A diagnostic and surgical approach to masses of the buccal ...
Upload
independentCategory
view
1download
0
ORIGINAL PAPER
A population of pedal–buccal projection neurons associatedwith appetitive components of Aplysia feeding behavior
Received: 3 May 2002 / Revised: 5 September 2002 / Accepted: 11 January 2003 / Published online: 18 February 2003� Springer-Verlag 2003
Abstract Backfills of the cerebral-buccal connective(CBC) of Aplysia californica revealed a cluster of five toseven pedal-buccal projection neurons in the anterolat-eral quadrant of the ventral surface of each pedal gan-glion. Intra- and extracellular recordings showed thatthe pedal-buccal projection neurons shared commonelectrophysiological properties and synaptic inputs.However, they exhibited considerable heterogeneity withrespect to their projection patterns. All pedal-buccalprojection neurons that were tested received a slow ex-citatory postsynaptic potential from the ipsi- and con-tralateral cerebral-pedal regulator (C-PR) neuron, a cellthat is thought to play a key role in the generation of afood-induced arousal state. Tests were conducted toidentify potential synaptic follower neurons of the pedal-buccal projection neurons in the cerebral and buccalganglia, but none were detected. Finally, nerve record-ings revealed projections from the pedal-buccal projec-tion neurons in the nerves associated with the buccalganglion. In tests designed to determine the functionalproperties of these peripheral projections, no evidencewas obtained supporting a mechanosensory or pro-prioceptive role and no movements were observed whenthey were fired. It is proposed that peripheral elementsutilized in consummatory phases of Aplysia feeding maybe directly influenced by a neuronal pathway that isactivated during the food-induced arousal state.
Keywords Buccal ganglion Æ ConsummatoryC-PR Æ Molluscan feeding Æ P-BPN
Abbreviations BCI buccal-cerebral interneuron Æ BMPbuccal motor program Æ Bn buccal nerve Æ CAIcerebral-abdominal interneuron Æ CBI cerebral-buccalinterneuron Æ CBC cerebral-buccal connective Æ C-Pd Ccerebral-pedal connective Æ CPG central patterngenerator Æ C-PR cerebral-pedal regulator Æ EPSPexcitatory post-synaptic potential Æ IPSP inhibitorypost-synaptic potential Æ MCC metacerebral cell ÆP-BPN pedal-buccal projection neuron Æ PC pedalcommissure Æ Pl-Pd C pleural-pedal connective Æ PSPpostsynaptic potential
Introduction
The feeding-related behaviors of Aplysia exhibit severalproperties that are characteristic of motivated behaviorsin a broad spectrum of animals (Kupfermann et al. 1991,1992; Kupfermann 1994). For instance, Aplysia feedingis associated with a central motive state that is referredto as food-induced arousal (Kupfermann 1974a).Moreover, this complex behavior manifests a highlyvariable appetitive phase (head-raised posture, head-waving) that precedes a more stereotyped consumma-tory phase (biting, swallowing, rejection movements).Consequently, the neural circuit organization and cel-lular mechanisms that have been disclosed by intensiveinvestigation in this model system are likely to revealcomprehensive principles with broad applicability tomotivated animal behaviors.
The distributed organization of molluscan nervoussystems, in which control of explicit functions can beattributed to specific loci or ganglia, is one feature thathas facilitated analyses of the neuronal regulation ofbehavior (Kupfermann 1974b, Kandel 1976, 1979;Arshavsky et al. 1992, Norekian and Satterlie 1996,Sumbre et al. 2001). In the feeding system of Aplysia, forinstance, the motor patterns that produce rhythmicmovements of the intrinsic muscles of the buccal massduring consummatory behaviors are primarily producedby central pattern generator (CPG) circuits located
J Comp Physiol A (2003) 189: 231–244DOI 10.1007/s00359-003-0396-0
Alice Robie Æ Manuel Dı́az-Rı́os Æ Mark W. Miller
A. Robie Æ M. Dı́az-Rı́os Æ M.W. Miller (&)Institute of Neurobiology, University of Puerto Rico,201 Blvd. del Valle, 00901, San Juan, Puerto RicoE-mail: [email protected].: +1-787-724-1024Fax: +1-787-725-3804
A. Robie Æ M. Dı́az-Rı́os Æ M.W. MillerDepartment of Anatomy, University of Puerto Rico,201 Blvd. del Valle, 00901, San Juan, Puerto Rico

within the paired buccal ganglia (Susswein and Byrne1988; Kirk 1989; Plummer and Kirk 1990). These CPGcircuits are regulated by a limited number of sensoryneurons and interneurons that project from the cerebralganglion (Rosen et al. 1982, 1991; Perrins and Weiss1998; Hurwitz et al. 1999; Xin et al. 1999). Frequently,the cerebral interneurons that act as command elementsfor buccal motor programs are, in turn, influenced byfeedback from those circuits (Chiel et al. 1988; Rosenet al. 1991; Perrins and Weiss 1998; see also Gillette et al.1978; Delaney and Gelperin 1990).
The postural movements associated with the appeti-tive phase of feeding are mediated primarily by circuitslocated in the pedal and pleural ganglia (Bablanian et al.1987; Nagahama et al. 1993, 1994). These circuits mayalso be regulated by a limited number of interneuronsprojecting from the cerebral ganglion, one of which, thecerebral-pedal regulator (C-PR) has been the subject ofparticularly intensive study (Teyke et al. 1990, 1991,1997; Nagahama et al. 1993). The widespread synapticeffects that are produced by the C-PR neuron on circuitsassociated with body posture, cardiovascular regulation,and defensive behaviors support the proposal that thisneuron acts as a command-like element in the generationof the food-induced arousal state (Teyke et al. 1990,1997; Kupfermann et al. 1991). Interestingly, the syn-aptic actions of C-PR can influence circuits affiliatedwith both the appetitive and consummatory phases offeeding (Teyke et al. 1990; Hurwitz et al. 1999). While itsactivation of appetitive behaviors appears to result fromdirect projections to neurons in the pedal ganglion, itsexcitatory actions on consummatory neurons [e.g., theserotonergic metacerebral cell (MCC) and the cerebral-buccal interneurons (CBIs)] occur via a circuitous andpolysynaptic route through the pedal ganglion. It hasbeen proposed that this organization serves to ensureappropriate sequencing of feeding behavior patterns,i.e., an appetitive phase followed by a consummatoryphase (Teyke et al. 1997; Hurwitz et al. 1999).
The present study was prompted by the identificationof an increasing number of Aplysia interneurons withmultiple and distant interganglionic projections (Mac-key et al. 1989; Wright et al. 1995; Xin et al. 2001).Nerve tracing and electrophysiological methods wereused to identify a cluster of neurons in the pedal gan-glion with projections to the nerves innervating the in-trinsic buccal musculature. The additional observationthat these cells are activated by the C-PR neuron sug-gests that they provide a pathway via which peripheralelements utilized in consummatory phases of Aplysiafeeding may be directly regulated during early phases ofthe food-induced arousal state.
Materials and methods
Histological experiments were conducted on small (10–30 g) spec-imens of Aplysia californica that were purchased from the AplysiaResource Facility and Experimental Hatchery (University ofMiami, Miami, Fla., USA). Experiments requiring electrophysio-
logical recording and neuron identification were facilitated by usingganglia from larger specimens (150–250 g). Animals were main-tained in refrigerated aquaria (14–16�C) and fed dried seaweedtwice per week. All of the results reported in this study wereobserved on at least four specimens.
Connective backfills
The biotin-avidin protocol followed the methods of Xin et al.(1999) with modifications based upon Dı́az-Rı́os et al. (1999, 2002).Specimens were immobilized with an injection of isotonic MgCl2equal to 50% of body weight and the entire central nervous systemwas dissected, taking care to avoid damage to the connectives be-tween ganglia. The cerebral ganglion was pinned out near a smallVaseline well that was formed on the Sylgard surface. The cerebral-buccal connective (CBC) was cut and drawn into the well. Care wastaken to avoid contact between the end of the connective and theVaseline. The tip of the CBC was cut one more time and then theartificial seawater inside the well was withdrawn and replaced witha saturated aqueous solution (1.6 mg/30 ll) of biocytin (SigmaChemical, St. Louis Mo., USA). The walls of the well were thenbuilt up with successive layers of Vaseline, forming an ’igloo’ thateffectively isolated the biocytin pool from the artificial seawatersurrounding the ganglion. The preparation was covered and incu-bated overnight at 14�C. The well was then removed, and gangliawere washed three to five times in artificial seawater, repinned, andfixed in cold 4% paraformaldehyde (2–4 h). The fixed ganglia weretransferred to micro-centrifuge tubes, washed five times (30 mineach) with PTA (0.1 mol l)1 phosphate buffer containing 2% Tri-ton X-100 and 0.1 mmol l)1 sodium azide) solution and incubatedovernight (room temperature, with shaking) in Rhodamine600Avidin D (Vector Laboratories, Burlingame, Calif., USA) diluted1:3000 in PTA (24–48 h, room temperature).
Electrophysiology and neurobiotin injections
Neurons were identified in preparations consisting of the buccal,cerebral, and pedal ganglia. Extracellular recordings were acquiredwith polyethylene suction electrodes and a.c.-coupled amplifiers(Model 1700, AM Systems). The initial configuration typicallyconsisted of two en passant recordings from the cerebral-buccalconnectives and a single cut-end recording from a selected buccalnerve. In some experiments, maps of the pedal-buccal projectionneuron (P-BPN) clusters based upon CBC projections were estab-lished and then CBC electrodes were transferred to additionalbuccal nerves. The nomenclature for the buccal nerves correspondsto that of Gardner (1971). Intracellular microelectrodes filled with2 mol l)1 KCl were used to record from neurons in the P-BPNclusters and from potential pre- and postsynaptic cells. The normalartificial seawater solution had the following composition (inmmol l)1): 460 NaCl, 10 KCl, 11 CaCl2, 55 MgCl2, 10 HEPES. Ahigh-divalent (3· Ca2+, 3· Mg2+ with compensatory decreases ofNa+) saline was used to suppress polysynaptic activity in someexperiments. Experiments were conducted at room temperature(20–23�C).
