
Surface-structure-regulated cell-membrane penetration by monolayer-protected nanoparticles
15
Surface Structure-Regulated Cell Membrane Penetration by Monolayer Protected Nanoparticles Ayush Verma 1 , Oktay Uzun 1 , Yuhua Hu 2 , Ying Hu 1 , Hee-Sun Han 3 , Nicky Watson 4 , Suelin Chen 1 , Darrell J. Irvine 1,5,* , and Francesco Stellacci 1,* 1Department of Materials Science and Engineering, MIT 2 Department of Chemical Engineering, MIT 3Department of Chemistry, MIT 4Department of Biology, MIT 5Department of Biological Engineering, MIT Abstract Intracellular trafficking of nanoscale objects is regulated by cell membranes through interactions that are under intense investigation. These materials are typically internalized by cells into membrane- bounded endosomes and never gain access to the cell machinery located in the cytosol. While some bio-macromolecules can penetrate or fuse with cell membranes without overt membrane disruption, no synthetic material of comparable size has shown this property yet. Cationic nanoscale objects (mostly assisted by some degree of hydrophobicity) pass through cell membranes by generating transient holes, a process associated with cytotoxicity. To date, studies aimed at generating cell- penetrating nanomaterials have focused on the effect of size, shape, and composition. Here we compare membrane penetration by two nanoparticle ‘isomers’ with the same composition (same hydrophobic content), one coated with sub-nanometer striations of alternating anionic and hydrophobic groups, and the other coated with the same moieties but in a random distribution. We show that the former particles penetrate the plasma membrane without bilayer disruption while the latter were instead mostly trapped in endosomes. Our results highlight a new design rule for materials that interact with cells and offer a paradigm for analyzing the fundamental problem of cell membrane penetrating bio- and macro-molecules. We anticipate consequences for designing materials for biomedical applications and for understanding potential safety issues associated with synthetic nanomaterials. Introduction Nanomaterials are of great interest for use in biomedicine as imaging tools 1-3 , phototherapy agents 4,5 , and gene delivery carriers 6,7 . Their interactions with cell membranes (CM) are of central importance for all such applications. For example, many drug delivery systems are based on the transport of therapeutic agents to the cytosol or nucleus of cells by nanoparticles; efficient delivery must be achieved while avoiding cytotoxicity during passage through cell *Correspondence and requests for materials should be addressed to Darrell J. Irvine (E-mail: [email protected]) and Francesco Stellacci (E-mail: [email protected]).. This paper is dedicated to Anne Mayes in recognition of the mentor role she had for both corresponding authors. Competing Interest Statement The authors declare no competing financial interests. Supplementary Information accompanies the paper on www.nature.com/nature/naturematerials. NIH Public Access Author Manuscript Nat Mater. Author manuscript; available in PMC 2009 May 19. Published in final edited form as: Nat Mater. 2008 July ; 7(7): 588–595. doi:10.1038/nmat2202. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Surface-structure-regulated cell-membrane penetration by monolayer-protected nanoparticles

Surface Structure-Regulated Cell Membrane Penetration byMonolayer Protected Nanoparticles
Ayush Verma1, Oktay Uzun1, Yuhua Hu2, Ying Hu1, Hee-Sun Han3, Nicky Watson4, SuelinChen1, Darrell J. Irvine1,5,*, and Francesco Stellacci1,*
1Department of Materials Science and Engineering, MIT
2 Department of Chemical Engineering, MIT
3Department of Chemistry, MIT
4Department of Biology, MIT
5Department of Biological Engineering, MIT
AbstractIntracellular trafficking of nanoscale objects is regulated by cell membranes through interactions thatare under intense investigation. These materials are typically internalized by cells into membrane-bounded endosomes and never gain access to the cell machinery located in the cytosol. While somebio-macromolecules can penetrate or fuse with cell membranes without overt membrane disruption,no synthetic material of comparable size has shown this property yet. Cationic nanoscale objects(mostly assisted by some degree of hydrophobicity) pass through cell membranes by generatingtransient holes, a process associated with cytotoxicity. To date, studies aimed at generating cell-penetrating nanomaterials have focused on the effect of size, shape, and composition. Here wecompare membrane penetration by two nanoparticle ‘isomers’ with the same composition (samehydrophobic content), one coated with sub-nanometer striations of alternating anionic andhydrophobic groups, and the other coated with the same moieties but in a random distribution. Weshow that the former particles penetrate the plasma membrane without bilayer disruption while thelatter were instead mostly trapped in endosomes. Our results highlight a new design rule for materialsthat interact with cells and offer a paradigm for analyzing the fundamental problem of cell membranepenetrating bio- and macro-molecules. We anticipate consequences for designing materials forbiomedical applications and for understanding potential safety issues associated with syntheticnanomaterials.
IntroductionNanomaterials are of great interest for use in biomedicine as imaging tools1-3, phototherapyagents4,5, and gene delivery carriers6,7. Their interactions with cell membranes (CM) are ofcentral importance for all such applications. For example, many drug delivery systems arebased on the transport of therapeutic agents to the cytosol or nucleus of cells by nanoparticles;efficient delivery must be achieved while avoiding cytotoxicity during passage through cell
*Correspondence and requests for materials should be addressed to Darrell J. Irvine (E-mail: [email protected]) and Francesco Stellacci(E-mail: [email protected])..This paper is dedicated to Anne Mayes in recognition of the mentor role she had for both corresponding authors.Competing Interest StatementThe authors declare no competing financial interests.Supplementary Information accompanies the paper on www.nature.com/nature/naturematerials.
NIH Public AccessAuthor ManuscriptNat Mater. Author manuscript; available in PMC 2009 May 19.
Published in final edited form as:Nat Mater. 2008 July ; 7(7): 588–595. doi:10.1038/nmat2202.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

membranes to reach intracellular target compartments8,9. Indeed, membrane penetration bysynthetic10 as well as by biologically-derived11 molecules/particles is currently under intenseinvestigation. Some biomacromolecules, e.g., cell penetrating peptides (CPPs), may be capableof penetrating membranes without overt lipid bilayer disruption/poration12-15. Likewise,synthetic nanomaterials with very small dimensions (molecules, metal nanoclusters16, smalldendrimers17, and carbon nanotubes18) can also pass through CMs. However, to the best ofour knowledge, no synthetic material larger than a few nm in size can pass through membraneswithout disrupting the integrity of these biological barriers. For example, charged particles(e.g., cationic quantum dots or dendrimers) induce transient poration of cell membranes toenter cells, a process associated with cytotoxicity19. Alternatively, nanoparticles have beendesigned to explicitly disrupt endolysosomal membranes to enter the cell by force20 or enterthe cell aided by exogenous agents such as CPP chaperones21. By contrast, most nanoparticlesare trapped in endosomes22 and hence do not reach the cytosol.
