm
tmww1sl2chL
December 2005 EPITHELIA–DERIVED MMP–9 IN COLITIS 1993
nfected with 108 colony-forming units of S typhimurium (50L of suspension in phosphate-buffered saline [PBS], orally) or
reated with vehicle (control). Thereafter, food and water wereffered immediately. Mice were killed by carbon dioxide in-alation, and tissue samples were processed as described for theSS colitis model. Systemic salmonellosis was induced by the
dministration of S typhimurium SL3201 (104 colony-formingnits/mouse) intraperitoneally. Mice were weighed daily andollowed up for clinical signs of sepsis, as well as mortality.
Clinical Activity Score
Assessment of body weights, stool consistency, and theresence of occult/gross blood by a guaiac test (Hemoccultensa; Beckman Coulter, Fullerton, CA) were determined dailyor each mouse. Colitis was quantified with a clinical score, asescribed by Cooper et al,33 by using the parameters of weightoss, stool consistency, and fecal blood. Briefly, no weight lossas considered as 0 points, weight loss of 1% to 5% was scored
s 1 point, loss of 5% to 10% was scored as 2 points, loss of0% to 20% was scored as 3 points, and a loss of more than0% was scored as 4. The stool was characterized as normalscore 0), soft with well-formed pellets (score 1), soft withoutellets (score 2), or diarrhea (score 4). For occult blood, nolood was scored 0, positive hemoccult was scored as 2 points,nd gross bleeding was scored as 12. These scores were addedo get a clinical activity score ranging from 0 to 4. Six daysfter the induction of colitis, mice were killed by carbonioxide/hypothermia. The abdominal cavity was exposed by aidline laparotomy, and the entire colon was removed from
he cecum to the anus. The colon was flushed with PBS andpened longitudinally for morphological studies. The lengthnd weight of the colon were measured, and tissue obtainedrom each colon was processed for further assays.
Histological Assessment of Colitis
Colonic specimens obtained as described previouslyere fixed in formalin and coded for blind microscopic assess-ent of mucosal lesions (descending colon for DSS colitis and
ecum for S typhimurium colitis). Sections were stained with&E. Microscopic sections were analyzed by a pathologist
G.O.), and histological scoring was performed as described byooper et al33 on the basis of 3 variables, according to the
everity of the induced damage. Briefly, for inflammation, rarenflammatory cells in the lamina propria were counted as 0,ncreased numbers of granulocytes in the lamina propria as 1,onfluence of inflammatory cells extending into the submucosaere counted as 2, and a score of 3 was given for transmural
xtension of the infiltrate. For crypt damage, an intact cryptas scored as 0, loss of the basal one third counted as 1, loss of
he basal two thirds was counted as 2, entire crypt loss wascored as 3, a change of epithelial surface with erosion wascored as 4, and a score of 5 was given for confluent erosion. Forvaluation of ulcers, an absence of ulcer was scored 0, 1 or 2oci of ulcerations were scored as 1, 3 or 4 foci of ulcerations
ere scored as 2, and confluent or extensive ulceration was gcored as 3. These values were added to give a total histologicalcore of 11.
Myeloperoxidase Activity in the Colon
Neutrophil infiltration into the colon was quantifiedy measuring myeloperoxidase (MPO) activity as describedreviously.34 Five animals were studied in each group. Briefly,portion of colon was homogenized in 1:20 (wt/vol) 50mol/L phosphate buffer (pH 6.0) containing 0.5% hexade-
yltrimethyl ammonium bromide on ice by using a Polytronomogenizer. The homogenate was sonicated for 10 seconds,reeze-thawed 3 times, and centrifuged at 14,000 rpm for 15inutes. Supernatant (14 �L) was added to 1 mg/mL of
-dianisidine hydrochloride and 0.0005% hydrogen peroxide,nd the change in absorbance at 460 nm was measured. Onenit of MPO activity was defined as the amount that degraded�mol of peroxidase per minute at 25°C. The results were
xpressed as absorbance per gram of tissue.
Neutrophil Isolation and TransmigrationStudies
Polymorphonuclear neutrophils (PMNs) were isolatedrom the bone marrow.35 Briefly, the femur and tibia wereemoved and stripped of all muscle and sinew, and the bonesere placed in Hank’s balanced salt solution (HBSS) withouta2� or Mg2� (HBSS�) on ice. The bone marrow was flushed
rom each bone with HBSS�. Cells were pelleted, and maturerythrocytes were removed by hypotonic lysis. PMNs wereonfirmed by Giemsa staining and fluorescence-activated cell-orting analysis by using anti-CD11b. Isolated PMNs wereesuspended in HBSS� and used within 4 hours of isolation.eutrophil transmigration experiments across filters or the84 monolayer were performed as previously described.36
ransmigration was initiated by adding 1–10 �mol/L-formyl methionine leucine phenylalanine (fMLP) in HBSSnto the lower chamber of 5-�m-pore transwell filters (Corn-ng Costar, Cambridge, MA) coated with collagen followed byncubation at 37°C for 2 hours. PMN migration across filtersr epithelial monolayers into the fMLP-containing lowerhambers was quantified by MPO assay as described previous-y.36 A total of 1 � 106 neutrophils per well were used forransmigration studies, and typically 1%–5% of neutrophilsigrated in response to fMLP 10 �mol/L.36
The capacity of PMNs to migrate through Matrigel (Bec-on-Dickinson Labware, Bedford, MA), a prototype basementembrane secreted by the Engelbreth–Holm–Swarm tumor,as assessed in a modified Boyden chamber assay.37 PMNsere isolated from bone marrow and suspended at 1.5 �06/mL in HBSS with Ca2� and Mg2� containing 0.1% bovineerum albumin and placed in the upper compartment. Theower compartment contained fMLP 10 �mol/L in HBSS. The
compartments were separated by an 8-�m-pore membraneoated with Matrigel. After a 3-hour incubation at 37°C inumidified 5% carbon dioxide/air, the filter was stained witheukoStat (Fisher Scientific, Fairlawn, NJ) and mounted on a
lass slide. The number of cells that migrated to the underside
om
amMTaesAw5iG2tdbe
tsh(dAiimalae(mobim(1pnCrwi4tmM
(mpfwe3�sHcdpa
mrc4tacsdthsm
upec0pmatpmuR(wsbC3bndd
1994 CASTANEDA ET AL GASTROENTEROLOGY Vol. 129, No. 6
f the filter in 5 random high-power fields (400�) was deter-ined for each of the filters in experimental condition.
Bone Marrow Transplantation
Bone marrow cells were harvested from both femursnd tibias by flushing the bone cavity with basal bonearrow medium: Iscove’s medium (Cambrex, Walkersville,D) supplemented with 30% fetal bovine serum (Stem Cellechnologies, Vancouver, BC, Canada), 1% bovine serumlbumin (Sigma, St Louis, MO), 100 mmol/L 2-mercapto-thanol (Sigma), 2 mmol/L L-glutamine, 1% penicillin/treptomycin, and 1% fungisone (Irvine Scientific, Santana, CA).38 After washing with PBS, bone marrow cellsere resuspended in basal bone marrow medium. A total of� 106 cells in 50 �L were transplanted retro-orbitally
nto lethally irradiated animals from a cesium source (7.5y): 6 animals per cohort. The mice were given neomycinmg/mL for the first week after transplantation, after which
hey were switched to tap water. All nontransplanted miceied 7 to 14 days after irradiation. Engraftment was verifiedy performing genotyping on the bone marrow cells at thend of 4 weeks after transplantation.
Wound Healing and Cell Adhesion Assay
The electrical cell-substrate impedance sensor sys-em (ECIS; Applied Biophysics, Troy, NY) was used totudy the role of MMP-9 on cell attachment and woundealing.39,40 This system uses small gold film electrodesdiameter, 250 �mol/L) deposited on the bottom of cultureishes and measures the electrode impedance in live cells.s the cells attach and spread on the electrode surface, the
mpedance is altered. This impedance is then recorded as anncrease in resistance by the computer system. Cell-attach-ent and wound-healing measurements were performed on
n 8-well culture dish (ECIS 8W1E plate) coated withaminin 1 (10 �g/mL) kept at 37°C in 5% carbon dioxidend 90% humidity. Each well contained a small activelectrode (area, 5 � 10�4 cm2) and a large counter electrodearea, 0.15 cm2) at the bottom of the well. For cell-attach-ent assay, suspensions of Caco2-BBE cells for inoculation
f ECIS electrodes were prepared from confluent monolayersy first washing the cells with PBS without divalent cat-ons. Monolayers were then incubated with PBS supple-
ented with 1 mmol/L ethylenediaminetetraacetic acidEDTA; Sigma) for 10 minutes at 37°C followed by a0-minute treatment with 0.05% (wt/vol) trypsin in theresence of 1.5 mmol/L EDTA. Trypsinization was termi-ated by adding an excess of complete culture medium.ells were spun down at 100g for 10 minutes and were then
esuspended in serum-free culture medium. The cell densityas monitored by using a standard hemacytometer, and the
noculum was adjusted to give a final cell density between� 105 and 5 � 105/cm2. Before inoculation, ECIS elec-
rodes were equilibrated with a small amount of serum-freeedium under incubator conditions. In experiments using
MP-9, appropriate volumes of the MMP-9 stock solutions din PBS) were added to the electrode-containing wells im-ediately before cells were seeded. In vehicle control ex-
eriments, only PBS was added to the medium. High-requency capacitance measurements (f � 40 kHz; 1 V)ere used to follow the increasing surface coverage of the
lectrode. Capacitance measurements were obtained every0 seconds. The slope of the curve (s � �C/�T) between C
4 nF and C � 2nF, equivalent to the apparent rate ofpreading, was extracted from the data by linear regression.alf times (t1/2), corresponding to the time required for the
ells to spread out on half the available electrode area, wereetermined from these data.41 At the end of the experiment,ictures of the small active electrodes were taken to visu-lize cell repair and migration.