Following identification and electrophysiological characteriza-tion of specific neurons, the KCl microelectrode was withdrawnand replaced with one containing neurobiotin. Injections weremodified from the methods described by Delgado et al. (2000)—seealso Dı́az-Rı́os et al. (2002). The microelectrode tips were filled with4% neurobiotin (Vector Laboratories) dissolved in 0.5 mol l)1
KCl, 50 mmol l)1 TRIS (pH 7.6). The electrode shafts were filledwith 2 mol l)1 KCl, resulting in resistances ranging from 10 to30 MX. Depolarizing current pulses (1–2 nA; 0.5 s; 1 Hz; 10–30 min) were used to eject the neurobiotin. Some preparations wereincubated overnight (4–14�C) in order to allow material to diffusefrom the injection site (cell body). They were then repinned ifnecessary, and fixed in paraformaldehyde as described above. Thefixed ganglia were transferred to microcentrifuge tubes, washed five
232
times (30 min each) with PTA solution and incubated in Rhod-amine600 Avidin D diluted (1:2,000–1:3,000) in PTA (24–48 h,room temperature).
Tests for P-BPN function were conducted in preparations inwhich the buccal mass remained innervated by the buccal nerves. Inall cases, the identity of intracellularly recorded P-BPNs was con-firmed with en passant recordings from the CBCs. A mechano-sensory function was tested by touching the lips, jaw, toothedradula surface, odontophore, oral cavity, and insertion points ofselected muscles with the closed point of a dull forceps. A possibleproprioceptive role was further examined during motor programsthat occurred spontaneously or in response to these tactile stimuli.Their potential function as motor neurons was tested by visualassessment of buccal muscle movements in response to firingindividual P-BPNs.
Results
The P-BPN cell cluster
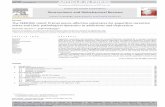
Examination of the cerebral ganglion following biocytinbackfills of the CBC revealed a group of fibers projectingtoward the ipsilateral cerebral-pedal connective (C-PdC). The majority of these fibers could be followed to aprominent cluster of five to seven P-BPNs on the ventralsurface of the ipsilateral pedal ganglion (Fig. 1A, ar-row). It is unlikely that these cells correspond to pedalneurons that have been previously examined, the ma-jority of which project to the periphery via pedal nerves(Rock et al. 1977; Hening et al. 1979; McPherson and
Blankenship 1991a, 1991b, 1991c; Skelton and Koester1992; Nagahama et al. 1993, 1994; Xin et al. 1996).Projections from identified pedal neurons to the C-Pd Chave not been reported, although several behavioral andelectrophysiological observations suggest that such pro-jections exist (see Discussion). The P-BPN cluster maybe most closely related to the ventral counterpart of thedorsal quadrant designated region II by Hening et al.(1979). However, it occupies a distinctly superficial po-sition, often protruding deeply into the overlyingganglionic sheath. No indication of bilateral asymmetryin the number or size of the P-BPNs was observed.
A single neuron on the ventral surface of the ipsi-lateral pleural ganglion was also consistently labeled inbackfills of the CBC (Fig. 1A, arrowhead). The axon ofthis cell projected to the pedal ganglion via the pleural-pedal connective (Pl-Pd C). It then turned and projectedto the cerebral ganglion via the C-Pd C. A single inter-neuron with similar axonal projections is found in thepleural ganglia of many pulmonate molluscs (Murphy1990; Alania 1995). It was not further examined in thisstudy.
The cell bodies of the neurons comprising the P-BPNcluster were heterogeneous in size, ranging from 40 to100 lm in diameter (Fig. 1B). They often had a slightelongation that gave them the appearance of being ori-ented along the rostral-caudal axis. Dye (neurobiotin)injections typically revealed two major processes origi-nating near the cell body (Fig. 1C, arrows), one givingrise to the axon projecting to the C-Pd C and the otherprojecting in the ventral direction, into the central coreof the ganglion.
Electrophysiological properties
The cells belonging to the P-BPN clusters shared severalelectrophysiological properties. Following a sufficientperiod to recover from microelectrode penetration, themajority of the P-BPNs exhibited a stable resting po-tential of )44.2±6.0 mV (mean±SD, n=15). However,some cells remained active for long periods (2–3 h), ex-hibiting a ‘beating’ mode of firing (�1 Hz–�1 Hz) thatdid not appear to be driven by synaptic input. Whilesmall amounts of depolarizing current were sufficient to
Fig. 1A–C Neurons in the pleural and pedal ganglia project to thecerebral-buccal connective (CBC). A Biocytin backfill of the CBCproduced labeling of one neuron in the pleural ganglion (arrow-head) and a cluster of neurons in the pedal ganglion (arrow). Imageshows ventral surface of the left pleural-pedal ganglia followingbackfill of the left CBC (not visible). The pleural cell has a singleaxon that projects into the cerebral-pedal connective (C-Pd C).Fluorescence in the C-Pd C at the top of this image originates fromthe pleural cell axon as it recedes from the focal plane. Calibrationbar=200 lm. B Pedal ganglion cluster from A shown at highermagnification reveals a group of seven somata that are heteroge-neous in size, ranging from 40 lm to 100 lm in diameter. C Asingle pedal neuron was injected with neurobiotin followingdemonstration of its projection to the CBC. Two major processesoriginate from the cell body, one projecting toward the C-PD C(left arrow) and the second projecting into the central neuropil(right arrow). Calibration bar=100 lm
233

bring the P-BPN cells to threshold, it was generally notpossible to produce firing at frequencies greater than10 Hz. Their somatic action potentials were large (peak-to-peak: 82.7±3.1 mV, n=6), relatively slow (durationat half maximal depolarization: 7.5±1.0 ms, n=6), andthey exhibited prominent after-hyperpolarizations.
The P-BPNs received a common synaptic input con-sisting of a train of high-frequency excitatory post-syn-aptic potentials (EPSPs) that occurred periodically andcaused them to fire two or three impulses (not shown).This input appeared to originate within the pedal gan-glion, but due to its low frequency of occurrence (periodof �0.5–�5 min) its source was not identified. There wasno indication of electrotonic or chemical synaptic sig-naling between pairs of neurons within a P-BPN ag-gregate or between members of bilateral clusters (notshown).
En passant extracellular recordings from the CBCswere used to determine the projection patterns of 50P-BPN neurons (Fig. 2). Of these, 30 projected to theipsilateral CBC (Fig. 2A), 4 projected to the contralat-eral CBC (Fig. 2B), and 6 had bilateral CBC projections(Fig. 2C). Ten neurons that appeared to belong to thecell cluster did not produce detectable signals in eitherCBC. The extracellular signals recorded from P-BPNcells in the CBC were often small, so their absence mayreflect the limits of the sensitivity of our recordingmethods. Moreover, the practice of desheathing the ce-rebral ganglia, although necessary for testing synapticconnections, could have produced a disruption ofP-BPN fibers of passage. These considerations not-withstanding, cells that do not project to either CBCmay in fact be present in the clusters that contain theP-BPNs.
The demonstration of contralateral and bilateralCBC projections by P-BPNs established that some ofthese cells give rise to commissural fibers. It was of
interest to determine whether the contralateral CBCprojections of the P-BPN cells originated from fiberstraversing the cerebral ganglion or whether their cross-ing occurred in the pedal commissure (PC). As the CBCbackfills rarely revealed fibers emerging from the con-tralateral C-Pd C or the C-Pl C, it was deduced that thebilateral P-BPNs were likely to accomplish their con-tralateral projections via the PC. This conclusion wassupported by experiments in which the contralateralCBC projections of bilaterally projecting P-BPNs wereeliminated upon bisection of the PC (Fig. 3, upper re-cords). As expected, this manipulation had no effect onprojections to the ipsilateral CBC (Fig. 3, middle re-cords). These findings prompted a careful reexaminationof the CBC biocytin backfill preparations, to determinewhether dye could be observed in contralateral P-BPNs.Although a few fibers could occasionally be discernedprojecting from the ipsilateral P-BPN cluster toward thePC, no contralateral cells were filled. Likewise, individ-ual neurons marked with intracellular injection of neu-robiotin (e.g., Fig. 1C) never revealed clear instances ofprojections into the contralateral pedal ganglion. Theseobservations are likely to reflect methodological limita-tions associated with the migration of biocytin andneurobiotin.
