
Role of isostaticity and load-bearing microstructure in ... - PNAS
Upload
khangminh22Category
view
6download
0
Supplementary Information 1. Supplementary Methods Phylogenetic and age justifications for fossil calibrations The justifications for each fossil calibration are presented here for the ‘hornworts-sister’ topology (summarised in Table S2). For variations of fossil calibrations for the other hypothetical topologies, see Supplementary Tables S1-S7. Node 104: Viridiplantae; Chlorophyta – Streptophyta: 469 Ma – 1891 Ma. Fossil taxon and specimen: Tetrahedraletes cf. medinensis [palynological sample 7999: Paleopalynology Unit, IANIGLA, CCT CONICET, Mendoza, Argentina], from the Zanjón - Labrado Formations, Dapinigian Stage (Middle Ordovician), at Rio Capillas, Central Andean Basin, northwest Argentina [1]. Phylogenetic justification: Permanently fused tetrahedral tetrads and dyads found in palynomorph assemblages from the Middle Ordovician onwards are considered to be of embryophyte affinity [2-4], based on their similarities with permanent tetrads and dyads found in some extant bryophytes [5-7] and the separating tetrads within most extant cryptogams. Wellman [8] provides further justification for land plant affinities of cryptospores (sensu stricto Steemans [9]) based on: assemblages of permanent tetrads found in deposits that are interpreted as fully terrestrial in origin; similarities in the regular arrangement of spore bodies and size to extant land plant spores; possession of thick, resistant walls that are chemically similar to extant embryophyte spores [10]; some cryptospore taxa possess multilaminate walls similar to extant liverwort spores [11]; in situ cryptospores within Late Silurian to Early Devonian bryophytic-grade plants with some tracheophytic characters [12,13]. The oldest possible record of a permanent tetrahedral tetrad is a spore assigned to Tetrahedraletes cf. medinensis from an assemblage of cryptospores, chitinozoa and acritarchs collected from a locality in the Rio Capillas, part of the Sierra de Zapla of the Sierras Subandinas, Central Andean Basin, north-western Argentina [1]. Minimum age: 469 Ma. Soft maximum age: 1891 Ma. Age justification: The minimum age constraint is based on a palynological sample containing the oldest tetrahedral tetrad, collected from strata in the transition between the upper part of the Zanjón Formation and the Laja Morada Member of the Labrado Formation (level 7999, fig. 1, [1]). This sample was assigned to the Dapinigian Stage of the Middle Ordovician, based on the presence of the chitinozoan Lagenochitina combazi, a biostratigraphical marker of the L. combazi Biozone of southern China [14]. At the Dapingian type sections in the Yichang area of China, the local L. combazi Biozone begins within 1m above the GSSP of the base of Dapingian, as defined by the FAD of the conodont Baltoniodus triangularis [15, 16], currently dated at 470.0 Ma ± 1.4 Myr [17]. The top of the combazi Biozone has been correlated closely to the boundary between the chitinozoan D. ornensis and B. henryi Biozones of Northern Gondwana [14, 18]. Dated at 469 Ma [17], this boundary is the youngest possible age for the top of the correlated L. combazi Biozone, and thus the minimum age of the tetrahedral tetrad record from Argentina, 1 Myr after the beginning of the Dapingian Stage/ Middle Ordovician. The soft maximum constraint is based on the earliest record of eukaryotes [19-21] when, despite the presence of simple eukaryotes, there is no evidence of anything as complex as a multicellular alga. Though the fossils present have been suggested by some to represent some kind of green algae [22]. The maximum age for this formation is based on the igneous and metamorphic rocks that it overlies. These rocks are dated at 1823 Ma ± 68 Myr [23]. Discussion: Parfrey et al. [24] identify Palaeastrum and Proterocladus, from the early Neoproterozoic Svenbergfjellet flora of Spitsbergen [25] as charophytes. However, though the characteristics of these fossils are compatible with Chlorophyaceae, their simple morphology precludes unequivocal assignment to Viridiplantae [26]. Claims of chlorophytes in the Ediaracan Miahoe Biota [27] are even more speculative. Amgaella, Palaeoporella, Yakutina and Seletonella, among many other Cambrian and Ordovician records [28, 29], have variously been considered the oldest Dasycladales, or siphonous algae (Bryopsidales and Dasycladales) [28-30], however, even their algal affinity has been considered uncertain [28, 29]. Palaeocymopolia silurica is considerably younger, but its anatomy is well characterized. Node 205: Chlorophyceae – Prasinophyaceae: 438.3 Ma – 1891 Ma. Fossil taxon and specimen: Palaeocymopolia silurica [TUG 1269-9 (holotype): Natural History Museum of the University of Tartu, Estonia], from the Aeronian (Llandovery; Silurian) Raikküla Stage at a quarry near Kalana, Estonia [31].

Phylogenetic justification: Mastik and Tinn [31] draw comparison between Palaeocymopolia silurica and living dasycladealean Cympolia species based on the shared presence of ‘thalli with dichotomously branching, serially segmented form, segment size, and shape, and size and morphology of filaments in the tufts’ [p. 6: 31]. Minimum age: 438.3 Ma. Soft maximum age: 1891 Ma. Age justification: The Kalana Lagerstätte occurs within the Pribylograptus leptotheca graptolite Biozone [32]. The minimum age can therefore be determined by the base of the succeeding Lituigraptus convolutes graptolite Biozone, dated to 439.21 Ma ± 0.9031 Myr, thus, 438.3 Ma. The soft maximum constraint is based on the earliest record of eukaryotes [19-21] when, despite the presence of simple eukaryotes, there is no evidence of anything as complex as a multicellular alga. Though the fossils present have been suggested by some to represent some kind of green algae [22]. The maximum age for this formation is based on the igneous and metamorphic rocks that it overlies. These rocks are dated at 1823 Ma ± 68 Myr [23]. Discussion: This is the divergence between Uronema versus Nephroselmis, Monomastix, and Pyramimonas, the former a member of the monophyletic Chlorophyceae, and the latter members of the paraphyletic Prasinophyaceae. Hence, we base our node calibration on the oldest certain Chlorophyceae, for which we use the oldest certain record of Dasycladales. Node 105: Streptophyta; Charophyta – Embryophyta: 469 Ma – 1891 Ma. Fossil taxon and specimen: Tetrahedraletes cf. medinensis [palynological sample 7999: Paleopalynology Unit, IANIGLA, CCT CONICET, Mendoza, Argentina], from the Zanjón - Labrado Formations, Dapinigian Stage (Middle Ordovician), at Rio Capillas, Central Andean Basin, northwest Argentina [1]. Phylogenetic justification: Permanently fused tetrahedral tetrads and dyads found in palynomorph assemblages from the Middle Ordovician onwards are considered to be of embryophyte affinity [2-4], based on their similarities with permanent tetrads and dyads found in some extant bryophytes [5-7] and the separating tetrads within most extant cryptogams. Wellman [8] provides further justification for land plant affinities of cryptospores (sensu stricto Steemans [9]) based on: assemblages of permanent tetrads found in deposits that are interpreted as fully terrestrial in origin; similarities in the regular arrangement of spore bodies and size to extant land plant spores; possession of thick, resistant walls that are chemically similar to extant embryophyte spores [10]; some cryptospore taxa possess multilaminate walls similar to extant liverwort spores [11]; in situ cryptospores within Late Silurian to Early Devonian bryophytic-grade plants with some tracheophytic characters [12,13]. The oldest possible record of a permanent tetrahedral tetrad is a spore assigned to Tetrahedraletes cf. medinensis from an assemblage of cryptospores, chitinozoa and acritarchs collected from a locality in the Rio Capillas, part of the Sierra de Zapla of the Sierras Subandinas, Central Andean Basin, north-western Argentina [1]. Minimum age: 469 Ma. Soft maximum age: 1891 Ma. Age justification: The minimum age constraint is based on a palynological sample containing the oldest tetrahedral tetrad, collected from strata in the transition between the upper part of the Zanjón Formation and the Laja Morada Member of the Labrado Formation (level 7999, fig. 1, [1]). This sample was assigned to the Dapinigian Stage of the Middle Ordovician, based on the presence of the chitinozoan Lagenochitina combazi, a biostratigraphical marker of the L. combazi Biozone of southern China [14]. At the Dapingian type sections in the Yichang area of China, the local L. combazi Biozone begins within 1m above the GSSP of the base of Dapingian, as defined by the FAD of the conodont Baltoniodus triangularis [15, 16], currently dated at 470.0 Ma ± 1.4 Myr [17]. The top of the combazi Biozone has been correlated closely to the boundary between the chitinozoan D. ornensis and B. henryi Biozones of Northern Gondwana [14, 18]. Dated at 469 Ma [17], this boundary is the youngest possible age for the top of the correlated L. combazi Biozone, and thus the minimum age of the tetrahedral tetrad record from Argentina, 1 Myr after the beginning of the Dapingian Stage/ Middle Ordovician. The soft maximum constraint is based on the earliest record of eukaryotes [1-3] when, despite the presence of simple eukaryotes, there is no evidence of anything as complex as a multicellular alga. Though the fossils present have been suggested by some to represent some kind of green algae [4]. The maximum age for this formation is based on the igneous and metamorphic rocks that it overlies. These rocks are dated at 1823 Ma ± 68 Myr [5]. Node 110: Embryophyta; Anthocerotophyta – [Marchantiophyta + Bryophyta] + Tracheophyta; 469 Ma – 515.5 Ma. Fossil taxon and specimen: Tetrahedraletes cf. medinensis [palynological sample 7999: Paleopalynology Unit, IANIGLA, CCT CONICET, Mendoza, Argentina], from the Zanjón - Labrado Formations, Dapinigian Stage (Middle Ordovician), at Rio Capillas, Central Andean Basin, northwest Argentina [1]. Phylogenetic justification: Permanently fused tetrahedral tetrads and dyads found in palynomorph assemblages from the Middle Ordovician onwards are considered to be of embryophyte affinity [2-4], based on their similarities with

permanent tetrads and dyads found in some extant bryophytes [5-7] and the separating tetrads within most extant cryptogams. Wellman [8] provides further justification for land plant affinities of cryptospores (sensu stricto Steemans [9]) based on: assemblages of permanent tetrads found in deposits that are interpreted as fully terrestrial in origin; similarities in the regular arrangement of spore bodies and size to extant land plant spores; possession of thick, resistant walls that are chemically similar to extant embryophyte spores [10]; some cryptospore taxa possess multilaminate walls similar to extant liverwort spores [11]; in situ cryptospores within Late Silurian to Early Devonian bryophytic-grade plants with some tracheophytic characters [12,13]. The oldest possible record of a permanent tetrahedral tetrad is a spore assigned to Tetrahedraletes cf. medinensis from an assemblage of cryptospores, chitinozoa and acritarchs collected from a locality in the Rio Capillas, part of the Sierra de Zapla of the Sierras Subandinas, Central Andean Basin, north-western Argentina [1]. Minimum age: 469 Ma. Soft maximum age: 515.5 Ma. Age justification: The minimum age constraint is based on a palynological sample containing the oldest tetrahedral tetrad, collected from strata in the transition between the upper part of the Zanjón Formation and the Laja Morada Member of the Labrado Formation (level 7999, fig. 1, [1]). This sample was assigned to the Dapinigian Stage of the Middle Ordovician, based on the presence of the chitinozoan Lagenochitina combazi, a biostratigraphical marker of the L. combazi Biozone of southern China [14]. At the Dapingian type sections in the Yichang area of China, the local L. combazi Biozone begins within 1m above the GSSP of the base of Dapingian, as defined by the FAD of the conodont Baltoniodus triangularis [15, 16], currently dated at 470.0 Ma ± 1.4 Myr [17]. The top of the combazi Biozone has been correlated closely to the boundary between the chitinozoan D. ornensis and B. henryi Biozones of Northern Gondwana [14, 18]. Dated at 469 Ma [17], this boundary is the youngest possible age for the top of the correlated L. combazi Biozone, and thus the minimum age of the tetrahedral tetrad record from Argentina, 1 Myr after the beginning of the Dapingian Stage/ Middle Ordovician. A soft maximum age constraint for crown group Embryophyta must encompass all total group Embryophyta, including all cryptospores (sensu stricto Strother et al. [33]). The oldest deposits to yield probable non-marine palynomorphs include the Rogersville Shale and Pumpkin Valley Shale of the Conasauga Group of east Tennessee [34], and the Bright Angel Shale of the Tonto Group of Arizona [33, 35], correlated to the Glossopleura and Ehmaniella Laurentian trilobite Biozones, dated to Stage 5 of Series 3, the oldest part of the Mid Cambrian [36]. Cryptospores have recently been reported from the Rome Formation [36] that underlies the Conasauga Group and is correlated to the Olenellus trilobite Biozone of Stage 4 of Series 2, the latest part of the Early Cambrian. The base of the Olenellus Biozone is dated to approximately 515.5 Ma [37]. Discussion: This node represents the divergence of crown group Embryophyta into Anthocerotophyta (hornworts) and the remaining land plants; Marchantiophyta and Bryophyta (liverworts and mosses) and Tracheophyta. There are no unequivocal records of bryophyte fossils before the Devonian. The oldest possible fossils include Longfengshania, from the Mesoproterozoic Little Dal Group of Canada and the Qingbaikou Group of North China [38-41], and Parafunaria sinensis from the Early to Middle Cambrian Kaili Formation of Taijiang County, Guizhou Province China [42]. However, the interpretation of bryophyte affinity for both these fossils is questioned [41, 43-47]. The fossil record of hornworts is scant, the first accepted body fossils not appearing until the Cretaceous, e.g. Dendroceros victoriensis [48] and Notothylacites filiformis [49]. In the study of wall ultrastructure, Taylor et al. [50] recognised that the extinct spore genus Emphanisporites possess a zone of weakness at the spore equator (termed pseudosuture), which is a feature of modern hornwort spores [51]. Although this evidence is equivocal, reports of emphanoid spores go back to the Wenlock e.g. E. neglectus, E. protophanus [50], which provides the same minimum age as the first unequivocal record of a total group tracheophyte Cooksonia spp. from the late Wenlock (Silurian) of Ireland [52, 53] (see node 111 for further discussion). Cardona-Correa et al. [54] describe sphagnopsid peat moss-like fragments extracted from the Upper Ordovician Platteville Formation dolomitized carbonates. However, these fragments are pale in colour, compared with other non-oxidised organic material of Palaeozoic age that has darkened as a result of diagenesis. It cannot be ruled out that they are later contaminants, encased within the carbonate matrix after dissolution and recrystallisation. Reports of other Palaeozoic mosses, e.g. Lower Devonian Sporogonites [55], Carboniferous Muscites plumatus [56], M. polytrichaceus [57], M. amplexifolius [58] and Dwykea [59], are all equivocal. Hübers and Kerp [60] report fossil moss fragments identical to Sphagnum from the Visean (Middle Mississippian) of Germany. The oldest fossil to represent the total Marchantiophyta group is Riccardiothallus devonicus from the Early Devonian (Pragian) of the Yunnan Province, China [61] (see nodes 173 and 184 for further discussion). This is approximately 20 Ma after the first record of Cooksonia spp. The plant body fossil record is meagre in contrast to the significant palynomorph records of unequivocal embryophyte spores, which date back to over 40 Ma prior to first appearance of Cooksonia. Clarke et al. [62] proposed trilete spores as the minimum age providing fossils for the Embryophyta node, the oldest record known from Late Ordovician strata of Saudi Arabia (see node 112 for discussion on trilete spores). However, it is now accepted that permanently fused tetrahedral tetrads and dyads found in palynomorph assemblages from the Middle Ordovician onwards are of embryophyte affinity [2-4]. In contrast, the affinity of cryptospores from various Cambrian deposits [33, 36, 63, 64) is equivocal, as they lack regular geometrical arrangements of younger cryptospores, some occurring as ‘packets’ of

multiple spore-like bodies surrounded by synoecosporal walls [65]. They do possess some similarities to younger cryptospores, including multilaminate walls, and as such have been interpreted as the spores of aeroterrestrial charophytic algae [33, 66]. However, an embryophytic affinity cannot be completely ruled out, thus the first appearance of cryptospores is the basis of the soft maximum age constraint for the Embryophyta node. Node 111: [Marchantiophyta + Bryophyta] – Tracheophyta; 426.9 Ma – 515.5 Ma. Fossil taxon and specimen: Cooksonia cf. pertoni [TCD 22944-22956: Geology Museum of the Geology Department, Trinity College Dublin], from the Cloncannon Formation, Homerian Stage (Wenlock, Silurian), at the top of Borrisnoe Mountain, in the Devil’s Bit mountain range of County Tipperary, Ireland (18S 052782) [52, 53]. Phylogenetic justification: The genus Cooksonia possess synapomorphies of crown group tracheophytes, as evidenced by younger specimens of Cooksonia pertoni, namely water-conducting cells with differentially thickened secondary walls (tracheids) [67]. However, as discussed by Clarke et al. [62], other characters, such as the sterome [68], are considered to be a synapomorphy of total group Tracheophyta, but not crown group. Cooksonia can be placed in the total group Tracheophyta with confidence, with additional synapomorphies such as stomata [68] and sporophyte branching [52, 69]. Minimum age: 426.9 Ma. Soft maximum age: 515.5 Ma. Age justification: The earliest records of Cooksonia occur in the Cloncannon Formation in the Devil’s Bit mountain range of County Tipperary, Ireland. This formation has been dated to the Homerian Stage (Wenlock, Silurian) based on the presence of the graptolite Colonograptus (Monograptus) ludensis. The top of the ludensis graptolite Biozone is correlated to the top of the Homerian Stage, dated at 427.4 Ma ± 0.5 Myr [70]; thus the minimum age constraint for this node is 426.9 Ma. A soft maximum age constraint for this node must encompass all cryptospore records (sensu stricto Strother et al. [33]). The oldest deposits to yield probable non-marine palynomorphs include the Rogersville Shale and Pumpkin Valley Shale of the Conasauga Group of east Tennessee [34], and the Bright Angel Shale of the Tonto Group of Arizona [33, 35], correlated to the Glossopleura and Ehmaniella Laurentian trilobite Biozones, dated to Stage 5 of Series 3, the oldest part of the Mid Cambrian [36]. Cryptospores have recently been reported from the Rome Formation [36] that underlies the Conasauga Group and is correlated to the Olenellus trilobite Biozone of Stage 4 of Series 2, the latest part of the Early Cambrian. The base of the Olenellus Biozone is dated to approximately 515.5 Ma [37]. Discussion: This node represents the divergence between [Marchantiophyta (liverworts) and Bryophyta (mosses)] and Tracheophyta (vascular plants). The fossil record of mosses is poor. Cardona-Correa et al. [54] describe sphagnopsid peat moss-like fragments extracted from the Upper Ordovician Platteville Formation dolomitized carbonates. However, these fragments are pale in colour, compared with other non-oxidised organic material of Palaeozoic age that has darkened as a result of diagenesis. It cannot be ruled out that they are later contaminants, encased within the carbonate matrix after dissolution and recrystallisation. Reports of other Palaeozoic mosses, e.g. Lower Devonian Sporogonites [55], Carboniferous Muscites plumatus [56], M. polytrichaceus [57], M. amplexifolius [58] and Dwykea [59], are all equivocal. Hübers and Kerp [60] report fossil moss fragments identical to Sphagnum from the Visean (Middle Mississippian) of Germany. A few additional unequivocal specimens are known from the Permian e.g. Polyssaievia spinulifolia [71] and Palaeocampylopus buragoase [72] (see node 175 for further discussion). The oldest fossil to represent the total Marchantiophyta group is Riccardiothallus devonicus from the Early Devonian (Pragian) of the Yunnan Province, China [61] (see nodes 173 and 184 for further discussion). However, in all cases these fossils are younger than the first appearance of Cooksonia. Trilete spores, monads that develop from the natural dissociation of a tetrahedral tetrad, have long been considered a synapomorphy of tracheophytes. However, Gray [2] estimates that 2.5% of extant bryophyte genera also produce monads with trilete marks. In some cases, particularly within the mosses, these are non-functional and inaperturate (e.g. cryptotrilete spores of Andreaea), that developed as a result of physical constraints on adjacent enlarging spores within the confines of a common sporocyte wall [73]. However, true triradiate sutures are observed in the spores of Sphagnum [74] and Takakia [75], hence trilete spores are not used here as a minimum age calibration for Tracheophyta. Node 173: Marchantiophyta – Bryophyta; 405.0 Ma – 515.5 Ma. Fossil taxon and specimen: Riccardiothallus devonicus [CBYn9004008 (holotype): Museum of Plant History, Institute of Botany, Chinese Academy of Sciences], from the Posongchong Formation, Pragian Stage (Early Devonian), near Zhichang Village, Gumu Town, Wenshan District, Yunnan Province, SW China (23°17ʹN, 104°16ʹE) [61]. Phylogenetic justification: The oldest liverwort fossil is Riccardiothallus devonicus, from the Gondwanian Posongchong Formation, Yunnan Province, southwest China [61]. It is described as a simple, irregularly bifurcating, ecostate thallus comprising several layers of cells with entire margins. No costa, air chambers, rhizoids or stomata were observed. The

specimen closely resembles the extant genus Riccardia, within the Aneuraceae family of the order Metzgeriales, Jungermanniopsida [76, 77], thus is considered to be the earliest representative of the Jungermanniopsida group. Minimum age: 405.0 Ma. Soft maximum age: 515.5 Ma. Age justification: Riccardiothallus devonicus was collected from the second unit of the Posongchong Formation, at a locality near Zhichang Village, Wenshan District, Yunnan Province, Southwest China [61]. The Posongchong Formation is considered by many to be middle to late Pragian in age, including Hao et al. [78], who provide the following justification. The Posongchong Formation underlies the marine Pojiao Formation at the Zhichang section. In the Guangnan and Funing Counties it has yielded the conodont Polygnathus dehiscens [79], a zonal conodont species of the early Emsian [80]. In the overlying Bajiaoquing Formation, the presence of Polygnathus dehiscens abyssus further demonstrates that the underlying Pojiao Formation is at least lowermost Emsian in age [81]. This age is further supported by the presence of the brachiopod Euryspirifer-Dicoelostrophia fauna and corals Xystriphylloides nobilis and Siphonophrentis angusta, which have been correlated well with the lower part of the Yujiang Formation at the Liujing section section in Guangxi [82]. The Yujiang Formation is of early Emsian age, based on the presence of the conodont Polygnathus dehiscens and tentaculite Nowakia praecursor. It overlies the Nagaoling Formation that has yielded conodonts Spathognathodus linearis postclinatus and S. salcatus, correlated to the middle-late Pragian [80, 83, 84]. Thus, the correlated Pojiao Formation is early Emsian in age, and its underlying Posongchong Formation is late Pragian in age. Although Hao et al. [78] consider Chinese spore zonation schemes as deficient, a spore assemblage described by Wang [85], placed the Posongchong Formation in the polygonalis – emsiensis (PE) spore assemblage Biozone of Richardson and McGregor [86], which supports a late Pragian age. The top of the Pragian is dated at 407.6 Ma ± 2.6 Myr [87], thus the minimum age constraint for the Marchantiophyta crown group is 405.0 Ma. A soft maximum age constraint for the Marchantiophyta crown group must encompass all cryptospore records (sensu stricto Strother et al. [33]). The oldest deposits to yield probable non-marine palynomorphs include the Rogersville Shale and Pumpkin Valley Shale of the Conasauga Group of east Tennessee [34], and the Bright Angel Shale of the Tonto Group of Arizona [33, 35], correlated to the Glossopleura and Ehmaniella Laurentian trilobite Biozones, dated to Stage 5 of Series 3, the oldest part of the Mid Cambrian [36]. Cryptospores have recently been reported from the Rome Formation [36] that underlies the Conasauga Group and is correlated to the Olenellus trilobite Biozone of Stage 4 of Series 2, the latest part of the Early Cambrian. The base of the Olenellus Biozone is dated to approximately 515.5 Ma [37]. Discussion: This node represents the divergence between Marchantiophyta and Bryophyta. The fossil record of mosses is poor. Cardona-Correa et al. [54] describe sphagnopsid peat moss-like fragments extracted from the Upper Ordovician Platteville Formation dolomitized carbonates. However, these fragments are pale in colour, compared with other non-oxidised organic material of Palaeozoic age that has darkened as a result of diagenesis. It cannot be ruled out that they are later contaminants, encased within the carbonate matrix after dissolution and recrystallisation. Reports of other Palaeozoic mosses, e.g. Lower Devonian Sporogonites [55], Carboniferous Muscites plumatus [56], M. polytrichaceus [57], M. amplexifolius [58] and Dwykea [59], are all equivocal. Hübers and Kerp [60] report fossil moss fragments identical to Sphagnum from the Visean (Middle Mississippian) of Germany. A few additional unequivocal specimens are known from the Permian e.g. Polyssaievia spinulifolia [71] and Palaeocampylopus buragoase [72] (see node 175 for further discussion). The oldest fossil to represent the total Marchantiophyta group is Riccardiothallus devonicus from the Early Devonian (Pragian) of the Yunnan Province, China [61] (see node 184 for further discussion). Node 184: Marchantiophyta; Jungermanniopsida – Marchantiopsida; 405.0 Ma – 515.5 Ma. Fossil taxon and specimen: Riccardiothallus devonicus [CBYn9004008 (holotype): Museum of Plant History, Institute of Botany, Chinese Academy of Sciences], from the Posongchong Formation, Pragian Stage (Early Devonian), near Zhichang Village, Gumu Town, Wenshan District, Yunnan Province, SW China (23°17ʹN, 104°16ʹE) [61]. Phylogenetic justification: The oldest liverwort fossil is Riccardiothallus devonicus, from the Gondwanian Posongchong Formation, Yunnan Province, southwest China [61]. It is described as a simple, irregularly bifurcating, ecostate thallus comprising several layers of cells with entire margins. No costa, air chambers, rhizoids or stomata were observed. The specimen closely resembles the extant genus Riccardia, within the Aneuraceae family of the order Metzgeriales, Jungermanniopsida [76, 77], thus is considered to be the earliest representative of the Jungermanniopsida group. Minimum age: 405.0 Ma. Soft maximum age: 515.5 Ma. Age justification: Riccardiothallus devonicus was collected from the second unit of the Posongchong Formation, at a locality near Zhichang Village, Wenshan District, Yunnan Province, Southwest China [61]. The Posongchong Formation is considered by many to be middle to late Pragian in age, including Hao et al. [78], who provide the following justification. The Posongchong Formation underlies the marine Pojiao Formation at the Zhichang section. In the Guangnan and Funing Counties it has yielded the conodont Polygnathus dehiscens [79], a zonal conodont species of the early Emsian [80]. In the overlying Bajiaoquing Formation, the presence of Polygnathus dehiscens abyssus further

demonstrates that the underlying Pojiao Formation is at least lowermost Emsian in age [81]. This age is further supported by the presence of the brachiopod Euryspirifer-Dicoelostrophia fauna and corals Xystriphylloides nobilis and Siphonophrentis angusta, which have been correlated well with the lower part of the Yujiang Formation at the Liujing section section in Guangxi [82]. The Yujiang Formation is of early Emsian age, based on the presence of the conodont Polygnathus dehiscens and tentaculite Nowakia praecursor. It overlies the Nagaoling Formation that has yielded conodonts Spathognathodus linearis postclinatus and S. salcatus, correlated to the middle-late Pragian [80, 83, 84]. Thus, the correlated Pojiao Formation is early Emsian in age, and its underlying Posongchong Formation is late Pragian in age. Although Hao et al. [78] consider Chinese spore zonation schemes as deficient, a spore assemblage described by Wang [85], placed the Posongchong Formation in the polygonalis – emsiensis (PE) spore assemblage Biozone of Richardson and McGregor [86], which supports a late Pragian age. The top of the Pragian is dated at 407.6 Ma ± 2.6 Myr [87], thus the minimum age constraint for the Marchantiophyta crown group is 405.0 Ma. A soft maximum age constraint for the Marchantiophyta crown group must encompass all cryptospore records (sensu stricto Strother et al. [33]). The oldest deposits to yield probable non-marine palynomorphs include the Rogersville Shale and Pumpkin Valley Shale of the Conasauga Group of east Tennessee [34], and the Bright Angel Shale of the Tonto Group of Arizona [33, 35], correlated to the Glossopleura and Ehmaniella Laurentian trilobite Biozones, dated to Stage 5 of Series 3, the oldest part of the Mid Cambrian [36]. Cryptospores have recently been reported from the Rome Formation [36] that underlies the Conasauga Group and is correlated to the Olenellus trilobite Biozone of Stage 4 of Series 2, the latest part of the Early Cambrian. The base of the Olenellus Biozone is dated to approximately 515.5 Ma [37]. Discussion: This node represents the divergence of Marchantiophyta into two of the three known classes within the phylum, Marchantiopsida and Jungermanniopsida, after Crandall-Stotler et al. [88]. There are no body fossils of liverworts prior to the Devonian, with only a handful of unequivocal specimens between the Early Devonian and Carboniferous, the majority from Laurentian deposits (Metzgeriothallus metzgerioides [89], Metzgeriothallus sharonae [90], Pallaviciniites devonicus [91] and Treubiites kidstonii [91]). The oldest liverwort fossil is Riccardiothallus devonicus, from the Gondwanian Posongchong Formation, Yunnan Province, southwest China [61]. As discussed under node 110, some unequivocal land plant spores as far back as the Ordovician possess multi-laminate walls, similar to extant spores of liverworts [92], specifically spores produced by the Sphaerocarpales of Marchantiopsida, but this remains too equivocal to use as a calibration. The oldest possible fossil record of crown Marchantiopsida is Treubiites (Hepaticites) kidstonii, from the Lower Carboniferous of Scotland [91]. Walton [93] initially described this fossil and named the genus after the extant taxon Treubia, a member of Haplomitriopsida, while Schuster [91] re-assigned it to Blasiales of Marchantiopsida. Anyhow, regardless of the uncertainties surrounding the affinity of this fossil, this record is younger than the first occurrence of Jungermanniopsida and hence Riccardiothallus devonicus is the minimum age providing fossil for the Marchantiophyta crown group. Node 185: Marchantiopsida: Sphaerocarpales – Marchantiales; 227 Ma – 515.5 Ma. Fossil taxon and specimen: Marchantites cyathodoides [13929: South African Museum, Cape Town], from the Molteno Formation, Carnian Stage (Late Triassic), at the Upper Umkomaas locality, Karoo Basin, South Africa [99, 100]. Phylogenetic justification: The oldest unequivocal fossil belonging to Marchantiales is Marchantites cyathodoides [99, 100] from the Upper Triassic Molteno Formation of South Africa, as discussed in Heinrichs et al. [101] and Villarreal et al. [102]. Anderson’s assigned of this species to Marchantiaceae was based on “the prostrate, forked thallus, a conspicuous midrib, rhizoides [sic], possible air chambers and central scales” [p. 22, 100]. Minimum age: 227 Ma. Soft maximum age: 515.5 Ma. Age justification: Marchantites cyathodoides was collected from the Molteno Formation at the Upper Umkomaas locality, Karoo Basin, South Africa [100]. The exact position of this locality within the Molteno Formation is unknown, so the youngest minimum age for this fossil must be taken at the top of the formation, which is considered to be Carnian (Upper Triassic) in age [103]. The top of the Carnian is currently dated at 227 Ma (International Chronostratigraphic Chart, v.2016/10). Therefore, the minimum age constraint for this node is 227 Ma. A soft maximum age constraint for this node must encompass all cryptospore records (sensu stricto Strother et al. [33]). The oldest deposits to yield probable non-marine palynomorphs include the Rogersville Shale and Pumpkin Valley Shale of the Conasauga Group of east Tennessee [34], and the Bright Angel Shale of the Tonto Group of Arizona [33, 35], correlated to the Glossopleura and Ehmaniella Laurentian trilobite Biozones, dated to Stage 5 of Series 3, the oldest part of the Mid Cambrian [36]. Cryptospores have recently been reported from the Rome Formation [36] that underlies the Conasauga Group and is correlated to the Olenellus trilobite Biozone of Stage 4 of Series 2, the latest part of the Early Cambrian. The base of the Olenellus Biozone is dated to approximately 515.5 Ma [37]. Discussion: This node represents the divergence of Marchantiidae, one of two subclasses within Marchantiopsida, into two of the four known orders, Sphaerocarpales and Marchantiales, after Crandall-Stotler et al. [88]. The oldest possible fossil of Marchantiopsida is Treubiites (Hepaticites) kidstonii, from the Lower Carboniferous of Scotland [91]. However, as discussed in node 184, there are uncertainties over its affinity, possibly belong to Blasiales, an order not represented

