
Macro-regions & Regions further development planning and statistical regions of Hungary

BIODIVERSITYRESEARCH
Systematic, large-scale nationalbiodiversity surveys: NeoMaps as amodel for tropical regionsJose R. Ferrer-Paris1,†, Jon P. Rodrıguez1*, Tatjana C. Good1,
Ada Y. Sanchez-Mercado1,†, Kathryn M. Rodrıguez-Clark1,
Gustavo A. Rodrıguez1 and Angel Solıs2
1Centro de Ecologıa, Instituto Venezolano de
Investigaciones Cientıficas (IVIC), Apartado
20632, Caracas, 1020-A, Venezuela,2Instituto Nacional de Biodiversidad
(INBio), Apartado 22-3100, Santo Domingo
de Heredia, Costa Rica
*Correspondence: Jon P. Rodrıguez, Centro
de Ecologıa, Instituto Venezolano de
Investigaciones Cientıficas (IVIC), Apartado
20632, Caracas 1020-A, Venezuela.
E-mail: [email protected]
†Present address: Centro de Estudios
Botanicos y Agroforestales, Instituto
Venezolano de Investigaciones Cientıficas
(IVIC), Sede IVIC-Zulia, Apartado 20632,
Caracas, 1020-A, Venezuela
ABSTRACT
Aim To test a method for rapidly and reliably collecting species distribution
and abundance data over large tropical areas [known as Neotropical Biodiversity
Mapping Initiative (NeoMaps)], explicitly seeking to improve cost- and time-
efficiencies over existing methods (i.e. museum collections, literature), while
strengthening local capacity for data collection.
Location Venezuela.
Methods We placed a grid over Venezuela (0.5 9 0.5 degree cells) and applied
a stratified sampling design to select a minimum set of 25 cells spanning envi-
ronmental and biogeographical variation. We implemented standardized field
sampling protocols for birds, butterflies and dung beetles, along transects on
environmental gradients (‘gradsects’). We compared species richness estimates
from our field surveys at national, bioregional and cell scales to those calculated
from data compiled from museum collections and the literature. We estimated
the variance in richness, composition, relative abundance and diversity between
gradsects that could be explained by environmental and biogeographical
variables. We also estimated total survey effort and cost.
Results In one field season, we covered 8% of the country and recorded 66%
of all known Venezuelan dung beetles, 52% of Pierid butterflies and 37% of
birds. Environmental variables explained 27–60% of variation in richness for all
groups and 13–43% of variation in abundance and diversity in dung beetles
and birds. Bioregional and environmental variables explained 43–58% of the
variation in the dissimilarity matrix between transects for all groups.
Main conclusions NeoMaps provides reliable estimates of richness, composi-
tion and relative abundance, required for rigorous monitoring and spatial
prediction. NeoMaps requires a substantial investment, but is highly efficient,
achieving survey goals for each group with 1-month fieldwork and about US$
1–8 per km2. Future work should focus on other advantages of this type of
survey, including the ability to monitor the changes in relative abundance and
turnover in species composition, and thus overall diversity patterns.
Keywords
Birds, butterflies, dung beetles, Neotropical Biodiversity Mapping Initiative,
richness, Venezuela.
INTRODUCTION
Tropical Latin America houses an enormous share of the
world’s biodiversity (Myers et al., 2000; Rahbek & Graves,
2001; Pimm & Brown, 2004). Developing measures to
describe spatial patterns and detect temporal trends in bio-
diversity is fundamental for systematic conservation planning,
management and monitoring, as well as for meeting many
DOI: 10.1111/ddi.12012ª 2012 Blackwell Publishing Ltd http://wileyonlinelibrary.com/journal/ddi 215
Diversity and Distributions, (Diversity Distrib.) (2013) 19, 215–231

international legal conservation obligations (Dobson, 2005;
UNEP, 2007). However, even the simplest measures of biodi-
versity based on species distributions, richness and relative
abundance are lacking for the vast majority of Neotropical
taxa and ecosystems (Prance, 1994; Castro & Locker, 2000;
Patterson, 2001; Rodrıguez, 2003).
A solution to this lack of basic knowledge is to carry out
large-scale, systematic surveys (e.g. Duro et al., 2007; Nielsen
et al., 2009). Various random sampling techniques and
exhaustive surveys have been tested in the industrialized
world, with notable examples in Australia, Europe and North
America (e.g. Margules, 1989; Sauer & Droege, 1990; Gib-
bons et al., 1993; Margules & Redhead, 1995; Sauer et al.,
2008; Eaton et al., 2009; EuMon, 2009). However, such
large-scale systematic monitoring efforts are scarce in the
tropical regions, where most initiatives have been concen-
trated in networks of field stations across several countries,
but with low sampling intensity within any individual region
(Ahumada et al., 2011; Jurgens et al., 2012).
Taxonomists and conservation-oriented ecologists and
biologists are overwhelmingly concentrated in industrialized
nations (Gaston & May, 1992; UNPD, 2003; Rodrıguez et al.,
2005). Unless developing countries can build up their own
institutions and cadre of competent researchers, little effec-
tive biodiversity monitoring will be accomplished (Barret
et al., 2001; Rodrıguez et al., 2006). A major conservation
research problem is thus how best to accomplish such moni-
toring within time-scales relevant to urgent management
needs, at the lowest possible cost and effort (Pereira &
Cooper, 2006; Sutherland et al., 2009).
A common response to this sampling efficiency question
has been to make use of surrogate measures of environmen-
tal diversity or to rely on previously collected data. Environ-
mental diversity summarizes the variability in environmental
conditions across a given area along continuous gradients,
usually based on ordination of multiple variables, and can be
estimated using maps of climate variables, geophysical char-
acteristics and remotely sensed land cover (Faith & Walker,
1996). However, environmental diversity may not accurately
capture the complex patterns in species richness and compo-
sition resulting from biogeographical processes and historical
events (Hortal & Lobo, 2006). Previously collected data have
been used in an attempt to overcome this weakness, and
come from specimens deposited in natural history museums
or herbaria, field guides and in-depth surveys at few loca-
tions (Ponder et al., 2001; Chernoff et al., 2003; Ridgely
et al., 2003). Statistical modelling can combine sparse histori-
cal data with environmental variables for predictive mapping
of species distribution or richness, but taxonomic, temporal
and spatial biases in sampling effort can impose serious limi-
tations on this approach (Yesson et al., 2007; Sastre & Lobo,
2009).
As an alternative, we propose a cost-effective, systematic
monitoring programme for the assessment of tropical biodi-
versity, which we tested in Venezuela and may serve as a
model for other tropical countries. With the Neotropical
Biodiversity Mapping Initiative (NeoMaps), we aimed to pro-
duce conservation-relevant estimates of species richness and
relative abundance while explicitly addressing the problems of
cost and time required for large-scale surveys and strengthen-
ing the local capacity necessary to carry them out (Rodrıguez
& Sharpe, 2002). Our aim was not to provide complete inven-
tories, which existing methods accomplish well, but rather to
provide estimates of indexes based on community composi-
tion and relative abundance. These are difficult to derive from
existing information, but are valuable for monitoring (Linden-
mayer & Likens, 2010), modelling (Ferrier & Guisan, 2006)
and to complement distributional and species richness data at
the national, regional and local levels (Hortal & Lobo, 2005a).
We present results from work conducted between 2001
and 2010 and evaluate three specific questions in the context
of our larger aims: (1) How much does NeoMaps add to
prior knowledge of species inventories in Venezuela? (2)
How reliable is the NeoMaps baseline for future monitoring
goals? (3) How representative is the NeoMaps sample for
community-level modelling and prediction? We first devel-
oped a biogeographically and environmentally stratified sam-
pling design to minimize the effort necessary to characterize
diversity within our sampling universe. Next, we compiled
prior knowledge of birds, butterflies and dung beetles from
the literature and natural history collections. Then, we con-
ducted field surveys for these same groups. Finally, we analy-
sed and compared the patterns derived from these diverse
data to assess the relative benefits of each source of informa-
tion and highlight directions for future research.
The present study focused on birds (Aves), butterflies (Lepi-
doptera: Rhopalocera) and dung beetles (Coleoptera: Scara-
baeinae) in Venezuela. We chose these groups for three
principal reasons. First, existing standardized sampling meth-
ods make large-scale sampling feasible for them (Newmark,
1985; Gardner et al., 2008b; Sauer et al., 2008; Schmeller et al.,
2009; Ferrer-Paris et al., 2013; Rodrıguez et al., 2012). Second,
they are found throughout the ecosystems they occupy, play-
ing a variety of roles as herbivores, pollinators, predators, pri-
mary and secondary seed dispersers and ‘compost recyclers’,
among others (Kremen, 1994; Nichols et al., 2007; Roy et al.,
2007; Gardner et al., 2008a; Sirami et al., 2009). Third, several
global and regional initiatives focus on the study of these three
groups (Avian Knowledge Network, 2009; TABDP, 2009; Sca-
rabNet, 2011), but Neotropical and particularly Venezuelan
data are scarce in most of them. Venezuela, with its wide vari-
ety of ecosystems within a fairly large geographical region
(< 1,000,000 km2) and a good road network comparable in
density to most Neotropical countries (Digital Chart of the
World Data retrieved from Hijmans et al., 2012), represents
an ideal pilot location for testing NeoMaps protocols (MARN,
2000; Szeplaki et al., 2001; Aguilera et al., 2003).
METHODS
Our sampling universe consisted of the 170 cells with at least
30% terrestrial coverage that were accessible by road within
216 Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd
J. R. Ferrer-Paris et al.

