
Rigorous calculation method for resonance frequencies in ...

ARTICLE IN PRESS
Microbiological Research 161 (2006) 187—202
KEYWORDAnabaena;Akinete;Genetic vaHeterocysMolecularpolymorphRepetitive
0944-5013/$ - sdoi:10.1016/j.
�CorrespondE-mail addr
www.elsevier.de/micres
Morphological, physiochemical and molecularcharacterization of Anabaena strains
Radha Prasannaa,�, Ramesh Kumara, Anjuli Sooda,B.M. Prasannab, P.K Singhc
aCentre for Conservation and Utilization of Blue-Green Algae, Indian Agricultural Research Institute (IARI), New Delhi110012, IndiabDivision of Genetics, IARI, New Delhi 110012, IndiacChandra Shekhar Azad University of Agriculture and Technology, Kanpur 205002, India
Accepted 4 August 2005
S
riability;t;
ism;sequence
ee front matter & 200micres.2005.08.001
ing author.ess: [email protected]
SummaryA set of 30 Anabaena strains, isolated from diverse geographical regions of India, werecharacterized using morphological and physiochemical attributes as well as molecularmarker profiles. Significant differences were observed among the Anabaena strainswith regard to the shape and size of trichomes and individual cells within a filament,besides qualitative and quantitative aspects of phycobiliprotein accumulation andactivities of enzymes involved in nitrogen metabolism. Analyses of molecularpolymorphisms in a selected set of 13 Anabaena strains, using primers based onrepetitive sequences in the genome, led to unambiguous differentiation of the strainsas well as understanding of their genetic relationships. Informative morphological,physio-chemical and molecular characters have been identified that could aid indifferentiation and utilization of Anabaena strains as bioinoculants or as sources ofpigments.& 2005 Elsevier GmbH. All rights reserved.
Introduction
Cyanobacteria constitute one of the largestgroups of photosynthetic prokaryotes having aworldwide distribution in diverse ecosystems. Theyhave a wide-ranging impact on natural ecosystems,and many have both beneficial and harmful effects
5 Elsevier GmbH. All rights rese
m (R. Prasanna).
on geochemical cycles and human health. Besidestheir ecological importance, cyanobacteria are alsorecognized as potentially rich sources of pharma-cological and structurally interesting secondarymetabolites. However, the widespread exploita-tion and genetic improvement of cyanobacteriahave been hampered by constant changes in
rved.

ARTICLE IN PRESS
R. Prasanna et al.188
nomenclature and confusion regarding the authen-ticity of a strain being utilized by one group ofresearchers against a similar or dissimilar strainbeing investigated by another.
Anabaena, an environmentally important cyano-bacterium, has been identified mainly on the basisof morphological characteristics, such as shape andsize of trichomes, cell types, size and location ofheterocysts and akinetes, and plane(s) of division(Rippka et al., 1979). According to the acceptedmorphological and ecological descriptions by Desi-kachary (1959), the genus Anabaena is identifiedbased on the ‘‘Presence of uniform trichomes,absence of sheath or presence of more or lessdiffluent sheath forming free or floccose or softmucilaginous thallus. Heterocysts, generally inter-calary, and presence of a single or series of sporesnear the heterocyst or between the heterocysts’’.Since the taxonomic entries of trichome form andcharacteristics of the vegetative cell, heterocystand akinete are controversial due to the occasionalabsence of these characters and due to phenotypicchanges under different environmental conditions,a more reliable approach to the taxonomy ofcyanobacteria is required that also takes intoconsideration physiological, biochemical and mole-cular attributes.
Biochemical studies in cyanobacteria (Holtonet al., 1968) demonstrated a significant correlationbetween the morphological complexity of speciesand their fatty acid composition. Rippka et al.(1979) observed that biochemical diversity allowedthe distinction of subgroups within several generaincluding Nostoc and Anabaena. Analysis of enzymesrelated to nitrogen-assimilation in cyanobacteriamay also provide valuable pointers regarding theirutility as biofertilizers. Nitrogen-fixing cyanobac-teria utilize a number of assimilatory pathwaysinvolving extracellular N-sources to cellular N-containing components (Flores and Herrero, 1994).Although cyanobacteria preferentially utilize fixednitrogen and express nitrogenase only in theabsence of a suitable source of combined nitrogen,the ability to regulate the expression of differentenzymes (GS, GOGAT, NR, NiR) are highly specific.Since the variability in the expression of enzymes inN-related pathways are under a global controlmechanism involving ntcA (Flores and Herrero,1994), this can be an extremely valuable tool indiscriminating and screening genotypes for theirfurther characterization by molecular techniques.
The nitrogen-fixing cyanobacteria are known toconstitute about 12% of the total algal flora south of35 1N latitude, and nearly 2% of the flora north of it.Extensive surveys in India carried out in 1975 on2213 soil samples from paddy fields, showed that
only 33% harboured nitrogen-fixing forms. Regionaldifferences could also be seen in the distributionpattern of dominant forms, such as Aulosirafertilissima and species of Nostoc (Venkataraman,1981). However, comparison of profiles of strains ofa particular genus from various geographic loca-tions and their in-depth characterization has notbeen undertaken. The cyanobacterial germplasmmaintained at the Centre for Conservation andUtilization of Blue-Green Algae (CCUBGA), IARI,New Delhi (India) includes an array of heterocys-tous and non-heterocystous strains. Efforts ofcharacterising this vast genetic resource wereconfined mainly to morphological and to someextent, biochemical and physiological levels. Withthe advent of powerful DNA-based marker technol-ogies, there is an opportunity for comprehensivecharacterization of cyanobacterial germplasm atintra- and inter-specific levels.
In recent years, the use of molecular markers foridentification of cyanobacteria in diverse niches hasgained considerable significance. Molecular data-sets can effectively complement morphologicalcharacterization. Molecular markers such as re-striction fragment length polymorphism (RFLP) and16S rRNA, have been employed in phylogeneticanalysis and characterization of cyanobacterialdiversity (Giovannoni et al., 1990). The presenceof highly conserved repetitive sequences in thegenomes of microorganisms makes them highlyuseful for strain differentiation and diversityanalysis. Repeat elements, such as short tandemlyrepeated repetitive (STRR), long tandemly re-peated repetitive (LTRR) sequences (Mazel et al.,1990) or palindromic sequences such as highlyiterated palindrome (Hip1), have been found in anumber of cyanobacteria, especially heterocystousgenera. PCR fingerprinting techniques based onsuch sequences have proved valuable in theidentification and analysis of symbiotic cyanobac-teria (Rasmussen and Svenning, 1998; Smith et al.,1998). Molecular techniques have also been utilizedto ascertain the genetic relationship of the genusAnabaena with genera such as Aphanizomenon andNostoc (Gugger et al., 2002).
The present study was taken up (i) to compre-hensively characterize the variability in a selectedset of Anabaena strains collected from diversegeographical regions in India, besides international‘reference strains’, in terms of morphological, andbiochemical attributes; and (ii) to utilize molecularmarkers for unambiguous differentiation of thestrains and for analysis of genetic diversity. Thebroad goal is to identify useful attributes forclassification of members of the genus Anabaena,as well as identification of a core set of strains for
ARTICLE IN PRESS
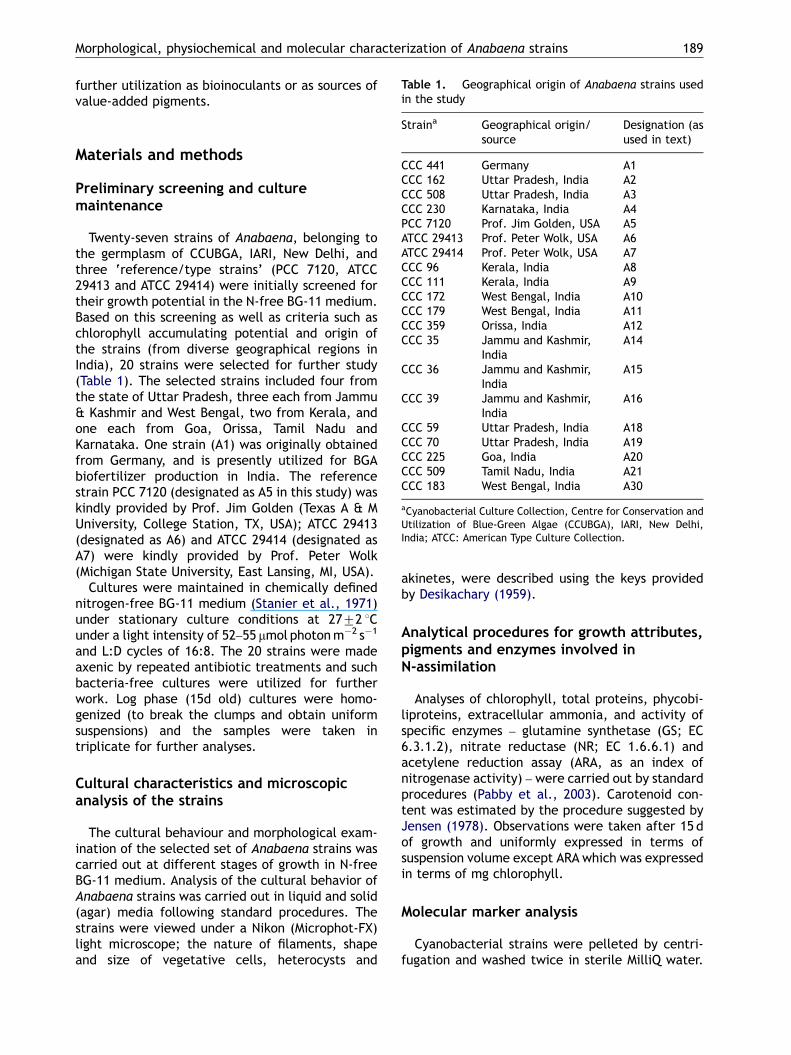
Table 1. Geographical origin of Anabaena strains usedin the study
Morphological, physiochemical and molecular characterization of Anabaena strains 189
further utilization as bioinoculants or as sources ofvalue-added pigments.