The P-BPN cells receive excitatory synapsesfrom the C-PR
The C-PR interneurons have been shown to influencenumerous cells in the pedal ganglia via direct and poly-synaptic connections (Teyke et al. 1990, 1991; Nagaha-ma et al. 1993, 1994). In consideration of these knownsynaptic actions, and the proposed role of C-PR in theinitiation of head-lifting components of the appetitivephase of feeding (Teyke et al. 1990; Kupfermann et al.1991; Nagahama et al. 1994), tests were conducted todetermine whether the P-BPNs were affected by firingC-PR. All of the ipsilateral P-BPNs that were examined(n=15) exhibited a slow and long-lasting depolarizingpostsynaptic potential (PSP) in response to a train ofhigh frequency C-PR firing (Fig. 4A). These responsesappeared to result from the summation of small(<1 mV) long-lasting PSPs that fused at particularplateau level (Fig. 4B). Following fusion, which typi-cally occurred within the initial six to eight PSPs, distinctinflections corresponding to the impulses of the C-PRwere not detected (Fig. 4B). When the membrane
Fig. 2A–C Individual pedal-buccal projection neurons (P-BPNs)project to the ipsilateral, contralateral, or both CBCs. A Intracel-lular recording from a P-BPN (upper recording) that produced one-for-one impulses with a brief and constant latency in the ipsilateralCBC (lower recording). Dotted lines were drawn to aid inidentification of extracellular spikes corresponding to the P-BPNimpulses. B Intracellular recording from a P-BPN (upper recording)that produced one-for-one impulses in the contralateral CBC(lower recording). C Intracellular recording from a P-BPN (upperrecording) that produced one-for-one impulses with brief andconstant latencies in the ipsilateral (middle recording) and contra-lateral CBCs (lower recording)
234
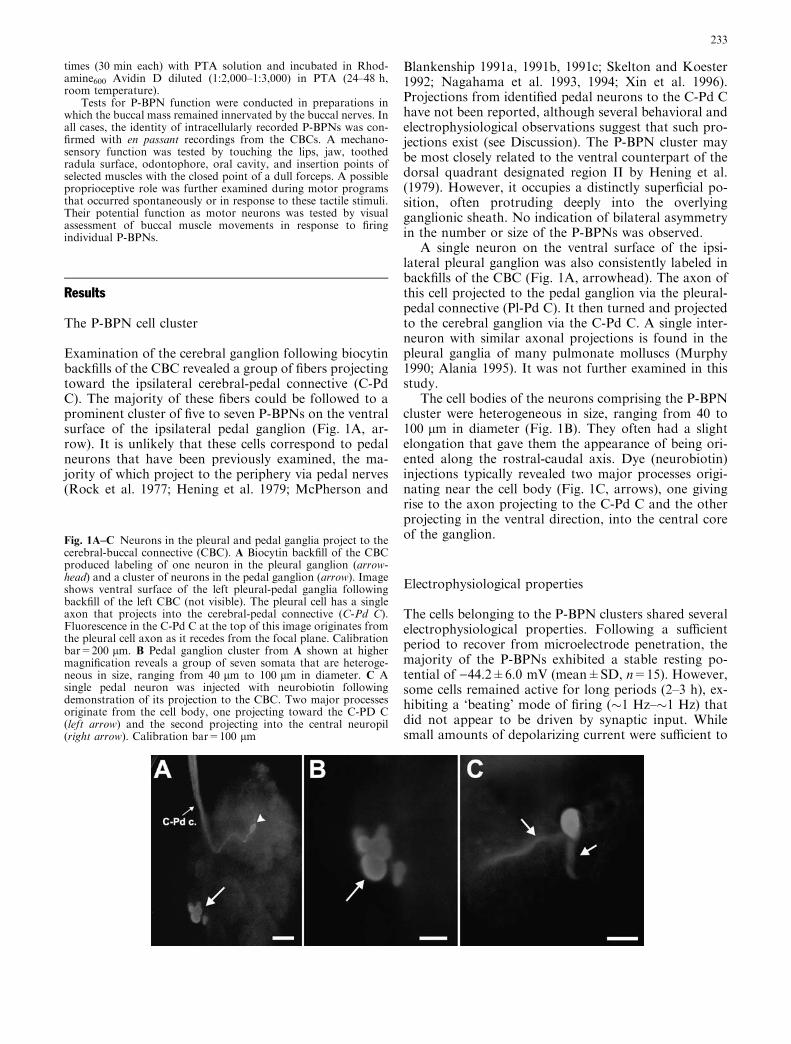
potential of a P-BPN was near its resting level, thesynaptic input from C-PR could produce repetitive fir-ing, demonstrating the excitatory nature of these depo-larizing PSPs (Fig. 4C). No reciprocal synapticconnections from the P-BPNs to the C-PR neuron wereobserved.
Each C-PR neuron projects to the contralateral pedalganglion via the PC (Teyke et al. 1997). Many pedalfollower neurons have been found to receive inputs ofthe same sign from both the ipsi- and the contralateralC-PR (Nagahama et al. 1993, 1994), suggesting that thetwo C-PR neurons possess substantial convergence andtarget overlap. It was therefore of interest to examinewhether the C-PR neurons produced comparable re-sponses in members of the contralateral P-BPN cluster.When C-PR neurons were fired at a high frequency viainjection of depolarizing current, slow EPSPs were ob-served in contralateral P-BPNs (Fig. 4D; n=6). Whilecomparable to their ipsilateral counterparts in directionand duration, the contralateral C-PR to P-BPN EPSPsappeared to be slightly reduced in amplitude (see alsoTeyke et al. 1997).
The slow depolarization observed in the P-BPNs inresponse to firing of the C-PR persisted in an artificialseawater solution in which Ca2+ and Mg2+ concentra-tions were raised (Fig. 5A). This observation indicatesthat the C-PR to P-BPN EPSP is produced, at least inpart, by direct monosynaptic signaling. When the
P-BPN membrane potential was pre-set at progressivelyhyperpolarized levels by injection of current, the EPSPwas found to decrease in amplitude (Fig. 5B–D). Al-though these responses were suggestive of an extremelyhigh reversal potential (greater than )90 mV; Fig. 5D),no instances of hyperpolarizing PSPs were observed so itwas not possible to reach conclusions regarding theirconductance mechanism.
Tests for synaptic interactions between the P-BPNsand neurons that participate in consummatoryphases of feeding behavior
The activity of the intrinsic muscles of the buccal massduring biting, swallowing, and rejection movements arelargely controlled by motor neurons and central pattern-generator circuits that are located within the pairedbuccal ganglia (Cohen et al. 1978; Susswein and Byrne1988; Kirk 1989; Morton and Chiel 1993; Church andLloyd 1994). These muscles and circuits are regulated bya relatively limited number of neurons that project to thebuccal ganglion from the cerebral ganglion via the CBCs(Weiss et al. 1978; Rosen et al. 1991; Perrins and Weiss1998; Hurwitz et al. 1999; Xin et al. 1999). The presenceof projections to the CBCs suggested that the P-BPNscould likewise be acting as interganglionic interneuronsin the regulation of consummatory behaviors.
Two interneurons that play key roles in the genera-tion of buccal motor patterns are the buccal-cerebralinterneurons (BCIs) designated B19 and B20. B19 pro-duces numerous synaptic actions within the cerebralganglion, where it is thought to provide a ‘read out’ ofthe buccal motor program that contributes to its coor-dination with the motor control of the extrinsic buccalmuscles (Chiel et al. 1988; Rosen et al. 1991, 2000a,2000b). No synaptic connections were detected from theP-BPNs to B19 (Fig. 6A, panel 1). In view of recentdemonstrations of remote excitation and antidromic
Fig. 3A,B Cutting the pedal commissure eliminates the P-BPNprojection to the contralateral CBC. A With the pedal commissureintact, firing a P-BPN with depolarizing intracellular current (barunder lower record) produced extracellular impulses with short andconstant latencies in extracellular recordings from the ipsilateral(middle record) and the contralateral (upper record) CBCs. Dottedlines were drawn to facilitate identification of extracellular spikescorresponding to P-BPN impulses. B After cutting the pedalcommissure, stimulation of the same P-BPN still producedimpulses in the ipsilateral CBC (middle record), but no longerproduced spikes in the contralateral CBC
235

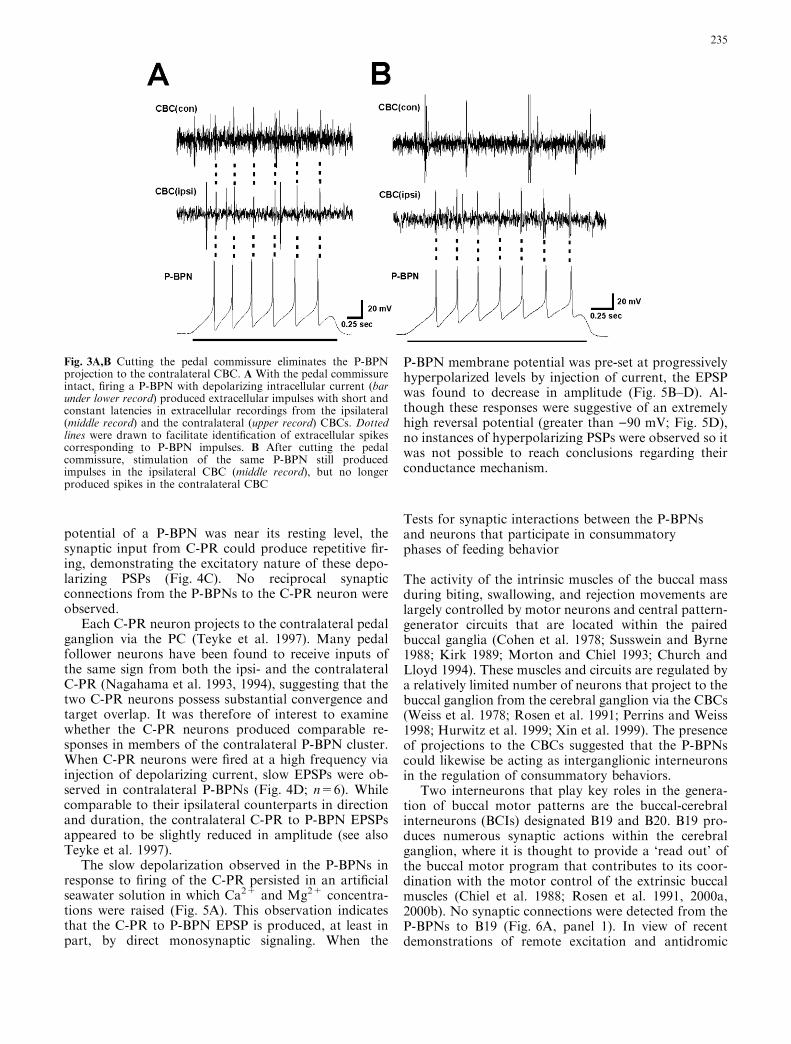
signaling in Aplysia interneurons (Perrins and Weiss1998), it was of interest to examine whether firing B19could affect the excitability of the P-BPNs via synapticconnections on their processes within the buccal or ce-rebral ganglia. Moreover, the presence of projectionsfrom B19 beyond the cerebral ganglion to the pedalganglion remains plausible. Tests were performed todetermine whether firing B19 exerted synaptic or mod-ulatory actions on the P-BPN cells, but none weredetected (Fig. 6A, panel 2).