The surface properties of nanomaterials play a critical role in determining the outcome of theirinteractions with cells23. Recently we found that when gold nanoparticles are coated withbinary mixtures of hydrophobic and hydrophilic organic molecules, ribbon-like domains ofalternating composition spontaneously form in the ligand shell24-26. These domains are onan average less than 6 Å wide, which is on the order of or even smaller than the distributionof chemical functionalities on biomolecules such as proteins and peptides. Ordered, rather thanrandom organization of the functional groups on these particle surfaces leads to unexpectedsurface properties due to the molecularly-close apposition of hydrophilic and hydrophobicmoieties at all points on the particle surfaces: the particles exhibit unusual solubility trends andlow protein binding even with high contents of hydrophobic groups24,25. With the orderedamphiphilic structure of some CPPs in mind, we explored the interaction of these nanoparticleswith living cells. Here we show that ∼6 nm nanoparticles, coated with a shell of hydrophobicand anionic ligands regularly arranged in ribbon-like domains of alternating composition (Fig.1), penetrate CMs at 37°C and 4°C without evidence of membrane disruption. Particles withidentical hydrophobic content but lacking structural order in the ligand shell for the most partdo not penetrate CMs, hence we conclude that the structural organization of surface chemicalgroups plays a key role in regulating CM penetration.
To create amphiphilic nanoparticles, we utilized mixed monolayer-protected goldnanoparticles27 surrounded by a self-assembled monolayer of organic ligands that determinesthe particles’ interactions with the external environment. They can resemble biomolecules andbiomolecular assemblies in terms of size and chemical composition and sometimes function(e.g., enzymatic activity)28,29, but their tightly-packed ligand shell25 lacks the structuralflexibility of proteins or peptide assemblies and instead is composed of an ordered layer ofsurface functional groups. Simple reactions allow for the synthesis of gold particles coatedwith single as well as multi-component ligand shells, whose composition and morphology canbe easily and precisely controlled30. Utilizing the unique physical chemistry offered by thissystem, we created water-soluble particles with ordered or disordered amphiphilic ligandshells31 to investigate CM penetration of nanoparticles as a function of the spatial distributionof chemical groups.
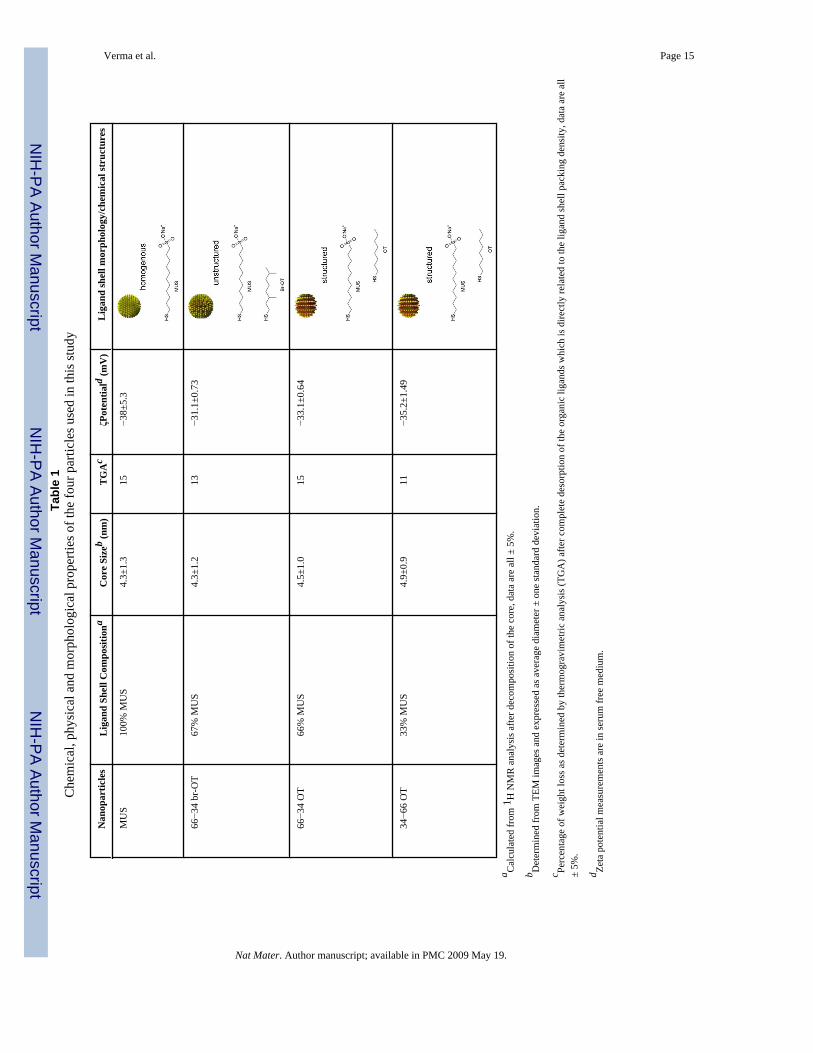
Four types of ligand-coated gold nanoparticles were synthesized32 and studied (Table 1):particles were coated with (i) 11-mercapto-1-undecanesulfonate (MUS), (ii) a 2:1 molarmixture of MUS and 1-octanethiol (OT) [hereafter referred to as 66−34 OT], (iii) a 1:2 molarmixture of MUS and OT [34−66 OT], and (iv) a 2:1 molar mixture MUS: 3,7 dimethyl octane1-thiol (br-OT) [66−34 br-OT], respectively. We used sulfonate groups because they impartedremarkable water solubility to all the particles studied irrespective of the fraction ofhydrophobic ligands used, with saturation concentrations in water in excess of 250 mg/mL31, orders of magnitude larger than the concentrations used here. 1H NMR was used to
Verma et al. Page 2
Nat Mater. Author manuscript; available in PMC 2009 May 19.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

confirm the absence of unbound ligands and the particle ligand shell composition wasdetermined after decomposing the gold core with iodine33. The size distributions, ligand shellpacking densities, and zeta potentials of the four particles studied had negligible differences(Table 1) (Supplementary Information Online, SIO). Thus, these particles had nearly identicalphysical characteristics, except for the composition and structure of the ligand shell, asdetermined by extensive scanning tunnelling microscopy (STM) studies (SIO) andschematically illustrated in Table 1. MUS particles had a homogeneous hydrophilic ligandshell, while 34−66 OT and 66−34 OT particles had a hydrophilic-hydrophobic striated ligandshell; 66−34 br-OT particles had a hydrophilic-hydrophobic disordered ligand shell. The lattertwo particles had nearly identical hydrophilic to hydrophobic ratio, the key difference residingin the ordering of hydrophilic and hydrophobic moieties of the ligand shell.
