
Synapsin-like Molecules in Aplysia punctata and Helix pomatia : Identification and Distribution in...
14
European hurnal of Neuroscience, Vol. 8, pp. 2530-2543, 1996 0 European Neuroscience Association Synapsin-like Molecules in Ap/ysia punctata and Helix pornatia: Identification and Distribution in the Nervous System and During the Formation of 6 Synaptic Contacts In Wtro Giuseppe Cibelli’, Mirella Ghirardi2, Franco Onofri3, Andrea Casadio215,Fabio Benfenati4, Pier Giorgio Montarolo2 and Francesco Vitiello’ ’Istituto di Fisiologia Umana, Universita di Bari, Policlinico, Piazza Giulio Cesare, 1-70124 Bari, Italy 2Dipartimentodi Neuroscienze, Sezione di Fisiologia, Universita di Torino, Torino, Italy 3Dipartimentodi Scienze Biomediche, Sezione di Fisiologia, Universita di Modena, Modena, Italy 4Dipartimentodi Medicina Sperimentale, Sezione di Fisiologia Umana, Universita di Roma ‘Tor Vergata’, Roma, Italy 5Present address: Center for Neurobiology and Behavior, Columbia University College of Physicians and Surgeons, 722 West 168th Street, New York, NY 10032, USA Keywords: synaptic vesicle proteins, invertebrate nervous system, neuronal cultures, synaptic plasticity Abstract The distribution and biochemical features of the synapsin-like peptides recognized in Aplysia and Helix by various antibodies directed against mammalian synapsins were studied. The peptides can be extracted at low pH and are digested by collagenase; further, they can be phosphorylated by both protein kinase A and Ca2+/calmodulin-dependent protein kinase II. In the ganglia of both snails, they are associated with the soma of most neurons and with the neuropil; punctate immunostaining is present along the neurites. Using cocultures of a Helix serotoninergic neuron and of its target cell, we analysed the redistribution of the synapsin-like peptides during the formation of active synaptic contacts. When the presynaptic neuron is plated in isolation, both synapsin and serotonin immunoreactivitiesare restricted to the distal axonal segments and to the growth cones; in the presence of the target, the formation of a chemical connection is accompanied by redistribution of the synapsin and serotonin immunoreactivities that concentrate in highly fluorescent round spots scattered along the newly grown neurites located close to the target cell. Almost every spot that is stained for serotonin is also positive for synapsin. In the presynaptic cell plated alone, the number of these varicosity-like structures is substantially stable throughout the whole period; by contrast, when the presynaptic cell synapses the target, their number increases progressively parallel to the increase in the mean amplitude of cumulative excitatory postsynaptic potentials recorded at the same times. The data indicate that mollusc synapsin-like peptides to some extent resemble their mammalian homologues, although they are not exclusively localized in nerve terminals and their expression strongly correlates with the formation of active synaptic contacts. Introduction A major step in the elucidation of synaptic function has been the discovery of the proteins associated with presynaptic specializations and involved in the storage and release of neurotransmitters. Most of these proteins have been highly conserved during evolution from yeasts to mammalian neurons, suggesting the idea that neuronal exocytosis is a particular aspect of the more general processes of intracellular vesicular fusion and constitutive secretion (Bajjalieh and Scheller, 1995 ). Although neurotransmitter release shares basic features with other kinds of exocytosis, it is a unique process in terms of efficiency, rapidity, plasticity and resistance to exhaustion. Many of these outstanding properties are contributed by the particular cytoarchitecture of the nerve terminal and, possibly, by the involve- ment of neuron-specific proteins (Valtorta and Benfenati, 1995). In addition, many of the nerve terminal proteins involved in exocytosis are phosphoproteins, suggesting that changes in the levels of second messengers and in the activity of protein kinases and phosphatases may affect the functional properties of neurotransmitter release (Greengard et al., 1993). A family of neuron-specific, synaptic vesicle-associated phospho- proteins, synapsins I and 11, has been implicated in the regulation of neurotransmitter release from mature nerve terminals and in the maturation of developing synapses. The synapsins are targets of the CAMP- and Ca’+/calmodulin-dependent phosphorylation processes in the brain. Due to their molecular diversity, both synapsins are phosphorylated by CAMP-dependent protein kinase (PKA) and Ca’+/calmodulin-dependent protein lunase I, but only synapsin 1 Correspondence rn: Prof. Francesco Vitiello, as above Received 20 March 1996, revised 26 July 1996, uccepted 2 August 1996
Transcript of Synapsin-like Molecules in Aplysia punctata and Helix pomatia : Identification and Distribution in...

European hurnal of Neuroscience, Vol. 8, pp. 2530-2543, 1996 0 European Neuroscience Association
Synapsin-like Molecules in Ap/ysia punctata and Helix pornatia: Identification and Distribution in the Nervous System and During the Formation of 6 Synaptic Contacts In Wtro
Giuseppe Cibelli’, Mirella Ghirardi2, Franco Onofri3, Andrea Casadio215, Fabio Benfenati4, Pier Giorgio Montarolo2 and Francesco Vitiello’ ’Istituto di Fisiologia Umana, Universita di Bari, Policlinico, Piazza Giulio Cesare, 1-70124 Bari, Italy 2Dipartimento di Neuroscienze, Sezione di Fisiologia, Universita di Torino, Torino, Italy 3Dipartimento di Scienze Biomediche, Sezione di Fisiologia, Universita di Modena, Modena, Italy 4Dipartimento di Medicina Sperimentale, Sezione di Fisiologia Umana, Universita di Roma ‘Tor Vergata’, Roma, Italy 5Present address: Center for Neurobiology and Behavior, Columbia University College of Physicians and Surgeons, 722 West 168th Street, New York, NY 10032, USA
Keywords: synaptic vesicle proteins, invertebrate nervous system, neuronal cultures, synaptic plasticity
Abstract
The distribution and biochemical features of the synapsin-like peptides recognized in Aplysia and Helix by various antibodies directed against mammalian synapsins were studied. The peptides can be extracted at low pH and are digested by collagenase; further, they can be phosphorylated by both protein kinase A and Ca2+/calmodulin-dependent protein kinase II. In the ganglia of both snails, they are associated with the soma of most neurons and with the neuropil; punctate immunostaining is present along the neurites. Using cocultures of a Helix serotoninergic neuron and of its target cell, we analysed the redistribution of the synapsin-like peptides during the formation of active synaptic contacts. When the presynaptic neuron is plated in isolation, both synapsin and serotonin immunoreactivities are restricted to the distal axonal segments and to the growth cones; in the presence of the target, the formation of a chemical connection is accompanied by redistribution of the synapsin and serotonin immunoreactivities that concentrate in highly fluorescent round spots scattered along the newly grown neurites located close to the target cell. Almost every spot that is stained for serotonin is also positive for synapsin. In the presynaptic cell plated alone, the number of these varicosity-like structures is substantially stable throughout the whole period; by contrast, when the presynaptic cell synapses the target, their number increases progressively parallel to the increase in the mean amplitude of cumulative excitatory postsynaptic potentials recorded at the same times. The data indicate that mollusc synapsin-like peptides to some extent resemble their mammalian homologues, although they are not exclusively localized in nerve terminals and their expression strongly correlates with the formation of active synaptic contacts.
Introduction
A major step in the elucidation of synaptic function has been the discovery of the proteins associated with presynaptic specializations and involved in the storage and release of neurotransmitters. Most of these proteins have been highly conserved during evolution from yeasts to mammalian neurons, suggesting the idea that neuronal exocytosis is a particular aspect of the more general processes of intracellular vesicular fusion and constitutive secretion (Bajjalieh and Scheller, 1995 ). Although neurotransmitter release shares basic features with other kinds of exocytosis, it is a unique process in terms of efficiency, rapidity, plasticity and resistance to exhaustion. Many of these outstanding properties are contributed by the particular cytoarchitecture of the nerve terminal and, possibly, by the involve- ment of neuron-specific proteins (Valtorta and Benfenati, 1995). In
addition, many of the nerve terminal proteins involved in exocytosis are phosphoproteins, suggesting that changes in the levels of second messengers and in the activity of protein kinases and phosphatases may affect the functional properties of neurotransmitter release (Greengard et al., 1993).
A family of neuron-specific, synaptic vesicle-associated phospho- proteins, synapsins I and 11, has been implicated in the regulation of neurotransmitter release from mature nerve terminals and in the maturation of developing synapses. The synapsins are targets of the CAMP- and Ca’+/calmodulin-dependent phosphorylation processes in the brain. Due to their molecular diversity, both synapsins are phosphorylated by CAMP-dependent protein kinase (PKA) and Ca’+/calmodulin-dependent protein lunase I, but only synapsin 1
Correspondence rn: Prof. Francesco Vitiello, as above
Received 20 March 1996, revised 26 July 1996, uccepted 2 August 1996

Synapsin-like molecules in Aplysia and Helix 253 1
is phosphorylated by Ca*+/calmodulin-dependent protein kinase I1 (CaMPKII) (Dc Camilli et al., 1990; Greengard et al., 1993).
A large body of experimental evidence points to the role of synapsin I in the regulation of neurotransmitter release. The introduction of dephosphorylated synapsin I, but not of synapsin I phosphorylated by CaMPKII, into the squid giant axon, the goldfish Mauthner neuron or nerve terminals isolated from the mammalian brain induces marked inhibition of the number of neurotransmitter quanta released in response to a constant stimulus (Llinis et al., 1985, 1991; Hackett et al., 1990; Nichols et al., 1992). Genetically altered mice lacking synapsin I, synapsin I1 or both synapsins (Chin et al., 1995; Li et al., 1995; Rosahl et al., 1995) exhibit marked alterations in the processes of presynaptic plasticity involving adaptive changes in the availability of synaptic vesicles for exocytosis. Indeed, synapsin I binds to synaptic vesicle membrane actin filaments and actin monomers in a phosphorylation-dependent manner (for reviews see De Camilli et al., 1990; Greengard et al., 1993). Through these activities, synapsin I has been demonstrated to cause the clustering of synaptic vesicles and to cross-link them to the actin cytoskeleton in a complex which is disrupted when synapsin I is phosphorylated by CaMPKII (Benfenati et al., 1992, 1993; Ceccaldi et al., 1995). Presynaptic injection of synapsin I antibodies results in the disappearance of synaptic vesicle clusters and is associated with a marked depression of high frequency- evoked neurotransmitter release (Pieribone et al., 1995). These data strongly implicate synapsin I in the formation of a reserve pool of vesicles and in the regulation of their availability for exocytosis (Greengard et al., 1993).
In addition to regulating neurotransmitter release, synapsins also participate in the formation of developing synapses. In cultured hippocampal neurons, synapsin I is precociously sorted to the axon and then it concentrates in presynaptic specializations as soon as synaptic contacts are formed (Fletcher et al., 1991 j. Further, functional data directly implicate synapsins in the formation of synaptic contacts. Overexpression of synapsin IIb in a neuroblastoma-glioma cell line induces, during differentiation, a dramatic increase in the number of varicosities along neurites and in the number of synaptic vesicles within the varicosities (Han et al., 1991). Loading of embryonic Xenopus motor neurons with either synapsin I or synapsin I1 induces accelerated structural and functional maturation of developing nerve terminals (Lu et al., 1992; Schaeffer et aE., 1994; Valtorta et al., 1995). On the other hand, cultured hippocampal neurons depleted of synapsin LI by antisense oligonucleotides bring about an impairment in the development of neurites and a decrease in the amount of synaptic vesicle proteins (Ferreira et aL, 1994).