on this tree. Naiadita lanceolata, is a liverwort fossil from the Late Triassic of England [104], closely allied to the extant liverwort Riella of the Sphaerocarpales [105-107]. The similarity with Sphaerocarpales is based on the sporophyte of Naiadita, which is short-stalked, with a unistratose capsule wall, bearing large permanent tetrad spores and no elaters [91]. However, this fossil differs from Sphaerocarpales, as discussed by Schuster [91] and recently in a diagnosis of Naiadita by Katagiri and Hagborg [107], in having an unusual combination of characters, such as spirally-arranged leaves, terminal gemmae cups, and stalked archegonia arising singly on the stem and other vegetative features that are more similar to the Calobryales. It has therefore been assigned to a monogeneric order Naiaditales within the family Naiaditaceae, placed in the Marchantiidae [107], thus not a suitable fossil for calibration of this tree. The extinct spore taxon Aequitriradites [108] is morphologically similar to extant spores of liverworts belong to the Sphaerocarpales order [109]. The oldest records of Aequitriradites are Anisian in age [110, 111]; the top of the Anisian Stage is currently dated at 242 Ma (International Chronostratigraphic Chart, v.2016/10). However, while this record is older than Marchantites cyathodoides, the affinity of Aequitriradites to Sphaerocarpales is too equivocal to use this taxon to calibrate this node. Node 174: Bryophyta: Sphagnopsida – [Polytrichiopsida + Bryopsida]; 330.7 Ma – 515.5 Ma. Fossil taxon and specimen: Sphagnales [KS27-074, KS27-031, KSgrPr-068, KS11-090, KS11-029, KS27-048: Forschungsstelle für Paläobotanik, University of Münster, Germany], from the upper part of the Ortelsdorf Formation, Hainchen Basin, Visean Stage (middle Mississippian, Carboniferous), from a roadcut of the A4 motorway near Chemnitz-Glösa, Saxony, Germany (50°52ʹ34.67ʹʹN; 12°54ʹ38.73ʹʹE) [59]. Phylogenetic justification: Although Hübers and Kerp [59] do not assign these fossils to a specific genus due to their fragmentary nature, they unequivocally assigned them to Sphagnales, based on: unistratose fragments; absence of vascular tissue; absence of stomata; small pores in the periclinal walls; dimorphic leaf cells and cell network patterns identical to those of Protosphagnales from the Permian of Russia [112, 113]; elongated cells with fibrils identical to those of leucocysts, the water-storage cells of the extant taxon Sphagnum. Although the fragments represent eight different forms, each possessing some, but not all, of these synapomorphies, the fragments could represent different parts of the same species. Minimum age: 330.7 Ma. Soft maximum age: 515.5 Ma. Age justification: As described in Hübers and Kerp [59], these fossil mosses were collected from the upper part of the Ortelsdorf Formation (Hainichen Subgroup, Hainichen Basin). At Glösa, spore assemblages from the Ortelsdorf Formation belong to the NM and VF Biozones [114] of Clayton et al. [115]. The moss-bearing samples are from the uppermost part of the formation that yielded assemblages belonging to the VF Biozone of late Visean age. This is consistent with a single-zircon Pb/Pb age of 330 Ma ± 4 Myr from a rhyolitic tuff at the base from the upper part of the formation [116, 117]. The top of the Visean is dated to 330.9 Ma ± 0.2 Myr (International Chronostratigraphic Chart v.2016/10), thus the minimum age is 330.7 Ma. A soft maximum age constraint for this node must encompass all cryptospore records (sensu stricto Strother et al. [33]). The oldest deposits to yield probable non-marine palynomorphs include the Rogersville Shale and Pumpkin Valley Shale of the Conasauga Group of east Tennessee [34], and the Bright Angel Shale of the Tonto Group of Arizona [33, 35], correlated to the Glossopleura and Ehmaniella Laurentian trilobite Biozones, dated to Stage 5 of Series 3, the oldest part of the Mid Cambrian [36]. Cryptospores have recently been reported from the Rome Formation [36] that underlies the Conasauga Group and is correlated to the Olenellus trilobite Biozone of Stage 4 of Series 2, the latest part of the Early Cambrian. The base of the Olenellus Biozone is dated to approximately 515.5 Ma [37]. Discussion: This node represents the divergence of Bryophyta into 2 of the 5 known Superclasses, II (Sphagnopsida) and V (including Polytrichiopsida and Bryopsida), after Goffinet et al. [118]. The oldest fossil that is assigned unequivocally to Polytrichiopsida is the Lower Permian (Cisuralian) Palaeocampylopus buragoase, of the Pospelovo Formation, Russky Island, Russia [72] (see node 175 for further discussion). The oldest possible fossil that is assigned to Bryopsida is the Middle Permian (Guadalupian) Capimirinus riopretensis, of the Teresina Formation, Paraná, Brazil [119] (see node 176 for further discussion). These are both younger than the Carboniferous mosses of Hübers and Kerp [59]. Node 175: Polytrichiopsida – Bryopsida; 271.8 Ma – 515.5 Ma Fossil taxon and specimen: Palaeocampylopus buragoase [PIN no. 5328/1-B (holotype): Paleontological Institute, Russian Academy of Sciences, Moscow, Russia], from the lower subformation of the Pospelovo Formation, middle Kungurian Stage (late Cisuralian, Permian) at a coastal cliff locality 0.3 km SSW of the Cape Novosilsky, Russky Island, near Vladivostok, Russia (ca. 43°00’N; 131°50’E) [72]. Phylogenetic justification: Following Laenen et al. [120], Palaeocampylopus buragoase [72] is the oldest unequivocal fossil belonging to Polytrichiopsida. This is based on gametophytic remains of foliate shoots, some bearing cup-like structures. Leaf shape is lanceolate proximate portions and subulate acumens, and they are arranged in a dense spiral.

Cup-like structures are interpreted as perigonial leaves around the gametangia [72] and these are similar to those in extant mosses belonging to Polytrichaceae. Minimum age: 271.8 Ma. Soft maximum age: 515.5 Ma. Age justification: Based on Siberian floral assemblages, the lower sub-formation of the Pospelovo Formation is assigned to the local Abrek Horizon, which is correlated to the upper part of the Kungurian Stage (Cisuralian, Permian) [121, 122]. The top of the Kungurian is dated at 272.3 Ma ± 0.5 Myr [123], thus the minimum age is 271.8 Ma. A soft maximum age constraint for this must encompass all cryptospore records (sensu stricto Strother et al. [33]). The oldest deposits to yield probable non-marine palynomorphs include the Rogersville Shale and Pumpkin Valley Shale of the Conasauga Group of east Tennessee [34], and the Bright Angel Shale of the Tonto Group of Arizona [33, 35], correlated to the Glossopleura and Ehmaniella Laurentian trilobite Biozones, dated to Stage 5 of Series 3, the oldest part of the Mid Cambrian [36]. Cryptospores have recently been reported from the Rome Formation [36] that underlies the Conasauga Group and is correlated to the Olenellus trilobite Biozone of Stage 4 of Series 2, the latest part of the Early Cambrian. The base of the Olenellus Biozone is dated to approximately 515.5 Ma [37]. Discussion: This node represents the divergence of the Polytrichiopsida and Bryopsida total groups, after Goffinet et al. [118]. The next possible oldest fossil assigned to Polytrichiopsida is the Campanian (Late Cretaceous) Eopolytrichum antiquum, from the Buffalo Creek Member of the Gaillard Formation, Allon Quarry, Crawford County, Georgia [124]. The oldest possible fossil that is assigned to Bryopsida is the Middle Permian (Guadalupian) Capimirinus riopretensis, of the Teresina Formation, Paraná, Brazil [119] (see node 176 for further discussion). Node 176: Bryopsida: Funariidae – [Dicraniidae + Bryidae]; 268.3 Ma – 515.5 Ma. Fossil taxon and specimen: Capimirinus riopretensis [CP1/193 (holotype): Scientific Paleontologic Collection – Paleobotany (CP1) of the Institute of Geosciences of the University of Campinas (UNICAMP)], from the Teresina Formation, Paraná Basin, Guadalupian (Middle Permian), at Rio Preto Quarry, by the Irati – Guaramirim road section, state of Paraná, Brazil (25°31’31.3”S; 50°44’54.0”W) [119]. Phylogenetic justification: Following Laenen et al. [119], Capimirinus riopretensis is the oldest unequivocal fossil belonging to Hypnaceae of the order Hypnales, Bryidae [119]. This is based on the similarities between pleurocarpous C. riopretensis and the extant genus Hypnum [125]; gametophytic dichotomous stems of variable thickness that decrease in thickness towards the apex; erect-patent leaves arranged in a spiral; leaf shape ovate and slightly asymmetrical; leaf bases decurrent, tips pointed with entire margin; well defined, straight costa; sporophyte with elongate seta, with inclined ovate capsule and perchaerial leaves surround the seta. However, Christiano de Souza et al. [119] were reluctant to place the fossil specimens in Hypnum directly, as the leaves of Hypnum always possess double short costae and have acuminate tips, while those of C. riopretensis possess a single costa and are only slightly pointed at the tips. Christiano de Souza et al. [119] also note differences in sporophyte shape. Based on these differences, C. riopretensis was proposed. Minimum age: 268.3 Ma. Soft maximum age: 515.5 Ma. Age justification: The Teresina Formation is part of the Passa Dois Group. The base of the lowermost formation in the group, the Irati Formation, is dated to 278.4 Ma ± 2.2 Myr, based on zircon U/Pb isotopes taken from bentonitic ash layers [126]. The younger Teresina Formation is correlated to the Roadian Stage, Guadalupian (Middle Permian) by Santos et al. [126]. The top of the Roadian Stage is currently dated at 268.8 Ma ± 0.5 Myr [123], thus the minimum age constraint for this node is 268.3 Ma. A soft maximum age constraint for this node must encompass all cryptospore records (sensu stricto Strother et al. [33]). The oldest deposits to yield probable non-marine palynomorphs include the Rogersville Shale and Pumpkin Valley Shale of the Conasauga Group of east Tennessee [34], and the Bright Angel Shale of the Tonto Group of Arizona [33, 35], correlated to the Glossopleura and Ehmaniella Laurentian trilobite Biozones, dated to Stage 5 of Series 3, the oldest part of the Mid Cambrian [36]. Cryptospores have recently been reported from the Rome Formation [36] that underlies the Conasauga Group and is correlated to the Olenellus trilobite Biozone of Stage 4 of Series 2, the latest part of the Early Cambrian. The base of the Olenellus Biozone is dated to approximately 515.5 Ma [37]. Discussion: This node represents the divergence of Bryopsida into three of the six known subclasses, Funariidae, plus Dicranidae and Bryidae, after Goffinet et al. [118]. Although many extant families of these subclasses are represented in the Cenozoic record, such as those preserved within the Eocene Baltic amber [127, 128] and Miocene amber from the Dominican Republic [129], the pre-Cenozoic record of these groups is poor. The next oldest members of the Hypnanae superorder of Bryidae are known from the Early Cretaceous Apple Bay Flora [130, 131]. A possible oldest fossil of Bryanae superorder of Bryidae is Intia vermicularis, described by Neuburg [71] from the Permian Angaraland. Of Dicraniidae, Campylopodium allonense from the Buffalo Creek Member of the Gaillard Formation, Georgia, USA, is the oldest representative, of Late Santonian (Late Cretaceous) age [132]. There is no fossil record of the Funariidae. Node 112: Tracheophyta: Lycopodiophyta – Euphyllophyta; 420.7 Ma – 451 Ma.

Fossil taxon and specimen: Zosterophyllum sp. [US384-8137: University of Saskatchewan Collections, Canada], from beds equivalent to the Bathurst Island and Stuart Bay Formations, Ludlow (Silurian), exposed 14 km north of Polar Bear Pass research station, eastern Bathurst Island, Arctic Canada (US384; ca. 75°50’N, 98°35’W) [134]. Phylogenetic justification: Following Clarke et al. [62], Zosterophyllum is considered to be a member of the Lycopodiophyta total group, based on the lateral insertion of reniform sporangia along stems, which dehisce along their distal margins into two valves [69]. Minimum age: 420.7 Ma. Soft maximum age: 451 Ma. Age justification: Zosterophyllum sp. on Bathurst Island co-occurs with the conodont Ozarkodina douroensis [134], which is restricted to the mid-late Ludlow (as O. n. sp. B, [135-138]). Therefore, the minimum age of these beds is the top of the Ludlow Epoch, dated at 423.0 Ma ± 2.3 Myr [70], thus the minimum age constraint for this node is 420.7 Ma. Following Clarke et al. [62], the soft maximum age constraint is based on the first appearance of trilete spores, in the Qusaiba-1 core from the Qasim Formation, northern Saudi Arabia [139]. This formation is Katian (Late Ordovician) in age based on the presence of Acanthochitina barbata, of the barbata Biozone. The oldest trilete spores precede the earliest occurrences of A. barbata, thus it is a possibility that they occur in the preceding Tanuchitina fistulosa Biozone, though T. fistulosa does not occur. The oldest trilete spores within the core co-occur with Armoricochitina nigerica, known to extend into the British Caradoc stage, within the Fungochitina spinifera Biozone (= F. fungiformis) [140]. The base of the F. spinifera Biozone falls within the Dicranograptus clingani Biozone (Dicellograptus morrisi sub-biozone) [141], the base of which is estimated to be 451 Ma [17]. Discussion: This node represents the divergence of Tracheophyta into Lycopodiophyta and Euphyllophyta. The oldest possible records of total group Lycopodiophyta are Cooksonia spp. from the Cloncannon Formation, County Tipperary, Ireland of Homerian age (Wenlock, Silurian) [52, 53]. This is based on the characters of younger specimens, including naked stems with terminal sporangia and water-conducting cells with differentially thickened secondary walls (tracheids) [67]. However, as discussed by Clarke et al. [62], while Cooksonia can be placed in the total group Tracheophyta with confidence, with additional synapomorphies such as stomata [68] and sporophyte branching [52, 69], other characters, such as the sterome [68], are not considered to be a synapomorphies of the crown group Tracheophyta. In addition, until recently only one species of Cooksonia bore terminal bivalved, reniform sporangia (C. caledonica Edwards [142]) [143], which has since been re-assigned to Aberlemnia caledonica [144]. Therefore, Cooksonia is not appropriate for the calibration of Lycopodiophyta. The next oldest possible record of total group Lycopodiophyta is Baragwanathia longifolia, a lycopsid from the ‘Lower Plant Assemblage’ in the Yea District, central Victoria, Australia [145]. The age of this horizon is also dated to the Ludlow (Silurian) [146, 147] (although it has been dated as Pragian in the past [148]). Wutubulaka multidichotoma from the Wutubalake Formation, late Pridoli of Xinjiang, China [149] is tentatively placed within total group Euphyllophyta, based only on one synapomorphy, pseudomonopodial branching [69]. Therefore, this fossil is not a suitable for calibration for total group Euphyllophyta or crown group Tracheophyta. Eophyllophyton bellum, Psilophyton primitivum and Pauthecophyton gracile from the Pragian Posongchong Formation of Yunnan, China [150-152] are considered to be the oldest unequivocal stem group Euphyllophyta based on characters such as pseudomonopodial branching, sporangial pairs grouped into terminal trusses and dichotomous appendages. However, they are significantly younger than the Ludlow Zosterophyllum sp. from Bathurst Island. Node 169: Lycopodiophyta: Isoetopsida – Lycopodiopsida; 392.1 Ma – 451 Ma. Fossil taxon and specimen: Leclercqia complexa [NBMG 13025 – 13034, 13069 – 13071: New Brunswick Museum, St. John’s, New Brunswick, Canada] from Campbellton Formation, Emsian (Early Devonian), ‘Locality F’ on the south shore of the Restigouche River, between Dalhousie and Campbellton, New Brunswick, eastern Canada [153, 154]. Phylogenetic justification: Leclercqia complexa is interpreted as a protolepidodendralean lycopsid and the oldest member of Isoetopsida and crown group Lycopodiophyta [95]. This is based on isotomously or pseudomonopodially branched stems, with helically-arranged forked leaves and sporophylls with isovalvate elongate-oval sporangia [95, 153]. In a study of wall ultrastructure of the in situ spores, Acinosporites lindlarensis, multilamellate regions in the interradial areas of the spore proximal surface, known to occur in the spores of other extinct and extant ligulate lycopsids, thus a further synapomorphy for the ligulate lycopsid clade [154]. Minimum age: 392.1 Ma. Soft maximum age: 451 Ma. Age justification: The specimens of Leclerqia complexa were collected from ‘Locality F’ on the south shore of the Restigouche River, between Dalhousie and Campbellton, New Brunswick, eastern Canada [153, 154]. Dispersed spore assemblages belong to the middle part of the Emphanisporites annulatus – Camarozonotriletes sextantii Spore Assemblage Biozone of Richardson and McGregor [86], indicating a mid Emsian age [153, 154]. The top of the Emsian Stage is dated at 393.3 Ma ± 1.2 Myr [87], thus the minimum age constraint for the Lycopodiophyta node is 392.1 Ma. Following Clarke et al. [62], the soft maximum age constraint is based on the first appearance of trilete spores, in the Qusaiba-1 core from the Qasim Formation, northern Saudi Arabia [139]. This formation is Katian (Late Ordovician) in age based on the presence of Acanthochitina barbata, of the barbata Biozone. The oldest trilete spores precede the

earliest occurrences of A. barbata, thus it is a possibility that they occur in the preceding Tanuchitina fistulosa Biozone, though T. fistulosa does not occur. The oldest trilete spores within the core co-occur with Armoricochitina nigerica, known to extend into the British Caradoc stage, within the Fungochitina spinifera Biozone (= F. fungiformis) [140]. The base of the F. spinifera Biozone falls within the Dicranograptus clingani Biozone (Dicellograptus morrisi sub-biozone) [141], the base of which is estimated to be 451 Ma [17]. Discussion: This node represents the divergence of Lycopodiophyta into Isoetopsida and Lycopodiopsida. Total group Lycopodiopsida include: Baragwanathia longifolia, possible of Ludlow (Silurian) age, from the Yea District, central Victoria, Australia [145]; Drepanophycus spinaeformis, a cosmopolitan taxon known from the Lochkovian to Frasnian (Devonian) [155-157]; and Asteroxylon mackiei, Rhynie Chert, Scotland [158], of Pragian (Early Devonian) age [159]. However, these taxa do not possess all synapomorphies of the crown group Lycopodiopsida [69], and are assigned to Drepanophycales, forming a sister group to all other lycopsids [69], and thus are not suitable for the calibration of this node. Another cited possible oldest fossil of Lycopodiopsida is the upper Devonian Lycopodites [160, 161], but there are no clear synapomorphies to support this [69]. Of Isoetopsida, one possible older fossil than Leclercqia complexa is Zhenglia radiata, from the Pragian Posongchong Formation of southeastern Yunnan, China [162]. However, as evidence for this is limited, this systematic placement is tentative. Node 113: Euphyllophyta: Monilophyta – Spermatophyta; 385.571 Ma – 451 Ma. Fossil taxon and specimen: Rellimia thomsonii [335.34: Paleobotanical Collection of the State University of New York at Binghamton], from the Panther Mountain Formation, Givetian Stage (Middle Devonian), from Blenheim-Gilboa Power Plant, Schoharie County, New York State (42° 4’N, 74°4’W) [163]. Phylogenetic justification: After Clarke et al. [62], Rellimia thomsonii is considered to be the oldest unequivocal aneurophytalean progymnosperm fossil [163], thus the oldest record of crown Euphyllophyta. The axes Rellimia thomsonii are helically-arranged branches up to four orders, with ultimate appendages of leaves or fertile organs, mesarch primary xylem and a bifacial cambium producing secondary xylem, phloem and vascular rays. Minimum age: 385.571 Ma. Soft maximum age: 451 Ma. Age justification: Rellimia thomsonii was collected from the Panther Mountain Formation of Schoharie County, New York, which is equivalent to Skaneateles and Ludlowville Formations of the Hamilton Group of northwestern New York. The Moscow Formation overlies the Ludlowville Formation [164], the boundary occurring deep within the lower varcus conodont Biozone [165], but at the very least is below the rhenanus-ansatus biozonal boundary, which has been dated to 386.25 Ma ± 0.679 Myr [87]. Therefore, the minimum age constraint for this node is 385.571 Ma. Following Clarke et al. [62], the soft maximum age constraint is based on the first appearance of trilete spores, in the Qusaiba-1 core from the Qasim Formation, northern Saudi Arabia [139]. This formation is Katian (Late Ordovician) in age based on the presence of Acanthochitina barbata, of the barbata Biozone. The oldest trilete spores precede the earliest occurrences of A. barbata, thus it is a possibility that they occur in the preceding Tanuchitina fistulosa Biozone, though T. fistulosa does not occur. The oldest trilete spores within the core co-occur with Armoricochitina nigerica, known to extend into the British Caradoc stage, within the Fungochitina spinifera Biozone (= F. fungiformis) [140]. The base of the F. spinifera Biozone falls within the Dicranograptus clingani Biozone (Dicellograptus morrisi sub-biozone) [141], the base of which is estimated to be 451 Ma [17]. Discussion: This node represents the divergence of Euphyllophyta into Monilophyta (after Cantino et al. [166] and Spermatophyta. One possible monilophyte is Estinnophyton yunnanense from the Pragian Posongchong Formation of Yunnan, China, which is considered to be a putative sphenopsid based on the pseudowhorled arrangement of leaves [167, 168]. However, this assignment is tentative, with an absence of anatomical data. A second possible oldest monilophyte is Foozia, from the Emsian Grés de Whépion Formation, Belgium [169]. However, the only synapomorphy of Monilophyta, a mesarch protoxylem confined to lobes of the xylem strand, has not been demonstrated in Foozia, and it is unclear if it is a crown or stem euphyllophyte. The oldest unequivocal monilophyte is considered to be Ibyka amphikoma [170], based on the presence of a mesarch protoxylem. It was recovered from the Manorkill Shale Member at Gilboa, Schoharie County, New York State. The Manorkill Shale Member of the Moscow Formation is the lateral equivalent of the marine Windom Member [171, 172], which falls within the conodont Polygnathus ansatus Biozone [173, 174], the top of which is dated to 385.41 Ma ± 0.704 Myrs [87]. The miniumum age for Ibyka amphikoma is therefore 384.706 Ma, slightly younger than the minimum age for Rellimia. The oldest possible record of total group Spermatophyta is Pertica [175], interpreted as a trimerophyte, but is only tentatively placed as a stem spermatophyte by Kenrick and Crane [69] because of a lack of anatomical data. Other candidates for stem spermatophytes include Polythecophyton demissum [176] and Hedeia sinica [151] from the Pragian Posongchong Formation, Yunnan, China, interpreted as putative aneurophytalean progymnosperms [168], based on branching patterns, but a lack of anatomical data means that they remain equivocal. Node 164: Monilophyta: Equisetopsida – [Polypodiopsida + [Psilotopsida + Marattiopsida]]; 384.706 Ma – 451 Ma.

Fossil taxon and specimen: Ibyka amphikoma [No. 179 (holotype): Paleobotanical Collection of Cornell University, NY], from the Manorkill Shale Member of the Moscow Formation, Givetian Stage (Middle Devonian), from a locality on the eastern bank of Schoharie Creek, directly below the spillway of Gilboa dam, Gilboa, Schoharie County, New York State [170]. Phylogenetic justification: The oldest unequivocal monilophyte is considered to be Ibyka amphikoma [170], based on the presence of a mesarch protoxylem and phylogenetic analysis [69]. Minimum age: 384.706 Ma. Soft maximum age: 451 Ma. Age justification: Ibyka amphikoma was recovered from the Manorkill Shale Member of the Moscow Formation, near Gilboa dam, Schoharie County, New York State. The Manorkill Shale Member is the lateral equivalent of the marine Windom Member [171, 172], which falls within the conodont Polygnathus ansatus Biozone [173, 174], the top of which is dated to 385.41 Ma ± 0.704 Myr [87]. Therefore, the minimum age for Ibyka amphikoma is 384.706 Ma. Following Clarke et al. [62], the soft maximum age constraint is based on the first appearance of trilete spores, in the Qusaiba-1 core from the Qasim Formation, northern Saudi Arabia [139]. This formation is Katian (Late Ordovician) in age based on the presence of Acanthochitina barbata, of the barbata Biozone. The oldest trilete spores precede the earliest occurrences of A. barbata, thus it is a possibility that they occur in the preceding Tanuchitina fistulosa Biozone, though T. fistulosa does not occur. The oldest trilete spores within the core co-occur with Armoricochitina nigerica, known to extend into the British Caradoc stage, within the Fungochitina spinifera Biozone (= F. fungiformis) [140]. The base of the F. spinifera Biozone falls within the Dicranograptus clingani Biozone (Dicellograptus morrisi sub-biozone) [141], the base of which is estimated to be 451 Ma [17]. Discussion: This node represents the divergence of Monilophyta into Equisetopsida (horsetails) and the remaining ferns; Polypodiopsida (leptosporangiate ferns), and the Psilotopsida and Marattiopsida. The oldest unequivocal record of Polypodiopsida is Senftenbergia plumosa, assigned to the extinct fern Tedeleaceae family, from the Pennsylvanian (Carboniferous) Kladno Formation of the Pilsen Basin, Bohemian Massif [177]. This is younger than the oldest unequivocal record of Equisetopsida, Ibyka amphikoma of Givetian (Middle Devonian) age. The oldest record of Marattiophyta is Psaronius simplicicaulis, a tree fern with a crown of pinnate fronds, from the Middle Pennsylvanian (Carboniferous) of Illinois, USA [178]. The fossil record of Psilotopsida is poor, the oldest unequivocal record of Ophiogloassales being Botrychium from the Palaeocene of Canada [179]. Node 165: [Psilotopsida + Marattiopsida] – Polypodiopsida; 318.71 Ma – 451 Ma. Fossil taxon and specimen: Psaronius simplicicaulis [10646, 10648, 10649: Paleobotanical Collections, Morrill Hall, Dept. of Botany, University of Illinois, Illinois, U.S.A.], from the Caseyville Formation of the McCormick Group, Morrowan (middle Bashkirian), Early Pennslyvanian (Carboniferous), from a locality in the Allied Stone Company quarry, Vandruff Island, Milan, Rock Island County, Illinois Basin, U.S.A. [178]. Phylogenetic justification: Psaronius is considered to be a marattialean tree fern [178]. Minimum age: 318.71 Ma. Soft maximum age: 451 Ma. Age justification: Psaronius simplicicaulis was collected from the Caseyville Formation of the McCormick Group, from a locality in the western part of the Illinois Basin [178]. At this locality the formation comprises shale channel fills lying unconformably over the Devonian Cedar Valley Limestone [178, 182]. The exact age of this formation at this locality is unknown, but similar channel fill shales overlying the Cedar Valley Limestone in Brown County, Il. are dated as Morrowan in age, based on palynology and compression floras [183-185]. The boundary between the North American Morrowan and the younger Atokan Series, is based on the FAD of the fusulinid Eoschubertella [181]. This is correlated to the base of the Western European Westphalian A, the base of which is defined by the Gastrioceras subcrenatum marine band (Pa4) [181]. This is correlated to the base of the Declinograptus margindosus (Pc5) Biozone. According to the Carboniferous Composite Standard of Davydov et al. [181], a Composite Level dated at 319.09 Ma ± 0.38 Myr occurs between the older Idiognathodus sinuosus Biozone (Pc4) and Pc5, within the middle part of the Bashkirian [181, 186]. Thus, the minimum age of the base of Pc5 is estimated as 318.71 Ma. Following Clarke et al. [62], the soft maximum age constraint is based on the first appearance of trilete spores, in the Qusaiba-1 core from the Qasim Formation, northern Saudi Arabia [139]. This formation is Katian (Late Ordovician) in age based on the presence of Acanthochitina barbata, of the barbata Biozone. The oldest trilete spores precede the earliest occurrences of A. barbata, thus it is a possibility that they occur in the preceding Tanuchitina fistulosa Biozone, though T. fistulosa does not occur. The oldest trilete spores within the core co-occur with Armoricochitina nigerica, known to extend into the British Caradoc stage, within the Fungochitina spinifera Biozone (= F. fungiformis) [140]. The base of the F. spinifera Biozone falls within the Dicranograptus clingani Biozone (Dicellograptus morrisi sub-biozone) [141], the base of which is estimated to be 451 Ma [17]. Discussion: The oldest unequivocal record of Polypodiopsida is Senftenbergia plumosa, assigned to the extinct fern Tedeleaceae family, from the Pennsylvanian (Carboniferous) Kladno Formation of the Pilsen Basin, Bohemian Massif, with a minimum age of 315.0 Ma [177]. The oldest record of Marattiophyta is Psaronius simplicicaulis, a tree fern with a