Straina Geographical origin/source
Designation (asused in text)
CCC 441 Germany A1CCC 162 Uttar Pradesh, India A2CCC 508 Uttar Pradesh, India A3CCC 230 Karnataka, India A4PCC 7120 Prof. Jim Golden, USA A5ATCC 29413 Prof. Peter Wolk, USA A6ATCC 29414 Prof. Peter Wolk, USA A7CCC 96 Kerala, India A8CCC 111 Kerala, India A9CCC 172 West Bengal, India A10CCC 179 West Bengal, India A11CCC 359 Orissa, India A12CCC 35 Jammu and Kashmir,
IndiaA14
CCC 36 Jammu and Kashmir,India
A15
CCC 39 Jammu and Kashmir,India
A16
CCC 59 Uttar Pradesh, India A18CCC 70 Uttar Pradesh, India A19CCC 225 Goa, India A20CCC 509 Tamil Nadu, India A21CCC 183 West Bengal, India A30
aCyanobacterial Culture Collection, Centre for Conservation andUtilization of Blue-Green Algae (CCUBGA), IARI, New Delhi,India; ATCC: American Type Culture Collection.
Materials and methods
Preliminary screening and culturemaintenance
Twenty-seven strains of Anabaena, belonging tothe germplasm of CCUBGA, IARI, New Delhi, andthree ‘reference/type strains’ (PCC 7120, ATCC29413 and ATCC 29414) were initially screened fortheir growth potential in the N-free BG-11 medium.Based on this screening as well as criteria such aschlorophyll accumulating potential and origin ofthe strains (from diverse geographical regions inIndia), 20 strains were selected for further study(Table 1). The selected strains included four fromthe state of Uttar Pradesh, three each from Jammu& Kashmir and West Bengal, two from Kerala, andone each from Goa, Orissa, Tamil Nadu andKarnataka. One strain (A1) was originally obtainedfrom Germany, and is presently utilized for BGAbiofertilizer production in India. The referencestrain PCC 7120 (designated as A5 in this study) waskindly provided by Prof. Jim Golden (Texas A & MUniversity, College Station, TX, USA); ATCC 29413(designated as A6) and ATCC 29414 (designated asA7) were kindly provided by Prof. Peter Wolk(Michigan State University, East Lansing, MI, USA).
Cultures were maintained in chemically definednitrogen-free BG-11 medium (Stanier et al., 1971)under stationary culture conditions at 2772 1Cunder a light intensity of 52–55 mmol photonm�2 s�1
and L:D cycles of 16:8. The 20 strains were madeaxenic by repeated antibiotic treatments and suchbacteria-free cultures were utilized for furtherwork. Log phase (15d old) cultures were homo-genized (to break the clumps and obtain uniformsuspensions) and the samples were taken intriplicate for further analyses.
Cultural characteristics and microscopicanalysis of the strains
The cultural behaviour and morphological exam-ination of the selected set of Anabaena strains wascarried out at different stages of growth in N-freeBG-11 medium. Analysis of the cultural behavior ofAnabaena strains was carried out in liquid and solid(agar) media following standard procedures. Thestrains were viewed under a Nikon (Microphot-FX)light microscope; the nature of filaments, shapeand size of vegetative cells, heterocysts and
akinetes, were described using the keys providedby Desikachary (1959).
Analytical procedures for growth attributes,pigments and enzymes involved inN-assimilation
Analyses of chlorophyll, total proteins, phycobi-liproteins, extracellular ammonia, and activity ofspecific enzymes – glutamine synthetase (GS; EC6.3.1.2), nitrate reductase (NR; EC 1.6.6.1) andacetylene reduction assay (ARA, as an index ofnitrogenase activity) – were carried out by standardprocedures (Pabby et al., 2003). Carotenoid con-tent was estimated by the procedure suggested byJensen (1978). Observations were taken after 15 dof growth and uniformly expressed in terms ofsuspension volume except ARA which was expressedin terms of mg chlorophyll.
Molecular marker analysis
Cyanobacterial strains were pelleted by centri-fugation and washed twice in sterile MilliQ water.

ARTICLE IN PRESS
R. Prasanna et al.190
The pellet was dissolved in an appropriate volumeof sterile water to ensure that at least a fewfilaments or cells were present in 1 ml, which wasused directly as a template for PCR. The sequencesof the oligonucleotide primers used for PCR arelisted in Table 6. Primers were synthesized byBangalore Genei (India) Ltd. The PCR cyclingconditions, primarily based on the proceduresuggested by Rasmussen and Svenning (1998), wereas follows: 1 cycle at 95 1C for 6min; 35 cycles of94 1C for 1min, 56 1C for 1min, and 65 1C for 5min,1 cycle at 65 1C for 16min, and a final step at 4 1C.For the LTRR primer, the PCR cycling program wasthe same, except that the annealing temperaturewas optimized at 45 1C for 1min with an extensionat 65 1C for 5min. Each PCR was carried out in a25 ml volume containing 50 pmol of each primer,1.25mM dNTPs, and 1U of Taq polymerase (B. GeneiLtd.). The amplifications were performed in a PCR-DYAD cycler (MJ Research Inc., USA).
The amplified products were resolved on 1.5%agarose gel in TAE buffer (0.04M Tris-acetate,0.001M EDTA, pH 8.0) and electrophoresed at80–100 V for about 1.5 h using a submarine gelelectrophoresis system (Biorad Model-96 or SunriseLife Technologies Model 12.16). In all, 500 and100 bp molecular size standards were run besidesthe amplified products to determine the approx-imate band size. The gels were photographed usinga CCD camera (Sony XC-75 CE) attached to a geldocumentation system with the Biocapture soft-ware (Vilber Lourmat, France). PCR reactions andelectrophoreses were repeated at least thrice foreach primer to ascertain the reproducibility of thebanding patterns.
Statistical analyses
The data recorded in triplicate for the para-meters in various strains were subjected to analysisof variance (ANOVA) in accordance with theexperimental design (completely randomized blockdesign) using MSTAT-C statistical package to quan-tify and evaluate the sources of variation. Duncan’smultiple range test (DMRT) was employed tocompare the mean performances of differenttreatments for the specific parameters under studyand the rankings are denoted by superscripts in therelevant tables.
For each molecular marker, a binary matrix of allbands present in each strain was generated, using‘1’ for the presence of an amplicon, and ‘0’ for itsabsence. Efficiency of discrimination was assessedin terms of the number of polymorphic markers
generated and the ability to generate uniquebanding profiles, the latter represented as ‘geno-type index’ (McGregor et al., 2000). Genotypeindex (GI), calculated as the number of genotypeswith unique banding profiles, expressed as afraction of the total genotypes fingerprinted,has a range of 0–1. In case of the geneticallydominant LTRR and STRR markers, PIC is synon-ymous with the term ‘gene diversity’ as describedby Weir (1990), and was calculated using theformula: 2[(1�q)2� (OQ)2], where Q is the pro-portion of genotypes showing the absence of anamplicon and q represents the frequency of therecessive allele.
Pairwise genetic similarities among the geno-types under study were determined usingJaccard’s coefficient (Jaccard, 1908), J ¼ N11/(N11+N10+N01), where N11 is the number of bandspresent in both individuals i and j, N10 is thenumber of bands present only in the individual i,and N01 is the number of bands present only in theindividual j. Unweighted pair group method usingarithmetic averages (UPGMA) clustering algorithm(Sneath and Sokal, 1973) was employed for gen-erating a dendrogram that depicts the geneticrelationships among the materials under study,using NTSYS-pc Version 2.11. Cophenetic correla-tion coefficients were computed to ascertain thecorrespondence between the genetic similaritymatrix and the dendrogram (Mohammadi andPrasanna, 2003) using the COPH and MXCOMPoptions in NTSYS.
Principal component analysis (PCA) is an ordina-tion technique that produces a visual representa-tion of the relative positions of genotypes in aspace of reduced dimensions, thus indicatingspatial relationships among genotypes (Mohammadiand Prasanna, 2003). In the present investigation,PCA was carried out using NTSYS-pc Version 2.11, tosummarize the variation in the molecular markerdataset and to make a graphical presentation of thegenetic relationships among the Anabaena strainsanalyzed.