The large size of neurons and the well-characterized neuronal networks make the mollusc nervous system extremely versatile for experimental manipulations of synaptic transmission (Schacher and Proshanski, 1983; Haydon el al., 1984; Ghirardi et al., 1996). Pre- and postsynaptic cells identified by morphological and electrophysio- logical criteria can be plated in culture, where they reliably form chemical synapses which show the same in vivo features (Schacher, 1985; Montarolo et al., 1986, 1988; Rayport and Schacher, 1986). Furthermore, in this model the influence of the target on neurite growth and synaptic plasticity has been identified and analysed (Glanzman et al., 1989; Schacher and Montarolo, 1991).
A previous phylogenetic survey of synapsin-like proteins has shown the presence in several non-mammalian vertebrate species of detectable immunoreactive proteins resembling mammalian synapsin I in terms of ti ssue-specific expression, subcellular distribution and biochemical properties (Goelz et al., 1985; Volknandt et al., 1987). Synapsin-like immunoreactivity (SLI) has been identified in the nervous system of Aplysia califarnica (Bongiovi et al., 1992). This
study, however, indicated that the synapsin I-like antigens are not as ubiquitous in the Aplysia nervous system as i n the mammalian brain, and that they fulfil only some of the defining criteria for synapsin I. Finally, a gene encoding various synapsin-like proteins has recently been cloned in Drnsophila (Klagges et al., 1996).
Taking advantage of the technique of invertebrate neuron culture, we have now extended these findings and studied the distribution of SLI in identified Aplysia and Helix neurons. We addressed the following questions: (i) are the synapsin-like proteins found in molluscs phosphorylatable by the same enzymes as those that phosphorylate synapsin I in mammals? (ii) does the sub- cellular distribution of synapsin-like proteins differ in the intact ganglion and in dissociated ganglion neurons in culture? (iii) does the distribution of SLI differ in the presynaptic neuron plated in isolation or co-cultured in the presence of its target cell? (iv) does the number of the SLI puncta increase as synaptic strength increases?
Some of these results have been presented i n abstract form (Cibelli et al., 1994).
Materials and methods Materials Juvenile and adult Aplvsia punctata were taken from the Adriatic sea; juvenile and adult Helix pomatia were obtained from a local breeder. Synapsin I was purified from bovinc brain as described by B a l e r and Greengard (1987). Recombinant rat synapsin IIa was expressed in Sf9 insect cells using the baculovirus expression system and purified by affinity chromatography as described by Siow et al. (1992). The catalytic subunit of PKA was purified from bovine heart as described by Kaczmarek et al. (1980), and CaMPKII was prepared from rat forebrain according to McGuinness et nl. (1985). Protein kinase activity was assessed using purified bovine synapsin I as substrate; both kinases phosphorylated synapsin I stoichiometrically at the appropriate sites. The CaMPKII inhibitory peptidc (CKIP), corresponding to residues 273-302 of the subunit of CaMPKII with Ala substituted for Thr-286, was kindly provided by Dr A. J. Czernik (Rockefeller University, New Yorkj. [y-”P]ATP (>3000 Ci/mmol), [‘251]anti-rabbit IgG (Fab fragments), biotinylated anti-rabbit IgG and biotinylated peroxidase-streptavidin complex were from Amersham (Little Chalfont, Bucks, UK). Calmodulin was from Boehringer Mannheim (Mannheim, Germany), highly purified collagenase form I11 was from Advance Biofactures Co. (Lynbrook, NY); protease IX, glutamine-free L15 medium, poly-L-lysine and fluorescein isothiocya- nate- or tetrmethyl rhodoamine isothiocyanate-conjugated secondary antibodies were from Sigma (St Louis, MO); the mouse anti-serotonin (5-HT) monoclonal antibody was from Dako (Glostrup, Denmark); and nitrocellulose membranes (0.2 pm pore size) were from Scleicher and Schuell (Dassel, Germany). The G-177 and G-178 antisera were obtained in rabbits by injecting purified bovine synapsin I [prepared by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS- PAGE) and electroeluted from the gel] with Freund’s complete adjuvant. The other anti-synapsin antibodies (G-94 and G-423) were kindly provided by Dr P. Greengard (Rockefeller University, New York). The antidplysia synaptotagmin antibody was a generous gift of Dr K. C. Martin (Center for Neurobiology and Behavior, Columbia University College of Physicians and Surgeons, New York).
All other chemicals of analytical grade were obtained from Sigma or Merck (Darmstadt, Germany).
Extraction of proteins from mollusc ganglia Molluscs were killed under anaesthesia with isotonic MgC12. Freshly dissected Aplysia and Helix ganglia or rat cerebral cortices were

2532 Synapsin-like molecules in Aplysia and Helix
homogenized with a Teflodglass homogenizer in warm electro- phoresis sample buffer (Laemmli, 1970). The homogenates were boiled for 4 min and the insoluble material was sedimented by centrifugation at 12 000 g for 30 min and discarded. The acid extracts were prepared essentially according to the method of Nestler and Greengard (1980). Briefly, ganglia and rat cerebral tissue were homogenized in ice-cold 5 mM HEPES, pH 7.4, containing 5 mM Zn2+-acetate. The homogenate was then centrifuged at 4000 g for 20 min, and the resulting pellet was resuspended in 1-3 ml of ice- cold 10 mM citric acid0.18 Nonidet P-40. After incubation in ice for 15 min, the samples were centrifuged at 7200 g for 20 min and the supernatant was neutralized by adding 1/10 (volhol) of 0.5 M Na2HP04. The neutralized acid extracts were either used for post hoc phosphorylation and digestion experiments or added to two-fold concentrated Laemmli sample buffer and boiled for 4 min.
Protein phosphorylation studies
The presence of phosphoproteins and protein kinase activities in nervous tissue from Aplysia and Helix was assessed by endogenous phosphorylation, post hoc phosphorylation and blot phosphorylation assays, and was evaluated as the incorporation of 32P into protein bands separated by SDS-PAGE.
Endogenous phosphorylation
Homogenates of mollusc ganglia (15-50 pg proteidsample) were incubated for 10 min at 30°C in a buffer containing 50 mM HEPES, 10 mM MgC12, 1 mM EGTA, 1 mM dithiothreitol, 50 pM [y-32P]ATP (1-2 pCi/sample) and protease inhibitors (0.2 mM phenylmethyl sulphonyl chloride, 10 pg/ml pepstatin, 10 p g h l chymostatin, 10 pglml antipain and 2 pg/ml leupeptin) in the absence or presence of CaCl, (2 mM), CaC1, (2 mh4) plus calmodulin (3 pg/sample), or CaClz (2 mM)/calmodulin (3 pg/ sample) plus CKIP (100 pM). The incubation was terminated by the addition of concentrated Laemmli sample buffer followed by boiling the samples for 4 min before SDS-PAGE and Western blotting.
Post hoc phosphor);lation
Freshly prepared, neutralized acid extracts of ganglia (15-50 pg protein/sample) were incubated for 30 min at 30°C under the same buffer conditions used for endogenous phosphorylation in the absence or the presence of either purified catalytic subunit of (1 pg/sample) or purified CaMPKII (50 nglsample) activated by CaCI2 (2 mM)/ calmodulin (3 pglsample).
Blot phosphovylation
The procedure for the assay of immobilized protein substrates for PKA and CaMPKII was that described by Valtorta et al. (1986). Briefly, acid extracts from ganglia (10-30 pg total protein) were separated by SDS-PAGE and electrophoretically transferred to nitro- cellulose membranes (see below). The blots were incubated overnight in a blocking solution containing 0.4% Ficoll 400 and 0.1% Triton X-100 in Tris-buffered saline. The phosphorylation reaction was carried out under the same conditions described for the post hoc phosphorylation assay except that the concentration of LY-~~P]ATP was 20 nM (10 pCi/ml) and that CaMPKII was previously autophos- phorylated with cold ATP prior to the addition to the overlay buffer. After incubation at room temperature for 90 min, the blots were extensively washed in PBS/O.l% Triton X-100, dried and exposed. After exposure, blots containing phosphorylated proteins were sub-
jected to immunolabelling of synapsin-like proteins as described below.
Digestion of synapsin I with collagenase
Acid extracts containing synapsin I or SLI (50 pg of protein) were incubated for 2 h at 30°C with 300 U purified collagenase (type 111) in a buffer containing 50 mM Tris-HC1 (pH 7 . 3 , 2.5 mM Ca2+- acetate and protease inhibitors (see above). The incubation was terminated by the addition of 10 mM EGTA followed by concentrated Laemmli sample buffer, and boiling for 4 min.
Electrophoresis and immunodetection
Proteins extracted from mollusc or rat nervous tissue were separated by SDS-PAGE according to Laemmli (1970) on 7-9% (wt/vol) polyacrylamide gels. In most cases proteins from unfixed gels were electrophoretically transferred (200 mA for 12 h) to nitrocellulose membranes (Towbin et al., 1979). Synapsin I immunoreactivity was detected by using specific rabbit polyclonal antisera and '251-labelled anti-rabbit Fab as the s e c o n d q antibody. When proteins had been phosphorylated with radioactive ATP, the blots were first auto- radiographed and subsequently subjected to immunolabelling with biotinylated secondary antibodies coupled with the biotin-streptavidin detection system using 4-chloro-1-naphthol as the dye. A battery of polyclonal antibodies directed against mammalian synapsin I and/or synapsin I1 (G-94, G-177, G-178, G-423) was used to detect SLI in mollusc nervous tissue. In order to improve the specificity of the sera, either the antibodies were affinity-purified on a synapsin I- Sepharose 4B column or the sera were passed through a human keratin-Sepharose 4B column to remove the contaminating anti- keratin antibodies, as described by Girault et al. (1989). The specificity of immunolabelling was determined by preadsorbing the antibodies with a 10- to 20-fold molar excess of purified synapsin I or synapsin I1 for 24 h at 4°C before immunoblotting.