crown of pinnate fronds, from the Middle Pennsylvanian (Carboniferous) of Illinois, USA [178]. The fossil record of Psilotopsida is poor, the oldest unequivocal record of Ophiogloassales being Botrychium from the Palaeocene of Canada [179]. Node 114: Spermatophyta: Acrogymnospermae – Angiospermae; 308.14 Ma – 365.629 Ma. Fossil taxon and specimen: Cordaixylon iowensis [UIC 12,233, OUPH 9616-9742: Ohio University Paleobotanical Herbarium, Department of Botany, Ohio University, Athens, Ohio] from coal balls from the Laddsdale Coal, Lower Cherokee Group, Desmoinesian (upper Moscovian – Kasimovian), Middle – Upper Pennsylvanian (Carboniferous), near What Cheer, Iowa [187]. Phylogenetic justification: Clarke et al. [62] identify cordaitean coniferophytes as the oldest records of the crown group of the spermatophyte clade. The oldest whole plant reconstruction is Cordaixylon iowensis from the Laddsdale Coals, Cherokee Group, near What Cheer, Iowa [187]. Minimum age: 308.14 Ma. Soft maximum age: 365.629 Ma. Age justification: Janousek and Pope [188] argue that the Laddsdale Coal is equivalent to the Bluejacket Coal of Oklahoma, which occurs as part of the Bluejacket Sandstone Member, underlying the Inola Limestone, part of the Inola Cyclothem of the Krebs subgroup of the Cherokee Group, characterized by the occurrence of the conodonts Idiognathodus amplificus, Idiognathodus podolskensis and Neognathodus asymmetricus [189]. The Inola cyclothem falls fully within the Idiognathodus amplificus/ Idiognathodus obliquus Biozone [190]. This is indicative of the Neognathodus medexultimus – Streptognathodus concinnus (Pc10) Biozone, certainly older than the Neognathodus roundyi – Streptognathodus cancellosus (Pc11) Biozone [181, 190]. The base of Pc10 is bracketed by an older age constraint of 312.01 Ma ± 0.37 Myr and the base of Pc11 is bracketed by a younger age constraint of 308.5 Ma ± 0.36 Myr in the Composite Standard of Davydov et al. [181], yielding a minimum constraint of 308.14 Ma. The soft maximum constraint follows Clarke et al. [62], who based theirs on the first records of seeds in the form of preovules that satisfy the criteria of the seed habit, which occur in the Upper Fammenian (Late Devonian) VCo Spore Biozone [191], a well-documented b example of which being Elkinsia polymorpha [192]; E. polymorpha has been recovered from the Hampshire Formation, West Virginia, from which the palynomorphs Grandispora cornuta, Retispora macroreticulata, Retusotriletes phillipsii and Rugospora radiata have been reported [193], which substantiate assignment to the VCo Biozone [194]. The VCo Biozone is not directly dated but its base falls within the Palmatolepis trachytera conodont Biozone [195], the base of which is dated to 364.19 Ma ± 1.439 Myr [87], yielding a soft maximum constraint on the divergence of crown Spermatophyta at 365.629 Ma. Node 151: Acrogymnospermae: Pinopsida – [Ginkgoopsida + Cycadopsida]; 308.14 Ma – 365.629 Ma Fossil taxon and specimen: Cordaixylon iowensis [UIC 12,233, OUPH 9616-9742: Ohio University Paleobotanical Herbarium, Department of Botany, Ohio University, Athens, Ohio] from coal balls from the Laddsdale Coal, Lower Cherokee Group, Desmoinesian (upper Moscovian – Kasimovian), Middle – Upper Pennsylvanian (Carboniferous), near What Cheer, Iowa [187]. Phylogenetic justification: Clarke et al. [62] identify cordaitean coniferophytes as the oldest records of the crown group of the Acrogymnospermae clade. The oldest whole plant reconstruction is Cordaixylon iowensis from the Laddsdale Coals, Cherokee Group, near What Cheer, Iowa [187]. Minimum age: 308.14 Ma. Soft maximum age: 365.629 Ma. Age justification: Janousek and Pope [188] argue that the Laddsdale Coal is equivalent to the Bluejacket Coal of Oklahoma, which occurs as part of the Bluejacket Sandstone Member, underlying the Inola Limestone, part of the Inola Cyclothem of the Krebs subgroup of the Cherokee Group, characterized by the occurrence of the conodonts Idiognathodus amplificus, Idiognathodus podolskensis and Neognathodus asymmetricus [189]. The Inola cyclothem falls fully within the Idiognathodus amplificus/ Idiognathodus obliquus Biozone [190]. This is indicative of the Neognathodus medexultimus – Streptognathodus concinnus (Pc10) Biozone, certainly older than the Neognathodus roundyi – Streptognathodus cancellosus (Pc11) Biozone [181, 190]. The base of Pc10 is bracketed by an older age constraint of 312.01 Ma ± 0.37 Myr and the base of Pc11 is bracketed by a younger age constraint of 308.5 Ma ± 0.36 Myr in the Composite Standard of Davydov et al. [181], yielding a minimum constraint of 308.14 Ma. The soft maximum constraint follows Clarke et al. [62], who based theirs on the first records of seeds in the form of preovules that satisfy the criteria of the seed habit, which occur in the Upper Fammenian (Late Devonian) VCo Spore Biozone [191], a well-documented example of which being Elkinsia polymorpha [192]; E. polymorpha has been recovered from the Hampshire Formation, West Virginia, from which the palynomorphs Grandispora cornuta, Retispora macroreticulata, Retusotriletes phillipsii and Rugospora radiata have been reported [193], which substantiate assignment to the VCo Biozone [194]. The VCo Biozone is not directly dated but its base falls within the Palmatolepis trachytera

conodont Biozone [195], the base of which is dated to 364.19 Ma ± 1.439 Myr [87], yielding a soft maximum constraint on the divergence of crown Acrogymnospermae at 365.629 Ma. Node 161: Ginkgoopsida – Cycadopsida; 264.7 Ma – 365.629 Ma. Fossil taxon and specimen: Crossozamia chinensis [GP0027: Beijing Graduate School, China Institute of Mining, Beijing, China], Lower Shihhotse Formation, Roadian-Wordian, (Middle Permian) at Simugedong, Dongshan (East Hills), Taiyuan, north China [196]. Phylogenetic justification: Nagalingum et al. [197] identify Crossozamia as the oldest record of the Cycas lineage, based on megasporophylls that exhibit similarity to extant Cycas [198]. They argue against the interpretation of Crossozamia as the sister lineage of Cycas based on the presence of an estipulate leaf base and a terminal pinna found in the seedlings [198], instead favouring its assignment to the cycad stem. The arguments presented clearly raise doubts about the assignment of Crossozamia to crown-cycads, however, they do not provide definitive evidence of its exclusion from this clade and so Crossozamia may more appropriately be assigned to the cycad total group (i.e. we cannot discriminate between a stem or crown-cycad affinity based on the available evidence). In either instance, Crossozamia is the oldest record of the minimal clade comprised of Gingko and Cycas. Minimum age: 264.7 Ma. Soft maximum age: 365.629 Ma. Age justification: The Lower Shihhotse Formation at Simugedong, Dongshan (East Hills), Taiyuan, north China [196] has been established biostratigraphically as Roadian-Wordian (middle Permian) [199] and, thus a minimum age constraint can be established on the Wordian-Capitanian Boundary which has been dated to 265.1 Ma ± 0.4 Myr [123]. Thus, the minimum age constraint on the Cycas-Ginkgo clade is 264.7 Ma. The soft maximum constraint follows Clarke et al. [62], who based theirs on the first records of seeds in the form of preovules that satisfy the criteria of the seed habit, which occur in the Upper Fammenian (Late Devonian) VCo Spore Biozone [191], a well-documented example of which being Elkinsia polymorpha [192]; E. polymorpha has been recovered from the Hampshire Formation, West Virginia, from which the palynomorphs Grandispora cornuta, Retispora macroreticulata, Retusotriletes phillipsii and Rugospora radiata have been reported [193], which substantiate assignment to the VCo Biozone [194]. The VCo Biozone is not directly dated but its base falls within the Palmatolepis trachytera conodont Biozone [195], the base of which is dated to 364.19 Ma ± 1.439 Myr [87], yielding a soft maximum constraint of 365.629 Ma. Node 152: Pinopsida: [Pinaceae + Gnetales] – Cupressophyta; 147 Ma – 321.38 Ma. Fossil taxon and specimen: Araucaria mirabilis [NHM V.30953: Natural History Museum, London, UK], represented by cones from the Cerro Cuadrado Petrified Forest, La Matilde Formation, Late Jurassic, Patagonia, Argentina [200 – 203]. Phylogenetic justification: These fossils possess a ‘vascular plexus’ at the ovule base, ovuliferous scale vascularization, two vascular strands to the conescale complex and an embryo with two cotyledons, all characters established to distinguish Araucaria Section Bunya of the Araucariaceae (Cupressophyta) [202, 204], to which only extant Araucaria bidwillii belongs. Minimum age: 147 Ma. Soft maximum age: 312.38 Ma. Age justification: The age of La Matilde Formation is poorly constrained, as the stratigraphy is complex, although the volcanic deposits do allow radiometric dating. La Matilde Formation is overlain by volcanics dated to 157 Ma ± 10 Myr [205], and thus the minimum constraint on the crown group Pinopsida is 147 Ma, within the Late Jurassic. A soft maximum constraint can be based on Cordaixylon iowensis, a cordaitean coniferophyte from the Laddsdale Coals, Cherokee Group, near What Cheer, Iowa [187], as the oldest whole plant reconstruction for the Acrogymnospermae clade. Janousek and Pope [188] argue that the Laddsdale Coal is equivalent to the Bluejacket Coal of Oklahoma, which occurs as part of the Bluejacket Sandstone Member, underlying the Inola Limestone, part of the Inola Cyclothem of the Krebs subgroup of the Cherokee Group, characterized by the occurrence of the conodonts Idiognathodus amplificus, Idiognathodus podolskensis and Neognathodus asymmetricus [189]. The Inola cyclothem falls fully within the Idiognathodus amplificus/ Idiognathodus obliquus Biozone [190]. This is indicative of the Neognathodus medexultimus – Streptognathodus concinnus (Pc10) Biozone, certainly older than the Neognathodus roundyi – Streptognathodus cancellosus (Pc11) Biozone [181, 190]. The base of Pc10 is bracketed by an older age constraint of 312.01 Ma ± 0.37 Myr and the base of Pc11 is bracketed by a younger age constraint of 308.5 Ma ± 0.36 Myr in the Composite Standard of Davydov et al. [181], yielding a soft maximum constraint of 321.38 Ma. Discussion: This is the fundamental divergence of Pinopsida into Pinaceae plus Gnetales and Cupressophyta (Araucariaceae, Podocarpaceae, Taxaceae, Cupressaceae and Sciadopityaceae; after Cantino et al. [166]). The oldest possible records of the gnepine total group occur within the Yixian Formation of Liaoning, China, e.g. Liaoxia chenii

[206], the minimum age of which is 121.8 Ma [207]. The oldest possible records of Cupressophyta total group include Triassic Rissikia media [208], but it lacks the Podocarpaceae diagnostic feature of one ovule per cone scale, instead possessing two [62]. Other Triassic and Jurassic records are equally problematic [209-211]. The next unequivocal member of Cupressophyta is Araucaria mirabilis. Node 153: Pinaceae – Gnetales; 119.6 Ma – 312.38 Ma. Fossil taxon and specimen: Eoantha zherikhinii [N TB 15-600: Repository of the Institute of Biology and Pedology, Vladivostok, Russia], from the Zaza Formation, Aptian (Early Cretaceous), at the Baisa locality in the upper reaches of the Vitim River in Lake Baikal [212]. Phylogenetic justification: This fossil is assigned to Gnetales because of the presence of an ovule with an extending micropylar tube and the presence of polyplicate pollen of Ephedritopes-type [213]. Eoantha is one out of several Early Cretaceous fossils unequivocally assigned to the crown Gnetales [214]. Minimum age: 119.6 Ma. Soft Maximum age: 312.38 Ma. Age justification: The Zaza Formation can be correlated with the Turga Formation, also of Transbaikalia based principally on common elements of their floral assemblages, including Asteropollis asteroids, Dicotylophyllum pusilum, Baisa hirsuita, Podozamites, Schizolepis, Pseudolarix, Phoenicopsis, Czekanowskia rigida and Sphenobaiera [212, 215-217]. The age of the Turga flora and Formation is based on the chronological distribution of Asteropollis type pollen, but correlation with the Yixian Formation of China is also supported strongly [216], allowing for refinement of the Asteropollis-derived ages. Correlation between Turga and Yixian is based on similarities in the floral assemblages of these two formations, with the shared presence of the species Baisa hirsuita, Botrychites reheensis, Neozamites verchojanensis, Pityolepis pseudotsugaoides, Brachyphyllum longispicum, Scarbugia hilii, Ephedrites chenii, Carpolithus multiseminalis, Carpolithus pachythelis, Schizolepis, Baiera, Coniopteris, Ginkoites, Pityocladus, Pityospermum and Elatocladus [212, 215, 218]. The shared presence of Asteropollis asteroides in Turga and Zaza can be used to constrain their age. The last appearance of Asteropollis pollen is in Antarctica [219] and is dated to the end-Campanian, at the latest 72.1 Ma ± 0.2 [220]. This minimum may be constrained further based on the correlation of the Zaza Formation through the Turga Formation to the Yixian Formation. The main fossil bearing beds in the Yixian Formation have been recently dated and may be as old as 129.2 Ma [221], however, in the absence of knowledge of the position of the fossils within the stratigraphy, relative to the sources of the absolute dates, a minimum age constraint can be derived from the Jiufontang Formation which overlies it. 40Ar/39Ar dating of a number of samples from the Jiufontang Formation has yielded an age of 120.3 ± 0.7 Ma for the volcanic tuffs [222], establishing a minimum constraint of 119.6 Ma for the age of the Yixian, Formation and, thus ultimately the Zaza Formation. A soft maximum constraint can be based on Cordaixylon iowensis, a cordaitean coniferophyte from the Laddsdale Coals, Cherokee Group, near What Cheer, Iowa [187], as the oldest whole plant reconstruction for the Acrogymnospermae clade. Janousek and Pope [188] argue that the Laddsdale Coal is equivalent to the Bluejacket Coal of Oklahoma, which occurs as part of the Bluejacket Sandstone Member, underlying the Inola Limestone, part of the Inola Cyclothem of the Krebs subgroup of the Cherokee Group, characterized by the occurrence of the conodonts Idiognathodus amplificus, Idiognathodus podolskensis and Neognathodus asymmetricus [189]. The Inola cyclothem falls fully within the Idiognathodus amplificus/ Idiognathodus obliquus Biozone [190]. This is indicative of the Neognathodus medexultimus – Streptognathodus concinnus (Pc10) Biozone, certainly older than the Neognathodus roundyi – Streptognathodus cancellosus (Pc11) Biozone [181, 190]. The base of Pc10 is bracketed by an older age constraint of 312.01 Ma ± 0.37 Myr and the base of Pc11 is bracketed by a younger age constraint of 308.5 Ma ± 0.36 Myr in the Composite Standard of Davydov et al. [181], yielding a soft maximum constraint of 321.38 Ma. Node 115: Angiospermae: Amborellales – [[Nymphaeales + Austrobaileyales] + Mesoangiospermae]; 125 Ma – 247.2 Ma. Fossil taxon and specimen: Tricolpate pollen grain [palynological sample BRN 126], from the middle Atherfield Wealden Bed 35 of the Cowleaze Chine Member, Vectis Formation, Barremian (Early Cretaceous), of the Isle of Wight [223]. Phylogenetic justification: Following Clarke et al. [62], our minimum age constraint is based on the earliest occurrences Fischer’s rule tricolpate pollen, and knowledge of the distribution of tricoplate pollen across the phylogeny of angiosperms [224]. Minimum age: 125 Ma. Soft maximum age: 247.2 Ma. Age justification: Following Clarke at al. [62], the Cowleaze Chine Member of the Vectis Formation of the Isle of Wight [223] occurs within the M1n polarity chron at the top of the Barremian, the top of which is dated at 125 Ma (International Chronostratigraphic Chart v.2016/10).

The soft maximum age constraint is based on sediments devoid of angiosperm-like pollen below their first report in the Middle Triassic. Pollen types C – F of Hochuli and Feist-Burkhardt [225] from the Snadd Formation, Norwegian Arctic, and type C ‘Retisulcites’ from the older Steinkobbe Formation of Hochuli et al. [226], represent the oldest pollen which possess synapomorphy combinations most similar to Cretaceous grains. The Steinkobbe Formation is considered to be middle Anisian in age [227], thus the maximum age of these pollen grains is at the base of the Anisian, dated at 247.2 Ma (International Chronostratigraphic Chart, v.2016/10). Discussion: The recently described Euanthus panii [228], Juraherba bodae [229] and Yuhania dahugouensis [230] from the Jiulongshan Formation were considered but not assigned. At the current stage, the age of the formation appears to be still not fully settled despite most experts agree on a middle Jurassic age (see 229, 230), whereas the assignment to extant lineages also required further investigation using phylogenetic approaches to confirm the proposed relationships of Juraherba to Hydatellaceae - which are the sister to the remaining Nymphaeales lineage and Yuhania to monocots. Node 116: Nymphaeales – [Austrobaileyales + Mesoangiospermae]; 125 Ma – 247.2 Ma. Fossil taxon and specimen: Tricolpate pollen grain [palynological sample BRN 126], from the middle Atherfield Wealden Bed 35 of the Cowleaze Chine Member, Vectis Formation, Barremian (Early Cretaceous), of the Isle of Wight [223]. Phylogenetic justification: Following Clarke et al. [62], our minimum age constraint is based on the earliest occurrences Fischer’s rule tricolpate pollen, and knowledge of the distribution of tricoplate pollen across the phylogeny of angiosperms [224]. Minimum age: 125 Ma. Soft maximum age: 247.2 Ma. Age justification: Following Clarke at al. [62], the Cowleaze Chine Member of the Vectis Formation of the Isle of Wight [223] occurs within the M1n polarity chron at the top of the Barremian, the top of which is dated at 125 Ma (International Chronostratigraphic Chart v.2016/10). The soft maximum age constraint is based on sediments devoid of angiosperm-like pollen below their first report in the Middle Triassic. Pollen types C – F of Hochuli and Feist-Burkhardt [225] from the Snadd Formation, Norwegian Arctic, and type C ‘Retisulcites’ from the older Steinkobbe Formation of Hochuli et al. [226], represent the oldest pollen which possess synapomorphy combinations most similar to Cretaceous grains. The Steinkobbe Formation is considered to be middle Anisian in age [227], thus the maximum age of these pollen grains is at the base of the Anisian, dated at 247.2 Ma (International Chronostratigraphic Chart, v.2016/10). Discussion: The recently described Euanthus panii [228], Juraherba bodae [229] and Yuhania dahugouensis [230] from the Jiulongshan Formation were considered but not assigned. At the current stage, the age of the formation appears to be still not fully settled despite most experts agree on a middle Jurassic age (see 229, 230), whereas the assignment to extant lineages also required further investigation using phylogenetic approaches to confirm the proposed relationships of Juraherba to Hydatellaceae - which are the sister to the remaining Nymphaeales lineage and Yuhania to monocots. Node 117: Austrobaileyales – Mesoangiospermae; 125 Ma – 247.2 Ma.
Fossil taxon and specimen: Tricolpate pollen grain [palynological sample BRN 126], from the middle Atherfield Wealden Bed 35 of the Cowleaze Chine Member, Vectis Formation, Barremian (Early Cretaceous), of the Isle of Wight [223]. Phylogenetic justification: Following Clarke et al. [62], our minimum age constraint is based on the earliest occurrences Fischer’s rule tricolpate pollen, and knowledge of the distribution of tricoplate pollen across the phylogeny of angiosperms [224]. Minimum age: 125 Ma. Soft maximum age: 247.2 Ma. Age justification: Following Clarke at al. [62], the Cowleaze Chine Member of the Vectis Formation of the Isle of Wight [223] occurs within the M1n polarity chron at the top of the Barremian, the top of which is dated at 125 Ma (International Chronostratigraphic Chart v.2016/10). The soft maximum age constraint is based on sediments devoid of angiosperm-like pollen below their first report in the Middle Triassic. Pollen types C – F of Hochuli and Feist-Burkhardt [225] from the Snadd Formation, Norwegian Arctic, and type C ‘Retisulcites’ from the older Steinkobbe Formation of Hochuli et al. [226], represent the oldest pollen which possess synapomorphy combinations most similar to Cretaceous grains. The Steinkobbe Formation is considered to be middle Anisian in age [227], thus the maximum age of these pollen grains is at the base of the Anisian, dated at 247.2 Ma (International Chronostratigraphic Chart, v.2016/10). Discussion: The recently described Euanthus panii [228], Juraherba bodae [229] and Yuhania dahugouensis [230] from the Jiulongshan Formation were considered but not assigned. At the current stage, the age of the formation appears to be still not fully settled despite most experts agree on a middle Jurassic age (see 229, 230), whereas the assignment to extant

lineages also required further investigation using phylogenetic approaches to confirm the proposed relationships of Juraherba to Hydatellaceae - which are the sister to the remaining Nymphaeales lineage and Yuhania to monocots. Node 118: Mesangiospermae: Monocotyledoneae – [Magnoliids + [Chloranthales + Eudicotyledoneae]]; 125 Ma – 247.2 Ma. Fossil taxon and specimen: Tricolpate pollen grain [palynological sample BRN 126], from the middle Atherfield Wealden Bed 35 of the Cowleaze Chine Member, Vectis Formation, Barremian (Early Cretaceous), of the Isle of Wight [223]. Phylogenetic justification: Following Clarke et al. [62], our minimum age constraint is based on the earliest occurrences Fischer’s rule tricolpate pollen, and knowledge of the distribution of tricoplate pollen across the phylogeny of angiosperms [224]. Minimum age: 125 Ma. Soft maximum age: 247.2 Ma. Age justification: Following Clarke at al. [62], the Cowleaze Chine Member of the Vectis Formation of the Isle of Wight [223] occurs within the M1n polarity chron at the top of the Barremian, the top of which is dated at 125 Ma (International Chronostratigraphic Chart v.2016/10). The soft maximum age constraint is based on sediments devoid of angiosperm-like pollen below their first report in the Middle Triassic. Pollen types C – F of Hochuli and Feist-Burkhardt [225] from the Snadd Formation, Norwegian Arctic, and type C ‘Retisulcites’ from the older Steinkobbe Formation of Hochuli et al. [226], represent the oldest pollen which possess synapomorphy combinations most similar to Cretaceous grains. The Steinkobbe Formation is considered to be middle Anisian in age [227], thus the maximum age of these pollen grains is at the base of the Anisian, dated at 247.2 Ma (International Chronostratigraphic Chart, v.2016/10). Discussion: The recently described Euanthus panii [228], Juraherba bodae [229] and Yuhania dahugouensis [230] from the Jiulongshan Formation were considered but not assigned. At the current stage, the age of the formation appears to be still not fully settled despite most experts agree on a middle Jurassic age (see 229, 230), whereas the assignment to extant lineages also required further investigation using phylogenetic approaches to confirm the proposed relationships of Juraherba to Hydatellaceae - which are the sister to the remaining Nymphaeales lineage and Yuhania to monocots. Node 128: Magnoliids – [Chloranthales + Eudicotyledoneae]; 125 Ma – 247.2 Ma. Fossil taxon and specimen: Tricolpate pollen grain [palynological sample BRN 126], from the middle Atherfield Wealden Bed 35 of the Cowleaze Chine Member, Vectis Formation, Barremian (Early Cretaceous), of the Isle of Wight [223]. Phylogenetic justification: Following Clarke et al. [62], our minimum age constraint is based on the earliest occurrences Fischer’s rule tricolpate pollen, and knowledge of the distribution of tricoplate pollen across the phylogeny of angiosperms [224]. Minimum age: 125 Ma. Soft maximum age: 247.2 Ma. Age justification: Following Clarke at al. [62], the Cowleaze Chine Member of the Vectis Formation of the Isle of Wight [223] occurs within the M1n polarity chron at the top of the Barremian, the top of which is dated at 125 Ma (International Chronostratigraphic Chart v.2016/10). The soft maximum age constraint is based on sediments devoid of angiosperm-like pollen below their first report in the Middle Triassic. Pollen types C – F of Hochuli and Feist-Burkhardt [225] from the Snadd Formation, Norwegian Arctic, and type C ‘Retisulcites’ from the older Steinkobbe Formation of Hochuli et al. [226], represent the oldest pollen which possess synapomorphy combinations most similar to Cretaceous grains. The Steinkobbe Formation is considered to be middle Anisian in age [227], thus the maximum age of these pollen grains is at the base of the Anisian, dated at 247.2 Ma (International Chronostratigraphic Chart, v.2016/10). Discussion: The recently described Euanthus panii [228], Juraherba bodae [229] and Yuhania dahugouensis [230] from the Jiulongshan Formation were considered but not assigned. At the current stage, the age of the formation appears to be still not fully settled despite most experts agree on a middle Jurassic age (see 229, 230), whereas the assignment to extant lineages also required further investigation using phylogenetic approaches to confirm the proposed relationships of Juraherba to Hydatellaceae - which are the sister to the remaining Nymphaeales lineage and Yuhania to monocots. Node 129: Magnoliids: [Magnoliales + Laurales] – Piperales; 110.87 Ma – 247.2 Ma. Fossil taxon and specimen: Endressinia brasiliana [MB. PB. 2001/1455: Museum of Natural History, Institute of Paleontology, Berlin, Germany], from the Crato Formation, early Albian (Early Cretaceous), from Nova Olinda, Santana do Cariri, northeastern Brazil [231].

Phylogenetic justification: Massoni et al. [232] identify both Schenkeriphyllum glanduliferum and Endressinia brasiliana, both from the Crato Formation of Brazil [233, 233], as the oldest records of crown Magnoliineae, the sister clade of Myristicaceae [234], based on the phylogenetic analyses [233, 235, 236]. Minimum age: 110.87 Ma. Soft maximum age: 247.2 Ma. Age justification. Clarke et al. [62] argued that the age of the Crato Formation could not be constrained to being definitively older than Albian based on pollen [237], ostracod [238], and dinoflagellate [239] biostratigraphy and, in the absence of further evidence, established a minimum constraint on the Albian-Cenomanian boundary. Massoni et al. [232] argued for an Aptian age for the Crato Formation based on evidence from Heimhofer and Hochuli [239] but, unfortunately, these authors do not present evidence that can discriminate against a possible early Albian age for the Crato Formation, as acknowledged by Mohr et al. [233]. The evidence suggests at worst an early Albian age for the Crato Formation, and so it is possible to derive a minimum age interpretation for the Formation based on the Early-Middle Albian Boundary, which coincides approximately with the base of the Douvilleiceras mammillatum ammonite biozone, dated to 110.87 Ma [220]. The soft maximum age constraint is based on sediments devoid of angiosperm-like pollen below their first report in the Middle Triassic. Pollen types C – F of Hochuli and Feist-Burkhardt [225] from the Snadd Formation, Norwegian Arctic, and type C ‘Retisulcites’ from the older Steinkobbe Formation of Hochuli et al. [226], represent the oldest pollen which possess synapomorphy combinations most similar to Cretaceous grains. The Steinkobbe Formation is considered to be middle Anisian in age [227], thus the maximum age of these pollen grains is at the base of the Anisian, dated at 247.2 Ma (International Chronostratigraphic Chart, v.2016/10). Discussion: Archaeanthus linnenbergii was recognized as a further putative stem group Magnoliids but it is younger than Endressinia [232, 236]. Node 130: Persea (Laurales) – Liriodendron (Magnoliales); 110.87 Ma – 247.2 Ma. Fossil taxon and specimen: Endressinia brasiliana [MB. PB. 2001/1455: Museum of Natural History, Institute of Paleontology, Berlin, Germany], from the Crato Formation, early Albian (Early Cretaceous), from Nova Olinda, Santana do Cariri, northeastern Brazil [231]. Phylogenetic justification: Massoni et al. [232] identify both Schenkeriphyllum glanduliferum and Endressinia brasiliana, both from the Crato Formation of Brazil [233, 233], as the oldest records of crown Magnoliineae, the sister clade of Myristicaceae [234], based on the phylogenetic analyses [233, 235, 236]. Minimum age: 110.87 Ma. Soft maximum age: 247.2 Ma. Age justification. Clarke et al. [62] argued that the age of the Crato Formation could not be constrained to being definitively older than Albian based on pollen [237], ostracod [238], and dinoflagellate [239] biostratigraphy and, in the absence of further evidence, established a minimum constraint on the Albian-Cenomanian boundary. Massoni et al. [232] argued for an Aptian age for the Crato Formation based on evidence from Heimhofer and Hochuli [239] but, unfortunately, these authors do not present evidence that can discriminate against a possible early Albian age for the Crato Formation, as acknowledged by Mohr et al. [233]. The evidence suggests at worst an early Albian age for the Crato Formation, and so it is possible to derive a minimum age interpretation for the Formation based on the Early-Middle Albian Boundary, which coincides approximately with the base of the Douvilleiceras mammillatum ammonite biozone, dated to 110.87 Ma [220]. The soft maximum age constraint is based on sediments devoid of angiosperm-like pollen below their first report in the Middle Triassic. Pollen types C – F of Hochuli and Feist-Burkhardt [225] from the Snadd Formation, Norwegian Arctic, and type C ‘Retisulcites’ from the older Steinkobbe Formation of Hochuli et al. [226], represent the oldest pollen which possess synapomorphy combinations most similar to Cretaceous grains. The Steinkobbe Formation is considered to be middle Anisian in age [227], thus the maximum age of these pollen grains is at the base of the Anisian, dated at 247.2 Ma (International Chronostratigraphic Chart, v.2016/10). Discussion: Archaeanthus linnenbergii was recognized as a further putative stem group Magnoliales but it is younger than Endressinia [232, 236]. Node 131: Piperales: Saururus – Houttuynia; 44.3 Ma – 247.2 Ma. Fossil taxon and specimen: Saururus tuckerae [UAPC P1631 Bbot a: University of Alberta (Edmonton) Paleobotanical Collections] from the Middle Eocene Princeton Chert, British Columbia, Canada [240]. Phylogenetic justification: Massoni et al. [232] follow Smith and Stockey [240] in identifying Saururus tuckerae as the oldest record of total group Saururus. Based on tens of flowers and a partial inflorescence, the flower structure and pollen are characteristic of Saururaceae (Piperales), and phylogenetic analyses resolved S. tuckerae as the sister clade to extant Saururus.

