Differential expression of gap-junction gene connexin 31 in seminiferous epithelium of rat testes
Upload
independentCategory
view
0download
0
Sh
QYa
b
c
d
e
h
••••
a
ARRA
KCGGN
1
hb
h0
Neuroscience Letters 566 (2014) 194–199
Contents lists available at ScienceDirect
Neuroscience Letters
jo ur nal ho me p age: www.elsev ier .com/ locate /neule t
uppression of spinal connexin 43 expression attenuates mechanicalypersensitivity in rats after an L5 spinal nerve injury
ian Xua,b,1, Yong-Kwan Cheongb,c,1, Shao-Qiu Heb, Vinod Tiwarib, Jian Liud,un Wange, Srinivasa N. Rajab, Jinheng Lia, Yun Guanb, Weiyan Lid,∗
Department of Clinical Pharmacology, Jinling Hospital, School of Medicine, Nanjing University, Nanjing, Jiangsu, ChinaDepartment of Anesthesiology and Critical Care Medicine, Johns Hopkins University, School of Medicine, Baltimore, MD, USADepartment of Anesthesiology and Pain Medicine, School of Medicine, Wonkwang University, Iksan, Republic of KoreaDepartment of Anesthesiology, Jinling Hospital, School of Medicine, Nanjing University, Nanjing, Jiangsu, ChinaDepartment of Anesthesiology, Beijing Chaoyang Hospital, Capital Medical University, Beijing 100020, China
i g h l i g h t s
Intrathecal Cx43 siRNA attenuated neuropathic mechanical hypersensitivity in rats.Pain inhibition by Cx43 siRNA was correlated with downregulation of spinal Cx43 expression.Cx43 siRNA treatment did not decrease protein levels of Cx36 and GFAP in the spinal cord.The enhanced function of spinal astroglial gap junctions may be important to neuropathic pain.
r t i c l e i n f o
rticle history:eceived 30 January 2014eceived in revised form 27 February 2014ccepted 1 March 2014
eywords:onnexin 43liaap junctioneuropathic pain
a b s t r a c t
Activation of spinal astrocytes may contribute to neuropathic pain.Adjacent astrocytes can make directcommunication through gap junctions formed by connexin 43 (Cx43) in the central nervous system. Yet,the role of spinal astroglial gap junctions in neuropathic pain is not fully understood. Since Cx43 is theconnexin isoform expressed preferentially in astrocytes in the spinal cord, we used a small interferingRNA (siRNA) approach to examine whether suppression of spinal Cx43 expression inhibits mechanicalhypersensitivity in rats after an L5 spinal nerve ligation (SNL). SNL rats were administered intrathecalCx43 siRNA (3 �g/15 �l, twice/day) or an equal amount of mismatch siRNA (control) on days 14–17post-SNL. Cx43 siRNA, but not mismatch siRNA, alleviated mechanical hypersensitivity in SNL rats. Fur-thermore, Western blot analysis showed that the pain inhibition induced by Cx43 siRNA correlated withdownregulation of Cx43 expression, but not that of Cx36 (the neuronal gap junction protein) or glialfibrillary acidic protein (GFAP, a marker for reactive astrocytes) in the spinal cord of SNL rats. Western
blot analysis and immunohistochemistry also showed that SNL increased GFAP expression, but decreasedCx43 expression, in spinal cord. Our results provide direct evidence that selective suppression of spinalCx43 after nerve injury alleviates neuropathic mechanical hypersensitivity. These findings suggest thatin the spinal cord, the enhanced function of astroglial gap junctions, especially those formed by Cx43,may be important to neuropathic pain in SNL rats.. Introduction
Gap junctions allow second messengers, ions, and smallydrophilic molecules to pass freely and rapidly between neigh-oring cells in the central nervous system (CNS). They are usually
∗ Corresponding author. Tel.: +86 25 80860147; fax: +86 25 84806835.E-mail address: [email protected] (W. Li).
1 These authors contributed equally to this work.
ttp://dx.doi.org/10.1016/j.neulet.2014.03.004304-3940/© 2014 Elsevier Ireland Ltd. All rights reserved.