the Venezuelan Biodiversity Grid (VBG) (Fig. 1, Data S1 in
Supporting Information). Contemporary environmental fac-
tors are well known to affect the patterns of species richness
and relative abundance (Currie, 1991; Brown & Mehlman,
1995; Guegan et al., 1998; Gaston, 2000; Rahbek & Graves,
2001; Ricklefs, 2004), and provide a framework for an objec-
tive classification and stratification of the territory for eco-
logical surveys (Bunce et al., 1996). We compiled available
spatial datasets for altitude, temperature, precipitation, num-
ber of dry months, total forest cover and deciduous forest
cover and calculated the mean and range of values for each
cell in our sampling universe (Bliss & Olsen, 1996; DeFries
et al., 2000; Hijmans et al., 2005; Table 1). We used princi-
pal component analysis (PCA) to compact these variables
into three independent axes with a biophysical interpretation
(Jongman et al., 1995) that explained 71% of the total vari-
ance. The first, an ‘elevation-seasonality’ axis, was dominated
by mean elevation, precipitation mean and range, tempera-
ture range and the mean number of dry months per year.
The second, a ‘forest cover’ axis, was influenced by the mean
and range of the total and deciduous forest cover. The third,
a ‘humidity’ axis, reflected mean precipitation, the range of
the number of dry months and the range of the deciduous
forest cover (Table 1, Data S1).
Given that environmentally similar locations in different
regions may have different species assemblages due to their
particular histories (Ricklefs, 2004), we next divided the sam-
pling universe into major biogeographical regions (Linares,
1988; MARN, 2000; Hilty, 2003), further grouped by prox-
imity and similarity, to minimize size differences among
biogeographical units (Metzger et al., 2005). The final set
consisted of five bioregions (Fig. 1), from west to east:
(1) Occident (OC), a combination of the Perija mountain
range, the Maracaibo Lake basin and the Lara-Falcon
drylands; (2) Andean mountains (AM); (3) Coastal moun-
tains (CM), grouping the central and eastern segments of the
Cordillera de la Costa; (4) Orinoco floodplain (LL), known
in Venezuela as llanos, and (5) Guayana shield (GS).
Applying a random stratified sampling design, we selected
a minimum set of cells with equal representation of environ-
mental and biogeographical variation (Beever, 2006; Ruxton
& Colegrave, 2006), and up to four additional cells targeting
those regions or habitat types that were considered subre-
presented.
With limited resources, there is always a trade-off between
the number of cells sampled and the sampling effort in each
cell (Beever, 2006). To optimize sampling effort, a 40-km
road transect was identified within each cell covering the
largest possible gradient in environmental variation (Austin
& Heyligers, 1991; Wessels et al., 1998). Sampling sites were
located along these ‘gradsects’ according to the taxon-specific
protocols and were spaced sufficiently apart to minimize spa-
tial and temporal autocorrelation (Liebhold & Sharov, 1998;
Fisher, 1999). For butterflies, 8–12 sampling points were
Figure 1 The Venezuelan Biodiversity Grid, showing the sampling universe (shaded cells) divided into the bioregions considered:
Occident (OC), Andean mountains (AM), Coastal mountains (CM), Orinoco floodplain (LL), Guayana shield (GS). Inset shows
Venezuela at the northern tip of South America.
Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd 217
Neotropical biodiversity surveys

visited each day, with the starting point/direction chosen ran-
domly, and consecutive points located at least 4 km apart.
For dung beetles, baited traps were placed in groups, with at
least 5 km between groups. For birds, point counts were
performed 800 m apart, resulting in 50 stops per gradsect.
Sampling protocols were developed during 2001–05 to ensure
that fieldwork was simple, repeatable and relatively rapid
(Table 2, Ferrer-Paris et al., 2013; Rodrıguez et al., 2012).
Since 2001, NeoMaps has invested more than 2000 per-
son-hours sampling butterflies, 200,000 trap-hours sampling
dung beetles and 150 person-hours counting birds. The pres-
ent study examines a subset of these data (Table 2). For but-
terflies, we included the Pieridae collected in 2006, of which
all specimens were adequately identified. Sampling effort var-
ied from 23 to 70 person-hours per gradsect. For dung bee-
tles, we used all 2009 gradsects where specimens had been
completely identified, and complemented it with six grad-
sects from 2006 to generate a pooled dataset. Sampling effort
was usually 4500–6500 trap-hours per gradsect, except for
two cases with 3000–4000 trap-hours. For birds, we used the
data for the 27 gradsects visited in 2010, with 150 min of
effort by one person at each gradsect (50 point
counts 9 3 min/point count).
For the analysis, we first summarized results from field
surveys (‘NeoMaps’ data) on species richness at a national
and bioregional level and compared them with pre-existing
knowledge from museums and the literature (‘prior’ data,
Tables S1–S3) and with the values obtained using all sources
of data (‘total’ data). Then, we analysed the relationship
between environmental and biogeographical variables with
different measures of species diversity between transects
(abundance, richness, diversity and composition). All analy-
ses were performed using R (R Core Development Team,
2005) and the vegan add-on package (Dixon, 2003).
Relative performance of NeoMaps and prior data
We examined estimates of species richness and, where possi-
ble, diversity, composition and relative abundance in NeoM-
aps and prior data at three spatial scales: national,
bioregional and individual cells. Adequately surveyed cells
were defined as those with enough records for the applica-
tion of any richness estimator. Alternatively, inadequately
surveyed cells were those with few records leading to very
low (near zero) or very high (greater than Sobs) standard
errors, unrealistic estimates of richness, and/or to very wide
confidence intervals for completeness (Nakamura & Soberon,
2008).
National species accumulation curves were calculated from
a ‘species by site’ matrix S of the number of individuals of
each species detected in each cell (NeoMaps data) or the
number of records of each species in each cell (prior data).
We used the method of moments with an unconditioned
standard deviation to estimate the mean of each species
accumulation curve and its 95% confidence intervals
(Colwell et al., 2004).
At a bioregional scale, we compared the average and the
total number of species in each bioregion, using Chao’s for-
mula for incidence-based estimation of species richness for
each bioregion and for the entire sampling universe. We esti-
mated inventory completeness as Cchao = Sobs/Schao (Chao
et al., 2005; Nakamura & Soberon, 2008) for NeoMaps data,
prior data and total data.
At the individual cell scale, we tried to move beyond com-
parisons of the simple number of species detected to include
measures of diversity, composition and relative abundance.
However, this was not straightforward because data sources
had different units (e.g. number of records vs. number of
individuals). Thus, for prior data, we applied the frequency-
based formula of (Chao et al. 2005), but many cells were
inadequately surveyed (see Tables S4–S6; Nakamura &
Soberon, 2008). For birds, we assumed that the lists of
expected species based on the literature and expert knowl-
edge represented the ‘true’ species richness for the cells sam-
pled by NeoMaps, and calculated the Jaccard index of biotic
dissimilarity between them (Chao et al., 2005). For NeoMaps
data, we were able to estimate species richness, diversity,
Table 1 Biological, physical and climatic variables initially
considered for the environmental stratification of NeoMaps’
0.5 9 0.5° cells, showing their relative weight on the first three
principal components (PC1–3). The eigenvalue of the fourth
component was 0.75, and the sum of the eigenvalues of the
remaining components was 2.12.
Variable Abbreviation PC1 PC2 PC3
Longitude of
cell centre
long (*) – – –
Latitude of
cell centre
lat (*) – – –
Mean elevation elev.avg �0.41 0.16 0.18
Range of elevation (*) – – –
Mean total annual
precipitation
prec.avg �0.26 �0.33 �0.45
Range of total
annual precipitation
prec.rng �0.45 0.07 �0.17
Mean annual
temperature
(*) – – –
Range of annual
temperature
temp.rng �0.42 0.20 �0.23
Mean number of
dry months
Dry.avg 0.45 0.18 �0.17
Range of number of
dry months
Dry.rng �0.03 0.21 �0.60
Mean total forest
cover
cover.avg �0.25 �0.48 �0.09
Range of total
forest cover
cover.rng �0.19 �0.36 �0.22
Mean deciduous
forest cover
Dec.avg 0.18 �0.51 �0.24
Range of deciduous
forest cover
Dec.rng 0.22 �0.35 �0.42
Eigenvalue 3.05 2.45 1.58
(*) Collinear variables excluded from further analyses.
218 Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd
J. R. Ferrer-Paris et al.

composition and relative abundance per cell directly
(Tables S4–S6). These estimates included (1) a richness
estimator based on abundances of species [abundance-base
coverage estimation (ACE); Colwell & Coddington, 1994],
(2) the Shannon–Wiener index of diversity (H) (Luoto et al.,
2004), (3) Chao’s version of the Jaccard index of dissimilar-
ity (Dchao) between cells (Chao et al., 2005) and (4) the stan-
dard deviation of log-abundances (log r). We used the
species by site matrix described above (S) and calculated the
estimators (1), (2) and (4) for each row, using the raw num-
ber of individuals as a measure of relative species abundance
for each transect. We applied (3), Dchao, to each pair of rows
in S in order to estimate biotic dissimilarity or beta diversity
between gradsects, generating a new matrix, D.
Explanatory power of stratifying variables
For NeoMaps data, we examined the explanatory power of
environmental gradients, biogeographical strata and spatial
location with respect to the variation in species richness
(SACE), diversity (H) and relative abundance (log r). For
each variable, we fitted a series of linear models to examine
potential predictors in a ‘variable by site’ matrix (E). We fit-
ted all possible combinations of models using continuous val-
ues of the three PCA axes for each cell (regression on principal
components, Legendre & Legendre, 2003), with biogeographi-
cal region as a categorical variable, and a second-order polyno-
mial of the longitude (long) and latitude (lat) of each cell.
Thus, our full model took the form y = PC1 + PC2 +PC3 + BIOREG + (long + lat)2, in which y was either SACE, H
or log r. Because imperfect detection of species, especially in
more complex habitats, might lead to sampling errors whose
standard deviation is not constant across all values of explana-
tory variables, we used weighted least squares to improve
model performance (Carroll & Ruppert, 1988). In our case, we
used a measure of completeness, CACE, as an estimate of the
weights for each observation. We then calculated a small-sam-
ple-size-corrected version of the Akaike Information Criterion
(AICc) for each model, ranked the models according to their
relative differences in AICc (DAICc) and calculated their AICc
weights (wi Burnham et al., 2011).
Finally, to evaluate how environmental variables and/or
bioregions explained the variation in species composition, we
applied a permutational multivariate analysis of variance
(PMANOVA) to D using the contents of E as the indepen-
dent variables and applying a simple model with no interac-
tions. PMANOVA is a nonparametric analogue to MANOVA
and describes how variation in a multivariate distance matrix
may be attributed to different experimental treatments or
uncontrolled covariates (Anderson, 2001; McArdle & Ander-
son, 2001). This method relies on distance matrices rather
than least squares, and significance of the terms is assessed by
using pseudo-F-ratios based on sequential sums of squares
from permutations of the raw data (McArdle & Anderson,
2001).
RESULTS
Relative performance of NeoMaps and prior data
Published and museum records for all taxa were distributed
throughout the VBG, covering between 36% and 52% of the
country, but fewer than half of those cells could be consid-
ered to be adequately surveyed (Fig. 2). Montane regions
(AM and CM in Fig. 1) contained cells that were well cov-
ered by prior surveys for all three groups, while GS was well
surveyed for dung beetles and birds. However, in LL and
OC, prior data were only adequate for birds.
In the equivalent of 1 month of fieldwork, NeoMaps cov-
ered c. 15% of the sampling universe (8% of the country),
detected 52% of previously recorded pierid butterflies, 37%
of birds and 66% of dung beetles, and extended the coverage
of georeferenced data for these groups. NeoMaps surveys
nearly doubled the number of adequately surveyed cells
within the sampling universe for dung beetles (prior data:
32; NeoMaps added 25) and butterflies (prior data: 24;
NeoMaps added 27), including several cells that had no prior
data at all (15 new cells for butterflies and nine for dung
beetles, mostly in GS and LL). For birds, there was consider-
able overlap between NeoMaps and prior data: 20 of 27
NeoMaps cells had prior data, and 11 of those could be con-
sidered adequately surveyed prior to NeoMaps (Fig. 2).
Table 2 Summary of NeoMaps field surveys and sampling effort (2001–10). For butterflies and birds, fieldwork is measured in person-
hours; for dung beetles, it is measured in trap-hours.
Taxon Year Objective
Cells
(n)
Fieldwork
(h)
No. of
persons
Specimens or
observations
Identified
(%) Data used in this article
Butterflies 2003–05 Calibration 10 309 10 5413 69 All cells, one family:
5504 specimens2006 National survey 27 1268 20 23,796 57
2009–10 National survey 29 885 32 11,530 61
Dung beetles 2005 Calibration 5 25,419 10 16,457 100
2006 Pilot survey 9 44,111 20 24,738 100 6 cells: 12,688 specimens
2009–10 National survey 26 129,816 32 c. 70,000 74 19 cells: 58,590 specimens
Birds 2001–02 Calibration 4 52 3 5451 90
2010 National survey 27 107.5 24 12,518 97 All cells, 1 day of sampling:
8573 observations
Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd 219
Neotropical biodiversity surveys