Minimum age: 44.3 Ma. Soft maximum age: 247.2 Ma. Age justification: The Princeton Chert is part of the Allenby Formation which has been the subject of a number of absolute dating studies yielding age estimates of 48 Ma ± 2 Myr [241, 242], between 47 Ma ± 2 Myr and 50 Ma ± 2 Myr [243], between 46.2 Ma ± 1.9 Myr and 49.4 Ma ± 2 Myr [244], and 52.08 Ma ± 0.12 Myr [245] for the Allenby Formation. We follow Massoni et al. [232] in basing our minimum constraint based on the youngest age interpretation of the youngest radiometric age estimate, viz. 44.3 Ma. The soft maximum age constraint is based on sediments devoid of angiosperm-like pollen below their first report in the Middle Triassic. Pollen types C – F of Hochuli and Feist-Burkhardt [225] from the Snadd Formation, Norwegian Arctic, and type C ‘Retisulcites’ from the older Steinkobbe Formation of Hochuli et al. [226], represent the oldest pollen which possess synapomorphy combinations most similar to Cretaceous grains. The Steinkobbe Formation is considered to be middle Anisian in age [227], thus the maximum age of these pollen grains is at the base of the Anisian, dated at 247.2 Ma (International Chronostratigraphic Chart, v.2016/10). Node 132: Chloranthales – Eudicotyledoneae; 119.6 Ma – 128.63 Ma. Fossil taxon and specimen: Hyrcantha decussata [NJU-DES02001: Geological Institute, Chinese Academy of Sciences, Beijing], from the lower part of the Yixian Formation, Jehol Group, Aptian (Early Cretaceous), from the Liaoning Province, China [246]. Phylogenetic justification: Similar to Leefrutcus from the Yixian Formation of the Lower Cretaceous of China, Hycantha is considered to be a stem group representative of the Ranunculales [247]. Minimum age: 119.6 Ma. Soft maximum age: 128.63 Ma. Age justification: The main fossil bearing beds have been dated and may be as old as 129.2 Ma [221], however, in the absence of knowledge of the position of the fossils within the stratigraphy, relative to the sources of the absolute dates, a minimum age constraint can be derived from the Jiufontang Formation which overlies it. 40Ar/39Ar dating of a number of samples from the Jiufontang Formation has yielded an age of 120.3 ± 0.7 Ma for the volcanic tuffs [222], establishing a minimum constraint of 119.6 Ma. The soft maximum age constraint is based on the maximum age of the oldest potential age of tricolpate pollen, from the middle Atherfield Wealden Bed 35 of the Cowleaze Chine Member, Vectis Formation [223]. Following Clarke at al. [62], the Cowleaze Chine Member of the Vectis Formation of the Isle of Wight [223] occurs within the M1n polarity chron in the Late Barremian. The base of the M1n chron is approximately correlated with the base of the Gerhardtia sartousi Biozone, dated at 128.63 Ma. Node 133: Eudicotyledoneae: Ranunculales – [Vitales + Rosids + [Caryophyllales + Asterids]]; 119.6 Ma – 128.63 Ma. Fossil taxon and specimen: Hyrcantha decussata [NJU-DES02001: Geological Institute, Chinese Academy of Sciences, Beijing], from the lower part of the Yixian Formation, Jehol Group, Aptian (Early Cretaceous), from the Liaoning Province, China [246]. Phylogenetic justification: Similar to Leefrutcus from the Yixian Formation of the Lower Cretaceous of China, Hycantha is considered to be a stem group representative of the Ranunculales [247]. Minimum age: 119.6 Ma. Soft maximum age: 128.63 Ma. Age justification: The main fossil bearing beds have been dated and may be as old as 129.2 Ma [221], however, in the absence of knowledge of the position of the fossils within the stratigraphy, relative to the sources of the absolute dates, a minimum age constraint can be derived from the Jiufontang Formation which overlies it. 40Ar/39Ar dating of a number of samples from the Jiufontang Formation has yielded an age of 120.3 ± 0.7 Ma for the volcanic tuffs [222], establishing a minimum constraint of 119.6 Ma. The soft maximum age constraint is based on the maximum age of the oldest potential age of tricolpate pollen, from the middle Atherfield Wealden Bed 35 of the Cowleaze Chine Member, Vectis Formation [223]. Following Clarke at al. [62], the Cowleaze Chine Member of the Vectis Formation of the Isle of Wight [223] occurs within the M1n polarity chron in the Late Barremian. The base of the M1n chron is approximately correlated with the base of the Gerhardtia sartousi Biozone, dated at 128.63 Ma. Node 134: Vitales – [Rosids + [Caryophyllales + Asterids]]; 85.8 Ma – 128.63 Ma.

Fossil taxon and specimen: Paleoclusia chevalieri / Paleoenkianthus sayrevillensis [CUPC 1192 and CUPC 1100: L. H. Bailey Hortorium Paleobotanical Collection, Cornell University, Ithaca, NY, USA], Raritan – Lower Magothy Formations, Coniacian (Late Cretaceous), from the Old Crossman Pit locality, Sayreville New Jersey, USA [248, 249]. Phylogenetic justification: The phylogenetic interpretation follows Crepet et al. [249] and Ruhfels [250]. Minimum age: 85.8 Ma. Soft maximum age: 128.63 Ma. Age justification: Clarke et al. [62] argued that a minimum constraint on the age of this deposit could be established from Santonian-Campanian Boundary, however, Massoni et al. [232] argue that a tighter correlation can be established with better rocks attributable to the CC13-14 Nannofossil Biozones in South Carolina, indicating a minimum age of 86.3 Ma ± 0.5 Myr, thus, 85.8 Ma. The soft maximum age constraint is based on the maximum age of the oldest potential age of tricolpate pollen, from the middle Atherfield Wealden Bed 35 of the Cowleaze Chine Member, Vectis Formation [223]. Following Clarke at al. [62], the Cowleaze Chine Member of the Vectis Formation of the Isle of Wight [223] occurs within the M1n polarity chron in the Late Barremian. The base of the M1n chron is approximately correlated with the base of the Gerhardtia sartousi Biozone, dated at 128.63 Ma. Discussion: The oldest fossil representatives of both Rosids and Ericales (Asterids) are known from the Old Crossman Pit locality at Sayreville, New Jersey, hence both provide a minimum age for this node. Node 135: Rosids (minus Vitales) – [Caryophyllales + Asterids]; 85.8 Ma – 128.63 Ma. Fossil taxon and specimen: Paleoclusia chevalieri / Paleoenkianthus sayrevillensis [CUPC 1192 and CUPC 1100: L. H. Bailey Hortorium Paleobotanical Collection, Cornell University, Ithaca, NY, USA], Raritan – Lower Magothy Formations, Coniacian (Late Cretaceous), from the Old Crossman Pit locality, Sayreville New Jersey, USA [248, 249]. Phylogenetic justification: The phylogenetic interpretation follows Ruhfels [250]. Minimum age: 85.8 Ma. Soft maximum age: 128.63 Ma. Age justification: Clarke et al. [62] argued that a minimum constraint on the age of this deposit could be established from Santonian-Campanian Boundary, however, Massoni et al. [232] argue that a tighter correlation can be established with better rocks attributable to the CC13-14 Nannofossil Biozones in South Carolina, indicating a minimum age of 86.3 Ma ± 0.5 Myr, thus, 85.8 Ma. The soft maximum age constraint is based on the maximum age of the oldest potential age of tricolpate pollen, from the middle Atherfield Wealden Bed 35 of the Cowleaze Chine Member, Vectis Formation [223]. Following Clarke at al. [62], the Cowleaze Chine Member of the Vectis Formation of the Isle of Wight [223] occurs within the M1n polarity chron in the Late Barremian. The base of the M1n chron is approximately correlated with the base of the Gerhardtia sartousi Biozone, dated at 128.63 Ma. Node 142: Caryophyllales – Asterids; 85.8 Ma – 128.63 Ma. Fossil taxon and specimen: Paleoenkianthus sayrevillensis [CUPC 1100: L. H. Bailey Hortorium, Cornell University, Ithaca NY, USA] from the South Amboy Fire Clay of the Raritan – Lower Magothy Formations, Coniacian (Late Cretaceous), of which outcrops are exposed in the Old Crossman Clay Pit in Sayreville, New Jersey, USA [249]. Phylogenetic justification: The phylogenetic relationship of this fossil has been tested based on morphological evidence [249]. Minimum age: 85.8 Ma. Soft maximum age: 128.63 Ma. Age justification: Clarke et al. [62] argued that a minimum constraint on the age of this deposit could be established from Santonian-Campanian Boundary, however, Massoni et al. [232] argue that a tighter correlation can be established with better rocks attributable to the CC13-14 Nannofossil Biozones in South Carolina, indicating a minimum age of 86.3 Ma ± 0.5 Myr, thus, 85.8 Ma. The soft maximum age constraint is based on the maximum age of the oldest potential age of tricolpate pollen, from the middle Atherfield Wealden Bed 35 of the Cowleaze Chine Member, Vectis Formation [223]. Following Clarke at al. [62], the Cowleaze Chine Member of the Vectis Formation of the Isle of Wight [223] occurs within the M1n polarity chron in the Late Barremian. The base of the M1n chron is approximately correlated with the base of the Gerhardtia sartousi Biozone, dated at 128.63 Ma. Node 119: Monocotyledoneae: Acorales – [Dioscoreales + [Liliales + [Asparagales + Aracales + Poales]]]; 113 Ma – 128.63 Ma.

Fossil taxon and specimen: Liliacidites sp. A [palynological slide 71-8-1d], from the Potomac Group, Albian Zone I (Early Cretaceous), at the Trent’s Reach Locality, Virginia, U.S.A. [251-253]. Phylogenetic justification: Doyle et al. [251] identified pollen referred to the genus Liliacidites (but not Similipollis) as represesentative of the monocot stem, making it the oldest secure record of the monocot total group (see [236]). Minimum age: 113 Ma. Soft maximum age: 128.63 Ma. Age justification: In the absence of further stratigraphic constraint, these earliest records of Liliacidites can be constrained in age by the Aptian-Albian Boundary, dated to 113.0 Ma (International Chronostratigraphic Chart, v.2016/10). The soft maximum age constraint is based on the maximum age of the oldest potential age of tricolpate pollen, from the middle Atherfield Wealden Bed 35 of the Cowleaze Chine Member, Vectis Formation [223]. Following Clarke at al. [62], the Cowleaze Chine Member of the Vectis Formation of the Isle of Wight [223] occurs within the M1n polarity chron in the Late Barremian. The base of the M1n chron is approximately correlated with the base of the Gerhardtia sartousi Biozone, dated at 128.63 Ma. Discussion: Doyle et al. [252] highlight that, despite decades of sampling of the Hauterivian and Barremian of England, no clear representatives of Liliacidites pollen have been recovered [254], perhaps implying that the earliest records from the Albian are a close approximation of their antiquity. Because of the position of monocots in our molecular tree we consider Liliacidites to be nested within monocots, and use it to calibrate the monocot crown node. Node 120: Dioscoreales – [Liliales + Asparagales + Aracales + Poales]; 85.8 Ma – 128.63 Ma. Fossil taxon and specimen: Mabelia connatifila [CUPC 1255: L. H. Bailey Hortorium Paleobotanical Collection, Cornell University, Ithaca NY, USA] from the South Amboy Fire Clay Member of the Raritan – Lower Magothy Formations, Coniacian (Late Cretaceous), at the Old Crossman clay pit in Sayreville, New Jersey, USA [255]. Phylogenetic justification: The phylogenetic assignment is based on the phylogenetic hypothesis reconstructed by Gandolfo et al. [255]. Minimum age: 85.8 Ma. Soft maximum age: 128.63 Ma. Age justification: Clarke et al. [62] argued that a minimum constraint on the age of this deposit could be established from Santonian-Campanian Boundary, however, Massoni et al. [232] argue that a tighter correlation can be established with better rocks attributable to the CC13-14 Nannofossil zones in South Carolina, indicating a minimum age of 86.3 Ma ± 0.5 Myr, thus, 85.8 Ma The soft maximum age constraint is based on the maximum age of the oldest potential age of tricolpate pollen, from the middle Atherfield Wealden Bed 35 of the Cowleaze Chine Member, Vectis Formation [223]. Following Clarke at al. [62], the Cowleaze Chine Member of the Vectis Formation of the Isle of Wight [223] occurs within the M1n polarity chron in the Late Barremian. The base of the M1n chron is approximately correlated with the base of the Gerhardtia sartousi Biozone, dated at 128.63 Ma. Node 121: Liliales – [Asparagales + Aracales + Poales]; 83.41 – 128.63 Ma. Fossil taxon and specimen: Sabalites carolinensis [PAL 175717/P 38208: Smithsonian Museum of Natural History; Washington DC, USA] described from the Middendorf Arkose Member of Black Creek Formation near Langley, Aiken County, South Carolina [256]. Phylogenetic justification: The phylogenetic relationships of this fossil have discussed in Hertweck et al. [257] and Iles et al. [258]. Minimum age: 83.41 Ma. Soft maximum age: 128.63 Ma. Age justification: Berry’s view that the Middendorf was merely a distinct facies within the Black Creek Formation, rather than a stratigraphically distinct unit, has been rejected. Sohl and Owens [259] subdivided the Upper Cretaceous of Carolina coastal plain into three lithostratigraphic units, the Middendorf, Black Creek and Peedee Formations, raised the Black Creek to group status and subdivided this into three unconformity-bound formations, viz. in stratigraphic sequence, the Tar Heel, Bladen and Donoho Creek formations. Evidently, Sabalites carolinensis was recovered from what is now recognized as the Middendorf Formation, and a minimum age constraint can be established on the boundary between the Middendorf and Tar Heel Formations. The Middendorf is commonly considered Santonian in age, however, little material evidence has been presented in support of this, in part a consequent of the complex history of stratigraphic divisions at outcrop, in subsurface and offshore [260]. Habib and Miller [261] established an age ‘not younger than Campanian’ on the basis of dinoflagellate biostratigraphy, but following the stratigraphic scheme outlined Campbell and Grohn [260], the Middendorf is older that the Shepherd Grove Formation and, therefore, following the stratigraphy of Christopher and Prowell [262], must be no younger than Santonian. Thus, we may established a minimum age constraint

on the Sabalites carolinesis based on the Santonian-Campanian Boundary, coincident with the base of the Scaphites leei III Biozone, dated to 83.64 Ma ± 0.23 Myr [220], thus, 83.41 Ma. The soft maximum age constraint is based on the maximum age of the oldest potential age of tricolpate pollen, from the middle Atherfield Wealden Bed 35 of the Cowleaze Chine Member, Vectis Formation [223]. Following Clarke at al. [62], the Cowleaze Chine Member of the Vectis Formation of the Isle of Wight [223] occurs within the M1n polarity chron in the Late Barremian. The base of the M1n chron is approximately correlated with the base of the Gerhardtia sartousi Biozone, dated at 128.63 Ma.

2. Supplementary Figures
Supplementary Figure S1. The nodes to which age data from the fossil record were applied in analyses shown on the monophyletic and hornworts-sister topologies.
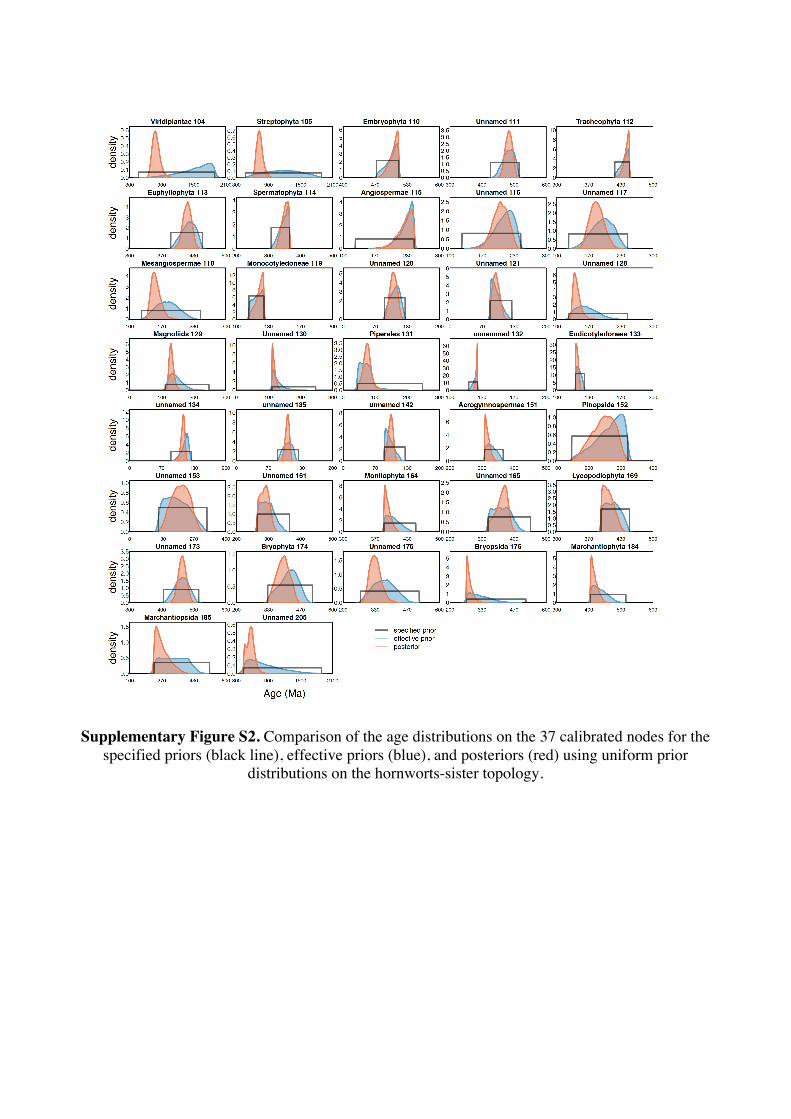
Supplementary Figure S2. Comparison of the age distributions on the 37 calibrated nodes for the specified priors (black line), effective priors (blue), and posteriors (red) using uniform prior
distributions on the hornworts-sister topology.

Supplementary Figure S3. Comparison of the age distributions on the 37 calibrated nodes for the specified priors (black line), effective priors (blue), and posteriors (red) using skew t prior
distributions on the hornworts-sister topology.
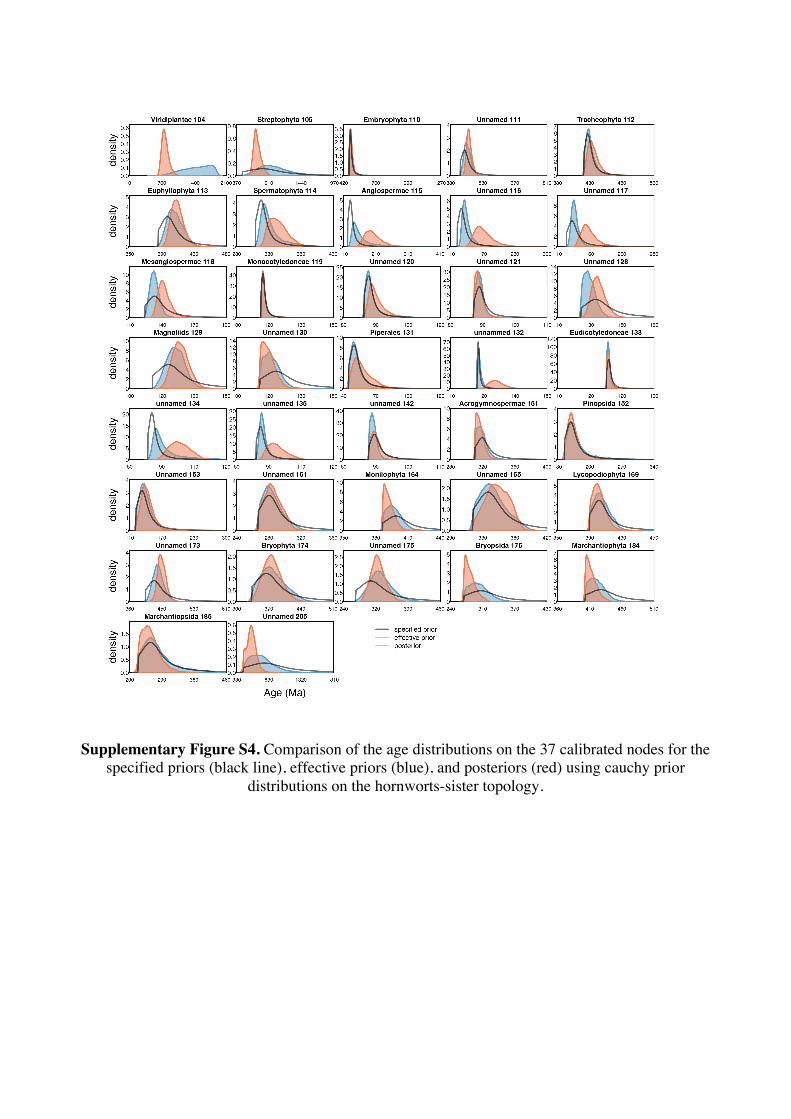
Supplementary Figure S4. Comparison of the age distributions on the 37 calibrated nodes for the
specified priors (black line), effective priors (blue), and posteriors (red) using cauchy prior distributions on the hornworts-sister topology.

Supplementary Figure S5. Comparison of ages using the monophyletic topology with different dataset sizes (number of nucleotides).
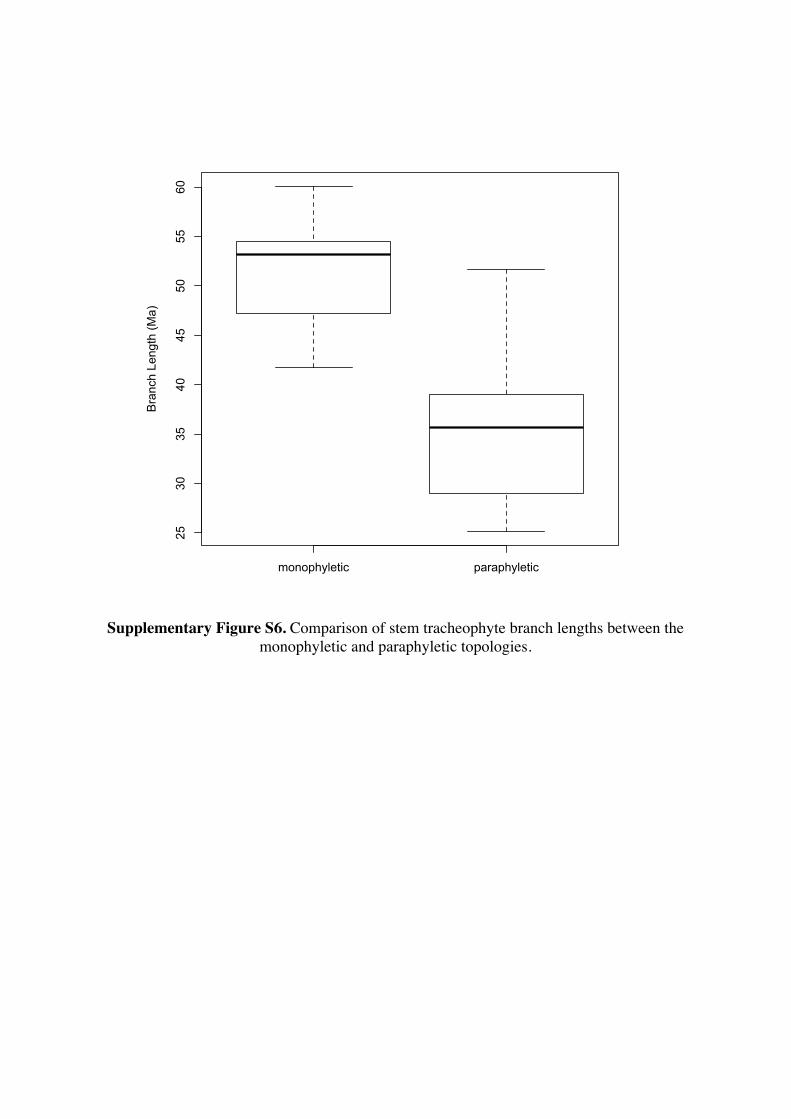
Supplementary Figure S6. Comparison of stem tracheophyte branch lengths between the
monophyletic and paraphyletic topologies.
monophyletic paraphyletic
2530
3540
4550
5560
Bra
nch
Leng
th (M
a)

Supplementary Figure S7. A comparison of the median and 95% HPD intervals for an analysis with all codon positions in a single partition (x axis) and each of the three codons partitioned into a
separate alignment (y axis). There are only minor and overlapping age estimates produced by the two analyses.
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●●
●●
●
●●
●
●●●
●●●●●
●
●●
●●●
●
●●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
0 2 4 6 8 10
0
2
4
6
8
10
single partition age (Ma)
parit
ition
for e
ach
codo
n ag
e (M
a)
y = 1.04x, R2 = 0.99