Results
Cultural characteristics
In general, the selected Anabaena strains grew asfree-floating, floccose, gelatinous, mucilaginousmasses, while some grew as suspension in liquidmedium. Most of the forms were planktonic innature, and the color of thalli ranged from palegreen, bluish green to dark green. The reference

ARTICLE IN PRESS
Morphological, physiochemical and molecular characterization of Anabaena strains 191
strains A5, A6 and A7 particularly formed a bluishgreen, evenly distributed suspension, while strainsA12, A19 and A21 grew as thick floccose, mucila-ginous masses that did not pellet even after highspeed centrifugation. A16 grew as pale greenishbrown mucilaginous balls, either evenly distributedin the medium or settled at the bottom. Diversegrowth patterns of the selected strains on agar-based media was observed, including small muci-laginous patches or balls, or spreading type, orsmall randomly distributed flakes. Many strains,including A1, exhibited thread-like growth ofspreading nature, resembling a network-like layeron the agar.
Microscopic analysis
The strains, in general, grew as straight fila-ments, except in case of A3, A11, A16 and A19.These strains exhibited slight coiling and sinuousappearance. A3, A5 and A6 showed beaded appear-ance of filaments. Some of the strains (A4, A7, A10and A18) showed characteristic pointed end cell inthe filaments, while filaments of A12 were char-acteristically flexuous and tapering (Table 2). Thenumber of cells in a filament, on an average,ranged from 10 to 400. A7, A11, A12 and A19exhibited short filaments of 30–40 cells, while A5and A6 grew as long chains of cells consisting ofaround 300 cells. The selected 20 strains exhibitedvegetative cells ranging in shape from square tospherical, barrel, elongate or cylindrical. A4 andA16 exhibited characteristic cylindrical cells. Fila-ments of A3 consisted of small spherical cells ofdiameter ranging from 4.2 to 4.4 mm while max-imum cell size (7.6–8.2� 2.0–3.5 mm) was observedin filaments of A18.
Heterocysts ranged in shape from oval, globularto elongate-barrel. The length of heterocysts wasmarginally higher compared to the vegetative cells.Mostly intercalary and rarely terminal heterocystswere observed in all strains. Terminal heterocystswere also associated with broken filaments.
Akinetes were observed in all strains at lateexponential stages of growth. The length ofakinetes was 1.5–4-fold higher as compared tovegetative cells while the breadth was slightlymore. The largest akinetes were observed in A20(18.2–20.0� 7.8–8.0 mm) and smallest in A5(5.6–6.5� 5.0–5.6 mm). The akinetes were gener-ally formed away from heterocyst except in A5.Akinetes also formed in series in most forms,except in A4 where a single akinete was generallyobserved next to the heterocyst.
Growth attributes
A wide range with respect to chlorophyll accu-mulation (0.11–3.40 mgml
�1
) was observed (Table3). The three top-ranked strains for chlorophyllaccumulation, based on analysis of mean valuesusing DMRT, were A21, A10 and A4. Comparison ofthe strains on the basis of total proteins revealedthe potential of A19, A4 and A9. A15 exhibited theextremely low value of 31.17 mgml
�1
, while thehighest ranked strain A19 displayed the highesttotal protein value (447.69 mgml
�1
). Doubling timewas evaluated after measuring chlorophyll after 12,24, 36 and 48 h of incubation in N-free BG-11medium. Most of the strains showed a doublingtime between 24 and 36 h, while A3, A5, A9, A12,A16 and A19 exhibited a doubling time of 18–24 h.
Carotenoid and phycobiliproteinaccumulation
The carotenoid content in the selected Anabaenastrains ranged from 0.4 to 21.63 mgml
�1
(Table 4).Strains A18, A19 and A7 exhibited the highestvalues for carotenoid accumulation. Similarly, thestrains also showed significant variability withrespect to phycobiliproteins, comprising phycocya-nin (PC), phycoerythrin (PE) and allophycocyanin(APC). The total phycobiliproteins ranged from1.4 mgml
�1
in A30 to 55.67 mgml�1
in A7. Most ofthe strains showed total phycobiliproteins in therange of 20–30 mgml�1. A15, with a value of59.39%, had the highest % Phycobiliproteins out oftotal proteins, while the lowest value was recordedin A30 (1.8%). A6, A11 and A7 were the top-rankedstrains for PC content, exhibiting values of 26.80,21.0 and 20.67 mgml�1, respectively, while A20, A7and A4 were ranked highest for APC content (Table4). PE content ranged from 0.43 (A30) to9.97 mgml�1 (A7). Among the strains analyzed, A7,A15 and A16 were top-ranked in that order for thePE content.
Activities of enzymes related toN-assimilation
Strain A20 exhibited the highest ARA value(22939.7� 10
3
nmoles C2H4mg chl�1 h�1), while thelowest activity (602.4 nmoles) was recorded in A30(Table 5). A1 and A18 were ranked second and thirdwith respect for this attribute. Nitrate reductase(NR) activity of the cultures ranged from2.83 mmoles NO2
� ml�1
(in A30) to 31.83 mmolesNO2�ml�1 (in A10). Among the strains analyzed,

ARTICLE IN PRESS
Table
2.
Morpho
logica
lch
arac
teristicsof
Ana
baen
astrainsreve
aled
bymicroscop
ican
alysis
Strain
Vege
tative
cell
Heteroc
yst
Akine
te
Shap
eSize
(mm)
Size
(mm)
Position
*Freq
uenc
y(%)
Shap
eSize
(mm)
Leng
thBread
thSh
ape
Leng
thBread
thLe
ngth
Bread
th
A1
Elon
gate
4.7–
5.3
3.8–
4.2
Sphe
rica
l–barrel
5.6–
6.0
5.0–
5.6
I&
T3.1
Cylindrica
l10
.2–11
.25.0–
5.8
A2
Barrel
6.0–
6.5
3.8–
4.0
Barrel
6.0–
6.5
4.0–
4.2
I&
T5.0
Elon
gate
8.4–
10.8
5.4–
6.0
A3
Sphe
rica
l4.2–
4.4
4.2–
4.4
Sphe
rica
l5.2–
6.4
5.2–
6.4
I&
T12
.5Barrel
11.4
–13
.37.8–
8.4
A4
Cylindrica
l6.2–
7.4
5.4–
6.2
Barrel
14.2
–16
.28.2–
10.4
I&
T6.5
Ovo
id–cylind
rica
l15
.2–17
.47.8–
8.9
A5
Short-barrel
4.2–
5.0
5.0–
5.6
Barrel
5.0–
5.6
4.7–
5.0
I&
T4.8
Barrel
5.6–
6.5
5.0–
5.6
A6
Short-barrel
3.3–
4.2
3.8–
4.8
Barrel
5.6–
6.7
4.5–
5.6
I&
T6.6
Ovo
id–barrel
7.6–
8.6
5.5–
6.6
A7
Barrel
5.7–
9.5
3.2–
4.4
Sphe
rica
l–barrel
5.7–
7.6
5.7–
6.0
I&
T4.0
Globular
10.2
–15
.04.7–
5.7
A8
Barrel
4.4–
6.6
3.8–
4.6
Barrel
4.7–
7.2
4.7–
5.7
I&
T4.1
Short–cylind
rica
l8.4–
10.8
5.4–
6.0
A9
Barrel
4.0–
5.6
3.8–
4.6
Sphe
rica
l–barrel
4.2–
5.6
3.8–
4.2
I&
T5.0
Obov
ate
7.2–
10.3
5.6–
6.2
A10
Barrel
5.7–
7.2
4.7–
5.7
Barrel
7.2–
9.5
5.7–
6.6
I&
T5.0
Barrel
5.7–
7.2
4.7–
5.7
A11
Barrel
3.8–
4.7
3.8–
4.7
Barrel
5.6–
6.5
4.7–
5.5
I&
T9.8
Barrel
7.6–
8.4
6.3–
6.8
A12
Barrel
6.2–
7.4
5.6–
6.2
Globular
6.6–
7.2
5.8–
6.2
I&
T5.0
Elon
gate
15.2
–18
.44.7–
5.7
A14
Barrel
7.3–
7.6
4.6–
5.3
Ova
l7.4–
8.6
4.8–
5.3
I&
T4.5
Ovo
id–glob
ular
14.3
–16
.06.3–
6.8
A15
Elon
gate
4.7–
5.6
3.5–
4.2
Elon
gate
4.7–
5.8
4.6–
5.7
I&
T5.0
Globular
6.8–
7.2
5.7–
6.2
A16
Cylindrica
l4.6–
5.8
4.6–
5.2
Sphe
rica
l6.8–
7.2
5.7–
6.1
I&
T2.1
Elon
gate
9.2–
10.0
6.0–
6.4
A18
Barrel
7.6–
8.2
2.0–
3.5
Elon
gate
11.2
–12
.44.8–
5.2
I&
T10
.0Cylindrica
l16
.7–17
.55.7–
6.1
A19
Barrel
3.8–
4.5
5.6–
6.3
Elon
gate
6.4–
6.8
5.2–
5.6
I&
T6.0
Ovo
id–cylind
rica
l11
.8–17
.511
.2–14
.7A20
Barrel
3.7–
4.5
3.0–
4.0
Elon
gate
–barrel
4.7–
5.8
3.8–
4.2
I&
T6.0
Ovo
id–cylind
rica
l18
.2–20
.07.8–
8.0
A21
Barrel
4.1–
4.4
3.6–
4.0
Sphe
rica
l4.8–
5.4
3.8–
4.2
I&
T9.0
Cylindrica
l10
.4–11
.29.4–
10.3
A30
Barrel
4.7–
5.5
4.7–
5.3
Sphe
rica
l7.6–
8.4
7.2–
7.6
I&
T5.0
Cylindrica
l14
.2–16
.310
.4–12
.6
*I&
T:bothintercalaryan
dterm
inal.