With respect to richness estimates, in general NeoMaps
had a slightly better performance than prior data at the scale
of the cells, but had mixed performance at larger geographi-
cal scales. For the Pieridae, NeoMaps richness estimates were
lower than values reported or estimated from prior data at
both national and bioregional levels, and bioregional esti-
mates were very similar despite the great differences observed
by previous data sources (Table 3). Disagreement between
data sources was more evident in bioregions that had been
better sampled in the past, such as CM, where NeoMaps
captured only a small fraction of known species and pro-
vided only one or two new species records, resulting in small
changes in the total estimates (Fig. 3, Table 3). NeoMaps’
cell averages for pierid species richness were higher and vari-
ation lower than for previous data (Fig. 4), and in direct
comparison of cells sampled by both data sources, NeoMaps
had larger values of Sobs in 14 of 18 cells, but differences
between sources were not significant (Wilcoxson-signed-rank
test, Z = 1.76, P-value = 0.08, Table S4). NeoMaps estimates
of completeness were very high at all scales (almost always
above 70%, Tables 3 and S4), and while prior data have
reached an almost complete national inventory, regional
completeness was comparable, between 67% and 88%
(Table 3).
For dung beetles, NeoMaps data performed almost uni-
formly better than prior data: the total numbers of species
at national, regional and local levels, as well as incidence-
based estimates, were the same or higher in NeoMaps data,
except in the Andes (Figs 3 and 4, Tables 3 and S5). At the
national level, completeness was similar for all sources
(Table 3). Both NeoMaps and prior data made similar pre-
dictions for LL and OC, but NeoMaps predicted the highest
richness in GS and CM, while prior data had the highest
estimates in AM. However, total Sobs and SChao were higher
than estimates from either of NeoMaps or prior data
(Table 3). In 15 of 17 cells with data from both sources,
NeoMaps achieved consistently higher values of Sobs (Wil-
coxson-signed-rank test, Z = 3.410, P-value < 0.001) and
very high CACE values that suggested nearly complete local
inventories (Table S5). NeoMaps had intermediate values of
Figure 2 Geographical distribution of samples used in this study for butterflies, dung beetles, and birds. Circles indicate cells visited
by NeoMaps survey teams since 2005; in black are those used in the present analysis (see Table 2). Grey cells are those with
georeferenced records from museums, literature or biodiversity databases: dark grey=adequately surveyed cells, pale grey=inadequately
surveyed.
220 Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd
J. R. Ferrer-Paris et al.

0 20 40 60 80
040
8012
0 Butterflies
Viloria (1990): 106 spp.
0 20 60 100
040
8012
0
Dung beetles
ScarabNet: 120 spp.
0 20 60 100
050
010
0015
00
Aves
Hilty (2003): 1383 spp.
VBG cells
Acc
umul
ated
spe
cies
rich
ness
Figure 3 Species accumulation curves
for all cells sampled in the sampling
universe by NeoMaps field surveys (dark
grey 95% confidence interval) and by
prior data sources (museums or
literature, pale grey 95% confidence
interval) for butterflies, dung beetles, and
birds. The dotted accumulation curve in
the dung beetle chart is for NeoMaps
data using only formally described
species (see methods). The horizontal
dotted line represents total described
species to date in each group according
to the best available published source.
Butterflies
020
4060 7681
Dung beetles
010
2030
40 42
Aves
010
020
030
0
AM LLOC CM GY
434
25%
50%
75%
1.5 interquantilerange
Outlyingobservations
NeoMaps Previousdata
Num
ber o
f spe
cies
per
cel
l
Figure 4 Species richness in different
bioregions (Fig. 1) estimated from
NeoMaps and previous data sources for
butterflies, dung beetles, and birds.
Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd 221
Neotropical biodiversity surveys