3. Supplementary Tables
Supplementary Table S1. Calibrations for monophyletic bryophytes topology (hypothesis i).
Node Clade Lineage 1 Lineage 2 Minimum providing fossil Min. date (Ma) Minimum age source Max. date (Ma) Soft maximum source 104 Viridiplantae Chlorophyta Streptophyta Tetrahedraletes cf. medinensis 469.0 Top of L. combazi Biozone 1823 ± 68 Igneous and metamorphic rocks
underlying deposits containing the earliest record of eukaryotes
205 Unnamed Chlorophyceae Prasinophyaceae Palaeocymopolia silurica 439.21 ± 0.9031 Top of P. leptotheca Biozone 105 Streptophyta Charophyta Embryophyta Tetrahedraletes cf. medinensis 469.0 Top of the L. combazi Biozone 110 Embryophyta Bryophytes Tracheophyta 515.5 Oldest cryptospores; base of the
Olenellus Biozone 172 Bryophytes Anthocerotophyta Marchantiophyta + Bryophyta Riccardiothallus devonicus 407.6 ± 2.6 Top of the Pragian Stage 173 Unnamed Marchantiophyta Bryophyta 184 Marchantiophyta Jungermanniopsida Marchantiopsida 185 Marchantiopsida Sphaerocarpales Marchantiales Marchantites cyathodoides 227.0 Top of the Carnian Stage 174 Bryophyta Sphagnopsida Polytrichopsida + Bryopsida Sphagnales 330.9 ± 0.2 Top of the VF Biozone 175 Unnamed Polytrichopsida Bryopsida Palaeocampylopus buragoase 272.3 ± 0.5 Top of the Kungurian Stage 176 Bryopsida Funariidae Dicranidae + Bryidae Capimirinus riopretensis 268.8 ± 0.5 Top of the Roadian Stage 111 Tracheophyta Lycopodiophyta Euphyllophyta Zosterophyllum sp. 423.0 ± 2.3 Top of the Ludlow Epoch 451 Trilete spores; base of the
morrisi sub-biozone of Dicranograptus clingani Biozone
168 Lycopodiophyta Isoetopsida Lycopodiopsida Leclercqia complexa 393.3 ± 1.2 Top of the Emsian Stage 112 Euphyllophyta Monilophyta Spermatophyta Rellimia thomsonii 386.25 ± 0.679 rhenanus-ansatus boundary 163 Monilophyta Equisetopsida Polypodiopsida + [Psilotopsida +
Marattiopsida] Ibyka amphikoma 385.41 ± 0.704 Top of the P. ansatus Biozone
164 Unnamed Psilotopsida + Marattiopsida Polypodiopsida Psaronius simplicicaulis 319.09 ± 0.4 Base of the Pc5 Biozone 113 Spermatophyta Acrogymnospermae Angiospermae Cordaixylon iowensis 308.5 ± 0.36 Top of the N. medexultimus -S.
concinnus (Pc10) Biozone 364.19 ± 1.439 First seeds; base of P.
trachytera Biozone 150 Acrogymnospermae Pinopsida Ginkgoopsida + Cycadopsida 151 Unnamed Ginkgoopsida Cycadopsida Crossozamia chinensis 265.1 ± 0.4 Top of the Wordian Stage 154 Pinopsida Cupressophyta Pinaceae + Gnetales Araucaria miribilis 157.0 ± 10 Igneous rocks overlying oldest
Araucaria fossils 321.01 ± 0.37 Cordaixylon iowensis bearing
strata; maximum age of Composite Level below conodont Pc10 Biozone
155 Unnamed Pinaceae Gnetales Eoantha zherikhinii 120.3 ± 0.7 Igneous rocks above oldest Eoantha zherikhinii
114 Angiospermae Amborellales Nymphaeales + [Austrobaileyales + Mesangiospermae]
Tricolpate pollen 125 Top of the Mn1 polarity chron / Barremian Stage
247.2 Sediments below the oldest occurrence of angiosperm-like pollen; base of the Anisian Stage
115 Unnamed Nymphaeales Austrobaileyales + Mesangiospermae 116 Unnamed Austrobaileyales Mesangiospermae 117 Mesangiospermae Magnoliids + [Chloranthales +
Eudicotyledoneae] Monocotyledoneae
127 Unnamed Magnoliids Chloranthales + Eudicotyledoneae 147 Magnoliids Magnoliales + Laurales Piperales Endressinia brasiliana 110..87 Early - Middle Albian
Boundary; base of D. mammillatum Biozone
148 Unnamed Persea Liriodendron
149 Piperales Saururus Houttuynia Saururus tuckerae 46.2 ± 1.9 Youngest radiometric date estimate for Allenby Formation
128 Unnamed Chloranthales Eudicotyledoneae Hyrcantha decussata 120.3 ± 0.7 Radiometric date from the overlying Jiufontang Formation
128.63 Oldest potential age of tricolpate pollen; base of the M1n chron/ G. sartousi Biozone
129 Eudicotyledoneae Ranunculales Vitales + Rosids + [Caryophyllales + Asterids]
130 Unnamed Vitales Rosids + [Caryophyllales + Asterids] Paleoclusia chevalieri / Paleoenkianthus sayrevillensis
86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage 131 Unnamed Rosids (minus Vitales) Caryophyllales + Asterids
138 Unnamed Caryophyllales Asterids Paleoenkianthus sayrevillensis 118 Monocotyledoneae Acorales [Dioscoreales + Liliales +Asparagales
+ Aracales + Poales] Liliacidites sp. A 113 Top of the Aptian Stage
119 Unnamed Dioscoreales Liliales + Asparagales + Aracales + Poales
Mabelia connatifila 86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage
120 Unnamed Liliales Asparagales + Aracales + Poales Sabalites carolinensis 83.64 ± 0.23 Top of the Santonian Stage / S. leei III Biozone
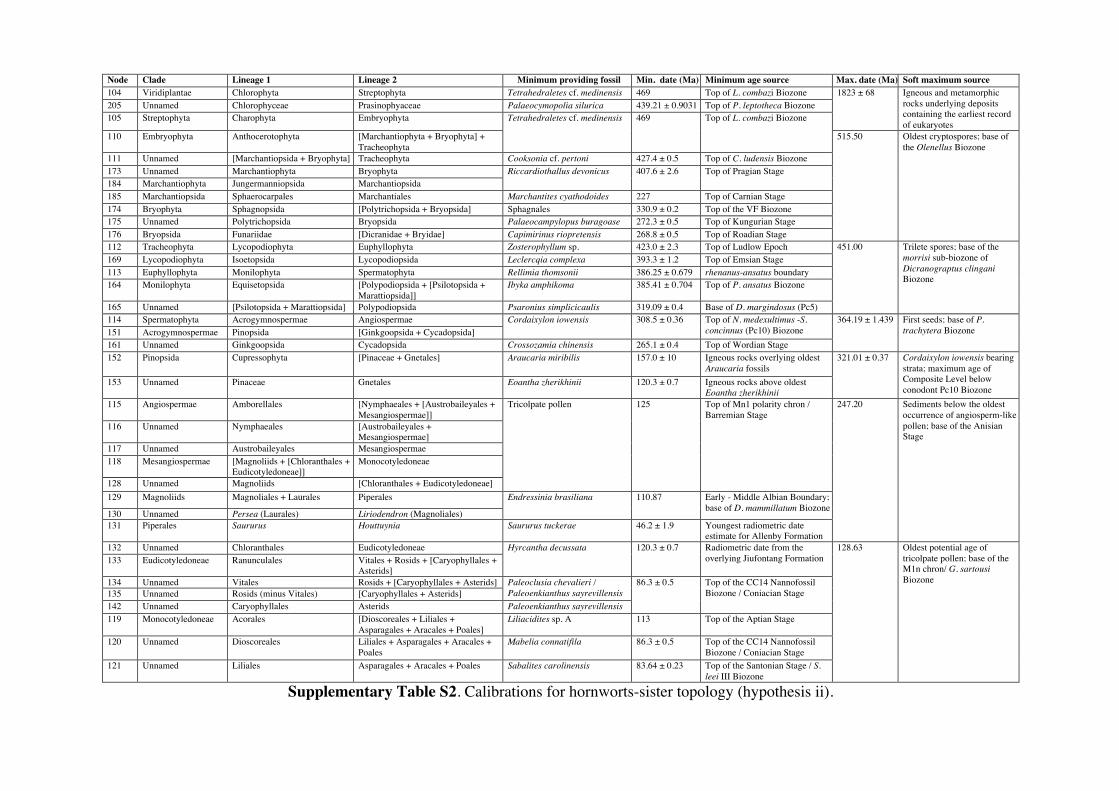
Node Clade Lineage 1 Lineage 2 Minimum providing fossil Min. date (Ma) Minimum age source Max. date (Ma) Soft maximum source 104 Viridiplantae Chlorophyta Streptophyta Tetrahedraletes cf. medinensis 469 Top of L. combazi Biozone 1823 ± 68 Igneous and metamorphic
rocks underlying deposits containing the earliest record of eukaryotes
205 Unnamed Chlorophyceae Prasinophyaceae Palaeocymopolia silurica 439.21 ± 0.9031 Top of P. leptotheca Biozone 105 Streptophyta Charophyta Embryophyta Tetrahedraletes cf. medinensis 469 Top of L. combazi Biozone
110 Embryophyta Anthocerotophyta [Marchantiophyta + Bryophyta] + Tracheophyta
515.50 Oldest cryptospores; base of the Olenellus Biozone
111 Unnamed [Marchantiopsida + Bryophyta] Tracheophyta Cooksonia cf. pertoni 427.4 ± 0.5 Top of C. ludensis Biozone 173 Unnamed Marchantiophyta Bryophyta Riccardiothallus devonicus 407.6 ± 2.6 Top of Pragian Stage 184 Marchantiophyta Jungermanniopsida Marchantiopsida 185 Marchantiopsida Sphaerocarpales Marchantiales Marchantites cyathodoides 227 Top of Carnian Stage 174 Bryophyta Sphagnopsida [Polytrichopsida + Bryopsida] Sphagnales 330.9 ± 0.2 Top of the VF Biozone 175 Unnamed Polytrichopsida Bryopsida Palaeocampylopus buragoase 272.3 ± 0.5 Top of Kungurian Stage 176 Bryopsida Funariidae [Dicranidae + Bryidae] Capimirinus riopretensis 268.8 ± 0.5 Top of Roadian Stage 112 Tracheophyta Lycopodiophyta Euphyllophyta Zosterophyllum sp. 423.0 ± 2.3 Top of Ludlow Epoch 451.00 Trilete spores; base of the
morrisi sub-biozone of Dicranograptus clingani Biozone
169 Lycopodiophyta Isoetopsida Lycopodiopsida Leclercqia complexa 393.3 ± 1.2 Top of Emsian Stage 113 Euphyllophyta Monilophyta Spermatophyta Rellimia thomsonii 386.25 ± 0.679 rhenanus-ansatus boundary 164 Monilophyta Equisetopsida [Polypodiopsida + [Psilotopsida +
Marattiopsida]] Ibyka amphikoma 385.41 ± 0.704 Top of P. ansatus Biozone
165 Unnamed [Psilotopsida + Marattiopsida] Polypodiopsida Psaronius simplicicaulis 319.09 ± 0.4 Base of D. margindosus (Pc5) 114 Spermatophyta Acrogymnospermae Angiospermae Cordaixylon iowensis 308.5 ± 0.36 Top of N. medexultimus -S.
concinnus (Pc10) Biozone 364.19 ± 1.439 First seeds; base of P.
trachytera Biozone 151 Acrogymnospermae Pinopsida [Ginkgoopsida + Cycadopsida] 161 Unnamed Ginkgoopsida Cycadopsida Crossozamia chinensis 265.1 ± 0.4 Top of Wordian Stage 152 Pinopsida Cupressophyta [Pinaceae + Gnetales] Araucaria miribilis 157.0 ± 10 Igneous rocks overlying oldest
Araucaria fossils 321.01 ± 0.37 Cordaixylon iowensis bearing
strata; maximum age of Composite Level below conodont Pc10 Biozone
153 Unnamed Pinaceae Gnetales Eoantha zherikhinii 120.3 ± 0.7 Igneous rocks above oldest Eoantha zherikhinii
115 Angiospermae Amborellales [Nymphaeales + [Austrobaileyales + Mesangiospermae]]
Tricolpate pollen 125 Top of Mn1 polarity chron / Barremian Stage
247.20 Sediments below the oldest occurrence of angiosperm-like pollen; base of the Anisian Stage
116 Unnamed Nymphaeales [Austrobaileyales + Mesangiospermae]
117 Unnamed Austrobaileyales Mesangiospermae 118 Mesangiospermae [Magnoliids + [Chloranthales +
Eudicotyledoneae]] Monocotyledoneae
128 Unnamed Magnoliids [Chloranthales + Eudicotyledoneae] 129 Magnoliids Magnoliales + Laurales Piperales Endressinia brasiliana 110.87 Early - Middle Albian Boundary;
base of D. mammillatum Biozone 130 Unnamed Persea (Laurales) Liriodendron (Magnoliales) 131 Piperales Saururus Houttuynia Saururus tuckerae 46.2 ± 1.9 Youngest radiometric date
estimate for Allenby Formation 132 Unnamed Chloranthales Eudicotyledoneae Hyrcantha decussata 120.3 ± 0.7 Radiometric date from the
overlying Jiufontang Formation 128.63 Oldest potential age of
tricolpate pollen; base of the M1n chron/ G. sartousi Biozone
133 Eudicotyledoneae Ranunculales Vitales + Rosids + [Caryophyllales + Asterids]
134 Unnamed Vitales Rosids + [Caryophyllales + Asterids] Paleoclusia chevalieri / Paleoenkianthus sayrevillensis
86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage 135 Unnamed Rosids (minus Vitales) [Caryophyllales + Asterids]
142 Unnamed Caryophyllales Asterids Paleoenkianthus sayrevillensis 119 Monocotyledoneae Acorales [Dioscoreales + Liliales +
Asparagales + Aracales + Poales] Liliacidites sp. A 113 Top of the Aptian Stage
120 Unnamed Dioscoreales Liliales + Asparagales + Aracales + Poales
Mabelia connatifila 86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage
121 Unnamed Liliales Asparagales + Aracales + Poales Sabalites carolinensis 83.64 ± 0.23 Top of the Santonian Stage / S. leei III Biozone
Supplementary Table S2. Calibrations for hornworts-sister topology (hypothesis ii).
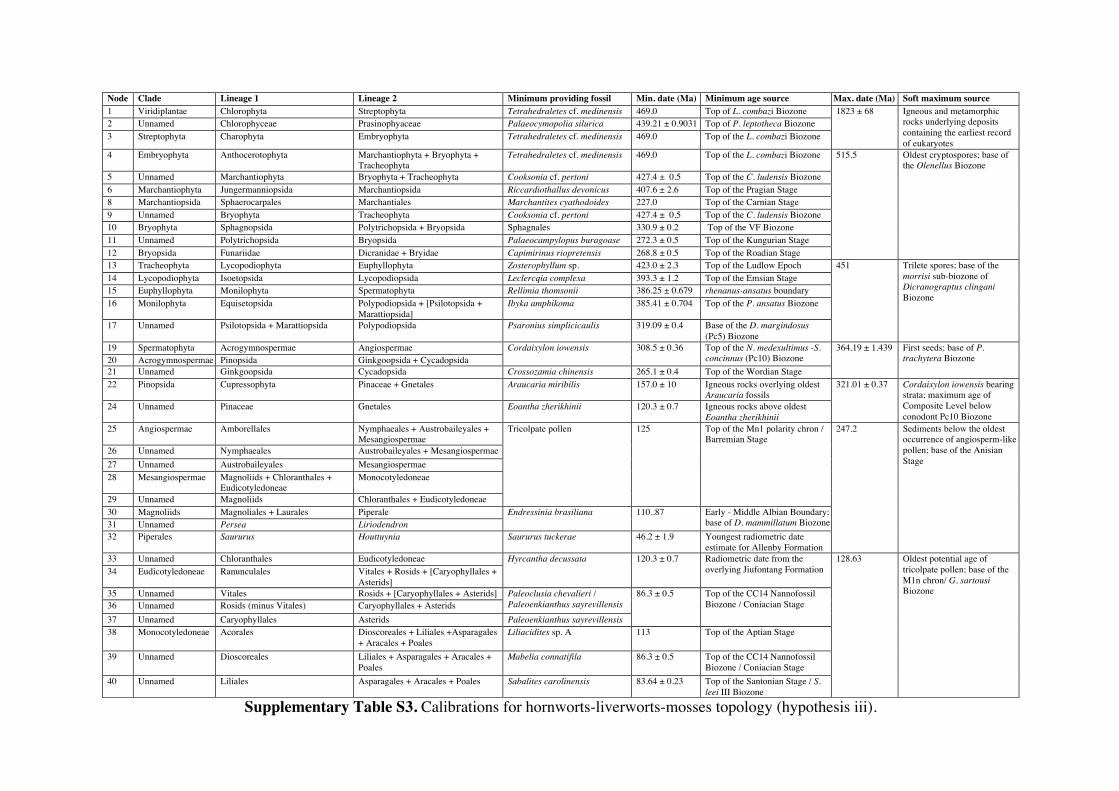
Node Clade Lineage 1 Lineage 2 Minimum providing fossil Min. date (Ma) Minimum age source Max. date (Ma) Soft maximum source 1 Viridiplantae Chlorophyta Streptophyta Tetrahedraletes cf. medinensis 469.0 Top of L. combazi Biozone 1823 ± 68 Igneous and metamorphic
rocks underlying deposits containing the earliest record of eukaryotes
2 Unnamed Chlorophyceae Prasinophyaceae Palaeocymopolia silurica 439.21 ± 0.9031 Top of P. leptotheca Biozone 3 Streptophyta Charophyta Embryophyta Tetrahedraletes cf. medinensis 469.0 Top of the L. combazi Biozone
4 Embryophyta Anthocerotophyta Marchantiophyta + Bryophyta + Tracheophyta
Tetrahedraletes cf. medinensis 469.0 Top of the L. combazi Biozone 515.5 Oldest cryptospores; base of the Olenellus Biozone
5 Unnamed Marchantiophyta Bryophyta + Tracheophyta Cooksonia cf. pertoni 427.4 ± 0.5 Top of the C. ludensis Biozone 6 Marchantiophyta Jungermanniopsida Marchantiopsida Riccardiothallus devonicus 407.6 ± 2.6 Top of the Pragian Stage 8 Marchantiopsida Sphaerocarpales Marchantiales Marchantites cyathodoides 227.0 Top of the Carnian Stage 9 Unnamed Bryophyta Tracheophyta Cooksonia cf. pertoni 427.4 ± 0.5 Top of the C. ludensis Biozone 10 Bryophyta Sphagnopsida Polytrichopsida + Bryopsida Sphagnales 330.9 ± 0.2 Top of the VF Biozone 11 Unnamed Polytrichopsida Bryopsida Palaeocampylopus buragoase 272.3 ± 0.5 Top of the Kungurian Stage 12 Bryopsida Funariidae Dicranidae + Bryidae Capimirinus riopretensis 268.8 ± 0.5 Top of the Roadian Stage 13 Tracheophyta Lycopodiophyta Euphyllophyta Zosterophyllum sp. 423.0 ± 2.3 Top of the Ludlow Epoch 451 Trilete spores; base of the
morrisi sub-biozone of Dicranograptus clingani Biozone
14 Lycopodiophyta Isoetopsida Lycopodiopsida Leclercqia complexa 393.3 ± 1.2 Top of the Emsian Stage 15 Euphyllophyta Monilophyta Spermatophyta Rellimia thomsonii 386.25 ± 0.679 rhenanus-ansatus boundary 16 Monilophyta Equisetopsida Polypodiopsida + [Psilotopsida +
Marattiopsida] Ibyka amphikoma 385.41 ± 0.704 Top of the P. ansatus Biozone
17 Unnamed Psilotopsida + Marattiopsida Polypodiopsida Psaronius simplicicaulis 319.09 ± 0.4 Base of the D. margindosus (Pc5) Biozone
19 Spermatophyta Acrogymnospermae Angiospermae Cordaixylon iowensis 308.5 ± 0.36 Top of the N. medexultimus -S. concinnus (Pc10) Biozone
364.19 ± 1.439 First seeds; base of P. trachytera Biozone 20 Acrogymnospermae Pinopsida Ginkgoopsida + Cycadopsida
21 Unnamed Ginkgoopsida Cycadopsida Crossozamia chinensis 265.1 ± 0.4 Top of the Wordian Stage 22 Pinopsida Cupressophyta Pinaceae + Gnetales Araucaria miribilis 157.0 ± 10 Igneous rocks overlying oldest
Araucaria fossils 321.01 ± 0.37 Cordaixylon iowensis bearing
strata; maximum age of Composite Level below conodont Pc10 Biozone
24 Unnamed Pinaceae Gnetales Eoantha zherikhinii 120.3 ± 0.7 Igneous rocks above oldest Eoantha zherikhinii
25 Angiospermae Amborellales Nymphaeales + Austrobaileyales + Mesangiospermae
Tricolpate pollen 125 Top of the Mn1 polarity chron / Barremian Stage
247.2 Sediments below the oldest occurrence of angiosperm-like pollen; base of the Anisian Stage
26 Unnamed Nymphaeales Austrobaileyales + Mesangiospermae 27 Unnamed Austrobaileyales Mesangiospermae 28 Mesangiospermae Magnoliids + Chloranthales +
Eudicotyledoneae Monocotyledoneae
29 Unnamed Magnoliids Chloranthales + Eudicotyledoneae 30 Magnoliids Magnoliales + Laurales Piperale Endressinia brasiliana 110..87 Early - Middle Albian Boundary;
base of D. mammillatum Biozone 31 Unnamed Persea Liriodendron 32 Piperales Saururus Houttuynia Saururus tuckerae 46.2 ± 1.9 Youngest radiometric date
estimate for Allenby Formation 33 Unnamed Chloranthales Eudicotyledoneae Hyrcantha decussata 120.3 ± 0.7 Radiometric date from the
overlying Jiufontang Formation 128.63 Oldest potential age of
tricolpate pollen; base of the M1n chron/ G. sartousi Biozone
34 Eudicotyledoneae Ranunculales Vitales + Rosids + [Caryophyllales + Asterids]
35 Unnamed Vitales Rosids + [Caryophyllales + Asterids] Paleoclusia chevalieri / Paleoenkianthus sayrevillensis
86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage 36 Unnamed Rosids (minus Vitales) Caryophyllales + Asterids
37 Unnamed Caryophyllales Asterids Paleoenkianthus sayrevillensis 38 Monocotyledoneae Acorales Dioscoreales + Liliales +Asparagales
+ Aracales + Poales Liliacidites sp. A 113 Top of the Aptian Stage
39 Unnamed Dioscoreales Liliales + Asparagales + Aracales + Poales
Mabelia connatifila 86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage
40 Unnamed Liliales Asparagales + Aracales + Poales Sabalites carolinensis 83.64 ± 0.23 Top of the Santonian Stage / S. leei III Biozone
Supplementary Table S3. Calibrations for hornworts-liverworts-mosses topology (hypothesis iii).

Node Clade Lineage 1 Lineage 2 Minimum providing fossil Min. date (Ma) Minimum age source Max. date (Ma) Soft maximum source 1 Viridiplantae Chlorophyta Streptophyta Tetrahedraletes cf. medinensis 469.0 Top of the L. combazi Biozone 1823 ± 68 Igneous and metamorphic
rocks underlying deposits containing the earliest record of eukaryotes
2 Unnamed Chlorophyceae Prasinophyaceae Palaeocymopolia silurica 439.21 ± 0.9031 Top of P. leptotheca Biozone 3 Streptophyta Charophyta Embryophyta Tetrahedraletes cf. medinensis 469.0 Top of the L. combazi Biozone
4 Embryophyta Marchantiophyta + Bryophyta Anthocerotophyta + Tracheophyta 515.5 Oldest cryptospores; base of the Olenellus Biozone 5 Unnamed Marchantiophyta Bryophyta Riccardiothallus devonicus 407.6 ± 2.6 Top of the Pragian Stage
6 Marchantiophyta Jungermanniopsida Marchantiopsida 8 Marchantiopsida Sphaerocarpales Marchantiales Marchantites cyathodoides 227.0 Top of the Carnian Stage 9 Bryophyta Sphagnopsida Polytrichopsida + Bryopsida Sphagnales 330.9 ± 0.2 Top of the VF Biozone 10 Unnamed Polytrichopsida Bryopsida Palaeocampylopus buragoase 272.3 ± 0.5 Top of the Kungurian Stage 11 Bryopsida Funariidae Dicranidae + Bryidae Capimirinus riopretensis 268.8 ± 0.5 Top of the Roadian Stage 12 Unnamed Anthocerotophyta Tracheophyta Cooksonia cf. pertoni 427.4 ± 0.5 Top of the C. ludensis Biozone 13 Tracheophyta Lycopodiophyta Euphyllophyta Zosterophyllum sp. 423.0 ± 2.3 Top of the Ludlow Epoch 451 Trilete spores; base of the
morrisi sub-biozone of Dicranograptus clingani Biozone
14 Lycopodiophyta Isoetopsida Lycopodiopsida Leclercqia complexa 393.3 ± 1.2 Top of the Emsian Stage 15 Euphyllophyta Monilophyta Spermatophyta Rellimia thomsonii 386.25 ± 0.679 rhenanus-ansatus boundary 16 Monilophyta Equisetopsida Polypodiopsida + Psilotopsida +
Marattiopsida Ibyka amphikoma 385.41 ± 0.704 Top of the P. ansatus Biozone
17 Unnamed Psilotopsida + Marattiopsida Polypodiopsida Psaronius simplicicaulis 319.09 ± 0.4 Base of the D. margindosus (Pc5) Biozone
19 Spermatophyta Acrogymnospermae Angiospermae Cordaixylon iowensis 308.5 ± 0.36 Top of the N. medexultimus -S. concinnus (Pc10) Biozone
364.19 ± 1.439 First seeds; base of P. trachytera Biozone 20 Acrogymnospermae Pinopsida Ginkgoopsida + Cycadopsida
21 Unnamed Ginkgoopsida Cycadopsida Crossozamia chinensis 265.1 ± 0.4 Top of the Wordian Stage 22 Pinopsida Cupressophyta Pinaceae + Gnetales Araucaria miribilis 157.0 ± 10 Igneous rocks overlying oldest
Araucaria fossils 321.01 ± 0.37 Cordaixylon iowensis bearing
strata; maximum age of Composite Level below conodont Pc10 Biozone
24 Unnamed Pinaceae Gnetales Eoantha zherikhinii 120.3 ± 0.7 Igneous rocks above oldest Eoantha zherikhinii
25 Angiospermae Amborellales Nymphaeales + Austrobaileyales + Mesangiospermae
Tricolpate pollen 125 Top of the Mn1 polarity chron / Barremian Stage
247.2 Sediments below the oldest occurrence of angiosperm-like pollen; base of the Anisian Stage
26 Unnamed Nymphaeales Austrobaileyales + Mesangiospermae 27 Unnamed Austrobaileyales Mesangiospermae 28 Mesangiospermae Magnoliids + Chloranthales +
Eudicotyledoneae Monocotyledoneae
29 Unnamed Magnoliids Chloranthales + Eudicotyledoneae 30 Magnoliids Magnoliales + Laurales Piperales Endressinia brasiliana 110..87 Early - Middle Albian Boundary;
base of D. mammillatum Biozone
31 Unnamed Persea Liriodendron
32 Piperales Saururus Houttuynia Saururus tuckerae 46.2 ± 1.9 Youngest radiometric date estimate for Allenby Formation
33 Unnamed Chloranthales Eudicotyledoneae Hyrcantha decussata 120.3 ± 0.7 Radiometric date from the overlying Jiufontang Formation
128.63 Oldest potential age of tricolpate pollen; base of the M1n chron/ G. sartousi Biozone
34 Eudicotyledoneae Ranunculales Vitales + Rosids + [Caryophyllales + Asterids]
35 Unnamed Vitales Rosids + [Caryophyllales + Asterids] Paleoclusia chevalieri / Paleoenkianthus sayrevillensis
86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage 36 Unnamed Rosids (minus Vitales) Caryophyllales + Asterids
37 Unnamed Caryophyllales Asterids Paleoenkianthus sayrevillensis 38 Monocotyledoneae Acorales Dioscoreales + Liliales +Asparagales
+ Aracales + Poales Liliacidites sp. A 113 Top of the Aptian Stage
39 Unnamed Dioscoreales Liliales + Asparagales + Aracales + Poales
Mabelia connatifila 86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage
40 Unnamed Liliales Asparagales + Aracales + Poales Sabalites carolinensis 83.64 ± 0.23 Top of the Santonian Stage / S. leei III Biozone
Supplementary Table S4. Calibrations for liverworts-mosses topology (hypothesis iv).
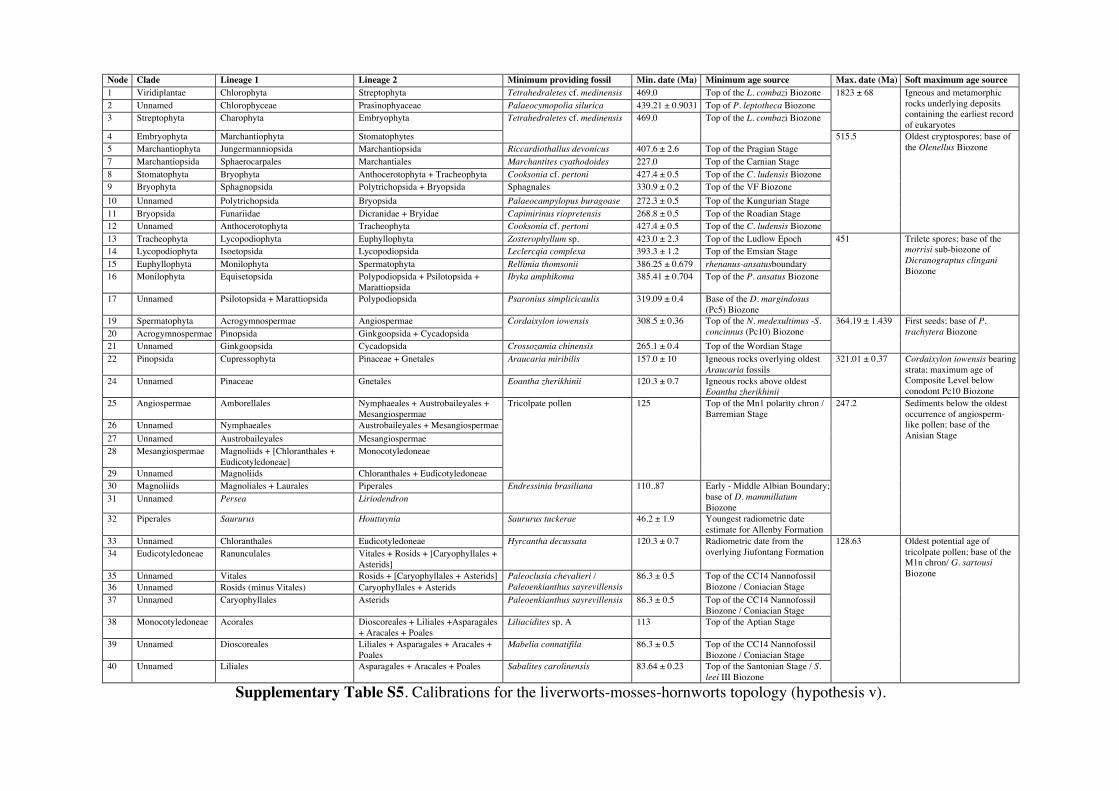
Node Clade Lineage 1 Lineage 2 Minimum providing fossil Min. date (Ma) Minimum age source Max. date (Ma) Soft maximum age source 1 Viridiplantae Chlorophyta Streptophyta Tetrahedraletes cf. medinensis 469.0 Top of the L. combazi Biozone 1823 ± 68 Igneous and metamorphic
rocks underlying deposits containing the earliest record of eukaryotes
2 Unnamed Chlorophyceae Prasinophyaceae Palaeocymopolia silurica 439.21 ± 0.9031 Top of P. leptotheca Biozone 3 Streptophyta Charophyta Embryophyta Tetrahedraletes cf. medinensis 469.0 Top of the L. combazi Biozone
4 Embryophyta Marchantiophyta Stomatophytes 515.5 Oldest cryptospores; base of the Olenellus Biozone 5 Marchantiophyta Jungermanniopsida Marchantiopsida Riccardiothallus devonicus 407.6 ± 2.6 Top of the Pragian Stage
7 Marchantiopsida Sphaerocarpales Marchantiales Marchantites cyathodoides 227.0 Top of the Carnian Stage 8 Stomatophyta Bryophyta Anthocerotophyta + Tracheophyta Cooksonia cf. pertoni 427.4 ± 0.5 Top of the C. ludensis Biozone 9 Bryophyta Sphagnopsida Polytrichopsida + Bryopsida Sphagnales 330.9 ± 0.2 Top of the VF Biozone 10 Unnamed Polytrichopsida Bryopsida Palaeocampylopus buragoase 272.3 ± 0.5 Top of the Kungurian Stage 11 Bryopsida Funariidae Dicranidae + Bryidae Capimirinus riopretensis 268.8 ± 0.5 Top of the Roadian Stage 12 Unnamed Anthocerotophyta Tracheophyta Cooksonia cf. pertoni 427.4 ± 0.5 Top of the C. ludensis Biozone 13 Tracheophyta Lycopodiophyta Euphyllophyta Zosterophyllum sp. 423.0 ± 2.3 Top of the Ludlow Epoch 451 Trilete spores; base of the
morrisi sub-biozone of Dicranograptus clingani Biozone
14 Lycopodiophyta Isoetopsida Lycopodiopsida Leclercqia complexa 393.3 ± 1.2 Top of the Emsian Stage 15 Euphyllophyta Monilophyta Spermatophyta Rellimia thomsonii 386.25 ± 0.679 rhenanus-ansatusboundary 16 Monilophyta Equisetopsida Polypodiopsida + Psilotopsida +
Marattiopsida Ibyka amphikoma 385.41 ± 0.704 Top of the P. ansatus Biozone
17 Unnamed Psilotopsida + Marattiopsida Polypodiopsida Psaronius simplicicaulis 319.09 ± 0.4 Base of the D. margindosus (Pc5) Biozone
19 Spermatophyta Acrogymnospermae Angiospermae Cordaixylon iowensis 308.5 ± 0.36 Top of the N. medexultimus -S. concinnus (Pc10) Biozone
364.19 ± 1.439 First seeds; base of P. trachytera Biozone 20 Acrogymnospermae Pinopsida Ginkgoopsida + Cycadopsida
21 Unnamed Ginkgoopsida Cycadopsida Crossozamia chinensis 265.1 ± 0.4 Top of the Wordian Stage 22 Pinopsida Cupressophyta Pinaceae + Gnetales Araucaria miribilis 157.0 ± 10 Igneous rocks overlying oldest
Araucaria fossils 321.01 ± 0.37 Cordaixylon iowensis bearing
strata; maximum age of Composite Level below conodont Pc10 Biozone
24 Unnamed Pinaceae Gnetales Eoantha zherikhinii 120.3 ± 0.7 Igneous rocks above oldest Eoantha zherikhinii
25 Angiospermae Amborellales Nymphaeales + Austrobaileyales + Mesangiospermae
Tricolpate pollen 125 Top of the Mn1 polarity chron / Barremian Stage
247.2 Sediments below the oldest occurrence of angiosperm-like pollen; base of the Anisian Stage
26 Unnamed Nymphaeales Austrobaileyales + Mesangiospermae 27 Unnamed Austrobaileyales Mesangiospermae 28 Mesangiospermae Magnoliids + [Chloranthales +
Eudicotyledoneae] Monocotyledoneae
29 Unnamed Magnoliids Chloranthales + Eudicotyledoneae 30 Magnoliids Magnoliales + Laurales Piperales Endressinia brasiliana 110..87 Early - Middle Albian Boundary;
base of D. mammillatum Biozone
31 Unnamed Persea Liriodendron
32 Piperales Saururus Houttuynia Saururus tuckerae 46.2 ± 1.9 Youngest radiometric date estimate for Allenby Formation
33 Unnamed Chloranthales Eudicotyledoneae Hyrcantha decussata 120.3 ± 0.7 Radiometric date from the overlying Jiufontang Formation
128.63 Oldest potential age of tricolpate pollen; base of the M1n chron/ G. sartousi Biozone
34 Eudicotyledoneae Ranunculales Vitales + Rosids + [Caryophyllales + Asterids]
35 Unnamed Vitales Rosids + [Caryophyllales + Asterids] Paleoclusia chevalieri / Paleoenkianthus sayrevillensis
86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage 36 Unnamed Rosids (minus Vitales) Caryophyllales + Asterids
37 Unnamed Caryophyllales Asterids Paleoenkianthus sayrevillensis 86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage
38 Monocotyledoneae Acorales Dioscoreales + Liliales +Asparagales + Aracales + Poales
Liliacidites sp. A 113 Top of the Aptian Stage
39 Unnamed Dioscoreales Liliales + Asparagales + Aracales + Poales
Mabelia connatifila 86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage
40 Unnamed Liliales Asparagales + Aracales + Poales Sabalites carolinensis 83.64 ± 0.23 Top of the Santonian Stage / S. leei III Biozone
Supplementary Table S5. Calibrations for the liverworts-mosses-hornworts topology (hypothesis v).
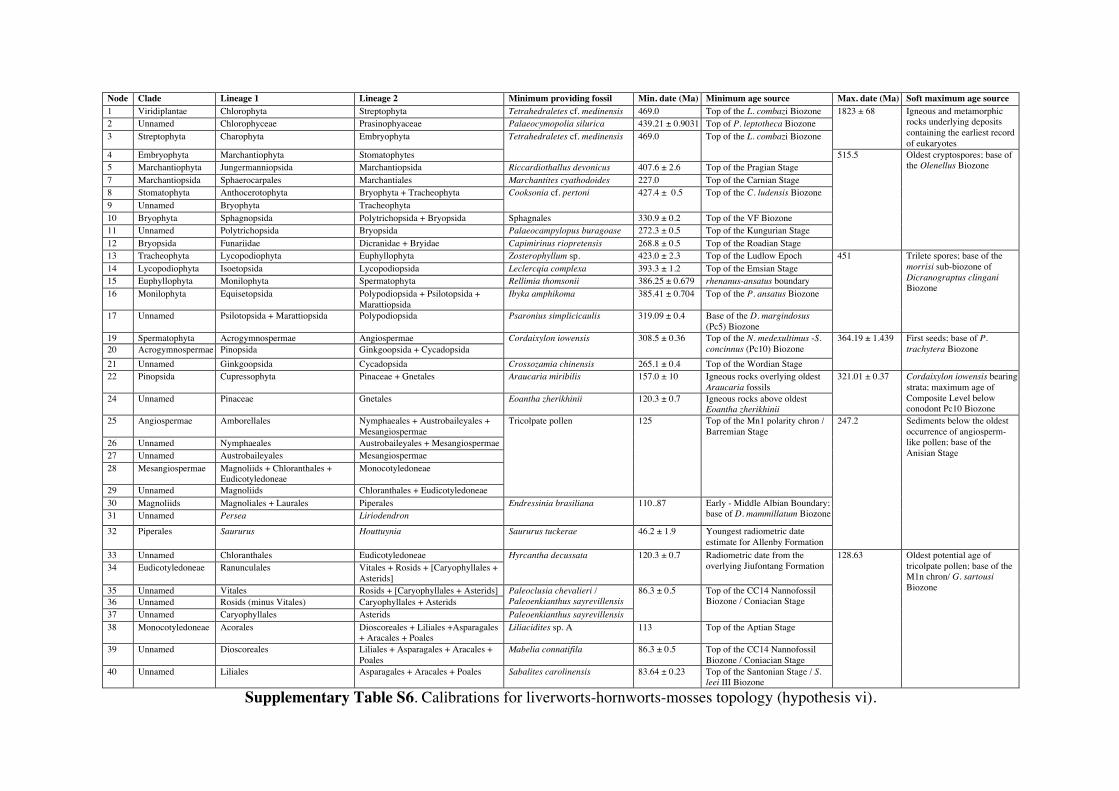
Node Clade Lineage 1 Lineage 2 Minimum providing fossil Min. date (Ma) Minimum age source Max. date (Ma) Soft maximum age source 1 Viridiplantae Chlorophyta Streptophyta Tetrahedraletes cf. medinensis 469.0 Top of the L. combazi Biozone 1823 ± 68 Igneous and metamorphic
rocks underlying deposits containing the earliest record of eukaryotes
2 Unnamed Chlorophyceae Prasinophyaceae Palaeocymopolia silurica 439.21 ± 0.9031 Top of P. leptotheca Biozone 3 Streptophyta Charophyta Embryophyta Tetrahedraletes cf. medinensis 469.0 Top of the L. combazi Biozone
4 Embryophyta Marchantiophyta Stomatophytes 515.5 Oldest cryptospores; base of the Olenellus Biozone 5 Marchantiophyta Jungermanniopsida Marchantiopsida Riccardiothallus devonicus 407.6 ± 2.6 Top of the Pragian Stage
7 Marchantiopsida Sphaerocarpales Marchantiales Marchantites cyathodoides 227.0 Top of the Carnian Stage 8 Stomatophyta Anthocerotophyta Bryophyta + Tracheophyta Cooksonia cf. pertoni 427.4 ± 0.5 Top of the C. ludensis Biozone 9 Unnamed Bryophyta Tracheophyta 10 Bryophyta Sphagnopsida Polytrichopsida + Bryopsida Sphagnales 330.9 ± 0.2 Top of the VF Biozone 11 Unnamed Polytrichopsida Bryopsida Palaeocampylopus buragoase 272.3 ± 0.5 Top of the Kungurian Stage 12 Bryopsida Funariidae Dicranidae + Bryidae Capimirinus riopretensis 268.8 ± 0.5 Top of the Roadian Stage 13 Tracheophyta Lycopodiophyta Euphyllophyta Zosterophyllum sp. 423.0 ± 2.3 Top of the Ludlow Epoch 451 Trilete spores; base of the
morrisi sub-biozone of Dicranograptus clingani Biozone
14 Lycopodiophyta Isoetopsida Lycopodiopsida Leclercqia complexa 393.3 ± 1.2 Top of the Emsian Stage 15 Euphyllophyta Monilophyta Spermatophyta Rellimia thomsonii 386.25 ± 0.679 rhenanus-ansatus boundary 16 Monilophyta Equisetopsida Polypodiopsida + Psilotopsida +
Marattiopsida Ibyka amphikoma 385.41 ± 0.704 Top of the P. ansatus Biozone
17 Unnamed Psilotopsida + Marattiopsida Polypodiopsida Psaronius simplicicaulis 319.09 ± 0.4 Base of the D. margindosus (Pc5) Biozone
19 Spermatophyta Acrogymnospermae Angiospermae Cordaixylon iowensis 308.5 ± 0.36 Top of the N. medexultimus -S. concinnus (Pc10) Biozone
364.19 ± 1.439 First seeds; base of P. trachytera Biozone 20 Acrogymnospermae Pinopsida Ginkgoopsida + Cycadopsida
21 Unnamed Ginkgoopsida Cycadopsida Crossozamia chinensis 265.1 ± 0.4 Top of the Wordian Stage 22 Pinopsida Cupressophyta Pinaceae + Gnetales Araucaria miribilis 157.0 ± 10 Igneous rocks overlying oldest
Araucaria fossils 321.01 ± 0.37 Cordaixylon iowensis bearing
strata; maximum age of Composite Level below conodont Pc10 Biozone
24 Unnamed Pinaceae Gnetales Eoantha zherikhinii 120.3 ± 0.7 Igneous rocks above oldest Eoantha zherikhinii
25 Angiospermae Amborellales Nymphaeales + Austrobaileyales + Mesangiospermae
Tricolpate pollen 125 Top of the Mn1 polarity chron / Barremian Stage
247.2 Sediments below the oldest occurrence of angiosperm-like pollen; base of the Anisian Stage
26 Unnamed Nymphaeales Austrobaileyales + Mesangiospermae 27 Unnamed Austrobaileyales Mesangiospermae 28 Mesangiospermae Magnoliids + Chloranthales +
Eudicotyledoneae Monocotyledoneae
29 Unnamed Magnoliids Chloranthales + Eudicotyledoneae 30 Magnoliids Magnoliales + Laurales Piperales Endressinia brasiliana 110..87 Early - Middle Albian Boundary;
base of D. mammillatum Biozone 31 Unnamed Persea Liriodendron
32 Piperales Saururus Houttuynia Saururus tuckerae 46.2 ± 1.9 Youngest radiometric date estimate for Allenby Formation
33 Unnamed Chloranthales Eudicotyledoneae Hyrcantha decussata 120.3 ± 0.7 Radiometric date from the overlying Jiufontang Formation
128.63 Oldest potential age of tricolpate pollen; base of the M1n chron/ G. sartousi Biozone
34 Eudicotyledoneae Ranunculales Vitales + Rosids + [Caryophyllales + Asterids]
35 Unnamed Vitales Rosids + [Caryophyllales + Asterids] Paleoclusia chevalieri / Paleoenkianthus sayrevillensis
86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage 36 Unnamed Rosids (minus Vitales) Caryophyllales + Asterids
37 Unnamed Caryophyllales Asterids Paleoenkianthus sayrevillensis 38 Monocotyledoneae Acorales Dioscoreales + Liliales +Asparagales
+ Aracales + Poales Liliacidites sp. A 113 Top of the Aptian Stage
39 Unnamed Dioscoreales Liliales + Asparagales + Aracales + Poales
Mabelia connatifila 86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage
40 Unnamed Liliales Asparagales + Aracales + Poales Sabalites carolinensis 83.64 ± 0.23 Top of the Santonian Stage / S. leei III Biozone
Supplementary Table S6. Calibrations for liverworts-hornworts-mosses topology (hypothesis vi).