R. Prasanna et al.192

ARTICLE IN PRESS
Table 3. Mean performances of the Anabaena strainsand their DMRT ranking for selected growth attributes
Strain Chlorophyll* Doubling time* Proteins*(mgml�1) (in h) (mgml�1)
A1 0.373 24–36 120.511
A2 1.209 24–36 89.676
A3 0.444 18–24 118.3310
A4 2.6117 24–36 212.0018
A5 1.3711 18–24 103.338
A6 1.9713 24–36 126.512
A7 1.3310 24–36 151.6716
A8 0.302 24–36 141.6713
A9 2.1715 18–24 190.3317
A10 2.7018 24–36 145.3314
A11 2.2016 24–36 112.679
A12 1.107 18–24 97.007
A14 1.138 24–36 63.002
A15 1.5712 24–36 31.171
A16 0.906 18–24 78.503
A18 2.0014 24–36 78.503
A19 2.1715 18–24 447.6919
A20 0.111 24–36 149.6715
A21 3.4019 24–36 80.174
A30 0.605 24–36 81.005
S.E (7mean) 0.35 — 74.73
*Superscripts denote DMRT ranking in ascending order.
Morphological, physiochemical and molecular characterization of Anabaena strains 193
A10, A18 and A9 were top-ranked for this para-meter.
A4, A14 and A7 were ranked highest in terms ofGS activity (Table 5). The values ranged from16.20 mmoles g-glutamyl hydroxamate ml�1 (inA11) to 272.07 mmoles g-glutamyl hydroxamateml�1 (in A4). Ammonia excretion (measured asmmoles NH+
4 ml�1) was observed to be negligible inseveral strains including A2, A3, A5, A6, A7, A8,A16, A18 and A21. The highest values of 0.7 mmoleswere recorded in A14 and A10, followed by A9(0.57 mmoles; Table 5).
Molecular profiling and genetic diversityanalysis of selected Anabaena strains
For molecular profiling, 13 strains, A1, A2, A3,A5, A6, A7, A8, A9, A10, A11, A14, A15 and A16)were selected based on their morphological andbiochemical attributes. Strains A5, A6 and A7 wereselected for this study as these are the interna-tional ‘reference or type strains’. A5 exhibits thedistinct cell morphology and pattern of akinetedevelopment, quite similar to Nostoc, whereas A8and A16 exhibit a growth pattern with slightlycoiled filaments. A6 and A7 were found to be high
phycobilin producers, while A20, A1 and A18 havethe highest nitrogen-fixing potential. Total proteincontent and NR activity were highest in case of A9,while A15 recorded the highest phycobiliproteincontent. The ammonia excreting ability was max-imum in the strains A14, A9 and A10, whereasGS activity was found to be highest in case of A4and A14.
Strain differentiation by molecular markers
The selected strains were analyzed with fourspecific primers, LTRR, STRRmod, STRR1A andHipTG. PCR amplification of the selected strainsusing the LTRR primer revealed 13 polymor-phic amplicons and nine distinct banding profiles(Fig. 1A), of which seven profiles were unique(in case of A3, A8, A9, A10, A11, A14 and A15). Theprimer was unable to differentiate A5 and A7 aswell as A6 and A16. STRR1A generated eightamplicons of which seven were polymorphic. Fivedistinct banding profiles were observed in thiscase, three of which were unique to A3, A5 andA11 (Fig. 1B). A6 and A7, as well as A8, A9, A10,A14, A15 and A16 exhibited common patterns.Strains A1 and A2 consistently showed no amplifica-tion with STRR1A.
Amplification with STRRmod resulted in six ampli-cons, all of which were polymorphic. Six differentbanding profiles were generated by this primer,of which unique profiles were limited to fourstrains (A3, A6, A8 and A10). No amplificationwas observed in strains A2, A9, A14 and A16. A1and A15 generated a similar profile with a singlelow molecular weight band (300 bp), while A5, A7and A11 strains yielded a common pattern. Ampli-fication using the extended Hip primer, HipTG,resulted in 8 polymorphic amplicons with 100%polymorphism. The primer generated as many as 9distinct banding profiles, eight of which wereunique (in A3, A5, A7, A8, A9, A11, A14 and A16).Only A1, A6, A10 and A15 displayed a commonbanding pattern.
It could be noticed that a combination of just twoprimers (LTRR and HipTG) could unambiguouslydifferentiate all the 13 Anabaena strains employedin this study. The GI of HipTG was found to be thehighest (0.61), followed by LTRR (0.54). HipTG alsodisplayed relatively higher PIC value (0.24). On thebasis of polymorphism information as well as GI,HipTG and LTRR primers are identified in thepresent study as highly suitable for differentiationof the Anabaena strains. Thus, the study highlightsthe utility of primers based on repetitive sequences
ARTICLE IN PRESS
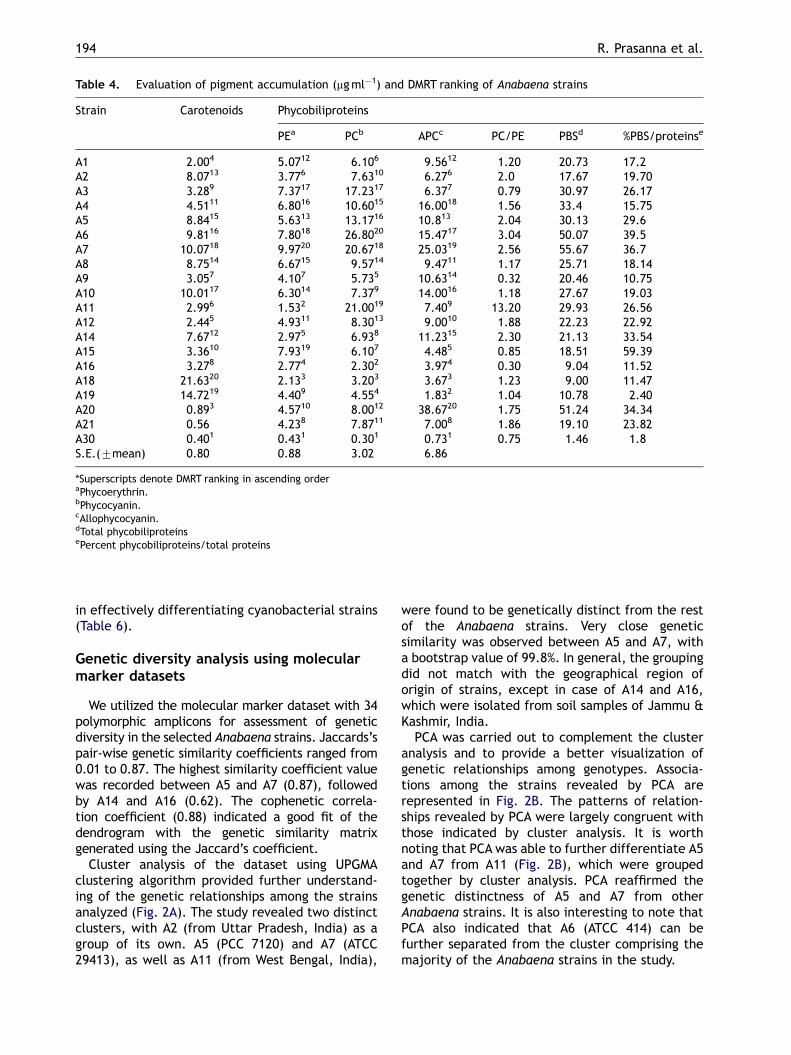
Table 4. Evaluation of pigment accumulation (mgml�1) and DMRT ranking of Anabaena strains
Strain Carotenoids Phycobiliproteins
PEa PCb APCc PC/PE PBSd %PBS/proteinse
A1 2.004 5.0712 6.106 9.5612 1.20 20.73 17.2A2 8.0713 3.776 7.6310 6.276 2.0 17.67 19.70A3 3.289 7.3717 17.2317 6.377 0.79 30.97 26.17A4 4.5111 6.8016 10.6015 16.0018 1.56 33.4 15.75A5 8.8415 5.6313 13.1716 10.813 2.04 30.13 29.6A6 9.8116 7.8018 26.8020 15.4717 3.04 50.07 39.5A7 10.0718 9.9720 20.6718 25.0319 2.56 55.67 36.7A8 8.7514 6.6715 9.5714 9.4711 1.17 25.71 18.14A9 3.057 4.107 5.735 10.6314 0.32 20.46 10.75A10 10.0117 6.3014 7.379 14.0016 1.18 27.67 19.03A11 2.996 1.532 21.0019 7.409 13.20 29.93 26.56A12 2.445 4.9311 8.3013 9.0010 1.88 22.23 22.92A14 7.6712 2.975 6.938 11.2315 2.30 21.13 33.54A15 3.3610 7.9319 6.107 4.485 0.85 18.51 59.39A16 3.278 2.774 2.302 3.974 0.30 9.04 11.52A18 21.6320 2.133 3.203 3.673 1.23 9.00 11.47A19 14.7219 4.409 4.554 1.832 1.04 10.78 2.40A20 0.893 4.5710 8.0012 38.6720 1.75 51.24 34.34A21 0.56 4.238 7.8711 7.008 1.86 19.10 23.82A30 0.401 0.431 0.301 0.731 0.75 1.46 1.8S.E.(7mean) 0.80 0.88 3.02 6.86
*Superscripts denote DMRT ranking in ascending orderaPhycoerythrin.bPhycocyanin.cAllophycocyanin.dTotal phycobiliproteinsePercent phycobiliproteins/total proteins
R. Prasanna et al.194
in effectively differentiating cyanobacterial strains(Table 6).