© 2014 Elsevier Ireland Ltd. All rights reserved.
formed by two hemichannels, each composed of a hexamer ofconnexin (Cx) protein and situated in the membrane of adjacentcells [1]. Cx43 is the connexin isoform expressed preferentially inastrocytes of the mammalian CNS [2–4]. Gap junctions composedof Cx43 may mediate the intercellular propagation of Ca2+ wavesbetween astrocytes, enabling these cells to generate rapid electrical
responses to neuronal activity [5,6].Increasing evidence suggests that gap junctions in astrocytesmay play an important role in various pathologic pain conditions[4,7,8], including neuropathic pain [3,9–11]. Studies suggest that

e Lett
ameineai[oaij
ttbtw
2
2
rwsasaU
2
brwIe
2
w1pm[fc
2
LrigobssnB
Q. Xu et al. / Neuroscienc
ctivation of astrocytes in the spinal cord may contribute to theaintenance of dorsal horn neuronal hyperexcitability that leads to
xaggerated spinal nociceptive transmission after tissue and nervenjury [12,13]. For example, patients with chemotherapy-inducedeuropathy exhibit increases in both astrocyte activation and Cx43xpression [4]. Blocking gap junctions by systemic or intrathecaldministration of carbenoxolone (CBX), a gap junction decoupler,nhibits neuropathic pain behavior in a variety of animal models4,7,14–16]. However, because CBX is nonselective and could havether effects on neuronal excitability [17,18], the specificity of drugction toward glial gap junctions in previous pharmacologic stud-es is uncertain. Consequently, the roles of spinal astroglial gapunctions in the maintenance of neuropathic pain are not fully clear.
In this study, our goal was to examine whether spinal Cx43 con-ributes to the manifestation of neuropathic pain in rats subjectedo L5 spinal nerve ligation (SNL). Because selective glial gap junctionlockers for animal studies are lacking, we knocked down gap junc-ion proteins using double-stranded small interfering RNA (siRNA),hich causes degradation of the cognate messenger RNA.
. Materials and methods
.1. Animals
Experiments were carried out in adult, male Sprague-Dawleyats (250–350 g, Harlan Laboratories, Inc., Indianapolis, IN). Effortsere made to minimize the number of animals used and their
uffering. All experimental procedures were conducted in strictccordance with the guidelines established by the Nanjing Univer-ity and Johns Hopkins University Animal Care and Use Committees consistent with the National Institutes of Health Guide for these of Experimental Animals.
.2. L5 SNL
The SNL model was produced as described previously [19]. Inrief, the rats were anesthetized with isoflurane (2%, Abbott Labo-atories, North Chicago, IL), and the left L5 spinal nerve was ligatedith a 6-0 silk suture and cut distally under aseptic conditions.
n the sham surgery group, the surgical procedure was identical,xcept that the left L5 spinal nerve was not ligated.
.3. Behavioral tests
Hypersensitivity of rats to punctuate mechanical stimulationith von Frey filaments (0.38, 0.57, 1.23, 1.83, 3.66, 5.93, 9.13,
3.1 g) was determined with the up-down method as describedreviously [20]. The paw withdrawal threshold (PWT) was deter-ined according to the method and formula provided by Dixon
21]. To minimize experimenter bias, the investigator who per-ormed the behavioral tests was blinded to the drug treatmentonditions.
.4. Western blotting
Ipsilateral and contralateral lumbar spinal segments (including4 and L5 segments) of SNL and sham-operated rats were sepa-ated and homogenized for immunoblotting. The tissues were lysedn ice-cold RIPA buffer (50 mM Tris [pH 7.5], 150 mM NaCl, 10%lycerol, 0.1% Triton X-100, 0.5 mg/ml BSA). After centrifugationf the homogenates, the protein concentration was determinedy using a detergent-compatible protein assay with a bovine
erum albumin standard. Samples were separated on a 7.5% (w/v)odium dodecyl sulfate-polyacrylamide gel and transferred onto aitrocellulose membrane (Amersham, Pittsburgh, PA) with a Trans-lot Transfer Cell system (Bio-Rad, Hercules, CA). Membranesers 566 (2014) 194–199 195
were incubated with the indicated primary antibody overnightat 4 ◦C, and immunoreactivity was detected by enhanced chemi-luminescence (ECL, Amersham). We used antibodies against Cx43(1:8000, Sigma-Aldrich, St. Louis, MO, Cat. #: C6219), Cx36 (1:1000,Life technologies, Grand Island, NY, Cat. #: 51-6200), glial fibril-lary acidic protein (GFAP; 1:2000, Millipore, Billerica, MA, Cat. #MAB360), and �-actin (1:10,000, Chemicon, Temecula, CA). Wethen quantified the intensity of immunoreactive bands of intereston the autoradiograms using Image J 1.46r software (NIH, Bethesda,MD). Actin staining was used as an internal control for proteinloading.