Node Clade Lineage 1 Lineage 2 Minimum providing fossil Min. date (Ma) Minimum age source Max, date (Ma) Soft maximum age source 1 Viridiplantae Chlorophyta Streptophyta Tetrahedraletes cf. medinensis 469.0 Top of the L. combazi Biozone 1823 ± 68 Igneous and metamorphic
rocks underlying deposits containing the earliest record of eukaryotes
2 Unnamed Chlorophyceae Prasinophyaceae Palaeocymopolia silurica 439.21 ± 0.9031 Top of P. leptotheca Biozone 3 Streptophyta Charophyta Embryophyta Tetrahedraletes cf. medinensis 469.0 Top of the L. combazi Biozone
4 Embryophyta Marchantiophyta Stomatophytes 515.5 Oldest cryptospores; base of the Olenellus Biozone 5 Marchantiophyta Jungermanniopsida Marchantiopsida Riccardiothallus devonicus 407.6 ± 2.6 Top of the Pragian Stage
7 Marchantiopsida Sphaerocarpales Marchantiales Marchantites cyathodoides 227.0 Top of the Carnian Stage 8 Stomatophyta Anthocerotophyta + Bryophyta Tracheophyta Cooksonia cf. pertoni 427.4 ± 0.5 Top of the C. ludensis Biozone 9 Unnamed Anthocerotophyta Bryophyta Sphagnales 330.9 ± 0.2 Top of the VF Biozone 10 Bryophyta Sphagnopsida Polytrichopsida + Bryopsida 11 Unnamed Polytrichopsida Bryopsida Palaeocampylopus buragoase 272.3 ± 0.5 Top of the Kungurian Stage 12 Bryopsida Funariidae Dicranidae + Bryidae Capimirinus riopretensis 268.8 ± 0.5 Top of the Roadian Stage 13 Tracheophyta Lycopodiophyta Euphyllophyta Zosterophyllum sp. 423.0 ± 2.3 Top of the Ludlow Epoch 451 Trilete spores; base of the
morrisi sub-biozone of Dicranograptus clingani Biozone
14 Lycopodiophyta Isoetopsida Lycopodiopsida Leclercqia complexa 393.3 ± 1.2 Top of the Emsian Stage 15 Euphyllophyta Monilophyta Spermatophyta Rellimia thomsonii 386.25 ± 0.679 rhenanus-ansatusboundary 16 Monilophyta Equisetopsida Polypodiopsida + Psilotopsida +
Marattiopsida Ibyka amphikoma 385.41 ± 0.704 Top of the P. ansatus Biozone
17 Unnamed Psilotopsida + Marattiopsida Polypodiopsida Psaronius simplicicaulis 319.09 ± 0.4 Base of the D. margindosus (Pc5) Biozone
19 Spermatophyta Acrogymnospermae Angiospermae Cordaixylon iowensis 308.5 ± 0.36 Top of the N. medexultimus -S. concinnus (Pc10) Biozone
364.19 ± 1.439 First seeds; base of P. trachytera Biozone 20 Acrogymnospermae Pinopsida Ginkgoopsida + Cycadopsida
21 Unnamed Ginkgoopsida Cycadopsida Crossozamia chinensis 265.1 ± 0.4 Top of the Wordian Stage 22 Pinopsida Cupressophyta Pinaceae + Gnetales Araucaria miribilis 157.0 ± 10 Igneous rocks overlying oldest
Araucaria fossils 321.01 ± 0.37 Cordaixylon iowensis
bearing strata; maximum age of Composite Level below conodont Pc10 Biozone
24 Unnamed Pinaceae Gnetales Eoantha zherikhinii 120.3 ± 0.7 Igneous rocks above oldest Eoantha zherikhinii
25 Angiospermae Amborellales Nymphaeales + Austrobaileyales + Mesangiospermae
Tricolpate pollen 125 Top of the Mn1 polarity chron / Barremian Stage
247.2 Sediments below the oldest occurrence of angiosperm-like pollen; base of the Anisian Stage
26 Unnamed Nymphaeales Austrobaileyales + Mesangiospermae 27 Unnamed Austrobaileyales Mesangiospermae 28 Mesangiospermae Magnoliids + Chloranthales +
Eudicotyledoneae Monocotyledoneae
29 Unnamed Magnoliids Chloranthales + Eudicotyledoneae 30 Magnoliids Magnoliales + Laurales Piperales Endressinia brasiliana 110..87 Early - Middle Albian
Boundary; base of D. mammillatum Biozone 31 Unnamed Persea Liriodendron
32 Piperales Saururus Houttuynia Saururus tuckerae 46.2 ± 1.9 Youngest radiometric date estimate for Allenby Formation
33 Unnamed Chloranthales Eudicotyledoneae Hyrcantha decussata 120.3 ± 0.7 Radiometric date from the overlying Jiufontang Formation
128.63 Oldest potential age of tricolpate pollen; base of the M1n chron/ G. sartousi Biozone
34 Eudicotyledoneae Ranunculales Vitales + Rosids + [Caryophyllales + Asterids]
35 Unnamed Vitales Rosids + [Caryophyllales + Asterids] Paleoclusia chevalieri / Paleoenkianthus sayrevillensis
86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage 36 Unnamed Rosids (minus Vitales) Caryophyllales + Asterids
37 Unnamed Caryophyllales Asterids Paleoenkianthus sayrevillensis 38 Monocotyledoneae Acorales Dioscoreales + Liliales +Asparagales +
Aracales + Poales Liliacidites sp. A 113 Top of the Aptian Stage
39 Unnamed Dioscoreales Liliales + Asparagales + Aracales + Poales
Mabelia connatifila 86.3 ± 0.5 Top of the CC14 Nannofossil Biozone / Coniacian Stage
40 Unnamed Liliales Asparagales + Aracales + Poales Sabalites carolinensis 83.64 ± 0.23 Top of the Santonian Stage / S. leei III Biozone
Supplementary Table S7. Calibrations for liverworts sister topology (hypothesis vii).
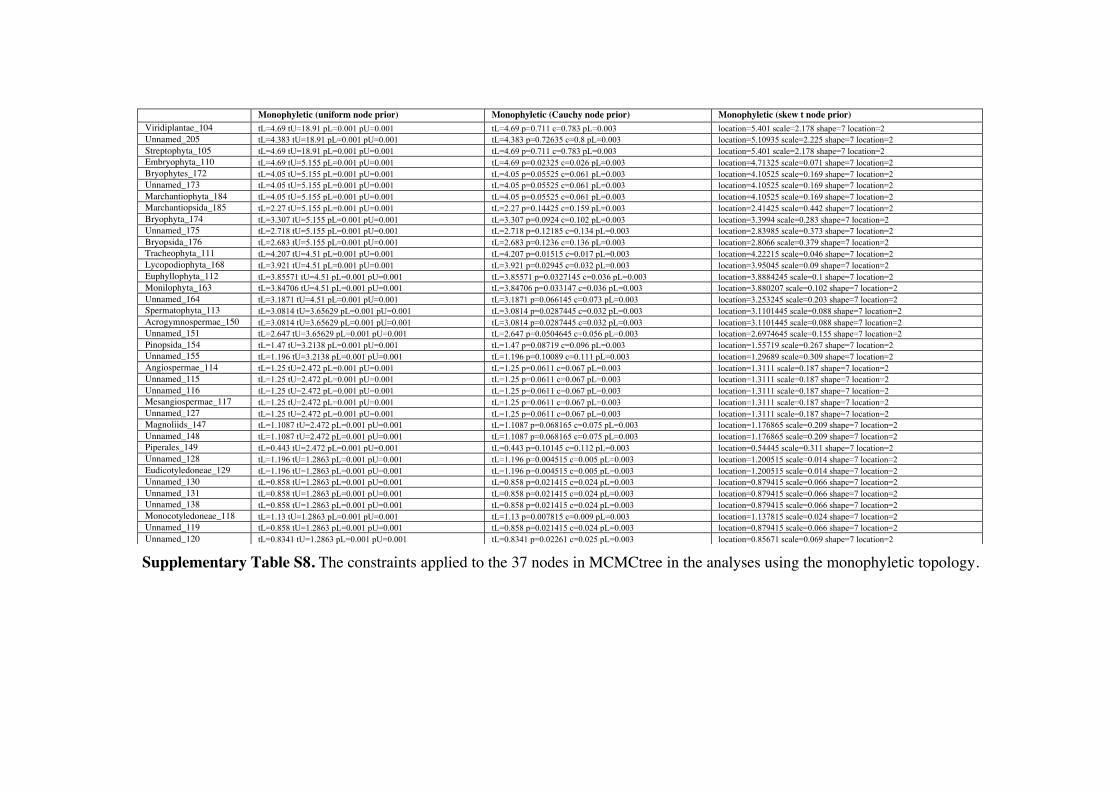
Supplementary Table S8. The constraints applied to the 37 nodes in MCMCtree in the analyses using the monophyletic topology.
Monophyletic (uniform node prior) Monophyletic (Cauchy node prior) Monophyletic (skew t node prior) Viridiplantae_104 tL=4.69 tU=18.91 pL=0.001 pU=0.001 tL=4.69 p=0.711 c=0.783 pL=0.003 location=5.401 scale=2.178 shape=7 location=2 Unnamed_205 tL=4.383 tU=18.91 pL=0.001 pU=0.001 tL=4.383 p=0.72635 c=0.8 pL=0.003 location=5.10935 scale=2.225 shape=7 location=2 Streptophyta_105 tL=4.69 tU=18.91 pL=0.001 pU=0.001 tL=4.69 p=0.711 c=0.783 pL=0.003 location=5.401 scale=2.178 shape=7 location=2 Embryophyta_110 tL=4.69 tU=5.155 pL=0.001 pU=0.001 tL=4.69 p=0.02325 c=0.026 pL=0.003 location=4.71325 scale=0.071 shape=7 location=2 Bryophytes_172 tL=4.05 tU=5.155 pL=0.001 pU=0.001 tL=4.05 p=0.05525 c=0.061 pL=0.003 location=4.10525 scale=0.169 shape=7 location=2 Unnamed_173 tL=4.05 tU=5.155 pL=0.001 pU=0.001 tL=4.05 p=0.05525 c=0.061 pL=0.003 location=4.10525 scale=0.169 shape=7 location=2 Marchantiophyta_184 tL=4.05 tU=5.155 pL=0.001 pU=0.001 tL=4.05 p=0.05525 c=0.061 pL=0.003 location=4.10525 scale=0.169 shape=7 location=2 Marchantiopsida_185 tL=2.27 tU=5.155 pL=0.001 pU=0.001 tL=2.27 p=0.14425 c=0.159 pL=0.003 location=2.41425 scale=0.442 shape=7 location=2 Bryophyta_174 tL=3.307 tU=5.155 pL=0.001 pU=0.001 tL=3.307 p=0.0924 c=0.102 pL=0.003 location=3.3994 scale=0.283 shape=7 location=2 Unnamed_175 tL=2.718 tU=5.155 pL=0.001 pU=0.001 tL=2.718 p=0.12185 c=0.134 pL=0.003 location=2.83985 scale=0.373 shape=7 location=2 Bryopsida_176 tL=2.683 tU=5.155 pL=0.001 pU=0.001 tL=2.683 p=0.1236 c=0.136 pL=0.003 location=2.8066 scale=0.379 shape=7 location=2 Tracheophyta_111 tL=4.207 tU=4.51 pL=0.001 pU=0.001 tL=4.207 p=0.01515 c=0.017 pL=0.003 location=4.22215 scale=0.046 shape=7 location=2 Lycopodiophyta_168 tL=3.921 tU=4.51 pL=0.001 pU=0.001 tL=3.921 p=0.02945 c=0.032 pL=0.003 location=3.95045 scale=0.09 shape=7 location=2 Euphyllophyta_112 tL=3.85571 tU=4.51 pL=0.001 pU=0.001 tL=3.85571 p=0.0327145 c=0.036 pL=0.003 location=3.8884245 scale=0.1 shape=7 location=2 Monilophyta_163 tL=3.84706 tU=4.51 pL=0.001 pU=0.001 tL=3.84706 p=0.033147 c=0.036 pL=0.003 location=3.880207 scale=0.102 shape=7 location=2 Unnamed_164 tL=3.1871 tU=4.51 pL=0.001 pU=0.001 tL=3.1871 p=0.066145 c=0.073 pL=0.003 location=3.253245 scale=0.203 shape=7 location=2 Spermatophyta_113 tL=3.0814 tU=3.65629 pL=0.001 pU=0.001 tL=3.0814 p=0.0287445 c=0.032 pL=0.003 location=3.1101445 scale=0.088 shape=7 location=2 Acrogymnospermae_150 tL=3.0814 tU=3.65629 pL=0.001 pU=0.001 tL=3.0814 p=0.0287445 c=0.032 pL=0.003 location=3.1101445 scale=0.088 shape=7 location=2 Unnamed_151 tL=2.647 tU=3.65629 pL=0.001 pU=0.001 tL=2.647 p=0.0504645 c=0.056 pL=0.003 location=2.6974645 scale=0.155 shape=7 location=2 Pinopsida_154 tL=1.47 tU=3.2138 pL=0.001 pU=0.001 tL=1.47 p=0.08719 c=0.096 pL=0.003 location=1.55719 scale=0.267 shape=7 location=2 Unnamed_155 tL=1.196 tU=3.2138 pL=0.001 pU=0.001 tL=1.196 p=0.10089 c=0.111 pL=0.003 location=1.29689 scale=0.309 shape=7 location=2 Angiospermae_114 tL=1.25 tU=2.472 pL=0.001 pU=0.001 tL=1.25 p=0.0611 c=0.067 pL=0.003 location=1.3111 scale=0.187 shape=7 location=2 Unnamed_115 tL=1.25 tU=2.472 pL=0.001 pU=0.001 tL=1.25 p=0.0611 c=0.067 pL=0.003 location=1.3111 scale=0.187 shape=7 location=2 Unnamed_116 tL=1.25 tU=2.472 pL=0.001 pU=0.001 tL=1.25 p=0.0611 c=0.067 pL=0.003 location=1.3111 scale=0.187 shape=7 location=2 Mesangiospermae_117 tL=1.25 tU=2.472 pL=0.001 pU=0.001 tL=1.25 p=0.0611 c=0.067 pL=0.003 location=1.3111 scale=0.187 shape=7 location=2 Unnamed_127 tL=1.25 tU=2.472 pL=0.001 pU=0.001 tL=1.25 p=0.0611 c=0.067 pL=0.003 location=1.3111 scale=0.187 shape=7 location=2 Magnoliids_147 tL=1.1087 tU=2.472 pL=0.001 pU=0.001 tL=1.1087 p=0.068165 c=0.075 pL=0.003 location=1.176865 scale=0.209 shape=7 location=2 Unnamed_148 tL=1.1087 tU=2.472 pL=0.001 pU=0.001 tL=1.1087 p=0.068165 c=0.075 pL=0.003 location=1.176865 scale=0.209 shape=7 location=2 Piperales_149 tL=0.443 tU=2.472 pL=0.001 pU=0.001 tL=0.443 p=0.10145 c=0.112 pL=0.003 location=0.54445 scale=0.311 shape=7 location=2 Unnamed_128 tL=1.196 tU=1.2863 pL=0.001 pU=0.001 tL=1.196 p=0.004515 c=0.005 pL=0.003 location=1.200515 scale=0.014 shape=7 location=2 Eudicotyledoneae_129 tL=1.196 tU=1.2863 pL=0.001 pU=0.001 tL=1.196 p=0.004515 c=0.005 pL=0.003 location=1.200515 scale=0.014 shape=7 location=2 Unnamed_130 tL=0.858 tU=1.2863 pL=0.001 pU=0.001 tL=0.858 p=0.021415 c=0.024 pL=0.003 location=0.879415 scale=0.066 shape=7 location=2 Unnamed_131 tL=0.858 tU=1.2863 pL=0.001 pU=0.001 tL=0.858 p=0.021415 c=0.024 pL=0.003 location=0.879415 scale=0.066 shape=7 location=2 Unnamed_138 tL=0.858 tU=1.2863 pL=0.001 pU=0.001 tL=0.858 p=0.021415 c=0.024 pL=0.003 location=0.879415 scale=0.066 shape=7 location=2 Monocotyledoneae_118 tL=1.13 tU=1.2863 pL=0.001 pU=0.001 tL=1.13 p=0.007815 c=0.009 pL=0.003 location=1.137815 scale=0.024 shape=7 location=2 Unnamed_119 tL=0.858 tU=1.2863 pL=0.001 pU=0.001 tL=0.858 p=0.021415 c=0.024 pL=0.003 location=0.879415 scale=0.066 shape=7 location=2 Unnamed_120 tL=0.8341 tU=1.2863 pL=0.001 pU=0.001 tL=0.8341 p=0.02261 c=0.025 pL=0.003 location=0.85671 scale=0.069 shape=7 location=2

Supplementary Table S9. The constraints applied to the 37 nodes in MCMCtree in the analyses using the hornworts-sister topology.
Hornworts-sister (uniform node prior) Hornworts-sister (Cauchy node prior) Hornworts-sister (skew t node prior) Viridiplantae_104 tL=4.69 tU=18.91 pL=0.001 pU=0.001 tL=4.69 p=0.711 c=0.783 pL=0.003 location=5.401 scale=2.178 shape=7 df=2 Unnamed_205 tL=4.383 tU=18.91 pL=0.001 pU=0.001 tL=4.383 p=0.72635 c=0.8 pL=0.003 location=5.10935 scale=2.225 shape=7 df=2 Streptophyta_105 tL=4.69 tU=18.91 pL=0.001 pU=0.001 tL=4.69 p=0.711 c=0.783 pL=0.003 location=5.401 scale=2.178 shape=7 df=2 Embryophyta_110 tL=4.69 tU=5.155 pL=0.001 pU=0.001 tL=4.69 p=0.02325 c=0.026 pL=0.003 location=4.71325 scale=0.071 shape=7 df=2 Unnamed_111 tL=4.269 tU=5.155 pL=0.001 pU=0.001 tL=4.269 p=0.0443 c=0.049 pL=0.003 location=4.3133 scale=0.136 shape=7 df=2 Unnamed_173 tL=4.05 tU=5.155 pL=0.001 pU=0.001 tL=4.05 p=0.05525 c=0.061 pL=0.003 location=4.10525 scale=0.169 shape=7 df=2 Marchantiophyta_184 tL=4.05 tU=5.155 pL=0.001 pU=0.001 tL=4.05 p=0.05525 c=0.061 pL=0.003 location=4.10525 scale=0.169 shape=7 df=2 Marchantiopsida_185 tL=2.27 tU=5.155 pL=0.001 pU=0.001 tL=2.27 p=0.14425 c=0.159 pL=0.003 location=2.41425 scale=0.442 shape=7 df=2 Bryophyta_174 tL=3.307 tU=5.155 pL=0.001 pU=0.001 tL=3.307 p=0.0924 c=0.102 pL=0.003 location=3.3994 scale=0.283 shape=7 df=2 Unnamed_175 tL=2.718 tU=5.155 pL=0.001 pU=0.001 tL=2.718 p=0.12185 c=0.134 pL=0.003 location=2.83985 scale=0.373 shape=7 df=2 Bryopsida_176 tL=2.683 tU=5.155 pL=0.001 pU=0.001 tL=2.683 p=0.1236 c=0.136 pL=0.003 location=2.8066 scale=0.379 shape=7 df=2 Tracheophyta_112 tL=4.207 tU=4.51 pL=0.001 pU=0.001 tL=4.207 p=0.01515 c=0.017 pL=0.003 location=4.22215 scale=0.046 shape=7 df=2 Lycopodiophyta_169 tL=3.921 tU=4.51 pL=0.001 pU=0.001 tL=3.921 p=0.02945 c=0.032 pL=0.003 location=3.95045 scale=0.09 shape=7 df=2 Euphyllophyta_113 tL=3.85571 tU=4.51 pL=0.001 pU=0.001 tL=3.85571 p=0.0327145 c=0.036 pL=0.003 location=3.8884245 scale=0.1 shape=7 df=2 Monilophyta_164 tL=3.84706 tU=4.51 pL=0.001 pU=0.001 tL=3.84706 p=0.033147 c=0.036 pL=0.003 location=3.880207 scale=0.102 shape=7 df=2 Unnamed_165 tL=3.1871 tU=4.51 pL=0.001 pU=0.001 tL=3.1871 p=0.066145 c=0.073 pL=0.003 location=3.253245 scale=0.203 shape=7 df=2 Spermatophyta_114 tL=3.0814 tU=3.65629 pL=0.001 pU=0.001 tL=3.0814 p=0.0287445 c=0.032 pL=0.003 location=3.1101445 scale=0.088 shape=7 df=2 Acrogymnospermae_151 tL=3.0814 tU=3.65629 pL=0.001 pU=0.001 tL=3.0814 p=0.0287445 c=0.032 pL=0.003 location=3.1101445 scale=0.088 shape=7 df=2 Unnamed_161 tL=2.647 tU=3.65629 pL=0.001 pU=0.001 tL=2.647 p=0.0504645 c=0.056 pL=0.003 location=2.6974645 scale=0.155 shape=7 df=2 Pinopsida_152 tL=1.47 tU=3.2138 pL=0.001 pU=0.001 tL=1.47 p=0.08719 c=0.096 pL=0.003 location=1.55719 scale=0.267 shape=7 df=2 Unnamed_153 tL=1.196 tU=3.2138 pL=0.001 pU=0.001 tL=1.196 p=0.10089 c=0.111 pL=0.003 location=1.29689 scale=0.309 shape=7 df=2 Angiospermae_115 tL=1.25 tU=2.472 pL=0.001 pU=0.001 tL=1.25 p=0.0611 c=0.067 pL=0.003 location=1.3111 scale=0.187 shape=7 df=2 Unnamed_116 tL=1.25 tU=2.472 pL=0.001 pU=0.001 tL=1.25 p=0.0611 c=0.067 pL=0.003 location=1.3111 scale=0.187 shape=7 df=2 Unnamed_117 tL=1.25 tU=2.472 pL=0.001 pU=0.001 tL=1.25 p=0.0611 c=0.067 pL=0.003 location=1.3111 scale=0.187 shape=7 df=2 Mesangiospermae_118 tL=1.25 tU=2.472 pL=0.001 pU=0.001 tL=1.25 p=0.0611 c=0.067 pL=0.003 location=1.3111 scale=0.187 shape=7 df=2 Unnamed_128 tL=1.25 tU=2.472 pL=0.001 pU=0.001 tL=1.25 p=0.0611 c=0.067 pL=0.003 location=1.3111 scale=0.187 shape=7 df=2 Magnoliids_129 tL=1.1087 tU=2.472 pL=0.001 pU=0.001 tL=1.1087 p=0.068165 c=0.075 pL=0.003 location=1.176865 scale=0.209 shape=7 df=2 Unnamed_130 tL=1.1087 tU=2.472 pL=0.001 pU=0.001 tL=1.1087 p=0.068165 c=0.075 pL=0.003 location=1.176865 scale=0.209 shape=7 df=2 Piperales_131 tL=0.443 tU=2.472 pL=0.001 pU=0.001 tL=0.443 p=0.10145 c=0.112 pL=0.003 location=0.54445 scale=0.311 shape=7 df=2 Unnamed_132 tL=1.196 tU=1.2863 pL=0.001 pU=0.001 tL=1.196 p=0.004515 c=0.005 pL=0.003 location=1.200515 scale=0.014 shape=7 df=2 Eudicotyledoneae_133 tL=1.196 tU=1.2863 pL=0.001 pU=0.001 tL=1.196 p=0.004515 c=0.005 pL=0.003 location=1.200515 scale=0.014 shape=7 df=2 Unnamed_134 tL=0.858 tU=1.2863 pL=0.001 pU=0.001 tL=0.858 p=0.021415 c=0.024 pL=0.003 location=0.879415 scale=0.066 shape=7 df=2 Unnamed_135 tL=0.858 tU=1.2863 pL=0.001 pU=0.001 tL=0.858 p=0.021415 c=0.024 pL=0.003 location=0.879415 scale=0.066 shape=7 df=2 Unnamed_142 tL=0.858 tU=1.2863 pL=0.001 pU=0.001 tL=0.858 p=0.021415 c=0.024 pL=0.003 location=0.879415 scale=0.066 shape=7 df=2 Monocotyledoneae_119 tL=1.13 tU=1.2863 pL=0.001 pU=0.001 tL=1.13 p=0.007815 c=0.009 pL=0.003 location=1.137815 scale=0.024 shape=7 df=2 Unnamed_120 tL=0.858 tU=1.2863 pL=0.001 pU=0.001 tL=0.858 p=0.021415 c=0.024 pL=0.003 location=0.879415 scale=0.066 shape=7 df=2 Unnamed_121 tL=0.8341 tU=1.2863 pL=0.001 pU=0.001 tL=0.8341 p=0.02261 c=0.025 pL=0.003 location=0.85671 scale=0.069 shape=7 df=2