Genetic diversity analysis using molecularmarker datasets
We utilized the molecular marker dataset with 34polymorphic amplicons for assessment of geneticdiversity in the selected Anabaena strains. Jaccards’spair-wise genetic similarity coefficients ranged from0.01 to 0.87. The highest similarity coefficient valuewas recorded between A5 and A7 (0.87), followedby A14 and A16 (0.62). The cophenetic correla-tion coefficient (0.88) indicated a good fit of thedendrogram with the genetic similarity matrixgenerated using the Jaccard’s coefficient.
Cluster analysis of the dataset using UPGMAclustering algorithm provided further understand-ing of the genetic relationships among the strainsanalyzed (Fig. 2A). The study revealed two distinctclusters, with A2 (from Uttar Pradesh, India) as agroup of its own. A5 (PCC 7120) and A7 (ATCC29413), as well as A11 (from West Bengal, India),
were found to be genetically distinct from the restof the Anabaena strains. Very close geneticsimilarity was observed between A5 and A7, witha bootstrap value of 99.8%. In general, the groupingdid not match with the geographical region oforigin of strains, except in case of A14 and A16,which were isolated from soil samples of Jammu &Kashmir, India.
PCA was carried out to complement the clusteranalysis and to provide a better visualization ofgenetic relationships among genotypes. Associa-tions among the strains revealed by PCA arerepresented in Fig. 2B. The patterns of relation-ships revealed by PCA were largely congruent withthose indicated by cluster analysis. It is worthnoting that PCA was able to further differentiate A5and A7 from A11 (Fig. 2B), which were groupedtogether by cluster analysis. PCA reaffirmed thegenetic distinctness of A5 and A7 from otherAnabaena strains. It is also interesting to note thatPCA also indicated that A6 (ATCC 414) can befurther separated from the cluster comprising themajority of the Anabaena strains in the study.

ARTICLE IN PRESS
Table 5. Mean performances and DMRT ranking of Anabaena strains for selected biochemical attributes related tonitrogen assimilation
Strain ARAa *(nmoles C2H4 mg chl�1 h�1) NRb *(mmoles NO2�) GSc *(mmoles gGHml�1) Ammonia* (mmolesml�1)
A1 17395.9019 21.3315 58.8316 0.083
A2 2350.108 16.3311 33.9011 0.001
A3 980.204 10.177 31.709 0.001
A4 2727.5010 18.5013 272.0720 0.176
A5 3143.0012 26.0016 25.335 0.001
A6 2193.197 18.8314 31.9310 0.001
A7 4185.0715 21.3315 88.5018 0.001
A8 687.592 14.5010 54.4714 0.001
A9 2969.2011 30.6717 30.578 0.5710
A10 1075.256 31.8319 77.8717 0.7011
A11 3465.3413 13.509 16.201 0.114
A12 4208.1516 16.6712 28.207 0.268
A14 3772.3714 6.004 104.3319 0.7012
A15 714.433 7.675 24.804 0.569
A16 1041.505 4.332 22.032 0.001
A18 6504.6418 31.1718 47.8313 0.001
A19 2469.229 5.003 42.3012 0.257
A20 22939.7020 12.838 26.976 0.175
A21 5533.4017 10.006 55.6015 0.001
A30 602.401 2.831 23.133 0.022
S.E.(7mean) 1641.11 1.81 4.40 0.005
*Superscripts denote DMRT ranking in ascending order.aAcetylene reduction assay.bNitrate reductase.cGlutamine synthetase.
Table 6. Molecular polymorphisms in the Anabaena strains revealed by primers based on repetitive sequences
Primer Sequence (50 to 30) Nf Np Nbp Nubp GI PIC
LTRR GACAGACAGACGACA 13 13 9 7 0.54 0.19STRR1A CCAATCCCCAATCCCC 8 7 5 3 0.23 0.20STRRmod GCGCCCCAATCC 6 6 6 4 0.31 0.22HipTG GCGATCACTG 8 8 9 8 0.61 0.24
Nf: number of amplicons; Np: number of polymorphic amplicons; Nbp: number of banding profiles revealed across the strains; Nubp:number of unique banding profiles; GI: genotype index; PIC; polymorphism information content.
Morphological, physiochemical and molecular characterization of Anabaena strains 195
Discussion
Cyanobacteria are an ancient group of prokar-yotes that played an important role in the evolutionof life on earth, both in the genesis of an aerobicatmosphere and as precursors of higher plantplastids. These oxygenic prokaryotes comprise asingle phylogenetic branch within the domainbacteria (Eubacteria). On the basis of most recent16S rDNA sequence analysis, this branch or clusterwas found most closely related to Gram-positivebacteria, particularly the low G+C group includingClostridium inocuum and Bacillus subtilis (Casten-holz, 2001). Cyanobacteria occupy a wide range of
illuminated niches in terrestrial, fresh water, marineand hypersaline environments. These organisms alsoexhibit a high degree of morphological, physiologicaland developmental complexity. Their distinct biolo-gical characteristics made them model systems inunderstanding metabolic processes in biology. Oneof the commonly observed genera in rice paddies isAnabaena which exists both in free living state andalso as symbiont with the water fern Azolla. ManyAnabaena strains exhibit high nitrogen fixing poten-tial and have been the favourite organisms forgenetic manipulation (Wolk et al., 1994). However,the identification of this genus is complex as somemorphological features change and may not be

ARTICLE IN PRESS
Figure 1. PCR-based DNA fingerprint patterns of the Anabaena strains revealed by (A) LTRR; and (B) STRR1A. M1 and M2represent 500 and 100 bp molecular size standards, respectively.
R. Prasanna et al.196
expressed always in culture (Li, 1998). A number ofresearchers attempted to utilize alternative meth-ods such as numericulture (numerical code based onmorphological and physiological characters) or DNAanalysis combined with morphological traits (Stulpand Stam, 1984; Hiroki et al., 1998) for identifica-tion and characterization of Anabaena. The presentstate of uncertainty in identification is mainly due totaxonomical treatises being based on Botanical Codeof Nomenclature. Although in recent studies, inclu-sion of other attributes provided better reliability,confusion still exists with regard to commonlyinvestigated genera and species. A classic example,in this context, can be seen in the most recentclassification (Rippka et al., 2001), wherein Anabae-na PCC 7120, the workhorse of cyanobacteriology,and other strains showing a high degree of DNA-DNAsimilarity with Nostoc were transferred to the genusNostoc (Rippka and Herdman, 1992).
Most of the earliest taxonomic studies have beenbased on protologues (field materials and preserved
specimens). A number of workers collected andinvestigated samples from different ecologicalhabitats and gave taxonomical assignments (Gei-tler, 1932; Fritsch, 1949; Desikachary, 1959).Desikachary and Anand (1974) identified a largenumber of isolates with different geographicaldistribution under identical cultural conditionsand emphasized the need for more cultural studies.Our investigation is on similar lines and provides anidea of the variability in cultural and biochemicalcharacteristics, observed as a result of theirgenetic background, despite identical culturalconditions. Also, by employing molecular markers,the Anabaena strains have been unambiguouslydifferentiated and the genetic diversity among thestrains ascertained.
The genus Anabaena was established by Bory in1922 (Geitler, 1932). In the first revision of thisgenus, 11 species were described; later, Geitler(1932) described 57 European species of Anabaena.Although this was one of the most comprehensive

ARTICLE IN PRESS
Figure 2. Genetic relationships revealed by analyses of the molecular marker dataset of the cyanobacterial strains: (A)cluster analysis using UPGMA, with bootstrap values at each node of the dendrogram; (B) principal component analysis.