2.5. Immunofluorescence
Animals were anesthetized with pentobarbital and perfusedwith 20 ml of 0.1 M phosphate-buffered saline (PBS; pH 7.4, 4 ◦C)followed by 25 ml of fixative (4% formaldehyde and 14% [v/v]saturated picric acid in PBS; 4 ◦C). Lumbar spinal cord tissueswere cryoprotected in 20% sucrose (24 h) before being serially cutinto 15-�m sections and placed onto slides. The slides were pre-incubated in blocking solution (5% normal donkey serum, 0.2%Triton X-100 in PBS) for 1 h at room temperature and then incu-bated overnight with the primary antibody at 4 ◦C. The primaryantibody was diluted to its final working concentration in PBS [pH7.4] with 1% BSA, 0.01% sodium azide, and 0.3% Triton X-100. Theslides were then incubated in secondary antibody at room temper-ature for 45 min and covered with Fluoroshield histology mountingmedium (Sigma-Aldrich). The primary antibodies used were rabbitanti-Cx43 (1:2000, Sigma-Aldrich, Cat. #: C6219), mouse anti-GFAP (1:5000, Millipore, Cat. #: MAB360), and mouse anti-OX42(1:2000, Millipore, Cat. # CBL1512). The secondary antibodies usedwere Rhodamine Red-X-conjugated AffiniPure donkey anti-rabbitIgG(H + L) (1:100, Jackson ImmunoResearch, West Grove, PA, Cat.#: 711-295-152) and Fluorescein-conjugated AffiniPure donkeyanti-mouse IgG(H + L) (1:100, Jackson ImmunoResearch, Cat. #:711-095-151).
Digitized images were captured using Olympus BX51 micro-scope system (Melville, NY). We then quantified the integratedoptical density (IOD) of immunostaining from the dorsal horn oflumbar spinal cord slice (three to four sections/animal) using ImageJ 1.46r software (NIH, Bethesda, MD). Similar areas were examinedin different animals. The background signal in each slice was sub-tracted. The mean IOD of sham-operated rats was calculated, andthe ratio of IOD in SNL rats was then normalized to the mean valueof sham-operated rats and illustrated as fold of sham for the pur-pose of comparison. Data were analyzed by an investigator blindedto experimental group.
2.6. siRNA
ON-TARGET plus SMARTpool Cx43 siRNA and mismatch siRNA(control) were purchased from Thermo Scientific (Pittsburgh, PA).Single deprotected strands were resuspended with an isotonicbuffer (100 mM potassium acetate, 30 mM HEPES-KOH, 2 mM mag-nesium acetate [pH 7.4 at 37 ◦C]; 26 mM NaCl was added to producea non-irritating, isotonic solution) to a concentration of 1 �g/�l.The strands were incubated at 90 ◦C for 5 min and then at 37 ◦Cfor 1 h. siRNAs were prepared immediately before administrationby mixing the RNA solution (1 �g/�l in annealing buffer) with thetransfection reagent i-Fect (v/v: 1/4; Neuromics, Edina, MN) to a
final siRNA/lipid complex concentration of 0.2 �g/�l. Cx43 siRNAor mismatch siRNA was infused into SNL rats (15 �l/injection, twicedaily) through an implanted intrathecal catheter (PE-10 tubing) for4 consecutive days.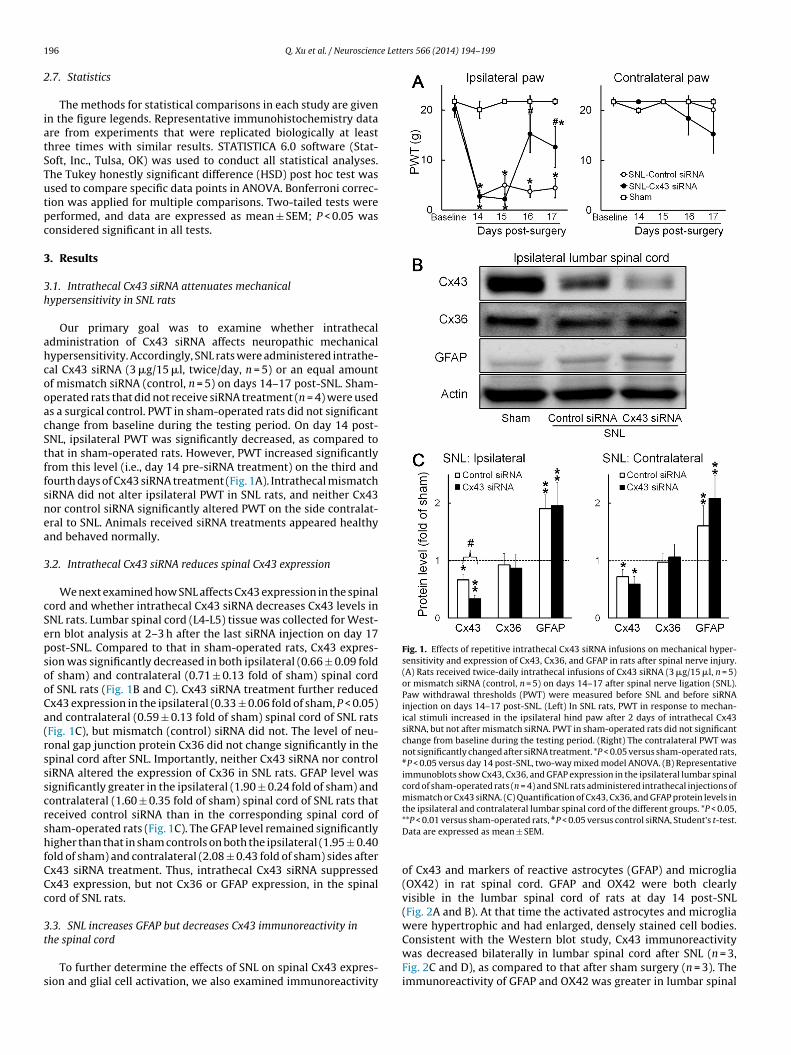
1 e Letters 566 (2014) 194–199
2
iatSTutpc
3
3h
ahcooacStffsnea