References 1. Rubenstein, C.V., Gerrienne, P., Puente de la, G. S., Astini, R.A., and Steemans, P., 2010. Early Middle
Ordovician evidence for land plants in Argentina (eastern Gondwana). New Phytologist, 188: 365-369. 2. Gray, J., 1985. The microfossil record of early land plants: advances in understanding of early terrestrialization,
1970 – 1984. Philosophical Transactions of the Royal Society of London, B, 309: 167 – 195. 3. Wellman, C.H., Gray, J., 2000. The microfossil record of early land plants. Phil. Trans. R. Soc. Lond. B., 355: 717
– 732. 4. Strother, P.K., Traverse, A., Vecoli, M., 2015. Cryptospores from the Hanadir Shale Member of the Qasim
Formation, Ordovician (Darriwilian) of Saudi Arabia: taxonomy and systematics. Review of Palaeobotany and Palynology, 212: 97-110.
5. Renzaglia, K.S., Crandall-Stotler, B., Pressel, S., Duckett, J.G., Schuette, S., Strother, P.K., 2015. Permanent spore dyads are not ‘a thing of the past’: on their occurrence in the liverwort Haplomitrium (Haplomitriopsida). Botanical Journal of the Linnean Society, 179: 658 – 669.
6. Renzaglia, K.S., Lopez, R.A., Johnson, E.E., 2015. Callose is integral to the development of permanent tetrads in the liverwort Sphaerocarpos. Planta, 241: 615 – 627.
7. Brown, R.C., Lemmon, B.E., Shimamura, M., Villarreal, J.C., Renzaglia, K.S., 2015. Spores of relictual bryophytes: diverse adaptations to life on land. Review of Palaeobotany and Palynology, 216: 1 – 17.
8. Wellman, C.H., 2010. The invasion of the land by plants: when and where? New Phytologist, 188 (2): 306 – 309. 9. Steemans, P., 2000. Miospore evolution from the Ordovician to the Silurian. Review of Palaeobotany and
Palynology, 113: 189 – 196. 10. Steemans, P., Lepot, K., Marshall, C.P., Le Hérissé, A., Javaux, E.J., 2010. FTIR characterisation of the chemical
composition of Silurian miospores (cryptospores and trilete spores) from Gotland, Sweden. Review of Palaeobotany and Palynology, 162: 577 – 590.
11. Taylor, W.A., 1995. Ultrastructure of Tetrahedraletes medinensis (Strother and Traverse) Wellman and Richardson, from the Upper Ordovician of southern Ohio. Review of Palaeobotany and Palynology, 85: 183 – 187.
12. Wellman, C.H., Osterloff, P.L., Mohiuddin, U., 2003. Fragments of the earliest land plants. Nature, 425: 282 – 285.
13. Edwards, D., Morris, J.L., Richardson, J.B., Kenrick, P., 2014. Cryptospores and cryptophytes reveal hidden diversity in early land floras. Tansley Review, New Phytologist, 202: 50 – 78.
14. Chen, X., Paris, F., Wang, X., Zhang, M., 2009. Early and Middle Ordovician chitinozoans from the Dapingian type sections, Yichang area, China. Review of Palaeobotany and Palynology, 153: 310 – 330.
15. Wang, X., Stouge, S., Erdtmann, B.-D., Chen, X., Li, Z., Wang, C., Zeng, Q., Zhou, Z., Chen, H., 2005. A proposed GSSP for the base of the Middle Ordovician Series: the Huanghuachang section, Yichang, China. Episodes, 28 (2): 105 – 117.
16. Chen, X., Bergström, S.M., Zhang, Y.-D., Fan, J.-X., 2009. The base of the Middle Ordovician in China with special reference to the succession at Hengtang near Jiangshan, Zhejiang Province, southern China. Lethaia, 42: 218 – 231.
17. Cooper, R.A., Sadler, P.M., 2012. The Ordovician Period. In: Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M. (eds.), The Geological Time Scale 2012, Volume 2, Chapter 20, Elsevier Press: 489 – 523.
18. Paris, F., 1990. The Ordovician chitinozoan biozones of the Northern Gondwana Domain. Review of Palaeobotany and Palynology, 66: 190 – 191.
19. Zhang, Z.-Y., 1986. Clastic facies microfossils from the Chuanlinggou Formation (1800 Ma) near Jixian, North China. Journal of Micropalaeontology, 5: 9 – 16.
20. Lamb, D.M., Awramik, S.M., Chapman, D.J., Zhu, S., 2009. Evidence for eukaryotic diversification in the ~1800 million-year-old Changzhougou Formation, North China. Precambrian Research, 173: 93 – 104.
21. Peng, Y., Bao, H., Yuan, X., 2009. New morphological observations for Paleoproterozoic acritarchs from the Chuanlinggou Formation, North China. Precambrian Research, 168: 223 – 232.
22. Moczydlowska, M., Landing, E.D., Zang, W., Palacios, T., 2011. Proterozoic phytoplankton and timing of Chlorophyte algae origins. Palaeontology, 54: 721 – 733.
23. Lu, S., Yang, C., Zhu, S., 1996. The Precambrian continental crust from sastern Hebei to Jixian. Tianjin 30th International Geological Congress, Beijing: Geological Publishing House.
24. Parfrey, L.W., Lahr, D.J.G., Knoll, A.H., Katz, L.A., 2011. Estimating the timing of early eukaryotic diversification with multigene molecular clocks. Proceedings of the National Academy of Sciences, 108: 13624 – 13629.
25. Butterfield, N.J., Knoll, A.H., Swett, K., 1994. Paleobiology of the Neoproterozoic Svanbergfjellet Formation, Spitsbergen. Fossils & Strata, 34: 1 – 84.
26. Butterfield, N.J., 2015. Early evolution of the Eukaryota. Palaeontology, 58: 5 – 17.

27. Xiao, S., Yuan, X., Steiner, M., Knoll, A.H., 2002. Macroscopic carbonaceous compressions in a terminal Proterozoic shale: a systematic reassessment of the Miahoe Biota, south China. Journal of Paleontology, 76: 347 – 376.
28. Riding, R., 2001. Calcified algae and bacteria. In: Zhuravlev, A.Y., Riding, R., (eds.), The ecology of the Cambrian radiation. New York: Columbia Univrersity Press: 445 – 473.
29. Riding, R., 1992. Cambrian calcareous cyanobacteria and algae. In: Riding, R. (ed.), Calcareous algae and stromatolites. New York, Springer: 305 – 334.
30. Verbruggen, H., Ashworth, M., LoDuca, S.T., Vlaeminck, C., Cocquyt, E., Sauvage, T., Zechman, F.W., Littler, D.S., Littler, M.M., Leliaert, F., De Clerck, O., 2009. A multi-locus time-calibrated phylogeny of the siphonous green algae. Molecular phylogenetics and Evolution, 50: 642 – 653.
31. Mastik, V., Tinn, O., 2015. New dasycladalean algal species from the Kalana Lagerstätte (Silurian, Estonia). Journal of Paleontology, 89: 262 – 268.
32. Ainsaar, L., Loydell, D.K., Tinn, O., Männik, P., 2016. Age of the Kalana Lagerstätte, early Silurian, Estonia. Estonian Journal of Earth Sciences, 65: 105.
33. Strother, P.K., Wood, G., Taylor, W., Beck, J., 2004. Middle Cambrian cryptospores and the origin of land plants. AAP Memoir 29: 99 – 113.
34. Strother, P.K., Beck, J.H., 2000. Spore-like microfossils from Middle Cambrian strata: expanding the meaning of the term cryptospore. In: Harley, M.M., Morton, C.M., Blackmore, S. (eds.), Pollen and Spores: Morphology and Biology. The Royal Botanical Gardens Kew: 413 – 424.
35. Baldwin, C.T., Strother, P.K., Beck, J.H., Rose, E., 2004. Palaeoecology of the Bright Angel Shale in the eastern Grand Canyon, Arizona, USA, incorporating sedimentological, ichnological and palynological data. In: McIlroy, D. (ed.) The Application of Ichnology to Palaeoenvironmental and Stratigraphic Analysis. Geological Society, London, Special Publications, 228: 213 – 236.
36. Strother, P.K., 2016. Systematics and evolutionary significance of some new cryptospores from the Cambrian of eastern Tennessee, USA. Review of Palaeobotany and Palynology, 227: 28 – 41.
37. Peng, S., Babcock, L.E., Cooper, R.A., 2012. The Cambrian Period. In: Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M. (eds.), The Geological Time Scale 2012, Volume 2, Chapter 19, Elsevier Press: 437 – 488.
38. Du, R.L.,1982. The discovery of the fossils such as Chuaria in the Qingbaikou System in Northwestern Hebei and their significance. Geological Review, 28: 1 – 7.
39. Du, R.L., Tian, L.F., 1985. Preliminary research of the macroalgae Longfengshania from Qingbaikou System, Yanshan areas. Acta Geologica Sinica, 59: 183 – 190.
40. Hofmann, H.J., 1985. The mid-Proterozoic Little Dal macrobiota, Mackenzie Mountains, north-west Canada. Palaeontology, 28: 331 – 354.
41. Xu, Z.L., 2002. The occurrence of Longfengshania in the Early Cambrian from Haikou, Yunnan, China. Acta Botanica Sinica, 44: 1250–1254.
42. Yang, R.D., Mao, J.R., Zhang, W.H., Jiang, L.J., Gao, H., 2004. Bryophyte-like fossil (Parafunaria sinensis) from Early - Middle Cambrian Kaili Formation in Guizhou Province, China. Acta Botanica Sinica, 46: 180 – 185.
43. Duan, C., Xing, Y., Du, R.L., Yin, Y., Liu, G., 1985. Macroscopic fossil algae. In: Xing, Y., Duan, C., Liang, Y., Duan, C., Liang, Y, Cao, R., (eds.). Late Precambrian palaeontology of China. Geological Publishing House: 68 – 77.
44. Liu, Z.-L., Du, R.-L., 1991. Morphology and systematics of Longfengshania. Acta Palaeontogica Sinica, 30: 106 – 114.
45. Conway Morris, S., 2006. Darwin’s dilemma: the realities of the Cambrian ‘explosion’. Philosophical Transactions of the Royal Society B- Biological Sciencesi, 361: 1069 – 1083.
46. Kenrick, P., Vinther, J., 2006. Chaetocladus gracilis n. sp., a non-calcified Dasycladales from the Upper Silurian of Skåne, Sweden. Review of Palaeobotany and Palynology, 142: 153 – 160.
47. Tang, F., Song, X., Yin, C., Liu, P., Awramik, S., Wang, Z., Gao, L., 2007. Discoveries of new Longfengshaniaceae from the uppermost Ediacaran in eastern Yunnan, South China and the significance. Frontiers of Earth Science in China, 1: 142 – 149.
48. Drinnan, A.N., Chambers, T.C., 1986. Flora of the Lower Cretaceous Koonwarra Fossil Bed (Korumburra Group), South Gippsland, Victoria. Association of Australasian Palaeontologists, Memoir 3: 1 – 77.
49. Nĕmejc, F., Pacltova, B., 1974. Hepaticae in the Senonian of South Bohemia. Palaeobotantist, 21: 23 – 26. 50. Taylor, W.A., Gensel, P.G., Wellman, C.H., 2011. Wall ultrastructure in three species of the dispersed spore
Emphanisporites from the Early Devonian. Review of Palaeobotany and Palynology, 163: 264 – 280. 51. Villarreal, J.C., Renzaglia, K.S., 2006. Sporophyte structure in the neotropical hornwort Phaeomegaceros
fimbriatus: implications for phylogeny, taxonomy and character evolution. International Journal of Plant Sciences, 167: 413 – 427.
52. Edwards, D., Feehan, J., 1980. Records of Cooksonia-type sporangia from late Wenlock strata in Ireland. Nature, 287: 41 – 42.

53. Edwards, D., Feehan, J., Smith, D.G., 1983. A late Wenlock flora from Co. Tipperary, Ireland. Botanical Journal of the Linnean Society, 86: 19 – 36.
54. Cardona-Correa, C., Piotrowski, M.J., Knack, J.J., Kodner, R.E., Geary, D.H., Graham, L.E. 2016. International Journal of Plant Sciences, 177, 523-538.
55. Halle, T.G., 1916. A fossil sporogonium from the Lower Devonian of Röragen in Norway. Botaniska Notiser 1916: 79 – 81.
56. Thomas, B.A., 1972. A probable moss from the Lower Carboniferous of the Forest of Dean, Gloucestershire. Annals of Botany, 35: 155 – 161.
57. Renault, B., Zeiller, R., 1888. Études sur les terrains houiller de Commentry. Livre deuxième. Flore fossile. St. Étienne: Imprimateur Théolier and Cie.
58. Ottone, E.G., Archangelsky, S., 2001. A new bryophyte from the Upper Carboniferous of Argentina. Ameghiniana, 38: 219 – 223.
59. Amaral P.G.C., Bernardes-de-Oliveira M.E., Ricardi-Branco F., Broutin J., 2004. Primera ocurrencia de uma briófita fértil en el Subgrupo Itararé (Carbonífero), Estado de São Paulo, Brasil. Tropical Bryology, 25: 101 – 110.
60. Hübers, M., Kerp, H., 2012. Oldest known mosses discovered in Mississippian (late Visean) strata of Germany. Geology, 40 (8): 755 – 758.
61. Guo, C.-Q., Edwards, D., Wu, P.-C., Duckett, J.G., 2012. Riccardiothallus devonicus gen. et sp. nov., the earliest simple thalloid liverwort from the Lower Devonian of Yunnan, China. Review of Palaeobotany and Palynology, 176 – 177: 35 – 40.
62. Clarke, J.T., Warnock, R.C.M., Donoghue, P.C.J., 2011. Establishing a time-scale for plant evolution. New Phytologist, 192: 266 – 301.
63. Taylor, W.A., Strother, P.K., 2009. Ultrastructure, morphology, and topology of Cambrian palynomorphs from the Lone Rock Formation, Wisconsin, USA. Review of Palaeobotany and Palynology, 153: 296 – 309.
64. Yin, L., Zhao, Y., Bian, L., Peng, J., 2013. Comparison between cryptospores from the Cambrian Log Cabin Member, Pioche Shale, Nevada, USA and similar specimens from the Cambrian Kaili Formation, Guizhou, China. Science China Earth Sciences, 56: 703 – 709.
65. Taylor, W.A., Strother, P.K., 2008. Ultrastructure of some Cambrian palynomorphs from the Bright Angel Shale, Arizona, USA. Review of Palaeobotany and Palynology, 151: 41 – 50.
66. Graham, L.E., Arancibia-Avila, P., Taylor, W.A., Strother, P.K., Cook, M.E., 2012. Aeroterrestrial Coleochaete (Streptophyta, Coleochaetales) models early plant adaptation to land. American Journal of Botany, 99: 130 – 144.
67. Edwards, D., Davies, K.L., Axe, L., 1992. A vascular conducting strand in the early land plant Cooksonia. Nature, 357: 683 – 685.
68. Edwards, D., Fanning, U., Richardson, J.B., 1986. Stomata and sterome in early land plants. Nature, 323: 438 – 440.
69. Kenrick, P., Crane, P.R., 1997. The origin and early diversification of land plants. A cladistics study. Smithsonian Institution Press, Washington, London, 441pp.
70. Melchin, M.J., Sadler, P.M., Cramer, B.D., 2012. The Silurian Period. In: Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M. (eds.), The Geological Time Scale 2012, Volume 2, Chapter 21, Elsevier Press: 525 – 558.
71. Neuburg, M.F., 1956. The discovery of mosses in the Permian deposits of the USSR: Doklady Akademii Nauk SSSR, 107: 321 – 324.
72. Ignatov, M.S., Shcherbakov, D.E., 2009. A new fossil moss from the Lower Permian of the Russian Far East. Arctoa, 18: 201 – 212.
73. Brown, R.C., Lemmon, B.E., 1984. Spore wall development in Andreaea (Musci: Andreaeopsida). American Journal of Botany, 71 (3): 412 – 420.
74. Brown, R.C., Lemmon, B.E., Carothers, Z.B., 1982. Spore wall ultrastructure of Sphagnum lescurii Sull. Review of Palaeobotany and Palynology, 38: 99 – 107.
75. Renzaglia, K.S., McFarland, K.D., Smith, D.K., 1997. Anatomy and ultrastructure of the sporophyte of Takakia ceratophylla (Bryophyta). American Journal of Botany, 84: 1337 – 1350.
76. Brown, E.A., Braggins, J.E., 1989. A revision of the genus Riccardia S.F. Gray in New Zealand with notes on the genus Aneura Dunn. Journal of the Hattori Botanical Laboratory, 66: 1 – 132.
77. Paton, J.A., 1999. The liverwort flora of the British Isles. Harley Books, Colchester. 78. Hao, S., Xue, J., 2013. The Early Devonian Posongchong Flora of Yunnan – A Contribution to an understanding
of the evolution and early diversification of vascular plants. Science Press Beijing: 366pp. 79. Bai, S.L., Dong, Z.Z., Jin, S.Y., 1982. The Devonian conodont sequence of Yunnan. In Bai, S.L., Jin, S.Y., Ning,
Z.S. (eds.), The Devonian of Guangxi and adjacent area. Beijing: Peking University Press: 67 – 71. 80. Klapper, G., Ziegler, W., 1979. Devonian conodont biostratigraphy. In: House, M.R., Scrutton, C.T., Bassett,
M.G., (eds.). The Devonian System. Special Papers in Palaeontology, 23: 199 – 224. 81. Jin, S.Y., Shen, A.J., Chen, Z.L., Lu, J.M., Wei, M., Wang, Y.Q., Xie, F., 2005. Mixed biostratigraphy of
Devonian in Wenshan, Yunnan. Beijing: Petroleum Industry Press.

82. Liao, W.H., Xu, H.K., Wang, C.Y., Ruan, Y.P., Cai, C.Y., Mu, D.C., Lu, L.C., 1978. The subdivision and correlation of the Devonian stratigraphy of Southwest China. In: Institute of Geology and Mineral Resources, Chinese Academy of Geological Sciences (ed.). Symposium on the Devonian System of South China. Beijing: Geological Publishing House: 193 – 213.
83. Ruan, Y.P., Wang, C.Y., Wang, Z.H., Rong, J.Y., Mu, D.C., Kuang, G.D., Yin, B.A., Sue, Y.B., 1979. On the age of the Nahkaoling and Yukiang Formations. Acta Stratigraphica Sinica, 3: 225 – 229.
84. Zhong, K., Wu, Y., Yin, B.A., Liang, Y.L., Yao, Z.G., Peng, J.L., 1992. Stratigraphy of Guangxi, China, Part 1: Devonian of Guangxi. Wuhan: China University of Geosciences Press.
85. Wang, Y., 1994. Lower Devonian miospores from Gumu in the Wenshan District, Southeastern Yunnan. Acta Micropalaeontologica Sinica, 11: 319 – 332.
86. Richardson, J.B., McGregor 1986. Silurian and Devonian spore zones of the Old Red Sandstone Continent and adjacent regions. Geological Survey of Canada, Bulletin, 364: 1 – 79.
87. Becker, R.T., Gradstein, F.M., Hammer, O., 2012. The Devonian Period. Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M., (eds), The Geologic Time Scale 2012, Volume 2, Chapter 22, Elsevier Press: 559 – 601.
88. Crandall-Stotler, B., Stotler, R.E., Long, D., 2009. Morphology and classification of the Marchantiophyta. In: Goffinet, B., Shaw, A.J., (eds.), Bryophyte Biology, 2nd edition, Cambridge University Press: 1 – 54.
89. Schuster, R.M., 1981. Paleoecology, origin, distribution through time, and evolution of Hepaticae and Anthocerotae. In: Niklas, K.J. (ed.), Paleobotany, Paleoecology, and Evolution, vol. 2. Praeger, New York, pp. 129 – 191.
90. Hernick, L.V., Landing, E., Bartowski, K.E., 2008. Earth’s oldest liverworts – Metzgeriothallus sharonae sp. nov. from the Middle Devonian (Givetian) of eastern New York, USA. Review of Palaeobotany and Palynology, 148: 154 – 162.
91. Schuster, R.M., 1966. The Hepaticae and Anthocerotae of North America, east of the hundredth meridian. Volume I, Columbia University Press, New York, London, pp. 802.
92. Taylor, W.A., 1997. Ultrastructure of lower Paleozoic dyads from southern Ohio II: Dyadospora murusattenuata, functional and evolutionary considerations. Review of Palaeobotany and Palynology, 97: 1 – 8.
93. Walton, J., 1928. Carboniferous bryophyta. II. Hepaticae and Musci. Annals of Botany, 42: 707 – 716. 94. Poulsen, C., 1974. Further contributions to the knowledge of the Palaeozoic of Slagelse no. 1, Western Sealand.
Yderligere bidrag til kendskabet til palaeozoikum i Slagelse nr. 1, Vestsjaelland. Danmarks Geol. Undersøl., Raekke 1–5 101. 42 pp.
95. Steenbock, C.M., Stockey, R.A., Beard, G., Tomescu, A.M.F., 2011. A new family of leafy liverworts from the Middle Eocene of Vancouver Island, British Columbia, Canada. American Journal of Botany, 98: 998 – 1006.
96. Yang, X., Wu, X., 2011. Sinolejeunea yimaensis gen. et sp. nov. (Hepaticae) from the Middle Jurassic Yima Formation in Henan Province of China. Science China Earth Sciences, 54: 228 – 232.
97. Brown, J.T., Robison, C.R., 1974. Diettertia montanensis, gen. et sp. nov., a fossil moss from the Lower Cretaceous Kootenai Formation of Montana. Botanical Gazette, 135: 170 – 173.
98. Schuster, R.M., Janssens, J.A., 1989. On Diettertia, an isolated Mesozoic member of Jungermaniales. Review of Palaeobotany and Palynology, 57: 277 – 287.
99. Townrow, J.A., 1959. Two Triassic bryophytes from South Africa. Journal of South African Botany 25: 1 -22. 100. Anderson, H.M., 1976. A review of the Bryophyta from the Upper Triassic Molteno Formation, Karroo Basin,
South Africa. Palaeont. Afr., 19: 21 – 30. 101. Heinrichs, J., Hentschel, J., Wilson, R., Feldberg, K., Schneider, H., 2007. Evolution of leafy liverworts
(Jungermanniidae, Marchantiophyta): estimating divergence times from chloroplast DNA sequences using penalized likelihood with integrated fossil evidence. Taxon, 56 (1): 31 – 44.
102. Villarreal, J.C.A., Crandall-Stotler, B.J., Hart, M.L., Long, F.G., Forrest, L.L., 2015. Divergence times and the evolution of morphological complexity in an early land plant lineage (Marchantiopsida) with a slow molecular rate. New Phytologist, 209 (4): 1734 – 1746.
103. Anderson, J.M., Anderson, H.M., Cruickshank, A.R.I., 1998. Late Triassic ecosystem of the Molteno/ Lower Elliot Biome of Southern Africa. Palaeontology, 41 (3): 387 – 421.
104. Brodie, P.B., 1845. A history of the fossil insects in the secondary rocks of England (London). 105. Harris, T.M., 1938. The British Rhaetic Flora. The British Museum (Natural History), London. 84 pp. 106. Harris, T.M., 1939. Naiadita, a fossil bryophyte with reproductive organs. Annales Bryologici, 12: 57 – 70. 107. Katagiri, T., Hagborg, A., 2015. Validation of ordinal and family names for a Triassic fossil liverwort, Naiadita
(Naiaditaceae, Marchantiopsida). Phytotaxa, 222 (2): 165 – 166. 108. Cookson, I.C., Dettmann, M.E., 1961. Reappraisal of the Mesozoic microspore genus Aequitriradites.
Palaeontology, 4: 425 – 427. 109. Dettmann, M.E., 1963. Upper Mesozoic microfloras from South-Eastern Australia. Proceedings of the Royal
Society of Victoria, 77: 1 – 148.

110. Warrington, G., 1973. British Triassic stratigraphy in the light of palynological studies. In: Vozzhennikova, T.K., Timofeev, T.B., (eds.) Mikrofossilii Drevneishikh Otlozhenii. 3rd International Palynological Conference, Proceedings; Nauka, Moscow, Russia.
111. Eshet, Y., 1990. Paleozoic-Mesozoic Palynology of Israel: I: Palynological aspects of the Permo-Triassic succession in the subsurface of Israel. Geological Survey of Israel, 81: 1 – 57.
112. Neuburg, M.F., 1960. Leafy mosses from the Permian deposits of Angaraland. Trudy Geologičeskij Institut Akademija Nauk SSSR, 43: 3 – 64.
113. Ignatov, M.S., 1990. Upper Permian mosses from the Russian Platform. Palaeontographica Abteilung B, 217: 147 – 189.
114. Jäger, H., Wierich, F., 2006. Palynostratigraphie. In: Amler, A., Stoppel, D. (eds), Stratigraphie von Deutschland VI. Unterkarbon (Mississippium): Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften: 41: 294 – 318.
115. Clayton, G., Coquel, R., Doubinger, J., Gueinn, K.J., Loboziak, S., Owens, B., Streel, M., 1977. Carboniferous miospores of Western Europe: illustration and zonation. Mededelingen Rijks Geologische Dienst: 29: 71pp.
116. Gehmlich, M., Linnemann, U., Tichomirowa, M., Gaitzsch, B., Bombach, K., 1998. Geochronologie invers gestapelter Deckenreste im Frankenberger Zwischengebirge. Terra Nostra, 98: 52 – 54.
117. Gaitzsch, B., Egenhoff, S., Hesse, S., Ehling, B.C., 2010. Variscan early molasses in the Saxo-Thuringian. In: Linnemann, U., Romer, R.L., (eds.) Pre-Mesozoic geology of Saxo-Thuringia; from the Cadomian active margin to the Variscan Orogen. Stuttgart, Schweizerbart: 311 – 322.
118. Goffinet, B., Buck, W.R., Shaw, J., 2009. Morphology, anatomy, and classification of the Bryophyta. In: Goffinet, B., Shaw, A.J., (eds.), Bryophyte Biology, 2nd edition, Cambridge University Press: 55 – 138.
119. Christiano de Souza, I.C., Branco, F.S.R., Vargas, Y.L. 2012. Permian bryophytes of Western Gondwanaland from the Paraná Basin in Brazil. Palaeontology, 55: 229 – 241.
120. Laenen, B., Shaw, B., Scheider, H., Goffinet, B., Paradis, E., Désamoré, A., Heinrichs, J., Villarreal, J.C., Gradstein, S.R., McDaniel, S.F., Long, D.G., Forrest, L.L., Hollingsworth, M.L., Crandall-Stotler, B., Davis, E.C., Engel, J., Von Konrat, M., Cooper, E.D., Patiño, J., Cox, C.J., Vanderpoorten, A., Shaw, A.J., 2014. Extant diversity of bryophytes emerged from successive post-Mesozoic diversification bursts. Nature Communications, 5: 6134.
121. Kotlyar, G.V., Belyansky, G.C., Burago, V.I., Nikitina, A.P., Zakharov, Y.D., Zhuravlev, A.V., 2006. South Primorye, Far East Russia – a key region for global Permian correlation. Journal of Asian Earth Sciences, 26: 280 – 293.
122. Zakharov, Y.D., Sha, J., Popov, A.M., Safronov, P.P., Shorochova, S.A., Volynets, E.B., Biakov, A.S., Burago, V.I., Zimina, V.G., Konovalova, I.V., 2009. Permian to earliest Cretaceous climatic oscillations in the eastern Asian continental margin (Sikhote-Alin area), as indicated by fossils and isotope data. GFF, 31: 25 – 47.
123. Henderson, C.M., Davydov, V.I., Wardlaw, B.R., 2012. The Permian Period. In: Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M. (eds.), The Geological Time Scale 2012, Volume 2, Chapter 24, Elsevier Press: 653 – 679.
124. Konopka, A.S., Herendeen, P.S., Smith Merrill, G.L., Crane, P.R., 1997. Sporophytes and gametophytes of Polytrichaceae from the Campanian (Late Cretaceous) of Georgia, U.S.A. International Journal of Plant Sciences, 158 (4): 489 – 499.
125. Rösler, O., Yano, O., 1994. Musgo fossil (Musci, Bryophyta) na Formação Terezina (Permiano Superior), Grupo Passa Dois, bacia do Paraná, próximo a Irati, Estado do Paraná. In: Septima Reunião de Paleobotánicos e Palinólogos, Resumo das Comunicações. São Paulo, Brazil: 68 – 69.
126. Santos, R.V., Souza, P.A., Alvarenga, C.J.S., Dantas, E.L., Pimentel, M.M., Oliveira, C.G., Araujo, L.M., 2006. Shrimp U-Pb zircon dating and palynology of bentonitic layers from the Permian Irati Formation, Paraná Basin, Brazil. Gondwana Research, 9: 456 – 463.
127. Frahm, J.-P., 2010. Die Laubmoosflora des Baltischen Bernsteinwales. Jena (Weissdorn Verlag). 128. Frahm, J.-P., Gröhn, C., 2013. More fossil bryophytes from Baltic amber. Archive for Bryology, 159: 1 – 9. 129. Frahm, J.-P., Newton, A.E., 2005. A new contribution to the moss flora of Dominican amber. Bryologist 108: 526
– 536. 130. Shelton, G.W.K., Stockey, R.A., Rothwell, G.W., Tomescu, A.M.F., 2015. Exploring the fossil history of
pleurocarpous mosses: Tricostaceae fam. nov. from the Cretaceous of Vancouver Island, Canada. Am. J. Bot., 102: 1883 – 1900.
131. Shelton, G.W.K., Stockey, R.A., Rothwell, G.W., Tomescu, A.M.F., 2016. Krassiloviella limbelloides gen. et sp. nov.: additional diversity in the Hypnanaean moss family Tricostaceae (Valanginian, Vancouver Island, British Columbia). International Journal of Plant Sciences, 177: 792 – 808.
132. Konopka, A.S., Herendeen, P.S., Crane, P.R., 1998. Sporophytes and gametophytes of Dicranaceae from the Santonian (Late Cretaceous) of Georgia, USA. American Journal of Botany, 85: 714 – 723.