Another BCI, designated B20, is thought to containboth GABA and dopamine (Teyke et al. 1993; Dı́az-Rı́os et al. 2002) and plays a key role in motor programswitching within the buccal ganglion (Jing and Weiss
Fig. 4A–D Firing the cerebral-pedal regulator (C-PR) produces aslow excitatory response in the P-BPNs. A Firing a C-PR neuronwith depolarizing intracellular current (bar below lower recording)produced a depolarizing response in an ipsilateral P-BPN (upperrecord). The P-BPN remained depolarized until the cessation ofC-PR firing. B The initial portion of the C-PR to P-BPN synapticresponse shown in A examined with a faster time base. Small(<1 mV) long-lasting ( >50 ms) post-synaptic potentials (PSPs)summated during the first six to eight C-PR impulses. The PSPsbecame fused at a level approximately 4 mV depolarized fromrest. C A P-BPN cell that was near its threshold level for impulsegeneration responded to firing of the C-PR (bar below lowerrecord) with repetitive action potentials (upper record), demon-strating the excitatory nature of this synaptic response. D Firingthe C-PR (bar below lower record) also produced an excitatoryresponse in a P-BPN located in the contralateral pedal ganglion(upper record)
236
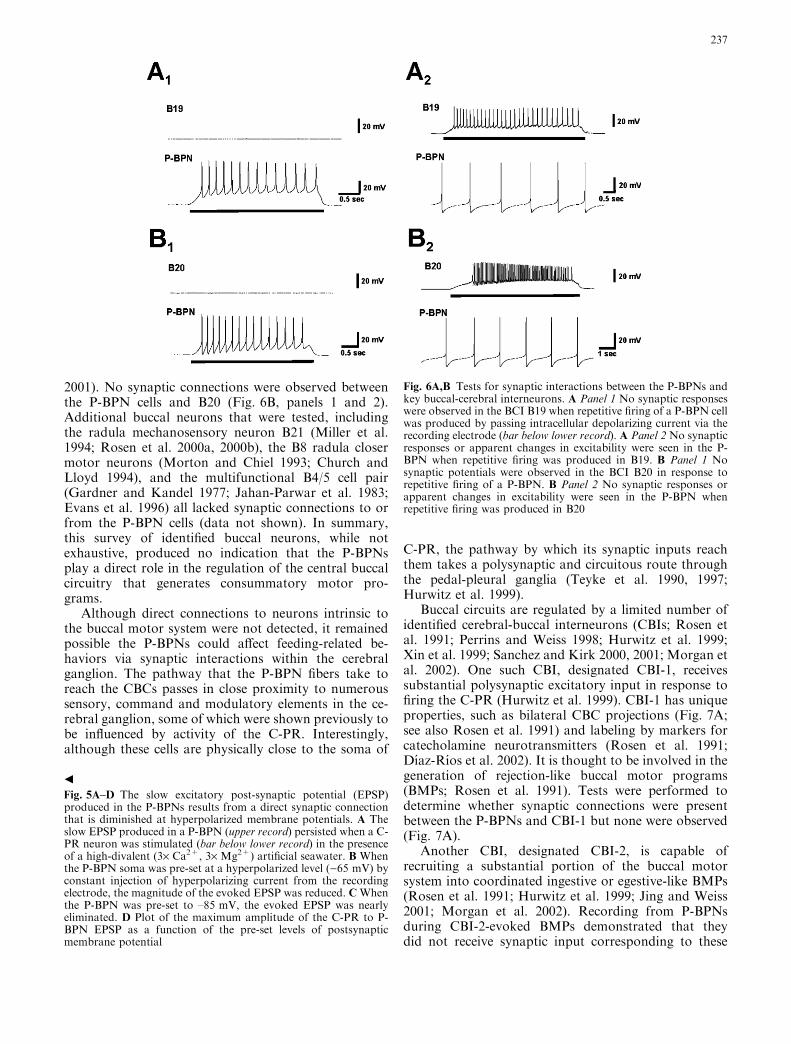
2001). No synaptic connections were observed betweenthe P-BPN cells and B20 (Fig. 6B, panels 1 and 2).Additional buccal neurons that were tested, includingthe radula mechanosensory neuron B21 (Miller et al.1994; Rosen et al. 2000a, 2000b), the B8 radula closermotor neurons (Morton and Chiel 1993; Church andLloyd 1994), and the multifunctional B4/5 cell pair(Gardner and Kandel 1977; Jahan-Parwar et al. 1983;Evans et al. 1996) all lacked synaptic connections to orfrom the P-BPN cells (data not shown). In summary,this survey of identified buccal neurons, while notexhaustive, produced no indication that the P-BPNsplay a direct role in the regulation of the central buccalcircuitry that generates consummatory motor pro-grams.
Although direct connections to neurons intrinsic tothe buccal motor system were not detected, it remainedpossible the P-BPNs could affect feeding-related be-haviors via synaptic interactions within the cerebralganglion. The pathway that the P-BPN fibers take toreach the CBCs passes in close proximity to numeroussensory, command and modulatory elements in the ce-rebral ganglion, some of which were shown previously tobe influenced by activity of the C-PR. Interestingly,although these cells are physically close to the soma of
C-PR, the pathway by which its synaptic inputs reachthem takes a polysynaptic and circuitous route throughthe pedal-pleural ganglia (Teyke et al. 1990, 1997;Hurwitz et al. 1999).
Buccal circuits are regulated by a limited number ofidentified cerebral-buccal interneurons (CBIs; Rosen etal. 1991; Perrins and Weiss 1998; Hurwitz et al. 1999;Xin et al. 1999; Sanchez and Kirk 2000, 2001; Morgan etal. 2002). One such CBI, designated CBI-1, receivessubstantial polysynaptic excitatory input in response tofiring the C-PR (Hurwitz et al. 1999). CBI-1 has uniqueproperties, such as bilateral CBC projections (Fig. 7A;see also Rosen et al. 1991) and labeling by markers forcatecholamine neurotransmitters (Rosen et al. 1991;Dı́az-Rı́os et al. 2002). It is thought to be involved in thegeneration of rejection-like buccal motor programs(BMPs; Rosen et al. 1991). Tests were performed todetermine whether synaptic connections were presentbetween the P-BPNs and CBI-1 but none were observed(Fig. 7A).
Another CBI, designated CBI-2, is capable ofrecruiting a substantial portion of the buccal motorsystem into coordinated ingestive or egestive-like BMPs(Rosen et al. 1991; Hurwitz et al. 1999; Jing and Weiss2001; Morgan et al. 2002). Recording from P-BPNsduring CBI-2-evoked BMPs demonstrated that theydid not receive synaptic input corresponding to these
b
Fig. 5A–D The slow excitatory post-synaptic potential (EPSP)produced in the P-BPNs results from a direct synaptic connectionthat is diminished at hyperpolarized membrane potentials. A Theslow EPSP produced in a P-BPN (upper record) persisted when a C-PR neuron was stimulated (bar below lower record) in the presenceof a high-divalent (3· Ca2+, 3· Mg2+) artificial seawater. B Whenthe P-BPN soma was pre-set at a hyperpolarized level ()65 mV) byconstant injection of hyperpolarizing current from the recordingelectrode, the magnitude of the evoked EPSP was reduced. CWhenthe P-BPN was pre-set to –85 mV, the evoked EPSP was nearlyeliminated. D Plot of the maximum amplitude of the C-PR to P-BPN EPSP as a function of the pre-set levels of postsynapticmembrane potential
Fig. 6A,B Tests for synaptic interactions between the P-BPNs andkey buccal-cerebral interneurons. A Panel 1 No synaptic responseswere observed in the BCI B19 when repetitive firing of a P-BPN cellwas produced by passing intracellular depolarizing current via therecording electrode (bar below lower record). A Panel 2 No synapticresponses or apparent changes in excitability were seen in the P-BPN when repetitive firing was produced in B19. B Panel 1 Nosynaptic potentials were observed in the BCI B20 in response torepetitive firing of a P-BPN. B Panel 2 No synaptic responses orapparent changes in excitability were seen in the P-BPN whenrepetitive firing was produced in B20
237
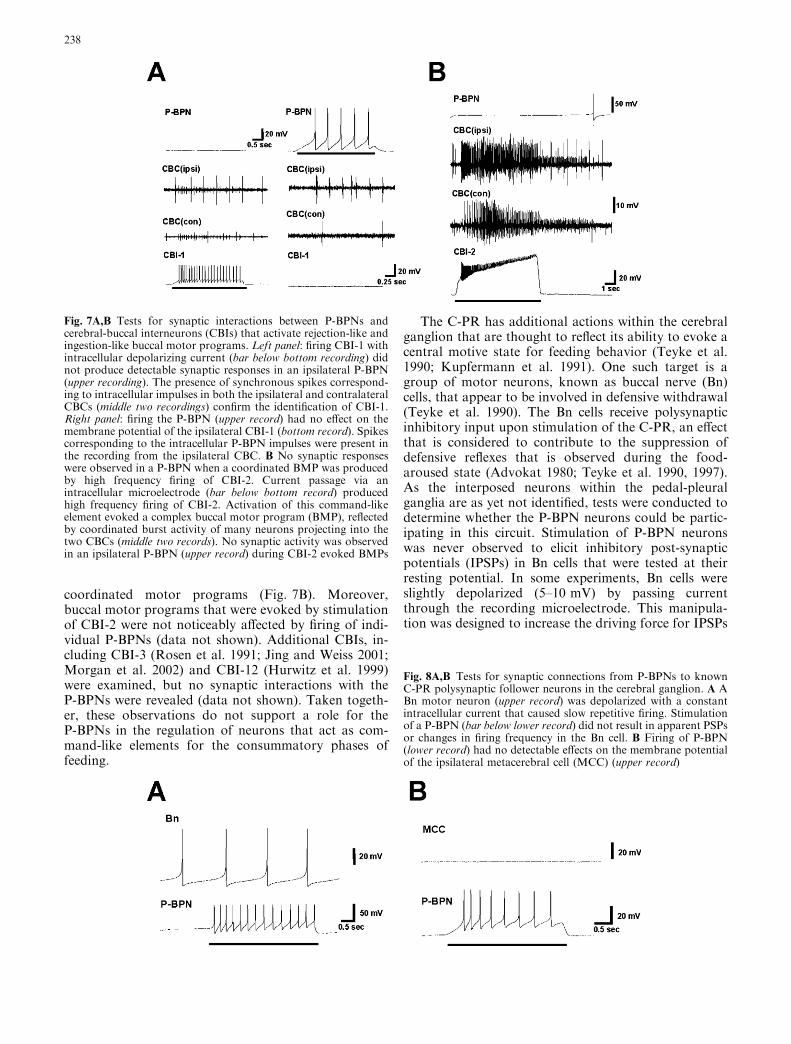
coordinated motor programs (Fig. 7B). Moreover,buccal motor programs that were evoked by stimulationof CBI-2 were not noticeably affected by firing of indi-vidual P-BPNs (data not shown). Additional CBIs, in-cluding CBI-3 (Rosen et al. 1991; Jing and Weiss 2001;Morgan et al. 2002) and CBI-12 (Hurwitz et al. 1999)were examined, but no synaptic interactions with theP-BPNs were revealed (data not shown). Taken togeth-er, these observations do not support a role for theP-BPNs in the regulation of neurons that act as com-mand-like elements for the consummatory phases offeeding.