To determine how the nature and organization of functional groups at the surface of thesenanoparticles influenced their interactions with cell membranes, we labelled the particles withan average upper limit of 14 molecules of thiolated BODIPY dye (BODIPY-SH) pernanoparticle (SIO) and performed confocal fluorescence microscopy studies of their uptakeand intracellular distribution in living cells. A mouse dendritic cell (DC) clone DC2.434 wasutilized for these studies, representing a key class of phagocytes guarding all peripheral tissuesand particularly the skin, lungs, and gut mucosa. DCs are professional antigen presenting cells,and actively sample particles/fluid/macromolecules from their environment (via phagocytosis,macropinocytosis, and endocytosis) as part of their normal function. Macropinocytosis, anactin-dependent process where the cell folds membrane around a small volume of fluid, allowsthese cells to even internalize particles that have no interaction with the cell membrane intointracellular vesicles. To avoid potential artifacts of cell fixation35, cells were imaged live ina temperature-controlled environmental chamber. DCs were incubated with nanoparticles (0.2mg/mL) in serum-containing or serum-free medium for 3 h, washed, and imaged live byconfocal microscopy. Fig. 1 illustrates the difference in behaviour between the various typesof nanoparticles. MUS nanoparticles bearing only the hydrophilic sulfonate ligand wereinternalized by DCs at 37 °C and displayed punctate fluorescence patterns indicative ofendosomal uptake (Fig. 1a), in agreement with previous findings for most types of syntheticnanoparticles22,36,37. 66−34 br-OT nanoparticles bearing a disordered mixed monolayer withsulfonate and methyl headgroups also showed a punctate distribution in DCs, though a lowlevel of background fluorescence in the cytosol of some cells was also detectable (Fig. 1b). Incontrast, 66−34 OT nanoparticles, which have an identical ratio of surface sulfonate and methylgroups with respect to 66−34 br-OT nanoparticles but in a ‘striped’ nanoscale organization,were detected in cells as a diffuse pattern of intracellular fluorescence overlaid on punctatesites of brighter fluorescence, suggesting passage of a fraction of these particles into the cytosol(Fig. 1c). A similar behaviour was observed for 34−66 OT nanoparticles (Supplementary Fig.3). Trends in nanoparticle uptake and intracellular distribution were qualitatively similar inserum-free and serum-containing medium, although a greater amount of cytosol-localizednanoparticles and fewer nanoparticles contained in vesicular structures were readily observedfor the serum-free condition (Supplementary Fig. 4). The similarity in the nanoparticle uptakein serum–containing and –free conditions is suggestive that ‘striped’ particles are resistant tonon-specific protein adsorption (as previously observed in similar systems)24, and hence retaintheir behaviour independent of the incubation medium. To confirm this assumption, we usedDynamic Light Scattering (DLS) to evaluate particle sizes in solution before and afterincubation in serum-containing medium. The radii obtained before incubation were all in goodagreement with TEM core size data, further confirming that the particles were well dispersedin solution. The particles were then incubated in serum-containing medium and separated fromunbound proteins using Gel Filtration Chromatography (GFC). The GFC traces showed that alarge fraction of the 66−34 OT nanoparticles retained their original size after the incubation,while the 66−34 br-OT and the MUS particles showed an increase in particle size (data notshown). In fact, the DLS data of the ‘purified’ particles following serum exposure showed an
Verma et al. Page 3
Nat Mater. Author manuscript; available in PMC 2009 May 19.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

increase in size for the non-striped particles and almost no change for the striped ones. Theincrease in size for the non-striped particles must be ascribed to non-specific protein adsorption,as TEM studies after incubation show nearly identical core size distribution (see Table 1, SIO).
To further explore the mechanism by which these particles entered the cells, we repeated theseinternalization experiments at 4 °C to inhibit mechanisms of active uptake by the cells. At thistemperature, endocytic/pinocytic uptake of tracer molecules such as calcein or labelleddextrans is completely blocked in DC2.4 cells20. BODIPY channel fluorescence following theincubation of cells with MUS or 66−34 br-OT particles at 4 °C (Fig. 1d and e, respectively)was indistinguishable from the low levels of autofluorescence observed within untreatedcontrol cells (Supplementary Fig. 5a), while 66−34 OT (Fig. 1f) and 34−66 OT nanoparticles(Supplementary Fig. 5e) still appeared to enter cells at substantial levels, taking on a diffusepattern of fluorescence consistent with cytosolic localization. A quantitative comparison ofcell-associated fluorescence at 4 °C is shown in Fig. 1g. The fluorescence of cells incubatedwith MUS or 66−34 br-OT particles was not statistically different from the backgroundautofluorescence of untreated cells, while the fluorescence of cells treated with ‘striped’nanoparticles was 8-fold greater than the background. All of the nanoparticles studied herewere uniformly excluded from the nucleus of cells at both 4 °C and 37 °C. The entry of theseparticles into the cytosol under conditions where endocytic processes are blocked suggests thatthe ‘striped’ nanoparticles are capable of directly passing through the plasma membrane of thecells, while 66−34 br-OT nanoparticles with an identical ratio of hydrophilic:hydrophobicmoieties but lacking the ordered arrangement of these groups do not penetrate. To further testthis possibility and rule out potential changes in membrane structure/fluidity induced byculturing the cells at 4 °C, we incubated DCs with nanoparticles at 37 °C in the presence ofsodium azide and 2-deoxyglucose, which block active forms of uptake such as endocytosis.To confirm the efficacy of the inhibitors, DCs were treated with the inhibitors and incubatedwith calcein, a small molecule membrane-impermeable fluorophore normally taken up byendocytosis/pinocytosis. After 20 min, control DCs cultured with calcein in the absence ofazide/deoxyglucose exhibited readily detectable endocytic vesicles of calcein throughoutconfocal z-sections (Fig. 2a). In contrast, DCs incubated with the inhibitors and calcein showedno detectable internalized dye in 3D confocal z-sections (Fig. 2b and data not shown). WhenDCs were incubated with both calcein and striped OT:MUS particles at 37 °C in the presenceof the internalization inhibitors, nanoparticle entry into the cytosol was readily detected after20 min incubation with the particles, while calcein was undetectable in the cells (Fig 2b). MUSparticles and br-OT nanoparticles failed to enter DCs under these conditions (data not shown).Thus, nanoparticles with an ordered surface ligand layer were found to access the cytosol at37 °C under conditions where no evidence for active uptake processes such as endocytosiscould be detected. Finally, consistent with an internalization mechanism involving directinteraction with the plasma membrane rather than a slower endocytic/pinocytic active uptakeroute, kinetic experiments showed that striped nanoparticles were readily detectable in cellswithin 5 min in the presence of azide/deoxyglucose (data not shown).