Morphological, physiochemical and molecular characterization of Anabaena strains 197
regarding blue-green algal taxonomy, the presenttaxonomic classification was considered ‘chaotic atthe genus and species levels’ (Rippka et al., 2001).Fritsch (1949) carried out taxonomic studies on thisgenus using species isolated from India and areasadjacent to Asiatic mainland. Rippka et al. (1979)and Herdman et al. (1979) proposed a taxonomicalclassification mainly based on biochemical char-acters like DNA base composition. A morphologicalapproach adopted by Drouet (1968) led to adramatic reduction in the number of Anabaenaspecies. The present confusion is mainly due toscepticism about the taxonomic value of thecriteria adopted by Geitler (1932). Stulp and Stam(1982, 1984) studied the morphological variability
of Anabaena strains under different light, tempera-ture and salinity conditions and observed that onlyunder adverse conditions, deviations in morphologyoccurred, in those characteristics that have beentraditionally accepted as useful in Anabaenataxonomy. A new and somewhat widely recognizedtaxonomic revision was later published (Anagnosti-dis and Komarek, 1985; Komarek and Anagnostidis,1989), based on the definition of smaller, morecoherent genera and integration of the availablebiochemical, ultrastructural and molecular char-acters. On the basis of all these treatises, Rippkaet al. (2001) provided the following description forthe form-genus 1. Anabaena Bory de St. Vincent1822 sensu Rippka et al. (1979) in Bergey’s Manual

ARTICLE IN PRESS
R. Prasanna et al.198
of Systematic Bacteriology: ‘‘Trichomes straight orslightly sinuous, vegetative cells spherical, cylind-rical, or barrel shaped, and separated by conspic-uous constrictions at the crosswalls. Heterocystspredominantly intercalary, though terminal het-erocysts may also occur. Akinetes located, singlyor in groups (2–3), adjacent to heterocysts oroccurring in chains, initiated distant from hetero-cysts. A firm sheath is absent, but mucilageproduction may be abundant; false branchinggenerally not observed. Reproduction by randomtrichome fragmentation. Hormogonia, distinct inmorphology from the parental trichomes, notproduced’’.
In the present study, we have taken intoconsideration all the above-mentioned treatises,and carried out comprehensive characterization ofa set of 20 Anabaena strains, including culturalcharacteristics, biochemical attributes and mole-cular marker profiles. Desikachary (1970) stressedthat cultural studies are essential to understandthe behavioural pattern of blue green algae,exhibiting a wide range of variability. In the presentinvestigation, most of the Anabaena strains ex-hibited planktonic—floccose thalli, which rangedfrom pale bluish green to dark green in colour.Colour of the plant mass has not been observed tohave much taxonomic value as there are too manyresemblances among the strains (Stulp and Stam,1982); however, this can be a preliminary distin-guishing feature. The Anabaena strains analyzed inthe present study consisted of filaments which weremostly straight with slight curvature, especially inthe forms exhibiting suspension like growth. Flexa-tion of trichomes was rather uniform among thestrains in the present study and this was inaccordance with earlier investigations (Anand,1979; Hiroki et al., 1998), although shown to havelow taxonomic value (Stulp, 1982). The growthpattern of cultures on solidified and liquid mediumwas distinct for the strains and groups could beidentified on the basis of spreading net like growthand patches type of growth. However, earlierworkers (Stulp, 1982; Stulp and Stam, 1982) havenot given much value to this criterion. Stulp (1982)concluded from his studies on morphologicalvariability of Anabaena strains that traditionallyaccepted criteria in the taxonomy of the genusAnabaena are stable over a wide range of environ-mental conditions and hence serve their utility fortaxonomy. The shape of terminal cell was observedto be a useful characteristic by Drouet (1968) andStulp (1982). In the present study, A4, A7, A10and A18 showed characteristic pointed/taperedend cell, which is a clear diagnostic feature foridentification at species level.
Trichome form and shape was almost uniform inour studies and important as an identificationcriterion (Table 2). Hiroki et al. (1998) consideredsimilar attributes for developing a database systemfor identification of planktonic Anabaena speciesbased on morphological characteristics; this has areference database containing 19 planktonic Ana-baena species along with morphological character-istics of 26 attributes. Of all the numerical criteriautilized in taxonomic investigations (Stulp andStam, 1982), vegetative cell width seems to bethe most stable and useful one. The cell width ofbotanical ‘species’ of Anabaena ranged between 2and 15 mm (Geitler, 1932). However, most repre-sentatives presently in culture show cylindrical tobarrel shaped cells of relatively small dimensions(3–6 mm). In the present study, most of theAnabaena strains showed a similar cell width ofvegetative cells, except A4 and A18 which exhib-ited slightly larger cells. Length/width ratio ofvegetative cells was observed to be quite stableand this is in consonance with earlier work (Stulpand Stam, 1982).
The dimensions of heterocysts were larger in A4and A18. The heterocysts of A4 filaments werealmost double in size as compared to vegetativecells. Such features were recorded earlier inAnabaena species (Geitler, 1932). Heterocyst fre-quency was observed to range from 2.2% to 12.5%.Interestingly, strain A4 exhibited very large hetero-cysts, and also a relatively higher heterocystfrequency of 6%.
According to Komarek and Anagnostidis (1989)heterocyst differentiation in Anabaena alwaysoccurs in an intercalary position, but in Anabaenacylindrica, conical end cells sometimes give rise tosimilarly shaped heterocysts and may becometerminal after trichome breakage. All Anabaenastrains investigated showed mostly intercalaryheterocysts but terminal heterocysts (possibleresult of breakage) were rarely observed.
Akinete morphology was observed to be a steadycharacteristic in light microscopic examination ofcultures for their identification (Hiroki et al.,1998). Stulp (1982) observed that incubation underdifferent light and temperature conditions hardlyaffected taxonomically important characters (suchas dimensions and shape of vegetative cells,heterocysts and akinetes, position of akinetes withregard to heterocyst, shape of terminal cells). Theposition of spores, whether adjacent to or awayfrom heterocyst was found to be a stable characterin Anabaena species (Anand, 1979; Stulp and Stam,1982). The characteristic formation of chains ofspores was observed to a stable diagnostic featurein our studies (Table 2). A4 exhibited distinctly

ARTICLE IN PRESS
Morphological, physiochemical and molecular characterization of Anabaena strains 199
large sized ovoid-cylindrical shaped akinetes, whilethe akinetes of A20 were much larger and similar inshape.
Another important feature that is utilized fordistinguishing Anabaena from Nostoc is the mode ofdevelopment of akinetes with respect to theheterocyst. Anabaena strains are differentiatedfrom Nostoc on the basis of their development ofakinetes distant from the heterocyst. The presenceand position of heterocysts and their relativearrangement with respect to spores have been ofmore taxonomic significance. A set of diagnostickeys of 19 species of Anabaena was proposed,based on cultural studies of the isolates (Anand,1979). Anabaena variabilis and other speciesexhibiting a different mode of akinete develop-ment were placed in Trichormus a new genusdeveloped by Komarek and Anagnostidis (1989).Interestingly, A20 showed characteristic develop-ment of akinetes on both sides of the heterocyst,which is an extremely important taxonomic trait(Desikachary, 1959). The position of akinetes in thetrichomes has been universally considered as animportant criterion in traditional taxonomy ofAnabaena due to its stable and reliable nature(Anand, 1979; Hiroki et al., 1998; Stulp and Stam,1982).
Characterization of the selected 20 strains ofAnabaena for their growth attributes providedinteresting information regarding the wide varia-tion in terms of chlorophyll, doubling time andprotein accumulation. A21, A10 and A4 were thetop ranked strains, accumulating up to 3 mgml�1
chlorophyll after 15 d of growth. Chlorophyllvalues, in general, ranged from 0.11 to 3.4 mgml�1.Such wide variations within a genus and also withinspecies have been observed earlier (Roychoudhuryet al., 1986; Mekonnen et al., 2002). Among thethree top-ranked strains for proteins, A4 found aplace again, indicating its utility in mass productionsystems because of its high growth potential interms of chlorophyll and total proteins synthesized.Cyanobacterial strains exhibit doubling times (dur-ing exponential growth) in the range of 18–36 h(Meeks et al., 1983). Out of the 20 selectedstrains, six strains (A3, A5, A9, A12, A16 and A19)were fast growers, exhibiting doubling time of18–24 h while the others showed a doubling time of24–36 h. Cyanobacterial doubling times duringexponential growth are often more than 12 h andusually 24 h or longer, even in cultures withunlimited nutrients and a saturating intensity ofcontinuous light. Meeks et al. (1983) observed thatthe doubling time of Anabaena 7120 and A.cylindrica in nitrogen free BG11 medium was 21.5and 18.2 h, respectively.
Besides chlorophyll, cyanobacteria possess car-otenoids and phycobiliproteins (PBS) which serve asaccessory pigments in photosynthesis. Carotenoidsprotect the cyanobacterial cell from photooxida-tive damage and are found associated with pro-teins. It was interesting to observe that strain A19exhibited the highest total proteins and the secondhighest rank for carotenoid content. Total phyco-biliproteins was highest in A7 which also exhibitedhighest PE, and figured among the top three strainsfor PC and APC content. Excluding A3, A9, A16 andA30, the other strains were generally PC-rich. A3was observed to be a PE-rich strain, with a PC/PEratio of 0.3. All naturally occurring cyanobacteriawere found to produce PC (Bryant 1982). InAnabaena PCC 7120 only a single set of set of PC-related genes have been identified, which areconstitutive in nature (Sidler, 1994). Late log phasecultures of A5 (PCC 7120) were observed to exudeblue colour into the medium under routine cultureconditions. In contrast, PE is not always found in allcyanobacteria and its chromophore contents isknown to differ for cyanobacteria living in eitherfreshwater or in soil or in marine environments(Bryant, 1982). Our observations were in conso-nance with those from an earlier work (Bryant,1982) in that the PC-rich strains were morecommonly found among cyanobacteria, especiallyAnabaena strains. The ranking of strains for PC, APCand PE content and also for their total phycobili-proteins and % PBS out of total proteins can be veryuseful in selection of strains for their potentialutilization as colouring agents, phycoflour probes oras additives in a range of cosmetic and pharma-ceutical products (MacColl and Guard-Friar, 1987).In the present study, A7 showing the highest totalphycobilproteins and ranking high for the individualpigments (PC, PE and APC), and A15 showing higher% PBS of total proteins are identified as potentialcandidates for exploitation in value-addition.