For the wound-healing assays,40 confluent Caco2-BBEonolayers were grown on ECIS 8W1E plates, and baseline
esistance measurements were obtained for 12–15 hours. Theells were then subjected to an increased voltage pulse of0-kHz frequency and 4.5-V amplitude for 30 seconds,42 andhis led to death and detachment of cells present on the smallctive electrode and resulted in a wound normally healed byells surrounding the small active electrode that were notubjected to the increased voltage pulse. MMP-9 was added atifferent concentrations after wounding. Wound healing washen assessed by continuous resistance measurements for 20ours at 500 Hz. At the end of the experiments, pictures of themall active electrodes were taken to visualize cell repair andigration.
Gelatin Zymography
The activity of MMPs was measured by zymographynder nonreducing conditions.43 Briefly, snap-frozen sam-les of colon were homogenized with homogenizer andxtracted in ice-cold extraction buffer (20 �L/mg tissue)ontaining (mmol/L) 50 Tris-HCl, 150 NaCl, 10 CaCl2 and.2 NaN3 and 0.01% Triton X-100 (Sigma; pH 7.6) in theresence of EDTA-free protease cocktail and stirred for 30inutes. After centrifugation, the supernatant was collected
nd stored at �80°C. A total of 10 �g of protein fromissue samples was electrophoretically separated on 7.5%olyacrylamide agarose gel copolymerized with gelatin (1.5g/mL) as a substrate. Electrophoresis was performed by
sing a mini-PROTEAN II electrophoresis system (Bio-ad Laboratories, Hercules, CA) under constant voltage
120 V).The gels were washed 3 times for 15 minutes eachith 2.5% Triton X-100 to remove the sodium dodecyl
ulfate and to allow the electrophoresed enzymes to renatureefore being incubated in zymography buffer (5 mmol/LaCl2 and 50 mmol/L Tris-HCl; pH 7.5) for 18 hours at7°C. The gels were then stained with 0.5% Coomassierilliant blue R-250 (Bio-Rad) and destained with metha-ol, acetic acid, and water (vol/vol 4:1:5). Prestained stan-ard high-range protein markers (Bio-Rad) were used toetermine the molecular weights of the gelatinases. Clear
igested regions representing MMP activity were quantified
we
itluLtf
dP(edp4am(m7taiIfi(bbiMdai
aDtaspcs
tiWtettasrMspD(eMFtwoaM
mW
FgktaswmbWpf
December 2005 EPITHELIA–DERIVED MMP–9 IN COLITIS 1995
ith an imaging densitometer (Camera Imaging Densitom-ter, Model 8300; Alpha Innotech Corp, San Leandro, CA).
Immunohistochemistry
Paraffin sections of colonic tissues were analyzedmmunohistologically by using an MMP-9 monoclonal an-ibody (1:500; Chemicon International, Temecula, CA) fol-owed by a second antibody. Staining was performed bysing the peroxidase-conjugated avidin ABC kit (Vectoraboratories, Burlingame, CA) with peroxidase-labeled an-i-mouse immunoglobulin G. Counterstaining was per-ormed with hematoxylin.
Protein Extraction and Western BlotAnalysis
For Western blot analysis, colon tissue, obtained asescribed previously, was homogenized and extracted withBS containing 1% Triton X-100 and 1% Nonidet P-40
vol/vol), protease inhibitor mixture (Roche Applied Sci-nce, Indianapolis, IN), 1 mmol/L EDTA, 1 mmol/L so-ium orthovanadate, and 1 mmol/L sodium fluoride. Sam-les were then centrifuged at 12,000 rpm for 10 minutes at°C, and the resulting supernatant was used for furtherssays. The total protein concentration of all samples waseasured by the Lowry method with a protein assay reagent
Bio-Rad). Thirty micrograms of protein was boiled for 5inutes in Laemmli’s sample buffer and electrophoresed in
.5%–12% sodium dodecyl sulfate-polyacrylamide gel elec-rophoresis gels. Proteins were transferred to nitrocellulose,nd the membrane was then blocked in 5% nonfat dry milkn Tris-buffered saline with 0.01% Tween-20 for 1 hour.ncubation was performed overnight at 4°C with antibodiesor tissue inhibitor of metalloproteinases 1 (1:1000), tissuenhibitor of metalloproteinases 2 (1:1000), and MMP-91:2000; Chemicon International). Subsequently, the mem-ranes were washed with Tris/NaCl/Tween 20 and incu-ated with a goat anti-rabbit immunoglobulin G horserad-sh peroxidase conjugate (1:4000; Cell Signaling, Beverly,
A) for 1 hour at room temperature. Membranes wereeveloped with Western Lightning Chemiluminescence Re-gent plus (Perkin Elmer, Boston, MA) and quantified bymage analysis.44
Statistical Analysis
The data are presented as mean � SEM. Statisticalnalysis was performed with GraphPad Instat 3 software (Saniego, CA). Groups were compared by using nonparametric
ests, and the significance of differences between groups wasssessed with the Mann–Whitney U-test or the Wilcoxonigned rank test for paired data. Multiple groups were com-ared by a 1-way analysis of variance with the Bonferroniorrection where appropriate. P values �.05 were considered
tatistically significant. aResults
MMP-9 Activity and Expression Are InducedDuring Experimental Colitis in Mice
We used the DSS-induced colitis model to inves-igate the level of gelatinolytic enzymes stimulated dur-ng the induction of colitis. DSS 3% was administered to
T and MMP-9�/� littermates for 6 days, after whichhe mice were killed, and colonic tissue was processed tostimate the gelatinolytic activity. To detect the gela-inolytic activity in protein extracts from water- or DSS-reated mice, we performed zymography by using gelatins a substrate as described in the Materials and Methodsection. A representative zymogram and its densitomet-ic analysis are shown in Figure 1. Lane 0 representsMP-9 recombinant protein (10 ng/mL) used as the
tandard. No MMP-9 activity was observed in extractsrepared from WT mice that had not been exposed toSS (Figure 1A, lanes 1–4) or from any MMP-9�/� mice
Figure 1B, lanes 1–4). However, extracts from DSS-xposed WT animals showed strong induction ofMP-9 activity (7.2 � 1-fold over control; P � .001;
igure 1A, lanes 5–9). MMP-2 was found to be consti-utively expressed in WT and MMP-9�/� mice fed withater, thus indicating its involvement in normal physi-logy. Tissue samples obtained from DSS-treated WTnd MMP-9�/� mice showed significantly increasedMP-2 activity compared with control.To confirm the identity of the enzymes seen in zy-ography, Western blot analysis of tissue extracts fromT and MMP-9�/� mice was performed. As expected,
igure 1. MMP-9 is induced after DSS-induced colitis. Mice wereiven water or 3% DSS (wt/vol) for 6 days, at which time mice wereilled. Colon was harvested and processed as described in the Ma-erials and Methods section. Colonic tissue homogenates from WTnd MMP-9�/� mice were analyzed by gelatin zymography. Each lanehows protein (30 �g per lane) from an individual mouse with orithout DSS treatment. (A and B) Representative zymogram of ho-ogenates from WT and MMP-9�/� mice, respectively. Gelatinolyticands of MMP-9 are indicated by arrows. (C and D) Representativeestern blot of protein from the colon of WT and MMP-9�/� micerobed with anti–MMP-9. Each lane shows protein (30 �g per lane)rom an individual mouse with or without DSS treatment.
nimals that had not been exposed to DSS showed no
cDcapD
pwmtomddm
tdcf2bFbgmps
ietem(
T
CIUT
Naa
b
1996 CASTANEDA ET AL GASTROENTEROLOGY Vol. 129, No. 6
onstitutive MMP-9 protein expression (Figure 1C and, lanes 1–4). MMP-9 protein was induced in WT
olitic samples (Figure 1C, lanes 5–8). Densitometricnalysis showed a 9.4 � 0.9-fold increase in MMP-9rotein expression in extracts from WT mice exposed toSS compared with controls (P � .001).
MMP-9�/� Mice Are Resistant to theDevelopment of Dextran Sodium Sulfate–Induced Colitis
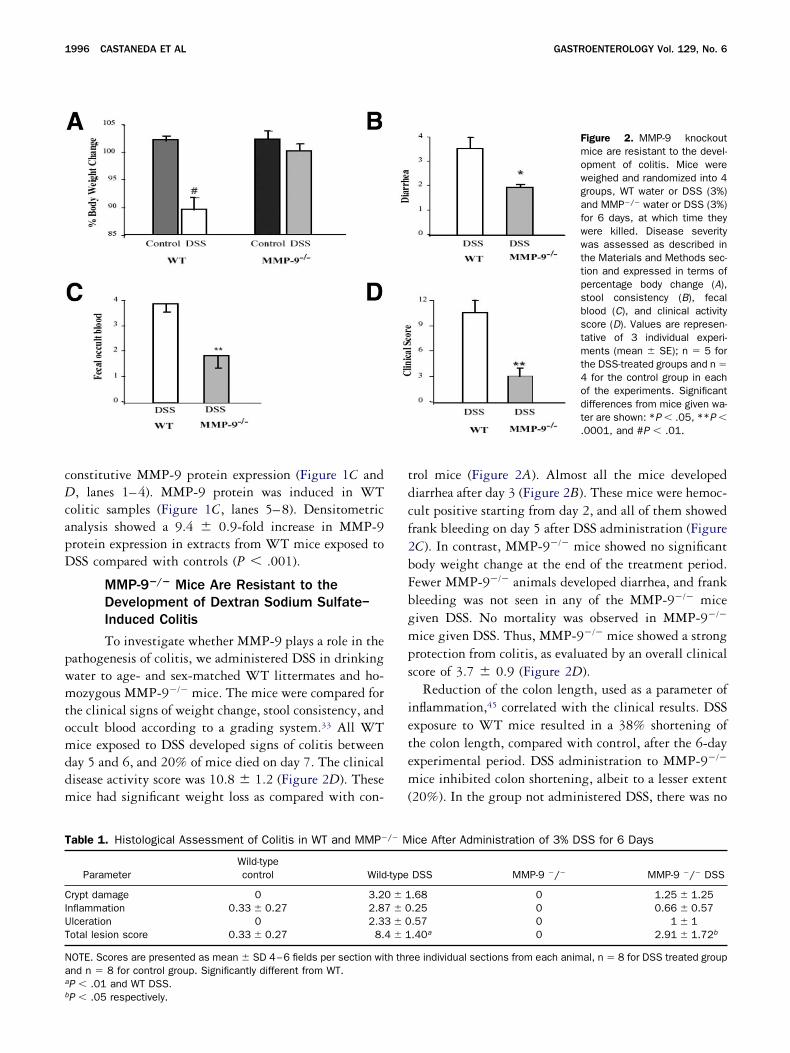
To investigate whether MMP-9 plays a role in theathogenesis of colitis, we administered DSS in drinkingater to age- and sex-matched WT littermates and ho-ozygous MMP-9�/� mice. The mice were compared for
he clinical signs of weight change, stool consistency, andccult blood according to a grading system.33 All WTice exposed to DSS developed signs of colitis between
ay 5 and 6, and 20% of mice died on day 7. The clinicalisease activity score was 10.8 � 1.2 (Figure 2D). Theseice had significant weight loss as compared with con-
able 1. Histological Assessment of Colitis in WT and MMP�
ParameterWild-typecontrol Wild
rypt damage 0 3.2nflammation 0.33 � 0.27 2.8lceration 0 2.3otal lesion score 0.33 � 0.27 8.