Two limitations of the confocal analysis are the resolution of optical microscopy and thepossibility of artifacts in the apparent subcellular distribution arising from the release of freedye from particles. Prior studies have shown that organic thiol ligands can be displaced fromgold nanoparticles by exchange reactions with cytosolic glutathione38, which is the mostabundant intracellular thiol39. This effect seems unlikely given that glutathione-mediateddisplacement has been shown to require timescales on the order of days40, compared with thepresent experiments carried out over 3 hours. However, to unequivocally interpret the confocalimages and also to rule out any possible effect of the fluorophore molecules attached to thenanoparticles, we performed an extensive TEM analysis of both fluorescently labelled andunlabelled nanoparticles taken up in cells. DC2.4 cells were incubated for 3 h with nanoparticlesas before, washed, fixed, and sectioned for TEM imaging (SIO). Several observations resulted
Verma et al. Page 4
Nat Mater. Author manuscript; available in PMC 2009 May 19.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

from analysis of TEM images. No significant difference in overall trends was observed forparticles either labelled with or without BODIPY-SH (Supplementary Fig. 13). Theintracellular distribution of each of the 4 types of ligand-coated nanoparticles studied was inqualitative agreement with the trends observed by confocal microscopy, as illustrated by theseries of TEM images in Fig. 3 for the case of unlabeled particles. MUS nanoparticles werealmost exclusively found in endosomal compartments (Fig. 3a); a similar distribution wasobserved for 66−34 br-OT nanoparticles (Fig. 3b). ‘Striped’ nanoparticles, in contrast, werereadily detected outside of defined membrane-enclosed compartments (Fig. 3c). As shown inFig. 3d, an enumeration of the number of free particles/μm3 localized outside of membrane-enclosed compartments revealed a higher density of cytosolic 66−34 and 34−66 OTnanoparticles as compared to the MUS and 66−34 br-OT particles. Interestingly, TEM imagesand examination of the number of cytosol localized particles/μm3 obtained from serum-containing condition displayed similar trends, suggesting that the differences in particlebehaviour are not mediated by differentially-adsorbed serum proteins41 (Supplementary Fig.12). Figure 4 shows high resolution TEM images that display nanoparticles very close to thecell membrane, probably frozen ‘in the act’ of passing through it.
Prior studies have shown that some cationic nanoparticles and macromolecules penetrate theplasma membrane of cells by generating transient holes17; in contrast, some data suggests thata subset of biological membrane-penetrating macromolecules (e.g., CPPs) may be capable ofpenetrating membranes without causing overt membrane poration12-15. The formermechanism can lead to cell death due to loss of membrane polarization and/or leakage of ionsand molecules into/out of the cell. To establish whether ‘striped’ nanoparticles penetrate cellmembranes through creation of transient holes, we tested whether the internalization of ligand-protected nanoparticles was accompanied by escape of cytosol-localized tracer dye orconversely, cytosolic entry of an initially extracellular tracer dye42,43. First, DC2.4 cells wereco-incubated with nanoparticles and calcein; calcein remains in an endolysosomalcompartments in cells unless it is co-internalized with a membrane-disrupting agent44. Cellswere incubated for 3 h with calcein and nanoparticles, washed, and imaged live by confocalmicroscopy; the localization of calcein in individual cells was then scored (endosomal vs.cytosolic). As a positive control, DCs were incubated with cationic gold nanoparticles bearing11-mercaptoundecane-tetramethylammonium chloride (TMA) ligands, which would beexpected to disrupt cell membranes based on previously published work10. As shown in Fig.5a and c, TMA nanoparticles caused calcein to escape to the cytosol of ∼50% of the DCs,whereas DCs incubated with calcein alone exhibited a solely punctate endosomal distributionof dye. Strikingly, 66−34 OT (Fig. 5a, b) and 34−66 OT nanoparticles (Supplementary Fig. 6)(both fluorescently labelled and unlabelled) did not promote calcein entry into the cytosol,despite the data described above showing that these particles clearly access the cytosoliccompartment. We next tested whether ‘striped’ nanoparticles caused an escape of tracermolecules from the cytosol of cells during uptake: DCs were pre-loaded with calcein-AM, anacetomethoxy form of the dye that is trapped in the cytosol by conversion from a membrane-permeable to –impermeable form by intracellular esterases. Again, 66−34 OT particles enteredcells without allowing any measurable leakage of calcein from the cells (Supplementary Fig.7). To further, albeit indirectly, confirm that ‘striated’ particles penetrate cell membraneswithout causing substantial membrane disruption, we measured their cytotoxicity relative tocationic TMA-coated particles using a colony-forming assay. DC2.4 cells were incubated withnanoparticles for 3 h, washed, replated at equal densities and allowed to grow for 24 h. Wefound that cells treated with 66−34 and 34−66 OT nanoparticles survived/proliferated at a levelthat was ∼ 90% of untreated control cells, while only 34% were recovered following treatmentwith TMA-coated particles (Supplementary Fig. 9). Similar results were obtained whenstudying immediate toxicity of cells with propidium iodide (PI) staining and MTT assay (Fig.10 and 11 SIO).