Cell cultures Aplysia neurons were isolated and cultured as described by Schacher and Proshansky (1983). Helix neurons were cultured according to Ghirardi et al. (1996). After digestion with protease IX, ganglia were pinned on Sylgard-coated dishes and desheathed. The metacerebral giant cell (MGC) from the cerebral ganglion and the B2 neuron from the buccal ganglion were isolated from juvenile animals. The cells were transferred to poly-L-lysine-coated dishes and cultured in a medium containing Aplysia haemolymph and isotonic L15 medium. Cultures were maintained up to 3 days at 18°C with changes of medium on alternate days.
lmmunohistochemistry on intact ganglia Abdominal, pleural and pedal ganglia of Aplysia and cerebral ganglia of Helix were fixed for 2 h at room temperature with 4% para- formaldehyde in PBS (Helix) or PBS/30% sucrose (Aplysia) and rinsed several times in the same buffer. Sections (20 pm thick) were cut with a Kryostat 1720 (Emst Leitz, Wetzlar, Germany), rinsed in PBS containing 0.25% saponin and air-dried onto gelatine-coated glass slides. The tissue was dehydrated in ethanol, bleached with 0.6% H202 in methanol, saturated in 5% bovine serum albumid 0.25% saponin in PBS (blocking buffer) for 1 hat 37"C, and incubated overnight at 4°C with the appropriate dilution of the primary antibody in blocking buffer (G-177,1:400; G-94,1:200; G-423,1:200). Sections were then sequentially incubated for 1.5 h at room temperature with biotinylated anti-rabbit IgGs, ( 1 :300 dilution in blocking buffer) and

Synapsin-like molecules in Aplysia and Helix 2533
for 1 h at room temperature with the avidin-biotin peroxidase complex. Between the different steps, the sections were washed with several changes of PBS containing 0.25% saponin. The antibody complex was revealed using a solution of 0.01% 3,3'-diaminobenzid- ine tetrahydrochloride in 50 mM Tns-HC1 buffer (pH 7.5) containing 0.01% H202. Stained sections were washed in 50 mM Tris-HC1 (pH 7 3 , dehydrated in ethanol and mounted in Canada balsam. Sections were observed under a Zeiss microscope equipped with Nomarsky optics and photographed with Kodak Technical Pan 135 film.
h'o labelling was observed in controls in which the primary antibodies were either omitted or replaced with immunoadsorbed antisera.
lmmunocytochemistry on cell cultures Cultures were washed in PBS (Helix) or in PBS/30% sucrose (Aplysia) for 1 h at room temperature, followed by fixation with 4% paraformaldehyde in the same buffer for 30 min at room temperature. After bleaching with 0.6% H202 in methanol for 10 min at room temperature, cells were rinsed in PBS and treated with blocking buffer for 1 h at 37°C before the addition of the primary antibodies. Double-labelling experiments were performed by incubating the cultures overnight at 4°C with an affinity-purified rabbit pol yclonal anti-synapsin I (G-177, 1:250 dilution) in blocking buffer and with a mouse anti-5-HT monoclonal antibody (1 :25 dilution) in the same buffer. Cultures were rinsed with several changes of PBS containing 0.25% saponin and then incubated for 1.5 h at room temperature with either a rhodamine-conjugated rabbit anti-mouse IgG or a fluorescein- conjugated goat anti-rabbit IgG diluted 1 :40 in blocking buffer. After a brief wash in PBS, cultures were examined under phase-contrast or fluorescence optics with a Leitz Orthoplan microscope and photo- graphed with Kodak Ektachrome 400 film.
No labelling was observed in controls in which the primary antibody was either omitted or preadsorbed with the purified antigen.
Electrop h ysiology Stimulation and recording were done using intracellular electrodes (10-15 M a ) connected to an Axoclamp A2 in bridge mode. The presence of synaptic contacts between the MGC and B2 neurons in culture was tested by recording the excitatory postsynaptic potential (EPSP) in the B2 cell during stimulation of the MGC presynaptic neuron. The electrical activity was recorded on tape and sent to a chart recorder for measurement of the amplitude of the EPSP.
Morphometric measurements Varicosities were counted on large montages obtained from high- magnification micrographs. Presynaptic varicosities were defined as round structures (5-10 Fm diameter) stained for both synapsin I and 5-HT. All counts were done blind.
Miscellaneous techniques Protein determination was performed by the Bio-Rad protein assay (Bio-Rad Laboratories, Hercules. CA) or by the bicinchonic acid assay, using bovine serum albumin as standard. The following prestained molecular weight markers were used: a2-macroglobulin, p- galactosidase, fructose-6-phosphate kinase, pyruvate kinase, fumarase, lactic dehydrogenase and triose phosphate isomerase. Autoradio- graphy was performed using Kodak X-o-Mat films exposed for various times at -80°C with an intensifying screen.
Statistical analysis was carried out by analysis of variance and the Newman-Keuls t-test.
FIG, 1. Synapsin-like immunoreactivity in Aplysia and H e h nervous tissue. Proteins from rat cerebral cortex (K, 10 pg), Aplysia ganglia (A, 30 pg) and Helix ganglia (H, 30 pg) were resolved on 9% SDS-polyacrylamide gels, transferred to nitrocellulose and reacted with different antisera to mammalian synapsins depleted of the anti-keratin IgGs. Serum dilutions ranged between 1: 150 and 1:300. Immunoreactivity was revealed using '251-labelled anti- rabbit Fab and autoradiography. The electrophoretic mobility of the four rat synapsin (rSYN) isoforms and of the mollusc synapsin-like immunorcactive (SYNLI) peptides are shown on the left. Molecular m a x markers (in ma) are shown on the right.
Results
Presence of SLI in Aplysia and Helix nervous tissue Homogenates of Aplysia and Helix ganglia were analysed for synapsin imniunoreactivity by immunoblotting with a battery of pol yclonal antibodies directed against mammalian synapsin I or synapsin I and synapsin 11. Due to the sequence homology between synapsins I and 11, antibodies specific for synapsin I (e.g. G-94) are mostly directed against epitopes within the basic, collagenase-sensitive COOH-terminal region of synapsin I, whereas antibodies recognizing both synapsins (e.g. G-423) are likely to recognize domains in the shared NH2-terminal region. The other antibodies used (G-177 and G-178) recognize mostly synapsin I and to a lesser extent synapsin 11.
The majority of the antibodies gave a very strong signal for rat synapsins but also recognized with lower sensitivity immunoreactive species in extracts from both Aplysia and Helix ganglia. In all the experiments performed using different snail preparations, an immunoreactive band or a doublet of apparent molecular mass 58-63 kDa was constantly present, although its intensity varied depending on the antibody used (Fig. 1). The various antibodies also recognized Aplysia and Helix synapsin-like peptides to a different extent. While G-94 and G-177 antibodies recognized Aplysia and Helix proteins to a similar extent, G- 178 preferentially recognized the Helix protein and G-423 selectively recognized the Aplysia protein. With some preparations, immunoreactive bands of higher molecular mass (90 and 110 kDa) were sometimes observed, but their immuno- reactivity was always less intense than that of the 58-63 kDa species. The acid extracts of homogenates exhibited similar immunoreactivity with a slightly higher specific activity.
As expected from the scarce sequence conservation that is likely to occur bemeen mollusc and mammalian proteins, monoclonal antibodies against mammalian synapsins or polyclonal antibodies directed against short peptide sequences of mammalian synapsin I did not detect any significant signal in Aplysia or Helix nervous tissue (not shown).

2534 Synapsin-like molecules in Aplysia and Helix
FIG. 2. Specificity of synapsin-like immunoreactivity in mollusc nervous tissue. (Upper panel) Antibody preadsorption with mammalian synapsins. Acid extracts of rat cerebral cortex (R, 10 pg) and Aplysia ganglia (A, 30 pg) were subjected to SDS-PAGE and immunoblotting with a polyclonal antibody recognizing both synapsin I and synapsin I1 (G-423). Prior to immunoblotting, the antibody was incubated at 4°C for 24 h with buffer (control), a 20-fold molar excess of purified bovine synapsin I (+SYN Iab), or a 10-fold excess of purified recombinant rat synapsin IIa (+SYN IIa). In both cases, preadsorption of the antibody with mammalian synapsins abolished the synapsin-like immunoreactivity in the Aplysia acid extract. (Lower panel) Collagenase digestion of synapsin-like peptides in mollusc nervous tissue. Neutralized acid extracts of rat cerebral cortex (R, 30 pg), Aplysia ganglia (A, 100 pg) and Helix ganglia (H, 100 pg) were incubated with purified collagenase type I11 as described in Materials and methods. After digestion, proteins were separated by SDS-PAGE and transferred to nitrocellulose. In nervous system extracts, the incubation with collagenase strongly decreased or abolished the immunoreactivity but did not generate the collagenase- resistant fragment typical of mammalian synapsin I.
Specificity of SLI in Aplysia and Helix nervous tissue The specificity of the immunoreactivity observed in the snail nervous tissue was ascertained by the following experiments: (i) extraction of synapsin-like proteins at low pH, a property shared by several extrinsic membrane proteins including the synapsins; (ii) preadsorption of the antibodies with mammalian proteins prior to immunolabelling ; (iii) specific proteolysis with collagenase which specifically cleaves the COOH-terminal region of synapsin I at multiple sites.
The extractability of the synapsin-like peptides from both Aplysia and Helix nervous tissue was very similar to that of mammalian synapsins. The immunoreactive signal was fully preserved after extraction of the homogenate at pH 3.0 and was enriched when expressed with respect to total homogenate protein (not shown).
Preadsorption of the antibodies with mammalian synapsin I and I1 strongly decreased or virtually abolished SLI in acid extracts of Aplysia nervous tissue. Interestingly, the separate preadsorption of the (3-423 antiserum with the different synapsin isoforms revealed
that it contained IgGs recognizing synapsin I, synapsin I1 and the homologous regions of the two proteins. These latter antibodies, which are inactivated by preadsorption with both synapsins, are likely to be responsible for the recognition of the Aplysia synapsin-like peptides (Fig. 2, upper panel).
The digestion of mammalian synapsin I with collagenase form LI1 induced the formation of several peptides of molecular mass ranging between 86 and 50 kDa, originating from the progressive cleavage of multiple sites in the COOH-terminal region of synapsin I, which are missing in synapsin 11. Extensive digestion generated a collagenase- resistant SO kDa peptide corresponding to the NH2-terminal region of synapsin I (Ueda and Greengard, 1977; Fig. 2, lower panel). Under the same conditions, both the Aplysia and Helix synapsin-like peptides are digested, with the disappearance (Aplysin) or a marked decrease (Helix) of the immunoreactivity of the 58-63 kDa bands. However, no lower molecular mass peptides preserving SLI could be detected; this suggests either the presence of additional collagenase cleavage sites or the loss of the immunoreactive epitopes during digestion.