Table
3Estim
ates
ofspeciesrichnessin
each
bioregionandthewhole
samplinguniverse
(see
Fig.1foracronym
s)from
NeoMaps,other
datasources
andalldatacombined.
NeoMaps
Prior
Total
nS o
bs
S chao
Cchao
nS o
bs
S chao
Cchao
nS o
bs
S chao
Cchao
Butterflies
OC
738
60.22±13.26
0.63
(0.56–0.71)
2157
75.38±10.53
0.76
(0.69–0.82)
2263
73.32±6.56
0.86
(0.80–0.92)
AM
329
38.85±6.45
0.75
(0.65–0.84)
1389
119.03
±14.24
0.75
(0.70–0.79)
1391
125.32
±16.45
0.73
(0.68–0.77)
CM
424
26.45±2.55
0.91
(0.80–1.01)
1781
92.12±7.05
0.88
(0.83–0.93)
1882
96.73±9.09
0.85
(0.80–0.90)
LL
824
30.4
±5.92
0.79
(0.67–0.91)
2761
91.15±15.17
0.67
(0.61–0.73)
3065
95.38±15.62
0.68
(0.63–0.74)
GS
525
28.6
±3.85
0.87
(0.76–0.99)
1828
38.08±8.00
0.74
(0.63–0.84)
2238
44.4
±5.92
0.86
(0.76–0.95)
SU27
5762.5
±4.39
0.91
(0.85–0.97)
96106
111.63
±4.18
0.95
(0.92–0.98)
105
110
115.63
±4.18
0.95
(0.92–0.98)
Dungbeetles
OC
335
55.05±11.54
0.64
(0.56–0.71)
1421
53±23.32
0.4(0.33–0.47)
1746
72.04±13.82
0.64
(0.57–0.70)
AM
543
64.13±10.85
0.67
(0.60–0.74)
1357
129.25
±36.59
0.44
(0.40–0.48)
1380
160.67
±34.85
0.5(0.46–0.53)
CM
451
94.68±21.53
0.54
(0.49–0.59)
1841
61.17±11.31
0.67
(0.60–0.74)
1871
120.85
±22.74
0.59
(0.54–0.63)
LL
447
67.17±11.31
0.7(0.63–0.77)
3332
62.08±19.27
0.52
(0.44–0.59)
3366
92.28±12.83
0.72
(0.66–0.77)
GS
991
127±15.87
0.72
(0.67–0.76)
2840
58.06±11.65
0.69
(0.61–0.77)
33115
151.75
±14.89
0.76
(0.72–0.80)
SU25
134
207.92
±27.82
0.64
(0.61–0.68)
106
105
155±21.36
0.68
(0.64–0.72)
114
193
283.74
±29.43
0.68
(0.65–0.71)
Birds
OC
5175
263.89
±26.52
0.66
(0.64–0.69)
24426
624.83
±38.77
0.68
(0.67–0.70)
25450
626.09
±34.67
0.72
(0.71–0.73)
AM
3149
393.16
±65.6
0.38
(0.36–0.39)
13512
634.24
±23.75
0.81
(0.80–0.82)
13526
644.22
±23.16
0.82
(0.80–0.83)
CM
5198
503.31
±82.76
0.39
(0.38–0.41)
19574
682.94
±22.04
0.84
(0.83–0.85)
22595
708.78
±22.81
0.84
(0.83–0.85)
LL
6203
285.22
±24.73
0.71
(0.69–0.74)
38475
799.11
±73.29
0.59
(0.58–0.61)
41492
757.69
±58.39
0.65
(0.64–0.66)
GS
8291
408.19
±27.14
0.71
(0.70–0.73)
26637
772.54
±25.9
0.82
(0.81–0.84)
28664
799.04
±26.09
0.83
(0.82–0.84)
SU27
526
684.9±30.04
0.77
(0.76–0.78)
120
1044
1133.39±18.22
0.92
(0.91–0.93)
129
1075
1164.65±18.42
0.92
(0.92–0.93)
n,number
ofcellssurveyed;S o
bs,speciesobserved
inallcellscombined;S c
hao,incidence-based
estimateofspeciesrichness±SE
;Cchao,estimated
completenessoftheinventory
(range),based
onS c
hao.
222 Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd
J. R. Ferrer-Paris et al.
Cchao (50–70%) at the bioregional level, which were similar
to total values and significantly higher than values for prior
data in OC, AM and LL, although they were significantly
lower in CM.
Finally, for birds, NeoMaps data performed similarly as
for Pieridae at national level, detecting fewer species than
prior data and adding few new species records per region
(Fig. 3, Table 3). At the bioregional level, NeoMaps data
improved inventories in OC and LL, where total complete-
ness was significantly higher than for prior data (with low to
medium values of CACE). NeoMaps’ cell values of Sobs had
slightly higher means and smaller variation than prior data,
especially in OC and LL (Fig. 4), but in direct comparison of
cells sampled by both data sources, NeoMaps had signifi-
cantly higher values in nine previously inadequately surveyed
cells and significantly lower values in nine of 11 previously
adequately surveyed ones (Wilcoxson-signed-rank test,
Z = 2.666, P-value = 0.007, and Z = �2.40, P-value = 0.016,
respectively, Table S6). Unlike other taxonomic groups,
though, observed and estimated richness corresponded
poorly for both data sources, because both sources detected
more species in GS, but predicted more species elsewhere
(CM for NeoMaps and LL for prior data), which does not
seem to be correct according to the relevant literature (Hilty,
2003 and references in Table S3).
At local level, NeoMaps data captured only a small pro-
portion of previously known richness: SACE for NeoMaps
data was only 40% of the expected species lists for each tran-
sect (range: 16–76%), and their correlation was weak (Pear-
son r = 0.40, t = 2.192, d.f. = 25, P = 0.037), particularly in
GS, where species lists included more than 400 species but
NeoMaps estimates were low (Table S6 and Fig. S1).
Excluding GS improved the correlation (Pearson r = 0.69,
t = 3.966, d.f. = 17, P = < 0.001). NeoMaps estimates of
biotic distance were also well correlated with estimates based
on prior data (Fig. S1, Mantel r = 0.639, P = 0.001 with GS,
and r = 0.706, P = 0.001 without GS, based on 1000 permu-
tations).
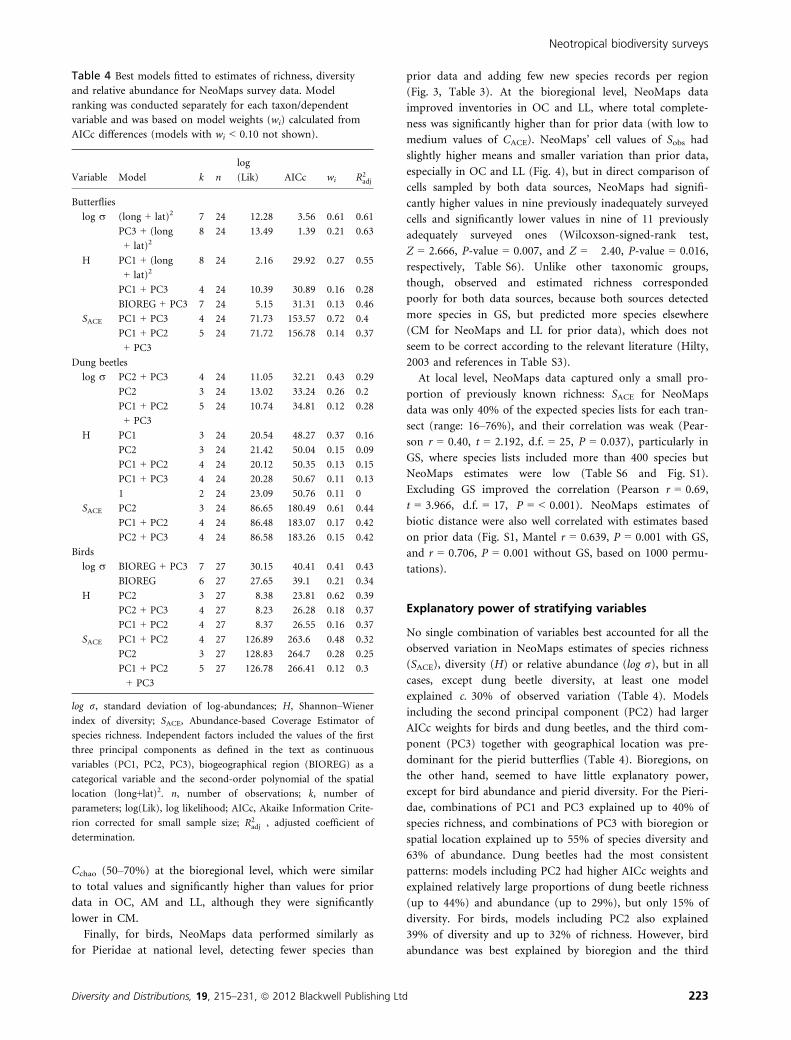
Explanatory power of stratifying variables
No single combination of variables best accounted for all the
observed variation in NeoMaps estimates of species richness
(SACE), diversity (H) or relative abundance (log r), but in all
cases, except dung beetle diversity, at least one model
explained c. 30% of observed variation (Table 4). Models
including the second principal component (PC2) had larger
AICc weights for birds and dung beetles, and the third com-
ponent (PC3) together with geographical location was pre-
dominant for the pierid butterflies (Table 4). Bioregions, on
the other hand, seemed to have little explanatory power,
except for bird abundance and pierid diversity. For the Pieri-
dae, combinations of PC1 and PC3 explained up to 40% of
species richness, and combinations of PC3 with bioregion or
spatial location explained up to 55% of species diversity and
63% of abundance. Dung beetles had the most consistent
patterns: models including PC2 had higher AICc weights and
explained relatively large proportions of dung beetle richness
(up to 44%) and abundance (up to 29%), but only 15% of
diversity. For birds, models including PC2 also explained
39% of diversity and up to 32% of richness. However, bird
abundance was best explained by bioregion and the third
Table 4 Best models fitted to estimates of richness, diversity
and relative abundance for NeoMaps survey data. Model
ranking was conducted separately for each taxon/dependent
variable and was based on model weights (wi) calculated from
AICc differences (models with wi < 0.10 not shown).
Variable Model k n
log
(Lik) AICc wi R2adj
Butterflies
log r (long + lat)2 7 24 12.28 �3.56 0.61 0.61
PC3 + (long
+ lat)28 24 13.49 �1.39 0.21 0.63
H PC1 + (long
+ lat)28 24 �2.16 29.92 0.27 0.55
PC1 + PC3 4 24 �10.39 30.89 0.16 0.28
BIOREG + PC3 7 24 �5.15 31.31 0.13 0.46
SACE PC1 + PC3 4 24 �71.73 153.57 0.72 0.4
PC1 + PC2
+ PC3
5 24 �71.72 156.78 0.14 0.37
Dung beetles
log r PC2 + PC3 4 24 �11.05 32.21 0.43 0.29
PC2 3 24 �13.02 33.24 0.26 0.2
PC1 + PC2
+ PC3
5 24 �10.74 34.81 0.12 0.28
H PC1 3 24 �20.54 48.27 0.37 0.16
PC2 3 24 �21.42 50.04 0.15 0.09
PC1 + PC2 4 24 �20.12 50.35 0.13 0.15
PC1 + PC3 4 24 �20.28 50.67 0.11 0.13
1 2 24 �23.09 50.76 0.11 0
SACE PC2 3 24 �86.65 180.49 0.61 0.44
PC1 + PC2 4 24 �86.48 183.07 0.17 0.42
PC2 + PC3 4 24 �86.58 183.26 0.15 0.42
Birds
log r BIOREG + PC3 7 27 30.15 �40.41 0.41 0.43
BIOREG 6 27 27.65 �39.1 0.21 0.34
H PC2 3 27 �8.38 23.81 0.62 0.39
PC2 + PC3 4 27 �8.23 26.28 0.18 0.37
PC1 + PC2 4 27 �8.37 26.55 0.16 0.37
SACE PC1 + PC2 4 27 �126.89 263.6 0.48 0.32
PC2 3 27 �128.83 264.7 0.28 0.25
PC1 + PC2
+ PC3
5 27 �126.78 266.41 0.12 0.3
log r, standard deviation of log-abundances; H, Shannon–Wiener
index of diversity; SACE, Abundance-based Coverage Estimator of
species richness. Independent factors included the values of the first
three principal components as defined in the text as continuous
variables (PC1, PC2, PC3), biogeographical region (BIOREG) as a
categorical variable and the second-order polynomial of the spatial
location (long+lat)2. n, number of observations; k, number of
parameters; log(Lik), log likelihood; AICc, Akaike Information Crite-
rion corrected for small sample size; R2adj , adjusted coefficient of
determination.
Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd 223
Neotropical biodiversity surveys

principal component (43%, Table 4). Alternative models fit-
ted with ordinary least squares (unweighted observations)
show similar results, but the percentage of explained varia-
tion was much lower for butterflies and moderately higher
for birds and dung beetles (results not shown).
For species composition, results were more consistent
across taxonomic groups: the explanatory power of environ-
mental variables and bioregions was slightly higher in all
cases, and the role of variables reversed. Bioregions explained
between 30% and 44% of the variance in species composi-
tion for all groups, with an additional 12–15% explained by
PCA scores (Table 5). Residual variance was lower in butter-
flies and birds than it was in dung beetles.
DISCUSSION
Here, we aimed to develop comparable and efficient esti-
mates of species richness, composition and relative abun-
dance at different geographical scales, so that major changes
may eventually be detected in subsequent sampling seasons.
In general, the NeoMaps protocols achieved these aims well:
although results varied across scales and taxonomic groups,
we sampled comparable numbers of species, with lower vari-
ance as prior data and with a lower time and resource
investment per unit area. Our sampling design produced
estimates of diversity and relative abundance that were
impossible to produce with prior data.
The performance of a systematic survey approach such as
NeoMaps must consider the taxonomic group, scale and
expected outputs. For example, if a survey is intended to
improve the knowledge of a taxonomic group at the national
level, the primary goal should be to maximize complemen-
tarity with existing sources, by targeting either poorly known
species groups (such as dung beetles in Venezuela) or poorly
surveyed regions for an otherwise well-known group (like
birds in OC and LL, or butterflies in GS). Alternatively, if
the main goal is to provide a baseline for monitoring, then
the survey should focus on indicator species or diversity pat-
terns that are expected to respond to human-driven change
(Lindenmayer & Likens, 2010). But if the intended output is
to infer or predict diversity patterns from the observed
locations to the entire region or country, then the focus
should be on robust estimation of correlations with environ-
mental variables and performance of a spatially representa-
tive sampling (Teder et al., 2007). Below, we examine the
relative merits of NeoMaps for inventory, monitoring and
modelling.
Contribution of NeoMaps sampling to a more
complete inventory
Inventories aim to obtain an accurate species list for a loca-
tion or a region, including the typically large number of rare
species in an area (Longino & Colwell, 1997). NeoMaps is of
limited use for inventories at large geographical scales (coun-
try or bioregion), because only a fraction of the species
recorded over many years of previous research were detected.
NeoMaps’ ability to fill in local and regional knowledge gaps
was higher, but depended on the level of prior knowledge
for each taxonomic group. For well-known groups such as
birds, NeoMaps made modest contributions to a larger
inventory: in previously well-surveyed cells, NeoMaps
detected only a fraction of the previously recorded birds spe-
cies (Table S6), but samples in new cells improved regional
estimates and completeness in regions that had less prior
study, such as LL and OC (Table 3).
Table 5 PMANOVA analysis of NeoMaps species composition estimate.
Taxon Variable d.f.
Sum of
squares
Mean
squares F R2 P
Pieridae BIOREG 4 1.881 0.470 4.948 0.434 0.001
PC1 1 0.420 0.420 4.424 0.097 0.005
PC2 1 0.181 0.180 1.900 0.042 0.097
PC3 1 0.047 0.048 0.502 0.011 0.741
Residuals 19 1.805 0.095 0.417
Total 26 4.334 1
Scarabaeinae BIOREG 4 2.333 0.583 2.221 0.300 0.002
PC1 1 0.436 0.435 1.658 0.056 0.106
PC2 1 0.164 0.164 0.624 0.021 0.769
PC3 1 0.392 0.392 1.494 0.050 0.146
Residuals 17 4.464 0.262 0.573
Total 24 7.789 1
Birds BIOREG 4 2.867 0.717 3.860 0.384 0.001
PC1 1 0.535 0.535 2.881 0.072 0.001
PC2 1 0.250 0.250 1.347 0.034 0.183
PC3 1 0.283 0.283 1.521 0.038 0.104
Residuals 19 3.529 0.186 0.473
Total 26 7.463 1
F, value of the F-statistic; d.f., degrees of freedom; R2, adjusted coefficient of determination; P, P-value.
224 Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd
J. R. Ferrer-Paris et al.