Verma et al. Page 5
Nat Mater. Author manuscript; available in PMC 2009 May 19.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

In these studies, we consistently observed no nanoparticle nuclear uptake, as well as noevidence for escape of nanoparticles from cells for up to 6 h after internalization occurred (datanot shown). We speculate that these two phenomena could be related; it is possible that whenparticles are in the cytosol they lose the delicate compositional balance and structural motifthat is needed to permeate cell membranes. However, given the unique structure of the nucleardouble membrane, further studies need to be performed to fully understand these observations.Moreover, to test the generality of our results, we repeated 37 °C and 4 °C uptake experimentswith a second common cell line, mouse embryonic fibroblasts (MEFs). The same trends inparticle uptake, with ‘striped’ particles accessing the cytosol, were observed (SupplementaryFig. 8).
Our results highlight the importance of structure in the interactions of nanoparticles with CMs.We synthesized two nanoparticles that could be loosely characterized as the nanoscaleequivalent of chemical isomers. They have the same size and shape, and are coated with thesame ratio of hydrophobic to hydrophilic molecules, the latter being identical (MUS) while theformer differing slightly (OT vs br-OT) in order to achieve different ligand shell structures.These two particles (66−34 OT and 66−34 br-OT) showed very similar solubility and wettingbehaviour as highlighted by nearly identical zeta potentials. They differed only in the moleculararrangement of surface chemical groups: 66−34 OT displaying a striated ligand organizationand 66−34 br-OT, lacking this structured arrangement. This seemingly small difference led todrastically different behaviour in CM penetration properties. Confocal studies of cell uptakeof these particles (dye functionalized to the same extent) in DC2.4 cells at 37°C (in both serum-containing and serum-free conditions) showed a pronounced punctate fluorescence distributionpattern indicative of endosomal entrapment for 66−34 br-OT nanoparticles, while, 66−34 OTparticles displayed a more diffuse fluorescence pattern indicating escape to the cytosol. Thesedifferences in intracellular localization were confirmed through TEM studies, which were alsoused to rule out any effect of the presence of the dye in the ligand shell. Nanoparticle uptakeexperiments at 4°C were used to prove that the 66−34 OT nanoparticles can directly passthrough cell membranes and do not require internalization by active endocytosis or pinocytosisto reach the cytosol, while at 4°C 66−34 br-OT are almost completely blocked from cell entry.This was additionally confirmed by detection of nanoparticle entry into cells within 5 mindespite actively blocking endocytosis by treating the cells with sodium azide and 2-deoxyglucose (marked by no calcein uptake). Finally we determined that 66−34 OTnanoparticles pass through cell membranes without causing overt membrane poration viastudies with tracer dyes (no noticeable leakage) and cytotoxicity assays (minimal toxicity).This is in stark contrast with cationic nanoparticles, that are known to penetrate cell membranesby creating pores17 and induce cytotoxicity19. At the moment we can only speculate on themechanism that leads to CM penetration. A possible explanation is that the ‘rigid’ arrangementof the amphiphilic domains on the nanoparticles permits non-disruptive fusion of thenanoparticle with cell membranes (a more fluid mixed layer) and subsequent penetrationthrough the bilayer, in a manner reminiscent of some models for entry of cell penetratingpeptides.
In conclusion, we believe that by highlighting the role of ligand structure and order in CMpenetration by nanoparticles, this work will shed new light on the understanding of themolecular interactions that lead to cell membrane permeation, certainly on nanoparticles,possibly on larger organic particles and hopefully in CPPs. Future work, besides thefundamental studies highlighted above, includes the engineering of novel and more efficientlow-toxicity drug delivery systems based on the design rules for membrane penetrationidentified here and on the analysis of potential safety issues associated with syntheticnanomaterials45.
Verma et al. Page 6
Nat Mater. Author manuscript; available in PMC 2009 May 19.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgementsThis work was supported by the National Science Foundation (CAREER Award) and the NHLBI TPEN program(U01-HL080731). F.S. is grateful to the Packard Foundation for their generous award.
References1. Weissleder R. Molecular imaging in cancer. Science 2006;312:1168–1171. [PubMed: 16728630]2. Somers RC, Bawendi MG, Nocera DG. CdSe nanocrystal based chem-/bio-sensors. Chem. Soc. Rev
2007;36:579–591. [PubMed: 17387407]3. Lewin M, et al. Tat peptide-derivatized magnetic nanoparticles allow in vivo tracking and recovery of
progenitor cells. Nat. Biotechnol 2000;18:410–414. [PubMed: 10748521]4. West JL, Halas NJ. Engineered nanomaterials for biophotonics applications: Improving sensing,
imaging, and therapeutics. Annu. Rev. Biomed. Eng 2003;5:285–292. [PubMed: 14527314]5. El-Sayed IH, Huang XH, El-Sayed MA. Selective laser photo-thermal therapy of epithelial carcinoma
using anti-EGFR antibody conjugated gold nanoparticles. Cancer Lett 2006;239:129–135. [PubMed:16198049]
6. Rosi NL, Giljohann DA, Thaxton CS, Lytton-Jean AKR, Han MS, Mirkin CA. Oligonucleotide-modified gold nanoparticles for intracellular gene regulation. Science 2006;312:1027–1030. [PubMed:16709779]
7. Han G, et al. Light-regulated release of DNA and its delivery to nuclei by means of photolabile goldnanoparticles. Angew. Chem.-Int. Edit 2006;45:3165–3169.
8. Allen TM, Cullis PR. Drug delivery systems: Entering the mainstream. Science 2004;303:1818–1822.[PubMed: 15031496]
9. Choleris E, Little SR, Mong JA, Puram SV, Langer R, Pfaff DW. Microparticle-based delivery ofoxytocin receptor antisense DNA in the medial amygdala blocks social recognition in female mice.Proc. Natl. Acad. Sci. U. S. A 2007;104:4670–4675. [PubMed: 17360582]
10. Leroueil PR, Hong SY, Mecke A, Baker JR, Orr BG, Holl MMB. Nanoparticle interaction withbiological membranes: Does nanotechnology present a janus face? Accounts Chem. Res2007;40:335–342.