OTE. Scores are presented as mean � SD 4–6 fields per section witnd n � 8 for control group. Significantly different from WT.P � .01 and WT DSS.
P � .05 respectively.rol mice (Figure 2A). Almost all the mice developediarrhea after day 3 (Figure 2B). These mice were hemoc-ult positive starting from day 2, and all of them showedrank bleeding on day 5 after DSS administration (FigureC). In contrast, MMP-9�/� mice showed no significantody weight change at the end of the treatment period.ewer MMP-9�/� animals developed diarrhea, and frankleeding was not seen in any of the MMP-9�/� miceiven DSS. No mortality was observed in MMP-9�/�
ice given DSS. Thus, MMP-9�/� mice showed a strongrotection from colitis, as evaluated by an overall clinicalcore of 3.7 � 0.9 (Figure 2D).
Reduction of the colon length, used as a parameter ofnflammation,45 correlated with the clinical results. DSSxposure to WT mice resulted in a 38% shortening ofhe colon length, compared with control, after the 6-dayxperimental period. DSS administration to MMP-9�/�
ice inhibited colon shortening, albeit to a lesser extent20%). In the group not administered DSS, there was no
ice After Administration of 3% DSS for 6 Days
DSS MMP-9 �/� MMP-9 �/� DSS
.68 0 1.25 � 1.25
.25 0 0.66 � 0.57
.57 0 1 � 1
.40a 0 2.91 � 1.72b
ee individual sections from each animal, n � 8 for DSS treated group
Figure 2. MMP-9 knockoutmice are resistant to the devel-opment of colitis. Mice wereweighed and randomized into 4groups, WT water or DSS (3%)and MMP�/� water or DSS (3%)for 6 days, at which time theywere killed. Disease severitywas assessed as described inthe Materials and Methods sec-tion and expressed in terms ofpercentage body change (A),stool consistency (B), fecalblood (C), and clinical activityscore (D). Values are represen-tative of 3 individual experi-ments (mean � SE); n � 5 forthe DSS-treated groups and n �4 for the control group in eachof the experiments. Significantdifferences from mice given wa-ter are shown: *P � .05, **P �.0001, and #P � .01.
/� M
-type
0 � 17 � 03 � 04 � 1
h thr

Fo6MbmsWm
December 2005 EPITHELIA–DERIVED MMP–9 IN COLITIS 1997
igure 3. Targeted deletion of MMP-9 inhibits inflammation associated with DSS. Mice were weighed and randomized into 4 groups, WT waterr DSS (3%) and MMP-9�/� water or DSS (3%), for 6 days, at which time they were killed. (A) Colons were removed from individual mice on day, fixed in formalin, paraffin-embedded, sectioned, and stained with H&E. Representative sections of colon from each group are shown. WT andMP-9�/� mice given water show normal colonic mucosa and intact crypts; WT mice given DSS show severe colonic inflammation characterizedy ulceration (arrows), a dense cell inflammatory infiltrate in the mucosa and submucosa, and disruption of the crypt architecture; and MMP-9�/�
ice given DSS show minimal inflammatory cell infiltration and crypt architectural distortion. (B) One group of WT and MMP-9�/� mice waswitched to plain drinking water after 7 days of DSS. Mice were killed after 7 days of drinking water. Representative histological sections fromT and MMP-9�/� littermates treated with DSS are shown. Note the extensive ulceration, inflammatory infiltrate, and granulation tissue in WT
ice. MMP-9�/� mice showed healed mucosa and residual inflammatory infiltrate in the lamina propria.
s(m
ittsDtpTcMcWadml(cflnycmialn(1w�dotmssems�ta
tat
nWcawmcmhtcaicltlnwriwcMiw
fmmiafss(wo(ie
iac
1998 CASTANEDA ET AL GASTROENTEROLOGY Vol. 129, No. 6
ignificant difference between the colon lengths of WT7.5 � 0.3 cm) and MMP-9�/� (7.75 � 0.3 cm) mice (5ice per group).DSS-induced colitis is characterized by the presence of
nflammation of the colon manifested by crypt destruc-ion, mucosal damage, epithelial erosions, and infiltra-ion of inflammatory cells into the mucosal tissue. Tis-ues collected from WT and MMP-9�/� mice exposed toSS were examined histologically and compared with
hose from normal controls. Histological scoring waserformed in a blinded fashion as described previously.33
he data obtained corroborated the results obtained fromlinical analysis and confirmed the protective role ofMP-9–targeted deletion toward the development of
olitis. As shown in Table 1 and Figure 3, DSS-treatedT mice showed obvious signs of colon inflammation
nd tissue destruction. These mice had extensive cryptamage, epithelial erosion/ulceration, crypt abscess for-ation, and infiltration of inflammatory cells in the
amina propria and muscularis mucosa of colonic sectionsTable 1 and Figure 3A). The inflammatory cells in-luded neutrophils, lymphocytes, and eosinophils, asound at high magnification (Figure 3A). The muscu-aris mucosa of the WT mice showed edema andeutrophil infiltration. In contrast, histological anal-sis of the sections from MMP-9�/� showed signifi-antly reduced histological inflammation, and theseice seemed protected from DSS-associated mucosal
njury, with fewer inflammatory infiltrates and ulcer-tions (Figure 3A; Table 1). Consistently, the histo-ogical injury score obtained from WT mice was sig-ificantly higher than that from the MMP-9�/� miceTable 1 and Figure 3). A histological score of 2.91 �.82 was observed in MMP-9�/� DSS mice, comparedith a score of 8.40 � 1.5 in the WT DSS group (P
.05). Histological signs of inflammation were notetected in the non-DSS control groups. We furtherbserved mice for an additional week after switchinghem to plain water on day 7. Fifty percent of WTice died during this period, and histological analysis
howed progressive ulceration and granulation of tis-ue (Figure 3B). In contrast, MMP-9�/� mice showedvidence of healing (Figure 3B) and residual inflam-atory infiltrate in the lamina propria (histological
core: WT [DSS], 11.1 � 5.1; MMP-9�/� [DSS], 4.92.9; P � .02). These data suggest that WT mice
reated with DSS developed significantly worse injurynd inflammation compared with MMP-9�/� mice.
Accumulation of the inflammatory cell infiltrates intohe injured tissue is a cardinal feature of colitis that mayugment the damage to the inflamed tissue. To confirm
he histological findings, we next compared the extent of heutrophil infiltration into the colonic tissue mucosa ofT and MMP-9�/� mice with or without DSS. The
ontrol animals showed very low levels of neutrophilsnd lymphocytes in the colonic mucosa. However, asith human colitis, WT mice exposed to DSS showed aarked increase in the total number of inflammatory
ells and in the number of granulocytes in their colonicucosa. Targeted deletion of MMP-9 significantly in-
ibited the influx of neutrophils after DSS administra-ion, as shown by total cell count and differential cellounts, including granulocytes (Figure 4A). There wasttenuation of PMN infiltration throughout the mucosa,ncluding the lamina propria vasculature. To furtheronfirm the histological findings with respect to granu-ocyte accumulation, we measured, in the colonic tissue,he activity of MPO, an enzyme specific to granulocyteysosomes and therefore directly correlated with theumber of neutrophils. In contrast to MMP-9�/� mice,hich had MPO activity comparable to that of controls
eceiving water, WT mice fed with DSS had significantlyncreased MPO activity (5.3-fold; P � .001) comparedith controls (Figure 4B). Taken together, these data
learly show in mice that targeted deletion of theMP-9 gene attenuated not only the clinical character-
stics of, but also the histological features associatedith, DSS-induced colitis.