By contrast, for more poorly known groups, such as dung
beetles, NeoMaps’ contribution to species inventories was
significant at all geographical scales, helping to solve an
historic taxonomic bottleneck (Kim & Byrne, 2006). NeoM-
aps contributed many new species records for all regions and
increased the regional and national totals beyond estimates
based on prior data (Fig. 4, Table 3), this is true even when
excluding unnamed taxa (Fig. 3). For example, NeoMaps
detected only 40–60% of the known Eurysternus and Onth-
ophagus species (Pulido & Zunino, 2007; Genier, 2009), but
added three (E. impressicolis, O. acuminatus and O. rubres-
cens) previously reported from Colombia or Brazil. In the
genera Ateuchus and Uroxys, there were 23 unnamed mor-
phospecies compared to just seven and two named species in
the literature for Venezuela (Martınez, 1988; Martınez &
Martınez, 1990). Thus, NeoMaps made a significant contri-
bution to the knowledge of this understudied group.
For Pieridae, NeoMaps’ geographical coverage was compa-
rable to the previous coverage of well-surveyed cells, but it
did not improve current knowledge substantially. Although it
is a well-known group at the national scale with a nearly
complete inventory, there are many local gaps in geographi-
cal coverage, and regional lists are far from complete (Fig. 2,
Table 3). In three of five regions, > 68 years of prior data
resulted in twice as many species as NeoMaps (Fig. 4), due
primarily to rare species with a low detection probability
(Lobo, 2008). Among the species absent were eight relatively
widespread (seven or more cell records in prior data) and 19
very restricted species (one or two cells in prior data) (Le
Crom et al., 2004; Bollino & Costa, 2007). Even with or with-
out NeoMaps’ contribution, however, butterfly richness in
Guayana remains underrepresented (Table 3, Fig. 4). A close
examination shows that both common, widespread species (e.
g. Phoebis agarite and Itaballia pandosia) and uncommon ones
(e.g. several species in genera Dismorphia and Enantia) are
missing from both prior data sources and NeoMaps. Indeed,
this region is considered one of the most species-rich but
least-studied areas for Venezuelan butterflies (Viloria, 2000).
Although NeoMaps was not designed for this purpose, an
optimal sampling design for inventories would be much like a
gap analysis: first identifying well-surveyed sites and then
defining a set of complementary locations that best represent
environmental conditions in the non-surveyed portions of the
sampling universe (Hortal & Lobo, 2005b; Williams et al.,
2006). For groups with poor prior data, more cells would need
to be added, or sampling effort increased per cell, or comple-
mentary methods added to increase detectability.
Reliability of baseline for future monitoring
Monitoring species richness and diversity through time does
not require an exhaustive account of all species, but rather an
informative metric that is representative of the variable of
interest and sensitive enough to detect changes in time
(Beever, 2006); by these criteria, NeoMaps data clearly outper-
form prior data (Rivadeneira et al., 2009; Jaric & Ebenhard,
2010). When local samples are repeatable and low in variance,
and species detectability is high or at least constant, they can
be reliably used to estimate local richness and other measures
of species diversity and composition and to make compari-
sons in time and space (Yoccoz et al. 2001). Incomplete local
surveys may still be useful for monitoring, provided that they
achieve an acceptable level of completeness and remain repre-
sentative of the assemblage (Colwell & Coddington, 1994;
Beccaloni & Gaston, 1995). For example, NeoMaps dung
beetle samples captured high number of species with high
completeness (Table 3), while bird local estimates were
correlated with species richness in cells with low to moderate
richness (< 400 sp.), and measures of biotic distance based on
NeoMaps data were correlated with expected dissimilarity
(Table S6, Fig. S1; Rodrıguez et al., 2012).
The most straightforward approach to using NeoMaps
data for monitoring would be to focus on the subset of taxa
that it reliably detected, including common and widespread
species, and use them as biodiversity or ecological indicators
(Larsen et al., 2009). For example, common butterfly species
have experienced large changes in distribution and abun-
dance in parts of Europe, both declining or increasing, and
these changes have been related to habitat/landscape changes
(Van Dyck et al., 2009). Trends in populations of species
with high or moderately high abundance are easier to detect
than trends in rare species, or species with high fluctuations
in abundance (Meyer et al., 2010). Furthermore, the abun-
dance of moderately common species can be used either as
an direct indicator of changes in habitat or a predictor of
the likelihood of presence/absence of other species with cor-
related habitat-use patterns but much lower detectability
(Mac Nally et al., 2003). The richness and composition esti-
mates used here (SACE, SChao, DChao) estimated a proportion
of undetected species for each site, but did not consider sys-
tematic factors that could affect detectability within (e.g. dif-
ferences between species) or between sites (e.g. landscape or
habitat structures). Estimates of detection probability could
be built into the monitoring design through replicated sam-
ples across sites (Kery & Schmid, 2004; Dorazio et al., 2006)
or through a double sampling approach, which makes use of
intensive sampling in selected calibration sites (Rodrıguez
et al., 2012).
NeoMaps sampling could of course be improved using a
general conceptual framework for future monitoring. Includ-
ing a measure of threats (e.g. climate change, deforestation,
desertification) or management interventions (e.g. conserva-
tion areas, reforestation programmes) in the sampling design
would increase the capability of NeoMaps to evaluate their
effects on any detected change, rather than just document
general trends (Lindenmayer & Likens, 2010). For example,
the degree and timing of land use changes and fragmentation
could be used as a measure of habitat modification and loss,
which are considered major drivers of species extinction risk
for terrestrial arthropods and birds (Thomas & Abery, 1995;
Koh et al., 2004; Thomas et al., 2004; BirdLife International,
2008). In the case of dung beetles, abundance appears to
Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd 225
Neotropical biodiversity surveys

decline with increasing habitat modification, and open
habitat communities contain a hyperabundance of few small-
bodied species (Nichols et al., 2007). This expected relation-
ship between habitat fragmentation and the number of
individuals of small-bodied species could be easily explored
with NeoMaps data in a long-term monitoring programme.
Representative sampling for community-level
prediction
NeoMaps captured much of the range of variation in a set of
derived environmental and biogeographical variables, which
successfully explained a considerable proportion of the varia-
tion in species richness and relative abundance (Table 4) and
a greater proportion in composition (Table 5). Such vari-
ables were clearly influenced by attributes such as habitat
heterogeneity, precipitation and temperature (Table 1),
which have been highlighted as important determinants of
spatial richness patterns for birds, butterflies and dung bee-
tles (Beccaloni & Gaston, 1995; Rahbek & Graves, 2001; van
Rensburg et al., 2002; Hayes et al., 2009). NeoMaps’ design
aimed to balance the contribution of these ecological and
historical components to produce data suitable for several
techniques of species-level and community-level prediction
of diversity patterns (Ferrier & Guisan, 2006), which are use-
ful for extrapolation at national and bioregion scales, facili-
tating planning, monitoring and action (Colwell & Coddington,
1994; Gotelli & Colwell, 2001). In these applications, prior
data had major limitations due to incomplete geographical
and environmental coverage and unbalanced taxonomic
representation (Loiselle et al., 2008). NeoMaps data from
dung beetles at all scales and from birds at the local scale
were as representative as prior data – but not so for
butterflies. For species-level prediction, taxonomical bias
might still be an important limitation because several species
are underrepresented in our samples.
Again, our results point to ways for improving sampling
design to increase the number of sampled species and improve
their spatial representation. Bioregional and environmental
variables explained a great proportion of compositional pat-
terns, but sample selection might need to be optimized for
each group separately in future surveys, spreading samples
along the gradients with the strongest associations with varia-
tion in the group (Hortal & Lobo, 2005a).
As expected, forest cover (PC2) was the variable that best
correlated with NeoMaps estimates of bird species richness
and diversity per cell, even though completeness was lower in
increasingly forested areas. The distribution of bird abundances
(log r) was related to bioregion and the humidity gradient
(PC3). Indeed, higher species diversity and evenness is to be
expected in forested areas, where detectability is probably lower
(or more heterogeneous) due to the complexity of the habitat,
while higher concentrations of individuals were expected in the
seasonally flooded Llanos (Stotz et al., 1996).
For dung beetles, forest cover was also the most important
variable explaining richness and abundance, but elevation-
seasonality was more correlated with diversity. These results
are in agreement with studies in other Neotropical regions
(Escobar et al., 2006; Nichols et al., 2007), and suggest that
NeoMaps should increase sampling effort at low elevations
and in forested regions in future surveys. Finally, for butter-
flies, the spatial component and the humidity gradient (PC3)
were better related to the patterns observed, but other evi-
dence (like the absence of a great proportion of mountain
species) suggests that we are underestimating the importance
of the elevational (PC1) and the forest cover gradient (PC2)
to species richness (Beccaloni & Gaston, 1995; Le Crom
et al., 2004).
Cost-effectiveness
The efficacy of any biodiversity monitoring effort must be
evaluated not only in terms of the quantity or thoroughness
of the data collected, but also by the investments made to
obtain those data (Margules & Austin, 1991; Gardner et al.,
2008b; Bried, 2009). The total monetary cost of NeoMaps in
the present study (including field and laboratory equipment,
survey costs and salaries for experts in taxonomic identifica-
tion) was a substantial investment (US$ 362,820), but con-
sidering the area sampled, the investment was more efficient
than other initiatives (about US$ 1–8 per km2 sampled vs.
US$ c. 70–90 per km2 for other multitaxa Neotropical biodi-
versity assessments; Gardner et al., 2008b). For conservation
planning, monitoring and management, the area covered is
more relevant than the number of specimens sampled; the
most important conservation actions take place increasingly
within particular units of space (i.e. in protected areas)
rather than on a species-by-species basis (Beever, 2006; Legg
& Nagy, 2006). This is particularly the case when the taxa
being monitored are used as indicators, rather than as being
of particular conservation interest in their own right (Larsen
et al., 2009). Thus, the NeoMaps sampling design was clearly
preferable in cost-effectiveness, and its applicability in other
Neotropical countries with similar infrastructure should be
straightforward (also see Supporting Information).
None of these figures include transportation costs to or
between sampling localities, although this is likely to repre-
sent a relatively smaller investment (compared to the total) if
sampling relies on a good road network where costs would
rise proportionally to the number of sampled localities, the
distance between them and the mean transportation cost per
km, which is tightly linked to mean gas prices. Each of
NeoMaps’ surveys in Venezuela required around 18,000 km
of road travel between and within sampling units, which
would cost around 2000–3000 US$ in most South American
countries, assuming gas prices between 1.1 and 1.6 US$ per l,
and a consumption of 10 km l�1.
Sampling localities outside the road network might be very
important for inventory purposes, but would increase total
survey costs, limiting the number of localities that can be
sampled. For Venezuela, we estimated that sampling one
locality outside the road network could require a 10-fold
226 Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd
J. R. Ferrer-Paris et al.