C$+/calmodulin- and CAMP-dependent phosphorylation of Aplysia synapsin-like peptides
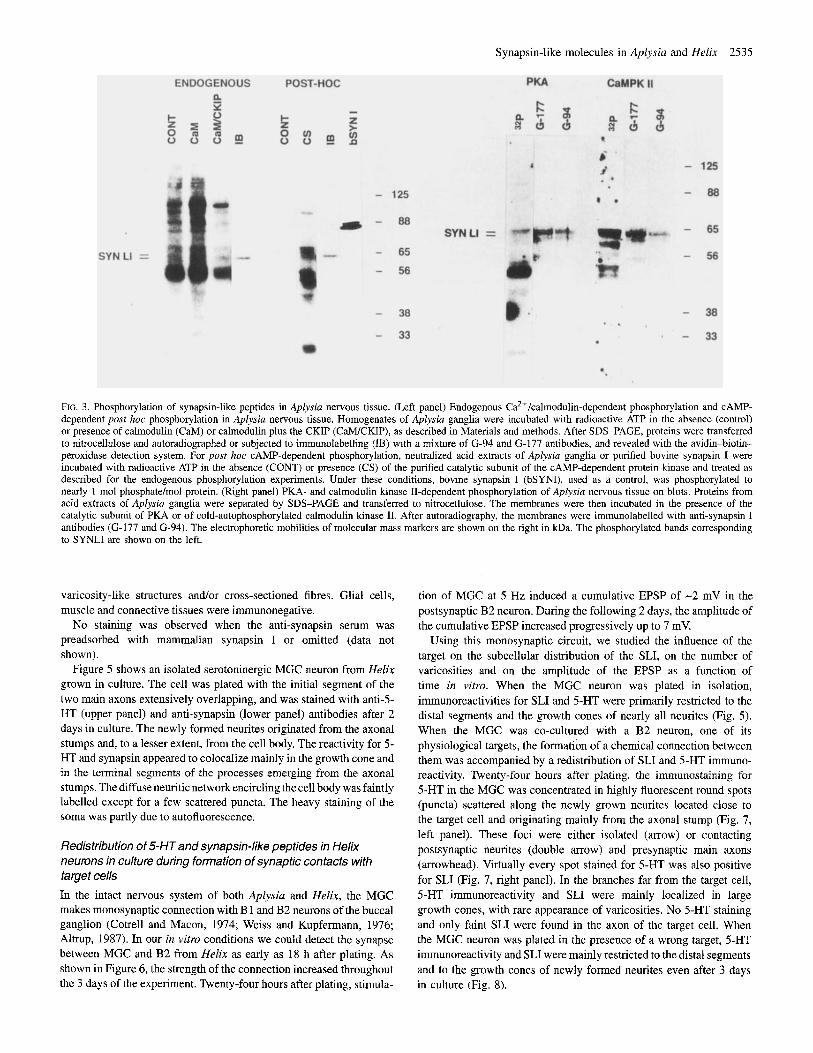
The presence of protein substrates for Ca2+/calmodulin- and CAMP- dependent phosphorylation in homogenates and acid extracts of Aplysia ganglia was studied using endogenous, post hoc and blot phosphorylation assays (Fig. 3). When freshly prepared homogenates of Aplysia ganglia were incubated in the presence of radioactive ATP and Ca*+/calmodulin, phosphate was incorporated into several protein bands, of which a 120 kDa band was the most prominent. Detectable phosphate incorporation was also present in a protein band comigrating with SLJ. Phosphorylation appeared to be mediated predominantly by a CaMPKII-like enzyme, since the addition of the specific peptide inhibitor CKIP markedly inhibited the phosphorylation of all proteins, including the Aplysin synapsin-like peptide (Fig. 3, left panel).
Phosphorylation of Aplysia acid extracts by the catalytic subunit of mammalian PKA induced marked incorporation of phosphate into bands of 25, 45-55, 58-63, 65 and 100 kDa. Among these phosphoproteins, most of which have been previously reported to be substrates for CAMP-dependent phosphorylation (Panchal et al., 1994), the 58-63 kDa doublet precisely overlapped with the SLI detected on the same blot (Fig. 3, left panel).
Phosphorylation of acid extracts of Aplysia ganglia was also tested after separation of the proteins by SDS-PAGE and Western blotting. Blotted protein bands corresponding to the SLI could be phosphorylated to a significant extent by both the purified catalytic subunit of PKA and the activated (autophosphorylated) CaMPKII, confirming the results obtained in the endogenous and post hoc phosphorylation assays (Fig. 3, right panel)
Cellular and subcellular distribution of synapsin-like peptides in Aplysia and Helix ganglia and in primary cultures of ganglion neurons
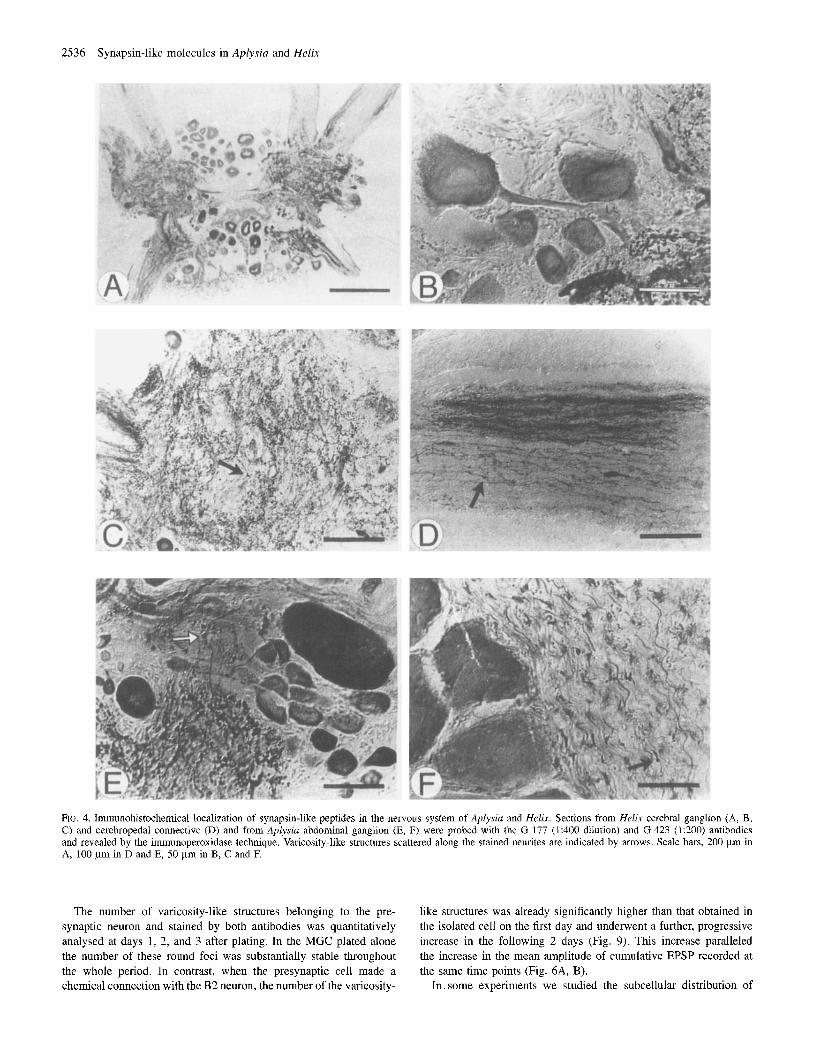
In ganglia of both Aplysia and Helix, SLI was associated with the soma of most of the neurons and with the neuropil. A low-magnification micrograph of the Helix cerebral ganglion shows that the cell body of most neurons and the neuropil were labelled, although the staining intensity varied from neuron to neuron (Fig. 4A). At a higher magnifica- tion (Fig. 4B, C), the cytoplasm of both the neuronal cell body and the axon hillock was immunopositive. Neurites running in the neuropil (Fig. 4D), in the connective (Fig. 4E) and in the ganglion sheath (Fig. 4F) showed strong SLI. Punctate immunostaining (arrow) was present along the stained neurites and in the neuropil (Fig. 4D, E), suggestive of
Synapsin-like molecules in Aplysiu and Helix 2535
RG. 3. Phosphorylation of synapsin-like peptides in Aplysia nervous tissue. (Left panel) Endogenous Ca2+/calmodulin-dependent phosphorylation and CAMP- dependent post hoc phosphorylation in Aplysia nervous tissue. Homogenates of Aplyssia ganglia were incubated with radioactive ATP in the absence (control) or presence of calmodulin (CaM) or calmodulin plus the CKIP (CMCKIP), as described in Materials and methods. After SDS-PAGE, proteins were transferred to nitrocellulose and autoradiographed or subjected to immunolabelling (IB) with a mixture of (3-94 and G-177 antibodies, and revealed with the avidin-biotin- peroxidase detection system. For post hoc CAMP-dependent phosphorylation. neutralized acid extracts of Aplysia ganglia or purified bovine synapsin I were incubated with radioactive ATP in the absence (CONT) or presence (CS) of the purified catalytic subunit of the CAMP-dependent protein kinase and treated as described for the endogenous phosphorylation experiments. Under these conditions, bovine synapsin I (bSYNI), used as a control, was phosphorylated to nearly 1 mol phosphate/mol protein. (Right panel) PKA- and calmodulin kinase II-dependent phosphorylation of Aplysia nervous tissue on blots. Proteins from acid extracts of Aplysia ganglia were separated by SDS-PAGE and transferred to nitrocellulose. The membranes were then incubated in the presence of the catalytic subunit of PKA or of cold-autophosphorylated calmodulin kinase 11. After autoradiography, the membranes were immunolabelled with anti-synapsin I antibodies (G-177 and (3-94). The electrophoretic mobilities of molecular mass markers are shown on the right in kDa. The phosphorylated bands corresponding to SYNLI are shown on the left.
varicosity-like structures and/or cross-sectioned fibres. Glial cells, muscle and connective tissues were immunonegative.
No staining was observed when the anti-synapsin serum was preadsorbed with mammalian synapsin I or omitted (data not shown).
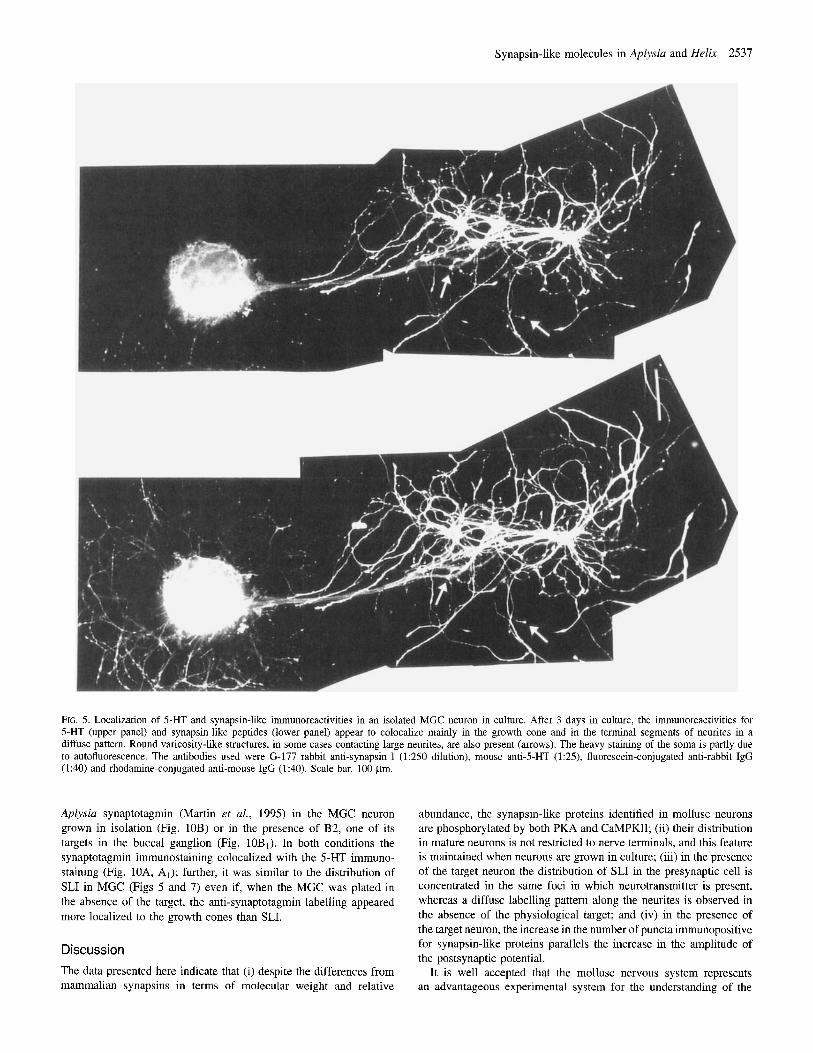
Figure 5 shows an isolated serotoninergic MGC neuron from Helix grown in culture. The cell was plated with the initial segment of the two main axons extensively overlapping, and was stained with anti-5- HT (upper panel) and anti-synapsin (lower panel) antibodies after 2 days in culture. The newly formed neurites originated from the axonal stumps and, to a lesser extent, from the cell body. The reactivity for 5- HT and synapsin appeared to colocalize mainly in the growth cone and in the terminal segments of the processes emerging from the axonal stumps. The diffuse neuritic network encircling the cell body was faintly labelled except for a few scattered puncta. The heavy staining of the soma was partly due to autofluorescence.