The C-PR has additional actions within the cerebralganglion that are thought to reflect its ability to evoke acentral motive state for feeding behavior (Teyke et al.1990; Kupfermann et al. 1991). One such target is agroup of motor neurons, known as buccal nerve (Bn)cells, that appear to be involved in defensive withdrawal(Teyke et al. 1990). The Bn cells receive polysynapticinhibitory input upon stimulation of the C-PR, an effectthat is considered to contribute to the suppression ofdefensive reflexes that is observed during the food-aroused state (Advokat 1980; Teyke et al. 1990, 1997).As the interposed neurons within the pedal-pleuralganglia are as yet not identified, tests were conducted todetermine whether the P-BPN neurons could be partic-ipating in this circuit. Stimulation of P-BPN neuronswas never observed to elicit inhibitory post-synapticpotentials (IPSPs) in Bn cells that were tested at theirresting potential. In some experiments, Bn cells wereslightly depolarized (5–10 mV) by passing currentthrough the recording microelectrode. This manipula-tion was designed to increase the driving force for IPSPs
Fig. 7A,B Tests for synaptic interactions between P-BPNs andcerebral-buccal interneurons (CBIs) that activate rejection-like andingestion-like buccal motor programs. Left panel: firing CBI-1 withintracellular depolarizing current (bar below bottom recording) didnot produce detectable synaptic responses in an ipsilateral P-BPN(upper recording). The presence of synchronous spikes correspond-ing to intracellular impulses in both the ipsilateral and contralateralCBCs (middle two recordings) confirm the identification of CBI-1.Right panel: firing the P-BPN (upper record) had no effect on themembrane potential of the ipsilateral CBI-1 (bottom record). Spikescorresponding to the intracellular P-BPN impulses were present inthe recording from the ipsilateral CBC. B No synaptic responseswere observed in a P-BPN when a coordinated BMP was producedby high frequency firing of CBI-2. Current passage via anintracellular microelectrode (bar below bottom record) producedhigh frequency firing of CBI-2. Activation of this command-likeelement evoked a complex buccal motor program (BMP), reflectedby coordinated burst activity of many neurons projecting into thetwo CBCs (middle two records). No synaptic activity was observedin an ipsilateral P-BPN (upper record) during CBI-2 evoked BMPs
Fig. 8A,B Tests for synaptic connections from P-BPNs to knownC-PR polysynaptic follower neurons in the cerebral ganglion. A ABn motor neuron (upper record) was depolarized with a constantintracellular current that caused slow repetitive firing. Stimulationof a P-BPN (bar below lower record) did not result in apparent PSPsor changes in firing frequency in the Bn cell. B Firing of P-BPN(lower record) had no detectable effects on the membrane potentialof the ipsilateral metacerebral cell (MCC) (upper record)
238

that could be obscured at membrane potentials close torest. Moreover, such a depolarization typically resultedin spontaneous firing at low frequency (beating), thusmaking it possible to explore possible subtle effects onmembrane excitability resulting from P-BPN firing. NoIPSPs or changes in excitability were detected in Bn cellswhen P-BPN cells were fired (Fig. 8A).
The serotonergic MCC is thought to increase theoverall efficiency of feeding responses via its widespreadmodulatory actions on the muscles and neurons that
participate in buccal motor programs (Weiss et al. 1978;Rosen et al. 1989; Alexeeva et al. 1998; Fox and Lloyd1998). The MCC receives a slow excitatory PSP whenthe C-PR is fired (Teyke et al. 1990, 1997). This PSP alsoresults from a polysynaptic route through the pedal-pleural ganglia, but the interposed cells appear to bedistinct from those influencing the Bn cells (Teyke et al.1997). Tests were therefore conducted to determinewhether the P-BPN neurons exerted synaptic effects onthe MCC but none were detected (Fig. 8B).
In summary, these experiments provided no indica-tion that the P-BPNs either influence or are influencedby buccal motor programs or the cerebral neurons thatregulate consummatory behaviors. Moreover, no inter-actions were detected with neurons that participate inthe expression of the central motive state for feeding.
The P-BPNs project fibers into buccal nerves: testsfor peripheral function
While the observations presented above revealed nocentral actions of the P-BPNs, the possibility remainedthat these cells could influence consummatory behaviorsvia peripheral projections. Extracellular nerve record-ings were therefore used to determine whether theP-BPNs could possess direct access to peripheral com-ponents of the buccal system. Upon firing individualP-BPNs, small impulses were detected with a brief andconstant latency in each of the buccal nerves with theexception of the esophageal nerve (Fig. 9). A particularP-BPN could project into one or more ipsilateral buccal
Fig. 9A–D Fibers from the P-BPNs project to buccal nerves (Bns).A Recording from the ipsilateral Bn 1 (nomenclature of Gardner1971) revealed one-for-one spikes corresponding to impulsesgenerated in a P-BPN during injection of depolarizing current(bar below lower record). B Recording from the contralateral Bn 2exhibited spikes corresponding to impulses generated in a P-BPN.C Recordings from both ipsilateral and contralateral Bn 3 bothincluded spikes corresponding to impulses in a P-BPN. DSchematic summary of some of the properties of the P-BPNs.The P-BPN clusters of cell bodies (black circles) are located on theventral surface of each pedal ganglion (shown only on right side offigure). They receive direct synaptic connections from the ipsilateraland contralateral C-PR (dashed gray lines). Some P-BPNs projectfibers into the ipsilateral cerebral-pedal connective (C-PD C; solidblack line) while others project into the contralateral C-PD C viathe pedal commissure (dotted black line). Still others have bilateralC-PD C projections (not shown for purposes of clarity). Within thebuccal ganglion, some P-BPN fibers project out one or more Bnsthat are ipsilateral to its side of its ascent (dotted black lines in Bn 1,2, and 3). Others deccussate via the buccal commissure, resulting inboth ipsilateral and contralateral buccal nerve projections (solidblack line). The neurons depicted are based upon actual recordingsand were chosen to illustrate the variability of P-BPN projectionpatterns
239
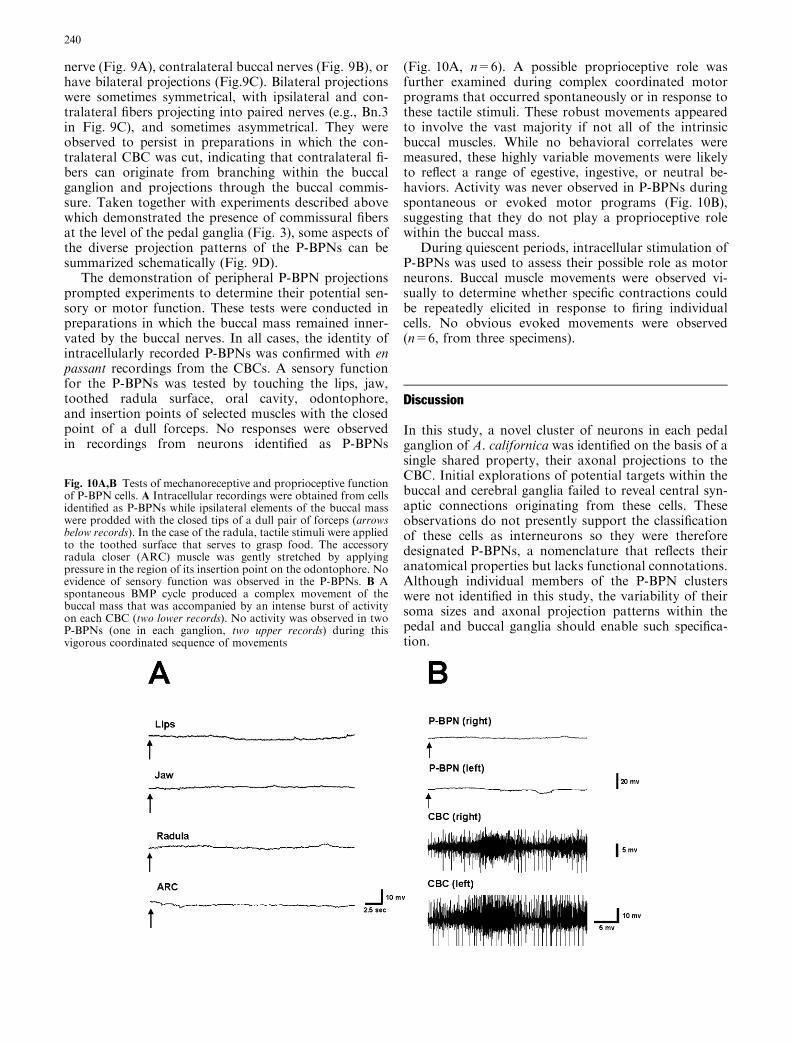
nerve (Fig. 9A), contralateral buccal nerves (Fig. 9B), orhave bilateral projections (Fig.9C). Bilateral projectionswere sometimes symmetrical, with ipsilateral and con-tralateral fibers projecting into paired nerves (e.g., Bn.3in Fig. 9C), and sometimes asymmetrical. They wereobserved to persist in preparations in which the con-tralateral CBC was cut, indicating that contralateral fi-bers can originate from branching within the buccalganglion and projections through the buccal commis-sure. Taken together with experiments described abovewhich demonstrated the presence of commissural fibersat the level of the pedal ganglia (Fig. 3), some aspects ofthe diverse projection patterns of the P-BPNs can besummarized schematically (Fig. 9D).