increase in costs, compared to a locality within the road net-
work. The difference is due mostly to higher transportation
costs, increased travelling time between localities and the lar-
ger personnel or time investment needed to achieve an
equivalent sampling effort.
Such an investment might be reasonable if the selected
localities contribute significantly to an improvement in the
overall data or information gathered by the survey. This is
arguably the case when the primary goal of the survey is
inventorying, and the chosen methods have high detectability
or completeness, or when the additional localities represent
very different environmental conditions that are complemen-
tary to those already sampled. However, we consider that the
actual extent and density of available roads in South America
would be sufficient to sample the principal ecosystems or
ecoregions in most countries, except Bolivia and the Guay-
anas (Hijmans et al., 2012).
CONCLUSIONS
The challenge of monitoring tropical biodiversity requires a
long-term, sustained effort and the continuous evaluation of
methods, procedures and goals (Lindenmayer & Likens,
2010). Ultimately, a comprehensive biodiversity survey strat-
egy for a tropical country like Venezuela should draw infor-
mation from all available complementary sources, including
large-scale national surveys, museum or literature records
and regional and local research initiatives. Systematic
national biodiversity surveys such as NeoMaps have a partic-
ularly important role to play, by providing estimates of rela-
tive abundance, and thus diversity and composition,
required for rigorous monitoring and spatial prediction of
community-level attributes. It is clear, however, that NeoM-
aps’ simple sampling design will need to be refined in future,
as data emerging from the surveys are examined in the light
of taxon-specific patterns and comprehensive sets of environ-
mental variables. Such analyses will allow the exploration of
modifications in either the spatial sampling design or the
field survey protocols (or both) to improve the likelihood of
detecting biodiversity changes, and thus support monitoring,
or provide data adequate for predicting spatial patterns at
the community level.
In future, it will also be important to evaluate additional
advantages of NeoMaps-type sampling efforts not considered
in depth here, including the ability to estimate relative abun-
dances and species absences, and thus overall diversity pat-
terns – all of which are difficult or impossible to estimate
with traditional museum data and the taxonomic literature
(Beck & Kitching, 2007).
ACKNOWLEDGEMENTS
We are grateful for support from the Biodiversity Analysis
Unit of the Andean Centre for Biodiversity Conservation
at Conservation International, the Conservation Technology
Support Program, the Disney Wildlife Conservation Fund,
EcoHealth Alliance, the Venezuelan Fondo Nacional de
Ciencia, Tecnologıa e Innovacion, the Instituto Venezolano
de Investigaciones Cientıficas, the Latin America and
Caribbean Program of the National Audubon Society, Pro-
vita and UNESCO. Major funding was provided by Total
Venezuela, S. A., as part of the Program for the Support
of the Conservation of the Biodiversity of Venezuela,
under the framework of the Ley Organica de Ciencia,
Tecnologıa e Innovacion (LOCTI). We are particularly
indebted to L. A. Solorzano, A. Grajal and C. J. Sharpe
for their involvement at earlier stages of NeoMaps. This
project would have been impossible without the help of
hundreds of national and international student and profes-
sional volunteers. Museum data were facilitated by R. A.
Briceno (MEJMO), J. Clavijo, L. J. Joly and Q. Arias
(MIZA), and J. Camacho (MALUZ). Comments from Lluis
Brotons and three anonymous referees helped improve the
manuscript.
REFERENCES
Aguilera, M., Azocar, A. & Gonzalez-Jimenez, E. (2003) Bio-
diversidad en Venezuela. Tomo I y II. Fundacion Polar.
Ministerios de Ciencia y Tecnologıa. Fondo Nacional de
Ciencia, Tecnologıa e Innovacion, Caracas, Venezuela.
Ahumada, J.A., Silva, C.E.F., Gajapersad, K., Hallam, C., Hur-
tado, J., Martin, E., McWilliam, A., Mugerwa, B., O’Brien,
T., Rovero, F., Sheil, D., Spironello, W.R., Winarni, N. &
Andelman, S.J. (2011) Community structure and diversity
of tropical forest mammals: data from a global camera trap
network. Philosophical Transactions of the Royal Society B:
Biological Sciences, 366, 2703–2711.
Anderson, M.J. (2001) A new method for non-parametric
multivariate analysis of variance. Austral Ecology, 26,
32–46.
Austin, M.P. & Heyligers, P.C. (1991) New approach to vege-
tation survey design: gradsect sampling. Nature conservation:
cost effective biological surveys and data analysis. CSIRO –
Commonwealth Scientific and Industrial Research Organi-
zation, Canberra, Australia.
Avian Knowledge Network (2009) Avian knowledge network:
an online database of bird distribution and abundance.
Available at: http://www.avianknowledge.net (accessed 3
October 2010).
Barret, C.B., Brandon, K., Gibson, C. & Gjertsen, H. (2001)
Conserving tropical biodiversity amid weak institutions.
BioScience, 51, 497–502.
Beccaloni, G.W. & Gaston, K.J. (1995) Predicting species
richness of Neotropical forest butterflies: ithomiinae (Lepi-
doptera: Nymphalidae) as indicators. Biological Conserva-
tion, 71, 77–86.
Beck, J. & Kitching, J. (2007) Estimating regional species
richness of tropical insects from museum data: a compari-
son of a geography-based and sample-based methods. Jour-
nal of Applied Ecology, 44, 672–681.
Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd 227
Neotropical biodiversity surveys

Beever, E.A. (2006) Monitoring biological diversity: strate-
gies, tools, limitations, and challenges. Northwestern Natu-
ralist, 87, 66–79.
BirdLife International (2008) State of the world’s birds: indi-
cators for our changing world. BirdLife International, Cam-
bridge, UK.
Bliss, N.B. & Olsen, L.M. (1996) Development of a 30-arc-
second digital elevation model of South America. Pecora
Thirteen, Human Interactions with the Environment – Per-
spectives from Space, Sioux Falls, SD.
Bollino, M. & Costa, M. (2007) An illustrated annotated
check-list of the species of Catasticta (s.l.) Butler (Lepidop-
tera: Pieridae) of Venezuela. Zootaxa, 1469, 1–42.
Bried, J.T. (2009) Information costs of reduced-effort habitat
monitoring in a butterfly recovery program. Journal of
Insect Conservation, 13, 615–626.
Brown, J.H. & Mehlman, D.W. (1995) Spatial variation in
abundance. Ecology, 76, 2028–2043.
Bunce, R.G.H., Barr, C.J., Clarke, R.T., Howard, D.C. & Lane,
A.M.J. (1996) Land classification for strategic ecological
survey. Journal of Environmental Management, 47, 37–60.
Burnham, K.P., Anderson, D.R. & Huyvaert, K.P. (2011)
AIC model selection and multimodel inference in behav-
ioral ecology: some background, observations, and compar-
isons. Behavioral Ecology and Sociobiology, 65, 23–35.
Carroll, R.J. & Ruppert, D. (1988) Transformation and
weighting in regression. Chapman & Hall, New York.
Castro, G. & Locker, I. (2000) Mapping conservation invest-
ments: an assessment of biodiversity funding in Latin Amer-
ica and the Caribbean. Biodiversity Support Program,
Washington, DC.
Chao, A., Chazdon, R.L., Colwell, R.K. & Shen, T.J. (2005) A
new statistical approach for assessing similarity of species
composition with incidence and abundance data. Ecology
Letters, 8, 148–159.
Chernoff, B., Machado-Allison, A., Riseng, K. & Montamba-
ult, J. (2003) Una evaluacion rapida de los ecosistemas
acuaticos de la cuenca del Rıo Caura, Estado Bolıvar, Vene-
zuela, no. 28. Conservation International, Washington, DC.
Colwell, R.K. & Coddington, J.A. (1994) Estimating terres-
trial biodiversity through extrapolation. Philosophical
Transactions of the Royal Society B: Biological Sciences, 345,
101–118.
Colwell, R.K., Mao, C.X. & Chang, J. (2004) Interpolating,
extrapolating, and comparing incidence-based species accu-
mulation curves. Ecology, 85, 2717–2727.
Currie, D.J. (1991) Energy and large-scale patterns of ani-
mal-species and plant-species richness. American Naturalist,
137, 27–49.
DeFries, R., Hansen, M., Townshend, J.R.G., Janetos, A.C. &
Loveland, T.R. (2000) A new global 1 km data set of per-
cent tree cover derived from remote sensing. Global Change
Biology, 6, 247–254.
Dixon, P.M. (2003) VEGAN, a package of R functions for
community ecology. Journal of Vegetation Science, 14,
927–930.
Dobson, A. (2005) Monitoring global rates of biodiversity
change: challenges that arise in meeting the Convention on
Biological Diversity (CBD) 2010 goals. Philosophical Trans-
actions of the Royal Society B: Biological Sciences, 360,
229–241.
Dorazio, R.M., Royle, J.A., Soderstrom, B. & Glimskar, A.
(2006) Estimating species richness and accumulation by
modeling species occurrence and detectability. Ecology, 87,
842–854.
Duro, D.C., Coops, N.C., Wulder, M.A. & Han, T. (2007)
Development of a large area biodiversity monitoring sys-
tem driven by remote sensing. Progress in Physical Geogra-
phy, 31, 235–260.
Eaton, M.A., Balmer, D.E., Conway, G.J., Grise, S., Hall, P.
V., Hearn, R., Musgrove, D., Risely, A.J. & Wootton, S.
(2009) The state of the UK’s birds 2008. T, Sandy, Bedford-
shire, RSPB, BTO, WWT, CCW, NIEA, JNCC, NE and
SNH, Bedfordshire, UK.
Escobar, F., Lobo, J.M. & Halffter, G. (2006) Assessing the
origin of Neotropical mountain dung beetle assemblages
(Scarabaeidae: Scarabaeinae): the comparative influence of
vertical and horizontal colonization. Journal of Biogeogra-
phy, 33, 1793–1803.
EuMon (2009) EU-wide monitoring methods and systems of
surveillance for species and habitats of Community interest,
Developed and maintained by EuMon, EBONE, and SCALES
for the European biodiversity monitoring community. Avail-
able at: http://eumon.ckff.si/index1.php (accessed 27 April
2010).
Faith, D.P. & Walker, P.A. (1996) Environmental diversity:
on the best-possible use of surrogate data for assessing the
relative biodiversity of sets of areas. Biodiversity and
Conservation, 5, 399–415.
Ferrer-Paris, J.R., Sanchez-Mercado, A. & Rodrıguez, J.P.
(2013) Optimizacion del muestreo de invertebrados tropi-
cales: un ejemplo con escarabajos coprofagos (Coleoptera:
Scarabaeinae) en Venezuela. Revista de Biologıa Tropical,
61, in press.
Ferrier, S. & Guisan, A. (2006) Spatial modelling of biodiver-
sity at the community level. Journal of Applied Ecology, 43,
393–404.
Fisher, B.L. (1999) Improving inventory efficiency: a case
study of leaf-litter ant diversity in Madagascar. Ecological
Applications, 9, 714–731.
Gardner, T.A., Hernandez, M.I.M., Barlow, J. & A., P.C.
(2008a) Understanding the biodiversity consequences of
habitat change: the value of secondary and plantation for-
ests for neotropical dung beetles. Journal of Applied Ecology,
45, 883–893.
Gardner, T.A., Barlow, J., Araujo, I.S. et al. (2008b) The
cost-effectiveness of biodiversity surveys in tropical forests.
Ecology Letters, 11, 139–150.
Gaston, K.J. (2000) Global patterns in biodiversity. Nature,
405, 220–227.
Gaston, K.J. & May, R.M. (1992) Taxonomy of taxonomists.
Nature, 356, 281–282.
228 Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd
J. R. Ferrer-Paris et al.