3
cSepsooCa(rssscrshfCCc
3t
s
Fig. 1. Effects of repetitive intrathecal Cx43 siRNA infusions on mechanical hyper-sensitivity and expression of Cx43, Cx36, and GFAP in rats after spinal nerve injury.(A) Rats received twice-daily intrathecal infusions of Cx43 siRNA (3 �g/15 �l, n = 5)or mismatch siRNA (control, n = 5) on days 14–17 after spinal nerve ligation (SNL).Paw withdrawal thresholds (PWT) were measured before SNL and before siRNAinjection on days 14–17 post-SNL. (Left) In SNL rats, PWT in response to mechan-ical stimuli increased in the ipsilateral hind paw after 2 days of intrathecal Cx43siRNA, but not after mismatch siRNA. PWT in sham-operated rats did not significantchange from baseline during the testing period. (Right) The contralateral PWT wasnot significantly changed after siRNA treatment. *P < 0.05 versus sham-operated rats,#P < 0.05 versus day 14 post-SNL, two-way mixed model ANOVA. (B) Representativeimmunoblots show Cx43, Cx36, and GFAP expression in the ipsilateral lumbar spinalcord of sham-operated rats (n = 4) and SNL rats administered intrathecal injections ofmismatch or Cx43 siRNA. (C) Quantification of Cx43, Cx36, and GFAP protein levels inthe ipsilateral and contralateral lumbar spinal cord of the different groups. *P < 0.05,
96 Q. Xu et al. / Neuroscienc
.7. Statistics
The methods for statistical comparisons in each study are givenn the figure legends. Representative immunohistochemistry datare from experiments that were replicated biologically at leasthree times with similar results. STATISTICA 6.0 software (Stat-oft, Inc., Tulsa, OK) was used to conduct all statistical analyses.he Tukey honestly significant difference (HSD) post hoc test wassed to compare specific data points in ANOVA. Bonferroni correc-ion was applied for multiple comparisons. Two-tailed tests wereerformed, and data are expressed as mean ± SEM; P < 0.05 wasonsidered significant in all tests.
. Results
.1. Intrathecal Cx43 siRNA attenuates mechanicalypersensitivity in SNL rats
Our primary goal was to examine whether intrathecaldministration of Cx43 siRNA affects neuropathic mechanicalypersensitivity. Accordingly, SNL rats were administered intrathe-al Cx43 siRNA (3 �g/15 �l, twice/day, n = 5) or an equal amountf mismatch siRNA (control, n = 5) on days 14–17 post-SNL. Sham-perated rats that did not receive siRNA treatment (n = 4) were useds a surgical control. PWT in sham-operated rats did not significanthange from baseline during the testing period. On day 14 post-NL, ipsilateral PWT was significantly decreased, as compared tohat in sham-operated rats. However, PWT increased significantlyrom this level (i.e., day 14 pre-siRNA treatment) on the third andourth days of Cx43 siRNA treatment (Fig. 1A). Intrathecal mismatchiRNA did not alter ipsilateral PWT in SNL rats, and neither Cx43or control siRNA significantly altered PWT on the side contralat-ral to SNL. Animals received siRNA treatments appeared healthynd behaved normally.