The demonstration of peripheral P-BPN projectionsprompted experiments to determine their potential sen-sory or motor function. These tests were conducted inpreparations in which the buccal mass remained inner-vated by the buccal nerves. In all cases, the identity ofintracellularly recorded P-BPNs was confirmed with enpassant recordings from the CBCs. A sensory functionfor the P-BPNs was tested by touching the lips, jaw,toothed radula surface, oral cavity, odontophore,and insertion points of selected muscles with the closedpoint of a dull forceps. No responses were observedin recordings from neurons identified as P-BPNs
(Fig. 10A, n=6). A possible proprioceptive role wasfurther examined during complex coordinated motorprograms that occurred spontaneously or in response tothese tactile stimuli. These robust movements appearedto involve the vast majority if not all of the intrinsicbuccal muscles. While no behavioral correlates weremeasured, these highly variable movements were likelyto reflect a range of egestive, ingestive, or neutral be-haviors. Activity was never observed in P-BPNs duringspontaneous or evoked motor programs (Fig. 10B),suggesting that they do not play a proprioceptive rolewithin the buccal mass.
During quiescent periods, intracellular stimulation ofP-BPNs was used to assess their possible role as motorneurons. Buccal muscle movements were observed vi-sually to determine whether specific contractions couldbe repeatedly elicited in response to firing individualcells. No obvious evoked movements were observed(n=6, from three specimens).
Discussion
In this study, a novel cluster of neurons in each pedalganglion of A. californica was identified on the basis of asingle shared property, their axonal projections to theCBC. Initial explorations of potential targets within thebuccal and cerebral ganglia failed to reveal central syn-aptic connections originating from these cells. Theseobservations do not presently support the classificationof these cells as interneurons so they were thereforedesignated P-BPNs, a nomenclature that reflects theiranatomical properties but lacks functional connotations.Although individual members of the P-BPN clusterswere not identified in this study, the variability of theirsoma sizes and axonal projection patterns within thepedal and buccal ganglia should enable such specifica-tion.
Fig. 10A,B Tests of mechanoreceptive and proprioceptive functionof P-BPN cells. A Intracellular recordings were obtained from cellsidentified as P-BPNs while ipsilateral elements of the buccal masswere prodded with the closed tips of a dull pair of forceps (arrowsbelow records). In the case of the radula, tactile stimuli were appliedto the toothed surface that serves to grasp food. The accessoryradula closer (ARC) muscle was gently stretched by applyingpressure in the region of its insertion point on the odontophore. Noevidence of sensory function was observed in the P-BPNs. B Aspontaneous BMP cycle produced a complex movement of thebuccal mass that was accompanied by an intense burst of activityon each CBC (two lower records). No activity was observed in twoP-BPNs (one in each ganglion, two upper records) during thisvigorous coordinated sequence of movements
240

The P-BPNs appear to participate in circuits thatexhibit both divergent and convergent synaptic signaling.In most preparations, all members of the cluster receiveda rhythmic burst of EPSPs that originated from anuninvestigated oscillator residing within the pedal gan-glia. An additional divergent presynaptic element in thiscircuit, the identified C-PR interneuron, produced a slowEPSP in each of the P-BPNs. The presence of conver-gence in this network is illustrated by the similarresponses of individual P-BPNs to firing of the ipsilateraland contralateral C-PRs. Such bilateral convergence ofC-PR signaling has been observed previously in motorneurons (Nagahama et al. 1993) and modulatory neu-rons (Nagahama et al. 1994) that innervate the neck andanterior foot. This convergence was interpreted as sup-porting a role for the C-PR in the execution of headlifting, rather than in head waving or other stronglylateralized movements (see also Kupfermann et al. 1991).
Previous investigations have revealed a limited num-ber of direct postsynaptic follower cells for the C-PR(Nagahama et al. 1993, 1994). The dynamics of theEPSP observed in the P-BPN neurons, with its pro-longed onset and decay, differs from the more conven-tional PSPs observed previously. These findings indicatethat the C-PR can exert specific forms of signaling (fastconventional or slow modulatory) in distinct targets.The capability of individual interneurons to modifyCPG circuits via signals that vary over a broad temporalrange may be widely used in the regulation and selectionof motor programs (Getting 1989; Katz et al. 1994; Blitzand Nusbaum 1999; Jing and Weiss 2001). Such sig-naling can result from distinct postsynaptic responses toa particular neurotransmitter (Katz and Frost 1995;Norekian 1997), or from dissimilar actions of cotrans-mitters (Blitz and Nusbaum 1999). While the identitiesof neurotransmitters that may participate in C-PR sig-naling are presently unknown, immunohistochemicaltechniques indicate that it contains an epitope that isrecognized by antibodies generated against crustaceancardioactive peptide (CCAP; Delgado et al. 2000).
The paired C-PRs are thought to contribute to theactivation of a food-induced arousal state includingappetitive components (specifically lifting of the headfrom the substrate) of feeding behavior that are medi-ated primarily by circuits within the pedal-pleural gan-glia (Teyke et al. 1990, 1991; Hurwitz et al. 1999). TheP-BPN cells possess the apparently paradoxical propertyof projecting directly to the intrinsic buccal musculaturethat controls protraction, retraction, opening, andclosing of the radula, movements that are considered tooccur during consummatory phases of feeding. Severalprior observations, however, support the notion thatconsummatory responses are influenced by activityoriginating from distant sources. For instance, speci-mens were observed to exhibit deficits in the intensityand rate of biting responses following bilateral lesions ofthe C-Pd Cs (Kupfermann 1974b; Jahan-Parwar andFredman 1979; Teyke et al. 1991). Moreover, in vivorecordings from the C-Pd C showed an increase in
activity during biting and swallowing (Teyke et al. 1991).Recently, it was shown that increases in CBC activitycould be produced by firing cerebral-abdominal inter-neurons (CAIs) and that this evoked activity was elim-inated by cutting the C-Pd and C-Pl connectives (Xinet al. 2001).
The C-PR neuron is known to have access to thecentral circuitry that regulates consummatory behaviors.Its targets include CBIs that are capable of evokingbiting responses (Teyke et al. 1990; Hurwitz et al. 1999)and the serotonergic MCC (Teyke et al. 1990), whichacts in a modulatory fashion to increase the speed andmagnitude of successive bites during consummatoryarousal (Rosen et al. 1989). The circuitous and indirectroute of CBI and MCC activation by the C-PR has beenproposed to contribute to the sequencing of feedingbehaviors, ensuring that the postural head-up compo-nents are achieved prior to initiation of ingestive pro-grams (Teyke et al. 1997; Hurwitz et al. 1999; see alsoMorgan et al. 2000). Although these pathways areknown to include synaptic connections in the pedal-pleural ganglia, they do not appear to involve theP-BPNs. Rather, the P-BPNs seem to provide an inde-pendent and parallel pathway that projects directly tothe peripheral plant that controls radula movements.
The projections of the P-BPNs to the buccal nervesare likely to produce an extensive innervation of theintrinsic buccal musculature (Cohen et al. 1978; Scott etal. 1991; Jordan et al. 1993; Church and Lloyd 1994).While the cell bodies of several classes of sensory neu-rons are located within the CNS of Aplysia (Byrne et al.1974; Rosen et al. 1979; Walters et al. 1983; Weiss et al.1986a, 1986b; Miller et al. 1994; Borovikov et al. 2000),the absence of detectable mechanosensory and pro-prioceptive responses, coupled with the activation of theP-BPNs by the C-PR, suggests that these cells are morelikely to be involved in some aspect of motor control.Although the C-PR is known to innervate motor neu-rons in the pedal ganglion (Nagahama et al. 1993), nobuccal muscle movements were observed in response tofiring the P-BPNs. However, in view of the predominantrole of modulatory neurons in the production of food-induced postural behaviors (Nagahama et al. 1994), thepossibility remains that these cells act to modify orregulate the peripheral actions of buccal motor neurons.The presence of numerous identified buccal motor neu-rons with known regions of innervation in specific buc-cal muscles (Cohen et al. 1978; Jordan et al. 1993;Church and Lloyd 1994; Hurwitz et al. 1996; Evans et al.1996) should facilitate investigation of the potentialmodulatory role of the P-BPNs.
The branching patterns of the P-BPNs within thebuccal ganglion and their radiation into each of thebuccal nerves bear some similarity to the serotonergicMCC (Weiss and Kupfermann 1976; Goldstein et al.1982). However, in contrast to theMCC, in which a singleneuron projects a large caliber fiber into each of thebuccal nerves, it appears that distinct P-BPNs have morerestricted projections. Collectively, the P-BPNs are likely
241

to reach many of the peripheral targets that are known tobe innervated by the MCC (Weiss et al. 1978; Alexeevaet al. 1998; Fox and Lloyd 1998; Hurwitz et al. 2000).Although neuron-specific synaptic inputs to the P-BPNswere not observed, the multicellular organization of thissystem may enable more selective responses than can beachieved with the MCC. While the identification of theneurotransmitters that are present in the P-BPNs remainslargely unexplored, double-labeling experiments (notshown) demonstrated that they do not contain serotonin.This finding is consistent with previous observationsindicating that the MCC provides the sole serotonergicinput to the buccal system (Ono and McCaman 1984;Longley and Longley 1986; Alexeeva et al. 1998).