133. Renzaglia, K.S., Villarreal, J.C., Duff, J., 2009. New insights into morphology, anatomy, and systematics of hornworts. In: Goffinet, B., Shaw, A.J., (eds.) Bryophyte Biology, 2nd edition, Cambridge University Press: 139 – 171.
134. Kotyk, M.E., Basinger, J.F., Gensel, P.G., de Frietas, T.A., 2002. Morphologically complex plant microfossils from the Late Silurian of Arctic Canada. American Journal of Botany, 89 (6): 1004 – 1013.
135. Klapper, G., Murphy, M. A., 1974. Silurian - lower Devonian conodont sequence in the Roberts Mountains Formation of central Nevada. University of California Publications in Geological Sciences, 111: 1 – 62.
136. Thorsteinsson, R., 1980. Stratigraphy and conodonts of Upper Silurian and Lower Devonian rocks in the environs of the Boothia Uplift Canadian Arctic Archipelago. 1. Contributions to stratigraphy. Geological Survey of Canada Bulletin, VIII – 38.
137. Uyeno, T. T., 1990. Biostratigraphy and conodont faunas of Upper Ordovician through Middle Devonian rocks, eastern Arctic Archipelago. Geological Survey of Canada, Bulletin, 401: 1 – 211.
138. Mayr, U., Brent, T.A., de Freitas, T., Frisch, T., Nowlan, G.S., Okulitch, A.V., 2004. Geology of Eastern Prince of Wales Island and Adjacent Smaller Islands, Nunavut. Geological Survey of Canada, Bulletin, 574: 1 – 99.
139. Steemans, P., Le Hérissé, A., Melvin, J., Miller, M.A., Paris, F., Verniers, J., Wellman, C.H., 2009. Origin and radiation of the earliest vascular land plants. Science, 324: 353.
140. Paris, F., Le Hérisse, A., Monod, O., Kozlu, H., Ghienne, J.-F., Dean, W.T., Vecoli, M., Günay, Y., 2007. Ordovician chitinozoans and acritarchs from southern and southeastern Turkey. Revue de micropaléontologie, 50: 81 – 107.
141. Vandenbroucke, T. R. A., Williams, M., Zalasiewicz, J. A., Davies, J. R., Waters, R. A., 2008. Integrated Upper Ordovician graptolite-chitinozoan biostratigraphy of the Cardigan and Whitland areas, southwest Wales. Geological Magazine, 145: 199 – 214.
142. Edwards, D., 1970. Fertile Rhyniophytina from the Lower Devonian of Britain. Palaeontology, 13: 451 – 461. 143. Gonez, P., Gerrienne, P., 2010. A new definition and a lectotypification of the genus Cooksonia Lang 1937.
International Journal of Plant Sciences, 171: 199 – 215. 144. Gonez, P., Gerrienne, P., 2010. Aberlemnia caledonica gen. et comb. nov., a new name for Cooksonia caledonica
Edwards 1970. Review of Palaeobotany and Palynology, 163: 64 – 72. 145. Lang, W.H., Cookson, I.C., 1935. On a flora, including vascular land plants, associated with Monograptus, in
rocks of Silurian age, from Victoria, Australia. Phil. Trans. R. Soc. Lond., Ser. B, 219: 133 – 163. 146. Garratt, M.J., 1978. New evidence for a Silurian (Ludlow) age for the earliest Baragwanathia flora. Alcheringa, 2:
217 – 224. 147. Rickards, R.B., 2000. The age of the earliest club mosses: the Silurian Baragwanthia flora in Victoria, Australia.
Geological Magazine, 137: 207 – 209. 148. Jaeger, H., 1966. Two late Monograptus species from Victoria, Australia, and their significance for datiing the
Baragwanathia flora. Proc. R. Soc. Vict., 79: 393 – 413. 149. Wang, Y., Fu, Q., Xu, H., Hao, S., 2007. A new Late Silurian plant with complex branching from Xinjiang,
China. Alcheringa, 31: 111 – 120. 150. Hao, S.G., Beck, C.B., 1993. Further observations on Eophyllophyton bellum from the Lower Devonian
(Siegenian) of Yunnan, China. Palaeontographica Abteilung B-Palaophytologie, 230: 27 41. 151. Hao, S.G., Gensel, P.G., 1998. Some new plant finds from the Posongchong Formation of Yunnan, and
consideration of a phytogeographic similarity between south China and Australia during the Early Devonian. Science in China Series D-Earth Sciences, 41: 1 – 13.
152. Xue, J.-Z., Hao, S.-G., Zhu, X., Wang, D., 2012. A new basal euphyllophyte, Pauthecophyton gen. nov., from the Lower Devonian (Pragian) of Yunnan, China. Review of Palaeobotany and Palynology, 183: 9 – 20.
153. Gensel, P.G., Albright, V.M., 2006. Leclercqia complexa from the Early Devonian (Emsian) of northern New Brunswick, Canada. Review of Palaeobotany and Palynology, 142: 103 – 121.
154. Wellman, C.H., Gensel, P.G., Taylor, W.A., 2009. Spore wall ultrastructure in the early lycopsid Leclerqia (Protolepidodendrales) from the Lower Devonian of North America: evidence for a fundamental division in the Lycopsids. American Journal of Botany, 96: 1849 – 1860.
155. Schweitzer, H.-J., 1980. Uber Drepanophycus spinaeformis Göppert. Bonner Paläobotanische Mitteilungen 7: 1 – 29.
156. Li, C.-S., Edwards, D., 1995. A re-investigation of Halle’s Drepanophycus spinaeformis Göpp. from the Lower Devonian of Yunnan Province, southern China. Botanical Journal of the Linnean Society, 118: 163 – 192.
157. Banks, H.P., Grierson, J.D., 1968. Drepanophycus spinaeformis Göppert in the early Upper Devonian of New York State. Palaeontographica, B, 123: 113-120.
158. Kidston, R., Lang, W.H., 1920. On Old Red Sandstone plants showing structure, from the Rhynie Chert Bed, Aberdeenshire. Part 3. Asteroxylon mackiei, Kidston and Lang. Transactions of the Royal Society of Edinburgh, 52: 643 – 680.
159. Parry, S.F., Noble, S.R., Crowley, Q.G., Wellman, C.H., 2011. A high precision U-Pb age constraint on the Rhynie Chert Konservat-Lagerstätte: a time scale and other implications. Journal of the Geological Society of

London, 168: 863 – 872. 160. Kräusel, R., Weyland, H., 1937. Pflanzenreste aus dem Devon. X. Zwei Pflanzenfunde im Oberdevon der Eifel.
Senckenbergiana, 19: 338 – 355. 161. Chaloner, W.G., 1967. Lycophyta. In: Boureau, E., Jovet-Ast, S., Høeg, O.A., Chaloner, W.G., (eds.), Traité de
paléobotanique, vol. 2, Paris, Masson et Cie. 162. Hao, S., Wang, D., Wang, Q., Xue, J., 2006. A new lycopsid, Zhenglia radiata gen. et sp. nov., from the Lower
Devonian Posongchong Formation of Southeastern Yunnan, China, and its evolutionary significance. Acta Geologica Sinica, 80: 11 – 19.
163. Bonamo, P.M., 1977. Rellimia thomsonii (Progymnospermopsida) from the Middle Devonian of New York State. American Journal of Botany, 64 (10): 1272 – 1285.
164. Bartholomew, A.J., Brett, C.E., 2007. Correlation of Middle Devonian Hamilton Group-equivalent strata in east-central North America: implications for eustasy, tectonics and faunal provinciality. Geological Society, London, Special Publications, 278: 105-131.
165. Johnson, J.G., Klapper, G., Sandberg, C.A., 1985. Devonian eustatic fluctuations in Euramerica. Geological Society of America Bulletin 96, 567.
166. Cantino, P.D., Doyle, J.A., Graham, S.W., Judd, W.S., Olmstead, R.G., Soltis, D.E., Soltis, P.S., Donoghue, M.J., 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon, 56: 822 – 846.
167. Hao, S.G., Wang, D.M., Wang, Q., 2004. A new species of Estinnophyton from the Lower Devonian Posongchong Formation, Yunnan, China; its phylogenetic and palaeophytogeographical significance. Botanical Journal of the Linnean Society, 146: 201 – 216.
168. Hao, S.G, Xue, J.Z., 2013. The Early Devonian Posongchong Flora of Yunnan – a contribution to an understanding of the evolution and early diversification of vascular plants. Science Press Beijing: 366pp.
169. Gerrienne, P., 1992. The Emsian plants from Fooz-Wépion (Belgium). III. Foozia minuta gen. et spec. nov., a new taxon with probable cladoxylalean affinities. Review of Palaeobotany and Palynology, 74: 139 – 157.
170. Skog, H.J, Banks, H.P. 1973. Ibyka amphikoma, gen. et sp. n., a new protoarticulate precursor from the late Middle Devonian of New York State. American Journal of Botany, 60: 366 – 380.
171. Fisher, D.W., Isachsen, Y.W., Rickard, L. V., Broughton, J. G., Offield, T. W., 1962. Geologic map of New York. New York State Museum Sciece Service, Geological Survey.
172. Rickard, L.V., 1964. Correlation of the Devonian rocks in New York State. Map and Chart Series 4. New York State Musuem Science Service Geological Survey.
173. Klapper, G., 1981. Review of New York Devonian conodont biostratigraphy. In: Oliver, W. A., Jr., Klapper, G., (eds.), Devonian biostratigraphy of New York, Part I. IUGS SDS: 57 – 68.
174. Kirchgasser, W. T., 2000. Correlation of stage boundaries in the Appalachian Devonian, Eastern United States. Courier Forschungsinstitut Senckenberg, 225: 271 – 284.
175. Banks, H.P., 1975. Reclassification of Psilophyta. Taxon, 24 (4): 401 – 413. 176. Hao, S.G., Gensel, P.G., Wang, D.M., 2001. Polythecophyton demissum, gen. et sp. nov., a new plant from the
Lower Devonian (Pragian) of Yunnan, China and its phytogeographic significance. Review of Palaeobotany and Palynology, 116: 55 – 71.
177. Bek, J., Pšenička, J., 2001. Senftenbergia plumosa (Artis) emend. and its spores from the Carboniferous of the Kladno and Pilsen Basins, Bohemian Massif, and some related and synonymous taxa. Review of Palaeobotany and Palynology, 116: 213 – 232.
178. DiMichele, W.A., Phillips, T.L., 1977. Monocyclic Psaronius from the lower Pennsylvanian of the Illinois Basin. Canadian Journal of Botany, 55: 2514 – 2524.
179. Rothwell, G.W., Stockey, R.A., 1989. Fossil Ophioglossales in the Paleocene of western North America. American Journal of Botany, 76: 637 – 644.
180. Pšenička, J., Bek, J., 2003. Cuticles and spores of Senftenbergia plumosa (Artis) Bek and Pšenička from the Carboniferous of Pilsen Basin, Bohemian Massif. Review of Palaeobotany and Palynology, 125: 299 – 312.
181. Davydov, V. I., Korn, D., Schmitz, M. D., 2012. The Carboniferous Period. In: Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M., (eds), The Geologic Time Scale 2012, Volume 2, Chapter 23, Elsevier Press: 603 – 651.
182. Anderson, R.C., Malone, D.H., Nelson, R.S., Frankie, W.T., Jacobson, R.J., 1999. Guide to the Geology of the Rock Island – Milan Area, Rock Island County. Field Trip Guidebook 1999B, Dept. of Natural Resources, Illinois State Geological Survey.
183. Leary, R.L., 1974. Stratigraphy and floral chararcteristics of the basal Pennsylvanian strata in west central Illinois. Compte Rendu, 7th International Congress on Stratigraphy and Geology of the Carboniferous, Kredfield, Germany, 1971, 3: 341 – 350.
184. Leary, R.L., 1974. Two early Pennsylvanian floras of western Illinois. Trans. III. State Acad. Sci., 67: 430 – 440. 185. Leary, R.L., 1975. Topography and sedimentation at the Mississippian-Pennsylvanian unconformity in Rock
Island, Ill. Abst. 68th Annu. Meet. Ill. State Acad. Sci. Trans., 68: 189.

186. Richards, B.C., 2013. Current status of the International Carboniferous time scale. In: Lucas, S.G., DiMichele, W.A., Barrick, J.E., Schneider, J.W., Spielmann, J.A., (eds.), The Carboniferous – Permian Transition, New Mexico Museum of Natural History and Science, Bulletin 60, Albuquerque: 348 – 353.
187. Trivett, M.L., 1992. Growth, architecture, structure, and relationships of Cordaixylon iowensis nov. comb. (Cordaitales). International Journal of Plant Sciences, 153, 273 – 287.
188. Janousek, T.J., Pope, J.P., 2014. Petrology, petrography and conodont biostratigraphy of the Laddsdale Coal interval, along Whitebreast Creek, Bauer, Iowa. GSA North-Central Section, 48th Annual Meeting, Abstracts,16: 5.
189. Heckel, P.H., 2013. Pennsylvanian stratigraphy of Northern Midcontinent Shelf and biostratigraphic correlation of cyclothems. Stratigraphy, 10: 3 – 39.
190. Barrick, J.E., Lambert, L.L., Heckel, P.H., Rosscoe, S.J., Boardman, D.R., 2013. Midcontinent Pennsylvanian conodont zonation. Stratigraphy, 10: 55 – 72.
191. Prestianni, C., 2005. Early diversification of seeds and seed-like structures. In: Steemans, P., Javaux, E., (eds.), Precambrian to Palaeozoic Palaeopalynology and Palaeobotany, Carnets de Geologie/Notebooks on Geology, Brest, Memoir 2005/02, 33-38.
192. Rothwell, G.W., Scheckler, S.E., Gillespie, W.H., 1989. Elkinsia gen nov, a late Devonian gymnospermn with cupulate ovules. Botanical Gazette, 150: 170 – 189.
193. Streel, M., Scheckler, S.E., 1990. Miospore lateral distribution in upper Fammenian alluvial lagoonal to tidal facies from eastern United States and Belgium. Review of Palaeobotany and Palynology, 64: 315 – 324.
194. Streel, M., Higgs, K., Loboziak, S., Riegel, W., Steemans, P., 1987. Spore stratigraphy and correlation with faunas and floras in the type marine Devonian of the Ardenne-Rhenish regions. Review of Palaeobotany and Palynology, 50: 211 – 229.
195. House, M.R., Gradstein, F.M., 2004. The Devonian Period. In: Gradstein, F. M., Ogg J. G., Smith, A. G., (eds.) A Geologic Time Scale 2004, Cambridge University Press: 202-221.
196. Gao, Z., Thomas, B.A., 1989. A review of fossil cycad megasporophylls, with new evidence of Crossozamia Pomel and its associated leaves from the lower Permian of Taiyuan, China. Review of Palaeobotany and Palynology, 60: 205 – 223.
197. Nagalingum, N.S., Marshall, C.R., Quental, T.B., Rai, H.S., Little, D.P., Mathews, S., 2011. Recent synchronous radiation of a living fossil. Science, 334: 796 – 799.
198. Hermsen, E.J., Taylor, T.N., Taylor, E.L., Stevenson, D.W., 2006. Cataphylls of the Middle Triassic cycad Antarcticycas schopfii and new insights into cycad evolution. American Journal of Botany, 93: 724 – 738.
199. Wang, J., 2010. Late Paleozoic macrofloral assemblages from Weibei Coalfield, with reference to vegetational change through the Late Paleozoic Ice-age in the North China Block. International Journal of Coal Geology, 83: 292 – 317.
200. Wieland, G.W., 1935. The Cerro Cuadrado petrified forest. Carnegie Institution of Washington Publication, 449: 1 – 183.
201. Calder, M.G. 1953. A coniferous petrified forest in Patagonia. Bulletin of the British Museum (Natural History): Geology, 2: 99 – 138.
202. Stockey, R.A., 1975. Seeds and embryos of Araucaria mirabilis. American Journal of Botany, 62: 856 – 868. 203. Stockey, R.A., 1978. Reproductive biology of Cerro Cuadrado fossil conifers: Ontogeny and reproductive
strategies in Araucaria mirabilis (Spegazzini) Windhausen. Palaeontographica Abteilung B, 166: 1 – 15. 204. Wilde, M.H., Eames, A.J. 1948. The ovule and seed of Araucaria bidwillii with discussion of the taxonomy of the
genus. I. Morphology. Annals of Botany, 12, 311 – 326. 205. Spalleti, L., Iñiguez Rodríguez, A.M., Masón, M., 1982. Edades radimétricas de piroclastitas y volvanitas del
Grupo Bahía Laura, Gran Bajo de San Julián, Santa Cruz. Revista de la Asociación Geológica Argentina, 37: 483 – 485.
206. Rydin, C., Wu, S.Q., Friis, E.M., 2006. Liaoxia Cao et S Q Wu (Gnetales): Ephedroids from the Early Cretaceous Yixian Formation in Liaoning, Northeastern China. Plant Systematics and Evolution, 262: 239 – 265.
207. Chang, S.C., Zhang, H.C., Renne, P.R., Fang, Y., 2009. High precision Ar-40/Ar-39 age for the Jehol Biota. Palaeogeography, Palaeoclimatology, Palaeoecology, 280: 94 – 104.
208. Townrow, J.A., 1967. On Rissikia and Mataia podocarpaceous conifers from the lower Mesozoic of Southern lands. Papers and Proceedings of the Royal Society of Tasmania, 101: 103 – 136.
209. Florin, R., 1951. Evolution in cordaites and conifers. Acta Horti Bergiani, 15: 285-388. 210. Yao, X.L., Taylor, T.N., Taylor, E.L., 1997. A taxodiaceous seed cone from the Triassic of Antarctica. American
Journal of Botany, 84: 343 – 354. 211. Axsmith, B.J., Taylor, T.N., Taylor, E.L., 1998. Anatomically preserved leaves of the conifer Notophytum
krauselii (Podocarpaceae) from the Triassic of Antarctica. American Journal of Botany, 85: 704 – 713. 212. Krassilov, V.A., 1986. New floral structure from the Lower Cretaceous of Lake Baikal Area. Review of
Palaeobotany and Palynology, 47: 9 – 16.

213. Rydin, C., Pedersen, K.R., Friis, E.M., 2004. On the evolutionary history of Ephedra: Cretaceous fossils and extant molecules. Proceedings of the National Academy of Sciences, USA, 101: 16571 – 16576.
214. Friis, E.M., Pedersen, K.R., Crane, P.R., 2009. Early Cretaceous mesofossils from Portugal and eastern North America related to the Bennettitales-Erdtmanithecales-Gnetales group. American Journal of Botany, 96: 252 – 283.
215. Godefroit, P., 2012. Bernissart dinosaurs and Early Cretaceous terrestrial ecosystems. Indiana University Press. 216 Vakhrameev, V., Kotova, I., 1977. Ancient angiosperms and accompanying plants from the Lower Cretaceous of
Transbaikalia. Paleontological Journal, 4: 487 – 495. 217. Vakhrameev, V., 1991. Jurassic and Cretaceous floras and climates of the Earth. Cambridge University Press. 218. Chen, P., Wang, Q., Zhang, H., Cao, M., Li, W., Wu, S., Shen, Y., 2005. Jianshangou Bed of the Yixian
Formation in West Liaoning, China. Science in China Series D: Earth Sciences, 48: 298 – 312. 219. Dettmann, M.E., Thomson, M.R.A., 1987. Cretaceous palynomorphs from the James-Ross Island area, Antarctica
- a pilot-study. British Antarctic Survey Bulletin, 77: 13 – 59. 220. Ogg, J.G., Hinnov, L.A., 2012. The Cretaceous Period. In: Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M.
(eds.), The Geological Time Scale 2012, Volume 2, Chapter 27, Elsevier Press: 793 – 853. 221. Chang, S.-C., Zhang, H., Hemming, S.R., Mesko, G.T., Fang, Y., 2012. Chronological evidence for extension of
the Jehol Biota into Southern China. Palaeogeography, Palaeoclimatology, Palaeoecology, 344–345: 1 – 5. 222. He, H.Y., Wang, X.L., Zhou, Z.H., Wang, F., Boven, A., Shi, G.H., Zhu, R.X., 2004. Timing of the Jiufotang
Formation (Jehol Group) in Liaoning, northeastern China, and its implications. Geophysical Research Letters, 31: 1 – 4.
223. Hughes, N. F., McDougall, A.B., 1990. Barremian–Aptian angiospermid pollen records from southern England. Review of Palaeobotany and Palynology, 65: 145 – 151.
224. Judd, W.S., Olmstead, R.G., 2004. A survey of tricolpate (eudicot) phylogenetic relationships. American Journal of Botany, 91: 1627 – 1644.
225. Hochuli, P.A., Feist-Burkhardt, S., 2004. A boreal early cradle of angiosperms? Angiosperm-like pollen from the Middle Triassic of the Barents Sea (Norway). Journal of Micropalaeontology, 23: 97 – 104.
226. Hochuli, P.A., Colin, J.P., Vigran, J.O., 1989. Triassic biostratigraphy of the Barents Sea area. In: Collinson, J.D., (ed.), Correlation in Hydrocarbon Exploration. London, UK: Norwegian Petroleum Society, Graham and Trotman Ltd.: 131 – 153.
227. Vigran, J.O., Mangerud, G., Mørk, A., Bugge, T., Weitschat, W., 1998. Biostratigraphy and sequence stratigraphy of the Lower and Middle Triassic deposits from the Svalis Dome, central Barents Sea, Norway. Palynology, 22: 89 – 141.
228. Liu, Z. J., Wang, X., 2016. A perfect flower from the Jurassic of China. Historical Biology, 28: 707 – 719, doi:10.1080/08912963.2015.1020423.
229. Han, G. Liu, Z., Liu, X., Mao, L., Jacques, F.M.B., Wang, X., 2016. A whole plant herbaceous angiosperm from the Middle Jurassic of China. Acta Geologica Sinica, 90: 19 – 29.
230. Liu, Z.-J., Wang, X., 2016. Yuhania: a unique angiosperm from the Middle Jurassic of Inner Mongolia, China. Historical Biology: 1 – 11, doi:10.1080/08912963.2015.1020423.
231. Mohr, B.A.R., Bernardes-de-Oliveira, M.E.C., 2004. Endressinia brasiliana, a magnolialean angiosperm from the lower Cretaceous Crato Formation (Brazil). International Journal of Plant Sciences, 165: 1121 – 1133.
232. Massoni, J., Doyle, J.A., Sauquet, H., 2014. Fossil calibration of Magnoliidae, an ancient lineage of angiosperms. Palaeontologica Electronica: 18.1.2FC.
233. Mohr, B.A.R., Coiffard, C., Bernardes-de-Oliveira, M.E.C., 2013. Schenkeriphyllum glanduliferum, a new magnolialean angiosperm from the Early Cretaceous of Northern Gondwana and its relationships to fossil and modern Magnoliales. Review of Palaeobotany and Palynology, 189: 57 – 72, doi:10.1016/j.revpalbo.2012.08.004.
234. Sauquet, H., Doyle, J.A., Scharaschkin, T., Borsch, T., Hilu, K.W., Chatrou, L.W., Le Thomas, A., 2003. Phylogenetic analysis of Magnoliales and Myristicaceae based on multiple data sets: implications for character evolution. Botanical Journal of the Linnean Society, 142: 125 – 186.
235. Doyle, J.A., Endress, P.K., 2010. Integrating Early Cretaceous fossils into the phylogeny of living angiosperms: Magnoliidae and eudicots. Journal of Systematics and Evolution, 48: 1 – 35, doi:10.1111/j.1759-6831.2009.00058.x.
236. Doyle, J.A., 2015. Recognising angiosperm clades in the Early Cretaceous fossil record. Historical Biology, 27: 414 – 429, doi:10.1080/08912963.2014.938235.
237. Batten, D.J., 2007. Spores and pollen from the Crato Formation: biostratigraphic and palaeoenvironmental implications. In: Martill D.M., Bechly, G., Loveridge R.F., (eds), The Crato fossil beds of Brazil - window into an ancient world. Cambridge University Press: 566 – 573.
238. Martill, D. M., 2007. The age of the Cretaceous Santana Formation fossil Konservat Lagerstatten of north-east Brazil: a historical review and an appraisal of the biochronostratigraphic utility of its palaeobiota. Cretaceous Research, 28: 895 – 920, doi:10.1016/j.cretres.2007.01.002.

239. Heimhofer, U., Hochuli, P.-A., 2010. Early Cretaceous angiosperm pollen from a low-latitude succession (Araripe Basin, NE Brazil). Review of Palaeobotany & Palynology, 161: 105 – 126, doi:10.1016/j.revpalbo.2010.03.010.
240. Smith, S.Y., Stockey, R.A., 2007. Establishing a fossil record for the perianthless Piperales: Saururus tuckerae sp. nov. (Saururaceae) from the Middle Eocene Princeton Chert. American Journal of Botany, 94: 1642 – 1657.
241. Rouse, G.E., Mathews, W.H., 1961. Radioactive dating of Tertiary plant-bearing deposits. Science, 133: 1079 – 1080.
242. Mathews, W.H., 1964. Potassium-argon age determinations of Cenozoic volcanic rocks from British Columbia. Geological Society of America Bulletin, 75: 465 – 468.
243. Hills, L. V., Baadsgaard, H., 1967. Potassium-argon dating of some Lower Tertiary strata in British Columbia. Bulletin of Canadian Petroleum Geology, 15: 138 – 149.
244. Read, P.B., 2000. Geology and industrial minerals of the Tertiary basins, south-central British Columbia. British Columbia Geological Survey Geo-File 2000.
245. Moss, P.T., Greenwood, D.R., Archibald, S.B. 2005. Regional and local vegetation community dynamics of the Eocene Okanagan Highlands (British Columbia – Washington State) from palynology. Canadian Journal of Earth Sciences, 42: 187 – 204.
246. Dilcher, D. L., Sun, G., Ji, Q. & Li, H. Q., 2007. An early infructescence Hyrcantha decussata (comb. nov.) from the Yixian Formation in northeastern China. Proceedings of the National Academy of Sciences of the United States of America, 104: 9370 – 9374, doi:10.1073/pnas.0703497104.
247. Wang, W., Dilcher, D.L., Sun, G., Wang, H.-S., Chen, Z.-D., 2016. Accelerated evolution of early angiosperms: Evidence from ranunculalean phylogeny by integrating living and fossil data. Journal of Systematics and Evolution, 54: 336 – 341, doi:10.1111/jse.12090.
248. Crepet, W.L., Nixon, K.C., 1998. Fossil Clusiaceae from the Late Cretaceous (Turonian) of New Jersey and implications regarding the history of bee pollination. American Journal of Botany, 85: 1122 – 1133.
249. Crepet, W.L., Nixon, K.C., Daghlian, C.P., 2012. Fossil Ericales from the Upper Cretceous of New Jersey. International Journal of Plant Sciences, 174: 572 – 584.
250. Ruhfel, B.R., Stevens, P.F., Davis, C.C., 2013. Combined morphological and molecular phylogeny of the clusioid clade (Malpighiales) and the placement of the ancient rosid macrofossil Paleoclusia. International Journal of Plant Sciences, 174: 910 – 936, doi:10.1086/670668.
251. Hickey, L.J., Doyle, J.A., 1977. Early Cretaceous fossil evidence for angiosperm evolution. Botanical Review, 43: 1 – 104
252. Doyle, J.A., Endress, P.K., Upchurch, G.R., Jr., 2008. Early Cretaceous monocots: a phylogenetic evaluation. Sbornik Narodniho Muzea v Praze Rada B Prirodni Vedy, 64: 59 – 87.
253. Hochuli, P.A., Heimhofer, U., Weissert, H., 2006. Timing of early angiosperm radiation: recalibrating the classical succession. Journal of the Geological Society, 163: 587 – 594.
254. Hughes, N.F., 1994. The enigma of angiosperm origins. Cambridge University Press. 255. Gandolfo, M.A., Nixon, K.C., Crepet, W.L., 2002. Triuridaceae fossil flowers from the Upper Cretaceous of New
Jersey. American Journal of Botany, 89: 1940 – 1957. 256. Berry, E.W., 1914. The Upper Cretaceous and Eocene floras of South Carolina and Georgia. United States
Geological Survey Professional Paper, 84: 1 – 200. 257. Hertweck, K.L., Kinney, M.S., Stuart, S.A., Maurin, O., Mathews, S., Chase, M.W., Gandolfo, M.A., Pires, J.C.,
2015. Phylogenetics, divergence times and diversification from three genomic partitions in monocots. Botanical Journal of the Linnean Society, 178: 375 – 393, doi:10.1111/boj.12260.
258. Iles, W.J.D., Smith, S.Y., Gandolfo, M.A., Graham, S.W., 2015. Monocot fossils suitable for molecular dating analyses. Botanical Journal of the Linnean Society, 178: 346 – 374, doi:10.1111/boj.12233.
259. Sohl, N.F., Owens, J.P., 1991. In: Horton, J.W.Jr., Zullo, V.A. (eds.,), The Geology of the Carolinas: Knoxville. University of Tennessee Press: 191 – 220.
260. Campbell, B.G., Gohn, G.S., 1994. Stratigraphic framework for geologic and geohydrologic studies of the subsurface Cretaceous section near Charleston, South Carolina. United States Geological Survey Map MF-2273, 1 – 11.
261. Habib, D., Miller, J.A., 1989. Dinoflagellate species and organic facies evidence of marine transgression and regression in the atlantic coastal plain. Palaeogeography, Palaeoclimatology, Palaeoecology, 74: 23 – 47, doi:http://dx.doi.org/10.1016/0031-0182(89)90018-7.
262. Christopher, R.A., Prowell, D. C., 2010. A palynological biozonation for the uppermost Santonian and Campanian Stages (Upper Cretaceous) of South Carolina, USA. Cretaceous Research, 31: 101 – 129, doi:10.1016/j.cretres.2009.09.004.
Copyright © 2022 FDOKUMEN




