.2. Intrathecal Cx43 siRNA reduces spinal Cx43 expression
We next examined how SNL affects Cx43 expression in the spinalord and whether intrathecal Cx43 siRNA decreases Cx43 levels inNL rats. Lumbar spinal cord (L4-L5) tissue was collected for West-rn blot analysis at 2–3 h after the last siRNA injection on day 17ost-SNL. Compared to that in sham-operated rats, Cx43 expres-ion was significantly decreased in both ipsilateral (0.66 ± 0.09 foldf sham) and contralateral (0.71 ± 0.13 fold of sham) spinal cordf SNL rats (Fig. 1B and C). Cx43 siRNA treatment further reducedx43 expression in the ipsilateral (0.33 ± 0.06 fold of sham, P < 0.05)nd contralateral (0.59 ± 0.13 fold of sham) spinal cord of SNL ratsFig. 1C), but mismatch (control) siRNA did not. The level of neu-onal gap junction protein Cx36 did not change significantly in thepinal cord after SNL. Importantly, neither Cx43 siRNA nor controliRNA altered the expression of Cx36 in SNL rats. GFAP level wasignificantly greater in the ipsilateral (1.90 ± 0.24 fold of sham) andontralateral (1.60 ± 0.35 fold of sham) spinal cord of SNL rats thateceived control siRNA than in the corresponding spinal cord ofham-operated rats (Fig. 1C). The GFAP level remained significantlyigher than that in sham controls on both the ipsilateral (1.95 ± 0.40
old of sham) and contralateral (2.08 ± 0.43 fold of sham) sides afterx43 siRNA treatment. Thus, intrathecal Cx43 siRNA suppressedx43 expression, but not Cx36 or GFAP expression, in the spinalord of SNL rats.
.3. SNL increases GFAP but decreases Cx43 immunoreactivity in
he spinal cordTo further determine the effects of SNL on spinal Cx43 expres-ion and glial cell activation, we also examined immunoreactivity
**P < 0.01 versus sham-operated rats, #P < 0.05 versus control siRNA, Student’s t-test.Data are expressed as mean ± SEM.
of Cx43 and markers of reactive astrocytes (GFAP) and microglia(OX42) in rat spinal cord. GFAP and OX42 were both clearlyvisible in the lumbar spinal cord of rats at day 14 post-SNL(Fig. 2A and B). At that time the activated astrocytes and microgliawere hypertrophic and had enlarged, densely stained cell bodies.Consistent with the Western blot study, Cx43 immunoreactivity
was decreased bilaterally in lumbar spinal cord after SNL (n = 3,Fig. 2C and D), as compared to that after sham surgery (n = 3). Theimmunoreactivity of GFAP and OX42 was greater in lumbar spinal
Q. Xu et al. / Neuroscience Letters 566 (2014) 194–199 197
Fig. 2. Spinal nerve injury increases glial activation but decreases Cx43 immunoreactivity in the spinal cord. (A) In rats on day 14 after spinal nerve ligation (SNL), astrocytesin the spinal cord exhibited increased immunoreactivity of GFAP, especially on the side ipsilateral (left) to nerve injury. (B) Meanwhile, microglia exhibited increasedimmunoreactivity of OX42. Compared to spinal astrocytes and microglia in sham-operated rats, those in SNL rats were hypertrophic. (C) In contrast, Cx43 immunoreactivitywas decreased on both sides of the spinal cord in SNL rats compared to that in sham-operated rats. (D) Quantification of Cx43, GFAP, and OX42 immunoreactivity in theipsilateral and contralateral dorsal horn. The immunostaining density of SNL rats (three to four sections/rat, n = 3) was normalized to that of sham-operated rats (n = 3).*P < 0.05 versus sham-operated rats, Student’s t-test. Data are expressed as mean ± SEM. (E) The overlap (yellow) of GFAP (green) and Cx43 (red) immunoreactivity wasincreased after SNL. (F) OX42 (green) and Cx43 (red) did not colocalize. (For interpretation of the references to color in this figure legend, the reader is referred to the webv
ceTbic
4
cYut
ersion of this article.)
ord of SNL rats than in lumbar spinal cord of sham-operated rats,specially on the side ipsilateral to the injury (Fig. 2A, B and D).he Cx43 immunoreactivity overlapped with that of GFAP (Fig. 2E),ut not OX42 (Fig. 2F), in SNL rats. These findings suggest that SNL
ncreases the activation of microglia and astrocytes but paradoxi-ally reduces the expression of Cx43 in the spinal cord.
. Discussion
Preclinical studies suggest that spinal glial cells, especially astro-
ytes, may play an important role in neuropathic pain [8,22,23].et, the roles of glial gap junctions in neuropathic pain are not fullynderstood. Because there is no selective blocker of glial gap junc-ions, conventional pharmacologic studies have not yet provideddirect evidence that pain inhibition from drug treatment (e.g., CBX)occurs by blockade of glial gap junctions. Cx43 is the gap junctionprotein that is preferentially expressed in astrocytes of the CNS[24,25]. In our study, knockdown of Cx43 by intrathecal adminis-tration of Cx43 siRNA alleviated mechanical hypersensitivity in SNLrats. Further, the pain inhibition correlated with downregulationof spinal Cx43, but not the neuronal gap junction protein Cx36 orreactive astrocyte marker GFAP. These results may represent directevidence that selective suppression of spinal astroglial gap junctioncan alleviate neuropathic mechanical hypersensitivity.