MMP-9 Localizes to Epithelial Cells DuringDextran Sodium Sulfate Colitis
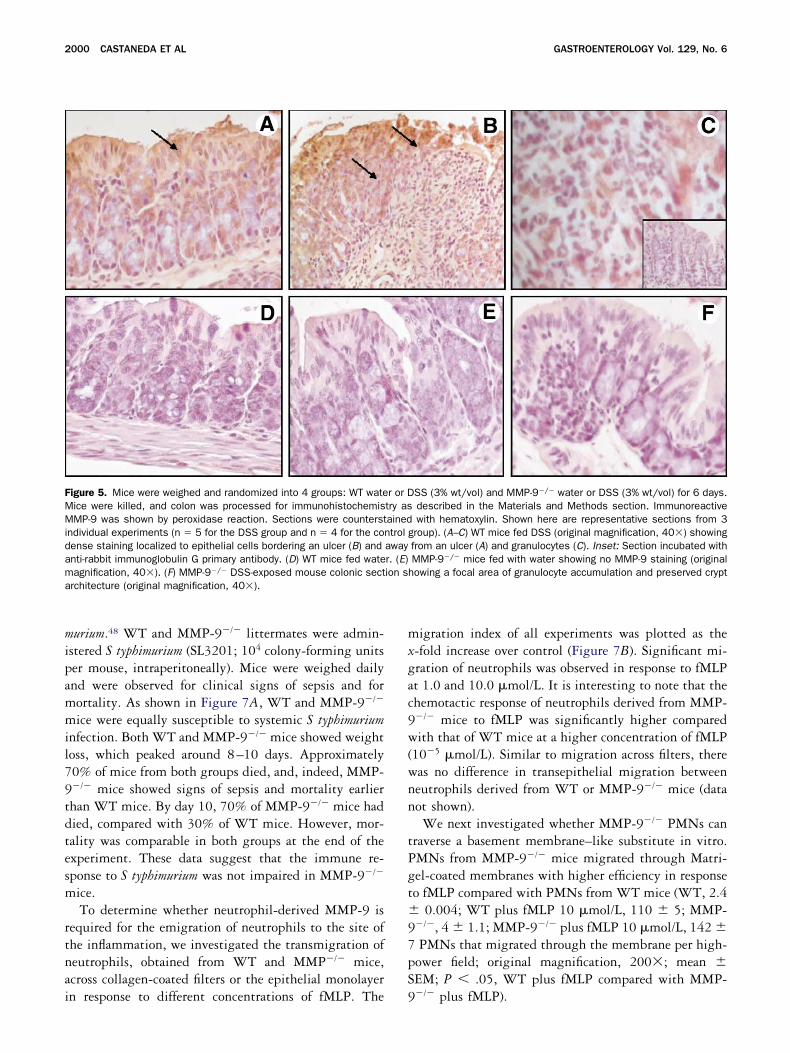
Immunohistochemistry of colon sections per-ormed with affinity-purified antibody directed againstouse MMP-9 confirmed the presence of this enzymeainly in the crypt epithelial cells—both those border-
ng ulcerated areas (Figure 5B) and those in nonulceratedreas (Figure 5A)—and in the granulocytes of WT miceed with DSS. No immunoreactivity was observed inections from MMP�/� mice fed DSS (Figure 5C) or inections from WT or MMP-9�/� animals given waterFigures 5D and E, respectively). Control sections stainedith isotype antibody (anti-rabbit immunoglobulin G)r buffer instead of primary antibody showed no stainingFigure 5C, inset). Thus, the expression of MMP-9 isnduced in mice exposed to DSS and is localized topithelium and granulocytes.
MMP-9�/� Mice Are Protected From OralSalmonella typhimurium–Induced Colitis
As an alternate model of colitis, we used oralnfection with S typhimurium wherein S typhimurium wasdministered after pretreatment of mice with streptomy-in.32 In this model, S typhimurium induces clinical and
istological features of enterocolitis that predominantly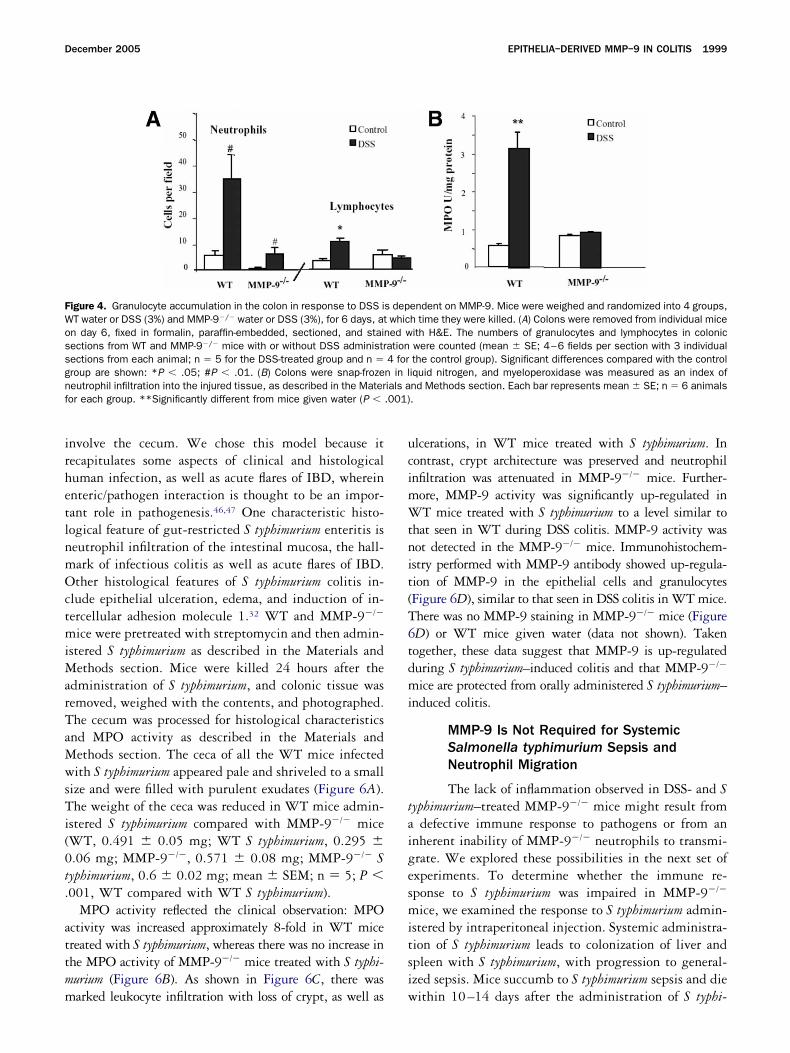
irhetlnmOctmiMarTaMwsTi(0t.
attmm
ucimWtnit(T6tdmi
taigesmitsi
FWossgnf .001)
December 2005 EPITHELIA–DERIVED MMP–9 IN COLITIS 1999
nvolve the cecum. We chose this model because itecapitulates some aspects of clinical and histologicaluman infection, as well as acute flares of IBD, whereinnteric/pathogen interaction is thought to be an impor-ant role in pathogenesis.46,47 One characteristic histo-ogical feature of gut-restricted S typhimurium enteritis iseutrophil infiltration of the intestinal mucosa, the hall-ark of infectious colitis as well as acute flares of IBD.ther histological features of S typhimurium colitis in-
lude epithelial ulceration, edema, and induction of in-ercellular adhesion molecule 1.32 WT and MMP-9�/�
ice were pretreated with streptomycin and then admin-stered S typhimurium as described in the Materials and
ethods section. Mice were killed 24 hours after thedministration of S typhimurium, and colonic tissue wasemoved, weighed with the contents, and photographed.he cecum was processed for histological characteristicsnd MPO activity as described in the Materials andethods section. The ceca of all the WT mice infectedith S typhimurium appeared pale and shriveled to a small
ize and were filled with purulent exudates (Figure 6A).he weight of the ceca was reduced in WT mice admin-
stered S typhimurium compared with MMP-9�/� miceWT, 0.491 � 0.05 mg; WT S typhimurium, 0.295 �.06 mg; MMP-9�/�, 0.571 � 0.08 mg; MMP-9�/� Syphimurium, 0.6 � 0.02 mg; mean � SEM; n � 5; P �001, WT compared with WT S typhimurium).
MPO activity reflected the clinical observation: MPOctivity was increased approximately 8-fold in WT micereated with S typhimurium, whereas there was no increase inhe MPO activity of MMP-9�/� mice treated with S typhi-urium (Figure 6B). As shown in Figure 6C, there was
igure 4. Granulocyte accumulation in the colon in response to DSS iT water or DSS (3%) and MMP-9�/� water or DSS (3%), for 6 days, atn day 6, fixed in formalin, paraffin-embedded, sectioned, and stainections from WT and MMP-9�/� mice with or without DSS administrections from each animal; n � 5 for the DSS-treated group and n �roup are shown: *P � .05; #P � .01. (B) Colons were snap-frozeeutrophil infiltration into the injured tissue, as described in the Materor each group. **Significantly different from mice given water (P �
arked leukocyte infiltration with loss of crypt, as well as w
lcerations, in WT mice treated with S typhimurium. Inontrast, crypt architecture was preserved and neutrophilnfiltration was attenuated in MMP-9�/� mice. Further-
ore, MMP-9 activity was significantly up-regulated inT mice treated with S typhimurium to a level similar to
hat seen in WT during DSS colitis. MMP-9 activity wasot detected in the MMP-9�/� mice. Immunohistochem-stry performed with MMP-9 antibody showed up-regula-ion of MMP-9 in the epithelial cells and granulocytesFigure 6D), similar to that seen in DSS colitis in WT mice.here was no MMP-9 staining in MMP-9�/� mice (FigureD) or WT mice given water (data not shown). Takenogether, these data suggest that MMP-9 is up-regulateduring S typhimurium–induced colitis and that MMP-9�/�
ice are protected from orally administered S typhimurium–nduced colitis.