Phototrophic cyanobacteria are capable of grow-ing on a variety of inorganic and organic nitrogensources. Many of these photosynthetic prokaryotesare also able to fix atmospheric nitrogen underconditions of deprivation for combined nitrogen.These properties account for their abundance inrice fields, among which the nitrogen fixing speciesconstitute half or more of the population (De,1939), and are considered responsible for thesustained fertility of the rice ecosystem. Nitrogenfixation by cyanobacteria is an economically viableinput in rice cultivation and selection and manip-ulation of high nitrogen fixing strains is an ongoingprocess in most rice growing countries. Heterocystdifferentiation is primarily a response to the N-status of the culture. In the present investigation,

ARTICLE IN PRESS
R. Prasanna et al.200
in the absence of combined N source in themedium, nitrogenase activity was high in most ofthe strains ranging from 22.94� 103 nmolesC2H4 mg chl�1 h�1 in A20 to a lowest value of0.602� 103 nmoles in A30. A20 is an interestingstrain with lowest chlorophyll value, moderateprotein content and 6% heterocyst frequency, butwith very high ARA value. This strain incidentallyalso showed low GS activity, which is consideredimportant for its utilization as a biofertilizer. Incontrast, A30 was lowly ranked for most attributesand as expected shows lowest ARA value too. A20needs to be critically evaluated not only as a modelsystem for the genetics and regulation of nitrogenfixation but also for its possible deployment as abioinoculant.
Nitrate assimilation has been exhaustively re-viewed by a number of researchers (Bagchi, 1994).A significant feature of all cyanobacterial NRs is itsclose association with photosynthetically activethylakoid membranes and dependence upon ferre-doxin as the sole natural electron donor. Meekset al. (1983) have observed that in comparisonwith unicellular cyanobacteria, Anabaena strainshave a high Km of 26–30 mmoles (as compared to1–4 mmoles for Synechococcus). The set of strainsanalyzed in the present study exhibited significantvariability for this parameter. Similar reports onAnabaena strains are available in literature (Me-konnen et al., 2002).
The selection of strains possessing a non-repres-sible nitrogenase along with rapid growth andliberation of substantial amount of ammonia (pre-sumably due to defective GS) is an importantstrategy for maximizing crop yields through cyano-bacterial nitrogen fixation (Rai and Prakasham,1991). Strains evaluated in our study such as A14(from Jammu & Kashmir soil samples), A10 (isolatedfrom soil samples of West Bengal), showing highammonia excretion coupled with moderate ARA(but high GS), indicated that a large amount ofN-fixed is released as ammonia. Such strains canhave tremendous potential as biofertilizers. A11(from West Bengal) is also of interest as it has a highheterocyst frequency of 9.8%. Thus, the presentinvestigation has aided in identifying region-speci-fic strains with good potential as biofertilizers.
Analysis of DNA base composition of nearly 200cyanobacterial strains (Herdman et al., 1979)was found to be of limited value in establishingtaxonomic relationships. Techniques based onhighly repetitive sequence have shown to be veryvaluable in this regard. Fingerprinting of therepetitive extragenic palindromic sequences (REP)or enterobacterial repetitive intergenic consensus(ERIC) elements have provided useful information
for identification of symbiotic and free-livingcyanobacteria (Rasmussen and Svenning, 1998). Incyanobacteria, a unique and distinct family ofrepetitive sequences, the STRR sequences, weredescribed (Mazel et al., 1990) in a number ofcyanobacterial genera and species, all belonging toheterocystous bacteria. Additionally a 37 bp LTRRsequences have been identified in Anabaena strainPCC7120 (Masepohl et al., 1996). In the presentinvestigation, analysis of molecular marker profileshelped in effectively discriminating the Anabaenastrains. Among the four PCR primers utilized, LTRRproved to be most informative, generating max-imum number of polymorphic amplicons and sevenunique banding patterns. However, HipTG was ableto differentiate these sets of strains.
HipTG, a primer, constructed on the basis of aconsensus sequence (an octa palindrome desig-nated Hip1), is known to be frequently occurring ina large number of cyanobacterial strains (Robinsonet al., 1995) and has been used as a cyanobacterialtyping technique akin to repetitive sequence-oriented PCR techniques. The presence of Hip1 ina number of cyanobacterial genomes permits itsutilization for fingerprinting based on amplificationof DNA between adjacent repeated Hip1 sequencesin the chromosomal DNA. However, its very highfrequency, leading to large number of discreteamplification products, has limited its use. There-fore, primers have been constructed in which twoadditional bases were added to the 30 end of Hip1sequence. Such extended Hip1 primers (like HipTGin the present study) have proved useful indistinguishing several species of cyanobacteria(Smith et al., 1998). In addition to LTRR and HipTG,two other primers – STRRmod and STRR1A – alsoprovided useful information for diversity analysesin the present study. Rasmussen and Svenning(1998) developed a fingerprinting technique basedon STRR and LTRR sequences, which was useful inrevealing heterogeneity among symbiotic isolatesfrom Gunnera species besides distinguishing themfrom the free-living isolates. Van Coppenolle et al.(1993) classified 25 Azolla acessessions using ran-dom primers, and showed that three primers weresufficient for routine identification. In the presentstudy, two primers – LTRR and HipTG – wereinformative enough to unambiguously differentiateall the 13 Anabaena strains.
Analysis of the genetic diversity in Anabaenastrains using the molecular marker dataset, basedon the 34 polymorphic amplicons helped in reveal-ing the genetic relationships among the strains. Thecophenetic correlation coefficient of 0.88 indicateda good fit between the dendrogram and the geneticsimilarity matrix based on Jaccard’s coefficient.

ARTICLE IN PRESS
Morphological, physiochemical and molecular characterization of Anabaena strains 201
Cluster analysis using UPGMA, supported by boot-strap analysis to ascertain the statistical support tothe internal branches in the dendrogram, revealedtwo distinct clusters and an outlier (A2). The highdegree of similarity between A5 (PCC 7120) and A7(ATCC 29414), the reference/type strains analyzedin this study, was further confirmed by PCA.Although cluster analysis and PCA utilize data setdifferently, their combined use proved highlyeffective in revealing the patterns of geneticrelationships among the Anabaena strains. Theanalyses also helped in distinguishing A6 (ATCC29413), another reference/type strain. In general,no correlation of the groupings with the geogra-phical origin of the isolate could be observed.However, the genetic relatedness of A5 and A7corresponds with their highly similar physio-bio-chemical characteristics, in terms of accumulationof carotenoids, PC and total PBS, besides enzymaticactivities of ARA and NR. Interestingly, strains A1and A15, which were found to be in the same groupwith ~40% similarity, exhibited similar PC and totalPBS values.
The present investigation clearly brought out thediversity existing among the Anabaena strainsanalyzed in terms of various morphological andbiochemical attributes. Molecular polymorphismsfurther aided in effective differentiation of theselected strains, besides revealing their geneticrelationships. A combination of attributes, includ-ing morphological (trichome form/shape, vegeta-tive cell width, position/shape/size of heterocystsand shape of terminal cell) and biochemical/physiological (PBS, nitrogenase, GS and ammoniaexcreting activity), besides molecular profiling,could provide useful taxonomic criteria for distin-guishing among the isolates belonging to the genusAnabaena, when analyzed under a uniform set ofconditions. Such attributes should also be includedin the descriptions of the genus/species/strains intaxonomic treatises, for providing a more compre-hensive characterization of the members placedunder this genus. This would facilitate not onlyselection of particular strain(s) for in-depth ana-lyses of various metabolic processes but also fortheir effective utilization for diverse purposes.
Acknowledgements
R. Kumar and A. Sood are thankful to IndianCouncil of Agricultural Research (ICAR) and Councilof Scientific and Industrial Research (CSIR), respec-tively, for providing financial assistance in the formof Research Fellowship.
References
Anagnostidis, K., Komarek, J., 1985. Modern approach tothe classification system of cyanophytes. I. Introduc-tion. Arch. Hydrobiol. (Suppl. 71) (Algol. Stud. 38/39),291–302.
Anand, N., 1979. Studies on the genus Anabaena (Bory).II. A key to the identification of some Anabaenaspecies in culture. Acta Bot. Indica 7, 22–28.
Bagchi, S.N., 1994. Cyanobacterial nitrate reduction:process and regulation. Proc. Indian Sci. Acad. 60,243–254.
Bryant, D.A., 1982. Phycoerythrocyanin and phycoery-thrin: properties and occurrence in cyanobacteria.J. Gen. Microbiol. 128, 835–844.
Castenholz, R.W., 2001. Phylum BX. Cyanobacteria –
oxygenic photosynthetic bacteria. In: Boone, D.R.,Castenholz, W.R. (Eds.), Bergey’s Manual of Systema-tic Bacteriology, second ed. Springer, Berlin.