Redistribution of 5-HT and synapsin-like peptides in Helix neurons in culture during formation of synaptic contacts with target cells In the intact nervous system of both Aplysiu and Helix, the MGC makes monosynaptic connection with B 1 and B2 neurons of the buccal ganglion (Cotrell and Macon, 1974; Weiss and Kupfermann, 1976; Altrup, 1987). In our in vitro conditions we could detect the synapse between MGC and B2 from Helix as early as 18 h after plating. As shown in Figure 6, the strength of the connection increased throughout the 3 days of the experiment. Twenty-four hours after plating, stimula-
tion of MGC at 5 Hz induced a cumulative EPSP of -2 mV in the postsynaptic B2 neuron. During the following 2 days, the amplitude of the cumulative EPSP increased progressively up to 7 mV.
Using this monosynaptic circuit, we studied the influence of the target on the subcellular distribution of the SLI, on the number of varicosities and on the amplitude of the EPSP as a function of time in vitro. When the MGC neuron was plated in isolation, immunoreactivities for SLI and 5-HT were primarily restricted to the distal segments and the growth cones of nearly all neurites (Fig. 5). When the MGC was co-cultured with a B2 neuron, one of its physiological targets, the formation of a chemical connection between them was accompanied by a redistribution of SLI and 5-HT immuno- reactivity. Twenty-four hours after plating, the immunostaining for 5-HT in the MGC was concentrated in highly fluorescent round spots (puncta) scattered along the newly grown neurites located close to the target cell and originating mainly from the axonal stump (Fig. 7, left panel). These foci were either isolated (arrow) or contacting postsynaptic neurites (double arrow) and presynaptic main axons (arrowhead). Virtually every spot stained for 5-HT was also positive for SLI (Fig. 7, right panel). In the branches f a from the target cell, 5-HT immunoreactivity and SLI were mainly localized in large growth cones, with rare appearance of varicosities. No 5-HT staining and only faint SLI were found in the axon of the target cell. When the MGC neuron was plated in the presence of a wrong target, 5-HT immunoreactivity and SLI were mainly restricted to the distal segments and to the growth cones of newly formed neurites even after 3 days in culture (Fig. 8).
2536 Synapsin-like molecules in Aplysia and Helix
FIG. 4. Imunohistochemical localization of synapsin-like peptides in the nervous system of Aplysia and He/i.w. Sections from Helix cerebral ganglion (A, B, C) and cerebropedal connectivc (D) and from Aplysin abdominal ganglion (E, F) were probed with the G-177 (1:400 dilution) and G-423 (1:200) antibodies and revealed by the immunoperoxidase technique. Varicosity-like structures scattered along the stained neurites are indicated by arrows. Scale bars, 200 pm in A, 100 pm in D and E, 50 pm in B, C and F.
The number of varicosity-like structures belonging to the pre- synaptic neuron and stained by both antibodies was quantitatively analysed at days 1, 2, and 3 after plating. In the MGC plated alone the number of these round foci was substantially stable throughout the whole period. In contrast, when the presynaptic cell made a chemical connection with the B2 neuron, the number of the varicosity-
like structures was already significantly higher than that obtained in the isolated cell on the first day and underwent a further, progressive increase in the following 2 days (Fig. 9). This increase paralleled the increase in the mean amplitude of cumulative EPSP recorded at the same time points (Fig. 6A, €3).
In. some experiments we studied the subcellular distribution of
Synapsin-like molecules in Aplysia and Helix 2537
FIG. 5. Localization of 5-HT and synapsin-like immunoreactivities in an isolated MGC neuron in culture. After 3 days in culture, the immunoreactivities for 5-HT (upper panel) and synapsin-like peptides (lower panel) appear to colocalize mainly in the growth cone and in the terminal segments of neurites in a diffuse pattern. Round varicosity-like structures, in some cases contacting large neurites, are also present (arrows). The heavy staining of the soma is partly due to autofluorescence. The antibodies used were G-177 rabbit anti-synapsin I (1250 dilution), mouse anti-5-HT (1:25), fluorescein-conjugated anti-rabbit IgG (1:40) and rhodamine-conjugated anti-mouse IgG (1:40). Scale bar, 100 pm.
Aplysia synaptotagmin (Martin et al., 1995) in the MGC neuron grown in isolation (Fig. 10B) or in the presence of B2, one of its targets in the buccal ganglion (Fig. 10Bl). In both conditions the synaptotagmin immunostaining colocalized with the 5-HT immuno- staining (Fig. 10A, A,); further, it was similar to the distribution of SLI in MGC (Figs 5 and 7) even if, when the MGC was plated in the absence of the target, the anti-synaptotagmin labelling appeared more localized to the growth cones than SLI.
Discussion The data presented here indicate that (i) despite the differences from mammalian synapsins in terms of molecular weight and relative
abundance, the synapsin-like proteins identified in mollusc neurons are phosphorylated by both PKA and CaMPKII; (ii) their distribution in mature neurons is not restricted to nerve terminals, and this feature is maintained when neurons are grown in culture; (iii) in the presence of the target neuron the distribution of SLI in the presynaptic cell is concentrated in the same foci in which neurotransmitter is present, whcreas a diffuse labelling pattern along the neurites is observed in thc absence of the physiological target; and (iv) in the presence of the target neuron, the increase in the number of puncta immunopositive for synapsin-like proteins parallels the increase in the amplitude of the postsynaptic potential.
It is well accepted that the mollusc nervous system represents an advantageous experimental system for the understanding of the
2538 Synapsin-like molecules in Aplysia and Helix
A B2
-#----A
MGC
day I day 2 day 3
B
l o 1 T
0 1 2 3
Time (days)
FIG. 6. Chemical connection between MGC and B2 neurons in culture. (A) Five-second train of action potentials at 5 Hz induced in the MGC neuron causes long-lasting depolarization, partly due to the summation of single EPSPs in the 8 2 neuron hyperpolarized 30 mV below the resting potential. The amplitude of this depolarization increases from day 1 to day 3. Often, on day 3 , under the same pattern of stimulation the B2 membrane potential reaches the threshold for finng a spike. Vertical bar, 10 mV; horizontal bar, 10 s. (B) Time-course of the amplitude of B2 depolarization shows a significant increase with time ( P < 0.001, analysis of variance). Each point in the plot is the mean 2 SE of the data for five cultures.
molecular mechanisms of synaptic function and plasticity. In these processes, a pivotal role is played by synaptic vesicle-specific proteins and by phosphorylatioddephosphorylation processes. Therefore, the identification and characterization of the mollusc homologues of the mammalian proteins participating in the machinery of neuroexocytosis can provide useful tools for the study of the physiological mechanisms involved. Recently, two synaptic vesicle proteins, vesicle-associated membrane protedsynaptobrevin (Yamasaki et al., 1994) and synapto- tagmin (Martin et al., 1995), have been cloned from Aplysia nervous tissue. Synapsins, a family of synaptic vesicle-associated proteins which in the mammalian brain are major substrates for CAMP- and Ca’+/calmodulin-dependcnt protein kinases, have not yet been cloned in molluscs. Due to the importance of synapsins in synapse formation and plasticity, we have analysed some features of synapsin-like peptides identified in the nervous system of Aplysia punctata and Helix pomatia using antibodies generated against mammalian synapsins, and
have described their distribution in the mature nervous tissue and in developing ganglion neurons in culture.
Identification and phosphorylation of synapsin-like proteins in Aplysia and Helix nervous tissue
A variety of well characterized polyclonal antibodies directed against mammalian synapsins reproducibly recognized immunoreactive bands in the 58-63 kDa region in the nervous system of both Aplysia and Helix, although the intensity of the immunostaining was different in the two molluscs. In all cases, however, the signal was considerably less intense than in mammalian nervous tissue, as can be expected from the evolutionary distance between mammals and invertebrates. In addition to extractability in acid medium and to preadsorption of the antibodies with the purified mammalian antigen, the specificity of the labelling of synapsin-like molecules was demonstrated by the identification of two characteristic properties of mammalian synapsins: their sensitivity to collagenase and their ability to be phosphorylated by CAMP- and Ca2+/calmodulin-depeiident kinases. The immuno- reactive proteins identified in both Aplysia and Helix fulfilled all the above-mentioned criteria. Although we have been unable to immunoprecipitate the proteins with our antibodies (which also precipitate poorly mammalian synapsin I), the blot phosphorylation assay demonstrated that the immunoreactive bands are phosphorylat- able by PKA and CaMPKII, similarly to the mammalian synapsins.
These findings confirm and extend those previously reported by other authors in Aplysia califomica. Bongiovi et al. (1992) detected an array of immunoreactive peptides of various molecular weights, including a 50-55 kDa species which appeared to be collagenase- sensitive and phosphorylatable by the catalytic subunit of PKA. DeRiemer et al. (1984) and Panchal et al. (1994) described several protein substrates of Aplysia PKA, including a 58-63 kDa doublet which probably corresponds to our immunoreactive bands. In addition to these observations, we have demonstrated, using various antibodies, invertebrate preparations and phosphorylation assays, the presence of a more restricted, selective and reproducible pattern of immuno- reactivity in proteins which are phosphorylatable not only by PKA but also by endogenous and exogenous CaMPKTT. The synapsin-like peptides described here exhibit a molecular mass intermediate between those of mammalian synapsins I and IIb, suggesting that these proteins could represent ancestors which have not yet evolved towards the distinct isoforms present in the vertebrate brain.
Recently, a Drosophila gene encoding various synapsin isoforms exhibiting some degree of homology with mammalian synapsins has been cloned. These isoforms appear to have a molecular mass ranging from 70 000 to 143 000 kDa and are widely expressed in the nervous tissue of the fly (Kbdgges et al., 1996). Mollusc synapsin-like protcins appear to be much less abundant. This may be attributable to a lower degree of tissue expression, to a more restricted cellular distribution, or to poor recognition by the antibodies directed against mammalian synapsins. The results of the endogenous phosphorylation assays, together with the tissue distribution described below, suggest that the concentration of the proteins in the mollusc nervous system is indeed lower than that found in the mammalian brain.