MMP-9 Is Not Required for SystemicSalmonella typhimurium Sepsis andNeutrophil Migration
The lack of inflammation observed in DSS- and Syphimurium–treated MMP-9�/� mice might result from
defective immune response to pathogens or from annherent inability of MMP-9�/� neutrophils to transmi-rate. We explored these possibilities in the next set ofxperiments. To determine whether the immune re-ponse to S typhimurium was impaired in MMP-9�/�
ice, we examined the response to S typhimurium admin-stered by intraperitoneal injection. Systemic administra-ion of S typhimurium leads to colonization of liver andpleen with S typhimurium, with progression to general-zed sepsis. Mice succumb to S typhimurium sepsis and die
endent on MMP-9. Mice were weighed and randomized into 4 groups,h time they were killed. (A) Colons were removed from individual miceith H&E. The numbers of granulocytes and lymphocytes in colonicwere counted (mean � SE; 4–6 fields per section with 3 individualthe control group). Significant differences compared with the controliquid nitrogen, and myeloperoxidase was measured as an index ofnd Methods section. Each bar represents mean � SE; n � 6 animals.
s depwhiced w
ation4 forn in lials a
ithin 10–14 days after the administration of S typhi-
mipammil79tdtesm
rtnai
mxgac9w(wnn
tPgt�97pS
FMMidama
2000 CASTANEDA ET AL GASTROENTEROLOGY Vol. 129, No. 6
urium.48 WT and MMP-9�/� littermates were admin-stered S typhimurium (SL3201; 104 colony-forming unitser mouse, intraperitoneally). Mice were weighed dailynd were observed for clinical signs of sepsis and forortality. As shown in Figure 7A, WT and MMP-9�/�
ice were equally susceptible to systemic S typhimuriumnfection. Both WT and MMP-9�/� mice showed weightoss, which peaked around 8–10 days. Approximately0% of mice from both groups died, and, indeed, MMP-�/� mice showed signs of sepsis and mortality earlierhan WT mice. By day 10, 70% of MMP-9�/� mice hadied, compared with 30% of WT mice. However, mor-ality was comparable in both groups at the end of thexperiment. These data suggest that the immune re-ponse to S typhimurium was not impaired in MMP-9�/�
ice.To determine whether neutrophil-derived MMP-9 is
equired for the emigration of neutrophils to the site ofhe inflammation, we investigated the transmigration ofeutrophils, obtained from WT and MMP�/� mice,cross collagen-coated filters or the epithelial monolayer
igure 5. Mice were weighed and randomized into 4 groups: WT wateice were killed, and colon was processed for immunohistochemisMP-9 was shown by peroxidase reaction. Sections were counterst
ndividual experiments (n � 5 for the DSS group and n � 4 for the coense staining localized to epithelial cells bordering an ulcer (B) andnti-rabbit immunoglobulin G primary antibody. (D) WT mice fed wateagnification, 40�). (F) MMP-9�/� DSS-exposed mouse colonic sectrchitecture (original magnification, 40�).
n response to different concentrations of fMLP. The 9
igration index of all experiments was plotted as the-fold increase over control (Figure 7B). Significant mi-ration of neutrophils was observed in response to fMLPt 1.0 and 10.0 �mol/L. It is interesting to note that thehemotactic response of neutrophils derived from MMP-�/� mice to fMLP was significantly higher comparedith that of WT mice at a higher concentration of fMLP
10�5 �mol/L). Similar to migration across filters, thereas no difference in transepithelial migration betweeneutrophils derived from WT or MMP-9�/� mice (dataot shown).We next investigated whether MMP-9�/� PMNs can
raverse a basement membrane–like substitute in vitro.MNs from MMP-9�/� mice migrated through Matri-el-coated membranes with higher efficiency in responseo fMLP compared with PMNs from WT mice (WT, 2.4
0.004; WT plus fMLP 10 �mol/L, 110 � 5; MMP-�/�, 4 � 1.1; MMP-9�/� plus fMLP 10 �mol/L, 142 �PMNs that migrated through the membrane per high-
ower field; original magnification, 200�; mean �EM; P � .05, WT plus fMLP compared with MMP-
SS (3% wt/vol) and MMP-9�/� water or DSS (3% wt/vol) for 6 days.described in the Materials and Methods section. Immunoreactivewith hematoxylin. Shown here are representative sections from 3roup). (A–C) WT mice fed DSS (original magnification, 40�) showingfrom an ulcer (A) and granulocytes (C). Inset: Section incubated withMMP-9�/� mice fed with water showing no MMP-9 staining (originalowing a focal area of granulocyte accumulation and preserved crypt
r or Dtry asainedntrol gawayr. (E)ion sh
�/� plus fMLP).

ttbmdibWst
MewTdiW�9b
FoaheSs
December 2005 EPITHELIA–DERIVED MMP–9 IN COLITIS 2001
Finally, we performed bone marrow transplantationo verify the role of MMP-9 in neutrophil transmigra-ion in vivo. All mice recovered uneventfully fromone marrow transplantation. Five weeks after bonearrow transplantation, mice were fed 3% DSS in
rinking water. The mice were compared for the clin-cal signs of weight loss, stool consistency, and occultlood, and mice were killed after 5 days of DSS. All
T mice that received WT bone marrow developedigns of colitis between days 5 and 6. It is interesting
igure 6. Targeted deletion of MMP-9 inhibits Salmonella typhimuriur S typhimurium and MMP-9�/� vehicle or S typhimurium. Mice weres described in the Materials and Methods section. Mice were killed 2arvested, photographed (A), and processed by MPO assay. (B) Thxperiments (n � 5 per group). (C) MPO values are represented as mtyphimurium. (D) MMP-9 immunohistochemistry performed on WT a
taining in the epithelial cells and inflammatory cells.
o note that WT mice that received bone marrow from s
MP-9�/� mice developed colitis that manifestedarlier (between days 4 and 5) and was worse comparedith that in WT mice that received WT bone marrow.hese mice lost 15% to 20% of their body weight andeveloped frank rectal bleeding within 4 days after thenitiation of DSS (clinical score— donor WT/recipient
T, 8.8 � 0.9; donor MMP-9�/�/recipient WT, 110.5; P � .05). Colitis was attenuated in all MMP-
�/� mice whether they received WT or MMP-9�/�
one marrow (Figure 8). MPO assay and histological
uced enterocolitis. Mice were randomized into 4 groups: WT vehicleeated with streptomycin before the administration of S typhimurium,urs after the administration of S typhimurium. Cecum and colon weretograph and histological results are representative of 2 individual
� SE (n � 5); P � .001 compared with MMP-9�/� mice treated withMP-9�/� mice infected with S typhimurium showing intense MMP-9
m–indpretr4 hoe phoeannd M
core reflected the clinical activity in that WT mice

toarrcmmtttCa
fniFMt
rimtpswlrtEataamktttttWatadbEsabc
smiFidt
FmmusoMdgHmqSn�
2002 CASTANEDA ET AL GASTROENTEROLOGY Vol. 129, No. 6
hat received MMP-9�/� or WT bone marrow showedbvious signs of colonic inflammation and tissue dam-ge, compared with MMP-9�/� mice (Figure 8) thateceived WT or MMP-9�/� bone marrow. The WTecipients of MMP-9�/� donor marrow had extensiverypt damage, epithelial ulceration, crypt abscess for-ation, and infiltration of inflammatory cells in theucosa. It is interesting to note that MMP-9�/� mice
hat received WT bone marrow showed increased neu-rophil infiltration compared with MMP-9�/� micehat received MMP-9�/� bone marrow (Figure 8B and). However, tissue damage as measured by clinical
igure 7. Lack of MMP-9 does not affect systemic Salmonella typhi-urium infection or neutrophil transmigration. (A) WT and MMP-9�/�
ice were administered S typhimurium SL3201 (104 colony-formingnits per mouse). Mice were weighed daily and observed for clinicaligns of sepsis, as well as mortality. Data represent the mean � SEf 2 independent experiments (n � 10). (B) Neutrophils from WT andMP-9�/� mice were isolated, and transmigration was performed asescribed in the Materials and Methods section. Neutrophil transmi-ration was initiated by 1 � 10�6 mol/L or 1 � 10�5 mol/L fMLP inBSS�. After 120 minutes at 37°C, migrated neutrophils acrossonolayers into chemoattractant-containing lower chambers wereuantitated by myeloperoxidase assay. Data represent the mean �E of 3 individual experiments, each consisting of duplicate determi-ations. Significant differences from the control group are shown: *P
.05; #P � .01; **P � .001.
ctivity index or total histological score was not dif- c
erent between these 2 groups. These data show thateutrophil MMP is not required for migration ornduction of tissue damage in DSS-induced colitis.urther, these data suggest that mucosal (epithelial)MP-9 expression during colitis is important for
issue damage.
MMP-9 Impairs Epithelial Cell Attachmentand Wound Healing
Taken together, the foregoing data show that theeduced inflammatory response to DSS or S typhimuriumn MMP-9�/� is likely not due to a defective neutrophil
igration or ability to mount an immune response to Syphimurium. They also suggest that epithelial MMP-9lays an important role in mediating the tissue damageeen during colitis. We next addressed the mechanism byhich MMP-9 expressed in the colonic mucosa modu-
ates tissue damage. Because MMPs play an importantole in epithelial cell–matrix interactions, we examinedhe hypothesis that during colitis, MMP-9 alters theCM, thus leading to impaired epithelial cell attachmentnd wound healing. The subepithelial ECM of the intes-inal mucosa is thought to play an important role in cellttachment and mobilization, and MMPs can adverselyffect the composition and integrity of the basementembrane, thus modulating cell attachment.49,50 It is
nown that for epithelial wound healing to occur, at-achment of epithelial cells at the edge of the wound siteo the ECM must be changed to facilitate mobilization ofhese cells. Once resealing of the wound is completed,he ECM provides important signals for the epitheliumo reestablish the barrier and other cellular functions.51,52
e therefore examined the effect of MMP-9 on cellttachment and wound healing by using the model in-estinal epithelial cell line, Caco2-BBE, and ECIS. ECISllows the measurement of cell substrate adhesion bothuring the cell attachment phase and after cells haveecome attached and are in a steady-state condition.lectrical impedance, resistance, and capacitance mea-urements are altered on the basis of the presence ofdherent cells on the electrode, and the plasma mem-ranes of cells act as insulating particles that blockurrent flow.