Previous studies have shown that Cx43 is upregulated in thetrigeminal ganglia after trigeminal nerve injury and in trigeminalnucleus after tooth pulp inflammation [26–28]. It was postulatedthat upregulation of Cx43 may increase the formation of new gap

1 e Lett
jItie(otshacsoitecsdcinoecftigpm
bwCmTtFn[aiGwcaFgtinsoimewr
olfiio
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
98 Q. Xu et al. / Neuroscienc
unction connections and hence increase membrane permeability.n this regard, it is surprising that our Western blot and immunohis-ochemistry studies showed that spinal Cx43 was downregulatedn rats after SNL. The mechanism for the decreased spinal Cx43xpression after SNL is unclear. It is possible that different insultse.g., nerve injury versus tissue inflammation) and different typesf nerve injury (e.g., SNL versus chronic constriction injury ofrigeminal nerve) may differentially regulate spinal Cx43 expres-ion. Intriguingly, lipopolysaccharide-activated microglia, mediumarvested from activated microglia, and mixed cytokines (TNF-�nd IL-1�) each downregulated Cx43 expression when added toultured astrocytes [29]. Previous studies showed that activation ofpinal microglia is prominent after nerve injury [9,30,31]. Likewise,ur immunohistochemistry study showed that immunoreactiv-ty of OX42, a marker of reactive microglia, was increased inhe spinal cord of SNL rats. Accordingly, future studies shouldxamine whether reactive microglia also release pro-inflammatoryytokines in spinal cord that lead to decreased Cx43 expres-ion, as observed in vitro [29]. Although Cx43 expression wasecreased in the spinal cord after SNL, activation of spinal astro-ytes was increased (i.e., increased GFAP level). This finding maymply a functional enhancement of astroglial gap junctions aftererve injury. Intriguingly, a previous study showed that additionf activated microglia or mixed cytokines downregulated Cx43xpression but increased the open probability of hemichannels inultured astrocytes [29]. This finding may partially explain whyurther suppression of Cx43 expression by siRNA in SNL rats ledo inhibition of mechanical hypersensitivity. Our findings maymply that enhanced function, rather than increased expression, oflial gap junctions formed by Cx43 may contribute to neuropathicain in SNL rats, presumably by increasing the release of activeolecules.In a study by Ohara et al. [26], the pain inhibition produced
y Cx43 siRNA treatment in the trigeminal ganglia was associatedith decreased GFAP in the ganglia. Yet, in our study, intrathecalx43 siRNA alleviated mechanical hypersensitivity without nor-alizing the upregulated GFAP in the spinal cord of SNL rats.
he reason for this discrepancy is unclear, but it should be notedhat the functional status of astrocytes may change after injury.or example, whereas inhibiting Cx43 expression in the trigemi-al ganglion after injury attenuated trigeminal neuropathic pain26], the same treatment elicited pain hypersensitivity in normalnimals [27,32]. Glial activation could also be beneficial, depend-ng upon the stage of activation [12,33,34]. One possibility is thatFAP is a marker of reactive astrocytes but is not indicative ofhether the astrocytes are releasing pro- or anti-inflammatory
ytokines. Hence, as with microglia, astrocyte reactivity may notlways correlate with neuropathic pain manifestations [35–37].inally, the Cx43 expressed in satellite glial cells in dorsal rootanglia and trigeminal ganglia increases after nerve injury and con-ributes to pain hypersensitivity [16,38]. Inhibiting Cx43 expressionn the trigeminal ganglion was shown to attenuate trigeminaleuropathic pain [26]. Accordingly, we cannot exclude the pos-ibility that the pain inhibition induced by intrathecal injectionf Cx43 siRNA may also involve inhibition of Cx43 expressionn dorsal root ganglia, owing to diffusion of siRNA. Future study
ay determine the time when the suppression of spinal Cx43xpression by siRNA may diminish after the last treatment, andhether it is associated with pain behavioral changes in SNL
ats.In summary, our study showed that selective downregulation
f astroglial gap junction protein by intrathecal Cx43 siRNA ame-
iorates neuropathic mechanical hypersensitivity in SNL rats. Thisnding suggests that enhanced activity of astroglial gap junctionsn the spinal cord might play an essential role in the maintenancef neuropathic pain.
[
ers 566 (2014) 194–199
Acknowledgements
The authors thank Claire F. Levine, MS (Scientific Editor, Depart-ment of Anesthesiology/CCM, the Johns Hopkins University) forediting the manuscript. This study was supported by a researchgrant to Q.X. from the China Scholarship Council (Beijing, China)and a grant to Y.G. from the National Institutes of Health (NS70814,Bethesda, MD, USA).
References
[1] J.C. Herve, M. Derangeon, Gap-junction-mediated cell-to-cell communication,Cell Tissue Res. 352 (2013) 21–31.