11. Zorko M, Langel U. Cell-penetrating peptides: mechanism and kinetics of cargo delivery. Adv. DrugDeliv. Rev 2005;57:529–545. [PubMed: 15722162]
12. Herbig ME, Assi F, Textor M, Merkle HP. The cell penetrating peptides pVEC and W2-pVEC inducetransformation of gel phase domains in phospholipid bilayers without affecting their integrity.Biochemistry 2006;45:3598–3609. [PubMed: 16533042]
13. Takeuchi T, et al. Direct and rapid cytosolic delivery using cell-penetrating peptides mediated bypyrenebutyrate. ACS Chem. Biol 2006;1:299–303. [PubMed: 17163758]
14. Thoren PEG, Persson D, Isakson P, Goksor M, Onfelt A, Norden B. Uptake of analogs of penetratin,Tat(48−60) and oligoarginine in live cells. Biochem. Biophys. Res. Commun 2003;307:100–107.[PubMed: 12849987]
15. Patel LN, Zaro JL, Shen WC. Cell penetrating peptides: Intracellular pathways and pharmaceuticalperspectives. Pharm. Res 2007;24:1977–1992. [PubMed: 17443399]
16. Yu J, Patel SA, Dickson RM. In vitro and intracellular production of peptide-encapsulated fluorescentsilver nanoclusters. Angew. Chem.-Int. Edit 2007;46:2028–2030.
17. Leroueil P, Hong S, Mecke A, Baker JR, Orr BG, Banaszak Holl MM. Nanoparticle Interaction withBiological Membranes: Does Nanotechnology Present a Janus Face? . Accounts Chem. Res. 2007
18. Kostarelos K, et al. Cellular uptake of functionalized carbon nanotubes is independent of functionalgroup and cell type. Nature Nanotechnology 2007;2:108–113.
Verma et al. Page 7
Nat Mater. Author manuscript; available in PMC 2009 May 19.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

19. Lovric J, Bazzi HS, Cuie Y, Fortin GRA, Winnik FM, Maysinger D. Differences in subcellulardistribution and toxicity of green and red emitting CdTe quantum dots. J. Mol. Med 2005;83:377–385. [PubMed: 15688234]
20. Hu Y, et al. Cytosolic Delivery of Membrane-Impermeable Molecules in Dendritic Cells Using pH-Responsive Core-Shell Nanoparticles. Nano Letters 2007;7:3056–3064. [PubMed: 17887715]
21. Tkachenko AG, et al. Multifunctional gold nanoparticle-peptide complexes for nuclear targeting. J.Am. Chem. Soc 2003;125:4700–4701. [PubMed: 12696875]
22. Shukla R, Bansal V, Chaudhary M, Basu A, Bhonde RR, Sastry M. Biocompatibility of goldnanoparticles and their endocytotic fate inside the cellular compartment: A microscopic overview.Langmuir 2005;21:10644–10654. [PubMed: 16262332]
23. Colvin V. Correlating the physical and chemical properties of nanoparticles to their biological activity.Environ. Mol. Mutagen 2007;48:533–533.
24. Jackson AM, Myerson JW, Stellacci F. Spontaneous assembly of subnanometre-ordered domains inthe ligand shell of monolayer-protected nanoparticles. Nat. Mater 2004;3:330–336. [PubMed:15098025]
25. Jackson AM, Hu Y, Silva PJ, Stellacci F. From homoligand- to mixed-ligand-monolayer-protectedmetal nanoparticles: A scanning tunneling microscopy investigation. J. Am. Chem. Soc2006;128:11135–11149. [PubMed: 16925432]
26. DeVries GA, et al. Divalent metal nanoparticles. Science 2007;315:358–361. [PubMed: 17234943]27. Daniel MC, Astruc D. Gold nanoparticles: Assembly, supramolecular chemistry, quantum-size-
related properties, and applications toward biology, catalysis, and nanotechnology. Chem. Rev2004;104:293–346. [PubMed: 14719978]
28. Manea F, Houillon FB, Pasquato L, Scrimin P. Nanozymes: Gold-nanoparticle-basedtransphosphorylation catalysts. Angew. Chem.-Int. Edit 2004;43:6165–6169.
29. Kisailus D, Najarian M, Weaver JC, Morse DE. Functionalized gold nanoparticles mimic catalyticactivity of a polysiloxane-synthesizing enzyme. Adv. Mater 2005;17:1234–+.
30. Templeton AC, Wuelfing MP, Murray RW. Monolayer protected cluster molecules. Accounts Chem.Res 2000;33:27–36.
31. Uzun O, Hu Y, Verma A, Chen S, Centrone A, Stellacci F. Water-soluble amphiphilic goldnanoparticles with structured ligand shells. Chem. Commun. 2008
32. Kang SY, Kim K. Comparative study of dodecanethiol-derivatized silver nanoparticles prepared inone-phase and two-phase systems. Langmuir 1998;14:226–230.
33. Tan H, Zhan T, Fan WY. Direct Functionalization of the Hydroxyl Group of the 6-Mercapto-1-hexanol (MCH) Ligand Attached to Gold Nanoclusters. J. Phys. Chem. B 2006;110:21690–21693.[PubMed: 17064127]
34. Shen ZH, Reznikoff G, Dranoff G, Rock KL. Cloned dendritic cells can present exogenous antigenson both MHC class I and class II molecules. J. Immunol 1997;158:2723–2730. [PubMed: 9058806]
35. Richard JP, et al. Cell-penetrating peptides - A reevaluation of the mechanism of cellular uptake. J.Biol. Chem 2003;278:585–590. [PubMed: 12411431]
36. Parak WJ, et al. Cell motility and metastatic potential studies based on quantum dot imaging ofphagokinetic tracks. Adv. Mater 2002;14:882–885.
37. Wilhelm C, Gazeau F, Roger J, Pons JN, Bacri JC. Interaction of anionic superparamagneticnanoparticles with cells: Kinetic analyses of membrane adsorption and subsequent internalization.Langmuir 2002;18:8148–8155.