De, P.K., 1939. The role of blue green algae in nitrogenfixation in rice fields. Proc. Roy. Soc. London Ser. B127, 121–139.
Desikachary, T.V., 1959. Cyanophyta. ICAR Monograph onAlgae. ICAR, New Delhi, India 686pp.
Desikachary, T.V., 1970. Taxonomy of blue-green algae:problems and prospects. Schweiz. Z. Hydrobiol. 32,490–494.
Desikachary, T.V., Anand, N., 1974. Preliminary studies onthe genus Anabaena Bory. In: Proceedings of theSecond International Symposium on Phycology: Tax-onomy of Algae. University of Madras, Madras.
Drouet, F., 1968. Revision of the Classification of theOscillatoriaceae. Monograph Acad. Nature Sci. Phila-delphia. Fulton Press, Lancaster, Pennsylvania.
Flores, E., Herrero, A., 1994. Assimilatory nitro-gen metabolism and its regulation. In: Bryant, D.A.(Ed.), The Molecular Biology of Cyanobacteria.Kluwer Academic Publishers, Dordrecht, Netherlands,pp. 769–823.
Fritsch, F.E., 1949. The genus Anabaena with specialreference to the species recorded from the areasadjacent to Asiatic mainland. J. Indian Bot. Soc. 28,135–161.
Geitler, L., 1932. Cyanophyceae. In: Robenhorst, E. (Ed.),Kryptogamen flora van Deutschland, Osterrich und derSchweiz, vol. XIV. Academische Verlags, Leipzig.
Giovannoni, S.J., Britschgi, T.B., Moyer, C.L., Field, K.G.,1990. Genetic diversity in sargasso sea bacterioplank-ton. Nature 345, 60–63.
Gugger, M., Lyra, L., Henriksen, P., Coute, A., Humbert,J.F., Sivonen, K., 2002. Phylogenetic comparison ofthe cyanobacterial genera Anabaena and Aphanizo-menon. Int. J. Syst. Evol. Microbiol. 52, 1867–1880.
Herdman, M., Janvier, M., Rippka, R., Stanier, R.Y., 1979.Genome size of cyanobacteria. J. Gen. Microbiol. 111,73–85.
Hiroki, M., Shimizu, A.L.R., Watanabe, M., Watanabe,M.M., 1998. Development of database system usefulfor identification of Anabaena spp. (Cyanobacteria).Phycol. Res. 46, 85–93.

ARTICLE IN PRESS
R. Prasanna et al.202
Holton, R.W., Blecker, H.H., Stevens, T.S., 1968. Fattyacid in blue-green algae: possible relation to phylo-genetic position. Science 160, 545–547.
Jaccard, P., 1908. Nouvelles recherches sur la distribu-tion florale. Bull. Soc. Vaud. Nat. 44, 223–270.
Jensen, A., 1978. Chlorophylls and carotenoids. In:Hellebust, J.A., Craige, J.S. (Eds.), PhycologicalMethods: Physiological and Biochemical Methods.Cambridge University Press, Cambridge, pp. 59–70.
Komarek, J., Anagnostidis, K., 1989. Modern approach tothe classification system of cyanophytes. 4. Nosto-cales. Arch. Hydrobiol. (Suppl. 83) (Algol. Stud.) 56,247–345.
Li, R., 1998. Taxonomic studies on planktonic species ofAnabaena (Cyanobacteria). Ph.D. thesis, TsukubaUniversity, Tsukuba.
MacColl, R., Guard-Friar, D., 1987. Phycobiliproteins.CRC Press, Boca Raton, FL.
Masepohl, B., Gorlitz, K., Bohmen, H., 1996. Longtandemly repetitive (LTRR) sequences in the filamen-tous cyanobacterium Anabaena sp. PCC 7120. Bio-chem. Biophys. Acta 1307, 26–30.
Mazel, D., Houmard, J., Castets, A.M., Tandeau deMarsac, N., 1990. Highly repetitive DNA sequencesin cyanobacterial genomes. J. Bacteriol. 172,2755–2761.
McGregor, C.E., Lambert, C.A., Greyliing, M.M., Louw,J.H., Warnich, L., 2000. A comparative assessment ofDNA fingerprinting techniques (RAPD, ISSR, AFLP, SSR)in tetraploid potato (Solanum tuberosum L.) germ-plasm. Euphytica 113, 135–144.
Meeks, J.C., Wycoff, K.L., Chapman, J.S., Enderlin, C.S.,1983. Regulation of expression of nitrate and dinitro-gen assimilation by Anabaena species. Appl. Environ.Microbiol. 54, 1351–1359.
Mekonnen, A.E., Prasanna, R., Kaushik, B.D., 2002.Response of Anabaena species to different nitrogensources. Acta Biol. Hungarica 53, 367–380.
Mohammadi, S.A., Prasanna, B.M., 2003. Analysis ofgenetic diversity in crop plants – salient statisticaltools and considerations. Crop Sci. 43, 1235–1248.
Pabby, A., Prasanna, R., Nayak, S., Singh, P.K., 2003.Physiological characterization of the cultured andfreshly isolated endosymbionts from different speciesof Azolla. Plant Physiol. Biochem. 41, 73–79.
Rai, A.N., Prakasham, R., 1991. Transport of inorganicnitrogen in cyanobacteria and its relevance in use ofcyanobacteria as bofertilizers. In: Dutta, S.K., Sloger,C. (Eds.), Biological Nitrogen Fixation Associated withRice Production. Oxford and IBH, India, pp. 177–189.
Rasmussen, U., Svenning, M.M., 1998. Fingerprinting ofcyanobacteria based on PCR with primers derivedfrom short and long tandemly repeated repetitivesequences. Appl. Environ. Microbiol. 64, 265–272.
Rippka, R., Herdman, M., 1992. Pasteur Culture Collec-tion of Cyanobacteria: Catalogue and TaxonomicHandbook: Catalogue of Strains, vol. I. InstitutePasteur, Paris 103pp.
Rippka, R., Deruelles, J., Waterbury, J.B., Herdman, M.,Stanier, R.Y., 1979. Generic assignments strain his-tories and properties of pure cultures of cyanobacter-ia. J. Gen. Microbiol. 111, 1–61.
Rippka, R., Castentholtz, R.W., Iteman, I., Herdman, M.,2001. Form-genus I. Anabaena Bory. In: Boone, D.R.,Castenholz, W.R. (Eds.), Bergey’s Manual of Sys-tematic Bacteriology, second ed. Springer, Berlin,pp. 566–568.
Robinson, N.J., Robinson, P.J., Gupta, A., Bleasby, J.A.,Whitton, B.A., Morby, A., 1995. Singular over-repre-sentation of an octameric palindrome, HIP1, in DNAfrom many cyanobacteria. Nucl. Acids Res. 23,729–773.
Roychoudhury, P., Kaushik, B.D., Singh, D., Venkatara-man, G.S., 1986. Pearson distribution of nitrogenfixing potential among Nostoc strains. Curr. Sci. 55,183–185.
Sidler, W.A., 1994. Phycobilisome and phycobiliproteinstructures. In: Bryant, D.A. (Ed.), The MolecularBiology of Cyanobacteria. Kluwer Academic, Dor-drecht, Netherlands, pp. 139–216.
Smith, J.K., Parry, J.D., Day, J.G., Smith, R.J., 1998. APCR technique based on Hip1 interspersed repetitivesequence distinguishes cyanobacterial species andstrains. Microbiology 144, 2791–2801.
Sneath, P.H.A., Sokal, R.R., 1973. Numerical Taxonomy.W.H. Truman, San Francisco.
Stanier, R.Y., Kunisawa, R., Mandel, M., Cohen-Bazire,G., 1971. Purification and properties of unicellularblue-green algae (order Chroococcales). Bacteriol.Rev. 35, 171–205.
Stulp, B.K., 1982. Morphological variability of Anabaenastrains (Cyanophyceae) under different culture condi-tions. Arch. Hyrdrobiol. 63, 165–176.
Stulp, B.K., Stam, W.T., 1982. General morphology andakinete germination of a number of Anabaena strains(Cyanophyceae) in culture. Arch. Hydrobiol. Suppl. 63(Algol, Stud. 30), 35–52.
Stulp, B.K., Stam, W.T., 1984. Genotypic relationshipsbetween strains of Anabaena (Cyanophyceae) andtheir correlation with morphological affinities. Br.Phycol. J. 19, 287–301.
Van Coppenolle, B., Watanabe, I., Van Hove, C., Second,G., Huang, N., McCouch, S.R., 1993. Genetic diversityand phylogeny analysis of Azolla based on DNAamplification by arbitrary primers. Genome 36,686–693.
Venkataraman, G.S., 1981. Blue-green algae for riceproduction. A manual for its promotion. FAO BulletinNo. 46, FAO, Rome, 102pp.
Weir, B.S., 1990. Genetic Data Analysis. Sinauer Associ-ates, Sunderland.
Wolk, C.P., Ernst, A., Elhai, J., 1994. Heterocystmetabolism and development. In: Bryant, D.A.(Ed.), The Molecular Biology of Cyanobacteria.Kluwer Academic, Dordrecht, Netherlands, pp.769–823.
Copyright © 2022 FDOKUMEN




