[2] J.I. Nagy, J.E. Rash, Connexins and gap junctions of astrocytes and oligodendro-cytes in the CNS, Brain Res. Brain Res. Rev. 32 (2000) 29–44.
[3] M.J. Chen, B. Kress, X. Han, K. Moll, W. Peng, R.R. Ji, M. Nedergaard, Astro-cytic CX43 hemichannels and gap junctions play a crucial role in developmentof chronic neuropathic pain following spinal cord injury, Glia 60 (2012)1660–1670.
[4] S.Y. Yoon, C.R. Robinson, H. Zhang, P.M. Dougherty, Spinal astrocyte gap junc-tions contribute to oxaliplatin-induced mechanical hypersensitivity, J. Pain 14(2013) 205–214.
[5] S. Burra, J.X. Jiang, Regulation of cellular function by connexin hemichannels,Int. J. Biochem. Mol. Biol. 2 (2011) 119–128.
[6] C. Giaume, L. Venance, Intercellular calcium signaling and gap junctional com-munication in astrocytes, Glia 24 (1998) 50–64.
[7] L.E. Spataro, E.M. Sloane, E.D. Milligan, J. Wieseler-Frank, D. Schoeniger, B.M.Jekich, R.M. Barrientos, S.F. Maier, L.R. Watkins, Spinal gap junctions: potentialinvolvement in pain facilitation, J. Pain 5 (2004) 392–405.
[8] Y.J. Gao, R.R. Ji, Targeting astrocyte signaling for chronic pain, Neurotherapeu-tics 7 (2010) 482–493.
[9] Z.Y. Zhuang, P. Gerner, C.J. Woolf, R.R. Ji, ERK is sequentially activated in neu-rons, microglia, and astrocytes by spinal nerve ligation and contributes tomechanical allodynia in this neuropathic pain model, Pain 114 (2005) 149–159.
10] R. Vallejo, D.M. Tilley, L. Vogel, R. Benyamin, The role of glia and the immunesystem in the development and maintenance of neuropathic pain, Pain Pract.10 (2010) 167–184.
11] X.F. Wu, W.T. Liu, Y.P. Liu, Z.J. Huang, Y.K. Zhang, X.J. Song, Reopening of ATP-sensitive potassium channels reduces neuropathic pain and regulates astroglialgap junctions in the rat spinal cord, Pain 152 (2011) 2605–2615.
12] E.D. Milligan, L.R. Watkins, Pathological and protective roles of glia in chronicpain, Nat. Rev. Neurosci. 10 (2009) 23–36.
13] R.R. Ji, T. Berta, M. Nedergaard, Glia and pain: is chronic pain a gliopathy? Pain(2013).
14] D.H. Roh, S.Y. Yoon, H.S. Seo, S.Y. Kang, H.J. Han, A.J. Beitz, J.H. Lee, Intrathecalinjection of carbenoxolone, a gap junction decoupler, attenuates the inductionof below-level neuropathic pain after spinal cord injury in rats, Exp. Neurol.224 (2010) 123–132.
15] H. Wang, Y. Cao, C.Y. Chiang, J.O. Dostrovsky, B.J. Sessle, The gap junction blockercarbenoxolone attenuates nociceptive behavior and medullary dorsal horn cen-tral sensitization induced by partial infraorbital nerve transection in rats, Pain(2013).
16] R.A. Warwick, M. Hanani, The contribution of satellite glial cells tochemotherapy-induced neuropathic pain, Eur. J. Pain (2012).
17] K.R. Tovar, B.J. Maher, G.L. Westbrook, Direct actions of carbenoxolone onsynaptic transmission and neuronal membrane properties, J. Neurophysiol. 102(2009) 974–978.
18] A.N. Chepkova, O.A. Sergeeva, H.L. Haas, Carbenoxolone impairs LTP and blocksNMDA receptors in murine hippocampus, Neuropharmacology 55 (2008)139–147.
19] Y. Guan, L.M. Johanek, T.V. Hartke, B. Shim, Y.X. Tao, M. Ringkamp, R.A. Meyer,S.N. Raja, Peripherally acting mu-opioid receptor agonist attenuates neuro-pathic pain in rats after L5 spinal nerve injury, Pain 138 (2008) 318–329.
20] S.R. Chaplan, F.W. Bach, J.W. Pogrel, J.M. Chung, T.L. Yaksh, Quantitative assess-ment of tactile allodynia in the rat paw, J. Neurosci. Methods 53 (1994) 55–63.
21] W.J. Dixon, Efficient analysis of experimental observations, Annu. Rev. Pharma-col. Toxicol. 20 (1980) 441–462.