38. Verma A, Simard JM, Worrall JWE, Rotello VM. Tunable reactivation of nanoparticle-inhibited beta-galactosidase by glutathione at intracellular concentrations. J. Am. Chem. Soc 2004;126:13987–13991. [PubMed: 15506760]
39. Meister A, Anderson ME. Glutathione. Annu. Rev. Biochem 1983;52:711–760. [PubMed: 6137189]40. Hong R, Han G, Fernandez JM, Kim BJ, Forbes NS, Rotello VM. Glutathione-mediated delivery and
release using monolayer protected nanoparticle carriers. J. Am. Chem. Soc 2006;128:1078–1079.[PubMed: 16433515]
41. Cedervall T, et al. Detailed identification of plasma proteins adsorbed on copolymer nanoparticles.Angew. Chem.-Int. Edit 2007;46:5754–5756.
Verma et al. Page 8
Nat Mater. Author manuscript; available in PMC 2009 May 19.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

42. Hong SP, et al. Interaction of polycationic polymers with supported lipid bilayers and cells: Nanoscalehole formation and enhanced membrane permeability. Bioconjugate Chem 2006;17:728–734.
43. Hong SP, et al. Interaction of poly(amidoamine) dendrimers with supported lipid bilayers and cells:Hole formation and the relation to transport. Bioconjugate Chem 2004;15:774–782.
44. Zhang LJ, Rozek A, Hancock REW. Interaction of cationic antimicrobial peptides with modelmembranes. J. Biol. Chem 2001;276:35714–35722. [PubMed: 11473117]
45. Nel A, Xia T, Madler L, Li N. Toxic potential of materials at the nanolevel. Science 2006;311:622–627. [PubMed: 16456071]
Verma et al. Page 9
Nat Mater. Author manuscript; available in PMC 2009 May 19.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
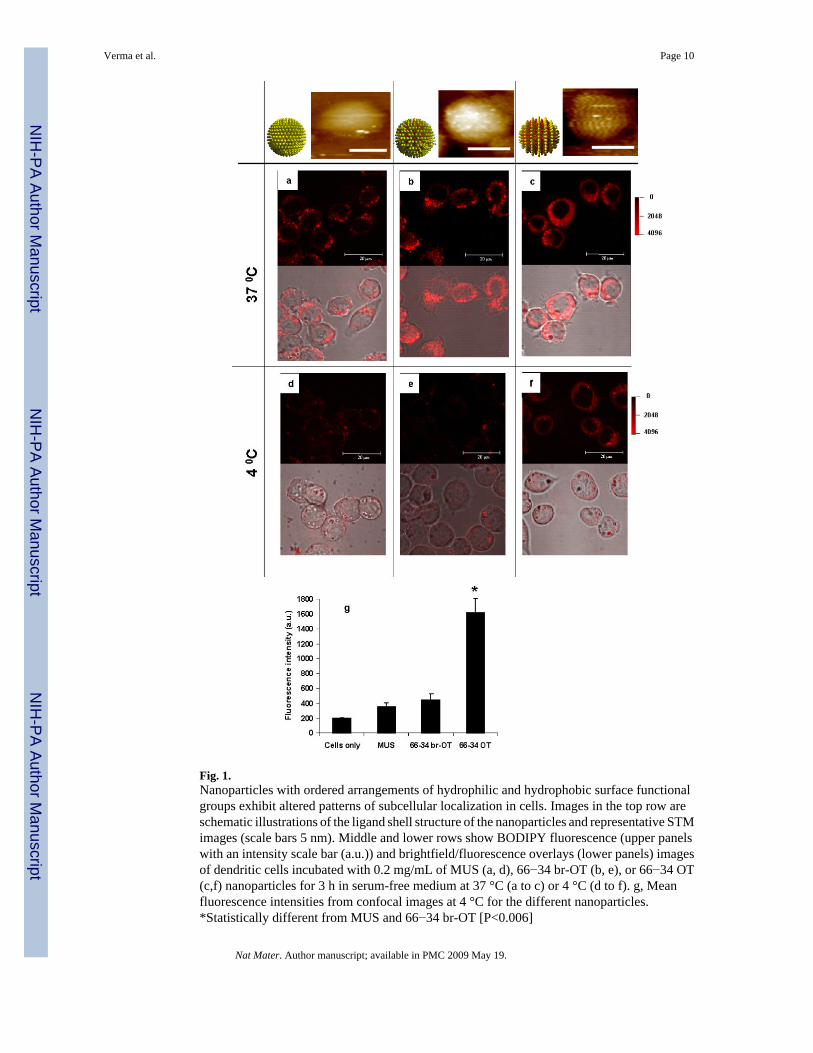
Fig. 1.Nanoparticles with ordered arrangements of hydrophilic and hydrophobic surface functionalgroups exhibit altered patterns of subcellular localization in cells. Images in the top row areschematic illustrations of the ligand shell structure of the nanoparticles and representative STMimages (scale bars 5 nm). Middle and lower rows show BODIPY fluorescence (upper panelswith an intensity scale bar (a.u.)) and brightfield/fluorescence overlays (lower panels) imagesof dendritic cells incubated with 0.2 mg/mL of MUS (a, d), 66−34 br-OT (b, e), or 66−34 OT(c,f) nanoparticles for 3 h in serum-free medium at 37 °C (a to c) or 4 °C (d to f). g, Meanfluorescence intensities from confocal images at 4 °C for the different nanoparticles.*Statistically different from MUS and 66−34 br-OT [P<0.006]
Verma et al. Page 10
Nat Mater. Author manuscript; available in PMC 2009 May 19.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 2.‘Striped’ nanoparticles enter cells at 37°C under conditions where active internalizationprocesses are blocked. DCs were incubated for 20 min at 37°C in serum-free medium withcalcein (a) or calcein and 66−34 OT nanoparticles in the presence of azide and 2-deoxyglucoseto block active internalization (b). Shown are confocal optical z-sections in 2 μm intervalstaken through the midplane of cells. Upper panels show nanoparticle fluorescence (red), middlepanels calcein fluorescence (green) and the lower panels show fluorescence/brightfieldoverlays.