Genier, F. (2009) Le genre Eurysternus Dalman, 1824 (Scara-
baeidae: Scarabaeinae: Oniticellini), revision taxonomique et
cles de determination illustrees. Pensoft Publishers, Sofia,
Bulgary.
Gibbons, D.W., Chapman, R. & Reid, J. (1993) The new atlas
of breeding birds in Britain and Ireland: 1988–91. T. & A.D.
Poyser, London, UK.
Gotelli, N.J. & Colwell, R.K. (2001) Quantifying biodiversity:
procedures and pitfalls in the measurement and compari-
son of species richness. Ecology Letters, 4, 379–391.
Guegan, J.F., Lek, S. & Oberdorff, T. (1998) Energy availabil-
ity and habitat heterogeneity predict global riverine fish
diversity. Nature, 391, 382–384.
Hayes, L., Mann, D.J., Monastyrskii, A.L. & Lewis, O.T.
(2009) Rapid assessments of tropical dung beetle and
butterfly assemblages: contrasting trends along a forest dis-
turbance gradient. Insect Conservation and Diversity, 2, 194
–203.
Hijmans, R.J., Cameron, S.E., Parra, J.L., Jones, P.G. & Jarvis,
A. (2005) Very high resolution interpolated climate sur-
faces for global land areas. International Journal of Clima-
tology, 25, 1965–1978.
Hijmans, R.J., Guarino, L. & Mathur, P. (2012) DIVA-GIS
version 7.5 manual. Available at: http://www.diva-gis.org/
gdata/ (accessed 2 February 2012).
Hilty, S.L. (2003) Birds of Venezuela, 2nd edn. Princeton
University Press, Princeton, NJ.
Hortal, J. & Lobo, J.M. (2005a) An ED-based protocol for
the optimal sampling of biodiversity. Biodiversity and Con-
servation, 14, 2913–2947.
Hortal, J. & Lobo, J.M. (2005b) A synecological framework
for systematic conservation planning. Biodiversity Informat-
ics, 9, 16–45.
Hortal, J. & Lobo, J.M. (2006) Towards a synecological
framework for systematic conservation planning. Biodiver-
sity Informatics, 3, 16–45.
Jaric, I. & Ebenhard, T. (2010) A method for inferring
extinction based on sighting records that change in fre-
quency over time. Wildlife Biology, 16, 267–275.
Jongman, R.H.G., Ter Braak, C.J.F. & van Tongeren, O.F.R.
(1995) Data analysis in community and landscape ecology.
Cambridge University Press, Cambridge, UK.
Jurgens, N., Schmiedel, U., Haarmeyer, D.H. et al. (2012)
The BIOTA Biodiversity Observatories in Africa—a
standardized framework for large-scale environmental
monitoring. Environmental Monitoring and Assessment, 184,
655–678.
Kery, M. & Schmid, H. (2004) Monitoring programs need to
take into account imperfect species detectability. Basic and
Applied Ecology, 5, 65–73.
Kim, K.C. & Byrne, L.B. (2006) Biodiversity loss and the tax-
onomic bottleneck: emerging biodiversity science. Ecology
Research, 21, 794–810.
Koh, L.P., Sodhi, N.S. & Brook, B.W. (2004) Co-extinctions
of tropical butterflies and their hostplants. Biotropica, 36,
272–274.
Kremen, C. (1994) Biological inventory using target taxa: a
case study of the butterflies of Madagascar. Ecological
Applications, 4, 407–422.
Larsen, F.W., Bladt, J. & Rahbek, C. (2009) Indicator taxa
revisited: useful for conservation planning? Diversity and
Distributions, 15, 70–79.
Le Crom, J.F., Llorente Bousquets, J., Constantino, L.M. &
Salazar, J.A. (2004) Mariposas de Colombia. Tomo II: pieri-
dae. Carlec Ltda, Bogota, Colombia.
Legendre, P. & Legendre, L. (2003) Numerical ecology. Else-
vier Science B.V., Amsterdam, the Netherlands.
Legg, C.J. & Nagy, L. (2006) Why most conservation moni-
toring is, but need not be, a waste of time. Journal of
Environmental Management, 78, 194–199.
Liebhold, A.M. & Sharov, A.A. (1998) Testing for correlation
in the presence of spatial autocorrelation in insect count
data. Population and community ecology for insect manage-
ment and conservation (ed. by J. Baumgartner, P. Brandm-
ayr and B.F.J. Manly), pp. 11–117. Balkema, Rotterdam,
the Netherlands.
Linares, O.J. (1988) Mamıferos de Venezuela. Sociedad Con-
servacionista Audubon de Venezuela, Caracas, Venezuela.
Lindenmayer, D.B. & Likens, G.E. (2010) The science and
application of ecological monitoring. Biological Conserva-
tion, 140, 1317–1328.
Lobo, J.M. (2008) Database records as a surrogate for sam-
pling effort provide higher species richness estimations.
Biodiversity and Conservation, 17, 873–881.
Loiselle, B.A., Jorgensen, P.M., Consiglio, T., Jimenez, I.,
Blake, J.G., Lohmann, L.G. & Montiel, O.M. (2008) Pre-
dicting species distributions from herbarium collections:
does climate bias in collection sampling influence model
outcomes? Journal of Biogeography, 35, 105–116.
Longino, J.T. & Colwell, R.K. (1997) Biodiversity assess-
ment using structured inventory: capturing the ant fauna
of a tropical rain forest. Ecological Applications, 7, 1263–
1277.
Luoto, M., Virkkala, R., Heikkinen, R.K. & Rainioc, K.
(2004) Predicting bird species richness using remote sens-
ing in boreal agricultural-forest mosaics. Ecological Applica-
tions, 14, 1946–1962.
Mac Nally, R., Fleishman, E., Fay, J.P. & Murphy, D.D.
(2003) Modelling butterfly species richness using mesoscale
environmental variables: model construction and validation
for mountain ranges in the Great Basin of western North
America. Biological Conservation, 110, 21–31.
Margules, C.R. (1989) Introduction to some Australian
developments in conservation evaluation. Biological Conser-
vation, 50, 1–11.
Margules, C.R. & Austin, M.P. (1991) Nature conservation:
cost effective biological surveys and data analysis. CSIRO –
Commonwealth Scientific and Industrial Research Organi-
zation, Canberra, Australia.
Margules, C.R. & Redhead, T.D. (1995) Guidelines for using
the BioRap methodology and tools. CSIRO and World Bank
BioRap Project, Dickson, Australia.
Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd 229
Neotropical biodiversity surveys