Modulatory regulation of motor systems is oftenreferred to as being ’intrinsic’ when it is derived frommotor neurons or interneurons comprising the CPGnetwork that is being modified (Cropper et al. 1987;Katz and Frost 1995). Modulation is considered to be‘extrinsic’ when it originates from neurons that do notparticipate directly in the mediation of the motor act(Weiss et al. 1978, 1982; Kupfermann et al. 1997;Hooper et al. 1999). It has been observed that extrinsicmodulation appears to play a predominant role in theregulation of appetitive feeding behaviors in Aplysia(Nagahama et al. 1994) while consummatory behaviorsseem to make extensive use of intrinsic modulation(Cropper et al. 1987; Church and Lloyd 1994; but seeMorgan et al. 2000). In view of the apparent absence ofsynaptic interactions with motor neurons, sensory neu-rons, interneurons, and command elements for con-summatory behaviors, the P-BPNs must presently beconsidered to be extrinsic to the buccal motor systems.Moreover, their sole demonstrated source of activationoriginates from the C-PR, a higher-order extrinsic in-terneuron that is thought to produce multiple manifes-tations of a food-induced central motive state.
Recent studies have shown that the precise phasing ofconsummatory behaviors differs in animals that arefeeding in the head-up position versus the head-down(grazing) posture (Saunders et al. 1998; Rosen et al. 2002;see also Kupfermann and Carew 1974). The centralconnections of the C-PR suggest that it may be selectivelyinvolved in promoting feeding in the head-up posture(Hurwitz et al. 1999). In view of their direct activation bythe C-PR, the P-BPNs could serve a role in regulating theperipheral buccal plant during behaviors that are spe-cifically associated with feeding in the head-up posture.
Acknowledgements Supported in part by NSF: CAREER AwardIBN-9722349; NIH: NS07464 (NINDS), NS039405 (SNRP,NINDS), NS045546-02 (NINDS), MH048190 (M-RISP, NIMH),RR03051 (RCMI, Division of Research Services), and GM08224(MBRS, NIGMS). M.D.-R. received support from the APA Mi-nority Fellowship Program in Neuroscience. The students andfaculty of the Tropical Neuroethology for Undergraduates summerprogram (2001) were instrumental in developing some of themethods used in this study. Experimental protocols followed es-tablished IACUC guidelines of the University of Puerto RicoMedical Sciences Campus.
References
Advokat C (1980) Modulation of defensive reflexes in Aplysiacalifornica by appetitive stimulation. Behav Neural Biol 28:253–265
Alania M (1995) Pleuro-buccal projections in pulmonate molluscs.Acta Biol Hung 46:267–270
Alexeeva V, Borovikov D, Miller MW, Rosen SC, Cropper EC(1998) Effect of a serotonergic extrinsic modulatory neuron(MCC) on radula mechanoafferent function in Aplysia. JNeurophysiol 80:1609–1622
Arshavsky YI, Deliagina TG, Orlovsky GN, Panchin YV, PopovaLB (1992) Interneurones mediating the escape reaction of themarine mollusc Clione limacina. J Exp Biol 164:307–314
Bablanian GM, Weiss KR, Kupfermann I (1987) Motor control ofthe appetitive phase of feeding behavior in Aplysia. BehavNeural Biol 48:394–407
Blitz DM, Nusbaum MP (1999) Distinct functions for cotrans-mitters mediating motor pattern selection. J Neurosci 19:6774–6783
Borovikov D, Evans CG, Jing J, Rosen SC, Cropper EC (2000) Aproprioceptive role for an exteroceptive mechanoafferent neu-ron in Aplysia. J Neurosci 20:1990–2002
Byrne J, Castellucci V, Kandel ER (1974) Receptive fields andresponse properties of mechanoreceptor neurons innervatingsiphon skin and mantle shelf in Aplysia. J Neurophysiol37:1041–1064
Chiel HJ, Kupfermann I, Weiss KR (1988) An identified hista-minergic neuron can modulate the outputs of buccal-cerebralinterneurons in Aplysia via presynaptic inhibition. J Neurosci8:49–63
Church PJ, Lloyd PE (1994) Activity of multiple identified motorneurons recorded intracellularly during evoked feeding-likemotor programs in Aplysia. J Neurophysiol 72:1794–1809
Cohen JL, Weiss KR, Kupfermann I (1978) Motor control ofbuccal muscles in Aplysia. J Neurophysiol 41:157–180
Cropper EC, Lloyd PE, Reed W, Tenenbaum R, Kupfermann I,Weiss KR (1987) Multiple neuropeptides in cholinergic motorneurons of Aplysia: evidence for modulation intrinsic to themotor circuit. Proc Natl Acad Sci USA 84:3486–3490
Delgado JY, Oyola E, Miller MW (2000) Localization of GABA-and glutamate-like immunoreactivity in the cardiac ganglion ofthe spiny lobster Panulirus argus. J Neurocytol 29:605–619
Delaney K, Gelperin A (1990) Cerebral interneurons controllingfictive feeding in Limax maximus. II. Initiation and modulationof fictive feeding. J Comp Physiol A 166:311–326
Dı́az-Rı́os M, Suess E, Miller MW (1999) Localization of GABA-like immunoreactivity in the central nervous system of Aplysiacalifornica. J Comp Neurol 413:255–270
Dı́az-Rı́os M, Oyola E, Miller MW (2002) Colocalization ofc-aminobutyric acid-like immunoreactivity and catecholaminesin the feeding network of Aplysia californica. J Comp Neurol445:29–46
Evans CG, Rosen S, Kupfermann I, Weiss KR, Cropper EC (1996)Characterization of a radula opener neuromuscular system inAplysia. J Neurophysiol 76:1267–1281
Fox LE, Lloyd PE (1998) Serotonergic neurons differentiallymodulate the efficacy of two motor neurons innervating thesame muscle fibers in Aplysia. J Neurophysiol 80:647–655
Gardner D (1971) Bilateral symmetry and internal organization inthe buccal ganglia of Aplysia. Science 173:550–553
Gardner D, Kandel ER (1977) Physiological and kineticproperties of cholinergic receptors activated by multiactioninterneurons in buccal ganglia of Aplysia. J Neurophysiol40:333–348
Getting PA (1989) Emerging principles governing the operation ofneural networks. Annu Rev Neurosci 12:185–204
Gillette R, Kovac MP, Davis WJ (1978) Command neurons inPleurobranchaea receive synaptic feedback from the motornetwork they excite. Science 199:798–801
242

Goldstein RS, Weiss KR, Schwartz JH (1982) Intraneuronal in-jection of horseradish peroxidase labels glial cells associatedwith the axons of the giant metacerebral neuron of Aplysia.J Neurosci 2:1567–1577
Hening WA, Walters ET, Carew TJ, Kandel ER (1979) Motor-neuronal control of locomotion in Aplysia. Brain Res 179:231–253
Hooper SL, Brezina V, Cropper EC, Weiss KR (1999) Flexibility ofmuscle control by modulation of muscle properties. In: Katz PS(ed) Beyond neurotransmission: neuromodulation and itsimportance for information processing. Oxford UniversityPress, New York, pp 241–274
Hurwitz I, Neustadter D, Morton DW, Chiel HJ, Susswein AJ(1996) Activity patterns of the B31/B32 pattern initiatorsinnervating the I2 muscle of the buccal mass during normalfeeding movements in Aplysia californica. J Neurophysiol75:1309–1326
Hurwitz I, Perrins R, Xin Y, Weiss, KR, Kupfermann I (1999) C-PR neuron of Aplysia has differential effects on ’feeding’ cere-bral interneurons, including myomodulin-positive CBI-12.J Neurophysiol 81:521–534
Hurwitz I, Cropper EC, Vilim FS, Alexeeva V, Susswein AJ,Kupfermann I, Weiss KR (2000) Serotonergic and peptidergicmodulation of the buccal mass protractor muscle (I2) in Aply-sia. J Neurophysiol 84:2810–2820
Jahan-Parwar B, Fredman SM (1979) Role of interganglionicsynaptic connections in the control of pedal and parapodialmovements in Aplysia. Brain Res Bull 4:407–420
Jahan-Parwar B, Wilson AH Jr, Fredman SM (1983) Role ofproprioceptive reflexes in control of feeding muscles of Aplysia.J Neurophysiol 49:1469–1480
Jing J, Weiss KR (2001) Neural mechanisms of motor programswitching in Aplysia. J Neurosci 21:7349–7362
Jordan R, Cohen KP, Kirk MD (1993) Control of intrinsic buccalmuscles by motoneurons B11, B15, and B16 in Aplysia cali-fornica. J Exp Zool 265:496–506
Kandel ER (1976) Cellular basis of behavior. Freeman, SanFrancisco
Kandel ER (1979) Behavioral biology of Aplysia. Freeman, SanFrancisco
Katz PS, Frost WN (1995) Intrinsic neuromodulation in theTritonia swim CPG: serotonin mediates both neuromodulationand neurotransmission by the dorsal swim interneurons.J Neurophysiol 74:2281–2294
Katz PS, Getting PA, Frost WN (1994) Dynamic neuromodulationof synaptic strength intrinsic to a central pattern generatorcircuit. Nature 367:729–731
Kirk MD (1989) Premotor neurons in the feeding system of Aplysiacalifornica. J Neurobiol 20:497–512
Kupfermann I (1974a) Feeding behavior in Aplysia: a simple sys-tem for the study of motivation. Behav Biol 10:1-26
Kupfermann I (1974b) Dissociation of the appetitive and con-summatory phases of feeding behavior in Aplysia: a lesionstudy. Behav Biol. 10:89–97
Kupfermann I (1994) Neural control of feeding. Curr OpinNeurobiol 4:869–876
Kupfermann I, Carew TJ (1974) Behavior patterns of Aplysiacalifornica in its natural environment. Behav Biol 12:317–337