Distribution of SLI in Aplysia and Helix nervous systems As already shown by Bongiovi et al. (1992) in Aplysia culifornica using a different set of antibodies, in Aplysia punctata and Helix pomatia the SLI is confined to the nervous system. Further, it does not have a ubiquitous distribution. For example, G-177 and G-423 antibodies do not recognize in Helix the somata of three giant buccal neurons (Bl, I32 and B3), whereas they notably label the cell body

Synapsin-like molecules in Aplysia and Helix 2539
FIG. 7. Localization of 5-HT and synapsin-like immunoreactivities in co-culture of Helix MGC and B2 neurons. Double immunostaining for 5-HT (left panel) and synapsin-like peptides (right panel) is shown. 5-HT and synapsin-like immunoreactivities colocalize in numerous and intensely labelled round spots scattered along the neurites. These varicosities were either isolated (arrows) or contacting postsynaptic neurites (double arrows) and presynaptic main axons (arrowheads). In the newly grown neurite emerging from the stump of the thinner and longer branch of the MGC and directed away from the target, the 5-HT and synapsin- like immunoreactivities were both diffusely located at the distal growth cones, with very few foci of varicosity-like strucmres. The faint SLI staining of the B2 neuron is probably attributable to the fact that this cell was plated without the axonal stump. Immunocytochemistry was performed 3 days after plating. For further details see legend to Figure 5. Scale bar, 100 pm.
of B4. This aspect is somewhat different in Aplysiu, where all these neurons were moderately labelled. The soma of the giant serotoninergic cell MGC is heavily stained in both species. In general, our results in Aplysia are in line with the findings of Bongiovi et al. (1992): the most intensely stained somata are those of the small neurons, while giant neurons are very faintly stained or not stained at all. In Helix, however, some giant neurons (e.g. Rpl, Lpl, Rp2, Lp2 and other large cells in the parietal and visceral ganglia) are very intensely labelled. The larger number of labelled giant neurons in Helix could be explained by considering that they are smaller than their counterparts from Aplysia. This could suggest a sort of ‘dilution effect’ in Aplysiu neurons by which the concentration of synapsins attained in the soma does not reach the detection level of the antibodies. In culture, Helix B2 neurons and the unidentified non- target neurons show negligible staining by the same antibodies as those used in the immunohistochemistry experiments, suggesting a real lack of staining.
Distribution of SLI in cultured neurons Invertebrate neurons are apparently non-polarized. In higher invertebrates, like Aplysiu and Helix, a typical neuron gives rise to one or two processes that have the properties of an axon. Within the neuropil these processes give off branches that are postsynaptic and can be considered as dendrites from a functional point of view (Bullock and Homdge, 1955; Coggeshall, 1967; Bailey et al., 1981; Bailey and Chen, 1988). This implies that, although the neuronal
branches are not organized in axonal and dendritic domains, the presynaptic and postsynaptic functions must be segregated, most probably relying on a differential distribution of membrane receptors and channels. When the MGC is cultured alone, both SLI and 5-HT immunoreactivity are uniformly distributed in the cell body, in the distal part of the newly formed branches originating from the stump and, although much less intensely, in the major axons and in the fine processes emerging from the cell body. This ubiquitous distribution of SLI and its colocalization with the neurotransmitter seems to confirm the lack of polarity of the invertebrate neurons, in which any process may be competent to become a presynaptic structure (Craig and Banker, 1994).
Distribution of SLI and 5-HT in the absence of the appropriate target cell Although Helix and Aplysia neurons appear to be different from vertebrate neurons in the location of the synapsin-like peptides in the processes, they are similar in the way the presence of the target cell influences SLI distribution within the branches. In the MGC cultured alone or co-cultured with a non-target neuron, immunostaining for both synapsin-like peptides and 5-HT is mostly concentrated in the distal part of the neurites and in the growth cones. Small labelled varicosity-like structures are present but in a much lesser amount compared with that observed when the presynaptic neuron contacts the target. Also, in Aplysiu sensory neurons cultured alone or in the presence of a non-target cell only a few varicosities are detected, in
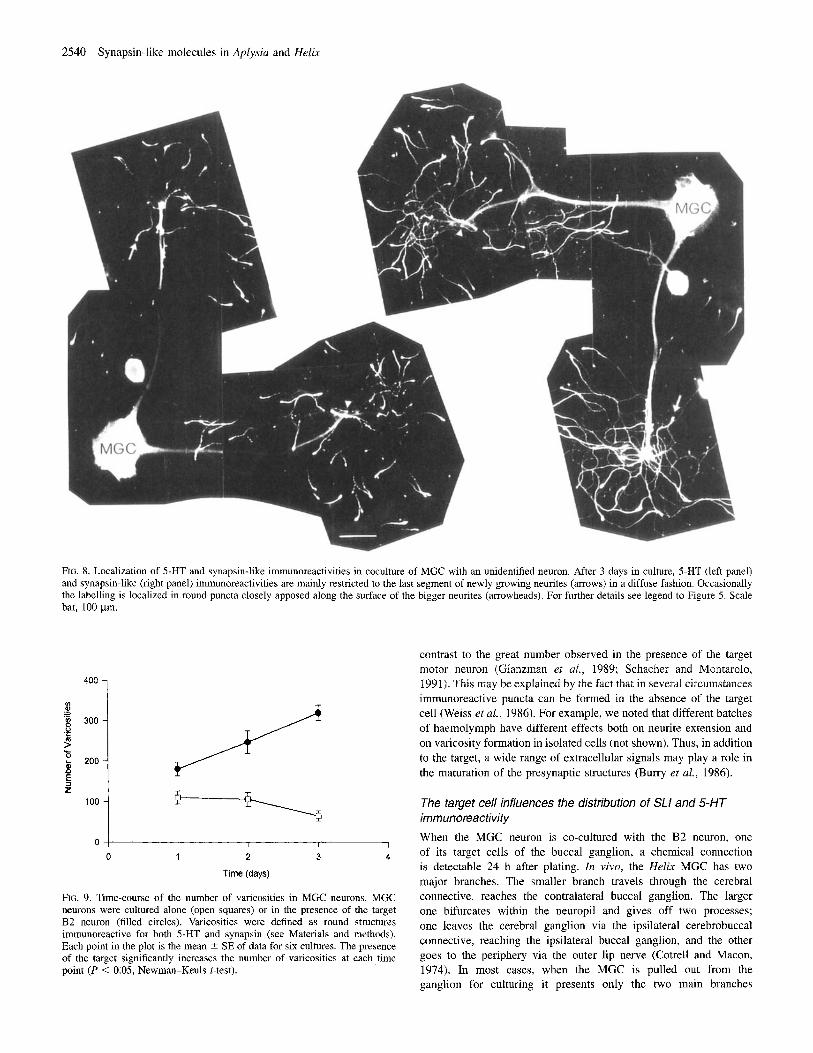
2540 Synapsin-like molecules in Aplysia and Helix
FIG. 8. LocaliLation of 5-HT and synapsin-like immunoreactivities in coculture of MGC with an unidentified neuron. After 3 days in culture, S-HT (left panel) and synapsin-like (right panel) immunoreactivities are mainly restricted 10 the last segment of newly growing neurites (arrows) in a diffuse fashion. Occaaionally the labelling is localized in round puncta closely apposed along the surface of the bigger neurites (arrowheads). For further details see legend to Figure 5. Scale bar, 100 pm.
L
400 1
200 ̂I loo 1 0 1 I I I I
0 1 2 3 4
Time (days)
FIG. 9. Time-course of the number of varicosities in MGC neurons. MGC neurons were cultured alone (open squares) or in the presence of the target B2 neuron (filled circles). Varicosities were defined as round structures immunoreactive for both 5-HT and synapsin (see Materials and methods). Each point in the plot is the mean 2 SE of data for six cultures. The presence of the target significantly increases the number of varicosities at each time point (P < 0.05, Newmm-Keuls t-test).
contrast to the great number observed in the presence of the target motor neuron (Glanzman et aL, 1989; Schacher and Montarolo, 1991). This may be explained by the fact that in several circumstances immunoreactive puncta can be formed in the absence of the target cell (Weiss e f aL, 1986). For example, we noted that different batches of haemolymph have different effects both on neurite extension and on varicosity formation in isolated cells (not shown). Thus, in addition to the target, a wide range of extracellular signals may play a role in the maturation of the presynaptic structures (Burry et al., 1986).
The target cell influences the distribution of SLI and 5-HT immunoreactivity
When the MGC neuron is co-cultured with the B2 neuron, one of its target cells of the buccal ganglion, a chemical connection is detectable 24 h after plating. In vivo, the Helix MGC has two major branches. The smaller branch travels through the cerebral connective. reaches the contralateral buccal ganglion. The larger one bifurcates within the neuropil and gives off two processes; one leaves the cerebral ganglion via the ipsilateral cerebrobuccal connective, reaching the ipsilateral buccal ganglion, and the other goes to the periphery via the outer lip nerve (Cotrell and Macon, 1974). In most cases, when the MGC is pulled out from the ganglion for culturing it presents only the two main branches
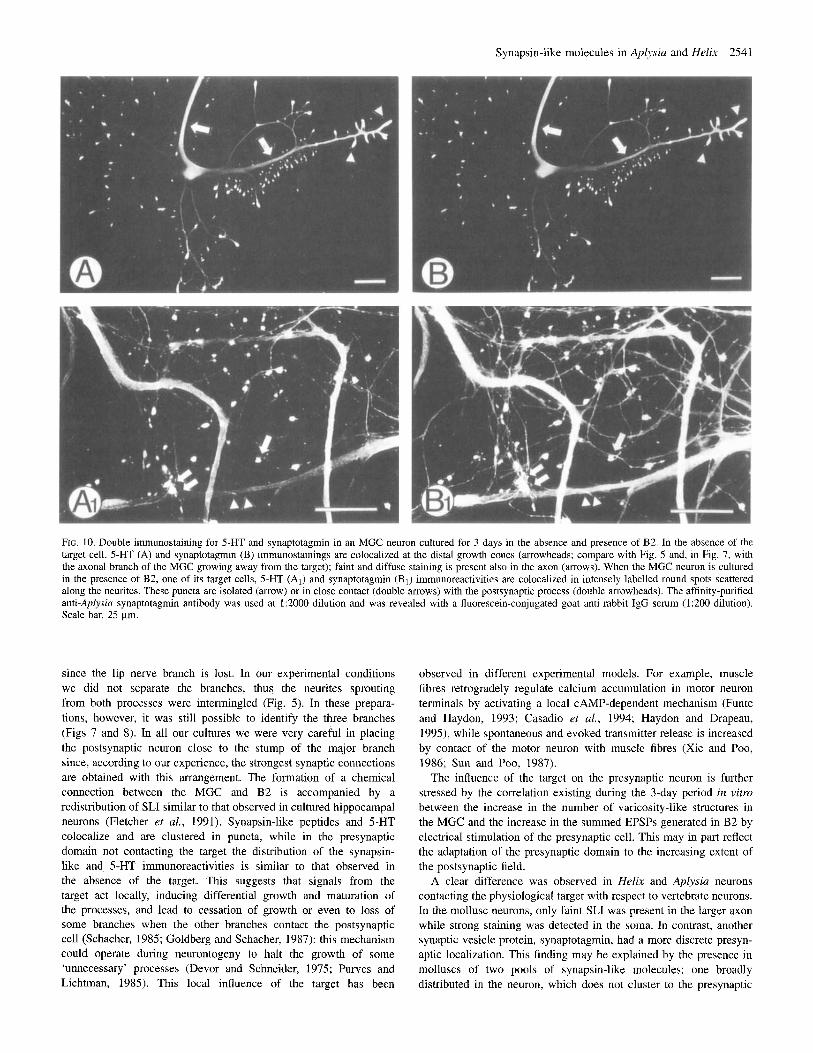
Synapsin-like molecules in Aplysia and Helix 2541
FIG. 10. Double immunostaining for 5-HT and synaptotagmin in an MGC neuron cultured for 3 days in the absence and presence of B2. In the absence of the target cell, 5-HT (A) and spnaptotagmn (B) immunostainings are colocalized at the distal growth cones (arrowheads; compare with Fig. 5 and, in Fig. 7, with the axonal branch of the MGC growing away from the target); faint and diffuse staining is present also in the axon (arrows). When the MGC neuron is cultured in the presence of B2, one of its target cells, 5-HT (AJ and synaptotagmin (B,) immunoreactivities are colocalized in intensely labelled round spots scattered along the neurites. These puncta are isolated (arrow) or in close contact (double arrows) with the postsynaptic process (double arrowheads). The affinity-purified anti-Aplysia synaptotagmin antibody was used at 1:2000 dilution and was revealed with a fluorescein-conjugated goat anti-rabbit IgG serum (1:200 dilution). Scale bar, 25 pm.