To study cell attachment, Caco2-BBE cells wereeeded into the ECIS culture plate wells, and capacitanceeasurements were performed for 16 hours as described
n the Materials and Methods section. As shown inigure 9A, the baseline capacitance of cells was approx-mately 5 nF. The capacitance of vehicle-treated cellsecreased over time, thus indicating attachment of cellso matrix (t1/2 vehicle, 2.6 � 0.2 hours). In contrast, the
apacitance measurements in wells treated with MMP-9
(wMmaodtC
rMCbc4c�twApw
FtEmEP4M
Fcshrc
December 2005 EPITHELIA–DERIVED MMP–9 IN COLITIS 2003
green, 100 ng/mL; red, 10 ng/mL; shown in duplicate)ere consistent with delayed cell attachment (t1/2:MP-9 10 ng/mL, 4.5 � 0.5 hours; MMP-9 100 ng/L, 4.5 hours; mean � SEM; n � 4). MMP-9 did not
ffect cell survival as measured by trypan blue exclusionf Caco2-BBE cells treated with MMP-9 (100 ng/mL;ata not shown). Photomicrographs taken at the end ofhe experiment reflected the inhibition of attachment ofaco2-BBE cells to the matrix by MMP-9 (98B).Because cell attachment to matrix plays an important
ole in wound healing, we next examined the effect ofMP-9 on wound healing by using the ECIS. Confluentaco2-BBE cell monolayers grown on an ECIS plate andaseline measurements were obtained for 16 hours. Theells were then subjected to an increased voltage of0-kHz frequency and 4.5-V amplitude for 30 seconds toreate the wound. Vehicle or MMP-9 (100 ng/mL or 1g/mL) was added, and resistance measurements were
aken every few seconds for 18 hours. Baseline resistancesere equal among various groups (31,500 � 4000 ).s shown in Figure 10, the application of the high-fieldulse led to a drastic decrease in the resistance in all the
igure 8. Neutrophil MMP-9 is not required for neutrophil migratransplantation was performed as described in the Materials and Meach bar represents the mean � SE (n � 6). a or c vs b: P � .0yeloperoxidase was measured as an index of neutrophil infiltration iach bar represents the mean � SE (n � 6 animals for each group). a� .05, respectively). (C) Histological assessment of colitis after ad–6 fields per section with 3 individual sections from each animal (nMP-9�/� recipient.
ion, and mucosal MMP-9 mediates tissue damage. (A) Bone marrowthods section (D, donor; R, recipient; WT, wild type; KO, MMP-9�/� mice).1; c vs a: P � .05. (B) Colons were snap-frozen in liquid nitrogen, andnto the injured tissue as described in the Materials and Methods section.vs b or c and b vs c: significantly different from each other (P � .001 and
ministration of 3% DSS for 5 days. Scores are presented as mean � SD:� 6). aP � .01, significantly different compared with MMP-9�/� donor and
ounded wells (4725 � 500 ). The resistances of e
igure 9. MMP-9 impairs cell attachment to matrix. (A) Caco2-BBEells were seeded into ECIS plates. MMP-9 was added at the time ofeeding, and capacitance was measured every few seconds for 16ours (green, 100 ng/mL; red, 10 ng/mL; shown in duplicate). Rep-esentative values from 2 individual experiments performed in dupli-ate are shown (n � 4). (B) Photomicrograph of the wells taken at the
nd of the experiment.
vrsvn8ctc1wi
mmMcdiphs
Sacsp9tS
eamtrafmWiifmbw
2004 CASTANEDA ET AL GASTROENTEROLOGY Vol. 129, No. 6
ehicle-treated cells increased over time and reached theesistance values of nonwounded cells (37,800 � 3800), thus indicating wound closure. In contrast, the re-
istance of MMP-9 treated cells did not reach controlalues (no wounding, 30,837 � 1598; MMP-9 100g/mL, 60% � 5% of baseline value; 1 �g/mL, 40% �% of baseline value; n � 4 per group). These valuesorrelated with the photomicrographs taken at the end ofhe experiment (Figure 10B). As shown in the photomi-rograph, vehicle-treated cells closed the wound within5 hours, and MMP-9–treated cells failed to close theound. Taken together, these data suggest that MMP-9
mpairs cell attachment and wound healing.
Discussion
To explore the direct evidence for the involve-ent and role of MMP-9 in the development of experi-ental colitis, mice with targeted disruption of theMP-9 gene and WT mice were evaluated for the
linical and histopathologic manifestations of colitis in-uced by DSS and S typhimurium. Colitis induced by DSSs a well-established preclinical model for studying theathophysiological aspects of intestinal inflammation,aving features of human colitis.45,53 The results pre-
ented here show that WT mice exposed to DSS or oral mtyphimurium showed significant weight loss, diarrhea,nd gross bleeding and developed inflammatory cell re-ruitment, epithelial ulceration, crypt damage, and de-truction of mucosal tissue. These changes mirror theathology of human colitis. Unlike WT mice, MMP-�/� mice showed significantly reduced clinical and his-ological manifestations of colitis in response to DSS andtyphimurium.Under most circumstances, active inflammatory dis-
ases, such as infectious colitis and acute flares of IBD,re characterized by neutrophil infiltration within theucosa and surface epithelium.54 PMN migration across
he epithelium is considered a key event because itesults in organ dysfunction and disease symptoms, suchs multiple organ failure and shock. In this study, weound robust accumulation of neutrophils in the colonicucosa of DSS and streptomycin/S typhimurium–treatedT mice. In contrast, MMP-9�/� mice had minimal
nflammatory infiltrates in the colonic mucosa and lam-na propria, thus suggesting that MMP-9 is importantor neutrophil recruitment. Because some basementembrane components are substrates for MMP-9, it has
een speculated that secretion of MMP-9 from the PMNsould facilitate their emigration. Indeed, MMP-9 is a
Figure 10. MMP-9 impairswound healing. (A) Caco2-BBEmonolayers plated in ECISplates were subjected to in-creased voltage pulses of 40-kHz frequency, 4.5-V amplitude,and 30-second duration to cre-ate a wound. Vehicle (black,phosphate-buffered saline) orMMP-9 (red, 100 ng/mL; green,1 �g/mL) was added. Thewound was then allowed toheal, and resistance was mea-sured every few seconds for 16hours. Representative normal-ized resistance values to abaseline resistance of 2 inde-pendent experiments areshown (n � 4). (B) Photomicro-graph of the wells taken at theend of the experiment.
ajor component of PMN granules, forming approxi-

mscitmficitcwm9phmMasMtMa
maMscrMkn
alafapnlMeMtLnipr
csEpvhttaIaiebd
iaMwaeMsMlaeodmcwhaphwrpimiOhvmmbii
December 2005 EPITHELIA–DERIVED MMP–9 IN COLITIS 2005
ately 30% of protein content.55 This hypothesis isupported by studies showing that human neutrophilhemotaxis through an artificial basement membrane ismpeded by blocking MMP-9 activity.56 However, inhis study, we found that MMP-9–deficient neutrophilsigrated in increased numbers across collagen-coated
lters and Matrigel (and colonic epithelial monolayer)ompared with WT neutrophils in response to fMLP. Its interesting that bone marrow chimera studies showedhat the neutrophil accumulation and clinical signs ofolitis in WT mice receiving MMP-9�/� bone marrowere significantly higher compared with those in WTice that received WT marrow or in any of the MMP-
�/� recipients. Although this finding is somewhat sur-rising, it is consistent with several previous reports thatave shown that MMP-9 is dispensable for neutrophiligration.37,57–60 Indeed, neutrophils deficient inMP-9 seem to be compensated for by unknown mech-
nisms in that they migrate in greater numbers in re-ponse to appropriate stimuli.59 Furthermore, neutrophilMP-9 has been shown to be intact as a proform until
he neutrophils have reached their target, where theMP-9 released plays an important role in tissue dam-
ge.59
Even though MMP-9 is not required for neutrophiligration, MMP-9–deficient mice show less neutrophil
ccumulation. One possible explanation could be thatMP-9 expressed by colonic epithelial cells after expo-
ure to DSS or S typhimurium generates a neutrophilhemotactic gradient that is responsible for neutrophilecruitment in response to DSS/S typhimurium. Indeed,MPs have been shown to induce cleavage of chemo-
ines, including interleukin 8, thus enhancing theireutrophilic chemotactic activity severalfold.61,62
Our data show that epithelial cells and granulocytesre the main source of MMP-9 in colitis tissue. In IBD,ocal concentrations of proinflammatory cytokines suchs interleukin 1�, interleukin 6, and tumor necrosisactor � are markedly increased, and there is a vastmount of data to show that cytokines can increase MMProduction.12 Also, colitis is characterized by an intenseeutrophil-dominated mucosal inflammation, and it isikely that neutrophils are an additional source of
MP-9 in the colonic compartment in these mice. How-ver, we did not observe the neutrophil lipocalin/pro–MP-9 complex (130 kilodaltons) in our zymograms
hat is characteristic of neutrophil-derived MMP-9.35
ipocalin, which is specifically stored in granules ofeutrophils, is associated with neutrophil gelatinases ands thus a marker for PMN-released MMP-9 in zymogra-hy. This suggests that other resident cells may be
esponsible for the induction of MMP-9 seen in our colitic samples. MMP-9 is produced by several cell types,uch as macrophages, epithelial cells, and fibroblasts.63,64
pithelial cells, including intestinal epithelial cells, havereviously been shown to secrete MMP-9 in response toarious stimuli, and strongly increased levels of MMP-9ave been found to be expressed in the migrating epi-helium of oral wounds and by repairing bronchial epi-helial cells.63,64 We have previously shown that MMP-9ctivity is detected in the luminal fluid of patients withBD and that interleukin 6 up-regulates MMP-9 activitynd protein expression in an intestinal epithelial cell linen vitro and during inflammation in vivo.65 It is inter-sting to note that MMP-2 and MMP-9 activity haveeen detected in lower crypt epithelial cells surroundingegraded matrix as well as underlying ulcerations.8,10,23
Our data show that MMP-9 expression that is inducedn the epithelial cells during colitis mediates tissue dam-ge. One possible mechanism of tissue damage byMP-9 is through its effect of cell attachment andound healing. The ability of epithelial cells to migrate
nd attach to appropriate matrix after damage is consid-red to be an important component of wound healing.MPs, particularly MMP-3 and MMP-9, have been
hown to modulate wound healing in epithelial cells.MP-3, which is constitutively expressed in the epithe-
ia, has been shown to modulate wound healing byltering the migratory substratum and to function asnzymatic machinery that drives the forward movementf the responding cells, modulating the attachment,islodgement, and, finally, resorption of the provisionalatrix. However, MMP-9 is induced at the front of the
orneal or skin epithelium migrating to resurface aound, and both underexpression and overexpressionave been shown to result in defective cell migration,ttachment, and wound healing.66,67 A prototype exam-le of the role of MMP-9 in cell migration and woundealing is exemplified in bullous pemphigus disease,herein defective wound healing in response to injury
esults in bullous lesions of the skin. Dysregulated ex-ression of MMP-9 has been shown to play a critical rolen the development of these skin lesions.16,68 MMP-9�/�
ice do not develop the classic skin lesions, and MMPnhibitors are used as a treatment in human disease.67
ur data show that exogenous MMP-9 significantly in-ibits epithelial cell adhesion and wound healing initro. The modulation of epithelial adhesion by MMP-9ay be due to alterations in cell– cell adhesion or cell–atrix interaction. MMP-9 has been shown to affect
oth of these. For example, MMP-9 expressed duringnflammation has been shown to play an important rolen the posttranslational regulation of cadherin and oc-
luding adhesive activities, wherein proteolytic cleavage
oam
rmbisaiFheshtt
1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
2006 CASTANEDA ET AL GASTROENTEROLOGY Vol. 129, No. 6
f occludin or E-cadherin ectodomian results tight anddhesion junction disassembly, leading to impaired celligration and wound healing.69
In summary, we have shown that MMP-9 is up-egulated during intestinal inflammation. The inflam-atory response to oral administration of toxin (DSS) or
acteria (S typhimurium) is attenuated in MMP-9�/� micen the face of a seemingly intact systemic immune re-ponse to S typhimurium and neutrophil transmigration toppropriate stimuli. MMP-9 is predominantly expressedn epithelial cells and inflammatory cells during colitis.inally, we show that exogenous MMP-9 impairs woundealing and cell attachment in the model intestinalpithelial cell line, Caco2-BBE. Collectively, these datauggest that increased MMP-9 activity impairs woundealing during intestinal inflammation and that selectiveargeting of MMP-9 holds a promising therapeutic po-ential in IBD.