MARN (2000) Primer informe de paıs para la Convencion
sobre Diversidad Biologica. Ministerio del Ambiente y los
Recursos Naturales, Caracas, Venezuela.
Martınez, A. (1988) Algunos Scarabaeinae neotropicales nue-
vos o poco conocidos. Revista Cientıfica UNET, 2, 85–90.
Martınez, A. & Martınez, A. (1990) Dos nuevas especies de
Ateuchus Weber (Coleoptera: Scarabaeinae: Ateuchina).
Boletın de Entomologıa Venezolana, 5, 159–163.
McArdle, B.H. & Anderson, M.J. (2001) Fitting multivariate
models to community data: a comment on distance-based
redundancy analysis. Ecology, 82, 290–297.
Metzger, M.J., Bunce, R.G.H., Jongman, R.H.G., Mucher, C.
A. & Watkins, J.W. (2005) A climatic stratification of the
environment of Europe. Global Ecology and Biogeography,
14, 549–563.
Meyer, C.F.J., Aguiar, L.M.S., Aguirre, L.F. et al. (2010)
Long-term monitoring of tropical bats for anthropogenic
impact assessment: gauging the statistical power to detect
population change. Biological Conservation, 143, 2797–
2807.
Myers, N., Mittermeier, R.A., Mittermeier, C.G., da Fonseca,
G.A.B. & Kent, J. (2000) Biodiversity hotspots for conser-
vation priorities. Nature, 403, 853–858.
Nakamura, M. & Soberon, J. (2008) Use of approximate
inference in an index of completeness of biological inven-
tories. Conservation Biology, 23, 469–474.
Newmark, W.D. (1985) Legal and biotic boundaries of Wes-
tern North American national parks: a problem of congru-
ence. Biological Conservation, 33, 197–208.
Nichols, E., Larsen, T. & Spector, S. (2007) Global dung bee-
tle response to tropical forest modification and fragmenta-
tion: a quantitative literature review and meta-analysis.
Biological Conservation, 137, 1–19.
Nielsen, S., Haughland, D., Bayne, E. & Schieck, J. (2009)
Capacity of large-scale, long-term biodiversity monitoring
programmes to detect trends in species prevalence. Biodi-
versity and Conservation, 18, 2961–2978.
Patterson, B.D. (2001) Fathoming tropical biodiversity: the
continuing discovery of Neotropical mammals. Diversity
and Distributions, 7, 191–196.
Pereira, H.M. & Cooper, H.D. (2006) Towards the global
monitoring of biodiversity change. Trends in Ecology and
Evolution, 21, 123–129.
Pimm, S.L. & Brown, J.H. (2004) Domains of diversity. Sci-
ence, 304, 831–833.
Ponder, W.F., Carter, G.A., Flemons, P. & Chapman, R.R.
(2001) Evaluation of museum collection data for use in
biodiversity assessment. Conservation Biology, 15, 648–
657.
Prance, G.T. (1994) A comparison of the efficacy of higher
taxa and species numbers in the assessment of biodiversity
in the neotropics. Philosophical Transactions of the Royal
Society B: Biological Sciences, 345, 89–99.
Pulido, L.A. & Zunino, M. (2007) Catalogo preliminar de los
Onthophagini de America (Coleoptera: Scarabaeine Scara-
baeinae). M3M Monografıas, 7, 93–129.
R Core Development Team (2005) The R project for statistical
computing. Version v 2.3.1. R Core Development Team,
Vienna, Austria.
Rahbek, C. & Graves, G.R. (2001) Multiscale assessment of
patterns of avian species richness. Proceedings of the
National Academy of Sciences of the United States of Amer-
ica, 98, 4534–4539.
van Rensburg, B.J., Chown, S.L. & Gaston, K.J. (2002) Spe-
cies richness, environmental correlates, and spatial scale: a
test using South African birds. The American Naturalist,
159, 566–577.
Ricklefs, R.E. (2004) A comprehensive framework for global
patterns in biodiversity. Ecology Letters, 7, 1–15.
Ridgely, R.S., Allnutt, T.F., Brooks, T., McNicol, D.K., Mehl-
man, D.W., Young, B.E. & Zook, J.R. (2003) Digital distri-
bution maps of the birds of the Western Hemisphere version
1.0. NatureServe, Arlington, VA.
Rivadeneira, M.M., Hunt, G. & Roy, K. (2009) The use of
sighting records to infer species extinctions: an evaluation
of different methods. Ecology, 90, 1291–1300.
Rodrıguez, J.P. (2003) Challenges and opportunities for sur-
veying and monitoring tropical biodiversity – a response to
Danielsen et al. Oryx, 37, 411–411.
Rodrıguez, J.P. & Sharpe, C.J. (2002) NeoMaps: the Neo-
tropical biodiversity mapping initiative. Cotinga, 17, 13–14.
Rodrıguez, J.P., Simonetti, J.A., Premoli, A. & Marini, M.A.
(2005) Conservation in Austral and Neotropical America:
building scientific capacity equal to the challenges. Conser-
vation Biology, 19, 969–972.
Rodrıguez, J.P., Rodrıguez-Clark, K.M., Oliveira-Miranda, M.
A., Good, T. & Grajal, A. (2006) Professional capacity
building: the missing agenda in conservation priority
setting. Conservation Biology, 20, 1340–1340.
Rodrıguez, G.A., Rodrıguez, J.P., Ferrer-Paris, J.R. & San-
chez-Mercado, A. (2012) A nation-wide standardized bird
survey scheme for Venezuela. The Wilson Journal of Orni-
thology, 124, 230–244.
Roy, D.B., Rothery, P. & Brereton, T. (2007) Reduced-effort
schemes for monitoring butterfly populations. Journal of
Applied Ecology, 44, 993–1000.
Ruxton, G.D. & Colegrave, N. (2006) Experimental design for
the life-sciences. Oxford University Press, New York, NY.
Sastre, P. & Lobo, J.M. (2009) Taxonomist survey biases and
the unveiling of biodiversity patterns. Biological Conserva-
tion, 142, 462–467.
Sauer, J.R. & Droege, S. (eds) (1990) Survey designs and sta-
tistical methods for the estimation of avian population trends.
U.S. Fish and Wildlife Service, Washington, DC.
Sauer, J.R., Hines, J.E. & Fallon, J. (2008) The North Ameri-
can Breeding Bird Survey, results and analysis 1966–2007.
version 5.15.2008. USGS Patuxent Wildlife Research Center,
Laurel, MD.
ScarabNet (2011) Scarabaeinae research network. Available at:
http://216.73.243.70/scarabnet/ (accessed 2 May 2011).
Schmeller, D.S., Henry, P.Y., Julliard, R., Clobert, J., Gruber,
B., Dziock, F., Lengyel, S., Nowicki, P., Deri, E., Budrys, E.,
230 Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd
J. R. Ferrer-Paris et al.

Kull, T., Tali, K. & Bauch, B. (2009) Advantages of volun-
teer-based biodiversity monitoring in Europe. Conservation
Biology, 23, 307–316.
Sirami, C., Brotons, L. & Martin, J.L. (2009) Do bird spatial
distribution patterns reflect population trends in changing
landscapes? Landscape Ecology, 24, 893–906.
Stotz, D.F., Fitzpatrick, J.W., Parker, T.A., III & Moskovits,
D.K. (1996) Neotropical birds. Ecology and conservation.
University of Chicago Press, Chicago, IL.
Sutherland, W.J., Adams, W.M., Aronson, R.B. et al. (2009)
One hundred questions of importance to the conservation
of global biological diversity. Conservation Biology, 23, 557–
567.
Szeplaki, E., Garcıa, L., Rodrıguez, J. & Gonzalez, E. (2001)
Estrategia nacional sobre diversidad biologica y su plan de
accion. Ministerio del Ambiente. Oficina Nacional de Div-
ersidad Biologica, Caracas, Venezuela.
TABDP (2009) Tropical Andean Butterfly Diversity Program.
Available at: http://www.andeanbutterflies.org/index.html
(accessed 1 April 2009).
Teder, T., Moora, M., Roosaluste, E., Zobel, K., Partel, M.,
Koljalg, U. & Zobel, M. (2007) Monitoring of biological
diversity: a common-ground approach. Conservation Biol-
ogy, 21, 313–317.
Thomas, C.D. & Abery, J.C.G. (1995) Estimating rates of
butterfly decline from distribution maps – The effect of
scale. Biological Conservation, 73, 59–65.
Thomas, J.A., Telfer, M.G., Roy, D.B., Preston, C.D., Green-
wood, J.J.D., Asher, J., Fox, R., Clarke, R.T. & Lawton, J.H.
(2004) Comparative losses of British butterflies, birds, and
plants and the global extinction crisis. Science, 303, 1879–
1881.
UNEP (2007) Global environment outlook: environment for
development (GEO4). United Nations Environment Pro-
gramme (UNEP) and Progress Press Ltd, Valletta, Malta.
UNPD (2003) UNDP Annual Report 2003. United Nations
Development Programme, New York, NY.
Van Dyck, H., Van Strien, A.J., Maes, D. & Van Swaay, C.A.
M. (2009) Declines in common, widespread butterflies in a
landscape under intense human use. Conservation Biology,
23, 957–965.
Viloria, A.L. (2000) Estado actual del conocimiento taxono-
mico de las mariposas (Lepidoptera: Rhopalocera) de Ven-
ezuela. Hacia un proyecto CYTED para el inventario y
estimacion de la diversidad entomologica en Iberoamerica,
PRIBES-2000, pp. 261–274. Monografıas Tercer Milenio,
Zaragoza, Espana.
Wessels, K.J., Jaarsveld, A.S.V., Grimbeek, J.D. & Linde, M.J.
V.D. (1998) An evaluation of the gradsect biological survey
method. Biodiversity and Conservation, 7, 1093–1121.
Williams, P., Faithb, D., Mannec, L., Sechrestd, W. & Pre-
stone, C. (2006) Complementarity analysis: mapping the
performance of surrogates for biodiversity. Biological
Conservation, 128, 253–264.
Yesson, C., Brewer, P.W., Sutton, T., Caithness, N., Pahwa, J.
S., Burgess, M., Gray, W.A., White, R.J., Jones, A.C., Bisby,
F.A. & Culham, A. (2007) How global is the global biodi-
versity information facility? PLoS ONE, 2, e1124.
Yoccoz, N.G., Nichols, J.D. & Boulinier, T. (2001) Moni-
toring of biological diversity in space and time. Trends
in Ecology and Evolution, 16, 446–453.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the
online version of this article:
Data S1 Methods
Figure S1 Estimated versus expected species richness per
transect (top) and biotic distance between transects (bottom)
for Birds
Table S1 List of published sources for Pieridae distribution
data in Venezuela
Table S2 List of published sources for Scarabaeinae distribu-
tion data in Venezuela
Table S3 List of published sources for bird’s distribution
data in Venezuela
Table S4 Richness, diversity and completeness estimates for
the Pieridae data set in NeoMaps and previous data sources
Table S5 Richness and completeness estimates for dung bee-
tles’ data set in NeoMaps and previous data sources
Table S6 Richness and completeness estimates for bird’s data
set in NeoMaps and previous data sources
As a service to our authors and readers, this journal provides
supporting information supplied by the authors. Such mate-
rials are peer-reviewed and may be re-organized for online
delivery, but are not copy-edited or typeset. Technical sup-
port issues arising from supporting information (other than
missing files) should be addressed to the authors.
BIOSKETCH
NeoMaps is collaborative research effort based at the Vene-
zuelan Institute for Scientific Investigation (www.neomaps.
org). Our aim is to create an efficient and cost-effective
method for quantifying the distribution and abundance of
tropical biodiversity at large geographical scales. Although
our initial focus is on Venezuela, we are exploring imple-
mentation of our spatial sampling design and field survey
protocols in other Neotropical regions.
Author contributions: J.R.F.P., J.P.R., T.G. and A.S.M.
conceived the study; J.R.F.P., T.G. and G.A.R. led the field
teams; J.R.F.P., A.S. and G.A.R. identified the specimens; J.R.
F.P. analysed the data; and T.G., A.S.M., K.M.R.C., J.R.F.P.
and J.P.R. planned and wrote the manuscript.
Editor: Lluıs Brotons
Diversity and Distributions, 19, 215–231, ª 2012 Blackwell Publishing Ltd 231
Neotropical biodiversity surveys
Copyright © 2022 FDOKUMEN




