since the lip nerve branch is lost. In our experimental conditions we did not separate the branches, thus the neurites sprouting from both processes were intermingled (Fig. 5). In these prepara- tions, however, it was still possible to identify the three branches (Figs 7 and 8). In all our cultures we were very careful in placing the postsynaptic neuron close to the stump of the major branch since, according to our experience, the strongest synaptic connections are obtained with this arrangement. The formation of a chemical connection between the MGC and B2 is accompanied by a redistribution of SLI similar to that observed in cultured hippocampal neurons (Fletcher et al., 1991). Synapsin-like peptides and 5-HT colocalize and are clustered in puncta, while in the presynaptic domain not contacting the target the distribution of the synapsin- like and. 5-HT immunoreactivities is similar to that observed in the absence of the target. This suggests that signals from the target act locally, inducing differential growth and maturation of the processes, and lead to cessation of growth or even to loss of some branches when the other branches contact the postsynaptic cell (Schacher, 1985; Goldberg and Schacher, 1987); this mechanism could operate during neurontogeny to halt the growth of some ‘unnecessary’ processes (Devor and Schneider, 1975; Purves and Lichtman, 1985). This local influence of the target has been
observed in different expcrimental modcls. For example, muscle fibres retrogradely regulate calcium accumulation in motor neuron terminals by activating a local CAMP-dependent mechanism (Funte and Haydon, 1993; Casadio et al., 1994; Haydon and Drapeau, 1995), while spontaneous and evoked transmitter release is increased by contact of the motor neuron with muscle fibres (Xie and Poo, 1986; Sun and Poo, 1987).
The influence of the target on the presynaptic neuron is further stressed by the correlation existing during the 3-day period in vilro between the increase in the number ol’ varicosity-like structures in the MGC and the increase in the summed EPSPs generated in B2 by electrical stimulation of the presynaptic cell. This may in part reflect the adaptation of the presynaptic domain to the increasing extent of the postsynaptic field.
A clear difference was observed in Helix and Aplysia neurons contacting the physiological target with respect to vertebrate neurons. In the mollusc neurons, only faint SLI was present in the larger axon while strong staining was detected in the soma. In contrast, another synaptic vesicle protein, synaptotagmin, had a more discrete presyn- aptic localization. This finding may be explained by the presence in molluscs of two pools of synapsin-like molecules: one broadly distributed in the neuron, which does not cluster to the presynaptic

2542 Synapsin-like molecules in Aplysia and Helix
specializations under the influence of the target, and the other localized in the presynaptic region, which is under the influence of the interaction between pre- and postsynaptic cell. Synapsin-like proteins belonging to the latter pool are likely to be associated with synaptic vesicles or with their precursors since they codistribute with the neurotransmitter and their expression correlates with axon growth, targct recognition, formation of synaptic contacts and synaptic strengthening. The exclusive nerve terminal localization of mamma- lian synapsins may be the result of evolution from a relatively ubiquitous cytoskeleton-organizing protein, such as that predomi- nantly found in the mollusc nervous system, into a purely synaptic vesicle-associated protein. A similar shift in subcellular distribution is also observed during the ontogeny of mammalian neurons (Fletcher et al., 1991). However, some degree of non-specific labelling cannot be ruled out since we used antibodies against vertebrate synapsins. The more restricted distribution of axonal 5-HT immunoreactivity supports this possibility.
By using an anti-Aplysia synaptotagmin antibody we have shown that, like synapsins, this integral synaptic vesicle-specific protein colocalizes with 5-HT, the neurotransmitter of the MGC neuron; furthermore, its distribution is strongly influenced by the presence of the target (Fig. 10). Using the same antibody, Martin et al. (1995) demonstrated that in Aplysia sensory neurons co-cultured with the target motor neuron, synaptotagmin immunoreactivity is localized in the presynaptic cell in punch similar to those observed in the MGC when cultured in the presence of the B2 neuron. These authors also reported that in their sensory-to-motor neuron preparation the staining pattern for synaptotagmin was indistinguishable from that obtained with anti-Aplysia synaptobrevin antibodies. All these data are in good agreement with our finding showing the presence of a synapsin-like immunoreactivity in axonal structures corresponding to synaptic contacts.
In conclusion, using a variety of polyclonal antibodies we have demonstrated the presence in the nervous systems of Aplysia and Helix of proteins sharing some properties with the mammalian synapsins, and have described their cellular and subcellular distribu- tions. Biochemical and cytochemical analyses indicate that these proteins are expressed in low amounts in the nervous system of these two molluscs and have a subcellular distribution wider than that observed in the mammalian nervous system. The localization of SLI at the tip of growth cones in developing neurons and in varicosities after the formation of synapses with the physiological target suggests that the mollusc synapsin homologues may play a role in cytoskeletal organization, neurite elongation and synapse formation similar to that proposed for the mammalian synapsins (Han et al., 1991; Ferreira et al., 1994; Chin et al., 1995).
Acknowledgements This work was supported by grants from Minister0 della Universith e della kcerca Scientifica e Tecnologica and Consiglio Nazionale delle Ricerche to E B., P. G. M. and E V. The financial support of Telethon-Italy (grant 753) and of Progetto Nazionale Sclerosi Multipla, Istituto Superiore di Sanita to F. B., and that of the Galileo programme (ami27-95WGS) and of Progetto Finalizzato FATMA, Consiglio Nazionale delle Ricerche to F. V. is also acknowledged. F. 0. is presently granted a fellowship from Anna Villa Rusconi Foundation (Varese, Italy). The authors are very grateful to Dr Rosalia Ferro for her help in culturing Aplysia and Helix neurons, to Dr Laura Morando for the synaptotagmin immunocytochemistry, and to Mr Gennara Diana for his skilful technical assistance and for the preparation of the photographs. Aplysia punctata were housed at the Istituto di Biologia Marina (Director, Prof. G. Marano). Helix potnutia at different developmental stages were provided by Mr Natale Griglione. The anti-Aplysia synaptotagmin antibody was a generous gift of Dr Kelsey C. Martin (Center for Neurobiology and Behavior, Columbia University College of Physicians and Surgeons, New York).
Abbreviations 5-HT CaMPKII CKIP EPSP MGC PKA SDS-PAGE SLI
serotonin Ca*+/calmodulin-dependent protein kinase I1 CaMPKIl inhibitory peptide excitatory postsynaptic potential metacerebral giant cell CAMP-dependent protein kinase sodium dodecyl sulphate-polyacrylatnide gel electrophoresis synapsin-like immunoreactivity
References
Altrup, U. (1987) Inputs and outputs of giant neurons B1 and B2 in the buccal ganglion of Helixpomatia: an electrophysiological and morphological study. Brain Res., 414, 271-284.
Bghler, M. and Greengard, P. (1987) Synapsin I bundles F-actin in a phosphorylation-dependent manner. Nature, 326, 704-707.
Bailey, C. H. and Chen, M. (1988) Long-term memory in Aplysia modulates the total varicosities of single identified sensory neurons. Proc. Nut1 Acad. Sci. USA, 85, 2373-2377.
Bailey, C. H., Kandel, E. R. and Chen, M. (1981) Active zone at Aplysia synapses: organization of presynaptic dense projections. J. Neurophysiol., 46, 356-367.
Bajjalieh, S. M. and Scheller, R. H. (1995) The biochemistry of neurotransmitter secretion. J. Biol. Chem., 270, 1971-1974.
Benfenati, F., Valtorta, E, Chieregatti, E. and Greengard, P. (1992) Interaction of free and synaptic vesicle-bound synapsin I with F-actin. Neuron, 8,
Benfenati, F., Valtorta, F., Rossi, M. C., Onofri, F., Sihra T. and Greengard, P. (1993) Interactions of synapsin I with phospholipids: possible role in synaptic vesicle clustering and in the maintenance of bilayer structures. J. Cell B i d , 123, 1845-1855.
Bongiovi, M. E., Ambron, R. T. and Silverman, A.-J. (1992) The morphological localization and biochemical characterization of a synapsin I-like antigen in the nervous system of Aplysia californica. J. Neurosci. Res., 32, 395406.
Bullock, T. H. and Horridge, G. A. (1955) Structure and Function in the Nervous System of Invertebrates. Freeman, San Francisco.
Burry, R. W., Raymond, H. H. and Mattew, W. D. (1986) Presynaptic elements formed on polylysine-coated beads contain synaptic vesicle antigens. J. Neurocytol., 15, 409419.
Casadio, A,, Levi, R., Morando, L., Naretto, G. and Montarolo, P. G. (1994,) Presynaptic calcium levels at the chemical connection between molluscan neurons in vitro. Soc. Neurosci. Absa , 20, 1087.
Ceccaldi, P.-E., Grohovaz, E , Benfenati, E, Chieregatti, E., Greengard, P. and Valtorta, F. (1995). Dephosphorylated synapsin I anchors synaptic vesicles to actin cytoskeleton: an analysis by videomicroscopy. J. Cell Biol., 128,
Chin, L.-S., Li, L., Ferreira, A., Kosik, K. S. and Greengard, P. (1995) Impairment of axonal development and of synaptogenesis in hippocampal neurons of synapsin I-deficient mice. Proc. Nafl Acad. Sci. USA, 92, 9230-9234.
Cibelli, G., Benfenati, E, Ghirardi, M., Vitiello, F. and Montarolo, P. G. (1994) Synapsin-like immunoreactivity in invertebrate neurons in vitm: redistribution following the establishment of synaptic contacts. SOC. Neurosci. Abstx, 20, 450.
Coggeshall, R. E. (1967) A light and electron microscopy study of the abdominal ganglion of Aplysia califarnica. J. Neurophysiol., 30, 1263-1287.
Cotrell, G. A. and Macon, J. B. (1974) Synaptic connections of two symmetrically placed giant serotonin-containing neurons. J . Physiol. (Lond. ), 236,435464.
Craig, A. M. and Banker, G. (1994) Neuronal polarity. Annu. Rev. Neurosci., 17, 267-310.
De Camilli, P., Benfenati, F., Valtorta, E and Greengard, P. (1990) The synapsins. Annu. Rev. Cell B i d , 6, 433460.
DeRiemer, S. A., Kaczmarek, L. K., Lai, Y., McCuinness, T. L. and Greengard, P. (1984) Calciudcalmodulin-dependent protein phosphorylation in the nervous system of Aplysia. J. Neurosci., 4, 1618-1625.