References1. Loftus EV Jr. Clinical epidemiology of inflammatory bowel dis-
ease: incidence, prevalence, and environmental influences. Gas-troenterology 2004;126:1504–1517.
2. Strober W, Fuss IJ, Blumberg RS. The immunology of mucosalmodels of inflammation. Annu Rev Immunol 2002;20:495–549.
3. Fiocchi C. Inflammatory bowel disease: etiology and pathogene-sis. Gastroenterology 1998;115:182–205.
4. Neurath MF, Finotto S, Glimcher LH. The role of Th1/Th2 polar-ization in mucosal immunity. Nat Med 2002;8:567–573.
5. von Lampe B, Barthel B, Coupland SE, Riecken EO, Rosewicz S.Differential expression of matrix metalloproteinases and theirtissue inhibitors in colon mucosa of patients with inflammatorybowel disease. Gut 2000;47:63–73.
6. Pender SL, Tickle SP, Docherty AJ, Howie D, Wathen NC, Mac-Donald TT. A major role for matrix metalloproteinases in T cellinjury in the gut. J Immunol 1997;158:1582–1590.
7. Salmela MT, MacDonald TT, Black D, Irvine B, Zhuma T, Saari-alho-Kere U, Pender SL. Upregulation of matrix metalloprotein-ases in a model of T cell mediated tissue injury in the gut:analysis by gene array and in situ hybridisation. Gut 2002;51:540–547.
8. Seifert WF, Wobbes T, Hendriks T. Divergent patterns of matrixmetalloproteinase activity during wound healing in ileum andcolon of rats. Gut 1996;39:114–119.
9. Saarialho-Kere UK. Patterns of matrix metalloproteinase andTIMP expression in chronic ulcers. Arch Dermatol Res 1998;290(Suppl):S47–S54.
0. Schuppan D, Hahn EG. MMPs in the gut: inflammation hits thematrix. Gut 2000;47:12–14.
1. Brinckerhoff CE, Matrisian LM. Matrix metalloproteinases: a tailof a frog that became a prince. Nat Rev Mol Cell Biol 2002;3:207–214.
2. Mott JD, Werb Z. Regulation of matrix biology by matrix metallo-proteinases. Curr Opin Cell Biol 2004;16:558–564.
3. Vermaelen KY, Cataldo D, Tournoy K, Maes T, Dhulst A, Louis R,Foidart JM, Noel A, Pauwels R. Matrix metalloproteinase-9-medi-ated dendritic cell recruitment into the airways is a critical step ina mouse model of asthma. J Immunol 2003;171:1016–1022.
4. Itoh T, Matsuda H, Tanioka M, Kuwabara K, Itohara S, Suzuki R.
The role of matrix metalloproteinase-2 and matrix metalloprotein-ase-9 in antibody-induced arthritis. J Immunol 2002;169:2643–2647.
5. Dubois B, Opdenakker G, Carton H. Gelatinase B in multiplesclerosis and experimental autoimmune encephalomyelitis. ActaNeurol Belg 1999;99:53–56.
6. Liu Z, Shipley JM, Vu TH, Zhou X, Diaz LA, Werb Z, Senior RM.Gelatinase B-deficient mice are resistant to experimental bullouspemphigoid. J Exp Med 1998;188:475–482.
7. Sternlicht MD, Werb Z. How matrix metalloproteinases regulatecell behavior. Annu Rev Cell Dev Biol 2001;17:463–516.
8. Parks WC, Wilson CL, Lopez-Boado YS. Matrix metalloprotein-ases as modulators of inflammation and innate immunity. NatRev Immunol 2004;4:617–629.
9. Baugh MD, Evans GS, Hollander AP, Davies DR, Perry MJ, LoboAJ, Taylor CJ. Expression of matrix metalloproteases in inflam-matory bowel disease. Ann N Y Acad Sci 1998;859:249–253.
0. Baugh MD, Perry MJ, Hollander AP, Davies DR, Cross SS, LoboAJ, Taylor CJ, Evans GS. Matrix metalloproteinase levels areelevated in inflammatory bowel disease. Gastroenterology 1999;117:814–822.
1. Kirkegaard T, Pedersen G, Saermark T, Brynskov J. Tumour ne-crosis factor-alpha converting enzyme (TACE) activity in humancolonic epithelial cells. Clin Exp Immunol 2004;135:146–153.
2. Bailey CJ, Hembry RM, Alexander A, Irving MH, Grant ME, Shuttle-worth CA. Distribution of the matrix metalloproteinases strome-lysin, gelatinases A and B, and collagenase in Crohn’s diseaseand normal intestine. J Clin Pathol 1994;47:113–116.
3. Tarlton JF, Whiting CV, Tunmore D, Bregenholt S, Reimann J,Claesson MH, Bland PW. The role of up-regulated serine pro-teases and matrix metalloproteinases in the pathogenesis of amurine model of colitis. Am J Pathol 2000;157:1927–1935.
4. Medina C, Videla S, Radomski A, Radomski MW, Antolin M,Guarner F, Vilaseca J, Salas A, Malagelada JR. Increased activityand expression of matrix metalloproteinase-9 in a rat model ofdistal colitis. Am J Physiol Gastrointest Liver Physiol 2003;284:G116–G122.
5. McKaig BC, McWilliams D, Watson SA, Mahida YR. Expressionand regulation of tissue inhibitor of metalloproteinase-1 andmatrix metalloproteinases by intestinal myofibroblasts in inflam-matory bowel disease. Am J Pathol 2003;162:1355–1360.
6. Gao Q, Meijer MJ, Kubben FJ, Sier CF, Kruidenier L, van Duijn W,van den Berg M, van Hogezand RA, Lamers CB, Verspaget HW.Expression of matrix metalloproteinases-2 and -9 in intestinaltissue of patients with inflammatory bowel diseases. Dig LiverDis 2005;37:584–592.
7. Kirkegaard T, Hansen A, Bruun E, Brynskov J. Expression andlocalisation of matrix metalloproteinases and their natural inhib-itors in fistulae of patients with Crohn’s disease. Gut 2004;53:701–709.
8. Kossakowska AE, Medlicott SA, Edwards DR, Guyn L, StabblerAL, Sutherland LR, Urbanski SJ. Elevated plasma gelatinase A(MMP-2) activity is associated with quiescent Crohn’s disease.Ann N Y Acad Sci 1999;878:578–580.
9. Di Sebastiano P, di Mola FF, Artese L, Rossi C, Mascetta G,Pernthaler H, Innocenti P. Beneficial effects of Batimastat (BB-94), a matrix metalloproteinase inhibitor, in rat experimentalcolitis. Digestion 2001;63:234–239.
0. Naito Y, Takagi T, Kuroda M, Katada K, Ichikawa H, Kokura S,Yoshida N, Okanoue T, Yoshikawa T. An orally active matrixmetalloproteinase inhibitor, ONO-4817, reduces dextran sulfatesodium-induced colitis in mice. Inflamm Res 2004;53:462–468.
1. Vu TH, Shipley JM, Bergers G, Berger JE, Helms JA, Hanahan D,Shapiro SD, Senior RM, Werb Z. MMP-9/gelatinase B is a keyregulator of growth plate angiogenesis and apoptosis of hyper-trophic chondrocytes. Cell 1998;93:411–422.
2. Barthel M, Hapfelmeier S, Quintanilla-Martinez L, Kremer M,
Rohde M, Hogardt M, Pfeffer K, Russmann H, Hardt WD. Pretreat-
3
3
3
3
3
3
3
4
4
4
4
4
4
4
4
4
4
5
5
5
5
5
5
5
5
5
5
6
6
6
6
6
6
6
6
6
December 2005 EPITHELIA–DERIVED MMP–9 IN COLITIS 2007
ment of mice with streptomycin provides a Salmonella entericaserovar Typhimurium colitis model that allows analysis of bothpathogen and host. Infect Immun 2003;71:2839–2858.
3. Cooper HS, Murthy SN, Shah RS, Sedergran DJ. Clinicopathologicstudy of dextran sulfate sodium experimental murine colitis. LabInvest 1993;69:238–249.