Verma et al. Page 11
Nat Mater. Author manuscript; available in PMC 2009 May 19.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
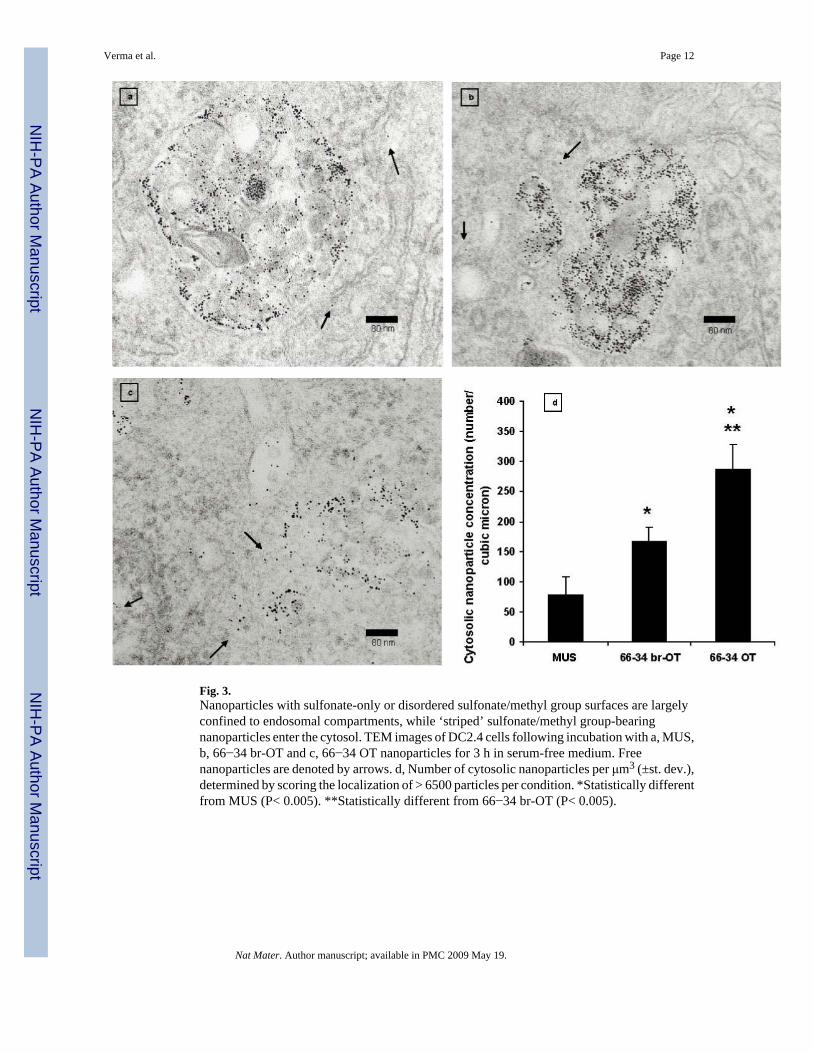
Fig. 3.Nanoparticles with sulfonate-only or disordered sulfonate/methyl group surfaces are largelyconfined to endosomal compartments, while ‘striped’ sulfonate/methyl group-bearingnanoparticles enter the cytosol. TEM images of DC2.4 cells following incubation with a, MUS,b, 66−34 br-OT and c, 66−34 OT nanoparticles for 3 h in serum-free medium. Freenanoparticles are denoted by arrows. d, Number of cytosolic nanoparticles per μm3 (±st. dev.),determined by scoring the localization of > 6500 particles per condition. *Statistically differentfrom MUS (P< 0.005). **Statistically different from 66−34 br-OT (P< 0.005).
Verma et al. Page 12
Nat Mater. Author manuscript; available in PMC 2009 May 19.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 4.‘Striped’ particles visualized at different stages of crossing the cell membrane of DCs. TEMimages of a-c, 66−34 OT and d-f, 34−66 OT incubated with cells in serum-free media at 37 °C. The arrows indicate few of the particles seen intracellular to the cell membrane.
Verma et al. Page 13
Nat Mater. Author manuscript; available in PMC 2009 May 19.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 5.‘Striped’ nanoparticles penetrate cell membranes without generating transient holes in contrastwith the known behaviour of cationic nanoparticles. a, Percentage of total cells showingcytosolic distribution of calcein. At least 100 cells were sampled for each particle and thecontrol. Confocal images showing calcein uptake and intracellular distribution in the presenceof b, 66−34 OT or c, TMA nanoparticles.
Verma et al. Page 14
Nat Mater. Author manuscript; available in PMC 2009 May 19.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Verma et al. Page 15Ta
ble
1C
hem
ical
, phy
sica
l and
mor
phol
ogic
al p
rope
rties
of t
he fo
ur p
artic
les u
sed
in th
is st
udy
Nan
opar
ticle
sL
igan
d Sh
ell C
ompo
sitio
naC
ore
Size
b (nm
)T
GA
cζP
oten
tiald (m
V)
Lig
and
shel
l mor
phol
ogy/
chem
ical
stru
ctur
es
MU
S10
0% M
US
4.3±
1.3
15−3
8±5.
3
66−3
4 br
-OT
67%
MU
S4.
3±1.
213
−31.
1±0.
73
66−3
4 O
T66
% M
US
4.5±
1.0
15−3
3.1±
0.64
34−6
6 O
T33
% M
US
4.9±
0.9
11−3
5.2±
1.49
a Cal
cula
ted
from
1H
NM
R a
naly
sis a
fter d
ecom
posi
tion
of th
e co
re, d
ata
are
all ±
5%
.
b Det
erm
ined
from
TEM
imag
es a
nd e
xpre
ssed
as a
vera
ge d
iam
eter
± o
ne st
anda
rd d
evia
tion.
c Perc
enta
ge o
f wei
ght l
oss a
s det
erm
ined
by
ther
mog
ravi
met
ric a
naly
sis (
TGA
) afte
r com
plet
e de
sorp
tion
of th
e or
gani
c lig
ands
whi
ch is
dire
ctly
rela
ted
to th
e lig
and
shel
l pac
king
den
sity
, dat
a ar
e al
l±
5%.
d Zeta
pot
entia
l mea
sure
men
ts a
re in
seru
m fr
ee m
ediu
m.
Nat Mater. Author manuscript; available in PMC 2009 May 19.




