Devor, M. and Schneider, G. (1975) Neuroanatomical plasticity: the principle of conservation of total axonal arborization. In Vidal, E and Jannerod, M. (eds), Aspects of Neural Plasticity, Vol. 43. INSERM, Pans, pp. 191-201,
Ferreira, A,, Kosik, K. S., Greengard, P. and Han, H. Q. (1994) Aberrant neurites and synaptic vesicle protein deficiency in synapsin 11-depleted neurons. Science, 264, 977-979.
377-386.
905-912.

Synapsin-like molecules in Aplysia and Helix 2543
Montarolo, P. G., Kandel, E. R. and Schacher, S. (1988) Long-term heterosynaptic inhibition in Aplysia. Nature, 333, 171-174.
Nestler, E. J. and Greengard, P. (1980) Dopamine and depolarizing agents regulate the state of phosphorylation of protein I in the mammalian superior cervical sympathetic ganglion. Proc. Nail Acad. Sci. USA, 77, 7479-7483.
Nichols, R. A,, Clulcote, T. J. , Czernik, A. J. and Greengard, P. (1992) Synapsin I regulates glutamate release from rat brain synaptosomes. J. Neurochem., 58, 783-785.
Panchal, R. G., Cheley, S. and Bayley, H. (1994) Differential phosphorylation of neuronal substrates by catalytic subunits of Aplysia CAMP-dependent protein kinase with alternativc N termini. J. Biol. Chem., 269,23722-23730.
Pieribone, V. A., Shupliakov, O., Brodin, L., Hilfiker-Rothenfluh, S., Czernik, A. J . and Greengard, P. (1995) Distinct pools of synaptic vesicles in neurotransmitter release. Nature, 375, 493497.
Purves, D. and Lichtman, J. W. (1985) Principles of Neural Development. Sinauer Associates, Sunderland, MA.
Rayport, S. G. and Schacher, S. (1986) Synaptic plasticity in viiro: cell culture of identified Aplysia neurons mediating short-term habituation and sensitization. J. Neurosci., 6, 759-763.
Rosahl, T. W., Spillane, D., Missler, M., Herz, J., Selig, D. K., Wolff, J. R., Hammer, R. E., Malenka, R. C. and Sudhof, T. C. (1995) Essential functions of synapsins I and 11 in synaptic vesicle regulation. Nature, 375, 488-493.
Schacher, S. and Proshansky, E. (1983) Neurite regeneration by Aplysia neurons in dissociated cell culture: modulation by Aplysia hemolymph and the presence of the initial segment. J. Neurosci., 3, 2403-2413.
Schacher, S. (1985) Differential synapse formation and neurite outgrowth at two branches of the metacerebra] cell of Aplysia in dissociated cell cullure. J. Neurosci., 5, 2028-2034.
Schacher, S. and Montarolo, P. G. (1991) Target-dependent structural changes in sensory neurons of Aplysia accompany long-term heterosynaptic inhibition. Neuron, 6, 679-690.
Schaeffer, E., Alder, J., Greengard, P. and Poo, M.-M. (1994) Synapsin IIa accelerates functional development of neuromuscular synapses. Proc. Nut1 Acad. Sci. USA, 91, 3882-3886.
Siow, C., Chilcote, T. J., Benfenati, F., Greengard, P. and Thiel, G. (1992) Synapsin IIa: expression in insect cells, purification and characterization. Biochemistry, 31, 42684275.
Sun, Y.-A. and Poo. M.-M. (1987) Evoked release of acetylcholine from growing embryonic neurons. Proc. Nafl Acud. Sci. USA, 84, 2 5 4 2 5 4 4 .
Towbin, H., Staehelin, T. and Gordon, J. (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedures and some applications. Proc. Nut1 Acad. Sci. USA, 76, 4350-4354.
Ueda, T. and Greengard, P. (1977) Adenosine 3’:5’-monophosphate-regulated phosphoprotein system of neuronal membrane. J. Biol. Chem., 252, 5155- 5163.
Valtorta, F. and Benfenati, F. (1995) Membrane trafficking in nerve terminals. Adv. Pharmacol., 32, 505-557.
Valtorta, F., Schiebler, W., Jahn, R., Ceccarelli, B. and Grcengard, P. (1986) A solid-phase assay for the phosphorylation of proteins blotted on nitrocellulose membrane filters. Anal. Biochem., 158, 130-137.
Valtorta, F., Iezzi, N., Benfenati, F., Lu, B., Poo, M.-M. and Greengard, P. (1995) Accelerated structural maturation induced by synapsin I at developing neurornuscular synapses of Xenopus laevis. Eur: J. Neurosci., 7, 261-270.
Volknandt, W., Naito, S., Ueda, T. and Zimmermann, H. (1987) Synapsin I is associated with cholinergic nerve terminals in the electric organs of Torpedo, Elecirophorus and Malapterurus and copurifies with Torpedo synaptic vesicles. J. Neurochem., 49, 342-347.
Weiss, K. R. and Kupfermann, I. (1976) Homology of the giant serotoninergic neurons (metacerebid cells) in Aplysia and pulmonate molluscs. Brain Res., 117, 3 3 4 9 .
Weiss, K. R., Pin, J.-P., Sebben, M., Kemp, D. E., Sladeczek, F., Gabrion, J. and Bodkaert, J. (1986) Synaptogeuesis of cultured striatal neurons in serum free medium: a morphological and biochemical study. Proc. Nut1 Acad. Sci. USA, 83, 2238-2242.
Xie, Z.-P. and Poo, M.-M. (1986) Initial events in the formation of neuromuscular synapse: rapid induction of acetylcholine release from embryonic neurons. Proc. Nut1 Acad. Sci. LISA, 83, 7068-7073.
Yamasaki, S . , Hu, Y., Binz, T., Kalkuhl, A., Kurazono, H., Tamura, T., Jahn, R., Kandel, E. R. and Niemann, H. (1994) Synaptobrevinlvesicle-associated membrane protein (VAMP) of Aplysia californica: structure and proteolysis by tetanus toxin and botulinal neurotoxins type D and F. Proc. Nail Acad. Sci. USA, 91, 46884692.
Fletcher, T. L., Cameron, P., De Camilli, P. and Banker, G. (1991) The distribution of synapsin I and synaptophysin in hppocampal neurons developing in culture. J. Neurosci., 11, 1617-1626.
Funte, L. R. and Haydon, P. G. (1993) Synaptic target contact enhances presynaptic calcium influx by activating CAMP-dependent protein lunase during synaptogenesis. Neuron, 10, 1069-1078.
Ghirardi, M., Casadio, A., Santarelli, L. and Montarolo, P. G. (1996) Aplysiu hemolymph promotes neurite outgrowth and synaptogenesis of identified Helix neurons in cell culture. Inv. Neurosci., 2, 4149.
Girault, J.-A., Gorelik, F. S . and Greengard, P. (1989) Improving the quality of immunoblots by chromatography of polyclonal antisera on keratin affinity columns. Anal. Biochem., 182, 193-198.
Glanzman, D. L., Kandel, E. R. and Schacher, S. (1989) Identified target motor neuron regulates neurite outgrowth and synapse formation of Aplysia sensory neurons in vitro. Neuron, 3, 441450.
Goelz, S. E., Nestler, E. J. and Greengard, P. (1985) Phylogenetic survey of proteins related to synapsin I and biochemical analysis of four such proteins from fish brain. J. Neurochem., 45, 63-72.
Goldberg, D. J. and Schacher, S. (1987) Differential growth of the branches of a regenerating bifurcate axon is associated with differential axonal transport of organelles. Dev. BioL, 124, 35-40.
Greengard, P., Valtorta, F., Czernik, A. J. and Benfenati, F. (1993) Synaptic vesicle phosphoproteins and regulation of synaptic function. Science, 259, 780-785.
Hackett, J. T., Cochran, S. L., Greenfield, J., Brosius, D. C. and Ueda, T. (1 990) Synapsin 1 injected presynaptically into goldfish Mauthner axons reduces quanta1 synaptic transmission. J. Neurophysiol., 63, 701-706.
Han, H. Q., Nichols, R. A., Rubin, M. R., B2hIer, M. and Greengard, P. (1991) Induction of formation of presynaptic terminals in neuroblastoma cclls by synapsin IIb. Nature, 349, 697-700.
Haydon, P. G., McCobh, D. P. and Kater, S. B. (1984) Serotonin selectively inhibits growth cone motility and synaptogenesis of specific identified neurons. Science, 226, 561-564.
Haydon, P. G. and Drapeau, P. (1995) From contact to connection: early events during synaptogenesis. Trends Neurosci., 18, 196201.
Kaczmarek, L. K., Jennings, K. R., Strumwasser, F., Nairn, A. C., Walter, U., Wilson, F. D. and Greengard, P. (1980) Microinjection of catalytic subunit of cyclic AMP-dependent protein kinase enhances calcium action potentials of bag cell neurons in cell culture. Proc. Nut1 Acad. Sci. USA, 77,
Klagges, B. R., Heimbeck, G., Godenschwege, T. A., Hofbauer, A,, Pflugfelder, G. O., Reifegerste, R., Reisch, D., Schaupp, M., Buchner, S. and Buchner, E. (1996) Invertebrate synapsins: a single gene codes for several isoforms in Drosophila. J. Neurosci., 16, 3154-3165.
Laemmli, U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227, 680-685.
Li, L., Chin, L.-S., Shupliakov, O., Brodin, L., Sihra, T. S., Hvalby, 0. E., Jensen, V., Zheng, D., McNamara, J. O., Greengard, P. and Andersen P. (1995) Impairment of synaptic vesicle clustering and of synaptic transmission, and increased seizure propensity, in synapsin I-deficient mice. Proc. Nut1 Acud. Sci. USA, 92, 923559239,
LlinBs, R. R., McGuinness, T. L., Leonard, C. S., Sugimori, M. and Greengard, P. (1985) Intraterminal injection of synapsin I or calciudcalmodulin dependent protein kinase 11 alters neurotransmitter release at the squid giant synapse. Proc. Nut1 Acad. Sci. USA, 82, 3035-3039.
Llinbs, R., Gruner, J. A., Sugimori, M., McGuinness, T. L. and Greengard, P. (1991) Regulation by synapsin I and Ca*+/calmodulin-dependent protein kinase I1 of transmitter release in squid giant synapse. J. Physiol. (Load.), 436, 257-282.
Lu, B., Greengard, P. and Poo, M.-M. (1992) Synapsin I promotes functional maturation of developing neuromuscular synapses. Neuron, 9, 759-768.
Martin, K. C., Hu, Y., Armitage, B. A,, Siegelbaum, S. A., Kandel, E. R. and Kaang, B.-K. (1995) Evidence for synaptotagmin as an inhibitory clamp on synaptic vesicle release in Aplysia neurons. Proc. Nail Acad. Sci. USA, 92, 11307-1 13 11.
McGuinness, T. L., Lai, Y. and Greengard, P. (1985) Ca2+/calmodulin- dependent protein kinase I1 isozymic forms from rat forebrain and cerebellum. J. B i d . Chern., 260, 1696-1704.
Montarolo, P. G.. Goelet, P., Castellucci, V. F., Morgan, J., Schacher, S. and Kandel, E. R. (1986) A critical time window for macromolecular synthesis on long-term heterosynaptic facilitation. Science, 234, 1249-1254.
7487-7491.




