Kupfermann I, Teyke T, Rosen SC, Weiss KR (1991) Studies ofbehavioral state in Aplysia. Biol Bull 180:262–268
Kupfermann I, Deodhar D, Teyke T, Rosen SC, Nagahama T,Weiss KR (1992) Behavioral switching of biting and of directedhead turning in Aplysia: explorations using neural networkmodels. Acta Biol Hung 43:315–328
Kupfermann I, Brezina V, Cropper EC, Deodhar D, Vilim FS,Weiss KR (1997) Reconfiguration of the peripheral plant dur-ing various forms of feeding behaviors in the mollusc Aplysia.In: Stein PSG, Grillner S, Selverston AI, Stuart DG (eds)Neurons, networks, and motor behavior. MIT Press, Cam-bridge MA, pp 217–222
Longley RD, Longley AJ (1986) Serotonin immunoreactivity ofneurons in the gastropod Aplysia californica. J Neurobiol17:339–58
Mackey SL, Kandel ER, Hawkins RD (1989) Identified seroton-ergic neurons LCB1 and RCB1 in the cerebral ganglia ofAplysia produce presynaptic facilitation of siphon sensoryneurons. J Neurosci 9: 4227–4235
McPherson DR, Blankenship JE (1991a) Neural control ofswimming in Aplysia brasiliana. I. Innervation of parapodialmuscle by pedal ganglion motoneurons. J Neurophysiol66:1338–1351
McPherson DR, Blankenship JE (1991b) Neural control of swim-ming in Aplysia brasiliana. II. Organization of pedal moto-neurons and parapodial motor fields. J Neurophysiol 66:1352–1365
McPherson DR, Blankenship JE (1991c) Neural control of swim-ming in Aplysia brasiliana. III. Serotonergic modulatoryneurons. J Neurophysiol 66:1366–1379
Miller MW, Rosen SC, Schissel SL, Cropper EC, Kupfermann I,Weiss KR (1994) A population of SCP-containing neurons inthe buccal ganglion of Aplysia are radula mechanoafferents andreceive excitation of central origin. J Neurosci 14:7008–7023
Morgan PT, Perrins R, Lloyd PE, Weiss KR (2000) Intrinsic andextrinsic modulation of a single central pattern generating cir-cuit. J Neurophysiol 84:1186–1193
Morgan PT, Jing J, Vilim FS, Weiss KR (2002) Interneuronal andpeptidergic control of motor pattern switching in Aplysia.J Neurophysiol 87:49–61
Morton DW, Chiel HJ (1993) The timing of activity in motorneurons that produce radula movements distinguishes ingestionfrom rejection in Aplysia. J Comp Physiol A 173:519–536
Murphy AD (1990) An identified pleural ganglion interneuron in-hibits patterned motor activity in the buccal ganglia of the snail,Helisoma. Brain Res 525:300–303
Nagahama T, Weiss KR, Kupfermann I (1993) Effects of cerebralneuron C-PR on body postural muscles associated with afood-induced arousal state in Aplysia. J Neurophysiol70:1231–1243
Nagahama T, Weiss KR, Kupfermann I (1994) Body posturalmuscles active during food arousal in Aplysia are modulated bydiverse neurons that receive monosynaptic excitation from theneuron C-PR. J Neurophysiol 72:314–325
Norekian TP (1997) Coordination of startle and swimmingneural systems in the pteropod mollusk Clione limacina: roleof the cerebral cholinergic interneuron. J Neurophysiol78:308–320
Norekian TP, Satterlie RA (1976) Cerebral serotonergic neuronsreciprocally modulate swim and withdrawal neural networks inthe mollusk Clione limacina. J Neurophysiol 75:538–546
Ono JK, McCaman RE (1984) Immunocytochemical localizationand direct assays of serotonin-containing neurons in Aplysia.Neuroscience 11:549–560
Perrins R, Weiss KR (1998) Compartmentalization of informationprocessing in an Aplysia feeding circuit interneuron throughmembrane properties and synaptic interactions. J Neurosci18:3977–3989
Plummer MR, Kirk MD (1990) Premotor neurons B51 and B52 inthe buccal ganglia of Aplysia californica: synaptic connections,effects on ongoing motor rhythms, and peptide modulation.J Neurophysiol 63:539–558
Rock MK, Blankenship JE, Lebeda FJ (1977) Penis-retractormuscle of Aplysia: excitatory motor neurons. J Neurobiol8:569–579
Rosen SC, Weiss KR, Kupfermann I (1979) Response propertiesand synaptic connections of mechanoafferent neurons in cere-bral ganglion of Aplysia. J Neurophysiol 42:954–974
Rosen SC, Weiss KR, Cohen JL, Kupfermann I (1982) Intergangl-ionic cerebral-buccal mechanoafferents of Aplysia: receptivefields and synaptic connections to different classes of neuronsinvolved in feeding behavior. J Neurophysiol 48:271–288
243

Rosen SC, Weiss KR, Goldstein RS, Kupfermann I (1989) The roleof a modulatory neuron in feeding and satiation in Aplysia:effects of lesioning of the serotonergic metacerebral cells. JNeurosci 9:1562–1578
Rosen SC, Teyke T, Miller MW, Weiss KR, Kupfermann I (1991)Identification and characterization of cerebral-to-buccal inter-neurons implicated in the control of motor programs associatedwith feeding in Aplysia. J Neurosci 11:3630–3655
Rosen SC, Miller MW, Evans CG, Cropper EC, Kupfermann I(2000a) Diverse synaptic connections between peptidergic rad-ula mechanoafferent neurons and neurons in the feeding systemof Aplysia. J Neurophysiol 83:1605–1620
Rosen SC, Miller MW, Cropper EC, Kupfermann I (2000b) Out-puts of radula mechanoafferent neurons in Aplysia are modu-lated by motor neurons, interneurons, and sensory neurons.J Neurophysiol 83:1621–1636
Rosen SC, Friedman AK, Kupfermann I (2002) Grasp to rasp:Aplysia neurons involved in switching from rasping (grazing) tograsping (browsing) modes of feeding behavior. Soc NeurosciAbstr 67:27
Sanchez JA, Kirk MD (2000) Short-term synaptic enhancementmodulates ingestion motor programs of Aplysia. J Neurosci20:RC85
Sanchez JA, Kirk MD (2001) Cerebral-buccal pathways in Aplysiacalifornica: synaptic connections, cooperative interneuronal ef-fects and feedback during buccal motor programs. J CompPhysiol A 187:801–815
Saunders RW, Rosen SC, Miller MW, Kupfermann I (1998)Kinematics analysis of radula movements during biting andgrazing behavior in Aplysia and Bursatella. Soc Neuroci Abstr24:1892
Scott ML, Govind CK, Kirk MD (1991) Neuromuscular organi-zation of the buccal system in Aplysia californica. J CompNeurol 312: 207–222
Skelton ME, Koester J (1992) The morphology, innervation andneural control of the anterior arterial system of Aplysia cali-fornica. J Comp Physiol A 171:141–155
Sumbre G, Gutfreund Y, Fiorito G, Flash T, Hochner B (2001)Control of octopus arm extension by a peripheral motor pro-gram. Science 293:1845–1848
Susswein AJ, Byrne JH (1988) Identification and characterizationof neurons initiating patterned neural activity in the buccalganglia of Aplysia. J Neurosci 6:1513–1527
Teyke T, Weiss KR, Kupfermann I (1990) An identified neuron(CPR) evokes neuronal responses reflecting food arousal inAplysia. Science 247:85–87
Teyke T, Weiss KR, Kupfermann I (1991) Activity of an identifiedcerebral neuron correlates with food-induced arousal in Aply-sia. Neurosci Lett 133:307–310
Teyke T, Rosen SC, Weiss KR, Kupfermann I (1993) Dopamin-ergic neuron B20 generates rhythmic neuronal activity in thefeeding motor circuitry of Aplysia. Brain Res 630:226–237
Teyke T, Xin Y, Weiss KR, Kupfermann I (1997) Ganglionicdistribution of inputs and outputs of C-PR, a neuron involvedin the generation of a food-induced arousal state in Aplysia.Invert Neurosci 2:235–244
Walters ET, Byrne JH, Carew TJ, Kandel ER (1983) Mechanoaf-ferent neurons innervating tail of Aplysia. I. Response proper-ties and synaptic connections. J Neurophysiol 50:1522–1542
Weiss KR, Kupfermann I (1976) Homology of the giant seroton-ergic neurons (metacerebral cells) in Aplysia and pulmonatemolluscs. Brain Res 117:33–49
Weiss KR, Cohen JL, Kupfermann I (1978) Modulatory control ofbuccal musculature by a serotonergic neuron (metacerebral cell)in Aplysia. J Neurophysiol 41:181–203
Weiss KR, Koch UT, Koester J, Rosen SC, Kupfermann I (1982)The role of arousal in modulating feeding behavior of Aplysia:neural and behavioral studies. In: Hoebel BG, Novin D (eds)The neural basis of feeding and reward. Haer Institute,Brunswick, ME, pp 25–57
Weiss KR, Chiel HJ, Kupfermann I (1986a) Sensory function andgating of histaminergic neuron C2 in Aplysia. J Neurosci6:2416–2426
Weiss KR, Chiel HJ, Koch U, Kupfermann I (1986b) Activity ofan identified histaminergic neuron, and its possible role inarousal of feeding behavior in semi-intact Aplysia. J Neurosci6:2403–2415
Wright WG, Jones K, Sharp P, Maynard B (1995) Widespreadanatomical projections of the serotonergic modulatory neuron,CB1, in Aplysia. Invert Neurosci 1:173–183
Xin Y, Weiss KR, Kupfermann I (1996) An identified interneuroncontributes to aspects of six different behaviors in Aplysia.J Neurosci 16:5266–5279
Xin Y, Hurwitz I, Perrins R, Evans CG, Alexeeva V, Weiss KR,Kupfermann I (1999) Actions of a pair of identified cerebral-buccal interneurons (CBI-8/9) in Aplysia that contain the pep-tide myomodulin. J Neurophysiol 81:507–520
Xin Y, Koester J, Jing J, Weiss KR, Kupfermann I (2001) Cere-bral-abdominal interganglionic coordinating neurons inAplysia. J Neurophysiol 85:174–186
244
Copyright © 2022 FDOKUMEN