22] X.T. Ji, N.S. Qian, T. Zhang, J.M. Li, X.K. Li, P. Wang, D.S. Zhao, G. Huang, L. Zhang,Z. Fei, D. Jia, L. Niu, Spinal astrocytic activation contributes to mechanical allo-dynia in a rat chemotherapy-induced neuropathic pain model, PLoS ONE 8(2013) e60733.
23] J. Scholz, C.J. Woolf, The neuropathic pain triad: neurons, immune cells and glia,Nat. Neurosci. 10 (2007) 1361–1368.
24] J.A. Orellana, P.J. Saez, K.F. Shoji, K.A. Schalper, N. Palacios-Prado, V. Velarde,C. Giaume, M.V. Bennett, J.C. Saez, Modulation of brain hemichannels and gapjunction channels by pro-inflammatory agents and their possible role in neu-rodegeneration, Antioxid. Redox Signal. 11 (2009) 369–399.
25] J.I. Nagy, F.E. Dudek, J.E. Rash, Update on connexins and gap junctions in neurons
and glia in the mammalian nervous system, Brain Res. Brain Res. Rev. 47 (2004)191–215.26] P.T. Ohara, J.P. Vit, A. Bhargava, L. Jasmin, Evidence for a role of connexin 43in trigeminal pain using RNA interference in vivo, J. Neurophysiol. 100 (2008)3064–3073.

e Lett
[
[
[
[
[
[
[
[
[
[
[
Q. Xu et al. / Neuroscienc
27] J.P. Vit, L. Jasmin, A. Bhargava, P.T. Ohara, Satellite glial cells in the trigeminalganglion as a determinant of orofacial neuropathic pain, Neuron Glia Biol. 2(2006) 247–257.
28] W. Guo, H. Wang, M. Watanabe, K. Shimizu, S. Zou, S.C. LaGraize, F. Wei, R. Dub-ner, K. Ren, Glial–cytokine–neuronal interactions underlying the mechanismsof persistent pain, J. Neurosci. 27 (2007) 6006–6018.
29] W. Meme, C.F. Calvo, N. Froger, P. Ezan, E. Amigou, A. Koulakoff, C.Giaume, Proinflammatory cytokines released from microglia inhibit gapjunctions in astrocytes: potentiation by beta-amyloid, FASEB J. 20 (2006)494–496.
30] R.R. Ji, Y. Kawasaki, Z.Y. Zhuang, Y.R. Wen, I. Decosterd, Possible role ofspinal astrocytes in maintaining chronic pain sensitization: review of cur-rent evidence with focus on bFGF/JNK pathway, Neuron Glia Biol. 2 (2006)259–269.
31] A. Okada-Ogawa, I. Suzuki, B.J. Sessle, C.Y. Chiang, M.W. Salter, J.O. Dostrovsky,Y. Tsuboi, M. Kondo, J. Kitagawa, A. Kobayashi, N. Noma, Y. Imamura, K. Iwata,Astroglia in medullary dorsal horn (trigeminal spinal subnucleus caudalis) areinvolved in trigeminal neuropathic pain mechanisms, J. Neurosci. 29 (2009)11161–11171.
[
ers 566 (2014) 194–199 199
32] M. Donegan, M. Kernisant, C. Cua, L. Jasmin, P.T. Ohara, Satellite glial cell pro-liferation in the trigeminal ganglia after chronic constriction injury of theinfraorbital nerve, Glia (2013).
33] D.M. Friend, K.A. Keefe, Glial reactivity in resistance to methamphetamine-induced neurotoxicity, J. Neurochem. 125 (2013) 566–574.
34] A. Williams, G. Piaton, C. Lubetzki, Astrocytes—friends or foes in multiple scle-rosis? Glia 55 (2007) 1300–1312.
35] B.A. Winkelstein, J.A. DeLeo, Nerve root injury severity differentially modulatesspinal glial activation in a rat lumbar radiculopathy model: considerations forpersistent pain, Brain Res. 956 (2002) 294–301.
36] R.W. Colburn, J.A. DeLeo, A.J. Rickman, M.P. Yeager, P. Kwon, W.F. Hickey, Dis-sociation of microglial activation and neuropathic pain behaviors followingperipheral nerve injury in the rat, J. Neuroimmunol. 79 (1997) 163–175.
37] V. Raghavendra, F. Tanga, J.A. DeLeo, Inhibition of microglial activation atten-
uates the development but not existing hypersensitivity in a rat model ofneuropathy, J. Pharmacol. Exp. Ther. 306 (2003) 624–630.38] H. Zhang, X. Mei, P. Zhang, C. Ma, F.A. White, D.F. Donnelly, R.H. Lamotte, Alteredfunctional properties of satellite glial cells in compressed spinal ganglia, Glia57 (2009) 1588–1599.
Copyright © 2022 FDOKUMEN