4. Gewirtz AT, Collier-Hyams LS, Young AN, Kucharzik T, Guilford WJ,Parkinson JF, Williams IR, Neish AS, Madara JL. Lipoxin a4analogs attenuate induction of intestinal epithelial proinflamma-tory gene expression and reduce the severity of dextran sodiumsulfate-induced colitis. J Immunol 2002;168:5260–5267.
5. Cowland JB, Borregaard N. Isolation of neutrophil precursorsfrom bone marrow for biochemical and transcriptional analysis.J Immunol Methods 1999;232:191–200.
6. Si-Tahar M, Sitaraman S, Shibahara T, Madara JL. Negativeregulation of epithelium-neutrophil interactions via activation ofCD44. Am J Physiol Cell Physiol 2001;280:C423–C432.
7. Betsuyaku T, Shipley JM, Liu Z, Senior RM. Neutrophil emigrationin the lungs, peritoneum, and skin does not require gelatinase B.Am J Respir Cell Mol Biol 1999;20:1303–1309.
8. Rojas M, Xu J, Woods CR, Mora AL, Spears W, Roman J, BrighamKL. Bone marrow-derived mesenchymal stem cells in repair of theinjured lung. Am J Respir Cell Mol Biol 2005;33:145–152.
9. Giaever I, Keese CR. A morphological biosensor for mammaliancells. Nature 1993;366:591–592.
0. Giaever I, Keese CR. Monitoring fibroblast behavior in tissueculture with an applied electric field. Proc Natl Acad Sci U S A1984;81:3761–3764.
1. Wegener J, Keese CR, Giaever I. Recovery of adherent cells afterin situ electroporation monitored electrically. Biotechniques2002;33:348, 350, 352 passim.
2. Charrier L, Yan Y, Driss A, Laboisse CL, Sitaraman SV, Merlin D.ADAM-15 inhibits wound healing in human intestinal epithelialcell monolayers. Am J Physiol Gastrointest Liver Physiol 2004;288:G346–G353.
3. Beurden S-V. Zymographic techniques for the analysis of matrixmetalloproteinases and their inhibitors. Biotechniques 2005;38:73–82.
4. Sitaraman SV, Merlin D, Wang L, Wong M, Gewirtz AT, Si-Tahar M,Madara JL. Neutrophil-epithelial crosstalk at the intestinal lume-nal surface mediated by reciprocal secretion of adenosine andIL-6. J Clin Invest 2001;107:861–869.
5. Okayasu I, Hatakeyama S, Yamada M, Ohkusa T, Inagaki Y,Nakaya R. A novel method in the induction of reliable experimen-tal acute and chronic ulcerative colitis in mice. Gastroenterology1990;98:694–702.
6. Sartor RB. Innate immunity in the pathogenesis and therapy ofIBD. J Gastroenterol 2003;38(Suppl 15):43–47.
7. Sartor RB. Therapeutic manipulation of the enteric microflora ininflammatory bowel diseases: antibiotics, probiotics, and prebi-otics. Gastroenterology 2004;126:1620–1633.
8. Richter-Dahlfors A, Buchan AM, Finlay BB. Murine salmonellosisstudied by confocal microscopy: Salmonella typhimurium residesintracellularly inside macrophages and exerts a cytotoxic effecton phagocytes in vivo. J Exp Med 1997;186:569–580.
9. Bouatrouss Y, Herring-Gillam FE, Gosselin J, Poisson J, BeaulieuJF. Altered expression of laminins in Crohn’s disease small in-testinal mucosa. Am J Pathol 2000;156:45–50.
0. Ito M, Hirata S, Arai S, Takahashi T. T cell adherence andmucosal injury in ulcerative colitis: involvement of integrin-fibronectin interaction in situ. J Gastroenterol 1995;30(Suppl8):70–72.
1. Simon-Assmann P, Kedinger M, De Arcangelis A, Rousseau V,Simo P. Extracellular matrix components in intestinal develop-
ment. Experientia 1995;51:883–900.2. Goke M, Zuk A, Podolsky DK. Regulation and function of extra-cellular matrix intestinal epithelial restitution in vitro. Am JPhysiol 1996;271:G729–G740.
3. Pizarro TT, Arseneau KO, Bamias G, Cominelli F. Mouse modelsfor the study of Crohn’s disease. Trends Mol Med 2003;9:218–222.
4. Parkos CA. Molecular events in neutrophil transepithelial migra-tion. Bioessays 1997;19:865–873.
5. Opdenakker G, Van den Steen PE, Van Damme J. Gelatinase B:a tuner and amplifier of immune functions. Trends Immunol2001;22:571–579.
6. Delclaux C, Delacourt C, D’Ortho MP, Boyer V, Lafuma C, Harf A.Role of gelatinase B and elastase in human polymorphonuclearneutrophil migration across basement membrane. Am J RespirCell Mol Biol 1996;14:288–295.
7. Opdenakker G, Masure S, Grillet B, Van Damme J. Cytokine-mediated regulation of human leukocyte gelatinases and role inarthritis. Lymphokine Cytokine Res 1991;10:317–324.
8. Mackarel AJ, Cottell DC, Russell KJ, FitzGerald MX, O’Connor CM.Migration of neutrophils across human pulmonary endothelialcells is not blocked by matrix metalloproteinase or serine pro-tease inhibitors. Am J Respir Cell Mol Biol 1999;20:1209–1219.
9. Allport JR, Lim YC, Shipley JM, Senior RM, Shapiro SD, MatsuyoshiN, Vestweber D, Luscinskas FW. Neutrophils from MMP-9- or neu-trophil elastase-deficient mice show no defect in transendothelialmigration under flow in vitro. J Leukoc Biol 2002;71:821–828.
0. Robinson SN, Pisarev VM, Chavez JM, Singh RK, Talmadge JE.Use of matrix metalloproteinase (MMP)-9 knockout mice demon-strates that MMP-9 activity is not absolutely required for G-CSF orFlt-3 ligand-induced hematopoietic progenitor cell mobilization orengraftment. Stem Cells 2003;21:417–427.
1. D’Haese A, Wuyts A, Dillen C, Dubois B, Billiau A, Heremans H,Van Damme J, Arnold B, Opdenakker G. In vivo neutrophil recruit-ment by granulocyte chemotactic protein-2 is assisted by gelati-nase B/MMP-9 in the mouse. J Interferon Cytokine Res 2000;20:667–674.
2. Van den Steen PE, Proost P, Wuyts A, Van Damme J, OpdenakkerG. Neutrophil gelatinase B potentiates interleukin-8 tenfold byaminoterminal processing, whereas it degrades CTAP-III, PF-4,and GRO-alpha and leaves RANTES and MCP-2 intact. Blood2000;96:2673–2681.
3. McCarthy GM, Macius AM, Christopherson PA, Ryan LM, Pour-motabbed T. Basic calcium phosphate crystals induce synthesisand secretion of 92 kDa gelatinase (gelatinase B/matrix metal-loprotease 9) in human fibroblasts. Ann Rheum Dis 1998;57:56–60.
4. Gan X, Wong B, Wright SD, Cai TQ. Production of matrix metallo-proteinase-9 in CaCO-2 cells in response to inflammatory stimuli.J Interferon Cytokine Res 2001;21:93–98.
5. Casaneda FE WB, Wang L, Sitaraman SV, Merlin D. Regulation ofexpression of matrix metalloproteinases (MMP-2, MT1- MMP)and their tissue inhibitor (TIMP-2) by IL-6 in intestinal epithelialcells. Gastroenterology 2004;126.
6. Fini ME, Parks WC, Rinehart WB, Girard MT, Matsubara M, CookJR, West-Mays JA, Sadow PM, Burgeson RE, Jeffrey JJ, RaizmanMB, Krueger RR, Zieske JD. Role of matrix metalloproteinases infailure to re-epithelialize after corneal injury. Am J Pathol 1996;149:1287–1302.
7. Mohan R, Chintala SK, Jung JC, Villar WV, McCabe F, Russo LA,Lee Y, McCarthy BE, Wollenberg KR, Jester JV, Wang M, WelgusHG, Shipley JM, Senior RM, Fini ME. Matrix metalloproteinasegelatinase B (MMP-9) coordinates and effects epithelial regener-ation. J Biol Chem 2002;277:2065–2072.
8. Liu Z, Zhou X, Shapiro SD, Shipley JM, Twining SS, Diaz LA,Senior RM, Werb Z. The serpin alpha1-proteinase inhibitor is acritical substrate for gelatinase B/MMP-9 in vivo. Cell 2000;102:
647–655.
6
o
Rs
nEA6C
2008 CASTANEDA ET AL GASTROENTEROLOGY Vol. 129, No. 6
9. Marambaud P, Shioi J, Serban G, Georgakopoulos A, Sarner S, NagyV, Baki L, Wen P, Efthimiopoulos S, Shao Z, Wisniewski T, RobakisNK. A presenilin-1/gamma-secretase cleavage releases the E-cad-herin intracellular domain and regulates disassembly of adherensjunctions. EMBO J 2002;21:1948–1956.
Received March 16, 2005. Accepted August 31, 2005.Address requests for reprints to: Shanthi V. Sitaraman, MD, Division
f Digestive Diseases, Room 201-F, 615, Michael Street, Whitehead
esearch Building, Emory University, Atlanta, Georgia 30322. e-mail:[email protected]; fax: (404) 727-5767.F.E.C., B.W., and M.V.-K. contributed equally to this article.Supported by National Institute of Diabetes and Digestive and Kid-
ey Diseases Grants DK 02802, DK 064644, and DK 06411; anlsevier Research Initiative Award; a Crohn’s and Colitis Foundation ofmerica Senior Research Award (S.V.S); Grants DK 02831 (D.M), DK1417 (A.T.G), and HL 68543 (J.R); and Digestive Disease Researchenter Grant 5R24DK064399-02.
We thank Dr Susan Voss for assistance with cell culture.Copyright © 2022 FDOKUMEN




















